 Kai Zhu
Kai Zhu Dongming Lin3,4,5,6
Dongming Lin3,4,5,6- 1Key Laboratory of Sustainable Utilisation of Technology Research for Fishery Resources, Zhejiang Marine Fisheries Research Institute, Zhoushan, China
- 2Scientific Observing and Experimental Station of Fishery Resources for Key Fishing Grounds, Ministry of Agriculture and Rural Affairs, Zhoushan, China
- 3College of Marine Sciences, Shanghai Ocean University, Shanghai, China
- 4The Key Laboratory of Sustainable Exploitation of Oceanic Fisheries Resources, Ministry of Education, Shanghai Ocean University, Shanghai, China
- 5Collaborative Innovation Center for National Distant-water Fisheries, Shanghai Ocean University, Shanghai, China
- 6National Engineering Research Center for Oceanic Fisheries, Shanghai Ocean University, Shanghai, China
Stock structure information is an important part of bases for understanding the dynamics of cephalopod populations. Purpleback flying squid Sthenoteuthis oualaniensis is an abundant and highly productive species in tropical and subtropical waters of the Indo-Pacific region. However, it is characterized by a complex stock structure, and the stock discrimination is an urgent priority to understand basic biology and for stock assessment and management purposes. Here, we used gladius morphology to identify and discriminate the dwarf without a dorsal photophore and middle-sized typical forms of S. oualaniensis in the South China Sea. Results showed that both forms had double axes on the gladius and females consistently had a larger gladius than males. Multivariate analyses using the gladius morphometric characteristics indicated that individuals of dwarf form without a dorsal photophore were distinguished from those of middle-sized typical form, which was evidenced by the obvious groups and significant dissimilarity of gladii of these two forms. The discrimination based on the gladius morphometric characteristics showed a high rate of accuracy, in which a global discrimination accuracy was estimated to be 92.36% for both forms without regarding sexes and 65.82% for the discrimination to form and sex. In combination, these lines of evidence indicated that individuals of dwarf without a dorsal photophore and middle-sized typical forms of S. oualaniensis can be accurately distinguished using the gladius morphometric characteristics, and these results will warrant the application of the gladius to study the stock structure of S. oualaniensis and other squid else.
Introduction
Purpleback flying squid Sthenoteuthis oualaniensis is an abundant and highly productive species that inhabits open waters of the Indo-Pacific region, including tropical and subtropical waters (Jereb and Roper, 2010). This species is characterized by fast growth, short lifespan, and semelparous reproduction (Liu et al., 2016). S. oualaniensis plays a critical role in epipelagic to mesopelagic waters, preying on a wide spectrum of food organisms from mesozooplankton to myctophids and supporting diverse marine predators including squids, fishes, sharks, whales, and seabirds (Jereb and Roper, 2010). S. oualaniensis is also becoming an important fishery species (Zhang et al., 2014). In the central South China Sea (SCS), for example, S. oualaniensis is a major target for the small-scale jigging fishery (Chen et al., 2008) as well as for the large-scale light falling-net fishery (Yang, 2002; Zhang et al., 2013). Similar to other Ommastrephidae squid such as Dosidicus gigas and Illex argentinus (Keyl et al., 2011; Rodhouse et al., 2013), however, stock discrimination is still uncertain and an important research priority (Chen et al., 2012), particularly for the case of increasing fishing pressure in the SCS (Zhang et al., 2014).
Stock structure information provides a basis for understanding the dynamics of cephalopod populations. Each stock may have unique demographic properties and responses to exploitation (Boyle and Rodhouse, 2005). Morphometric characteristics form the basis for one of the simplest, and most often used, tools to identify and characterize squid stocks, i.e., in determining population assemblages (Rodhouse et al., 2013) and assigning individuals to stock (Fang et al., 2014). Morphometric characteristics can also be used to identify traits with evolutionary significance (e.g., Wanninger and Wollesen, 2018). Therefore, information on how, and the extent to which, a species or one stock within a species evolves specific morphometric relationships can potentially contribute to better management and conservation (Clavel and Morlon, 2017), as well as leading to a better understanding of species evolution, ecology, and ultimately stock assessment (Laptikhovsky et al., 2017; Wright et al., 2018).
S. oualaniensis exhibits a complicated intraspecific stock structure, with multiple morphological (Nesis, 1993), geographic (Chen et al., 2007; Yan et al., 2015), molecular (Staaf et al., 2010), and/or even spawning forms (Liu et al., 2008). In the SCS, available information indicates that there are probably two stocks of purpleback flying squid—the middle-sized (with dorsal photophore) and dwarf (without dorsal photophore) forms (Zhu et al., 2016; Wang et al., 2017; Li et al., 2019). These two stocks are, however, unconfirmed, presumably owing to a significant geographic overlap in morphology (Li et al., 2019). According to the different types of gladius, middle-sized individuals are divided into two forms: the middle-sized, typical form with double axes and the middle-sized not typical form with single axes (Jereb and Roper, 2010). Hard structures such as gladius, statoliths, and beaks are increasingly applied for stock discrimination of cephalopods (e.g., Liu et al., 2008; Fang et al., 2014; Fan et al., 2015) and show greater promise than the mantle and other body organs (Zhu et al., 2016; Wang et al., 2019). The gladius grows continuously throughout the lifetime of the species. The morphological characteristics and chemical composition of the gladius among species are obviously different, which is an important basis for the classification of cephalopods (Liu and Chen, 2010; Chen and Qiu, 2014). Little attention has been paid to morphological variations in gladius between different forms of the same species; it still needs relative information for classification (Gong et al., 2018).
Here, we used “form” to refer to different stocks of S. oualaniensis, following Jereb and Roper (2010). We analyzed the morphometric characteristics of the gladius of the middle-sized form (with dorsal photophore) and dwarf without a dorsal photophore form of S. oualaniensis and used multivariate statistics to analyze the differences in morphometric characteristics of the gladius between the two forms, with the ultimate aim of acquiring novel insights into stock identification. Specifically, we examined the following questions: 1) how different are the gladius morphologic characteristics between forms and sexes of S. oualaniensis? and 2) can gladius morphologic characteristics be used to distinguish these forms and/or sexes? These results will help put forward our understanding of the structure of this species and also warrant consideration of using the work as a framework to be applied in other commercially exploited squid species.
Material and methods
Sample collection
S. oualaniensis was sampled at 20 stations in the SCS (9.80°N–17.25°N, 110.25°E–115.02°E) by the Chinese lighting falling-net vessel Gui Beiyu 61999, from May to June in 2017 and 2018 (Figure 1). Four hundred thirty-eight specimens were randomly caught and collected and immediately frozen (–18°C) on board for further laboratory analysis.
