 Luca Tonin1*
Luca Tonin1* Marco Pitteri2
Marco Pitteri2 Robert Leeb1
Robert Leeb1 Huaijian Zhang1
Huaijian Zhang1 Emanuele Menegatti3
Emanuele Menegatti3 Francesco Piccione4,5
Francesco Piccione4,5 José del R. Millán1*
José del R. Millán1*- 1Chair in Brain-Machine Interface, Center for Neuroprosthetics, École Polytechnique Fédérale de Lausanne, Geneva, Switzerland
- 2Neurology Section, Department of Neurosciences, Biomedicine and Movement Sciences, University of Verona, Verona, Italy
- 3Intelligent Autonomous Systems Laboratory, Department of Information Engineering, University of Padova, Padova, Italy
- 4Laboratory of Neuropsychology, IRCCS San Camillo Hospital Foundation, Venice, Italy
- 5Laboratory of Neurophysiology, IRCCS San Camillo Hospital Foundation, Venice, Italy
During the last years, several studies have suggested that Brain-Computer Interface (BCI) can play a critical role in the field of motor rehabilitation. In this case report, we aim to investigate the feasibility of a covert visuospatial attention (CVSA) driven BCI in three patients with left spatial neglect (SN). We hypothesize that such a BCI is able to detect attention task-specific brain patterns in SN patients and can induce significant changes in their abnormal cortical activity (α-power modulation, feature recruitment, and connectivity). The three patients were asked to control online a CVSA BCI by focusing their attention at different spatial locations, including their neglected (left) space. As primary outcome, results show a significant improvement of the reaction time in the neglected space between calibration and online modalities (p < 0.01) for the two out of three patients that had the slowest initial behavioral response. Such an evolution of reaction time negatively correlates (p < 0.05) with an increment of the Individual α-Power computed in the pre-cue interval. Furthermore, all patients exhibited a significant reduction of the inter-hemispheric imbalance (p < 0.05) over time in the parieto-occipital regions. Finally, analysis on the inter-hemispheric functional connectivity suggests an increment across modalities for regions in the affected (right) hemisphere and decrement for those in the healthy. Although preliminary, this feasibility study suggests a possible role of BCI in the therapeutic treatment of lateralized, attention-based visuospatial deficits.
Introduction
Spatial neglect (SN) is one of the most frequent and disabling neuropsychological syndromes following right-hemisphere damage (Heilman et al., 2003; Buxbaum et al., 2004; Adair and Barrett, 2008). SN patients usually fail to report stimuli in the contralesional side of space, rendering difficult their effective perception of the surrounding space. Although some spontaneous recovery occurs in the majority of patients after stroke, SN might remain severe in the chronic phase, limiting considerably the effectiveness of rehabilitation interventions (Katz et al., 1999; Battelli et al., 2001; Jehkonen et al., 2006; Kerkhoff and Schenk, 2012; Bowen et al., 2013; Riestra and Barrett, 2013).
One of the most accredited hypothesis explaining SN was introduced by Kinsbourne and it relies on the concept of inter-hemispheric rivalry (Kinsbourne, 1993). In terms of attentional vectors, it is assumed that lesions to the right-hemisphere provoke changes in neural activation inducing inter-hemispheric imbalance and, as a consequence, a hypoexploration of the left (neglected) space and a hyperattention toward the right.
Recently, it has been proposed that SN should be better attributed to abnormalities in the functional organization of large fronto-parietal attention networks, rather than lesions in local cerebral areas (Corbetta et al., 2005; He et al., 2007; Bartolomeo et al., 2012). Furthermore, it has been demonstrated that the normalization of the inter-hemispheric connectivity is a physiological signature of recovery from SN (Baldassarre et al., 2014; Ramsey et al., 2016).
Behavioral treatments of SN are heterogeneous, because they are based on different theoretical concepts (Riestra and Barrett, 2013). Most of them, however, are based on the concept of re-orienting the visuospatial attention toward the neglected side of space (Kortte and Hillis, 2011; Priftis et al., 2013; Azouvi et al., 2016). Recently, it has been demonstrated that such interventions may have direct effects in the neural mechanisms of SN patients (Saj et al., 2013). Furthermore, novel approaches have been proposed based on neurofeedback or brain stimulation techniques in order to suppress/enhance the hyper/hypo-activity in the healthy/affected hemisphere (Kortte and Hillis, 2011; Müri et al., 2013; Okazaki et al., 2014; Robineau et al., 2014).
The current study aims at evaluating the feasibility of a novel approach based on Brain-Computer Interface (BCI). The last years have seen a growing interest of the scientific community in identifying new directions and different target populations for BCI-driven control and rehabilitation (Brunner et al., 2015). Indeed, beyond the use as an assistive device (Birbaumer, 2006; Sellers et al., 2014; Leeb et al., 2015), evidences suggest that BCI can play a relevant role in motor rehabilitation by associating actual or imagined motor tasks to a coherent, real-time feedback provided to the patient (Daly and Wolpaw, 2008; Grosse-Wentrup et al., 2011; Ramos-Murguialday et al., 2013; Ang and Guan, 2017). Focusing on electroencephalography (EEG)-based studies, recent reviews have highlighted the potential benefits of BCI approaches in motor rehabilitation after stroke (Monge-Pereira et al., 2017; Remsik et al., 2017). Based on the same principles, we hypothesized that BCI systems might also be adopted in the case of cognitive rehabilitation of SN. Our BCI approach exploits the covert visuospatial attention (CVSA) orienting paradigm (Tonin et al., 2012; Marchetti et al., 2013; Tonin et al., 2013) that might offer a direct backdoor to the impaired visuospatial attention mechanisms of SN patients.
Herein, we report the cases of three SN patients who operated an EEG-based CVSA BCI during two consecutive weeks. We evaluated the ability of each patient to operate the BCI and we analyzed possible neurophysiological changes during online BCI operations. To the best of our knowledge, this is the first attempt of using a CVSA BCI and analyzing its contingency effects in SN patients.
Case Description
Three SN patients (P1-3, from 46 to 61 years old, median 57; two females) with unilateral right-hemisphere damage participated in the study. Patients did not have previous experience with BCI systems. Written informed consent was obtained from all patients to participate in the study, to collect data and to publish information appearing in this case report. Patients were tested in accordance with the Helsinki declaration. The study was approved by the institutional ethical committee (Nucleo per la Ricerca Clinica) of the IRCCS San Camillo Hospital Foundation, Venice, Italy. All patients had unilateral lesions because of first stroke (time from lesion: 4, 8, and 13 months, respectively; Supplementary Table 1). Lesion sites were confirmed by magnetic resonance imaging scans and localized in the fronto-parietal lobe and in the insula (P1); in the capsulo-thalamic area and in the insula (P2); and in the fronto-temporal-parietal lobe (P3).
