 Valentina Valdés-Castro
Valentina Valdés-Castro Camila Fernandez
Camila Fernandez- 1Centro de Investigación Oceanográfica COPAS Sur-Austral, Universidad de Concepción, Concepción, Chile
- 2Interdisciplinary Center for Aquaculture Research (INCAR), Universidad de Concepción, Concepción, Chile
- 3Sorbonne Universités, UPMC Univ Paris 06, CNRS, Laboratoire d’Océanographie Microbienne (LOMIC), Observatoire Océanologique, Banyuls-sur-Mer, France
- 4Centro FONDAP de Investigación en Dinámica de Ecosistemas Marinos de Altas Latitudes (IDEAL), Valdivia, Chile
Chile is the second largest global producer of farmed salmon. The growth of salmon production has not been free of environmental challenges, such as the increasing use of pesticides to control the parasitic load of the sea lice Caligus rogercresseyi. The lack of the specificity of pesticides can potentially affect non-target organisms, as well as the structure and functioning of aquatic ecosystems. The aim of this study, was to understand the effect of pesticides on natural microbial communities to the addition of the anti-lice pesticide azamethiphos, deltamethrin and emamectin benzoate, and their potential impact in ammonium uptake rates in the coast off central-southern Chile and Northern Patagonia. The addition of pesticides on natural microbial communities resulted in a rapid response in ammonium uptake, which was significant for the single use of pesticide, azamethiphos and emamectin benzoate, as well as the combination, azamethiphos, deltamethrin and emamectin benzoate. In northern Patagonia, azamethiphos addition produced a 53% decrease in photoautotrophic uptake. However, an increase, although variable, was observed in chemoautotrophic uptake. Emamectin benzoate produced a 36 to 77% decrease in chemo and photoautotrophic ammonium uptake, respectively. The combined use of pesticides, also produced up to 42% decrease in both photo and chemoautotrophic assimilation. We conclude that the use of pesticides in salmon farming produces diverse responses at the microbial level, stimulating and/or inhibiting microbial communities with subsequent impact on nitrogen budgets. Further studies are necessary to understand the impact of pesticides in the ecology of central-southern and northern Patagonia, Chile.
Introduction
Chile is the second largest global producer of farmed salmon, with an annual production of over 700 thousand tons (Avendaño-Herrera, 2018). The growth of Chilean salmon production has not been free of sanitary and environmental difficulties, such as benthic accumulation of organic matter, eutrophication, massive escapes of farmed salmon, input of chemicals such as antibiotic and pesticides into the ecosystem, fish diseases, and social issues (Burridge et al., 2010; Hargrave, 2010; Baskett et al., 2013; Dresdner et al., 2019; Quiñones et al., 2019).
The intensive use of pesticides to control sea lice infections of Caligus rogercresseyi has become a key environmental concern for Chilean aquaculture (Millanao et al., 2011; Núñez-Acuña et al., 2015; Avendaño-Herrera, 2018). Infestations result in severe skin erosion and epidermal hemorrhage, leading to chronic stress, reduced growth (González and Carvajal, 2003; Rozas and Asencio, 2007; González et al., 2015) and increasing the vulnerability of fish to other diseases (Bravo, 2003; Johnson et al., 2004; Dresdner et al., 2019).
Since the earliest reported infections, sea lice in Chile have been controlled using chemicals originally used in agriculture (Reyes and Bravo, 1983). Feed additives such as emamectin benzoate have been used since the 1990s (Bravo, 2003; Bravo et al., 2008) and later complemented by pyrethroids such as deltamethrin in 2007 due to the appearance of resistance of C. rogercresseyi to emamectin benzoate. In 2009, the Chilean authorities allowed the use of another pyrethroid, cypermethrin. Diflubenzuron was used between 2008 and 2012 and finally the organophosphate azamethiphos was introduced in Chile in 2013 (Helgesen et al., 2014).
Currently, the most commonly used products in Chile are the organophosphate azamethiphos and the pyrethroid deltamethrin. They are applied in situ by bath solution and are subsequently discharged into the surrounding environment in large volumes (Nash, 2003; Burridge et al., 2010; Gebauer et al., 2017). Helgesen et al. (2014) reported that 677 kg of active compound cypermethrin and 197 kg of deltamethrin were discarded in 2012.
The anti-lice treatments lack specificity and therefore may affect indigenous organisms with the potential to affect sensitive non-target species (Johnson et al., 2004). However, until now the studies available describing the impacts of pesticides in non-target organism are scarce and are mainly focus in invertebrates (Burridge et al., 2004; Knapp et al., 2005; Gebauer et al., 2017). Studies focusing on microorganism are even more scarce, and recently was reported that the use of pesticides in marine waters can produce changes in microbial photo and chemoautotrophic carbon uptake (Rain-Franco et al., 2018).
Microorganism are important habitants of aquatic ecosystems, playing a pivotal role in primary productivity and nutrient recycling. In the traditional view of the marine nitrogen (N) cycle, phytoplankton use dissolved inorganic N as nitrate (NO3–) or ammonium (NH4+) to create biomass while heterotrophic bacterioplankton remineralize dissolved organic N into the inorganic forms hence supporting primary production. By being in contact with pesticides, microbial community structure and metabolic fluxes can be modified, an effect that varies according to their chemical form and characteristics while interacting with the nutrient pool of the water column (Rain-Franco et al., 2018). This can result in an alteration of biogeochemical fluxes and local primary production.
To understand the impact of pesticides on non-target organisms and biogeochemistry, we investigated the potential effect of three pesticides used against C. rogercresseyi (azamethiphos, deltamethrin, and emamectin benzoate) on photo and chemoautotrophic ammonium uptake in two key regions of the Chilean coast that hold high levels of biological productivity but contrasting levels of influence from salmon farming.
Materials and Methods
Study Area and Sampling Strategy
The study was conducted in two sites in central-southern and norther Patagonia, Chile: Llico Bay and Caucahue Channel, respectively. Llico Bay is a close bay located south of Gulf of Arauco in central Chile (37.1°S 73.5°W; Figure 1A) in an area untouched by salmon farming while, Caucahue Channel was located in the inland sea of Chiloe island, Northern Patagonia (42.1°S 73.4°W; Figure 1B) and is intensely influenced by salmon farming. Llico Bay was visited 4 times on board of R/V Kay Kay II (Universidad de Concepción) between July 2015 and April 2016 (Table 1). Caucahue Channel was visited twice on board the L/M “Don José” in June 2014 and January 2015 (Table 1).
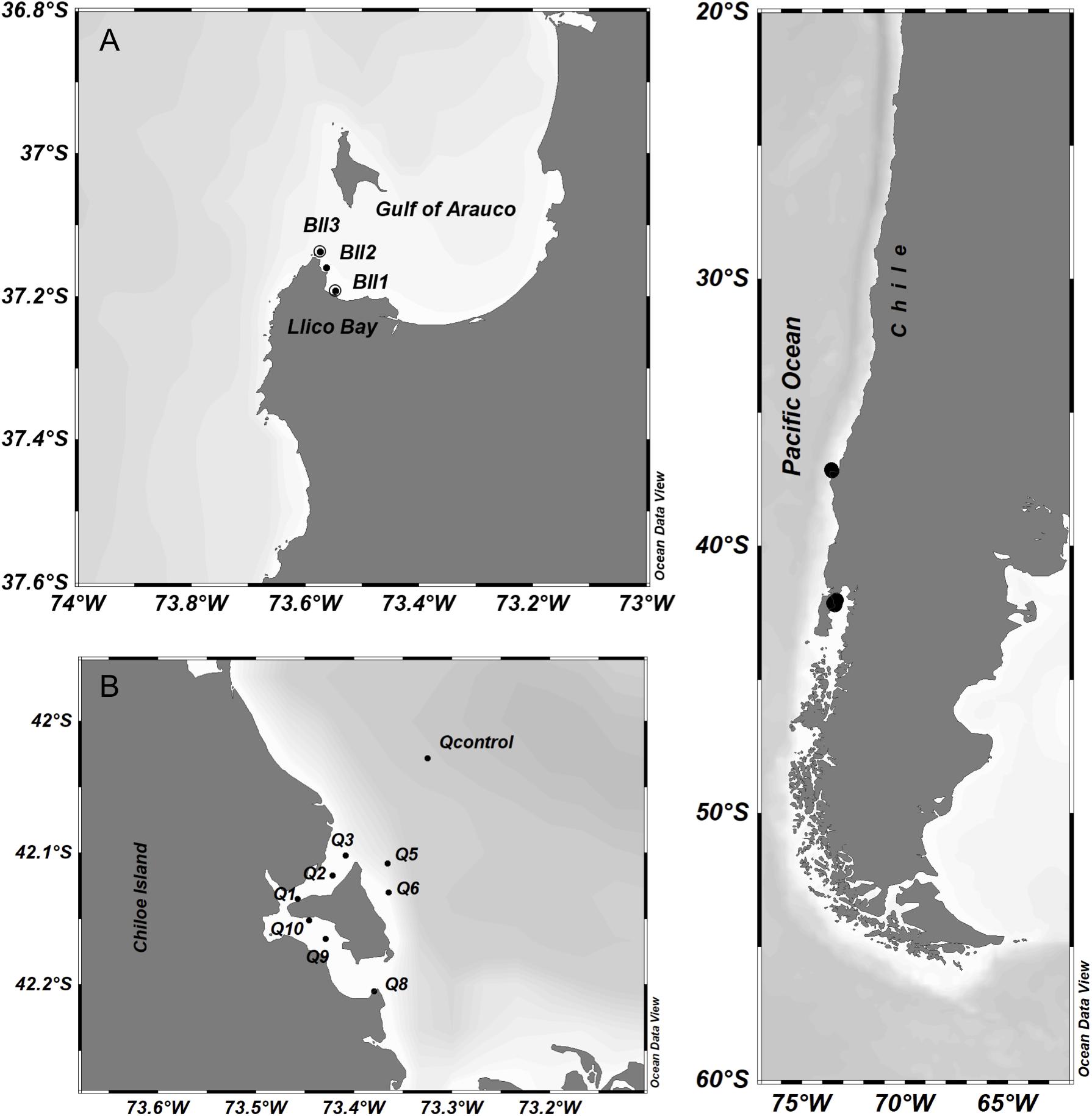
Figure 1. Study area in central-southern and Norther Patagonia Chile showing the sampling location in (A) Llico Bay in Gulf of Arauco and (B) Caucahue channel in the inner sea of Chiloé. Circles indicate sampling stations.
