Abstract
Intraspecific variation and trade-off in aerobic and anaerobic traits remain poorly understood in aquatic locomotion. Using gilthead sea bream (Sparus aurata) and Trinidadian guppy (Poecilia reticulata), both axial swimmers, this study tested four hypotheses: (1) gait transition from steady to unsteady (i.e., burst-assisted) swimming is associated with anaerobic metabolism evidenced as excess post exercise oxygen consumption (EPOC); (2) variation in swimming performance (critical swimming speed; Ucrit) correlates with metabolic scope (MS) or anaerobic capacity (i.e., maximum EPOC); (3) there is a trade-off between maximum sustained swimming speed (Usus) and minimum cost of transport (COTmin); and (4) variation in Usus correlates positively with optimum swimming speed (Uopt; i.e., the speed that minimizes energy expenditure per unit of distance traveled). Data collection involved swimming respirometry and video analysis. Results showed that anaerobic swimming costs (i.e., EPOC) increase linearly with the number of bursts in S. aurata, with each burst corresponding to 0.53 mg O2 kg−1. Data are consistent with a previous study on striped surfperch (Embiotoca lateralis), a labriform swimmer, suggesting that the metabolic cost of burst swimming is similar across various types of locomotion. There was no correlation between Ucrit and MS or anaerobic capacity in S. aurata indicating that other factors, including morphological or biomechanical traits, influenced Ucrit. We found no evidence of a trade-off between Usus and COTmin. In fact, data revealed significant negative correlations between Usus and COTmin, suggesting that individuals with high Usus also exhibit low COTmin. Finally, there were positive correlations between Usus and Uopt. Our study demonstrates the energetic importance of anaerobic metabolism during unsteady swimming, and provides intraspecific evidence that superior maximum sustained swimming speed is associated with superior swimming economy and optimum speed.
Introduction
Variation in locomotor performance and metabolism is linked to fitness, because both traits are often coupled with important behaviors such as predator evasion, prey capture, reproduction, migration, and dominance (Clobert et al., 2000; Walker et al., 2005; Langerhans, 2009a; Leis et al., 2009; Eliason et al., 2011; Seebacher et al., 2013; Wilson et al., 2013; Burnett et al., 2014; Killen et al., 2014; Scantlebury et al., 2014). Intraspecific variation in locomotor performance and metabolism is repeatable across time and environments (Chappell and Odell, 2004; Claireaux et al., 2005, 2007; Oufiero and Garland, 2009; Norin and Malte, 2011, 2012; Careau et al., 2014) and may be heritable and/or trans-generational (Rønning et al., 2007; Dalziel et al., 2011, 2012; Dalziel and Schulte, 2012; Gore and Burggren, 2012; McKenzie et al., 2013; Mattila and Hanski, 2014), indicating that traits related to locomotor performance and metabolism are subjected to natural selection and could evolve over time.
Performance trade-offs are central to understanding the vast phenotypic variation found among species, populations, and individuals. Trade-offs may occur when two antagonistic traits cannot be optimized simultaneously, because the two traits pose conflicting demands on the same design feature (Damme et al., 2002). Consequently, excellence in one trait will come at the cost of performance in the other trait (Vanhooydonck et al., 2014). Hence, an organism may specialize in one trait at the cost of the other, in which case a trade-off may cause phenotypic differentiation (DeWitt and Scheiner, 2004; Konuma and Chiba, 2007; Herrel et al., 2009). Alternatively, the conflicting demands may result in organisms performing sub-optimally for both traits and therefore, constrain evolution (Lewontin, 1978; Arnold, 1992). In fish, there is evidence of a trade-off between endurance capacity and sprint speed (Langerhans, 2009b; Oufiero et al., 2011); however, the trade-off is not ubiquitous at the whole-organism level (Wilson et al., 2002; Vanhooydonck et al., 2014; Fu et al., 2015).
Levels of swimming exercise in fishes have been divided into three categories on the basis of the time a given speed can be maintained before the onset of fatigue (Beamish, 1978): sustained (more than 200 min), prolonged (20 s to 200 min) and burst swimming (less than 20 s). In many teleosts, the segmented myotomal musculature is distinctively divided into red oxidative (slow-twitch) muscles and white glycolytic (fast-twitch) muscles. Red muscles are powered by oxidative phosphorylation, whereas white muscles are largely powered by anaerobic utilization of phosphocreatine, ATP and glycogen. At sustainable swimming speeds, the red musculature is powering propulsion, whereas white musculature is increasingly recruited for propulsion at faster speeds. Employing white musculature for burst-assisted swimming typically involves significant physiological perturbations including decreasing levels of intracellular pH (Lurman et al., 2007) and muscle glycogen (Peake and Farrell, 2004), and increasing levels of lactate (Martínez et al., 2004; Peake and Farrell, 2004) and inorganic phosphate (Lurman et al., 2007) indicating a non-steady state and partial reliance on anaerobic metabolism. While metabolic locomotor cost during sustained swimming has received considerable attention (Brett, 1964; Steinhausen et al., 2005; Ohlberger et al., 2006; Svendsen et al., 2013), the metabolic cost during non-steady burst swimming remains poorly understood. Metabolic locomotor cost during sustained swimming can be estimated using measurements of instantaneous oxygen consumption rates (exercise MO2), whereas metabolic cost during unsustainable swimming can be estimated by combining exercise MO2 with excess post exercise oxygen consumption (EPOC). The presence of EPOC is considered evidence of anaerobic activity in intact fish (Beamish, 1978), with many of the physiological perturbations related to anaerobic metabolism cleared during the period associated with EPOC (Peake and Farrell, 2004). In striped surf perch (Embiotoca lateralis), a labriform swimmer, there is a linear relationship between the number of bursts and EPOC, with EPOC constituting 25% of the total swimming costs (total MO2; i.e., exercise MO2 and EPOC combined) on average (Svendsen et al., 2010). In contrast, the metabolic cost of burst swimming in axial swimmers is largely unknown (Puckett and Dill, 1984; Farrell, 2007).
Standard metabolic rate (MO2stand) is a basic maintenance requirement measured as the minimum rate of oxygen consumption of postprandial unstressed animals at rest, below which physiological function is impaired. Long-term energy demands for swimming, food acquisition and treatment, regulation owing to environmental perturbations, and reproduction are additional to standard metabolism. These demands are met within the range set by the maximum metabolic rate (MO2max) (Priede, 1985). The difference between MO2stand and MO2max is termed the metabolic scope (MS). Because MS is strongly influenced by environmental variables, including temperature and oxygen availability, MS is predicted to be a major physiological variable in relation to climate change and aquatic hypoxia (Claireaux and Lefrançois, 2007; Chabot and Claireaux, 2008; Guderley and Pörtner, 2010; Pörtner, 2010; Pörtner and Peck, 2010; Di Santo, 2015). Nevertheless, intraspecific relationships between MS and other important physiological traits have rarely been explored in detail. MS and swimming performance correlate positively in intraspecific comparisons involving disparate populations of Atlantic silverside (Menidia menidia) (Arnott et al., 2006) and rainbow trout (Oncorhynchus mykiss) (Claireaux et al., 2005), but it remains uncertain to what extent the relationship is found in other species.
