 Andrés Hernández*
Andrés Hernández* Arthur Cheng and Håkan Westerblad
Arthur Cheng and Håkan Westerblad
- Department of Physiology and Pharmacology, Karolinska Institutet, Stockholm, Sweden
Antioxidants are assumed to provide numerous benefits, including better health, a reduced rate of aging, and improved exercise performance. Specifically, antioxidants are commonly “prescribed” by the media, supplement industry, and “fitness experts” for individuals prior to training and performance, with assumed benefits of improved fatigue resistance and recovery. This has provoked expansion of the supplement industry which responded by creation of a plethora of products aimed at facilitating the needs of the active individual. However, what does the experimental evidence say about the efficacy of antioxidants on skeletal muscle function? Are antioxidants actually as beneficial as the general populous believes? Or, could they in fact lead to deleterious effects on skeletal muscle function and performance? This Mini Review addresses these questions with an unbiased look at what we know about antioxidant effects on skeletal muscle, and what we still need to know before conclusions can be made.
Introduction
Commoner et al. (1954) reported that reactive oxygen species (ROS) intermediates were present in a wide range of animal tissues, including whole blood, brain, liver, and muscle. They proposed that ROS production was related to metabolic activity. It was later found that exercise results in elevated ROS, and that endurance was reduced by ∼40% in vitamin E deficient rats (Davies et al., 1982). The authors surmised that the peroxidative damage induced by ROS in the absence of vitamin E was responsible for the reduced exercise performance. Accordingly, a multitude of scientific investigations were launched to look at the effects of ROS, and also reactive nitrogen species (RNS), in relation to physical exercise and skeletal muscle fatigue (for in depth review, see Ferreira and Reid, 2008; Powers and Jackson, 2008; Westerblad and Allen, 2011).
It has become “common knowledge” that ROS generated during exercise are bad, and usage of antioxidant supplements to ameliorate their effects promotes health. The various makers of dietary supplements have taken full advantage of this phenomenon by including antioxidants in their supplements; or as concentrated products. However, experimental evidence shows that increased ROS production is not necessarily bad: ROS are important for a wide range of normal exercise-related physiological processes, including a role in contraction-mediated glucose uptake (e.g., Sandström et al., 2006) and promotion of the adaptive responses to training (Ristow et al., 2009). Accordingly, the use of antioxidants has been shown to blunt training responses (Ristow et al., 2009; Petersen et al., 2011; Strobel et al., 2011). But what does scientific evidence say about antioxidant usage prior to a single exercise bout? Do performance and recovery become enhanced and, if so, what are the underlying mechanisms? In this Mini Review we will survey potential effects on performance and recovery of antioxidants frequently used in association with physical exercise. We specifically focus on whether effects seen in the exercising human can be explained by effects observed in experiments on isolated muscle or muscle fibers. For more detailed descriptions of properties of different ROS and RNS that may increase during physical exercise and endogenous antioxidant systems we refer to more comprehensive reviews (Dröge, 2002; Powers and Jackson, 2008; Westerblad and Allen, 2011).
Antioxidant Supplementation and Muscle Fatigue
Ubiquinone-10
Ubiquinone-10 is a lipid soluble antioxidant found in high concentrations in meat and fish (Powers et al., 2004). Concentrated ubiquinone-10 supplements are readily available. Early evidence indicated intramuscular ubiquinone-10 content had a positive relationship with exercise capacity (Karlsson et al., 1996). However, the greater exercise capacity was more likely a function of decreased fatigability based on the muscle properties (e.g., oxidative capacity) and not ubiquinone-10 content. Whereas supplementation with ubiquinone-10 may provide assistance to individuals with mitochondrial disease (Glover et al., 2010), most investigations on healthy individuals show no effect (Braun et al., 1991; Mizuno et al., 1997; Weston et al., 1997; Bonetti et al., 2000) or a deleterious effect (Laaksonen et al., 1995; Malm et al., 1997) on exercise performance. The lone exception is a recent study with a dose ∼3× that used in previous studies (Mizuno et al., 2008). Thus at present time, there is not enough evidence to support a role for ubiquinone-10 as an antioxidant having an ergogenic effect in healthy individuals.
Vitamins C and E
Vitamin C is hydrophilic and widely distributed in mammalian tissues. It can act as a radical scavenger and recycles vitamin E (Powers et al., 2004; Powers and Jackson, 2008). Vitamin E is lipid soluble and the major chain-breaking antioxidant found in cell membranes (Powers et al., 2004; Powers and Jackson, 2008). These two vitamins are “expected” to improve exercise performance based on their antioxidant properties and are commonly used by athletes and active individuals. However, experimental evidence to support beneficial effects on physical performance does not exist. Neither vitamin C (Clarkson, 1995; Ashton et al., 1999) nor vitamin E supplementation (Shephard et al., 1974; Lawrence et al., 1975; Sumida et al., 1989; Rokitzki et al., 1994a,b; Bryant et al., 2003; Gaeini et al., 2006) improves exercise performance in humans. Further, no beneficial effects have been observed with the combination of vitamins C and E (Bryant et al., 2003), or vitamins C and E with ubiquinone-10 (Nielsen et al., 1999). Thus, claims as to the efficacy of vitamins C and E to improve exercise performance are without experimental support.