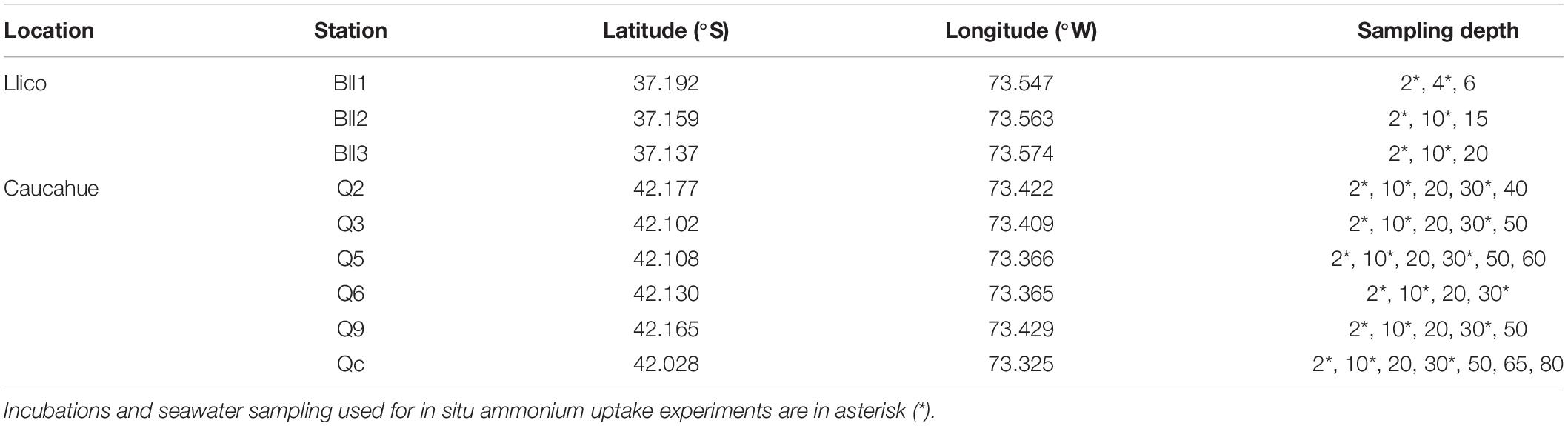
Table 1. Geographical location and seawater sampling depths for hydrographic characterization at Llico Bay and Caucahue channel stations.
During the sampling, we first assessed physical parameters such as temperature and salinity (CTD Minus; AML Oceanographic, Canada) at the three stations in Llico (Figure 1 and Table 1). During Caucahue Channel sampling, we used a CTDO data sensor (SAIV A/S, Norway; Figure 1 and Table 1). Chemical and biological samples were collected with Niskin bottles at all sampling stations (Table 1). Ammonium samples were taken in duplicate using 50 mL glass bottles (Duran Schott) and analyzed by the fluorometric method described by Holmes et al. (1999), using a Turner Designs fluorometer. Samples for nitrate (NO3–), nitrite (NO2–) and phosphate (PO43–) were stored in high-density polyethylene (HDPE) plastic bottles (11 mL in duplicate) at −20°C until analyses by standard colorimetric techniques (Aminot and Kérouel, 2007). Bacterioplankton abundance was analyzed by flow cytometry (Marie et al., 1997) at Laboratory for oceanographic Processes and Climate (PROFC; Universidad de Concepción, Chile) with a FACSCalibur flow cytometer (Becton Dickinson). Total chlorophyll-a (Chl-a) were estimated using a Turner design fluorometer according (Holm-Hansen et al., 1965).
Isotopic Analysis and in situ Ammonium Uptake Experiments
Rates of ammonium uptake (ρNH4+) were determined using the 13C/15NH4+ stable isotope technique (Fernández et al., 2009; Fernandez and Farías, 2012). Results of Carbon-13 tracer were published previously by Rain-Franco et al. (2018), which describes carbon fluxes for the same experiments. For ammonium uptake experiments, seawater samples taken from depths described in Table 1 were incubated in 600 mL polycarbonate bottles using 15NH4Cl (ICON isotopes IN 5037; 99% at 0.5 μmol mL–1). 15N-ammonium tracer additions were generally kept close to 10% of ambient ammonium concentration.
Incubations were done using an in situ mooring line deployed at sunrise and recovered at sunset (∼7 h later). Once recovered, the bottles were filtered through 0.7 μm GF/F filters (Whatman; pre-combusted at 540°C, 4 h) using a vacuum pump. Filters were stored at −20°C until analysis by isotope mass spectrometry at the Laboratory of Biogeochemistry and Applied Stable Isotopes (LABASI) of Pontificia Universidad Católica de Chile using a Thermo Delta V Advantage IRMS coupled with Flash2000 Elemental Analyzer.
Deltamethrin and azamethiphos were added to primary production incubations at dosses determined according to the concentrations used in sea lice treatments: 3 μg L–1 for deltamethrin (Siwicki et al., 2010) and 200 μg L–1 for azamethiphos (Davies, 2001; Canty et al., 2007; Burridge et al., 2010). Stock solutions of azamethiphos (Dr. Ehrenstorfer GmbH, 98.5% purity) and deltamethrin (Dr. Ehrenstorfer GmbH, 99.5% purity) were prepared by dissolving the reagents in dimethylsulphoxide (DMSO, MERCK, 99.9% analytical grade). The concentration of the pesticide solution added was 1.2 and 3 μmol L–1 for azamethiphos and deltamethrin, respectively, as described in the Table 2. Bottles without pesticides were incubated as controls (15NH4; Table 2).
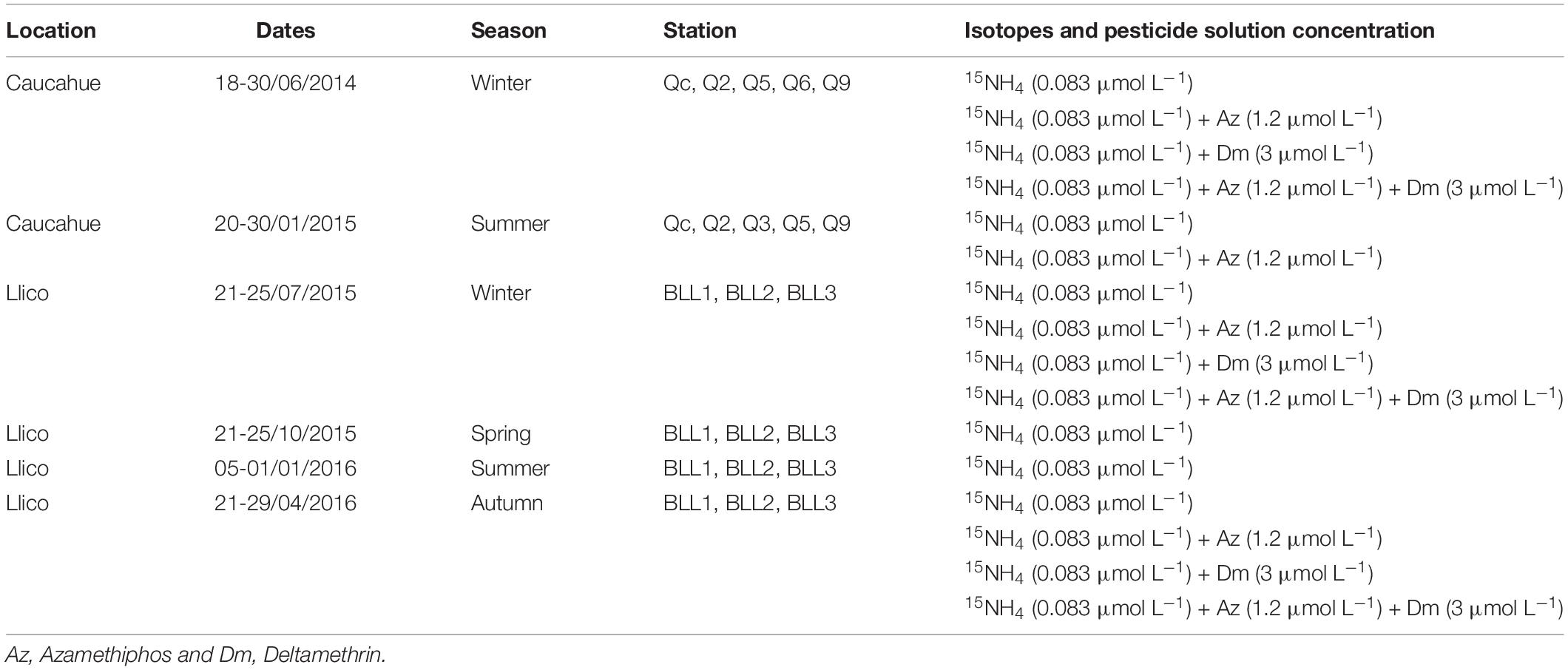
Table 2. Summary of in situ ammonium assimilation experiment setup during Llico and Caucahue campaigns.
Seawater sampling and incubation depths at the sampling points were set between surface and the compensation depth (which in this case coincided with app ∼2.7 times the depth of view of the Secchi disk). At this level the light availability allows phytoplankton growth and the oxygen produced is equal to the consumed by respiration. Incubation and sampling depths for Llico Bay were at 2 and 4 m (Bll1) and 2 and 10 m depth (Bll2 and Bll3). At Caucahue Channel the depths sampling and for incubation were 2, 10 and 30 m depth (Table 1). More details of treatments and controls for in situ ammonium uptake experiments were described in Table 2.
On-Deck Ammonium Uptake Experiments
To study ammonium photo and chemoautotrophic uptake, four on-deck experiments were performed using dual stable isotopes techniques (13C/15NH4). Results of Carbon-13 tracer were published in Rain-Franco et al. (2018). On-deck experiments were developed at Llico Bay (Bll2 station) with surface waters (2 m), during winter and spring 2015, and summer and autumn 2016.
In order to simulate solar radiation, we performed atmospheric measurements of incident Photosynthetically Active Radiation (PAR) using a portable radiometer (RM-21 Dr. Gröbel, Germany) and these results are available in Rain-Franco et al. (2018). Photoautotrophic samples were incubated under a combination of two cut off filters to simulate the irradiance at the sampling depths: 0.3 Neutral density 209 (47% of average transmittance between 400 and 700 nm, Lee filters) and Steel Blue 117 (Lee filters). Dark treatments to obtain chemoautotrophic ammonium uptake were put in a closed incubator. Incubations were done in Duran Schott bottles (580 mL) and amended with a 15NH4 solution (ICON isotopes IN 5037; 99% at 0.5 μmol mL–1; final concentration in the sample in the 83 nmol L–1) and pesticide solution as described in Table 3. The pesticide treatments used were azamethiphos (15NH4 + Az), deltamethrin (15NH4 + Dm) and emamectin benzoate (15NH4 + Em). The dose of emamectin benzoate (0.66 mmol L–1, Dr. Ehrenstorfer GmbH) was estimated in order to obtain the highest possible experimental concentration by dividing the standard into two equal parts. Also, combined treatments were evaluated, azamethiphos-deltamethrin (15NH4 + Az + Dm) and azamethiphos-deltamethrin-emamectin benzoate (15NH4 + Az + Dm + Em). All the incubations (light and dark) were done in duplicate for 6 h, with a subsample at 2 h. Bottles were incubated in 3 large aquariums of 50 L each, which were interconnected and received surface seawater with a continuous flow to maintain in situ temperature conditions (ca. 13°C). More details of treatments and controls of on-deck incubation are available at Table 3.