Patients were assessed with a screening test to exclude general cognitive impairment (Measso et al., 1993), and with a battery of neuropsychological tests to detect SN in the peripersonal space (Wilson et al., 1987; Vallar et al., 1994). Clinical signs of SN were present in each patient (P1: BIT-conventional = 104, cut-off < 130; P2: MMSE = 23.31, cut-off < 24; BIT-conventional = 122, cut-off < 130; Symbol cancelation: 3, -3 ≤ cut-off ≥ 3; P3: Symbol cancelation = 5, -3 ≤ cut-off ≥ 3; Supplementary Table 2).
Materials and Methods
Experimental Design
Patients were asked to control an online two-class BCI by means of a voluntary CVSA-orienting task (Figure 1A). Each trial started with a fixation cross (size 3.12°) in the middle of the screen and a random image of real object (4.8°) at the bottom left (neglected side). After 3000 ms, patients were instructed by a cue (300 ms) to covertly focus their attention to the left target (attention condition) or to keep fixating the center of the screen (rest condition). After a random time (3000–4000 ms), the target was outlined in red. In case of left target selection, the image started moving toward the center. Patients were required to press a button with their right hand as soon as they perceived the target selection.
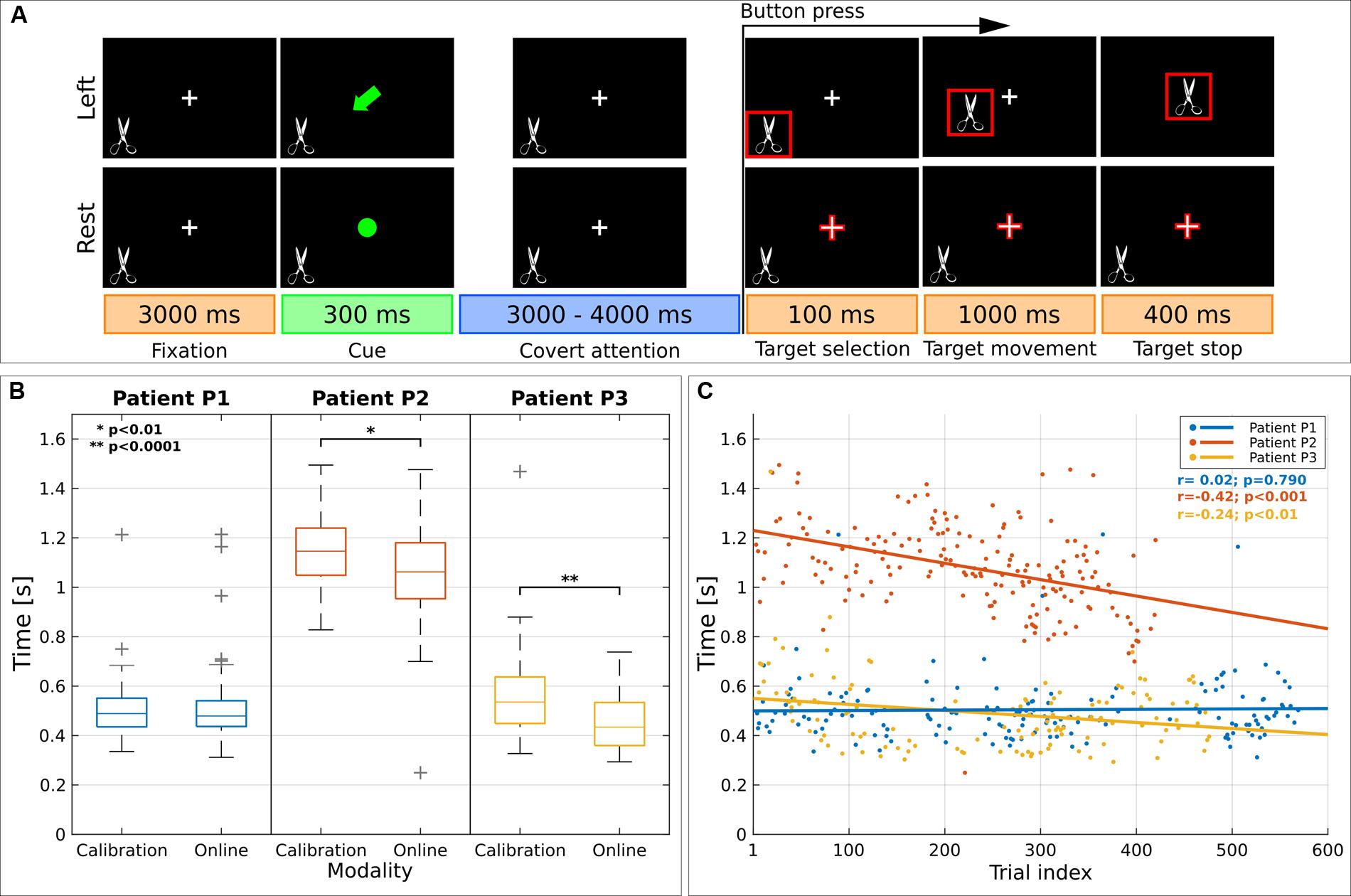
FIGURE 1. Visual paradigm and reaction time (RT). (A) Schematic representation of the paradigm presented to spatial neglect (SN) patients. Each trial started with a fixation period (3000 ms) where patients had to gaze at a cross, visible at the center of the screen. Then the cross was replaced by a symbolic cue (300 ms) indicating the to-be-attended location. The covert attention period lasted for 3000–4000 ms; afterward, one of the two to-be-attended locations was highlighted in red (target selection, 100 ms) as online feedback of the classification result. Immediately afterward—in the case of left target—the image started moving toward the center of the screen (target movement, 1000 ms) and it disappeared after 400 ms (target stop). SN patients were required to press a button with their right hand as soon as they perceived the images highlighted in red. (B) Analysis on RT for left covert visuospatial attention (CVSA) task. Distribution of RT for each patient across modality. For each box, median is reported. Box edges represent the 25th and 75th percentile of Individual α-Power (IAP). Student t-test outcomes are annotated in each plot. (C) Scatter plot shows the evolution of RT over time. Results are reported for each patient (in blue, red, and yellow for P1, P2 and P3, respectively). Lines represent the least-squares fit to the scatter plot for each patient. Spearman’s correlation and significance are annotated.