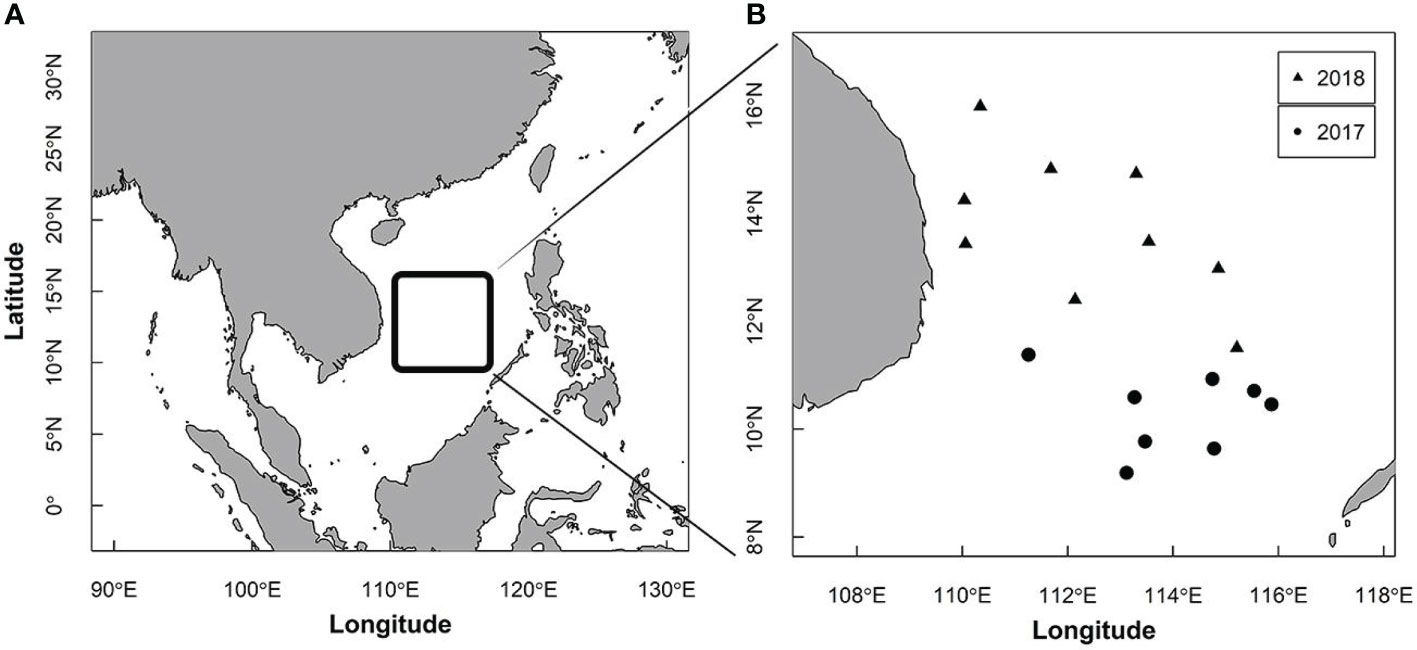
Figure 1 Study region (A) and stations (B) for Sthenoteuthis oualaniensis sampled in the South China Sea.
After being defrosted at room temperature in the laboratory, S. oualaniensis was categorized into the dwarf without a dorsal photophore and middle-sized forms based on the absence or presence of the dorsal photophore and the apparatus’ length (Zhu et al., 2016). The middle-sized forms were categorized into the typical and not typical forms based on the double axes or single axis of the gladius (Jereb and Roper, 2010). The dorsal mantle lengths (ML) were measured to the nearest 1 mm and body mass (BW) weighted to the nearest 1 g. Thereafter, a subsample of 275 specimens (130 females, 145 males) were used for gladius morphology analysis, whereas the gladius of other samples had been abandoned due to damage. The subsampled S. oualaniensis were dissected, sexed, and assigned a maturity stage on a macro scale following Lin (2015): I immature, II developing, III physiologically maturing, IV–V physiologically mature, VI functionally mature, VII spawning, and VIII spent.
Gladius morphometric measurement
The gladii of each subsampled S. oualaniensis were removed and measurements made of the following morphometric characteristics (Figure 2): total gladius length (GL), cone length (CL), greatest width of the cone (GWC), proostracum length (PL), length of the vane (VL), greatest width of the vane (GWV), the length of greatest width of the proostracum (GWPL), and greatest width of the proostracum (GWP). All these characteristics were measured accurately to be 0.01 mm.
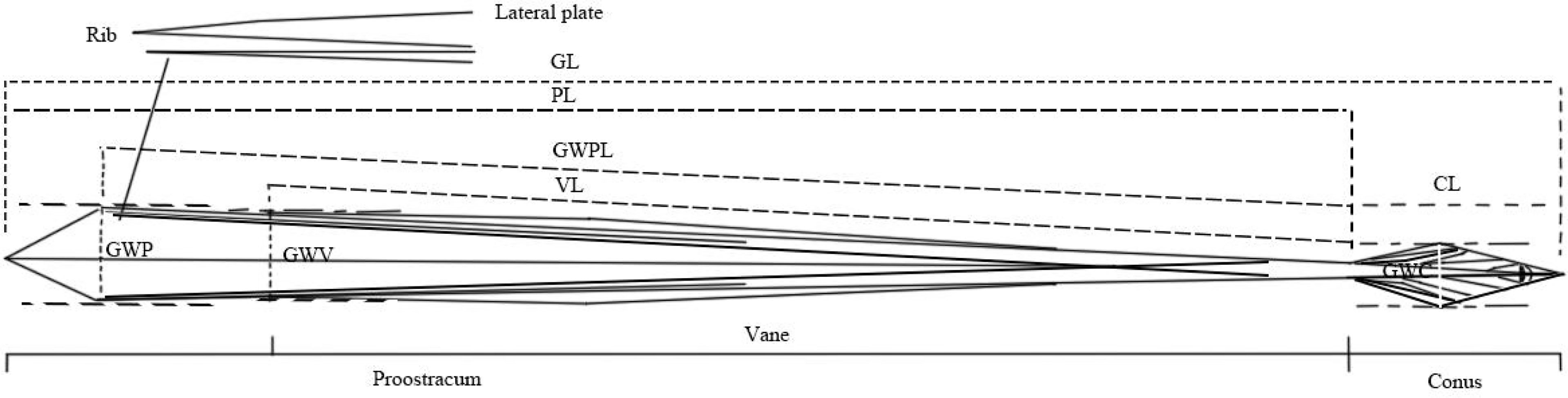
Figure 2 Scheme of gladius morphometric measurements for Sthenoteuthis oualaniensis. GL, total gladius length; CL, length of the cone; GWC, greatest width of the cone; PL, proostracum length; VL, length of the vane; GWV, greatest width of the vane; GWPL, length of the greatest width of the proostracum; GWP, greatest width of the proostracum.