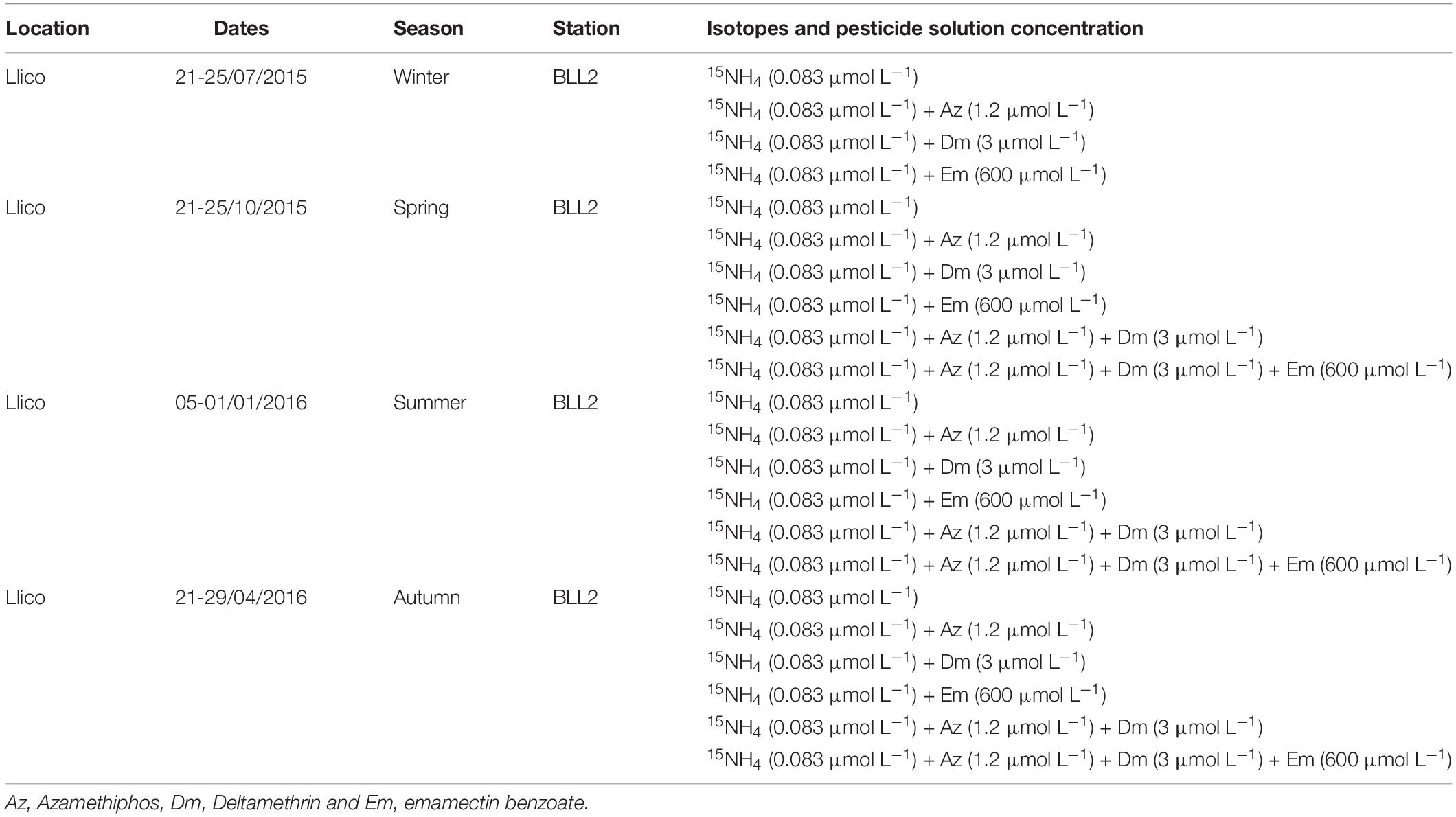
Table 3. Summary of on-deck ammonium uptake incubations in Llico campaigns.
Data Analysis
Ammonium uptake rates were estimated as described in previous studies (Fernández et al., 2009; Fernandez and Farías, 2012) following Eq. (1):
where Vf is the filtrated volume, PON represents the amount of particulate N obtained by mass spectrometry (μg) and%RPON is the enrichment in 15NH4 in the GF/F filter after the incubation. %RDIN is the excess of enrichment of 15N after the inoculation (T0) computed according to Eq. (2):
where 15N is the volume of isotopic solution (15NH4), 15DIN represents the concentration of the 15N added inoculums. The term 0.0036912 represents the natural abundance (average) of 15N. DINi represents the concentration of DIN in the sample before the addition of tracer. Vb represent the incubation bottle volume. For in situ incubations, hourly rates were multiplied by 12 in order to obtain daily ammonium fixation values. Initial ammonium concentrations were taken from a nutrient profile prior to incubation and subsamples for final ammonium concentrations were taken directly from the incubation bottle before filtration.
For Llico Bay and Caucahue Channel in situ incubations a paired t-test was done to compare treatments with their respective control conditions. For on-deck incubations (Llico Bay), a two-way ANOVA was computed to evaluate significant variations among treatments and between incubation times after checking for normality assumptions (Kolmogorov-Smirnov test) and homoscedasticity (Levene’s test). Whenever data did not pass the test, a log transformation was implemented. Pairwise multiple comparison was performed using a Tukey test as a posteriori analysis. All the statistical analyses were performed using the statistical software R1.
Results
Chemical and Biological Seasonal Variability in Llico Bay and Caucahue Channel
Temperature and salinity in Llico Bay varied between seasons, showed a marked thermal stratification during spring 2015 and summer 2016 in Bll1 and Bll2 stations, the maximum surface values was nearly 17°C at Bll1 station in summer 2016 and the maximum salinity values was ∼ 35 in summer 2016 for the three stations (Supplementary Figure 1). Whereas in winter and autumn lower temperature values was observed (∼ 12°C in winter) and as well lower salinity values (∼ 32°C in winter 2015). Ammonium showed higher concentrations in autumn 2016 and spring 2015 compared with winter 2015 and summer 2016 at Bll1 and Bll2 stations. Station Bll3 showed the highest ammonium concentration and reached 9 μmol L–1 at 20 m depth in autumn 2016 (Figures 2A,E,I). Nitrate concentration showed a marked seasonal variability with higher concentrations in spring 2015 followed by winter 2015. The lowest nitrate concentration was observed in summer 2016 in almost all depths (Figures 2B,F,J). Nitrite was higher in winter 2015 at all stations (Figures 2C,G,K). The N/P ratio varied seasonally, although maximum values were observed in spring and winter 2015, particularly Bll3 station showed an average ratio of 25 (Figures 2D,H,L).

Figure 2. Ammonium (μmol L–1), nitrate (μmol L–1), nitrite (μmol L–1) concentrations, N/P ratio, and Chl-a concentrations (mg m–3) profiles at Llico Bay station in winter and spring 2015, and summer and autumn 2016. (A–E) Bll1, (F–J) Bll2, and (K-O) Bll3.
The highest concentrations of Chl-a were observed in summer 2016 in the three stations (Figure 2), and the maximum value was observed in surface at Bll3 station (∼ 23 mg m–3; Figure 2O). and The lowest concentration was observed in autumn 2016 at Bll1, Bll2, and Bll3, at all depths. The highest bacterioplankton abundance was observed in spring 2015, with the maximum abundance at station Bll3 (3,990 × 103 cell mL–1). Minor changes were observed in winter 2015, summer 2016 and autumn 2016 (Supplementary Figure 2).
A seasonal variability was also observed at Caucahue channel stations, lower temperature was observed during winter (∼11°C) compared with summer (∼16°C; Supplementary Figure 3). Salinity showed a strong vertical variation in winter, particularly at the station Q6. Ammonium concentration was higher in winter compared to summer in Caucahue Channel (Figures 3A,F), specifically at station Q2 (5 μmol L–1) in surface waters, followed by station Q6 with 3 μmol L–1 at 20 m depth. The lowest concentration was found at the control station (Qc) with values close to zero throughout the water column (Figure 3A). In summer, the concentrations were generally below 0.3 μmol L–1 for all stations, with the exception of Q2 station (>1 μmol L–1; Figure 3F). As well of summer, in winter, ammonium concentrations at Qc station were nearly zero throughout the water column.