Locomotor performance and associated metabolic costs are often coupled with life history traits, which may involve trade-offs related to growth and MO2stand (Arnott et al., 2006; Rouleau et al., 2010). Recently, it was suggested that a trade-off between maximum sustained swimming speed (Usus) and minimum cost of transport (COTmin) may be driving morphological diversity in axial swimmers including teleosts and cetaceans (Tokić and Yue, 2012). The trade-off assumes constraints in optimizing both Usus and COTmin and suggests that aquatic species optimize either Usus or COTmin. Tokić and Yue (2012) applied the trade-off to models of morphological variation and reported congruent morphological variation in a number of extant aquatic species. While the trade-off may explain interspecific morphological variation, the trade-off has not been examined empirically at the intraspecific level. Likewise, it is not known if intraspecific diversity in Usus is a source of variation in optimum swimming speed (Uopt), i.e., the speed that minimizes energy expenditure per unit of distance traveled.
Using gilthead sea bream (Sparus aurata), E. lateralis and Trinidadian guppy (Poecilia reticulata), we employed swimming respirometry and video analyses to test four hypotheses: (1) burst activity is an indicator of anaerobic power production and correlates positively with the presence and magnitude of EPOC; (2) intraspecific diversity in MS or anaerobic capacity correlates positively with swimming performance; (3) there is a trade-off between Usus and COTmin such that a high value of Usus is associated with a high value of COTmin at the intraspecific level, and (4) variation in Usus correlates positively with Uopt. Data on S. aurata were collected for the present study, whereas data on E. lateralis and P. reticulata were derived from previous studies (Svendsen et al., 2010, 2013).
Materials and methods
Animals
A total of 13 gilthead sea bream (Sparus aurata) (body mass: 79.77 ± 2.38 g: standard length: 14.79 ± 0.24 cm (mean ± SE)) were obtained from a fish farm (Ferme Marine de Douhet) in France and kept in a flow-through holding tank (0.7 m3) with saltwater (30%) at 10 ± 1°C at the University of Copenhagen in Denmark. S. aurata were fed daily with commercial trout pellets (Biomar, Brande, Denmark). All methods applied in the present study were in agreement with current Danish regulations for the treatment and welfare of experimental animals. No fish were used more than once, and there was no mortality during any of the tests.
Respirometry
A swimming respirometer (8.24 L) was used to measure oxygen consumption rate (MO2; mg O2 kg−1 h−1) as a function of swimming speed (U). Water temperature inside the respirometer was maintained at 10.0°C (range: 9.9–10.1°C) using a temperature controlling instrument (TMP-REG; Loligo Systems; Tjele, Denmark). The respirometer was submerged in an ambient tank supplying water for the respirometer. Air stones maintained oxygen levels >95% air saturation in the ambient tank, and the water was recirculated through a loop consisting of a separate biological filter and a UV sterilizer (model UV-1000; Tetra Pond, Melle, Germany).
The swimming section of the respirometer was 32 × 9 × 11 cm (L × W × H). An impeller placed downstream of the swimming section was driven by an external electric motor that generated a re-circulating flow. Deflectors situated upstream of the swimming section collimated the flow. To promote rectilinear flow and a uniform velocity profile in the swimming section, water passed through an upstream honeycomb (7 mm cell diameter; Plascore Inc., Michigan, USA) producing a micro turbulent flow. A grid (10 mm) in the downstream direction bounded the swimming section. A vane wheel flow sensor (Höntzsch GmbH, Waiblingen, Germany) was used to measure water speeds in the swimming section. The measurements were used for a linear correlation between water speed and voltage output from the external motor controller.
Oxygen partial pressure (kPa) in the respirometer was measured using fiber optic sensor technology (PreSens, Regensburg, Germany). Intermittent flow respirometry was applied in accordance with previous studies (Steffensen, 1989). A computer-actuated pump was employed to replace water in the respirometer through a chimney as described previously (Svendsen et al., 2013). The software AutoResp (Loligo Systems Aps, Tjele, Denmark) was used to control the flush (240 s), wait (120 s) and measurement (540 s) phases. The settings provided one measurement of MO2 per 15 min. The declining oxygen partial pressure (kPa) during the measurement phase was used to calculate MO2 (mg O2 kg−1 h−1) using the equation: where K is the linear rate of decline (kPa h−1) in the oxygen content over time (h) in the respirometer, V is the volume of the respirometer (L) corrected for the volume of fish, β is the solubility of oxygen in the water (mg O2 L−1 kPa−1) (β = 0.4480) and M is the body mass of the fish (kg).
Preliminary trials demonstrated that the variation explained (R2) by the linear equation fitted to the declining oxygen content (kPa h−1), associated with each MO2 measurement, was always ≥0.95, similar to previous studies (Claireaux et al., 2006; Svendsen et al., 2012). The oxygen content never fell below 17.6 kPa. Levels of background respiration (i.e., microbial respiration) were estimated from blank runs and used to correct MO2 measurements (Jones et al., 2007; Svendsen et al., 2014).
Burst swimming
Individual fish in the swimming section were recorded dorsally using a Hitachi video camera (model VM-H630E; Düsseldorf, Germany), situated above the swimming respirometer. A Pinnacle frame grabber (model PCTV USB2; Corel Corporation, Ontario, Canada) continuously transferred recordings to a PC, and fish 2D position (x, y coordinates) was tracked at 25 Hz using the software LoliTrack (Loligo Systems, Tjele, Denmark). A burst was defined as a forward excursion (≥4 cm) with the swimming speed increasing ≥5 cm s−1. The number of bursts was determined over 3 min per respirometric loop (each 15 min) and used to estimate the total number of bursts per swimming speed (each 30 min; see below).
Experimental protocol
S. aurata for experiments were fasted for 48 h prior to respirometry to ensure a post-absorptive state. Fish mass (to nearest 0.01 g), length, depth and width (all to nearest 1 mm) were measured for pre-experimental calculation and correction of the solid blocking effects (Bell and Terhune, 1970; Gehrkel et al., 1990). Fish were acclimated to the respirometer for 12 h (overnight) while swimming at 0.5 body lengths per second (BL s−1) prior to collection of data.
After the acclimation period, routine MO2 (MO2routine) was estimated as the average MO2 during eight consecutive respirometric loops (i.e., 2 h) for each individual S. aurata swimming at 0.5 BL s−1 (i.e., acclimation speed) (Svendsen et al., 2010). At the individual level, the standard deviation (SD) of MO2routine was calculated using the eight MO2 measurements. Next, S. aurata were exposed to progressive increments in the swimming speed of 0.5 BL s−1 every 30 min up to 2 BL s−1. Using 30 min intervals for each swimming speed is a common approach (Schurmann and Steffensen, 1997; McKenzie et al., 2003, 2004; Lurman et al., 2007). Two measures of MO2 were collected at each swimming speed. After completing measurements at 2 BL s−1, S. aurata were exposed to speed increments of 0.25 BL s−1 every 30 min.