N-Acetylcysteine
The antioxidant N-acetylcysteine (NAC) got its start in the 1990s and has grown in popularity, now being readily available for daily use. NAC easily enters cells and contains a thiol group that can interact with ROS, RNS, and their derivatives (Aruoma et al., 1989; Dekhuijzen, 2004; Ferreira and Reid, 2008). As a thiol donor, NAC also supports resynthesis of one of the major endogenous antioxidant systems, glutathione (Dekhuijzen, 2004). The first report of beneficial effects of antioxidant supplementation on fatigue in humans came after NAC infusion (Reid et al., 1994). NAC was infused into the subjects for 1 h prior to low-(10 Hz) and high-frequency (40 Hz) stimulation of the tibialis anterior muscle. NAC infusion resulted in significantly less fatigue during 10-Hz stimulation; but not during 40-Hz stimulation. These data indicated two potential, important features of NAC supplementation: (1) fatigue can be reduced by NAC supplementation; and (2) the effect depends on the exercise protocol in that the effect is larger with submaximal contractions. Accordingly, a later study showed a beneficial effect of NAC during fatigue induced by repetitive submaximal handgrip exercise but not during maximal contractions (Matuszczak et al., 2005). The specific effect of NAC on submaximal contractile force has also been extended to cycling exercise (Medved et al., 2003, 2004a,b; McKenna et al., 2006; Corn and Barstow, 2011).
NAC has been shown to have beneficial effects on contractility and fatiguability of human ventilatory muscles (Travaline et al., 1997; Kelly et al., 2009). Using the murine diaphragm contracting in situ, Shindoh et al. (1990) measured a beneficial effect of NAC on fatigue resistance. They speculated that the mechanism of action could be through NAC effects on blood flow or directly on the muscle fibers themselves. Similar effects on fatigue resistance in the diaphragm have been reported by other groups (Diaz et al., 1994; Khawli and Reid, 1994; Supinski et al., 1997). Results from isolated diaphragm strips contracting in vitro indicate that the effects of NAC on fatigue resistance are at the muscle fiber level (Diaz et al., 1994; Khawli and Reid, 1994). Furthermore, using diaphragm bundles contracting in vitro, Mishima et al. (2005) reported less fatigue in fibers treated with NAC and this effect was independent of changes in sarcoplasmic reticulum (SR) Ca2+ release and uptake.
Mechanisms by Which ROS/RNS may Affect Fatigue
Proposed mechanisms intrinsic to the muscle fibers by which ROS/RNS can accelerate fatigue development include: (1) reduced membrane excitability, (2) impaired SR Ca2+ release, (3) inhibition of SR Ca2+-ATPase (SERCA), and (4) deleterious effects on myofibrillar function. Accordingly, antioxidants such as NAC may enhance fatigue resistance by hindrance of any of these proposed effects. NAC supplementation increased the time to fatigue in humans during submaximal cycling exercise and analyses of muscle biopsies suggest that the improved performance could be due to preserved function of Na+-K+ ATPase (McKenna et al., 2006). This indicates that ROS may accelerate fatigue development by impairing membrane excitability. However, studies on isolated intact muscle fibers do not show any evidence of action potential failure induced by exposure to ROS either in the unfatigued state (Andrade et al., 1998a, 2001) or during fatiguing stimulation (Place et al., 2009).
Results from experiments with intact single fast- and slow-twitch fibers from limb muscles do not support a role for ROS in decreasing SR Ca2+ release during high-intensity fatiguing stimulation (Moopanar and Allen, 2005; Bruton et al., 2008). For example, SR Ca2+ release, and hence contractile force (Figure 1), can be well maintained even when fatigue is induced in the presence of a high concentration of the ROS hydrogen peroxide (10 μM) and at high temperature (43°C; Place et al., 2009). Thus, these studies do not support an ability of antioxidants to prevent the reductions in SR Ca2+ release that occur during fatigue. Accordingly, if effects are seen, antioxidant supplementation must exert its beneficial effects on exercise performance via some other mechanism.