Each patient performed six recording sessions within two consecutive weeks. In average, each session consisted of 2.7 ± 0.8 runs and each run of 30 trials randomly shuffled between attention (20) and rest (10) condition (Supplementary Table 3). The first two sessions were devoted to the calibration of the BCI with a positive feedback always delivered to patients; in the following runs (online modality) the image selected at the end of the trial was based on the output of the CVSA BCI, as real-time feedback for the patients.
CVSA BCI
Electroencephalography signals were acquired with a 64-channel system at 2048 Hz (BioSemi, Amsterdam, Netherlands). Electrodes were placed according to the standard international 10–20 system. Eye movements were recorded by means of three electrodes placed at the outer canthi of the eyes and at the gabella. The CVSA BCI was similar to our previous work (Tonin et al., 2012, 2013). The envelope of the EEG was extracted in seven α sub-bands (8–14 Hz, with 3 Hz of bandwidth) by means of Hilbert transform and a Laplacian filter was applied. Channels were pre-selected in the parieto-occipital regions (17 electrodes: P7–8, PO7–8, O1–2). Trial classification was based on data from the first 3000 ms after the cue. This period was split into windows of 150 ms. For each window, a quadratic discriminant analysis classifier was trained with the most discriminant features (frequency-channel pairs) selected during the calibration. In the online modality, classifiers were evaluated and the resulting posterior probabilities were integrated over time to deliver the final decision at the end of the trial.
Neurophysiological Analysis
Data Processing
Data were spatially filtered with Common Average Reference and periodogram was extracted in the 4–48 Hz frequency range (1 Hz resolution, 0.0625 ms shift, 1000 ms window). Trials were extracted in the pre-cue interval. We defined six nodes (frontal, parietal, and occipital regions for the left- and right-hemisphere; Supplementary Table 4) according to (Baldassarre et al., 2014).
Individual α-Frequency (IAF) and Individual α-Power (IAP)
We selected symmetric nodes in the parieto-occipital regions (Thut et al., 2006). The spectrum of each channel was normalized for inter-trial comparisons. For each hemisphere, the IAF was defined as the first peak in the frequency range between 6 and 12 Hz (Supplementary Table 5). If no peak was found in the right (affected) hemisphere, the corresponding average peak computed in the same modality was selected. Therefore, Individual α-Power (IAP) was computed as the average around IAF (±1 Hz) for each trial and for each hemisphere.
Inter-Hemispheric Discriminancy
First, we selected those features that patients were modulating during BCI operations. Second, we extracted those related to the IAF of each patient. Third, for each run we computed the Fisher Score (FS) values of each feature as follows:
where k indicates the kth feature, μ the mean and σ the variance over the attention and rest conditions. This metric represents the participants’ ability to modulate their neural networks accordingly to the CVSA-orienting task. Fourth, for each run we computed the differences between the FS of each homotopic nodes (right minus left hemisphere).
Inter-Hemispheric Functional Connectivity (FC)
Analysis on FC was based on (Baldassarre et al., 2014). First, for each patient, IAP was extracted in the trial interval (left CVSA task). Second, we averaged the IAP of the channels belonging to each aforementioned nodes. Third, we generated a channel-wise FC map for each node by extracting the time-course of the IAP for the node and computing the Pearson’s correlation coefficient between that time-course and the time-course of all the other channels. Finally, we computed the inter-hemispheric connectivity for each node by averaging the FC values of those channels belonging to the nodes in the opposite hemisphere.
Statistical Tests
Statistics were based on Student’s t-test between data distributions from the two modalities (calibration vs. online). Statistical significance values reported in the paper are Bonferroni corrected.
Results and Discussion
CVSA BCI Online Accuracy
We computed BCI performance as the percentage of trials that the BCI classified correctly during each online session. The average performance was 55 ± 3.9%, 60 ± 2.8% and 58.3 ± 3.7% (median and standard error) for patients P1, P2, and P3. Although patients’ BCI performance was low, it was above random for most sessions with an individual maximum accuracy of 76.6, 70, and 70% in their best runs. Furthermore, such a level of BCI accuracy is similar to that achieved by stroke patients during BCI-based motor rehabilitation (Prasad et al., 2010; Ramos-Murguialday et al., 2013; Pichiorri et al., 2015).
A posteriori analysis of ocular artifacts showed that horizontal eye movements were moderate for all three patients (9.4, 3.6, and 25% of the trials for P1, P2, and P3). Furthermore, the mean square contingency coefficient between the direction of horizontal eye movements and the CVSA tasks was not significant (P1: φ2 = 0.53, p = 0.46; P2: φ2= 0.83, p = 0.36; P3: φ2= 0.29, p = 0.58).
Behavioral Outcome
Figures 1B,C report the analysis of the behavioral response to the button press across modalities (calibration or online) and over time for left CVSA task. Results showed a decrement of the reaction time (RT) for patients P2 and P3 (1.15 ± 0.15 s vs. 1.06 ± 0.17 s, p < 0.01 and 0.57 ± 0.20 s vs. 0.45 ± 0.10 s, p < 0.001). Patient P1 did not exhibit any change in RTs (0.50 ± 0.13 s vs. 0.50 ± 0.13 s; p = 0.72), however, he had the fastest initial RTs. A significant negative correlation between RTs and trial index was found for patients P2 and P3 (r = -0.42, p < 0.001 and r = -0.24, p < 0.01). The speedup in RTs cannot be explained as a consequence of behavioral training, since the duration of the trials was random.
Inter-Hemispheric Asymmetry of IAF and IAP
We analyzed the spectral and spatial locations of the IAF in the parieto-occipital regions during the pre-cue interval for each patient, modality and hemisphere. During calibration the percentage of trials with missing α-peak was significantly higher in the affected hemisphere (1.73 ± 2.71% vs. 10.02 ± 7.68%, p < 0.01; mean and standard deviation for healthy vs. affected hemisphere). However, during the online modality, a reduction of this percentage seems to occur (1.73 ± 3.1% vs. 7.52 ± 7.90%, p < 0.01). This spatial asymmetry substantially differs from healthy populations, where a symmetric α-response is expected along the two hemispheres (Klimesch et al., 1998; de Munck et al., 2007).