To examine the presence and magnitude of EPOC, the swimming speed was reduced to 0.5 BL s−1 (acclimation speed) after each exercise level from 2 BL s−1 and onwards. Specifically, detection of EPOC was carried out by comparing individual MO2routine + SD with the first post exercise MO2 measurement during the 0.5 BL s−1 period that followed each new swimming exercise (Svendsen et al., 2010). It was considered evidence of EPOC if the first post exercise MO2 was above MO2routine + SD. The measurements of MO2 at 0.5 BL s−1 were continued until the MO2 was below MO2routine + SD. When the MO2 stabilized below MO2routine + SD, the swimming speed was increased to the next exercise level (i.e., the previous exercise speed + 0.25 BL s−1). The protocol involving incrementally increasing swimming speeds followed by the procedure to detect EPOC was continued until fatigue.
Data acquisition and analysis
Exercise MO2 was recorded at increasing speeds from 0.5 BL s−1 to fatigue. Exercise MO2 as a function of U in individual fish was described by the exponential equation: where a is the MO2 at zero speed (U = 0) and b is the rate of increase in the MO2 as a function of U. The intercept with the y-axis (a) provides an estimate of the standard metabolic rate (MO2stand) (Brett, 1964; Arnott et al., 2006; Svendsen et al., 2013). The analyses included a comparable data set on P. reticulata from an earlier study (Svendsen et al., 2013) in addition to the collected data on S. aurata. Following Svendsen et al. (2013), model fittings were limited to swim speeds without burst-assisted swimming. The analysis disregarded the measurements of post exercise MO2 at 0.5 BL s−1 that were inserted to evaluate EPOC after swimming speeds ≥2 BL s−1. Equation (2) was fitted to the individual data sets using mixed-effect models to account for temporal autocorrelation due to the repeated measurements. The analysis included an AR1 (autoregressive of order 1) covariance structure.
Maximum sustained (or aerobic) metabolic rate (MO2sus) is defined as the maximum metabolic rate that can be maintained aerobically without the accumulation of anaerobic metabolic products that contribute to fatigue and negatively impact endurance (Hillman et al., 2014). In the present study, EPOC was detected when post exercise MO2 was above MO2routine + SD, indicating anaerobic metabolism. At the individual level, MO2sus was measured as the maximum recorded metabolic rate (over 0.5 h) at increasing swimming speeds without evidence of EPOC. The concurrent swimming speed was used as an estimate of the maximum sustained swimming speed (Usus).
Active metabolic rate (MO2active) was defined as the maximum exercise MO2 that S. aurata maintained for 0.5 h without fatigue (Schurmann and Steffensen, 1997; Claireaux et al., 2005). Maximum metabolic rate (MO2max) was defined as the highest exercise MO2 measured during the complete swimming protocol (McKenzie et al., 2003; Svendsen et al., 2013; Binning et al., 2014). MO2active and MO2max may be different, because MO2active is measured over 30 min, whereas MO2max is often measured over a shorter period of time (minimum 15 min; one respirometric loop) and at a higher swim speed.
MO2active is usually assumed to be the maximum aerobic metabolic rate (Schurmann and Steffensen, 1997); however, to what extent MO2active includes an anaerobic component remains uncertain. If MO2active is the maximum aerobic metabolic rate, MO2active should not differ significantly from MO2sus. To clarify differences between metabolic rates, a one way repeated measure ANOVA was used to compare MO2stand, MO2sus, MO2active, and MO2max. The test was followed by all pairwise comparison procedures (Holm-Šídák). The same test was employed to compare the swimming speeds associated with MO2sus, MO2active, and MO2max (i.e., Usus, Uactive, and Umax).
The method described by Brett (1964) was used to calculate the critical swimming speed (Ucrit). The protocol provides measurements that are repeatable in individual fish, suggesting that Ucrit represent a measure of performance, which is a lasting characteristic of the organism (Claireaux et al., 2007; Oufiero and Garland, 2009).
The magnitude of EPOC (mg O2 kg−1) was quantified using protocols published previously (Svendsen et al., 2010). When EPOC was detected, the individual relationship between time t (h) and post exercise MO2 was described using a double exponential equation: where a, b, c, and d are constants estimated using non-linear regression. Data included the exercise MO2 at t = 0. The recovery period was terminated when the fitted curve intercepted MO2routine + SD and provided an estimate of recovery time (h). EPOC magnitude was calculated as the integrated area between the fitted curve (Equation 3) and MO2routine from t = 0 to the end of the recovery period. At the individual level, EPOC was combined with the exercise MO2 to provide an estimate of the total cost of swimming (total MO2; mg O2 kg−1 h−1), covering both aerobic and anaerobic components. The anaerobic capacity was estimated as the maximum EPOC observed in individual fish. Anaerobic capacity was quantified as mg O2 kg−1 and mg O2 kg−1 h−1.
To test if the onset of burst swimming is a reliable predictor of the onset of EPOC, the minimum speed with burst swimming was correlated with the minimum speed with EPOC. The analysis was carried out using linear least square regression.
Linear mixed effects models were used to examine the relationship between the number of bursts and the magnitude of EPOC (mg O2 kg−1). Models included swimming speed as a covariate and interaction terms for swimming speed, burst number and fish identity. Temporal autocorrelation due to repeated measures was accounted for by including an AR1 covariance structure. The analysis included a comparable data set on E. lateralis from an earlier study (Svendsen et al., 2010).
The metabolic scope was calculated as MO2max–MO2stand in individual fish. The hypothesis that swimming performance (Ucrit) is correlated with metabolic scope or anaerobic capacity in individual fish was tested using linear least square regression.
Cost of transport (COT) was calculated as mg O2 kg−1 m−1 using the equation:
where MO2 is the metabolic rate (mg O2 kg−1 h−1), and U is the corresponding swimming speed (m h−1). The relationship between swimming speed and COT is usually U or 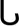 shaped with high COT values at low and high swimming speeds (Rouleau et al., 2010).
shaped with high COT values at low and high swimming speeds (Rouleau et al., 2010).
For each individual fish, COTmin was measured using two different approaches: (A) COTmin was estimated as the lowest recorded value of COT. Following this approach, the optimum swimming speed (Uopt; the speed that minimizes energy expenditure per unit of distance traveled) was estimated as the swimming speed that corresponded to COTmin; (B) COTmin was estimated by first determining Uopt using the equation: where b originates from Equation (2) describing the individual relationship between swimming speed (cm s−1) and MO2 (mg O2 kg−1 h−1). Next, MO2 at Uopt was calculated using Equation (2); and then COTmin was derived using Equation (4). Results from both approaches (A and B) to estimate COTmin and Uopt are reported, but figures are based on approach A. The analyses included a comparable data set on P. reticulata from an earlier study (Svendsen et al., 2013).
In a modeling study, Tokić and Yue (2012) presented evidence for a trade-off between Usus and COTmin. The trade-off predicts a positive correlation between Usus and COTmin, i.e., superior sustained swimming performance is associated with inferior swimming economy. To examine the trade-off in S. aurata, individual measures of Usus and COTmin were correlated using linear least square regression. Similarly, this study tested for a relationship between Usus and Uopt in individual fish. In addition to the data on S. aurata, the analyses of Usus, COTmin, and Uopt included a comparable data set derived from an earlier study on P. reticulata (Svendsen et al., 2013).