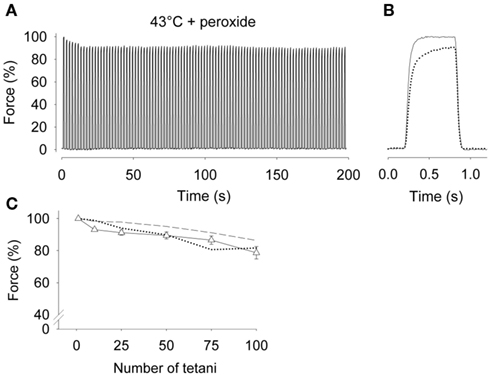
Figure 1. Tetanic force was well maintained in intact soleus fibers during fatiguing stimulation at 43°C in the presence of peroxide. (A) Typical continuous force records from a soleus fiber fatigued by 100 Hz, 600-ms tetanic contractions repeated every 2 s at 43°C in the presence of 10 μM hydrogen peroxide. Force is expressed relative to the first tetanus, which was set to 100%. (B) Superimposed force records on an expanded time axis from the first (solid) and last (dotted line) tetani of the fatigue run. (C) Mean data (±SEM) of relative force measured during the 1st, 10th, 25th, 50th, 75th, and 100th fatiguing tetani at 43°C in the presence of 10 μM hydrogen peroxide (Δ, n = 9). For comparison, mean data from soleus fibers fatigued at 37°C (dashed line) and 43°C (dotted line) in the absence of peroxide are also shown. Fatigue in fast-twitch fibers was unaffected by elevated temperature. Contractile force in rested fibers was unaffected by 5 min of 10 μM hydrogen peroxide exposure, i.e., 100% force did not differ between groups. Data are from Place et al. (2009).
The changes occurring during fatiguing stimulation of skeletal muscle fibers often include an elevation of baseline [Ca2+]i, which can be due to impaired SERCA function (Westerblad and Allen, 1991, 1993). Studies on muscle biopsies taken after exercise in humans have shown impaired SR Ca2+ uptake into the SR (Booth et al., 1997; Duhamel et al., 2007). Scherer and Deamer (1986) found that administration of oxidants to SR microsomes reduced SERCA function and Ca2+ transport. Moreover, prolonged exposure to high concentrations of oxidants resulted in impaired SR Ca2+ uptake and an elevation in baseline [Ca2+]i in isolated intact muscle fibers (Andrade et al., 1998a, 2001). Accordingly, antioxidant supplementations may improve performance by preventing adverse effects on SERCA function, but we are not aware of any studies where this potential mechanism has been shown to occur.
Impairment in the ability of the contractile elements to respond to Ca2+ (myofibrillar Ca2+ sensitivity) is a common feature of fatigue (Allen et al., 2008). In this case, force can become depressed in the absence of any impairment of SR Ca2+ handling. Prolonged exposure to oxidants reduces myofibrillar Ca2+ sensitivity in unfatigued fibers (Andrade et al., 1998a, 2001). In addition, Andrade et al. (1998b) showed that nitric oxide (NO) donors reduce myofibrillar Ca2+ sensitivity in unfatigued fast-twitch fibers. Pye et al. (2007) used dissociated fast-twitch skeletal muscle fibers from mice and fluorescent indicator to measure NO production during contractions and observed a marked increase after 5 min of contractions. Accordingly, the production of NO, leading to the production of RNS in contracting skeletal muscle, may contribute to the decrease in myofibrillar Ca2+ sensitivity during fatiguing stimulation. Thus, NAC and other antioxidant treatments could potentially enhance fatigue resistance by counteracting any ROS/RNS-induced decrease in myofibrillar Ca2+ sensitivity.
Taken together, the experimental evidence regarding a positive role for antioxidant supplementation during exercise indicates that, whereas diaphragm muscle fibers display beneficial effects, these effects have not been observed in muscles involved in locomotion. Accordingly, the positive effects of NAC observed in experiments on exercising humans seem not to be due to direct antioxidant effects on limb muscle fibers.
Antioxidants and Recovery
Depending on the nature of exercise, the time for recovery may vary between minutes to days. An increased rate of recovery is beneficial, e.g., by allowing bouts of exercise to be performed at short intervals. In this section, we will discuss the role of ROS in the recovery process and whether antioxidants can help improve recovery of force.
In humans, there is a rapid rate of recovery of maximum voluntary contraction force (Baker et al., 1993; Allman and Rice, 2001), and force at high frequencies (50–100 Hz) of electrical stimulation within the first 5 min after fatigue (Edwards et al., 1977; Allman and Rice, 2001). However, force is not fully restored by 5 min, and it can take several hours to recover maximal, and especially submaximal, force generating capacity (Edwards et al., 1977; Allman and Rice, 2001; Hill et al., 2001).