Statistical analysis
Each morphometric measurement of the gladius was checked for normal distribution with the one-sample Kolmogorov–Smirnoff test as well as for homogeneity of the variances with the Levene’s test (Zar, 1999). One-way ANOVA was used to test the difference between different forms. When significant differences were achieved, a Tukey’s post-hoc test was applied to determine where the differences occurred (Zar, 1999). When either of normal distribution or homoscedasticity was not achieved, data were subjected to a Kruskal–Wallis non-parametric one-way ANOVA test and a Games–Howell post-hoc test was performed (Zar, 1999).
Non-metric multidimensional scaling (nMDS) analysis and analysis of similarity (ANOSIM) employing the Bray–Curtis dissimilarity measure were used to access the dissimilarities of gladii between forms. This allowed for the potential identification of individuals that belong to the dwarf form without a dorsal photophore or middle-sized form. Furthermore, a stepwise discriminant analysis was performed to identify the gladius morphological characteristics that significantly classified individuals from the dwarf without a dorsal photophore and middle-sized forms (Rencher, 2002). The leave-one-out cross-validation was used to determine the rate of correct classification for different groups. Prior to nMDS, ANOSIM, and stepwise discriminant analyses, each morphometric measurement of the gladius was standardized by dividing by mantle length to eliminate the possible effects of allometric growth of animals, and then square root transformation was performed to all the standardized gladius morphometrics (Zar, 1999). The standardized morphometric measurements are represented by a subscript “s”, i.e., GLs, CLs, GWCs, PLs, VLs, GWVs, GWPLs, and GWPs.
Statistical analyses were obtained using OriginPro version 2015 (OriginLab Corporation 2015) and R version 3.5.0 (R Core Team, 2018). In all statistical tests used, significant differences were considered only when P < 0.05.
Results
The dwarf form without a dorsal photophore of S. oualaniensis was 80–124-mm mantle length (ML, mean 88 ± 6 mm) for females and 76–95 mm (mean 78 ± 5 mm) for males. Individuals of the middle-sized form were larger with mantle lengths of 132–173 mm (mean 135 ± 15 mm) for females and 116–158 mm (mean 118 ± 12 mm) for males.
Gladius morphology of dwarf without a dorsal photophore and middle-sized forms
The gladius of both dwarf and middle-sized forms had a pair of double ribs at the edge of the medial plate of the proostracum (Figure 2); it was reasonable to expect that the individuals of middle-sized form were affiliated to the middle-sized typical form. The total gladius length (GL) of the dwarf-form individuals, however, was significantly shorter than that of the middle-sized typical-form individuals (Kruskal–Wallis test, χ2 = 224.41, P < 0.05) (Table 1). Similar findings were obtained with the analyses of other morphological characteristics, namely, the cone length (CL, Kruskal–Wallis test, χ2 = 199.45, P < 0.05), greatest width of the cone (GWC, Kruskal–Wallis test, χ2 = 213.13, P < 0.05), proostracum length (PL, Kruskal–Wallis test, χ2 = 223.17, P < 0.05), length of the vane (VL, Kruskal–Wallis test, χ2 = 222.27, P < 0.05), greatest width of the vane (GWV, Kruskal–Wallis test, χ2 = 220.13, P < 0.05), the length of greatest width of the proostracum (GWPL, Kruskal–Wallis test, χ2 = 223.27, P < 0.05), and greatest width of the proostracum (GWP, Kruskal–Wallis test, χ2 = 225.07, P < 0.05). The gladius for female individuals of dwarf form was consistently larger than that for males, and there were significant differences in the morphological characteristics of the gladius between males and females, except for cone length (CL) (Table 1). Similarly, females of the middle-sized typical form had a larger gladius than males. Consequently, each gladius characteristic differed significantly between females and males of the middle-sized typical form, with females being consistently larger (Table 1).
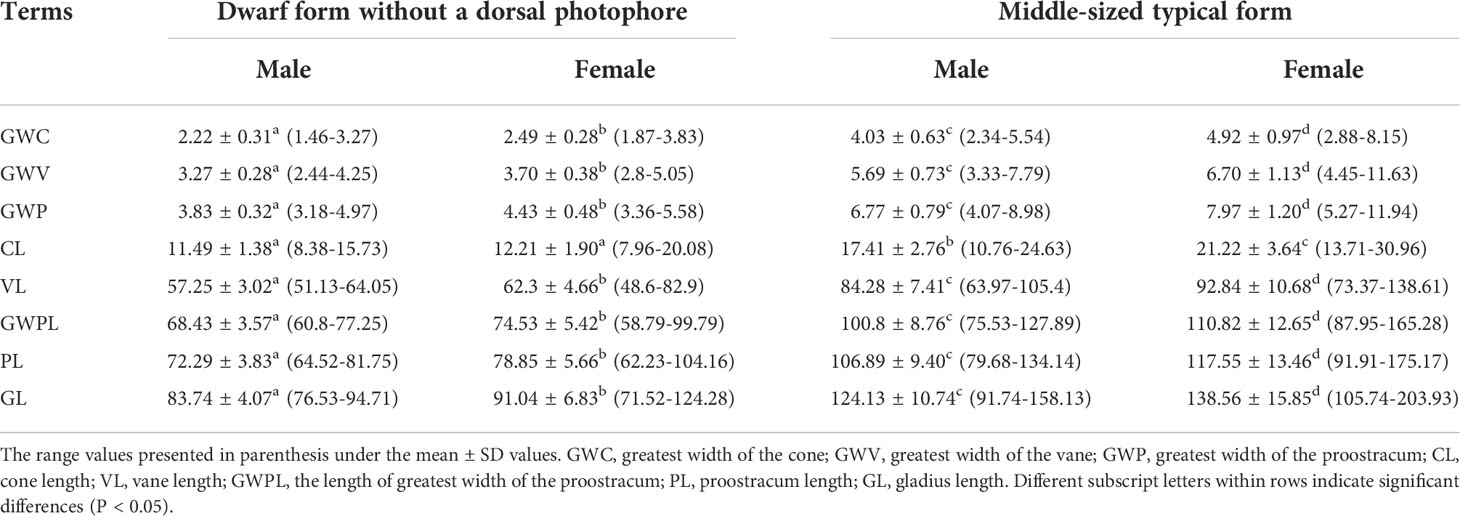
Table 1 Measurements of gladius morphometric characteristics (mean ± SD, mm) for dwarf without a dorsal photophore and middle-sized typical form Sthenoteuthis oualaniensis in the South China Sea.