Data were transformed [e.g., ln(x + 1)] to meet the normality and homoscedasticity requirements of parametric analyses. The free statistical software R (R Development Core Team, 2014) and SigmaPlot (Systat Software, Erkrath, Germany) were used for statistical analyses and graphing. The R package nlme (Pinheiro et al., 2011) was employed to fit models. Results were considered significant at P < 0.05. All values are reported as means ± SE unless otherwise noted.
Results
Metabolic rates and swimming performance
MO2stand, MO2sus, MO2active, and MO2max were measured at increasing speeds (Figure 1) and were all statistically different (P < 0.05). Notably, MO2sus was lower than MO2active, providing evidence of anaerobic metabolism (EPOC) in a significant number of S. aurata exercising at the level of MO2active (Figure 1A). The finding suggests that MO2sus is a more appropriate measure of maximum sustained (or aerobic) metabolic rate than MO2active. Similar to the metabolic values, the corresponding swimming speeds (Usus, Uactive, and Umax) differed significantly (P < 0.05) (Figure 1B). Interestingly, Usus varied twofold between individuals with measurements ranging between 27 and 53.2 cm s−1. Measures of Ucrit were not included in Figure 1, but ranged between 35.3 and 56.5 cm s−1, with an average value of 45.0 ± 1.6 cm s−1. MO2sus and Usus corresponded to 79.3 ± 3.3% of MO2max and 88.9 ± 1.9% of Ucrit, respectively, with anaerobic metabolism detected above these exercise levels.
Figure 1
Exercise MO2 and total MO2 in relation to Ucrit
EPOC was detected at all swimming speeds faster than Usus and was combined with the exercise MO2 to estimate the total MO2. Because of the observed intraspecific variation in swimming performance, exercise MO2 and total MO2 were plotted as a function of %Ucrit (Figure 2A) similar to previous studies (Lurman et al., 2007; Tudorache et al., 2008; Teulier et al., 2013). EPOC contributed to the total MO2 starting at 86% of Ucrit (Figure 2A). EPOC constituted 53.5 ± 4.9% of the total MO2, ranging from 14.2 to 86.4% of total MO2, at swimming speeds with evident EPOC. Thus, EPOC frequently constituted more than half of the swimming costs. Recovery time associated with EPOC lasted 7.8 ± 1.1 h, ranging from 1.0 to 20.9 h (Figure 2B).
Figure 2
Positive correlations between burst activity and anaerobic metabolism (EPOC)
There was a positive linear relationship (P < 0.0001; R2 > 0.95) between the minimum speed with EPOC and the minimum speed with burst swimming (Figure 3). The intercept with the y-axis was not significantly different from zero (P > 0.65). The relationship shows that the onset of burst swimming is a strong predictor of the onset of EPOC and anaerobic metabolism at increasing swimming speeds.
Figure 3
The relationship between the number of bursts and magnitude of EPOC was examined using a linear mixed effects model. The model included swimming speed as a covariate, but no significant effect (P > 0.25) or interactions (P > 0.64) related to swimming speed was detected. Model terms for swimming speed and interactions were therefore eliminated from further analyses. A comparable data set from a previous study on E. lateralis (Svendsen et al., 2010) was included in the analysis. For both data sets, the intercept with y-axis was not significantly different from zero (P > 0.34) and the slopes did not differ between the two data sets (P > 0.94). These findings indicated that the relationships between burst numbers and EPOC were similar in the two species, and the data were therefore, combined. The resulting common relationship (Figure 4) was described by the equation (P < 0.0001):
Figure 4
The relationship indicates that each burst corresponds to an average metabolic cost of 0.53 mg O2 kg−1 (Figure 4).
No correlation between Ucrit and metabolic scope or anaerobic capacity
Metabolic scope was estimated as MO2max–MO2stand, whereas anaerobic capacity was estimated as the maximum EPOC observed in individual fish. The maximum EPOC value was always associated with fish fatigue. There was no evidence that individual Ucrit correlated with metabolic scope (P > 0.87; R2 < 0.01) or with anaerobic capacity (P > 0.57; R2 < 0.04) (data not shown). The analyses of anaerobic capacity involved maximum EPOC quantified as mg O2 kg−1 and mg O2 kg−1 h−1.
No trade-off between Usus and COTmin
This study examined a trade-off between Usus and COTmin by comparing swimming performance and metabolism in S. aurata and P. reticulata. In terms of S. aurata, Usus was assumed to correspond to the highest swimming speed without EPOC (Figures 1, 2). Data on P. reticulata were derived from Svendsen et al. (2013). While EPOC was not measured in P. reticulata, the study quantified burst activity in individual P. reticulata at increasing speeds. Using the relationship between the onset of burst swimming and the onset of EPOC (Figure 3), EPOC occurrence at increasing speeds, and thereby Usus, were estimated in individual P. reticulata. COTmin in P. reticulata was estimated in the same fashions (approaches A and B) as in S. aurata (Equations 4 and 5). The relationships between Usus and COTmin were examined using linear least square regressions (Figure 5). For both species, there was no evidence of a trade-off between Usus and COTmin. In fact, there were significant negative correlations between Usus and COTmin, revealing that individuals exhibiting superior sustained swimming performance (i.e., high Usus) also exhibit superior swimming economy (i.e., low COTmin) (Figure 5). The negative correlations between Usus and COTmin were evident in both species and regardless of the approach (A and B) used to estimate COTmin (all P < 0.005; R2 > 0.53). Data in Figure 5 are based on approach A.
Figure 5
Positive correlations between Usus and Uopt
There were significant positive correlations between Usus and Uopt (Figure 6). The analyses included data on S. aurata (Figure 6A) and P. reticulata (Figure 6B) and revealed that individuals exhibiting superior sustained swimming performance (i.e., high Usus) also exhibit superior optimum swim speed (i.e., high Uopt). The positive correlations between Usus and Uopt were evident in both species and regardless of the approach used to estimate Uopt (approach A: all P < 0.005; R2 > 0.40; approach B: all P < 0.05; R2 > 0.26). Data in Figure 6 are based on approach A.
Figure 6
Discussion
This study demonstrated the energetic importance of anaerobic metabolism during unsteady locomotion. There was no evidence of Ucrit correlating with MS or anaerobic capacity. Moreover, we provided intraspecific evidence that a high Usus is coupled with low COTmin and high Uopt in individual fish. Specifically, our results reveal that burst swimming is associated with anaerobic metabolism and a substantial metabolic cost, which is expressed as EPOC. Our intraspecific results on two teleost species are at odds with the conjecture that there is a trade-off between Usus and COTmin as indicated by Tokić and Yue (2012). By applying the trade-off, the authors provided a model that explained variation in morphology in various teleost and cetacean species. In contrast, the present study is based on intraspecific data collected empirically. Our findings suggest that intraspecific variation in Usus and COTmin is not driven by a trade-off producing a high Usus in some individuals and a low COTmin in other individuals. Because the results suggest that Usus and COTmin are optimized concurrently, it is unlikely that the trade-off drives intraspecific morphological variation.