Edwards et al. (1977) were the first to report a pronounced delay in the recovery of force at low stimulation frequencies (10–20 Hz) lasting several hours or even days after repeated voluntary isometric contractions performed in humans. This delayed recovery was initially named “low-frequency fatigue,” but this name has unfortunately been used to describe many different situations with decreased force production and therefore it was recently re-defined as prolonged low-frequency force depression (PLFFD; Allen et al., 2008; Bruton et al., 2008; Lamb and Westerblad, 2011; Westerblad and Allen, 2011). The slow recovery of force at low frequencies could explain the sensation of muscle weakness at submaximal levels of voluntary effort that appears to last for a similar duration as PLFFD. Results from isolated muscles of rodents have determined that the primary mechanisms causing PLFFD in fast-twitch fibers are decreased SR Ca2+ release and decreased Ca2+ sensitivity (Bruton et al., 2008). In intact muscle fibers of wild-type mice it appears that the ROS superoxide induces impairments in SR Ca2+ release that can explain PLFFD in intact single fibers (Bruton et al., 2008). In contrast, a similar PLFFD was observed in intact single fibers of wild-type rats, but in this case it was due to decreased Ca2+ sensitivity (Bruton et al., 2008). Superoxide dismutase (SOD) converts superoxide into hydrogen peroxide, and hydrogen peroxide exposure has been shown to decrease myofibrillar Ca2+ sensitivity in rested fibers even at very low concentrations in muscle (Andrade et al., 2001; Murphy et al., 2008), and blunt the recovery of 50 Hz force in fatigued amphibian single fibers (Oba et al., 2002). Wild-type rat muscles displayed higher SOD activity and thus would produce more hydrogen peroxide during fatigue than wild-type mouse fibers (Bruton et al., 2008). Furthermore, Figure 2 shows a similar PLFFD in muscle fibers of wild-type and SOD2 overexpressing mice, but the underlying mechanism differs: in wild-type fibers, where superoxide would dominate, PLFFD is due to decreased SR Ca2+ release, whereas in SOD2 overexpressing fibers, where hydrogen peroxide would dominate, PLFFD is due to decreased myofibrillar Ca2+ sensitivity (Bruton et al., 2008). Decreased myofibrillar Ca2+ sensitivity might be related to ROS-induced oxidation of the contractile proteins since 0.5–1 mM dithiothreitol, a non-reversible reducing agent, has been shown to restore low frequency force in intact rat (Diaz et al., 1998; Bruton et al., 2008) and mouse intact skeletal muscle (Moopanar and Allen, 2006) without affecting SR Ca2+ release (Moopanar and Allen, 2006; Bruton et al., 2008).
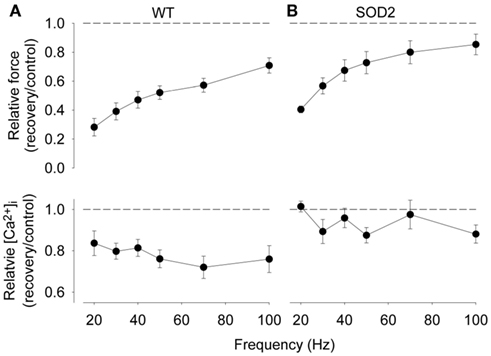
Figure 2. Both wild-type (WT) and superoxide dismutase 2 (SOD2) overexpressing fibers display marked PLFFD but the underlying mechanism differs. Mean data (±SEM) of the relative change in tetanic force (top) and [Ca2+]i (bottom) obtained in mouse WT (A) and SOD2 overexpressing (B) fibers (n = 4). Relative changes were calculated as ratio 30 min after (recovery) to before (control) fatiguing stimulation; dashed lines indicate no change. Contractile force and tetanic [Ca2+]i before fatigue did not differ between WT and SOD2 overexpressing fibers. Data are from Bruton et al. (2008).
To sum up, there is clear-cut experimental evidence supporting important effects of oxidants generated during fatiguing contractions on the recovery process. However, there are also many puzzling results in this respect. For instance, studies have shown that only the antioxidant NAC attenuates the low-frequency force decline during fatigue (Shindoh et al., 1990; Reid et al., 1994), but a consistent finding is that NAC does not improve the recovery after fatigue (Shindoh et al., 1990; Reid et al., 1994; Bruton et al., 2008). Thus, many questions need to be addressed in future investigations of the recovery processes, including the need to identify the specific site of action of ROS, and the extent to which different ROS are potentially responsible for the prolonged impairment in force during recovery.