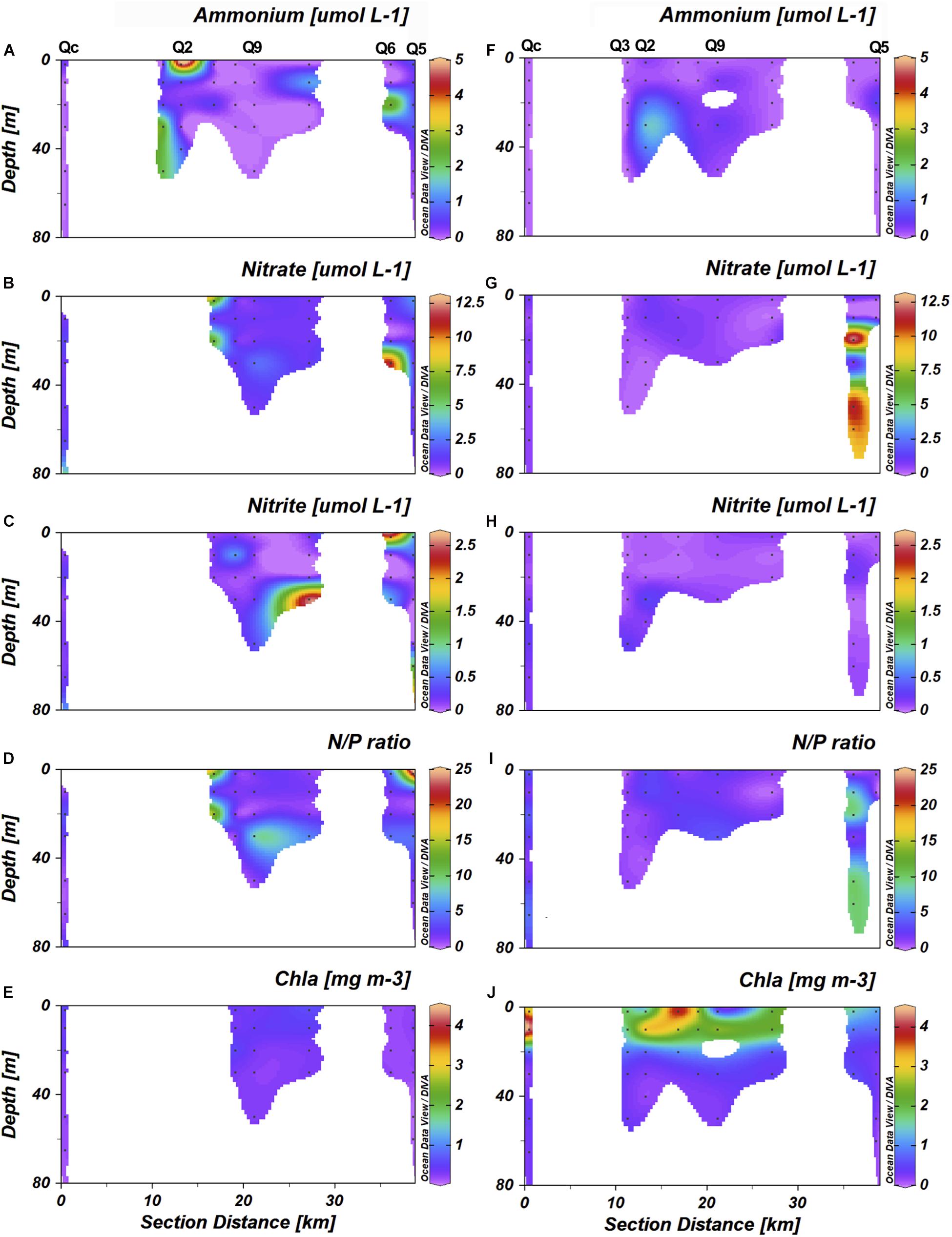
Figure 3. Ammonium (μmol L–1), nitrate (μmol L–1), nitrite (μmol L–1), N/P ratio and Chl-a concentrations transects at Caucahue channel stations in winter 2014 (A–E) and summer 2015 (F–J).
Nitrate concentrations in winter and summer were in general below 3 μmol L–1, except at station Q1 and Q6 (Figures 3B–G), with maximum concentration of 11 μmol L–1 at 30 m depth and during summer, station Q5 showed maximum concentration of 11.5 μmol L–1 at 20. Nitrite concentration were in general higher in winter compared to summer (Figures 3C–H). During winter, nitrite concentration tends to increase with the depth, the highest values was found at station Q6 (2 μmol L–1). In summer, the concentration was lower than 0.5 μmol L–1. The N/P ratio varied greatly depending on stations and depths. Station Qc had the lowest ratios, 1.71 ± 1.17 in winter and 2.84 ± 1.14 in summer (Figures 3D–I). The highest ratio was observed at station Q5 in winter 22.8 at 2 m.
Chl-a concentration was higher in summer at the station Qc and between stations Q3 and Q2, nearly 4 mg m–3 (Figures 3E–J). Concentration in winter were lower than 1 mg m–3 for all stations. Bacterioplankton in Caucahue Channel showed a marked seasonal influence. In winter showed abundances under the 500 × 103 cell mL–1 and during summer in general the abundances were over the 500 × 103 cell mL–1 (Supplementary Figures 2E–F). In winter bacterioplankton was more abundant at station Q9 and lowest at station Q5. In summer, maximum abundances were observed at stations Q9 and the lowest abundances were observed in general at control station (Supplementary Information; Figure 2B).
In situ Ammonium Uptake Rates at Llico Bay and Caucahue Channel
Ammonium uptake rates were higher in surface waters compared to deeper levels (10 m) at all stations and seasons (Figure 4). At station Bll1 assimilation rates observed in spring 2015, were 93 and 95% higher than winter 2015 and autumn 2016 (7.9 and 10.4 μmol L–1d–1 at 2 and 4 m depth, respectively; Figures 4A–C). Station Bll2 showed the highest ρNH4+ values during autumn 2016 reaching 3 μmol L–1d–1 at 2 m and the minimum rate was observed in spring at 10 m depth (0.5 μmol L–1d–1). The same pattern was observed in Bll3 with higher ammonium uptake during autumn 2016 at 2 m depth (2.5 μmol L–1d–1) and a decrease in 81% at 10 m depth. The lowest rate was observed in winter 2015 at 10 m depth (0.9 μmol L–1d–1; Figures 4H–J).
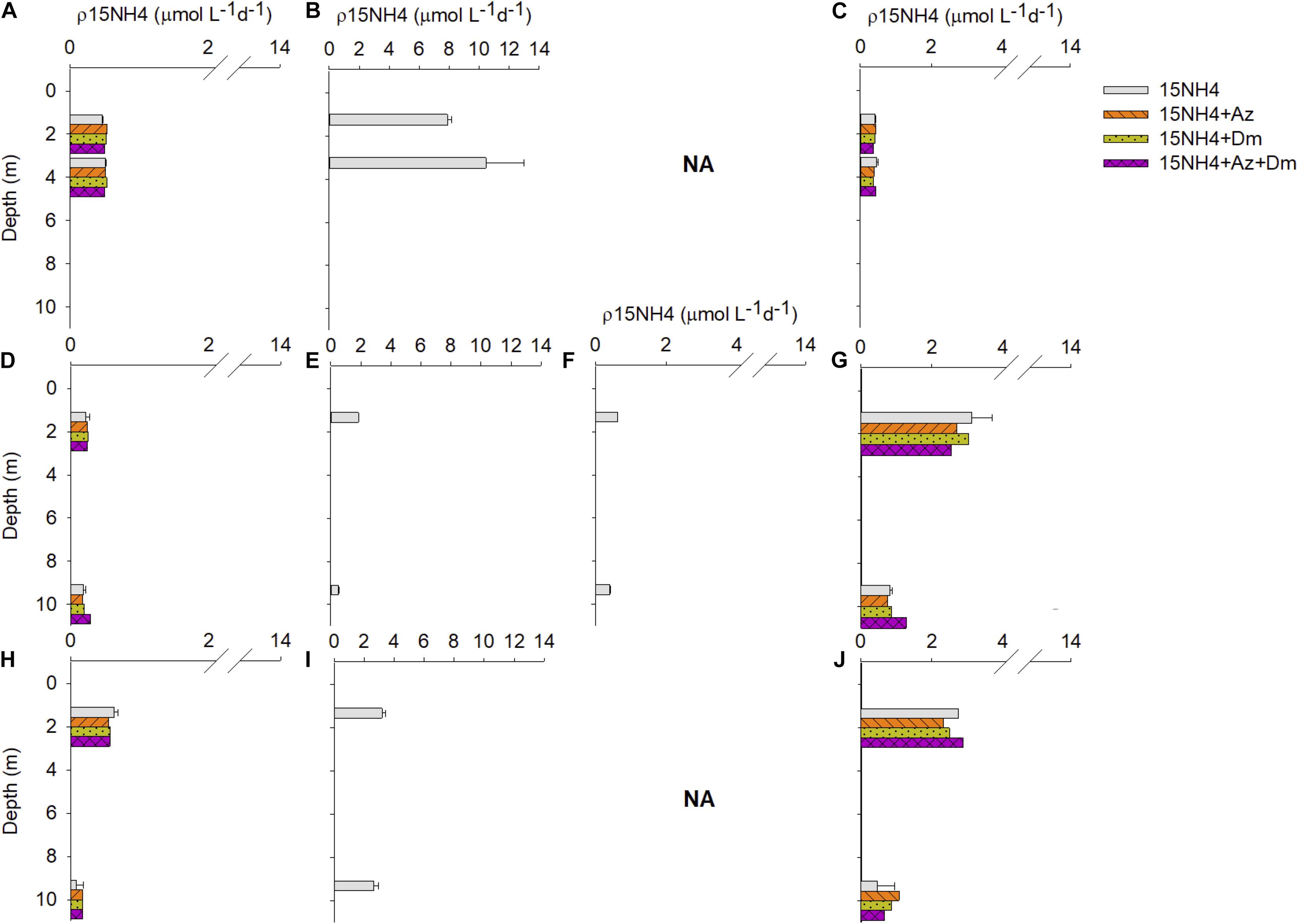
Figure 4. In situ ammonium uptake rates (μmol L–1 d–1) profiles at Llico Bay. Upper panels for Bll1 station, middle panels for Bll2 and lower panels for Bll3 station. (A,D,H) Winter 2015; (B,E,I) Spring 2015; (F) Summer 2016; (C,G,J) Autumn 2016. NA are not available data for Bll1 and Bll3 station in Summer 2016. Errors bars represent the standard deviations.
In treatments where pesticides were added, ammonium uptake showed variable responses compared to the control (15NH4; Table 2). The addition of pesticides caused an increase in ammonium assimilation in winter 2015 (Figures 4A,D,H) compared to controls, particularly at station Bll3 at 10 m depth, in which rates in all treatments (15NH4 + Az, 15NH4 + Dm, and 15NH4 + Az + Dm) increased by 42% compared to their controls. At Bll1 and Bll2 the rates increased between 2 and 15% compared to the controls (15NH4), except for the combined treatment 15NH4 + Az + Dm at Bll1 station and 15NH4 + Az at 10 m at station Bll2.
In general, the addition of pesticides in autumn 2016 resulted in a decrease of ammonium utilization rates compared to the controls. At Bll1 station rates were 20% lower than the control (15NH4) for the treatment 15NH4 + Dm (4 m; Figure 4C). At Bll2 station the rates decreased between 3 and 18%, in the treatment with deltamethrin and in the combined treatment 15NH4 + Az + Dm at 2 m, respectively. However, at this station was also observed an increase in a 55% the ρNH4+ values for 15NH4 + Az + Dm treatment at 10 m depth. In the station Bll3, an inhibition was only observed in the treatments with a single pesticide (15NH4 + Az and 15NH4 + Dm) at 2 m depth. In subsurface layer (10 m) the addition of pesticides caused an increase in the ammonium assimilation rates up to 132% compared to the control for the treatment 15NH4 + Az. This increase was also observed for 15NH4 + Dm (89%) and 15NH4 + Az + Dm (46%; Figure 4J). However, all these differences reported were non-significant (t-test > 0.05) for pesticide additions at any station during winter 2015 and autumn 2016. For spring 2015 and summer 2016 no data are available for treatments with pesticides (Figure 4).