Meanwhile, univariate statistics (ANOVA) showed that standardized gladius morphometric characteristics were significantly different between the dwarf and middle-sized typical forms (P <0.05), with the exception of GL/ML (P > 0.05). Within a single form, female and male individuals differed in seven of nine standardized gladius morphometric characteristics, dwarf (except GWC/ML, GWV/ML), and middle-sized typical forms (except GWV/ML, GWP/ML).
Dissimilarity analysis of the gladius between forms
In non-metric multidimensional scaling (nMDS) analysis, ordination of all graphs with stress values equal to or below 0.1 was considered fair, which could reflect the actual distribution characteristics (Figure 3). Regardless of sexes, nMDS analysis revealed that most of the gladii of dwarf without a dorsal photophore and middle-sized typical forms grouped together separately (Figure 3A) and the dissimilarity between them was significant, with an ANOSIM statistical R value of 0.24 (P = 0.001) (Table 2). For female and male individuals of both forms, the gladii were distinctly separate, with the exception of male dwarf and female middle-sized typical forms (Figures 3B–E). The difference between females and/or males of both forms was significant, and a higher dissimilarity value was obtained between female dwarf and female middle-sized typical forms (ANOSIM-R = 0.30) and between male dwarf and female middle-sized typical forms (ANOSIM-R = 0.37) (Table 2).
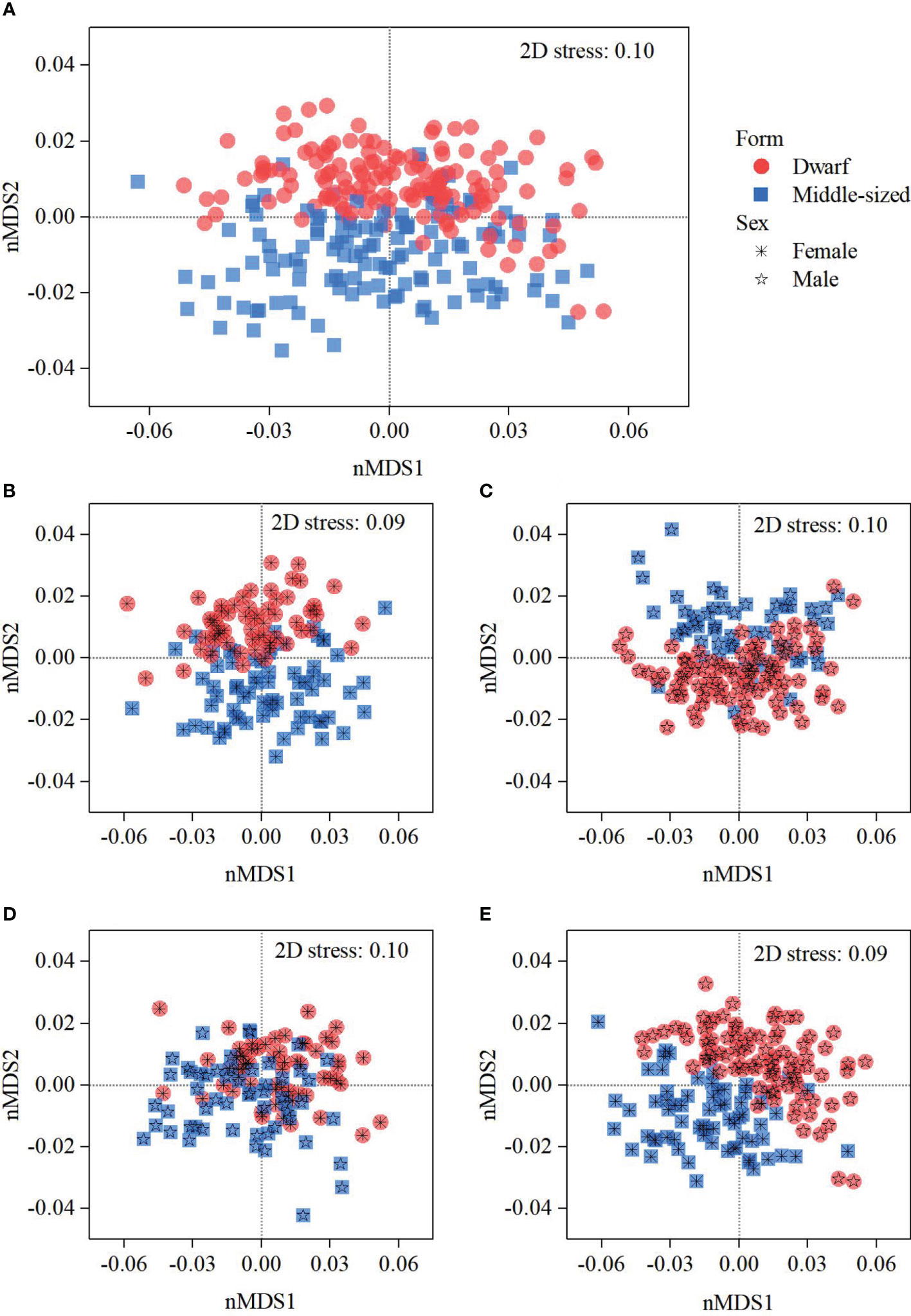
Figure 3 Non-metric multidimensional scaling (nMDS) ordination of gladius morphometric characteristics for dwarf without a dorsal photophore and middle-sized typical forms of Sthenoteuthis oualaniensis in the South China Sea. (A), nMDS plots for the analysis between forms without regarding sexes; (B) nMDS plots for the analysis between females of both forms; (C) nMDS plots for the analysis between males of both forms; (D) nMDS plots for the analysis between female dwarf without a dorsal photophore and male middle-sized typical forms; (E) nMDS plots for the analysis between male dwarf and female middle-sized typical forms.

Table 2 Results of analysis of similarities (ANOSIM) for gladius morphometric characteristics of the dwarf without a dorsal photophore and middle-sized typical forms Sthenoteuthis oualaniensis in the South China Sea.
Discriminant analysis of gladius morphology between forms
Stepwise discriminant analyses indicated that the difference in the gladius morphology between the dwarf without a dorsal photophore and middle-sized typical forms could be explained using four gladius morphometric characteristics, namely, GWCs, GWVs, GWPs, and GLs. Accordingly, the accuracy of the discriminant analyses was estimated to be 88.71% for the middle-sized typical form, 95.36% for the dwarf form, and the global accuracy and cross-validation accuracy were the same, being 92.36% (Table 3).