Conclusion
Experimental evidence does not support the “common knowledge” that antioxidant treatment greatly improves exercise performance and recovery. On the contrary, studies with antioxidant supplementations generally show no effect on muscle function during and after exercise. The exception is NAC treatment, which has been found to improve performance during submaximal exercise. The limited effects of ROS/RNS and antioxidants during exercise are unexpected in that increases in ROS/RNS are likely to occur and these are potentially harmful. It appears that muscle fibers are in some way protected against deleterious effects of oxidants during exercise and fibers are generally much more sensitive to exposure to oxidants in the rested state than during fatigue. For instance, experiments on single mouse muscle fibers have shown that application of 10 μM hydrogen peroxide did not affect fatigue development (Place et al., 2009), whereas concentrations as low as 100 pM hydrogen peroxide affected contraction and Ca2+ handling in rested fibers (Andrade et al., 2001). Thus, numerous questions remain to be answered in relation to the effects of oxidants during and after exercise. In order to do this, improved methods to measure ROS/RNS are essential, since the effects of these highly reactive substances are likely to strongly depend on both their temporal and spatial distribution. Furthermore, the effects are likely to show marked differences between slow-twitch fatigue resistant and fast-twitch easily fatigued muscle fibers due to a major difference in ROS/RNS production, endogenous oxidant levels and sensitivity of Ca2+ handling and contractile properties.
Conflict of Interest Statement
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Acknowledgments
The authors acknowledge support from the Swedish Research Council, the Swedish Center for Sports Research, Association Francaise Contre les Myopathies (AFM), and the National Institute of Arthritis and Musculoskeletal and Skin Diseases (NIAMS) grant 1F32AR057619-01A1 (to Andrés Hernández).
References
Allen, D. G., Lamb, G. D., and Westerblad, H. (2008). Skeletal muscle fatigue: cellular mechanisms. Physiol. Rev. 88, 287–332.
Allman, B. L., and Rice, C. L. (2001). Incomplete recovery of voluntary isometric force after fatigue is not affected by old age. Muscle Nerve 24, 1156–1167.
Andrade, F. H., Reid, M. B., Allen, D. G., and Westerblad, H. (1998a). Effect of hydrogen peroxide and dithiothreitol on contractile function of single skeletal muscle fibres from the mouse. J. Physiol. (Lond.) 509, 565–575.
Andrade, F. H., Reid, M. B., Allen, D. G., and Westerblad, H. (1998b). Effect of nitric oxide on single skeletal muscle fibres from the mouse. J. Physiol. (Lond.) 509, 577–586.
Andrade, F. H., Reid, M. B., and Westerblad, H. (2001). Contractile response of skeletal muscle to low peroxide concentrations: myofibrillar calcium sensitivity as a likely target for redox-modulation. FASEB J. 15, 309–311.
Aruoma, O. I., Halliwell, B., Hoey, B. M., and Butler, J. (1989). The antioxidant action of N-acetylcysteine: its reaction with hydrogen peroxide, hydroxyl radical, superoxide, and hypochlorous acid. Free Raic. Biol. Med. 6, 593–597.
Ashton, T., Young, I. S., Peters, J. R., Jones, E., Jackson, S. K., Davies, B., and Rowlands, C. C. (1999). Electron spin resonance spectroscopy, exercise, and oxidative stress: an ascorbic acid intervention study. J. Appl. Physiol. 87, 2032–2036.
Baker, A. J., Kostov, K. G., Miller, R. G., and Weiner, M. W. (1993). Slow force recovery after long-duration exercise: metabolic and activation factors in muscle fatigue. J. Appl. Physiol. 74, 2294–2300.
Bonetti, A., Solito, F., Carmosino, G., Bargossi, A. M., and Fiorella, P. L. (2000). Effect of ubidecarenone oral treatment on aerobic power in middle-aged trained subjects. J. Sports Med. Phys. Fitness 40, 51–57.
Booth, J., Mckenna, M. J., Ruell, P. A., Gwinn, T. H., Davies, G. M., Thompson, M. W., Harmer, A. R., Hunter, S. K., and Sutton, J. R. (1997). Impaired calcium pump function does not slow relaxation in human skeletal muscle after prolonged exercise. J. Appl. Physiol. 83, 511–521.
Braun, B., Clarkson, P. M., Freedson, P. S., and Kohl, R. L. (1991). Effects of coenzyme Q10 supplementation on exercise performance, VO2max, and lipid peroxidation in trained cyclists. Int. J. Sport Nutr. 1, 353–365.
Bruton, J. D., Place, N., Yamada, T., Silva, J. P., Andrade, F. H., Dahlstedt, A. J., Zhang, S. J., Katz, A., Larsson, N. G., and Westerblad, H. (2008). Reactive oxygen species and fatigue-induced prolonged low-frequency force depression in skeletal muscle fibres of rats, mice and SOD2 overexpressing mice. J. Physiol. (Lond.) 586, 175–184.
Bryant, R. J., Ryder, J., Martino, P., Kim, J., and Craig, B. W. (2003). Effects of vitamin E and C supplementation either alone or in combination on exercise-induced lipid peroxidation in trained cyclists. J. Strength Cond. Res. 17, 792–800.
Clarkson, P. M. (1995). Antioxidants and physical performance. Crit. Rev. Food Sci. Nutr. 35, 131–141.