A comparison of the related IAP is reported in Figure 2 (first row) for each modality and hemisphere. Statistical analysis showed an increment in the affected hemisphere for all patients during the online modality (statistically significant for P1 and P3: p < 0.00001; marginally significant for P2: p < 0.05 without correction). For patient P3 the increment was significant also in the healthy hemisphere.
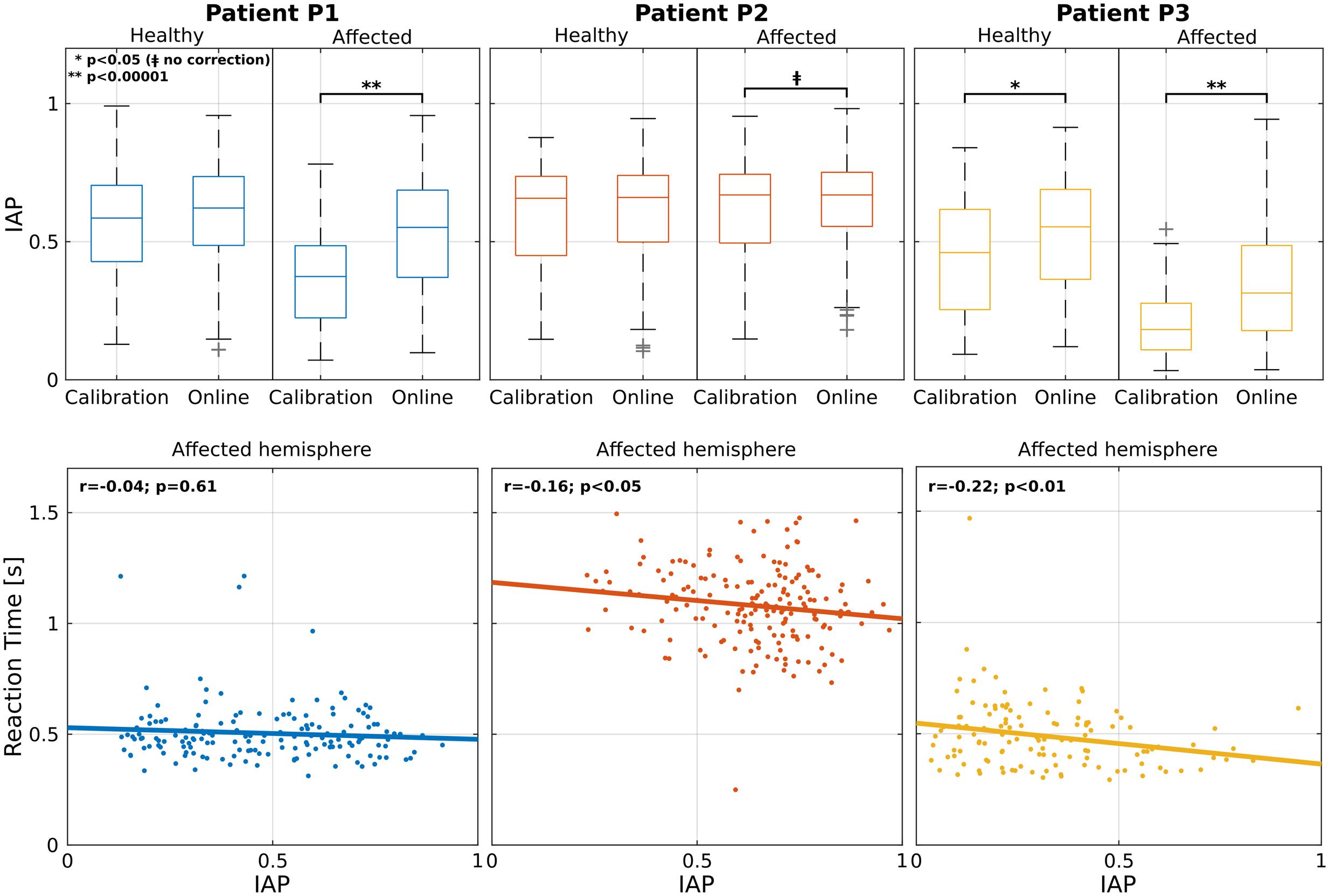
FIGURE 2. Analysis on the IAP. Individual patient results are grouped by column. First row shows the IAP averaged across calibration and online modalities for healthy (left) and affected (right) parieto-occipital nodes. For each box, median is reported. Box edges represent the 25th and 75th percentile of IAP. Student t-test outcomes are annotated in each plot. In the second row, the scatter plots show the relation between IAP and RT for the parieto-occipital nodes in the affected (right) hemisphere. Each point corresponds to a trial. Lines represent the least-squares fit to the scatter plot. Spearman’s correlation and significance are annotated in each plot.
Furthermore, the aforementioned restoration of the α-peak in the pre-cue interval seems to anticipate the decrement of the RTs in the correctly classified left trials (Figure 2, second row). In fact, patients P2 and P3 showed a significant negative correlation between the RTs and the IAP in the affected hemisphere (r = -016, p < 0.05 and r = -0.22, p < 0.01). This did not apply for patient P1 (r = -0.04, p = 0.61) but he/she was the only one who showed constantly low RTs over all runs. These results are in line with the literature, where it has been demonstrated that exists a positive correlation between the strength of α-power in the pre-stimulus interval and a faster RT (Klimesch et al., 1996; Nenert et al., 2012).
Inter-Hemispheric Asymmetry of Features Discriminancy
Feature discriminancy is a common BCI metric to assess the subject’s ability of modulating channel-frequency pairs during different mental tasks (Galán et al., 2007; Leeb et al., 2015). Herein, we investigated possible inter-hemispheric changes in the spatial distribution of discriminancy across the two experimental modalities (calibration and online).
Topographic maps in Figure 3 show the spatial distribution of discriminancy during calibration and online modalities (only parieto-occipital channels exploited in the online BCI are reported; normalized values are shown for comparison purposes; low discriminancy in blue, high in red). An initial imbalance of the discriminancy toward the healthy hemisphere (patients P1 and P3) and toward the affected one (patient P2) seems to be attenuated in the online modality. This hypothesis is supported by analysis on the difference of discriminancy between homotopic regions (Figure 3, second row). Negative values correspond to asymmetry in the modulation toward the left (healthy) hemisphere. A general asymmetry reduction (values toward zero) was reported for all patients in the parietal or the occipital regions with statistical significance for P1 and P3 (p < 0.05) and marginal significance for P2 (p < 0.05, without correction). Furthermore, for all patients, the asymmetry toward the healthy hemisphere significantly decreased across runs (P1, parietal regions: r = 0.59, p < 0.01; P2 and P3, occipital regions: r = 0.52, p < 0.05 and r = 0.51, p < 0.05) (Figure 3, third row).