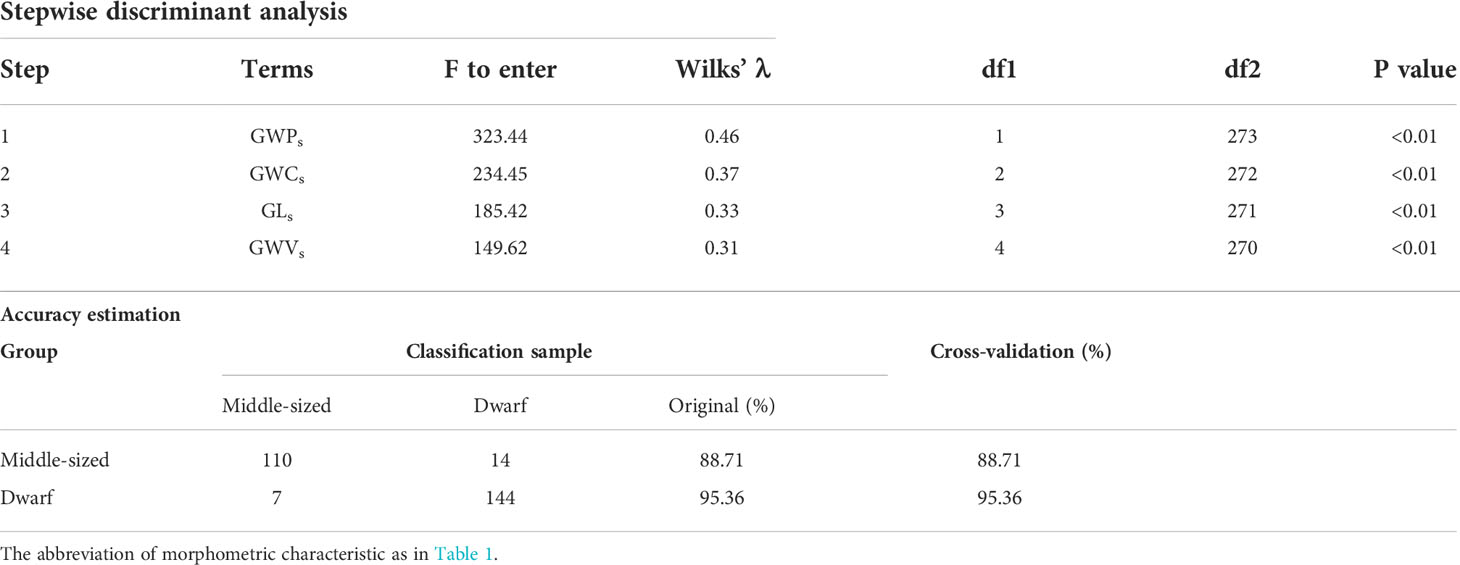
Table 3 The results of stepwise discriminant analyses for the gladius morphometric characteristics of the dwarf without a dorsal photophore and middle-sized typical forms Sthenoteuthis oualaniensis in the South China Sea.
With respect to sexes, stepwise discriminant analyses revealed that individuals from female middle-sized typical forms, male middle-sized typical forms, female dwarf forms, and male dwarf forms could be successfully discriminated by five gladius morphometric characteristics, namely, GWPs, PLs, GWCs, GWVs, and GLs. The accuracy of discrimination ranged from 58.18% to 74.44% for form and sex, respectively. The global accuracy and cross-validation accuracy were 65.82% and 63.64%, respectively.
Discussion
Gladius morphology
Because the gladius of cephalopods has the function of supporting its body, it grows continuously throughout the lifetime of the species. The morphological characteristics of the gladius among species are obviously different, which is an important basis for the classification of cephalopods. Our findings showed that the gladius had double axes at the edge of the medial plate of the proostracum for both dwarf forms without a dorsal photophore and middle-sized forms in the SCS. For the middle-sized form, this observation was in agreement with previous research that the feature of double axes on the gladius was typical for the middle-sized form purpleback flying squid (see Jereb and Roper, 2010). However, it was noteworthy that the gladius morphology of the middle-sized form appeared to vary with oceanic regions. For example, individuals of the middle-sized form in the Indian and Pacific Oceans had double axes on the gladius like those from the SCS, whereas the median form animals from the Red and Arabian Seas had a single axis on the gladius (Bizikov, 1995). The presence of double axes on the gladius was typical for middle-sized S. oualaniensis, which was the most abundant and widely distributed form within their distribution range (Jereb and Roper, 2010). In contrast, the middle-sized not typical forms with a single axis on the gladius are narrowly distributed in the Red and Arabian Seas and the Gulf of Aden (Jereb and Roper, 2010).
The gladius of middle-sized typical forms in the SCS was relatively smaller than those from the Indian Ocean and Hawaiian waters (Indian Ocean, Chen et al., 2007; Hawaiian water, Harman et al., 1989). This may be related to smaller mantle size for the animals in the SCS given that the gladius size was related closely to animal size, which in turn may be attributed to food availability (Seibel et al., 2000). Low primary productivity was well documented for the SCS (Shen et al., 2008), which could lead to slower growth rates and smaller body sizes, presumably owing to their “live for today” life history strategy (Pecl and Moltschaniwskyj, 2006). On the other hand, individuals of the middle-sized typical form with larger sizes in the SCS have been found migrating to areas such as the Taiwan waters (Tung et al., 1973), which would also be expected to lead to smaller middle-sized typical form animals in the SCS. However, it is not possible to confirm any hypothesis at present and further research is needed.
In contrast, little information appeared to be available on the gladius morphology for the dwarf form of S. oualaniensis, although an assumption of more than one type is expected based on Bizikov (1995). We found that dwarf form individuals without a dorsal photophore in the SCS had only one type of gladius morphology, which has double axes. This finding was in contrast to the assumption that the morphology of the gladius was variable for individuals from different types of dwarf form without a dorsal photophore (Zuyev et al., 2002). Thus, given that the gladius morphology is reflective of different forms among purpleback flying squid (Jereb and Roper, 2010), it is reasonable to expect only one type of dwarf form without a dorsal photophore of purpleback flying squid in the SCS.