Commoner, B., Townsend, J., and Pake, G. E. (1954). Free radicals in biological materials. Nature 174, 689–691.
Corn, S. D., and Barstow, T. J. (2011). Effects of oral N-acetylcysteine on fatigue, critical power, and W’ in exercising humans. Respir. Physiol. Neurobiol. 178, 261–268.
Davies, K. J., Quintanilha, A. T., Brooks, G. A., and Packer, L. (1982). Free radicals and tissue damage produced by exercise. Biochem. Biophys. Res. Commun. 107, 1198–1205.
Dekhuijzen, P. N. (2004). Antioxidant properties of N-acetylcysteine: their relevance in relation to chronic obstructive pulmonary disease. Eur. Respir. J. 23, 629–636.
Diaz, P. T., Brownstein, E., and Clanton, T. L. (1994). Effects of N-acetylcysteine on in vitro diaphragm function are temperature dependent. J. Appl. Physiol. 77, 2434–2439.
Diaz, P. T., Costanza, M. J., Wright, V. P., Julian, M. W., Diaz, J. A., and Clanton, T. L. (1998). Dithiothreitol improves recovery from in vitro diaphragm fatigue. Med. Sci. Sports Exerc. 30, 421–426.
Dröge, W. (2002). Free radicals in the physiological control of cell function. Physiol. Rev. 82, 47–95.
Duhamel, T. A., Stewart, R. D., Tupling, A. R., Ouyang, J., and Green, H. J. (2007). Muscle sarcoplasmic reticulum calcium regulation in humans during consecutive days of exercise and recovery. J. Appl. Physiol. 103, 1212–1220.
Edwards, R. H., Hill, D. K., Jones, D. A., and Merton, P. A. (1977). Fatigue of long duration in human skeletal muscle after exercise. J. Physiol. (Lond.) 272, 769–778.
Ferreira, L. F., and Reid, M. B. (2008). Muscle-derived ROS and thiol regulation in muscle fatigue. J. Appl. Physiol. 104, 853–860.
Gaeini, A. A., Rahnama, N., and Hamedinia, M. R. (2006). Effects of vitamin E supplementation on oxidative stress at rest and after exercise to exhaustion in athletic students. J. Sports Med. Phys. Fitness 46, 458–461.
Glover, E. I., Martin, J., Maher, A., Thornhill, R. E., Moran, G. R., and Tarnopolsky, M. A. (2010). A randomized trial of coenzyme Q10 in mitochondrial disorders. Muscle Nerve 42, 739–748.
Hill, C. A., Thompson, M. W., Ruell, P. A., Thom, J. M., and White, M. J. (2001). Sarcoplasmic reticulum function and muscle contractile character following fatiguing exercise in humans. J. Physiol. (Lond.) 531, 871–878.
Karlsson, J., Lin, L., Sylven, C., and Jansson, E. (1996). Muscle ubiquinone in healthy physically active males. Mol. Cell. Biochem. 156, 169–172.
Kelly, M. K., Wicker, R. J., Barstow, T. J., and Harms, C. A. (2009). Effects of N-acetylcysteine on respiratory muscle fatigue during heavy exercise. Respir. Physiol. Neurobiol. 165, 67–72.
Khawli, F. A., and Reid, M. B. (1994). N-acetylcysteine depresses contractile function and inhibits fatigue of diaphragm in vitro. J. Appl. Physiol. 77, 317–324.
Laaksonen, R., Fogelholm, M., Himberg, J. J., Laakso, J., and Salorinne, Y. (1995). Ubiquinone supplementation and exercise capacity in trained young and older men. Eur. J. Appl. Physiol. Occup. Physiol. 72, 95–100.
Lamb, G. D., and Westerblad, H. (2011). Acute effects of reactive oxygen and nitrogen species on the contractile function of skeletal muscle. J. Physiol. (Lond.) 589, 2119–2127.
Lawrence, J. D., Bower, R. C., Riehl, W. P., and Smith, J. L. (1975). Effects of alpha-tocopherol acetate on the swimming endurance of trained swimmers. Am. J. Clin. Nutr. 28, 205–208.
Malm, C., Svensson, M., Ekblom, B., and Sjodin, B. (1997). Effects of ubiquinone-10 supplementation and high intensity training on physical performance in humans. Acta Physiol. Scand. 161, 379–384.
Matuszczak, Y., Farid, M., Jones, J., Lansdowne, S., Smith, M. A., Taylor, A. A., and Reid, M. B. (2005). Effects of N-acetylcysteine on glutathione oxidation and fatigue during handgrip exercise. Muscle Nerve 32, 633–638.
McKenna, M. J., Medved, I., Goodman, C. A., Brown, M. J., Bjorksten, A. R., Murphy, K. T., Petersen, A. C., Sostaric, S., and Gong, X. (2006). N-acetylcysteine attenuates the decline in muscle Na+, K+-pump activity and delays fatigue during prolonged exercise in humans. J. Physiol. (Lond.) 576, 279–288.