In Caucahue channel, ammonium uptake varied throughout the stations and also between seasons and depths. The highest ρNH4+ values were observed at station Q2 in winter (0.5 μmol L–1 d–1) and in summer (0.6 μmol L–1 d–1) in surface layer (Figures 5, 6, respectively) inside of the area influenced by the farming centers, and the lowest rates was observed at station Q9 in both season (Figures 5E, 6E). In summer 2015, the chemoautotrophic ammonium utilization (dark treatment) was 10 times higher than the photoautotrophic nitrogen uptake in surface at Q9, but in general the both photo and chemoautotrophic are in the same range of values.
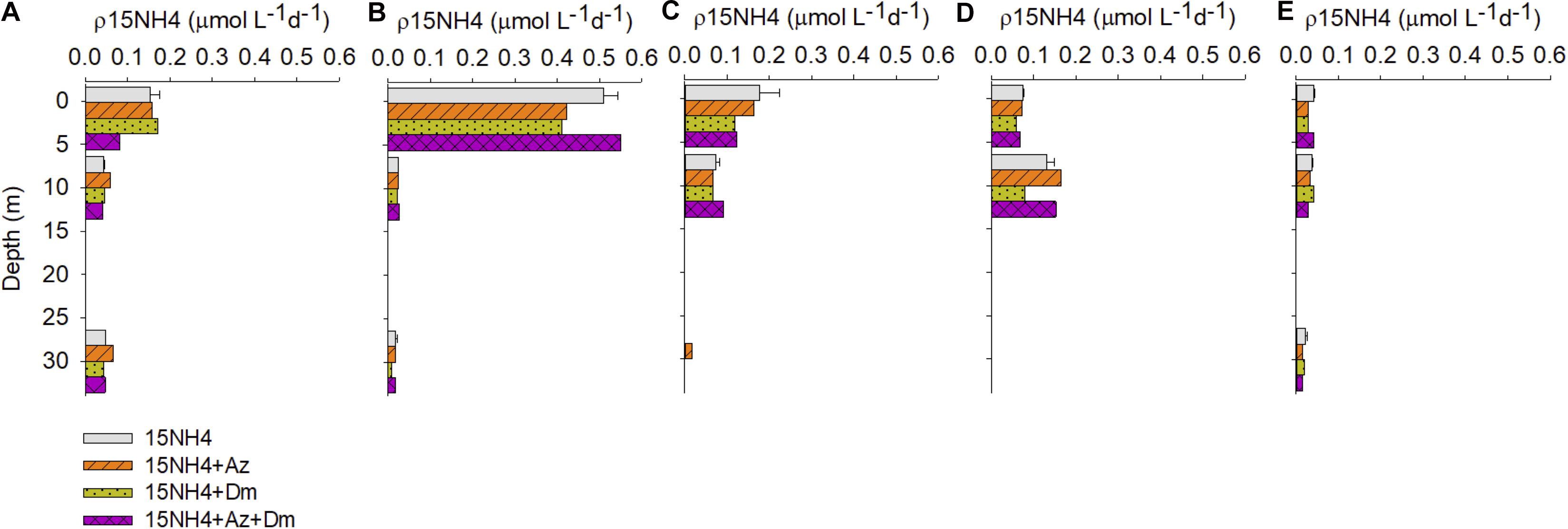
Figure 5. In situ ammonium uptake rates (μmol L–1 d–1) at Caucahue Channel stations during winter 2014. (A) Qc, (B) Q2, (C) Q5, (D) Q6, and (E) Q9 station.
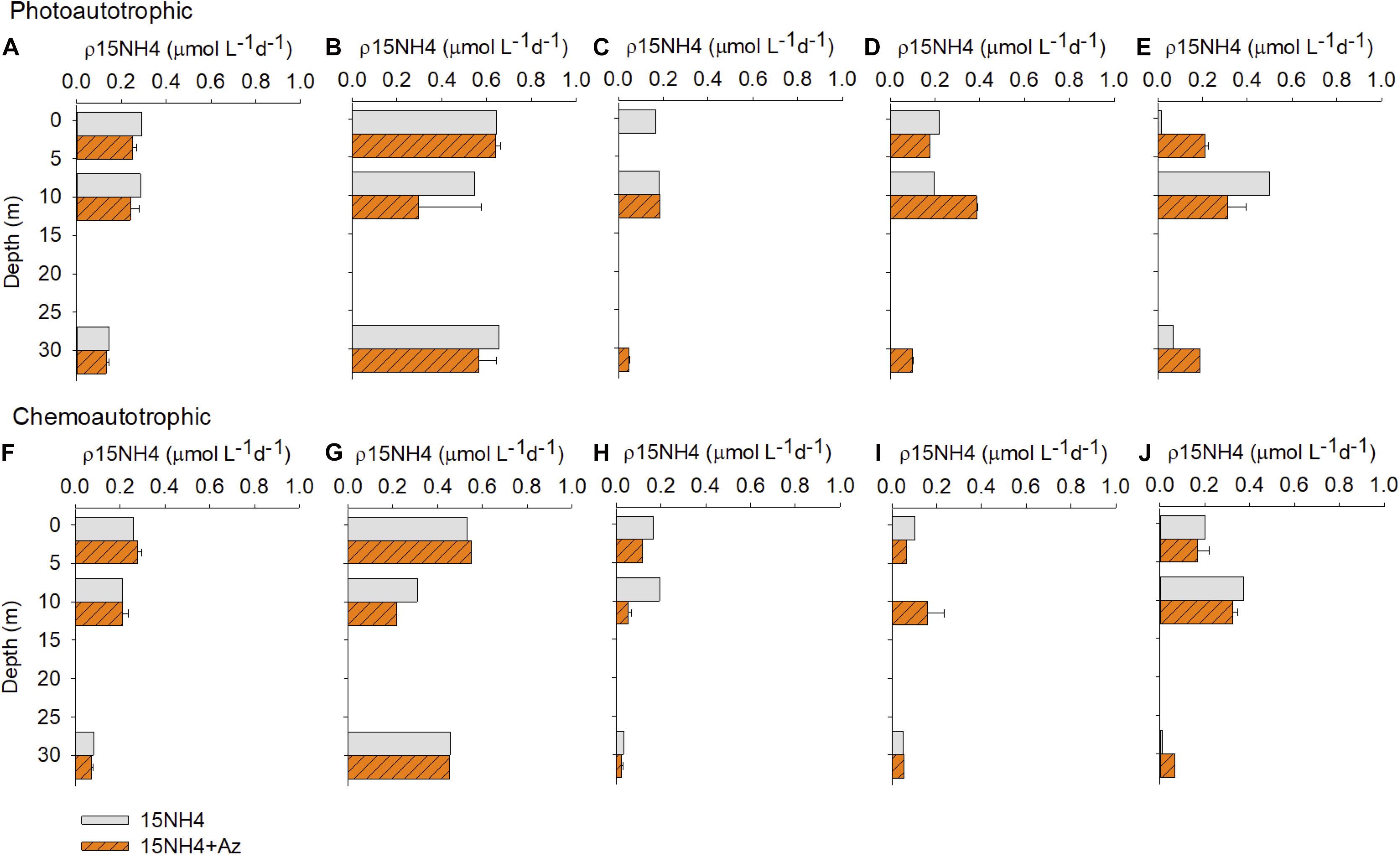
Figure 6. In situ photoautotrophic (upper panel) and chemoautotrophic (lower panel) ammonium uptake rates (μmol L–1 d–1) during summer 2015. (A,F) Qc; (B,G) Q2; (C,H) Q3; (D,I) Q5; (E,J) Q9 station.
In winter 2014 (Figure 5), pesticide additions resulted in an increase in ammonium uptake in the treatment 15NH4 + Az at station Qc between 3% (2 m) and 40% (30 m). In general, deltamethrin addition resulted in a decrease in ammonium assimilation rates, the highest difference found with the control (15NH4) was at station Q2 (Figure 5B). At this station which was located close to the farms, ρNH4+ decreased by 53% (30 m depth), followed by a reduction of 33% at the station Q5 in surface (Figure 5C). However, deltamethrin addition cause an increase in the rates of ammonium assimilation in the first 10 m depth at station Qc, between 7 and 12% higher than the controls (Figure 5A).
The combined treatment 15NH4 + Az + Dm also resulted in a variable response to the pesticide addition (Figure 5). Qc station showed a decrease in the ammonium assimilation rates up to 47% at the surface. This pattern was also observed at station Q9, observed rates up to 29% less than the controls. Higher rates compared to the control was observed at Q2, up to 13% higher at 10 m depth. At the same depth, an increase was observed at stations Q5 (13%) and Q6 (23%). However, these values were not significantly different from the control treatment (15NH4; paired t-test, p < 0.05).
In summer (Figure 6), the in situ incubations were made under light (photoautotrophic) and dark conditions (chemoautotrophic). At station Qc, the addition of azamethiphos resulted in a significant decrease in ammonium uptake under light conditions (in average 13%; paired t-test > 0.05). However, we also observed an increase in ρNH4+ values at 2 and 30 m in the station Q9, being up to 100 times higher the rates obtained after the addition of azamethiphos. However, these increases were not significant, since they were not observed for all depths (paired t-test > 0.05). The chemoautotrophic uptake showed a decrease thought the water column only at station Q3 between 23 and 72% lower than the controls. Slight increase was observed for the chemoautotrophic ammonium utilization (Figures 6F,G,I,J) after the addition of azamethiphos, except for station Q9 in which we observed rates 570% higher than the control (Figure 6J). But, these effects were not significant (paired t-test > 0.05).
Pesticide Effect on Surface Ammonium Uptake Under Light and Dark Conditions: On-Deck Experiments
On-deck ammonium uptake experiments (Figure 7) showed higher photoautotrophic control rates (15NH4) in autumn 2016 (0.14 μmol L–1 h–1) after 2 h of incubation followed by spring 2015 (0.1 μmol L–1 h–1) at the same incubation time (Figures 7A,B). On the other hand, the highest chemoautotrophic ammonium uptake rate was observed in spring 2015 (0.1 μmol L–1 h–1). This rate was equivalent to 100% of photoautotrophic ammonium assimilation after 2 h of incubation, and after 6 h of incubation the chemoautotrophic assimilation was 7% higher than photoautotrophic ammonium assimilation (Figures 7E–H). This was also observed in summer 2016 when the chemoautotrophic ammonium uptake was equivalent to an 85% of the photoautotrophic at 2 h of incubation.