Previous studies have demonstrated that the Ucrit protocol includes swimming powered by both aerobic and anaerobic metabolism (Burgetz et al., 1998; Richards et al., 2002). In E. lateralis, EPOC and anaerobic metabolism is present at 88% of Ucrit (Svendsen et al., 2010). Corroborating previous results, the present study found evidence of EPOC starting at swimming speeds corresponding to 86% of Ucrit. At higher speeds, EPOC increased rapidly and constituted up to 86% of the total MO2. The maximum value of EPOC was always associated with fatigue. Likewise, beginning at 89% of Ucrit in Atlantic cod (Gadus morhua), Lurman et al. (2007) found evidence of anaerobic metabolism as indicated by decreasing levels of phosphocreatine and intracellular pH and increasing levels of inorganic phosphate. Our study corroborates that the Ucrit protocol involves depletion of both aerobic and anaerobic resources, and shows that the metabolic costs associated with the recovery from the anaerobic perturbation (i.e., EPOC) may constitute the majority of the swimming costs. The results highlight the importance of measuring both exercise MO2 and EPOC to estimate the total metabolic costs of swimming in fish approaching prolonged and burst swimming speeds. In the absence of EPOC measurements, the metabolic cost of swimming may be significantly underestimated.
This study shows that the onset of burst-assisted swimming is closely related to the onset of EPOC at increasing swimming speeds in individual fish. The initiation of burst swimming is therefore a strong predictor of EPOC and anaerobic metabolism. Similarly, we found that the magnitude of EPOC increases linearly with the number of bursts. The present data are consistent with a previous study on E. lateralis (Svendsen et al., 2010). Combining the two data sets suggests that each burst corresponds to an energetic cost of 0.53 mg O2 kg−1. E. lateralis is a labriform swimmer (i.e., pectoral fins used for propulsion at low and medium swimming speeds) whereas S. aurata is an axial swimmer (i.e., axial undulation used for propulsion). The fact that we found no differences in the two relationships between bursts and EPOC indicates that the metabolic cost of burst swimming may be similar across fish species employing disparate types of locomotion.
MS is predicted to play a major role in relation to effects of climate change, and other anthropogenic stressors including hypoxia, on aquatic exothermic animals (Claireaux and Lefrançois, 2007; Chabot and Claireaux, 2008; Guderley and Pörtner, 2010; Pörtner, 2010; Pörtner and Peck, 2010; McBryan et al., 2013; Seth et al., 2013; Di Santo, 2015). It remains uncertain, however, to what extent intraspecific diversity in MS varies with other important physiological traits including locomotor performance. The present study measured intraspecific variation in MS and anaerobic capacity (i.e., maximal EPOC) in S. aurata and correlated data with individual variation in swimming performance (Ucrit). We found no evidence that diversity in MS or anaerobic capacity correlates with Ucrit (P > 0.57) indicating that other factors, including morphological (Rouleau et al., 2010) or biomechanical (Svendsen et al., 2013) traits, drive the variation in swimming performance.
It is possible that the lacking relationship between MS and Ucrit was caused by our method of measuring MO2max. Similar to previous studies (McKenzie et al., 2003; Svendsen et al., 2013; Binning et al., 2014), we used an Ucrit protocol to measure MO2max involving progressive increments in the swimming speed of 0.25 BL s−1 every 30 min, starting from 2 BL s−1 and until fatigue. Our protocol differed, however, from conventional protocols, because we inserted periods with swimming speeds adjusted to 0.5 BL s−1 (acclimation speed) for measurements of EPOC after each swimming speed ≥ 2 BL s−1. Although the mechanistic basis is unknown, it is possible that our protocol affected the measurements of MO2max. As an alternative to the Ucrit protocol, a number of recent studies have used a chase protocol to measure MO2max (Norin and Malte, 2011, 2012; Svendsen et al., 2014). The Ucrit protocol is often assumed to provide measures of MO2max (Farrell and Steffensen, 1987; Hammer, 1995) and may in fact elicit values of MO2max that are higher than the values elicited by the chase protocol (Roche et al., 2013). Therefore, it is unlikely that a significant relationship between MS and swimming performance would have been revealed if we had used a chase protocol instead of the Ucrit protocol to measure MO2max. In humans, MO2max is typically measured using test protocols that are much faster (Barker et al., 2011; Vanhatalo et al., 2011; Mauger et al., 2013) than the Ucrit protocol used in the present study. While a protocol that continuously steps up the swimming speed in much faster pace than the Ucrit protocol might produce higher values of MO2max (and therefore MS) and swimming performance (Farrell, 2008), it remains to be tested if the methodology would produce a significant relationship between MS and swimming performance. A faster protocol would rely more on anaerobic metabolism to power swimming (Farrell, 2008; Poulsen et al., 2012), and so a relationship between anaerobic capacity and swimming performance might be revealed.
A recent study emphasized a trade-off between Usus and COTmin driving morphological diversity in aquatic locomotion (Tokić and Yue, 2012). The trade-off assumes constraints in optimizing Usus and COTmin simultaneously, suggesting that aquatic species may optimize either Usus or COTmin. By applying the trade-off, Tokić and Yue (2012) modeled morphological variation and reported congruent morphological variation in several extant aquatic species. The present study examined the trade-off within two teleost species and found no support for the trade-off. In fact, data revealed a significant negative correlation between Usus and COTmin, suggesting that individuals with high Usus also exhibit low COTmin. The negative relationship indicates that the two traits are optimized simultaneously and could be related to the same mechanistic basis without constraints. Interestingly, studies are increasingly uncovering significant intraspecific variation in locomotor performance and metabolic rate (Nelson et al., 2003; Langerhans, 2008, 2009a; Dalziel et al., 2011, 2012; Dalziel and Schulte, 2012; Svendsen et al., 2013; Binning et al., 2014). The present study indicates that intraspecific morphological variation, associated with intraspecific variation in locomotor performance and metabolic rate, is not driven by a trade-off between Usus and COTmin.
There are a number of reasons why we may not observe a trade-off between Usus and COTmin in our intraspecific data. Variation between species is much more pronounced than between individuals of the same species. Therefore, interspecific variation may better reflect the full spectrum of functional trade-offs that influences morphological variation related to aquatic locomotion. It is also possible that a trade-off between Usus and COTmin is present in the two tested fish species, but not expressed at the whole-organism level, because of compensating or masking factors involving morphological, physiological and/or biomechanical traits. Moreover, our estimates of Usus and COTmin based on respirometry and video analysis might be misleading. For example, it is possible that estimates of Usus using measures of EPOC (S. aurata) and burst-assisted swimming (P. reticulata) do not accurately reflect maximum sustained swimming speeds. MO2 is, however, a well-established proxy for aerobic metabolic rate, and the gait transition from steady to unsteady (i.e., burst-assisted) swimming is a well-known indicator of the shift from aerobic to anaerobic power production (Peake and Farrell, 2004, 2006; Peake, 2008; Svendsen et al., 2010). Similarly, it is possible that the use of forced linear swimming to estimate Usus and COTmin provides results that do not necessarily reflect natural conditions, because fish typically swim spontaneously in a non-linear fashion with the relationship between swimming speed and metabolic rate differing from linear swimming (Steinhausen et al., 2010).