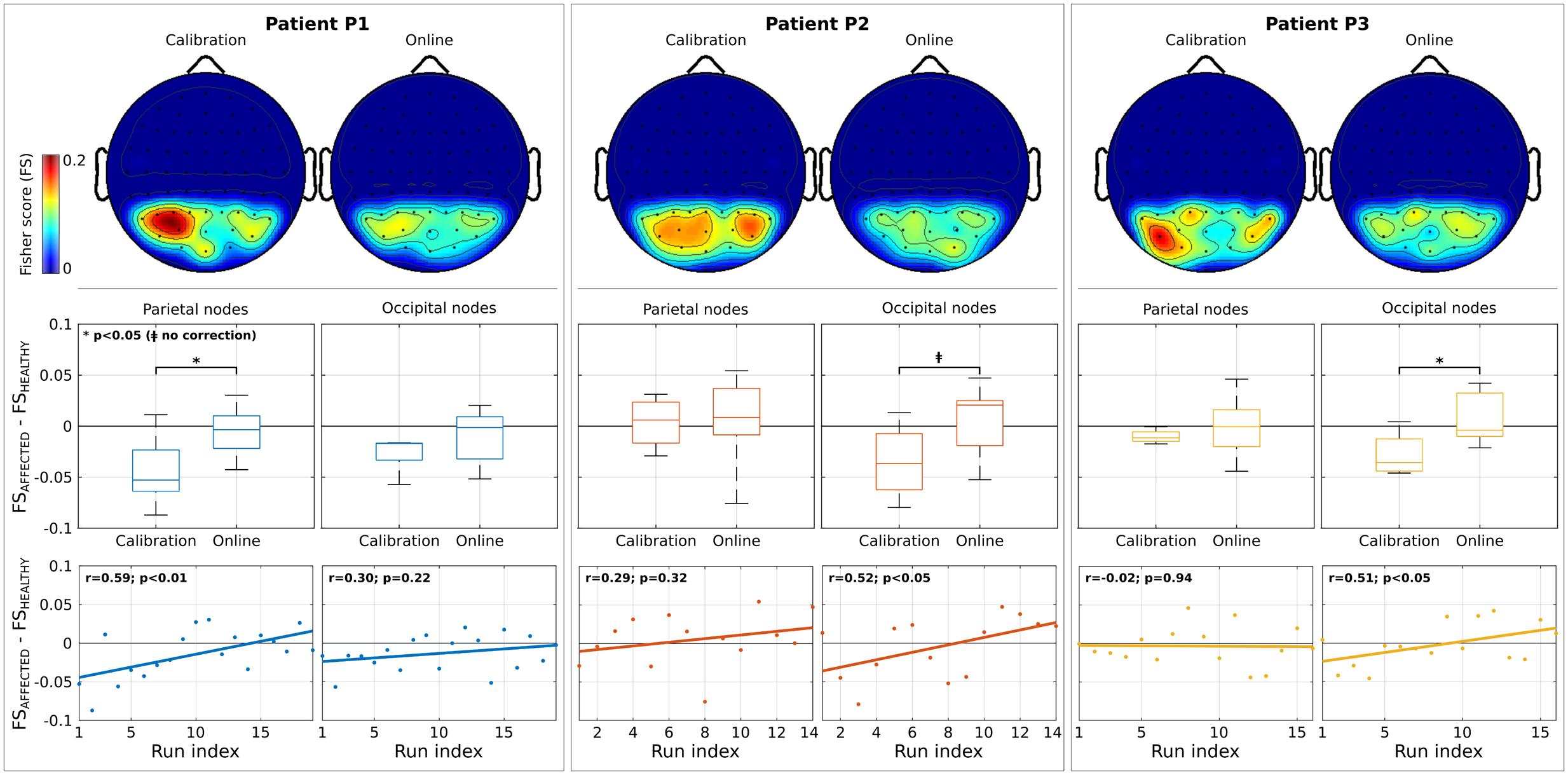
FIGURE 3. Analysis on features discriminancy. Individual patient results are grouped by column. First row illustrates the topographic maps of Fisher Score (FS) values normalized between 0 and 1 (for comparison purposes) for calibration and online modality. Low and high discriminancy is reported in blue and red, respectively. Only the parieto-occipital channels used as input for Brain-Computer Interface (BCI) online operations are shown. Second row depicts the difference of FS values between nodes belonging to affected and healthy hemisphere (parieto and occipital regions, separately) for each modality. In each box, median is reported. Box edges represent the 25th and 75th of FS. Negative values correspond to a stronger modulation in the healthy (right) hemisphere. Student t-test outcomes are annotated in each plot. Third row reports the evolution over runs of the difference of FS values. Each point corresponds to the value computed in a run. Lines represent the least-square fit to the scatter plot. Spearman’s correlation and significance are annotated in each plot.
Inter-patient differences might be explained by the fact that the most discriminant channels are strictly subject-dependent during CVSA tasks, as already reported in literature (Tonin et al., 2012, 2013). These results are in line with the rivalry hypothesis of SN (Kinsbourne, 1993) with the additional advantage of contingency with respect to the attention task performed by the patients.
Inter-Hemispheric FC
Several functional Magnetic Resonance Imaging (fMRI) studies reported a relation between FC in large scale resting-state networks and SN (Corbetta et al., 2005; He et al., 2007). On the other hand, our within-patient connectivity analyses focused on investigating the FC during the attention task (driven by the CVSA BCI) and possible changes between calibration and online trials by exploiting the high spectro-temporal resolution of EEG signals. A general inter-patient trend seems to appear, highlighting an increment of inter-hemispheric FC for nodes in the affected (right) hemisphere and decrement for those in the healthy (left) one (Figure 4). Topographic plots show the difference between online and calibration FC maps computed with respect to each of the selected nodes (electrodes highlighted in magenta). Boxplots illustrate the difference in calibration and online modality between the FC of each node and the FC averaged across the whole opposite hemisphere. Only statistically significant differences are reported. For patient P2 the inter-hemispheric FC increment happened in the right-occipital node (p < 0.05) and for patient P3 in right-frontal (p < 0.001) and right-parietal nodes (p < 0.001). Contrarily, for patient P1, decrement of FC occurred in left-parietal (p < 0.01) and left-occipital nodes (p < 0.05) and, again, for patient P2 marginally in left-frontal node (p < 0.05, without correction). Changes in FC did not correlate with RT for any patient or node. Previous studies in resting-state networks demonstrated that the increasing inter-hemispheric connectivity (from right parietal regions to left hemisphere) is a signature of recovery from SN (Ramsey et al., 2016). The missing correlation between connectivity changes and RT might be due to the task-dependent nature of our analysis and, consequently, to the task-induced specific correlation patterns between different cortical regions (Zanto et al., 2011) as already suggested in (Baldassarre et al., 2014).