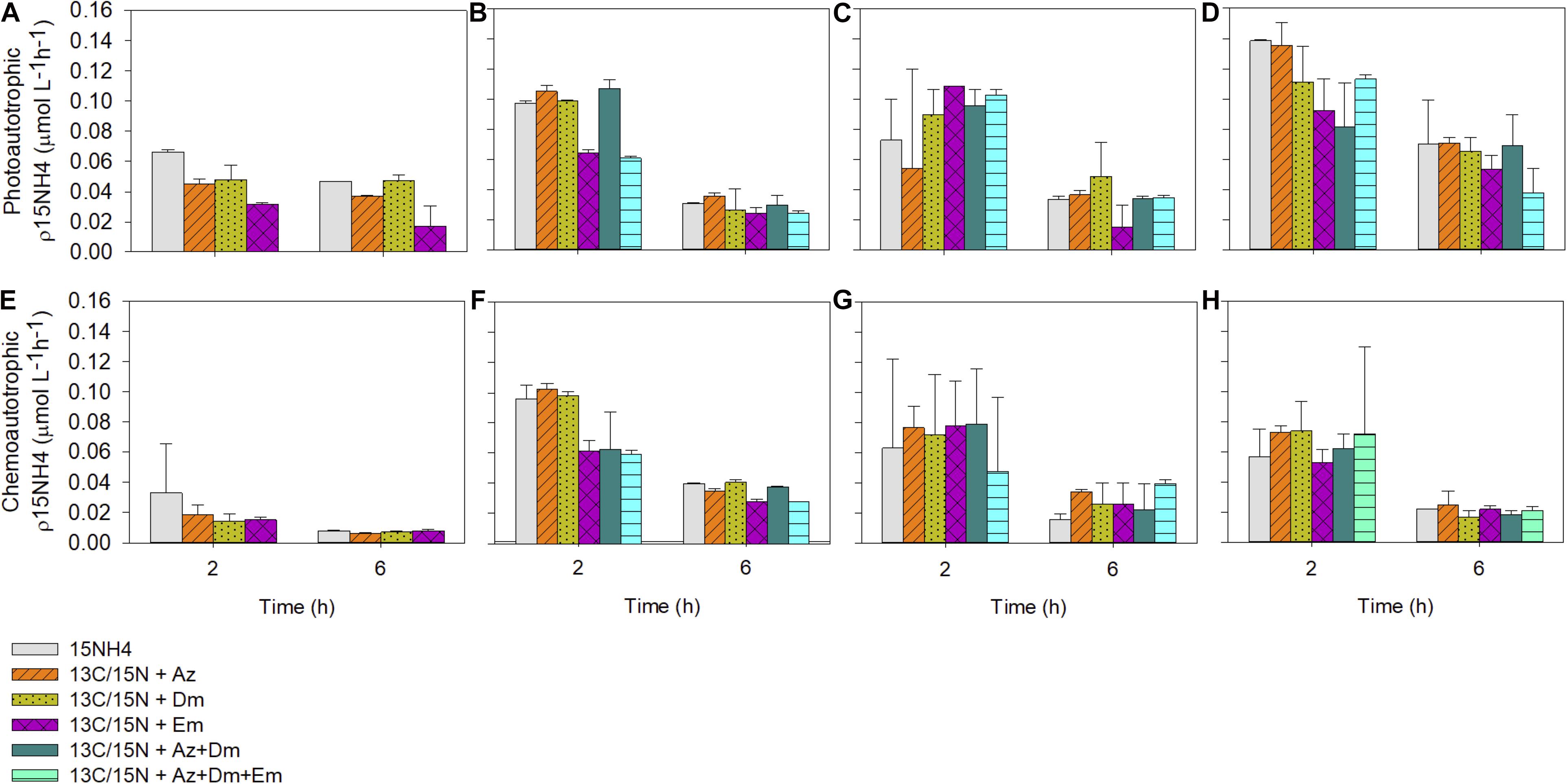
Figure 7. Photoautotrophic and chemoautotrophic ammonium uptake rates (μmol L–1h–1) during on-deck experiments under light and dark conditions. (A,E) Winter 2015; (B,F) Spring 2015; (C,G) Summer 2016; (D,H) Autumn 2016.
Results from a two-way ANOVA applied to light condition experiments showed significant differences in all the experiments between the treatments with pesticides and/or the time (Table 4). In winter 2015, the treatment 15NH4 + Az and 15NH4 + Dm were significantly different from the control (15NH4). The 15NH4 + Az was 32% lower after 2 h of incubation and dropped by 21% after 6 h of incubation compared to their control values (p = 0.001). The treatment 15NH4 + Dm was also a 53 and 64% lower after 2 and 6 h of incubation, respectively (p = 0.001). Photoautotrophic ρNH4+ values were significantly lower at the end of the incubation (6 h) compared to the first 2 h of incubation for control and treatments with pesticides (15NH4:30%; 15NH4 + Az:16%; 15NH4 + Em:47%; 15NH4 + Dm: 6% lower at 6 h of incubation; Table 4). In spring 2015, the treatment 15NH4 + Em and the combined pesticides treatment (15NH4 + Az + Dm + Em) were significantly different from their controls (p = 0.001), showing a decrease in ammonium assimilation rates. The 15NH4 + Em treatment rates were 77 and 23% lower than their control after 2 and 6 h of incubation, respectively. The combined treatment (15NH4 + Az + Dm + Em) was 39 and 23% lower compared to their controls at 2 and 6 h, respectively. Photoautotrophic ammonium uptake rates during spring 2015 were significantly lower (p = 0.001) after 6 h of incubation (69% average) in control and in the treatments amended with 15NH4 + Az, 15NH4 + Bm, 15NH4 + Em, 15NH4 + Az + Dm, 15NH4 + Az + Dm + Em. Light treatments during summer 2016 did not show significant differences between the treatments amended with pesticides and controls (p = 0.469) but values over time were significantly different from each other. After 6 h of incubation the light treatments were in average 58% lower than values obtained during the first 2 h of incubation. The same result was found in autumn 2016 when treatments did not show significant differences with pesticides compared to the controls but assimilation rates were significantly lower (average 59%) at 6 h compared to the first 2 h of incubation.
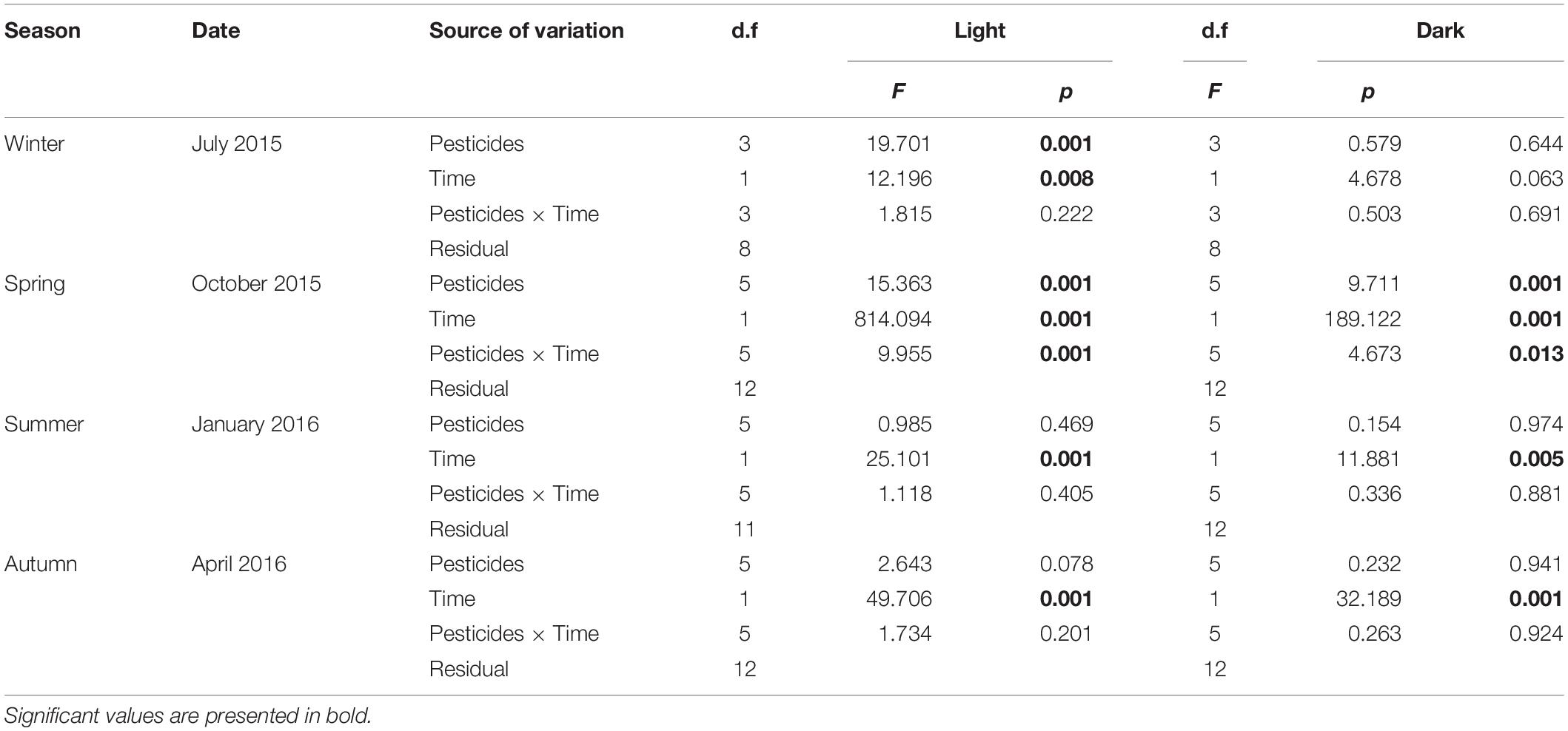
Table 4. Results from two-way ANOVA examining the effect of pesticides and the incubation time on N uptake rates obtained during on-deck experiments under light and dark conditions at Llico Bay.
A two-way ANOVA for dark incubation (Table 4) showed that ammonium uptake in spring 2015 was 36 and 30% lower in the treatments 15NH4 + Em and 15NH4 + Az + Dm + Em compared to the control (15NH4) at 2 and 6 h of incubation, respectively. The combined treatment 15NH4 + Az + Dm + Em was a 42 and 25% lower that the control after 2 and 6 h of incubation (p = 0.001). These differences were also found between time (p = 0.001) and also the interaction of the pesticides and time (p = 0.013). In average, dark ammonium utilization was 56% lower after 6 h of incubation that the first 2 h in spring 2015. In winter 2015, did not found significant differences in the chemoautotrophic ammonium assimilation between control and the treatments amended with pesticides (p = 0.6) and between the incubation times (p = 0.06). On-deck experiments in summer and autumn 2016 in dark conditions did not show differences between the treatments with pesticides and its controls (p = 0.97 and p = 0.94, respectively). However, values were significantly higher at 2 compared to 6 h of incubation by 59% in summer and by 69% in autumn 2016.