Diversity in locomotor performance and metabolism can be important sources of variation in animal behaviors. For example, Hillman et al. (2014) suggested that variation in physiological capacity for movement influences dispersal and therefore fine-scale genetic structure of several vertebrate groups. At the intraspecific level, physiological performance is an important determinant of behaviors related to schooling (Killen et al., 2011), territory acquisition and defense and foraging (Breau et al., 2011; Killen et al., 2014). Likewise, physiological and energetic states may influence behaviors in migratory species (Poulsen et al., 2010; Boel et al., 2014). Recent studies have shown that exercise training that increases swimming performance may change the behavior of animals and cause elevated boldness and exploratory tendency (Sinclair et al., 2014). The mechanistic basis of the relationship between exercise training and behavior could be related to the positive relationship between Usus and Uopt found in the present study. Because exercise training increases aerobic potentials in red and white musculature (Davison, 1997) and swimming performance (Farrell et al., 1990; Sinclair et al., 2014), exercise training should also elevate Usus and therefore Uopt. Typically, fish swim spontaneously at speeds corresponding to Uopt (Videler, 1993; Tudorache et al., 2011). This hypothesis suggests that exercise training increases spontaneous swimming speeds via the positive relationship between Usus and Uopt. It seems likely that increased spontaneous swimming speed is associated with elevated boldness and exploratory tendency as observed by Sinclair et al. (2014). Therefore, the positive relationship between Usus and Uopt could provide a mechanistic link between physiological and behavioral phenotypes. Nevertheless, this hypothetical framework warrants additional study to clarify the mechanistic basis of intraspecific correlations between physiological and behavioral phenotypes.
Statements
Author contributions
Conceived and designed the experiments: BT, JCS, JFS. Performed the experiments: BT. Analyzed the data: JCS, GAC, BT. Contributed reagents/materials/analysis tools: JFS. Wrote the paper: JCS. Revised the manuscript critically for important intellectual content: JCS, BT, GAC, JFS.
Acknowledgments
This research was supported by a grant (SFRH/BPD/89473/2012) from the Foundation for Science and Technology (FCT) in Portugal to JCS. The study was part of the project SUNFISH (sustainable fisheries, climate change and the North Sea ecosystem) and financially supported by the Danish Council for Strategic Research (grant number: 09-063096) and Department of Biology, University of Copenhagen (grant number: 102-0218/11-5550) to BT. Financial support from the Danish Research Council to JFS is also gratefully acknowledged. We thank C. Oufiero for helpful discussions. We thank two reviewers for constructive and helpful comments that improved the paper. This publication is supported by COST Action FA1304 “Swimming of fish and implications for migration and aquaculture (FITFISH).”
Conflict of interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
- EPOC
Excess post exercise oxygen consumption
- BL
Body length
- COT
Cost of transport
- COTmin
Minimum cost of transport
- Exercise MO2
Metabolic rate measured in swimming fish (i.e., instantaneous metabolic rate)
- MO2
Metabolic rate
- MO2active
Active metabolic rate defined as the maximum metabolic rate maintained for 0.5 h
- MO2max
Maximum metabolic rate defined as the maximum metabolic rate measured at increasing swimming speeds
- MO2routine
Routine metabolic rate defined as the average metabolic rate in fish swimming at 0.5 BL s−1
- MO2stand
Standard metabolic rate (i.e., a in Equation 2)
- MO2sus
Maximum sustained metabolic rate defined as the maximum metabolic rate (over 0.5 h) without any EPOC (i.e., no influence of anaerobic metabolism)
- Total MO2
Exercise MO2 and EPOC combined as an estimate of the total metabolic swimming cost
- Uactive
Swimming speed associated with the active metabolic rate (MO2active)
- Ucrit
Critical swimming speed
- Umax
Swimming speed associated with the maximum metabolic rate (MO2max)
- Uopt
Optimum swimming speed defined as the speed that minimizes energy expenditure per unit of distance traveled
- Usus
Maximum sustained swimming speed defined as the maximum recorded swimming speed (over 0.5 h) without any EPOC (i.e., no influence of anaerobic metabolism).
Abbreviations
References
1
ArnoldS. J. (1992). Constraints on phenotypic evolution. Am. Nat. 140, S85–S107. 10.1086/285398
2
ArnottS. A.ChibaS.ConoverD. O. (2006). Evolution of intrinsic growth rate: metabolic costs drive trade-offs between growth and swimming performance in Menidia menidia. Evolution60, 1269–1278. 10.1111/j.0014-3820.2006.tb01204.x
3
BarkerA. R.WilliamsC. A.JonesA. M.ArmstrongN. (2011). Establishing maximal oxygen uptake in young people during a ramp cycle test to exhaustion. Br. J. Sports Med. 45, 498–503. 10.1136/bjsm.2009.063180
4
BeamishF. W. H. (1978). Swimming capacity, in Fish Physiology, Vol. 7, eds HoarW. S.RandallD. J. (New York, NY: Academic Press), 101–187.
5
BellW. M.TerhuneL. D. B. (1970). Water tunnel design for fisheries research. Fish. Res. Board Can. Tech. Rep. 195, 1–69.
6
BinningS. A.RocheD. G.FultonC. J. (2014). Localised intraspecific variation in the swimming phenotype of a coral reef fish across different wave exposures. Oecologia174, 623–630. 10.1007/s00442-013-2794-5
7
BoelM.AarestrupK.BaktoftH.LarsenT.Søndergaard MadsenS.MalteH.et al. (2014). The physiological basis of the migration continuum in brown trout (Salmo trutta). Physiol. Biochem. Zool. 87, 334–345. 10.1086/674869
8
BreauC.CunjakR. A.PeakeS. J. (2011). Behaviour during elevated water temperatures: can physiology explain movement of juvenile Atlantic salmon to cool water?J. Anim. Ecol. 80, 844–853. 10.1111/j.1365-2656.2011.01828.x
9
BrettJ. R. (1964). The respiratory metabolism and swimming performance of young sockeye salmon. J. Fish. Board Can. 21, 1183–1226. 10.1139/f64-103
10
BurgetzI. J.Rojas-VargasA.HinchS. G.RandallD. J. (1998). Initial recruitment of anaerobic metabolism during sub-maximal swimming in rainbow trout (Oncorhynchus mykiss). J. Exp. Biol. 201, 2711–2721.