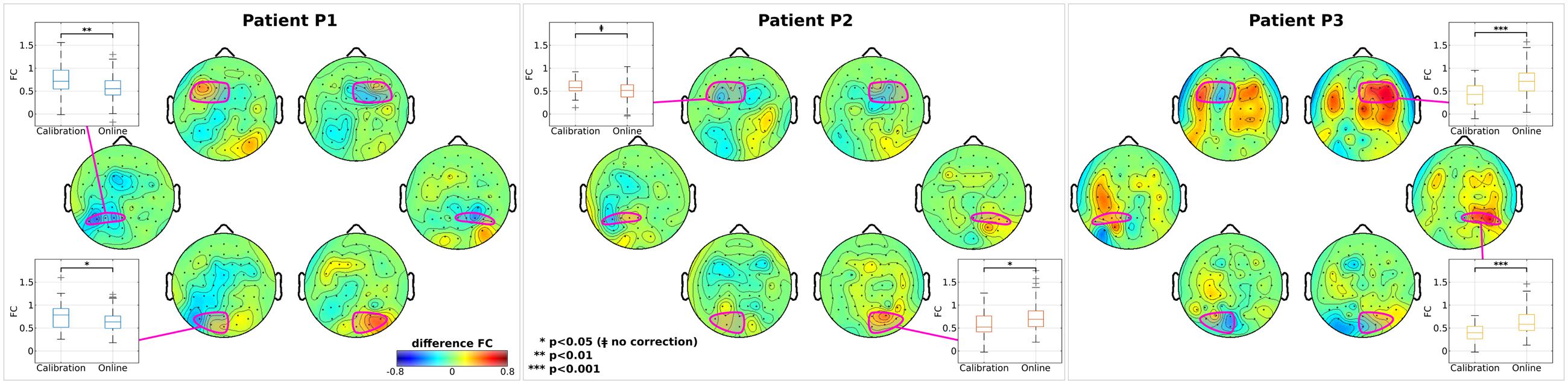
FIGURE 4. Analysis on inter-hemispheric connectivity. For each patient, topographic maps of z-transformed functional connectivity (FC) between each node and the whole opposite hemisphere are reported. Difference between online and calibration modality is reported for visualization purposes. In each map, nodes (and channels belonging to) are highlighted in magenta. Box plots represent the distributions of the z-transformed FC in the calibration and online modality. Only nodes with statistical significance difference between modality are shown. In each box, median is reported. Box edges represent the 25th and 75th of the z-transformed FC. Student t-test outcomes are annotated in each plot.
Concluding Remarks
To the best of our knowledge, for the first time cortical effects in EEG patterns have been monitored during online CVSA BCI operations performed by SN patients. As primary outcome, our study showed that three patients suffering from SN can actively control a CVSA BCI. Nonetheless, an improvement of such a level of control might be desirable to enhance the neurofeedback- driven reward and, as a consequence, the neuroplasticity response (Kaiser et al., 2012; Young et al., 2014). In this regard, additional neural correlates of CVSA [e.g., the lateralization index (Thut et al., 2006)] and new training paradigms can be investigated in future studies. From the neurophysiological point of view, inter-modality analyses on the restoration of the initial α-response asymmetry, as well as on the increment of modulation and connectivity in the affected hemisphere, suggest a positive effect of the online BCI feedback. However, it should be noticed that causality between online BCI operations and changes in the inter-hemispheric activity cannot be firmly established yet, given the limited population size and the uncontrolled experimental design. Further studies are required in order to verify such hypotheses. First, it is mandatory to run a long-term randomized controlled trial with larger groups to identify the effective contribution of the online BCI feedback. Second, it is also crucial to probe with other functional techniques, such as fMRI, the actual recruitment of perilesional areas and their role in inter-hemispheric activity. Finally, such a kind of CVSA BCI should be combined with tailored cognitive interventions to optimize the clinical outcomes and to reduce the confounding factors and the within-patient variability effects.
Author Contributions
LT, RL, and JdRM were responsible for the study conception; LT and MP implemented and executed the experiments; LT, MP, RL, HZ, EM, FP, and JdRM contributed to the methodology, data analysis, and manuscript preparation.
Funding
LT, RL, and JdRM reports grants from European Union, FP7-224631, during the conduct of the study.
Conflict of Interest Statement
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Acknowledgments
This work was partially supported by the European ICT Programme Project TOBI (FP7-224631). This paper only reflects the authors’ views and funding agencies are not liable for any use that may be made of the information contained herein.
Supplementary Material
The Supplementary Material for this article can be found online at: http://journal.frontiersin.org/article/10.3389/fnhum.2017.00336/full#supplementary-material
References
Adair, J. C., and Barrett, A. M. (2008). Spatial neglect: clinical and neuroscience review: a wealth of information on the poverty of spatial attention. Ann. N. Y. Acad. Sci. 1142, 21–43. doi: 10.1196/annals.1444.008
Ang, K. K., and Guan, C. (2017). EEG-based strategies to detect motor imagery for control and rehabilitation. IEEE Trans. Neural Syst. Rehabil. Eng. 25, 392–401. doi: 10.1109/TNSRE.2016.2646763
Azouvi, P., Jacquin-Courtois, S., and Luauté, J. (2016). Rehabilitation of unilateral neglect: evidence-based medicine. Ann. Phys. Rehabil. Med. doi: 10.1016/j.rehab.2016.10.006 [Epub ahead of print].