Medved, I., Brown, M. J., Bjorksten, A. R., Leppik, J. A., Sostaric, S., and McKenna, M. J. (2003). N-acetylcysteine infusion alters blood redox status but not time to fatigue during intense exercise in humans. J. Appl. Physiol. 94, 1572–1582.
Medved, I., Brown, M. J., Bjorksten, A. R., and McKenna, M. J. (2004a). Effects of intravenous N-acetylcysteine infusion on time to fatigue and potassium regulation during prolonged cycling exercise. J. Appl. Physiol. 96, 211–217.
Medved, I., Brown, M. J., Bjorksten, A. R., Murphy, K. T., Petersen, A. C., Sostaric, S., Gong, X., and McKenna, M. J. (2004b). N-acetylcysteine enhances muscle cysteine and glutathione availability and attenuates fatigue during prolonged exercise in endurance-trained individuals. J. Appl. Physiol. 97, 1477–1485.
Mishima, T., Yamada, T., Matsunaga, S., and Wada, M. (2005). N-acetylcysteine fails to modulate the in vitro function of sarcoplasmic reticulum of diaphragm in the final phase of fatigue. Acta Physiol. Scand. 184, 195–202.
Mizuno, K., Tanaka, M., Nozaki, S., Mizuma, H., Ataka, S., Tahara, T., Sugino, T., Shirai, T., Kajimoto, Y., Kuratsune, H., Kajimoto, O., and Watanabe, Y. (2008). Antifatigue effects of coenzyme Q10 during physical fatigue. Nutrition 24, 293–299.
Mizuno, M., Quistorff, B., Theorell, H., Theorell, M., and Chance, B. (1997). Effects of oral supplementation of coenzyme Q10 on 31P-NMR detected skeletal muscle energy metabolism in middle-aged post-polio subjects and normal volunteers. Mol. Aspects Med. 18(Suppl.), S291–S298.
Moopanar, T. R., and Allen, D. G. (2005). Reactive oxygen species reduce myofibrillar Ca2+ sensitivity in fatiguing mouse skeletal muscle at 37 degrees C. J. Physiol. (Lond.) 564, 189–199.
Moopanar, T. R., and Allen, D. G. (2006). The activity-induced reduction of myofibrillar Ca2+ sensitivity in mouse skeletal muscle is reversed by dithiothreitol. J. Physiol. (Lond.) 571, 191–200.
Murphy, R. M., Dutka, T. L., and Lamb, G. D. (2008). Hydroxyl radical and glutathione interactions alter calcium sensitivity and maximum force of the contractile apparatus in rat skeletal muscle fibres. J. Physiol. (Lond.) 586, 2203–2216.
Nielsen, A. N., Mizuno, M., Ratkevicius, A., Mohr, T., Rohde, M., Mortensen, S. A., and Quistorff, B. (1999). No effect of antioxidant supplementation in triathletes on maximal oxygen uptake, 31P-NMRS detected muscle energy metabolism and muscle fatigue. Int. J. Sports Med. 20, 154–158.
Oba, T., Kurono, C., Nakajima, R., Takaishi, T., Ishida, K., Fuller, G. A., Klomkleaw, W., and Yamaguchi, M. (2002). H2O2 activates ryanodine receptor but has little effect on recovery of releasable Ca2+ content after fatigue. J. Appl. Physiol. 93, 1999–2008.
Petersen, A. C., McKenna, M. J., Medved, I., Murphy, K. T., Brown, M. J., Della Gatta, P., and Cameron-Smith, D. (2011). Infusion with the antioxidant N-acetylcysteine attenuates early adaptive responses to exercise in human skeletal muscle. Acta Physiol. (Oxf.) 204, 382–392.
Place, N., Yamada, T., Zhang, S. J., Westerblad, H., and Bruton, J. D. (2009). High temperature does not alter fatigability in intact mouse skeletal muscle fibres. J. Physiol. (Lond.) 587, 4717–4724.
Powers, S. K., DeRuisseau, K. C., Quindry, J., and Hamilton, K. L. (2004). Dietary antioxidants and exercise. J. Sports Sci. 22, 81–94.
Powers, S. K., and Jackson, M. J. (2008). Exercise-induced oxidative stress: cellular mechanisms and impact on muscle force production. Physiol. Rev. 88, 1243–1276.
Pye, D., Palomero, J., Kabayo, T., and Jackson, M. J. (2007). Real-time measurement of nitric oxide in single mature mouse skeletal muscle fibres during contractions. J. Physiol. (Lond.) 581, 309–318.
Reid, M. B., Stokic, D. S., Koch, S. M., Khawli, F. A., and Leis, A. A. (1994). N-acetylcysteine inhibits muscle fatigue in humans. J. Clin. Invest. 94, 2468–2474.