Discussion
Studies oriented specifically on the impacts of anti-lice pesticides to microbial communities involved in primary production are quite limited (Knapp et al., 2005; Burridge et al., 2010; Rain-Franco et al., 2018). Our results complement a first report of the effects of pesticides on rates of carbon assimilation (Rain-Franco et al., 2018) for the same study area, and as part of the same experiment. Overall the results provide new evidence that anti-lice pesticides can affect fundamental processes of marine ecosystem functioning.
Here we report ammonium uptake rates at Llico Bay and Caucahue Channel. In central-southern Chile, ammonium utilization in spring-summer season is high (0.23 ± 0.4 μmol L–1d–1 on average; Fernandez and Farías, 2012). At Llico Bay, ammonium was always present in subsurface waters at relatively high concentrations (Figures 2A,E,I), and the rates of ammonium assimilation observed in subsurface waters suggest that the entire surface pool of ammonium cannot be taken within 24 h. This pattern, was also observed in Caucahue Channel, except at control station (Qc), outside of the influence of farms, in which the ammonium concentration was very lower in surface waters, suggesting that the ammonium pool could be used within 24 h. Inside the channel is expected found higher accumulation of organic matter due to the presence of salmon farms and consequently higher remineralization rates that can explain the higher ammonium concentration found in the channel (Olsen et al., 2014; e.g., Q2) and this is contrasted with the Qc station in which concentration was nearly zero.
Ammonium is highly bioavailable and is often the preferred N compound assimilated by bacterioplankton and phytoplankton (Glibert et al., 2016). During dark conditions, ammonium uptake was higher than the photoautotrophic uptake, being up to 10 times higher at station Q9 in Caucahue channel (summer 2015). This was also found during on-deck experiments in Llico, in which the chemoautotrophic ammonium uptake was equivalent to 100% of the photoautotrophic uptake in spring 2015. Dark N uptake has been reported that can be anywhere from 10 to 100% of the uptake rate measured in the light, and this varies with physiological conditions of the populations and the species (Mulholland and Lomas, 2008). Nevertheless, these results suggest a potentially important contribution to regenerated production in these areas, affecting surface N dynamics, modifying the balance between new and regenerated production and consequently the downward flux of N.
The impact of pesticide in ammonium uptake was highly variable between the different pesticides amendments, significant effects were detected for a single pesticide (azamethiphos) during in situ incubations and for combined pesticides (azamethiphos-deltamethrin-emamectin benzoate) during on-deck experiments. The response of ammonium utilization to the addition of azamethiphos revealed an inhibition of the photoautotrophic uptake at station control (Qc) in Caucahue Channel in summer 2015. These finding are in contrast to the response obtained in C uptake, in which the addition of azamethiphos trigger an increase in the phototrophic C utilization (Rain-Franco et al., 2018), and also, are in contrast with the findings reported by Burridge et al. (2010) that suggest neutral effects on primary production, since no changes in dissolved oxygen and chlorophyll-a concentrations were observed in field studies in Canada.
Azamethiphos is an organophosphate insecticide highly soluble in water (Canty et al., 2007; Burridge et al., 2010). A very recent study, showed higher microbial degradation capacity of the pesticide azamethiphos nearly to our study area in Caucahue channel, suggesting that microorganism can use organophosphate pesticides as a source of C and phosphorus for their metabolism (Garcés et al., 2020). Periods of N and phosphorus limitation has been reported for the inner Sea of Chiloé (Iriarte et al., 2007; Olsen et al., 2014, 2017), in our results, we observed a higher increase in ammonium assimilation in surface and deeper waters at station Q9 (summer) in Caucahue Channel. This station showed a lower concentration of N and phosphorus (Figure 3) and a high bacterioplankton abundance, so the addition of this organic phosphorus compounds should be used as a nutrients source. This also can occur in the limit of the photic zone in which the competition of nutrients increases, and pesticides, such as azamethiphos, can be used by the microbial communities. Such increases in phototrophic and also chemoautotrophic ammonium uptake were seen during winter at stations Q5, and Q2 (Figure 5), and at station Bll3 in Llico Bay (Figure 4).
On the other hand, emamectin benzoate produces a decrease in photo and chemoautotrophic ammonium utilization between a 22 and 77% as we observed in on-deck experiments at Llico bay. The emamectin benzoate is a semi-synthetic derivative of a chemical produced by the bacterium, Streptomyces avermitilis (Bravo et al., 2008). The effects of this pesticide on non-target organism has been reported by a number of authors using, however, a few species (e.g., mussels, fishes, american lobster and copepods; Willis and Ling, 2003; Burridge et al., 2010, 2004). Our results are in accordance with the results found in C uptake, suggesting that emamectin benzoate can potentially act as a depressor of C fixation (Rain-Franco et al., 2018) and now also for ammonium assimilation. The inhibition in ρNH4+ values was also found in the combination of the three pesticides treatment (15NH4 + Az + Dm + Em) in autumn. However, this effect was not detected in the combined treatment with azamethiphos and deltamethrin (15NH4 + Az + Dm), suggesting that the inhibitory effect was related to the addition of emamectin benzoate. Previous studies suggest that toxicity can be increased using a combination of compounds compared to the use of single chemical (Laetz et al., 2009), but in our case the higher inhibitory response was found with a single addition of emamectin benzoate and not after a combined addition.
There are no studies dealing directly with deltamethrin and microorganism. From our data, deltamethrin produce an inhibitory or toxic effect during winter 2015 reducing up to 64% the ammonium assimilation after 6 h of incubation. The toxic effects have been also observed in in amphibians, crustacean and mollusk (Bhanu et al., 2011; Ernst et al., 2014). However, due to the high variability found in our results further studies are needed.
The interaction between the salmon cages, local hydrodynamics and local hydrographic characteristic are important factors to evaluate the dispersal propagation and the impact on non-target species of pesticides in the aquatic environment. Dispersion and dilution rates of the pesticides depend on the chemical nature (e.g., solubility, adsorption capacity, and chemistry composition) and the hydrographic characteristics of the area. A residual chemical compound concentration can persists in the area which is likely to impact non-target species (Nash, 2003). Uneaten food is point sources of emamectin benzoate to the surrounding ambient, and it is estimated that approximately 5–15% of the administered food is uneaten (Chen et al., 1999). Bloodworth et al. (2019) demonstrated a widespread detection of emamectin benzoate in sediments around Scottish fish farms. As well, Herrera et al. (2018) through a modeling methodology to assess the hydrodynamic effects of a salmon farm in a Patagonia channel, found that a considerable part of the enclosed volume within the cages of a fish farms remains circulating in the area of channels in Patagonia, suggesting an impact in the dispersal of pesticides in the area. The topical bath treatment method applied for azamethiphos will most likely result in rapid dilution of active ingredients. Azamethiphos has been detected up to 1000 m away from the location treatment (Ernst et al., 2014). The dispersion and dilution rates of the different anti-lice treatments utilized in this study can explain the high variability found in our results through the different stations and depths sampled, and this variability was also found in C assimilation (Rain-Franco et al., 2018). These results suggest a strong impact of the choice of pesticide used, the seasonal variability of the study area and the distance of salmon farming on the impact on microbial community and consequently on local primary productivity.
The impact of pesticides can be either positive by enhancing ammonium uptake or could be inhibitory for microorganism but either way it will impact the natural N budget and this impact was not only detected in photoautotrophic ammonium uptake; pesticides can also affect the chemoautotrophic communities, and potentially can modulate the regenerated primary production and therefore the balance between new and regenerated production and organic matter export. Taken together, our findings and those of Rain-Franco et al. (2018) show significant alterations in C and N assimilation.
In conclusion, the use of anti-lice pesticides can produce changes in photo and chemoautotrophic ammonium uptake in central-southern and northern Patagonia in Chile. Azamethiphos act as depressor of ammonium assimilation but also can be used by phyto- and bacterioplankton under nutrient limitation conditions. Emamectin benzoate can act as a strong depressor as well for deltamethrin and these changes were strongest if a single pesticide is applied as opposed to a combination of two or more pesticides. We conclude that pesticides in marine waters have the potential for altering local primary production fluxes and therefore can impact biological ecosystem productivity.
Data Availability Statement
The original contributions presented in the study are included in the article/Supplementary Material, further inquiries can be directed to the corresponding author.
Author Contributions
CF designed the experiments and lead the research project, carried out the sample collection, and experiments. VV-C analyzed the data and participated in data interpretation. VV-C and CF wrote the manuscript. Both authors contributed to the article and approved the submitted version.
Funding
This work was supported by FONDAP grants 15110027 and 1515003, FONDECYT project 1180954, and by COPAS Sur-Austral ANID PIA APOYO CCTE AFB170006. This study was development in the frame of the French-Chilean project LIA-MORFUN and the current LIA-MAST project (CNRS).
Conflict of Interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Acknowledgments
The authors are grateful to Claudia Muñoz for assistance during field experiments.
Supplementary Material
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fmars.2020.602002/full#supplementary-material
Footnotes
References
Aminot, A., and Kérouel, R. (2007). Dossage Automatique Des Nutriments Dans Les Eaux Marins. France: Editions Quae.
Avendaño-Herrera, R. (2018). Proper antibiotics use in the Chilean salmon industry: policy and technology bottlenecks. Aquaculture 495, 803–805. doi: 10.1016/j.aquaculture.2018.06.072
Baskett, M. L., Burgess, S. C., and Waples, R. S. (2013). Assessing strategies to minimize unintended fitness consequences of aquaculture on wild populations. Evol. Appl. 6, 1090–1108. doi: 10.1111/eva.12089
Bhanu, S., Archana, S., Ajay, K., Singh, S., and Vandana, B. (2011). Impact of deltamethrin on environment, use as an insecticide and its bacterial degradation – a preliminary study. Int. J. Environ. Sci. 1, 977–985.
Bloodworth, J. W., Baptie, M. C., Preedy, K. F., and Best, J. (2019). Negative effects of the sea lice therapeutant emamectin benzoate at low concentrations on benthic communities around Scottish fish farms. Sci. Total Environ. 669, 91–102. doi: 10.1016/j.scitotenv.2019.02.430
Bravo, S., Sevatdal, S., and Horsberg, T. E. (2008). Sensitivity assessment of Caligus rogercresseyi to emamectin benzoate in Chile. Aquaculture 282, 7–12. doi: 10.1016/j.aquaculture.2008.06.011
Burridge, L., Weis, J. S., Cabello, F., Pizarro, J., and Bostick, K. (2010). Chemical use in salmon aquaculture: a review of current practices and possible environmental effects. Aquaculture 306, 7–23. doi: 10.1016/j.aquaculture.2010.05.020
Burridge, L. E., Hamilton, N., Waddy, S. L., Haya, K., Mercer, S. M., Greenhalgh, R., et al. (2004). Acute toxicity of emamectin benzoate (SLICEtm) in fish feed to American lobster, Homarus americanus. Aquac. Res. 35, 713–722. doi: 10.1111/j.1365-2109.2004.01093.x
Canty, M. N., Hagger, J. A., Moore, R. T. B., Cooper, L., and Galloway, T. S. (2007). Sublethal impact of short term exposure to the organophosphate pesticide azamethiphos in the marine mollusc Mytilus edulis. Mar. Pollut. Bull. 54, 396–402. doi: 10.1016/j.marpolbul.2006.11.013
Chen, Y.-S., Beveridge, M. C. M., and Telfer, T. C. (1999). Physical characteristics of commercial pelleted Atlantic salmon feeds and consideration of implications for modeling of waste dispersion through sedimentation. Aquacult. Int. 7, 89–100.