Gladius morphological characteristics in form identification
The size of the gladius of dwarf forms without a dorsal photophore was significantly smaller than that of middle-sized typical forms (Table 1). This could be associated with the individual’s body size, as it is obvious that dwarf forms had a smaller mantle length than middle-sized typical forms. Within forms, furthermore, there was a significant difference among sexes, in which the gladius of females was larger than that of males (Table 1). This observation could be explained by the sexual dimorphism (Snÿder, 1998) as well as faster growth for female individuals (Liu et al., 2016). Accordingly, the gladius morphometric is confident to identify individuals between dwarf and middle-sized typical forms, evidenced by the obvious groups of the gladii of dwarf and middle-sized typical forms (Figure 3A). Such evidence was further provided by the significant dissimilarity detected in the gladius morphometric characteristics between dwarf and middle-sized typical forms (ANOSIM-R = 0.24, P = 0.001). In comparison with female and male individuals, nMDS and ANOSIM analyses of gladius morphometric characteristics further highlighted that dwarf form individuals were distinguishable from middle-sized typical form ones (Figures 3B–E; Table 2).
Furthermore, the stepwise discriminate analyses revealed that the gladius morphometric characteristics were ideal to distinguish the dwarf and middle-sized typical form S. oualaniensis in the SCS, in which the individuals from these two forms can be accurately classified by four characteristics, namely, GWCs, GWVs, GWPs, and GLs (Table 3). In comparison with other methods such as body morphometric characteristics, the accuracy of discrimination for these two forms is relatively high, e.g., 93% in this study vs. 85% in Zhu et al. (2016) who used body morphometric characteristics. In addition, Liu et al. (2015) have reported that beak morphological characteristics achieved a discriminant accuracy around 79% for classifying the geographical forms of D. gigas. Therefore, using gladius morphological characteristics to identify forms of S. oualaniensis would be expected to achieve more accurate results, probably also for other cephalopods.
Regarding the sexes, individuals of dwarf form were successfully distinguished from middle-sized typical forms using gladius morphometric characteristics. The difference could be explained by five gladius morphometric characteristics, namely, GWPs, PLs, GWCs, GWVs, and GLs, even though the discrimination accuracy was relatively low (Table 4). The low discrimination accuracy could be partly attributed to the lack of dimorphism in gladius morphology between sexes within forms. This observation is very similar to the findings based on beak and statolith morphometric characteristics for other Ommastrephids. For example, it has been reported that female and male Ommastrephes bartramii can be somewhat separated using beak morphology, with a discrimination value of around 56% (Fang et al., 2014). Similar findings were found in the analyses using statolith morphology and beak morphology for Illex argentinus (Fang et al., 2012).
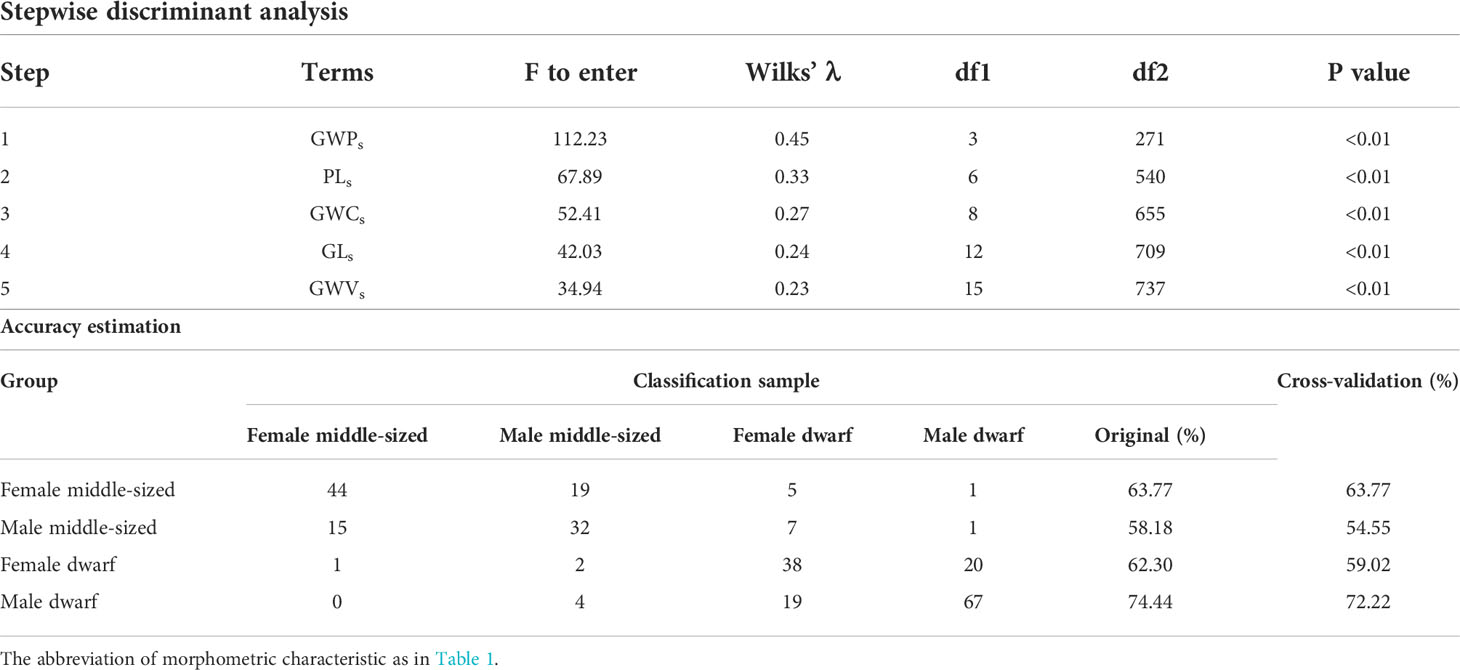
Table 4 The result of stepwise discriminant analyses for gladius morphometric characteristics of female and male dwarf without a dorsal photophore and middle-sized typical forms Sthenoteuthis oualaniensis in the South China Sea.
In conclusion, the dwarf without a dorsal photophore and middle-sized typical forms of S. oualaniensis in the SCS exhibited double axes on the gladius. The size of the gladius of dwarf forms was significantly smaller than that of middle-sized typical forms. Individuals of dwarf form were obviously identified from the middle-sized typical form using the gladius morphometric characteristics, evidenced by the obviously separate groups of gladii of these two forms. Significant differences in the morphometric characteristics between dwarf and middle-sized typical forms were confirmed by the significant dissimilarity values calculated by ANOSIM analysis. Furthermore, stepwise discriminant analyses revealed that there was a high rate of discriminant accuracy for distinguishing the dwarf and middle-sized typical forms using gladius morphometric characteristics. Because the gladius is easier to collect and measure, it is an ideal hard structure to identify forms of S. oualaniensis. Although there are still few studies using the gladius for stock/form identification in squids, the successful distinction of dwarf and middle-sized typical forms of S. oualaniensis here could warrant consideration of the gladius morphology and its morphometric characteristics to study the complicated population structure of squids.