11
BurnettN. J.HinchS. G.BraunD. C.CasselmanM. T.MiddletonC. T.WilsonS. M.et al. (2014). Burst swimming in areas of high flow: delayed consequences of anaerobiosis in wild adult sockeye salmon. Physiol. Biochem. Zool. 87, 587–598. 10.1086/677219
12
CareauV.GiffordM. E.BiroP. A. (2014). Individual (co)variation in thermal reaction norms of standard and maximal metabolic rates in wild-caught slimy salamanders. Funct. Ecol. 28, 1175–1186. 10.1111/1365-2435.12259
13
ChabotD.ClaireauxG. (2008). Environmental hypoxia as a metabolic constraint on fish: the case of Atlantic cod, Gadus morhua. Mar. Pollut. Bull. 57, 287–294. 10.1016/j.marpolbul.2008.04.001
14
ChappellM.OdellJ. (2004). Predation intensity does not cause microevolutionary change in maximum speed or aerobic capacity in trinidadian guppies (Poecilia reticulata Peters). Physiol. Biochem. Zool. 77, 27–38. 10.1086/378920
15
ClaireauxG.CouturierC.GroisonA.-L. (2006). Effect of temperature on maximum swimming speed and cost of transport in juvenile European sea bass (Dicentrarchus labrax). J. Exp. Biol. 209, 3420–3428. 10.1242/jeb.02346
16
ClaireauxG.HandelsmanC.StandenE.NelsonJ. A. (2007). Thermal and temporal stability of swimming performance in the European sea bass. Physiol. Biochem. Zool. 80, 186–196. 10.1086/511143
17
ClaireauxG.LefrançoisC. (2007). Linking environmental variability and fish performance: integration through the concept of scope for activity. Philos. Trans. R. Soc. Lond. B Biol. Sci. 362, 2031–2041. 10.1098/rstb.2007.2099
18
ClaireauxG.McKenzieD. J.GengeA. G.ChatelierA.AubinJ.FarrellA. P. (2005). Linking swimming performance, cardiac pumping ability and cardiac anatomy in rainbow trout. J. Exp. Biol. 208, 1775–1784. 10.1242/jeb.01587
19
ClobertJ.OppligerA.SorciG.ErnandeB.SwallowJ. G.GarlandT. (2000). Trade-offs in phenotypic traits: endurance at birth, growth, survival, predation and susceptibility to parasitism in a lizard, Lacerta vivipara. Funct. Ecol. 14, 675–684. 10.1046/j.1365-2435.2000.00477.x
20
DalzielA. C.OuM.SchulteP. M. (2012). Mechanisms underlying parallel reductions in aerobic capacity in non-migratory threespine stickleback (Gasterosteus aculeatus) populations. J. Exp. Biol. 215, 746–759. 10.1242/jeb.065425
21
DalzielA. C.SchulteP. M. (2012). Correlates of prolonged swimming performance in F2 hybrids of migratory and non-migratory threespine stickleback. J. Exp. Biol. 215, 3587–3596. 10.1242/jeb.071951
22
DalzielA. C.VinesT. H.SchulteP. M. (2011). Reductions in prolonged swimming capacity following freshwater colonization in multiple threespine stickleback populations. Evolution66, 1226–1239. 10.1111/j.1558-5646.2011.01498.x
23
DammeR.Van WilsonR. S.VanhooydonckB.AertsP. (2002). Performance constraints in decathletes. Nature415, 755–756. 10.1038/415755b
24
DavisonW. (1997). The effects of exercise training on teleost fish, a review of recent literature. Comp. Biochem. Physiol. Part A Physiol. 117, 67–75. 10.1016/S0300-9629(96)00284-8
25
DeWittT. J.ScheinerS. M. (2004). Phenotypic Plasticity: Functional and Conceptual Approaches. Oxford, UK: Oxford University Press.
26
Di SantoV. (2015). Ocean acidification exacerbates the impacts of global warming on embryonic little skate, Leucoraja erinacea (Mitchill). J. Exp. Mar. Biol. Ecol. 463, 72–78. 10.1016/j.jembe.2014.11.006
27
EliasonE. J.ClarkT. D.HagueM. J.HansonL. M.GallagherZ. S.JeffriesK. M.et al. (2011). Differences in thermal tolerance among sockeye salmon populations. Science332, 109–112. 10.1126/science.1199158
28
FarrellA. P.JohansenJ. A.SteffensenJ. F.MoyesC. D.WestT. G.SuarezR. K. (1990). Effects of exercise training and coronary ablation on swimming performance, heart size, and cardiac enzymes in rainbow trout, Oncorhynchus mykiss. Can. J. Zool. 68, 1174–1179. 10.1139/z90-174
29
FarrellA. P.SteffensenJ. F. (1987). An analysis of the energetic cost of the branchial and cardiac pumps during sustained swimming in trout. Fish Physiol. Biochem. 4, 73–79. 10.1007/BF02044316
30
FarrellA. P. (2007). Cardiorespiratory performance during prolonged swimming tests with salmonids: a perspective on temperature effects and potential analytical pitfalls. Philos. Trans. R. Soc. Lond. B Biol. Sci. 362, 2017–2030. 10.1098/rstb.2007.2111
31
FarrellA. P. (2008). Comparisons of swimming performance in rainbow trout using constant acceleration and critical swimming speed tests. J. Fish Biol. 72, 693–710. 10.1111/j.1095-8649.2007.01759.x
32
FuC.YuanX. Z.CaoZ. D.FuS. J. (2015). Predator-driven intra-species variation in locomotion, metabolism and water velocity preference in pale chub (Zacco platypus) along a river. J. Exp. Biol. 218, 255–264. 10.1242/jeb.109561
33
GehrkelP. C.FidlerL. E.MenseD. C.RandallD. J. (1990). A respirometer with controlled water quality and computerized data acquisition for experiments with swimming fish. Fish Physiol. Biochem. 8, 61–67. 10.1007/BF00004432
34
GoreM.BurggrenW. W. (2012). Cardiac and metabolic physiology of early larval zebrafish (Danio rerio) reflects parental swimming stamina. Front. Physiol. 3, 1–9. 10.3389/fphys.2012.00035
35
GuderleyH.PörtnerH. O. (2010). Metabolic power budgeting and adaptive strategies in zoology: examples from scallops and fish. Can. J. Zool. 88, 753–763. 10.1139/Z10-039
36
HammerC. (1995). Fatigue and exercise tests with fish. Comp. Biochem. Physiol. 112A, 1–20. 10.1016/0300-9629(95)00060-K
37
HerrelA.PodosJ.VanhooydonckB.HendryA. P. (2009). Force-velocity trade-off in Darwin's finch jaw function: a biomechanical basis for ecological speciation?Funct. Ecol. 23, 119–125. 10.1111/j.1365-2435.2008.01494.x
38
HillmanS. S.DrewesR. C.HedrickM. S.HancockT., V (2014). Physiological vagility and its relationship to dispersal and neutral genetic heterogeneity in vertebrates. J. Exp. Biol. 217, 3356–3364. 10.1242/jeb.105908
39
JonesE. A.LuceyK. S.EllerbyD. J. (2007). Efficiency of labriform swimming in the bluegill sunfish (Lepomis macrochirus). J. Exp. Biol. 210, 3422–3429. 10.1242/jeb.005744