Baldassarre, A., Ramsey, L., Hacker, C. L., Callejas, A., Astafiev, S. V., Metcalf, N. V., et al. (2014). Large-scale changes in network interactions as a physiological signature of spatial neglect. Brain 137, 3267–3283. doi: 10.1093/brain/awu297
Bartolomeo, P., Thiebaut de Schotten, M., and Chica, A. B. (2012). Brain networks of visuospatial attention and their disruption in visual neglect. Front. Hum. Neurosci. 6:110. doi: 10.3389/fnhum.2012.00110
Battelli, L., Cavanagh, P., Intriligator, J., Tramo, M. J., Hénaff, M. A., Michèl, F., et al. (2001). Unilateral right parietal damage leads to bilateral deficit for high-level motion. Neuron 32, 985–995. doi: 10.1016/S0896-6273(01)00536-0
Birbaumer, N. (2006). Breaking the silence: brain-computer interfaces (BCI) for communication and motor control. Psychophysiology 43, 517–532. doi: 10.1111/j.1469-8986.2006.00456.x
Bowen, A., Hazelton, C., Pollock, A., and Lincoln, N. B. (2013). Cognitive rehabilitation for spatial neglect following stroke. Cochrane Database Syst. Rev. 7:CD003586. doi: 10.1002/14651858.CD003586.pub3
Brunner, C., Birbaumer, N., Blankertz, B., Guger, C., Kübler, A., Mattia, D., et al. (2015). BNCI horizon 2020: towards a roadmap for the BCI community. Brain Comput. Interfaces 2, 1–10. doi: 10.1080/2326263X.2015.1008956
Buxbaum, L. J., Ferraro, M. K., Veramonti, T., Farne, A., Whyte, J., Ladavas, E., et al. (2004). Hemispatial neglect: subtypes, neuroanatomy, and disability. Neurology 62, 749–756. doi: 10.1212/01.WNL.0000113730.73031.F4
Corbetta, M., Kincade, M. J., Lewis, C., Snyder, A. Z., and Sapir, A. (2005). Neural basis and recovery of spatial attention deficits in spatial neglect. Nat. Neurosci. 8, 1603–1610. doi: 10.1038/nn1574
Daly, J. J., and Wolpaw, J. R. (2008). Brain-computer interfaces in neurological rehabilitation. Lancet Neurol. 7, 1032–1043. doi: 10.1016/S1474-4422(08)70223-0
de Munck, J. C., Gonçalves, S. I., Huijboom, L., Kuijer, J. P., Pouwels, P. J., Heethaar, R. M., et al. (2007). The hemodynamic response of the alpha rhythm: an EEG/fMRI study. Neuroimage 35, 1142–1151. doi: 10.1016/j.neuroimage.2007.01.022
Galán, F., Ferrez, P. W., Oliva, F., Guardia, J., and Millán, J. d. R. (2007). “Feature extraction for multi-class BCI using canonical variates analysis,” in Proceedings of the IEEE International Symposium on Intelligent Signal Processing, Alcalá de Henares, 1–6. doi: 10.1109/WISP.2007.4447615
Grosse-Wentrup, M., Mattia, D., and Oweiss, K. (2011). Using brain-computer interfaces to induce neural plasticity and restore function. J. Neural Eng. 8:25004. doi: 10.1088/1741-2560/8/2/025004
He, B. J., Snyder, A. Z., Vincent, J. L., Epstein, A., Shulman, G. L., and Corbetta, M. (2007). Breakdown of functional connectivity in frontoparietal networks underlies behavioral deficits in spatial neglect. Neuron 53, 905–918. doi: 10.1016/j.neuron.2007.02.013
Heilman, K. M., Watson, R. T., and Valenstein, E. (2003). “Neglect and related disorders,” in Clinical Neuropsychology, 4th Edn, eds K. M. Heilman and E. Valenstein (Oxford: Oxford University Press), 243–293.
Jehkonen, M., Laihosalo, M., and Kettunen, J. E. (2006). Impact of neglect on functional outcome after stroke: a review of methodological issues and recent research findings. Restor. Neurol. Neurosci. 24, 209–215.
Kaiser, V., Daly, I., Pichiorri, F., Mattia, D., Müller-Putz, G. R., and Neuper, C. (2012). Relationship between electrical brain responses to motor imagery and motor impairment in stroke. Stroke 43, 2735–2740. doi: 10.1161/STROKEAHA.112.665489
Katz, N., Hartman-Maeir, A., Ring, H., and Soroker, N. (1999). Functional disability and rehabilitation outcome in right hemisphere damaged patients with and without unilateral spatial neglect. Arch. Phys. Med. Rehabil. 80, 379–384. doi: 10.1016/S0003-9993(99)90273-3
Kerkhoff, G., and Schenk, T. (2012). Rehabilitation of neglect: an update. Neuropsychologia 50, 1072–1079. doi: 10.1016/j.neuropsychologia.2012.01.024
Kinsbourne, M. (1993). “Orientational bias model of unilateral neglect: evidence from attentional gradients within hemispace,” in Unilateral Neglect: Clinical and Experimental Studies (Brain Damage, Behaviour and Cognition), eds J. Marshall and I. Robertson (Hove: Psychology Press), 63–86.
Klimesch, W., Doppelmayr, M., Russegger, H., Pachinger, T., and Schwaiger, J. (1998). Induced alpha band power changes in the human EEG and attention. Neurosci. Lett. 244, 73–76.
Klimesch, W., Doppelmayr, M., Schimke, H., and Pachinger, T. (1996). Alpha frequency, reaction time, and the speed of processing information. J. Clin. Neurophysiol. 13, 511–518.
Kortte, K. B., and Hillis, A. E. (2011). Recent trends in rehabilitation interventions for visual neglect and anosognosia for hemiplegia following right hemisphere stroke. Fut. Neurol. 6, 33–43. doi: 10.2217/fnl.10.79
Leeb, R., Tonin, L., Rohm, M., Desideri, L., Carlson, T., and Millan, J. d. R. (2015). Towards independence: a BCI telepresence robot for people with severe motor disabilities. Proc. IEEE 103, 969–982. doi: 10.1109/JPROC.2015.2419736
Marchetti, M., Piccione, F., Silvoni, S., Gamberini, L., and Priftis, K. (2013). Covert visuospatial attention orienting in a brain-computer interface for amyotrophic lateral sclerosis patients. Neurorehabil. Neural Repair 27, 430–438. doi: 10.1177/1545968312471903
Measso, G., Cavarzeran, F., Zappalà, G., Lebowitz, B. D., Crook, T. H., Pirozzolo, F. J., et al. (1993). The mini-mental state examination: normative study of an Italian random sample. Dev. Neuropsychol. 9, 77–85. doi: 10.1080/87565649109540545
Monge-Pereira, E., Ibañez-Pereda, J., Alguacil-Diego, I. M., Serrano, J. I., Spottorno-Rubio, M. P., and Molina-Rueda, F. (2017). Use of electroencephalography brain computer interface systems as a rehabilitative approach for upper limb function after a stroke. A systematic review. Am. Acad. Phys. Med. Rehabil. doi: 10.1016/j.pmrj.2017.04.016 [Epub ahead of print].