Data availability statement
The original contributions presented in the study are included in the article. Further inquiries can be directed to the corresponding author.
Ethics statement
Ethical review and approval was not required for the animal study because the samples we studied were all from frozen squid caught in commercial fishing activities.
Author contributions
KZ: conception and design of this study, acquisition of data, visualization, software, writing—original draft. DL: Writing—review and editing. XC: investigation, resources. KX: Writing— review and editing. All authors contributed to the article and approved the submitted version.
Funding
This work was supported by the National Key R&D Program of China (2020YFD0901203), An exploratory fishery resources survey of Sthenoteuthis oualaniensis in northern Indian Ocean (CTZB-2020050277 (1)), National Natural Science Foundation of China (41876144, 41876141) and Natural Science Foundation of Shanghai (grant no. 16ZR1415400).
Acknowledgments
This work is a contribution of the Distant Squid Fisheries Sci-Tech Group, SHOU. We thank the staff members of the Key Laboratory of Sustainable Exploitation of Oceanic Fisheries Resources, Ministry of Education, Shanghai Ocean University, for providing assistance at laboratory. We are grateful to Bachelors Chengjie Sun, Lichuan Zhang, Jixiang Yao, Haitao Liu, and HaoGu for the biological data collected. We are grateful to Dr. André E. Punt for his cordial help with English edits.
Conflict of interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Publisher’s note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
References
Bizikov V. A. (1995). Growth of Sthenoteuthis oualaniensis, using a new method based on gladius microstructure. ICES marine. Sci. Symp. 199, 445–458.
Boyle P., Rodhouse P. (2005). Cephalopods: Ecology and fisheries Vol. 464 (Oxford, UK: Wiley-Blackwell).
Chen X. J., Liu B. L., Chen Y. (2008). A review of the development of Chinese distant-water squid jigging fisheries. Fish. Res. 89 (3), 211–221. doi: 10.1016/j.fishres.2007.10.012
Chen X. J., Liu B. L., Tian S. Q., Qian W. G., Zhao X. H. (2007). Fishery biology of purpleback squid, Sthenoteuthis oualaniensis, in the northwest Indian ocean. Fish. Res. 83 (1), 98–104. doi: 10.1016/j.fishres.2006.09.005
Chen X. J., Lu H. J., Liu B. L., Tian. S. Q. (2012). Current exploitation and some scientific issues in the sustainable utilization of ommastrephidae. J. Shanghai. Ocean. Univ. 21 (5), 831–840. doi: 10.1007/s11783-011-0280-z
Chen D. H., Qiu H. M. (2014). Surface microstructures of suckers and their endoskeletons in cephalopods as revealed by SEM. Chinese. J. Zoo. 49 (5), 736–743. doi: 10.13859/j.cjz.201405013
Clavel J., Morlon H. (2017). Accelerated body size evolution during cold climatic periods in the Cenozoic. P. Natl. Acad. Sci. U.S.A. 114 (16), 4183–4188. doi: 10.1073/pnas.1606868114
Fang Z., Chen X. J., Lu H. J. (2012). Morphological differences in statolith and beak between two spawning stocks for Illex argentinus. Acta Ecologica. Sinica. 32 (19), 5986–5997. doi: 10.5846/stxb201109101331
Fang Z., Liu B. L., Li J. H., Su H., Chen X. J. (2014). Stock identification of neon flying squid (Ommastrephes bartramii) in the north pacific ocean on the basis of beak and statolith morphology. Sci. Mar. 78 (2), 239–248. doi: 10.3989/scimar.03991.06A
Fan J. T., Qiu Y. S., Chen Z. Z., Feng X., Zhang J. (2015). Morphological difference of the beak between two stocks of Sthenoteuthis oualaniensis inhabiting south China Sea. Period. Ocean. Univ. China. 45 (10), 42–49. doi: 10.16441/j.cnki.hdxb.20140270
Gong Y., Chen X. J., Li Y. K., Fang Z. (2018). Geographic variations of jumbo squid (Dosidicus gigas) based on gladius morphology. Fish. Bull. 116 (1), 50–59. doi: 10.7755/FB.116.1.5
Harman R. F., Young R. E., Reid S. B., Mangold K. M., Suzuki T., Hixon R. F. (1989). Evidence for multiple spawning in the tropical oceanic squid Stenoteuthis oualaniensis (Teuthoidea: Ommastrephidae). Mar. Biol. 101 (4), 513–519. doi: 10.1007/BF00541653
Jereb P., Roper C. F. E. (Eds.) (2010). “Cephalopods of the world. an annotated and illustrated catalogue of cephalopod species known to Date.Vol 2. myopsid and oegopsid squids,” in FAO species catalogue for fishery purposes, vol. Vol. 2 (Rome: FAO), 4.
Keyl F., Argüelles J., Tafur R. (2011). Interannual variability in size structure, age, and growth of jumbo squid (Dosidicus gigas) assessed by modal progression analysis. ICES. J. Mar. Sci. 68 (3), 507–518. doi: 10.1093/icesjms/fsq167
Laptikhovsky V., Nikolaeva S., Rogov M. (2017). Cephalopod embryonic shells as a tool to reconstruct reproductive strategies in extinct taxa. Biol. Rev. 93, 270–283. doi: 10.1111/brv.12341
Lin D. M. (2015). Spawning strategy of Argentine shortfin squid, illex argentinus(Cephalopoda: Ommastrephidae) in the southwest Atlantic (Shanghai: Shanghai Ocean University).
Liu B. L., Chen X. J. (2010). Research progress on shells in cephalopids. Mar. Fish 32 (3), 332–339. doi: 10.13233/j.cnki.mar.fish.2010.03.001
Liu B. L., Chen X. J., Li J. H., Chen Y. (2016). Age, growth and maturation of Sthenoteuthis oualaniensis in the eastern tropical pacific ocean by statolith analysis. Mar. Freshwater. Res. 67 (12), 1973–1981. doi: 10.1071/mf14427
Liu B. L., Chen X. J., Zhong J. S. (2008). Statolith morphology of the purpleback flying squid Sthenoeuthis oualaniensis in the northwest Indian ocean. J. Shanghai. Ocean. Univ. 17 (5), 604–609. doi: 10.3724/SP.J.1035.2008.00038
Liu B. L., Fang Z., Chen X. J., Chen Y. (2015). Spatial variations in beak structure to identify potentially geographic populations of Dosidicus gigas in the Eastern pacific ocean. Fish. Res. 164, 185–192. doi: 10.1016/j.fishres.2014.12.001
Li M., Zhang P., Zhang J., Zhang K., Chen Z. Z. (2019). Genetic differentiation of the purpleback flying squid Sthenoteuthis oualaniensis in the south China Sea: population or species divergence. J. Fish. Sci. China. 26 (01), 133–140. doi: 10.3724/SP.J.1118.2019.18193
Nesis K. N. (1993). “Population structure of oceanic ommastrephids, with particular reference to sthenoteuthis oualaniensis: a review,” in Recent advances in fisheries biology. Eds. Okutani T., O’Dor R. K., Kubodera T. (Tokyo: Takai University Press), 375–383.