Ristow, M., Zarse, K., Oberbach, A., Klöting, N., Birringer, M., Kiehntopf, M., Stumvoll, M., Kahn, C. R., and Blüher, M. (2009). Antioxidants prevent health-promoting effects of physical exercise in humans. Proc. Natl. Acad. Sci. U.S.A. 106, 8665–8670.
Rokitzki, L., Logemann, E., Huber, G., Keck, E., and Keul, J. (1994a). Alpha-tocopherol supplementation in racing cyclists during extreme endurance training. Int. J. Sport Nutr. 4, 253–264.
Rokitzki, L., Logemann, E., Sagredos, A. N., Murphy, M., Wetzel-Roth, W., and Keul, J. (1994b). Lipid peroxidation and antioxidative vitamins under extreme endurance stress. Acta Physiol. Scand. 151, 149–158.
Sandström, M. E., Zhang, S. J., Bruton, J., Silva, J. P., Reid, M. B., Westerblad, H., and Katz, A. (2006). Role of reactive oxygen species in contraction-mediated glucose transport in mouse skeletal muscle. J. Physiol. (Lond.) 575, 251–262.
Scherer, N. M., and Deamer, D. W. (1986). Oxidative stress impairs the function of sarcoplasmic reticulum by oxidation of sulfhydryl groups in the Ca2+-ATPase. Arch. Biochem. Biophys. 246, 589–601.
Shephard, R. J., Campbell, R., Pimm, P., Stuart, D., and Wright, G. R. (1974). Vitamin E, exercise, and the recovery from physical activity. Eur. J. Appl. Physiol. Occup. Physiol. 33, 119–126.
Shindoh, C., DiMarco, A., Thomas, A., Manubay, P., and Supinski, G. (1990). Effect of N-acetylcysteine on diaphragm fatigue. J. Appl. Physiol. 68, 2107–2113.
Strobel, N. A., Peake, J. M., Matsumoto, A., Marsh, S. A., Coombes, J. S., and Wadley, G. D. (2011). Antioxidant supplementation reduces skeletal muscle mitochondrial biogenesis. Med. Sci. Sports Exerc. 43, 1017–1024.
Sumida, S., Tanaka, K., Kitao, H., and Nakadomo, F. (1989). Exercise-induced lipid peroxidation and leakage of enzymes before and after vitamin E supplementation. Int. J. Biochem. 21, 835–838.
Supinski, G. S., Stofan, D., Ciufo, R., and DiMarco, A. (1997). N-acetylcysteine administration alters the response to inspiratory loading in oxygen-supplemented rats. J. Appl. Physiol. 82, 1119–1125.
Travaline, J. M., Sudarshan, S., Roy, B. G., Cordova, F., Leyenson, V., and Criner, G. J. (1997). Effect of N-acetylcysteine on human diaphragm strength and fatigability. Am. J. Resp. Crit. Care Med. 156, 1567–1571.
Westerblad, H., and Allen, D. G. (1991). Changes in myoplasmic calcium concentration during fatigue in single mouse muscle fibers. J. Gen. Physiol. 98, 615–635.
Westerblad, H., and Allen, D. G. (1993). The contribution of [Ca2+]i to the slowing of relaxation in fatigued single fibres from mouse skeletal muscle. J. Physiol. (Lond.) 468, 729–740.
Westerblad, H., and Allen, D. G. (2011). Emerging roles of ROS/RNS in muscle function and fatigue. Antioxid. Redox Signal. 15, 2487–2499.
Keywords: muscle, antioxidants, reactive oxygen species, reactive nitrogen species, performance, exercise, recovery, fatigue
Citation: Hernández A, Cheng A and Westerblad H (2012) Antioxidants and skeletal muscle performance: “Common knowledge” vs. experimental evidence. Front. Physio. 3:46. doi: 10.3389/fphys.2012.00046
Received: 13 December 2011; Accepted: 20 February 2012;
Published online: 12 March 2012.
Edited by:
Christina Karatzaferi, University of Thessaly, GreeceReviewed by:
Gerhard Meissner, University of North Carolina at Chapel Hill, USANiels Ørtenblad, University of Southern Denmark, Denmark
Athanasios Jamurtas, University of Thessaly, Greece
Copyright: © 2012 Hernández, Cheng and Westerblad. This is an open-access article distributed under the terms of the Creative Commons Attribution Non Commercial License, which permits non-commercial use, distribution, and reproduction in other forums, provided the original authors and source are credited.
*Correspondence: Andrés Hernández, Department of Physiology and Pharmacology, Karolinska Institutet, SE-171 77 Stockholm, Sweden. e-mail:YW5kcmVzLmhlcm5hbmRlekBraS5zZQ==