Davies, I. (2001). Targeted environmental monitoring for the effects of medicines used to treat sea-lice infestation on farmed fish. ICES J. Mar. Sci. 58, 477–485. doi: 10.1006/jmsc.2000.1040
Dresdner, J., Chávez, C., Quiroga, M., Jiménez, D., Artacho, P., and Tello, A. (2019). Impact of Caligus treatments on unit costs of heterogeneous salmon farms in Chile. Aquac. Econ. Manag. 23, 1–27. doi: 10.1080/13657305.2018.1449271
Ernst, W., Doe, K., Cook, A., Burridge, L., Lalonde, B., Jackman, P., et al. (2014). Dispersion and toxicity to non-target crustaceans of azamethiphos and deltamethrin after sea lice treatments on farmed salmon, Salmo salar. Aquaculture 424–425, 104–112. doi: 10.1016/j.aquaculture.2013.12.017
Fernandez, C., and Farías, L. (2012). Assimilation and regeneration of inorganic nitrogen in a coastal upwelling system: ammonium and nitrate utilization. Mar. Ecol. Prog. Ser. 451, 1–14. doi: 10.3354/meps09683
Fernández, C., Farías, L., and Alcaman, M. E. (2009). Primary production and nitrogen regeneration processes in surface waters of the Peruvian upwelling system. Prog. Oceanogr. 83, 159–168. doi: 10.1016/j.pocean.2009.07.010
Garcés, D. V., Fuentes, M. E., and Quiñones, R. A. (2020). Effect of Azamethiphos on enzymatic activity and metabolic fingerprints of marine microbial communities from the water column. Aquaculture 529:735650. doi: 10.1016/j.aquaculture.2020.735650
Gebauer, P., Paschke, K., Vera, C., Toro, J. E., Pardo, M., and Urbina, M. (2017). Lethal and sub-lethal effects of commonly used anti-sea lice formulations on non-target crab Metacarcinus edwardsii larvae. Chemosphere 185, 1019–1029. doi: 10.1016/j.chemosphere.2017.07.108
Glibert, P. M., Wilkerson, F. P., Dugdale, R. C., Raven, J. A., Dupont, C. L., Leavitt, P. R., et al. (2016). Pluses and minuses of ammonium and nitrate uptake and assimilation by phytoplankton and implications for productivity and community composition, with emphasis on nitrogen-enriched conditions: pluses and minuses of NH4+ and NO3-. Limnol. Oceanogr. 61, 165–197. doi: 10.1002/lno.10203
González, L., and Carvajal, J. (2003). Life cycle of Caligus rogercresseyi, (Copepoda: Caligidae) parasite of Chilean reared salmonids. Aquaculture 220, 101–117. doi: 10.1016/s0044-8486(02)00512-4
González, M. P., Marín, S. L., and Vargas-Chacoff, L. (2015). Effects of Caligus rogercresseyi (Boxshall and Bravo, 2000) infestation on physiological response of host Salmo salar (Linnaeus 1758): establishing physiological thresholds. Aquaculture 438, 47–54. doi: 10.1016/j.aquaculture.2014.12.039
Hargrave, B. (2010). Empirical relationships describing benthic impacts of salmon aquaculture. Aquac. Environ. Interact. 1, 33–46. doi: 10.3354/aei00005
Helgesen, K. O., Bravo, S., Sevatdal, S., Mendoza, J., and Horsberg, T. E. (2014). Deltamethrin resistance in the sea louse Caligus rogercresseyi (Boxhall and Bravo) in Chile: bioassay results and usage data for antiparasitic agents with references to Norwegian conditions. J. Fish Dis. 37, 877–890. doi: 10.1111/jfd.12223
Herrera, J., Cornejo, P., Sepúlveda, H. H., Artal, O., and Quiñones, R. A. (2018). A novel approach to assess the hydrodynamic effects of a salmon farm in a Patagonian channel: coupling between regional ocean modeling and high resolution les simulation. Aquaculture 495, 115–129. doi: 10.1016/j.aquaculture.2018.05.003
Holm-Hansen, O., Lorenzen, C. J., Holmes, R. W., and Strickland, J. D. H. (1965). Fluorometric determination of chlorophyll. ICES J. Mar. Sci. 30, 3–15. doi: 10.1093/icesjms/30.1.3
Holmes, R., Aminot, A., Kérouel, R., Hooker, B., and Peterson, B. (1999). A simple and precise method for measuring ammonium in marine and freshwater ecosystems. Can. J. Fish. Aquat. Sci. 56, 1801–1808. doi: 10.1139/f99-128
Iriarte, J. L., González, H. E., Liu, K. K., Rivas, C., and Valenzuela, C. (2007). Spatial and temporal variability of chlorophyll and primary productivity in surface waters of southern Chile (41.5–43° S). Estuar. Coast. Shelf Sci. 74, 471–480. doi: 10.1016/j.ecss.2007.05.015
Johnson, S. C., Treasurer, J. W., Bravo, S., and Nagasawa, K. (2004). A review of the impact of parasitic copepods on marine aquaculture. Zool. Stud. 43, 8–19.
Knapp, C. W., Caquet, T., Hanson, M. L., Lagadic, L., and Graham, D. W. (2005). Response of water column microbial communities to sudden exposure to deltamethrin in aquatic mesocosms. FEMS Microbiol. Ecol. 54, 157–165. doi: 10.1016/j.femsec.2005.03.004
Laetz, C. A., Baldwin, D. H., Collier, T. K., Hebert, V., Stark, J. D., and Scholz, N. L. (2009). The synergistic toxicity of pesticide mixtures: implications for risk assessment and the conservation of endangered Pacific Salmon. Environ. Health Perspect. 117, 348–353. doi: 10.1289/ehp.0800096
Marie, D., Partensky, F., Jacquet, S., and Vaulot, D. (1997). Enumeration and cell cycle analysis of natural populations of marine picoplankton by flow cytometry using the nucleic acid stain SYBR Green I. Appl. Environ. Microbiol. 63, 186–193. doi: 10.1128/aem.63.1.186-193.1997
Millanao, B. A., Barrientos, H. M., Gómez, C. C., Tomova, A., Buschmann, A., Dölz, H., et al. (2011). Uso inadecuado y excesivo de antibióticos: salud pública y salmonicultura en Chile. Rev. Médica Chile 139, 107–118. doi: 10.4067/s0034-98872011000100015
Mulholland, M. R., and Lomas, M. W. (2008). “Chapter 7 - nitrogen uptake and assimilation,” in Nitrogen in the Marine Environment, 2nd Edn, eds D. G. Capone, D. A. Bronk, M. R. Mulholland, and E. J. Carpenter (San Diego: Academic Press), 303–384. doi: 10.1016/b978-0-12-372522-6.00007-4
Nash, C. E. (2003). Interactions of Atlantic salmon in the Pacific Northwest. Fish. Res. 62, 339–347. doi: 10.1016/s0165-7836(03)00068-7
Núñez-Acuña, G., Gonçalves, A. T., Valenzuela-Muñoz, V., Pino-Marambio, J., Wadsworth, S., and Gallardo-Escárate, C. (2015). Transcriptome immunomodulation of in-feed additives in Atlantic salmon Salmo salar infested with sea lice Caligus rogercresseyi. Fish Shellfish Immunol. 47, 450–460. doi: 10.1016/j.fsi.2015.09.009
Olsen, L., Hernández, K., Van Ardelan, M., Iriarte, J., Can Bizsel, K., and Olsen, Y. (2017). Responses in bacterial community structure to waste nutrients from aquaculture: an in situ microcosm experiment in a Chilean fjord. Aquac. Environ. Interact. 9:2017.
Olsen, L., Hernández, K., Van Ardelan, M., Iriarte, J., Sánchez, N., González, H., et al. (2014). Responses in the microbial food web to increased rates of nutrient supply in a southern Chilean fjord: possible implications of cage aquaculture. Aquac. Environ. Interact. 6, 11–27. doi: 10.3354/aei00114
Quiñones, R. A., Fuentes, M., Montes, R. M., Soto, D., and León-Muñoz, J. (2019). Environmental issues in Chilean salmon farming: a review. Rev. Aquac. 11, 375–402. doi: 10.1111/raq.12337
Rain-Franco, A., Rojas, C., and Fernandez, C. (2018). Potential effect of pesticides currently used in salmon farming on photo and chemoautotrophic carbon uptake in central – southern Chile. Aquaculture 486, 271–284. doi: 10.1016/j.aquaculture.2017.12.048
Reyes, X., and Bravo, S. (1983). Nota sobre una copepoidosis en salmones de cultivo. Invest. Mar. 11, 55–57.
Rozas, M., and Asencio, G. (2007). Evaluación de la situación epidemiológica de la caligiasis en chile: hacia una estrategia de control efectiva. Salmociencia 2, 43–59. doi: 10.1787/9789264284272-4-es
Siwicki, A. K., Terech-Majewska, E., Grudniewska, J., Malaczewska, J., Kazun, K., and Lepa, A. (2010). Influence of deltamethrin on nonspecific cellular and humoral defense mechanisms in rainbow trout (Oncorhynchus mykiss). Environ. Toxicol. Chem. 29, 489–491. doi: 10.1002/etc.75
Keywords: ammonium uptake, salmon farming, azamethiphos, deltamethrin, emamectin benzoate, Chile
Citation: Valdés-Castro V and Fernandez C (2021) Effect of Three Pesticides Used in Salmon Farming on Ammonium Uptake in Central-Southern and Northern Patagonia, Chile. Front. Mar. Sci. 7:602002. doi: 10.3389/fmars.2020.602002
Received: 09 October 2020; Accepted: 30 December 2020;
Published: 25 January 2021.
Edited by:
Jose Luis Iriarte, Austral University of Chile, ChileReviewed by:
Lasse Mork Olsen, University of Bergen, NorwayAlexander Godfrey Murray, Marine Scotland, United Kingdom
Copyright © 2021 Valdés-Castro and Fernandez. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Camila Fernandez, fernandez@obs-banyuls.fr