Pecl G. T., Moltschaniwskyj N. A. (2006). Life history of a short-lived squid (Sepioteuthis australis): resource allocation as a function of size, growth, maturation, and hatching season. ICES. J. Mar. Sci. 63, 995–1004. doi: 10.1016/j.icesjms.2006.04.007
R Core Team (2018). R: A language and environment for statistical computing (Vienna, Austria: R Foundation for Statistical Computing). Available at: http://www.R-project.org/.
Rencher A. C. (2002). Methods of multivariate analysis, 2nd edition (New York: John Wiley & Sons, Inc).
Rodhouse P. G. K., Arkhipkin A. I., Laptikhovsky V. (2013). “Illex argentines, Argentine shortfin squid,” in Advances in squid biology and fisheries (Part II.Oegopsid squids). Eds. Rosa R., O’Dor R., Pierce G. (New York: Nova Biomedical), 109–148.
Seibel B. A., Thuesen E. V., Childress J. J. (2000). Light-limitation on predator-prey interactions: consequences for metabolism and locomotion of deep-sea cephalopods. Biol. Bull-US. 198 (2), 284–298. doi: 10.1016/s1095-6433(99)90328-8
Shen S. H., Leptoukh G. G., Acker J. G., Yu. Z. J., Kempler S. J. (2008). Seasonal variations of chlorophyll a concentrations in the northern south China Sea. IEEE. Geosci. Remote. S. 5 (2), 315–319. doi: 10.1109/lgrs.2008.915932
Snÿder R. (1998). Aspects of the biology of the giant form of Sthenoteuthis oualaniensis (Cephalopoda: Ommastrephidae) from the Arabian Sea. J. Mollus. Stud. 64, 21–34. doi: 10.1093/mollus/64.1.21
Staaf D. J., Ruiz-Cooley R. I., Elliger C., Lebaric1 Z., Bernardita C., Markaida U., et al. (2010). Ommastrephid squids Sthenoteuthis oualaniensis and Dosidicus gigas in the eastern pacific show convergent biogeographic breaks but contrasting population structures. M. E. P. S. 418, 165–178. doi: 10.3354/meps08829
Tung I. H., Lan C. S., Hu C. C. (1973). The preliminary investigation for exploitation of common squid resources. Rep. Institute Fishery Biol. Ministry Economic Affairs Natl. Taiwan University. 3, 211.
Wang X. H., Liu B. L., Qiu Y. S., Zhang P., Du F. Y., Chen X. J., et al. (2019). Regional and intraspecific variation in Sthenoteuthis oualaniensis (Cephalopoda: Ommastrephidae) morphology from the western central pacific ocean. B. Mar. Sci. 95 (1), 1–12. doi: 10.5343/bms.2017.1173
Wang X. H., Qiu Y. S., Zhang P., Du F. Y. (2017). Natural mortality estimation and rational exploitation of purpleback flying squid Sthenoteuthis oualaniensisin the southern south China Sea. China. J. Oceanol. Limn. 35 (4), 902–911. doi: 10.1007/s00343-017-6115-4
Wanninger A., Wollesen T. (2018). The evolution of mollusks. Biol. Rev. 94 (1), 102–115. doi: 10.1111/brv.12439
Wright J., Bolstad G. H., Araya-Ajoy Y. G., Dingemanse N. J. (2018). Life-history evolution under fluctuating density-dependent selection and the adaptive alignment of pace-of-life syndromes. Biol. Rev. 94 (1), 230–247. doi: 10.1111/brv.12451
Yang D. K. (2002). The resources and its exploitation and utilization of two species of squid. J. Shanghai. Ocean. Univ. 11, 176–179. doi: 10.1111/j.1469-8137.1982.tb03304.x
Yan Y. R., Yi M. R., Feng B., Zhao C. X., Li Z. H., Lu H. S. (2015). Morphological variations and discriminant analysis of three different geographic cohorts of Sthenoteuthis oualaniensis in the south China Sea. J. Guangdong. Ocean. Univ. 35 (3), 43–50.
Zhang J., Chen G. B., Zhang P., Chen Z. Z., Fan J. T. (2014). Estimation of purpleback flying squid (Sthenoteuthis oualaniensis) resources in the central and southern south China Sea based on fisheries acoustics and light-falling net. J. Fishery Sci. China 21 (4), 822–831. doi: 10372/SP.J.1118.2014.00822
Zhang Y. M., Yan Y. R., Lu H. S., Zheng Z. W., Yi M. R. (2013). Study on feeding and reproduction biology of purple flying squid, Sthenoteuthis oualaniensis in the Western south China Sea. J. Guangdong Ocean Univ. 33 (3), 56–64.
Zhu K., Wang X. H., Zhang P., Du F. Y. (2016). A study on morphological variations and discrimination of medium and dwarf forms of purple flying squid Sthenoteuthis oualaniensis in the southern south China Sea. J. Trop. Oceanography 35 (6), 82–88. doi: 10.11978/2016011
Keywords: Sthenoteuthis oualaniensis, gladius, form discrimination, hard structure, South China Sea
Citation: Zhu K, Lin D, Chen X and Xu K (2022) Form identification of purple flying squid (Sthenoteuthis oualaniensis) based on gladius morphology. Front. Mar. Sci. 9:956461. doi: 10.3389/fmars.2022.956461
Received: 30 May 2022; Accepted: 21 October 2022;
Published: 11 November 2022.
Edited by:
Jan Strugnell, James Cook University, AustraliaReviewed by:
Cesar A. Salinas-Zavala, Centro de Investigación Biológica del Noroeste (CIBNOR), MexicoMariana Díaz-Santana-Iturrios, Andres Bello University, Chile
Jose Angel Alvarez Perez, Universidade do Vale do Itajaí, Brazil
Copyright © 2022 Zhu, Lin, Chen and Xu. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Kaida Xu, xukaidahys@126.com