Müri, R. M., Cazzoli, D., Nef, T., Mosimann, U. P., Hopfner, S., and Nyffeler, T. (2013). Non-invasive brain stimulation in neglect rehabilitation: an update. Front. Hum. Neurosci. 7:248. doi: 10.3389/fnhum.2013.00248
Nenert, R., Viswanathan, S., Dubuc, D. M., and Visscher, K. M. (2012). Modulations of ongoing alpha oscillations predict successful short-term visual memory encoding. Front. Hum. Neurosci. 6:127. doi: 10.3389/fnhum.2012.00127
Okazaki, Y. O., Horschig, J. M., Luther, L., Oostenveld, R., Murakami, I., and Jensen, O. (2014). Real-time MEG neurofeedback training of posterior alpha activity modulates subsequent visual detection performance. Neuroimage 107, 323–332. doi: 10.1016/j.neuroimage.2014.12.014
Pichiorri, F., Morone, G., Petti, M., Toppi, J., Pisotta, I., Molinari, M., et al. (2015). Brain-computer interface boosts motor imagery practice during stroke recovery. Ann. Neurol. 77, 851–865. doi: 10.1002/ana.24390
Prasad, G., Herman, P., Coyle, D., McDonough, S., and Crosbie, J. (2010). Applying a brain-computer interface to support motor imagery practice in people with stroke for upper limb recovery: a feasibility study. J. Neuroeng. Rehabil. 7:60. doi: 10.1186/1743-0003-7-60
Priftis, K., Passarini, L., Pilosio, C., Meneghello, F., and Pitteri, M. (2013). Visual scanning training, limb activation treatment, and prism adaptation for rehabilitating left neglect: who is the winner? Front. Hum. Neurosci. 7:360. doi: 10.3389/fnhum.2013.00360
Ramos-Murguialday, A., Broetz, D., Rea, M., Läer, L., Yilmaz, O., Brasil, F. L., et al. (2013). Brain-machine interface in chronic stroke rehabilitation: a controlled study. Ann. Neurol. 74, 100–108. doi: 10.1002/ana.23879
Ramsey, L. E., Siegel, J. S., Baldassarre, A., Metcalf, N. V., Zinn, K., Shulman, G. L., et al. (2016). Normalization of network connectivity in hemispatial neglect recovery. Ann. Neurol. 80, 127–141. doi: 10.1002/ana.24690
Remsik, A., Young, B., Vermilyea, R., Kiekoefer, L., Abrams, J., Elmore, S. E., et al. (2017). A review of the progression and future implications of brain-computer interface therapies for restoration of distal upper extremity motor function after stroke. Expert Rev. Med. Devices 13, 445–454. doi: 10.1080/17434440.2016.1174572.A
Riestra, A. R., and Barrett, A. M. (2013). Rehabilitation of spatial neglect. Handb. Clin. Neurol. 110, 347–355. doi: 10.1016/B978-0-444-52901-5.00029-0
Robineau, F., Rieger, S. W., Mermoud, C., Pichon, S., Koush, Y., Van De Ville, D., et al. (2014). Self-regulation of inter-hemispheric visual cortex balance through real-time fmri neurofeedback training. Neuroimage 100, 1–14. doi: 10.1016/j.neuroimage.2014.05.072
Saj, A., Cojan, Y., Vocat, R., Luauté, J., and Vuilleumier, P. (2013). Prism Adaptation Enhances Activity of Intact Fronto-Parietal Areas in Both Hemispheres in Neglect Patients. Cortex 49, 107–119. doi: 10.1016/j.cortex.2011.10.009
Sellers, E. W., Ryan, D. B., and Hauser, C. K. (2014). Noninvasive brain-computer interface enables communication after brainstem stroke. Sci. Transl. Med. 6, 257re7. doi: 10.1126/scitranslmed.3007801
Thut, G., Nietzel, A., Brandt, S. A., and Pascual-Leone, A. (2006). Alpha-band electroencephalographic activity over occipital cortex indexes visuospatial attention bias and predicts visual target detection. J. Neurosci. 26, 9494–9502. doi: 10.1523/JNEUROSCI.0875-06.2006
Tonin, L., Leeb, R., and Millán, J. d. R. (2012). Time-dependent approach for single trial classification of covert visuospatial attention. J. Neural Eng. 9:45011. doi: 10.1088/1741-2560/9/4/045011
Tonin, L., Leeb, R., Sobolewski, A., and Millán, J. d. R. (2013). An online EEG BCI based on covert visuospatial attention in absence of exogenous stimulation. J. Neural Eng. 10:56007. doi: 10.1088/1741-2560/10/5/056007
Vallar, G., Rusconi, M. L., Fontana, S., and Musicco, M. (1994). Tre test Di esplorazione visuo-spaziale: taratura Su 212 soggetti normali. Arch. Psicol. Neurol. Psichiatr. 55, 827–841.
Wilson, B. A., Cockburn, J., and Halligan, P. (1987). Behavioural Inattention Test: Manual. Bury St Edmunds: Thames Valley Test Company.
Young, B. M., Nigogosyan, Z., Walton, L. M., Song, J., Nair, V. A., Grogan, S. W., et al. (2014). Changes in functional brain organization and behavioral correlations after rehabilitative therapy using a brain-computer interface. Front. Neuroeng. 7:26. doi: 10.3389/fneng.2014.00026
Keywords: brain-computer interface, spatial neglect, covert visuospatial attention, electroencephalogram, alpha oscillations, functional connectivity
Citation: Tonin L, Pitteri M, Leeb R, Zhang H, Menegatti E, Piccione F and Millán JdR (2017) Behavioral and Cortical Effects during Attention Driven Brain-Computer Interface Operations in Spatial Neglect: A Feasibility Case Study. Front. Hum. Neurosci. 11:336. doi: 10.3389/fnhum.2017.00336
Received: 05 May 2017; Accepted: 12 June 2017;
Published: 28 June 2017.
Edited by:
Klaus Gramann, Technische Universität Berlin, GermanyReviewed by:
Brendan Z. Allison, University of California, San Diego, United StatesNoman Naseer, Air University, Pakistan
Copyright © 2017 Tonin, Pitteri, Leeb, Zhang, Menegatti, Piccione and Millán. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) or licensor are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Luca Tonin, bHVjYS50b25pbkBlcGZsLmNo José del R. Millán, am9zZS5taWxsYW5AZXBmbC5jaA==