40
KillenS. S.MarrasS.SteffensenJ. F.McKenzieD. J. (2011). Aerobic capacity influences the spatial position of individuals within fish schools. Proc. Trans. R. Soc. Lond. B Biol. Sci. 279, 357–364. 10.1098/rspb.2011.1006
41
KillenS. S.MitchellM. D.RummerJ. L.ChiversD. P.FerrariM. C. O.MeekanM. G.et al. (2014). Aerobic scope predicts dominance during early life in a tropical damselfish. Funct. Ecol. 28, 1367–1376. 10.1111/1365-2435.12296
42
KonumaJ.ChibaS. (2007). Trade-offs between force and fit: extreme morphologies associated with feeding behavior in carabid beetles. Am. Nat. 170, 90–100. 10.1086/518182
43
LangerhansR. B. (2008). Predictability of phenotypic differentiation across flow regimes in fishes. Integr. Comp. Biol. 48, 750–768. 10.1093/icb/icn092
44
LangerhansR. B. (2009a). Morphology, performance, fitness: functional insight into a post-Pleistocene radiation of mosquitofish. Biol. Lett. 5, 488–491. 10.1098/rsbl.2009.0179
45
LangerhansR. B. (2009b). Trade-off between steady and unsteady swimming underlies predator-driven divergence in Gambusia affins. J. Evol. Biol. 22:1057. 10.1111/j.1420-9101.2009.01716.x
46
LeisJ. M.HayA. C.HowarthG. J. (2009). Ontogeny of in situ behaviours relevant to dispersal and population connectivity in larvae of coral-reef fishes. Mar. Ecol. Prog. Ser. 379, 163–179. 10.3354/meps07904
47
LewontinR. C. (1978). Adaptation. Sci. Am. 239, 212–218. 10.1038/scientificamerican0978-212
48
LurmanG. J.BockC. H.PörtnerH.-O. (2007). An examination of the metabolic processes underpinning critical swimming in Atlantic cod (Gadus morhua L.) using in vivo 31 P-NMR spectroscopy. J. Exp. Biol. 210, 3749–3756. 10.1242/jeb.008763
49
MartínezM.BédardM.DutilJ.-D.GuderleyH. (2004). Does condition of Atlantic cod (Gadus morhua) have a greater impact upon swimming performance at Ucrit or sprint speeds?J. Exp. Biol. 207, 2979–2990. 10.1242/jeb.01142
50
MattilaA. L. K.HanskiI. (2014). Heritability of flight and resting metabolic rates in the Glanville fritillary butterfly. J. Evol. Biol. 27, 1733–1743. 10.1111/jeb.12426
51
MaugerA. R.MetcalfeA. J.TaylorL.CastleP. C. (2013). The efficacy of the self-paced VO2max test to measure maximal oxygen uptake in treadmill running. Appl. Physiol. Nutr. Metab. 38, 1211–1216. 10.1139/apnm-2012-0384
52
McBryanT. L.AnttilaK.HealyT. M.SchulteP. M. (2013). Responses to temperature and hypoxia as interacting stressors in fish: implications for adaptation to environmental change. Integr. Comp. Biol. 53, 648–659. 10.1093/icb/ict066
53
McKenzieD. J.EstivalesG.SvendsenJ. C.SteffensenJ. F. (2013). Local adaptation to altitude underlies divergent thermal physiology in tropical killifishes of the genus Aphyosemion. PLoS ONE8:e54345. 10.1371/journal.pone.0054345
54
McKenzieD. J.MartínezR.MoralesA.AcostaJ.MoralesR.TaylorE. W.et al. (2003). Effects of growth hormone transgenesis on metabolic rate, exercise performance and hypoxia tolerance in tilapia hybrids. J. Fish Biol. 63, 398–409. 10.1046/j.1095-8649.2003.00162.x
55
McKenzieD. J.WongS.RandallD. J.EggintonS.TaylorE. W.FarrellA. P. (2004). The effects of sustained exercise and hypoxia upon oxygen tensions in the red muscle of rainbow trout. J. Exp. Biol. 207, 3629–3637. 10.1242/jeb.01199
56
NelsonJ. A.GotwaltP. S.SnodgrassJ. W. (2003). Swimming performance of blacknose dace (Rhinichthys atratulus) mirrors home-stream current velocity. Can. J. Fish. Aquat. Sci. 60, 301–308. 10.1139/f03-023
57
NorinT.MalteH. (2011). Repeatability of standard metabolic rate, active metabolic rate and aerobic scope in young brown trout during a period of moderate food availability. J. Exp. Biol. 214, 1668–1675. 10.1242/jeb.054205
58
NorinT.MalteH. (2012). Intraspecific variation in aerobic metabolic rate of fish: relations with organ size and enzyme activity in brown trout. Physiol. Biochem. Zool. 85, 645–656. 10.1086/665982
59
OhlbergerJ.StaaksG.HölkerF. (2006). Swimming efficiency and the influence of morphology on swimming costs in fishes. J. Comp. Physiol. B176, 17–25. 10.1007/s00360-005-0024-0
60
OufieroC. E.GarlandT. (2009). Repeatability and correlation of swimming performances and size over varying time-scales in the guppy (Poecilia reticulata). Funct. Ecol. 23, 969–978. 10.1111/j.1365-2435.2009.01571.x
61
OufieroC. E.WalshM. R.ReznickD. N.GarlandT.Jr. (2011). Swimming performance trade-offs across a gradient in community composition in Trinidadian killifish (Rivulus hartii). Ecology92, 170–179. 10.1890/09-1912.1
62
PeakeS. J.FarrellA. P. (2004). Locomotory behaviour and post-exercise physiology in relation to swimming speed, gait transition and metabolism in free-swimming smallmouth bass (Micropterus dolomieu). J. Exp. Biol. 207, 1563–1575. 10.1242/jeb.00927
63
PeakeS. J.FarrellA. P. (2006). Fatigue is a behavioural response in respirometer-confined smallmouth bass. J. Fish Biol. 68, 1742–1755. 10.1111/j.1095-8649.2006.01052.x
64
PeakeS. J. (2008). Gait transition speed as an alternate measure of maximum aerobic capacity in fishes. J. Fish Biol. 72, 645–655. 10.1111/j.1095-8649.2007.01753.x
65
PinheiroJ.BatesD.DebRoyS.SarkarD.the R Development Core Team. (2011). nlme: Linear and Nonlinear Mixed Effects Models. R Package Version 3.1-101. Vienna: R Foundation for Statistical Computing.
66
PörtnerH. O.PeckM. A. (2010). Climate change effects on fishes and fisheries: towards a cause-and-effect understanding. J. Fish Biol. 77, 1745–1779. 10.1111/j.1095-8649.2010.02783.x
67
PörtnerH.-O. (2010). Oxygen- and capacity-limitation of thermal tolerance: a matrix for integrating climate-related stressor effects in marine ecosystems. J. Exp. Biol. 213, 881–893. 10.1242/jeb.037523
68
PoulsenS. B.JensenL. F.SchulzC.DeaconM.MeyerK. E.Jäger-KleinickeT.et al. (2012). Ontogenetic differentiation of swimming performance and behaviour in relation to habitat availability in the endangered North Sea houting (Coregonus oxyrinchus). Aquat. Living Resour. 25, 241–249. 10.1051/alr/2002019
69
PoulsenS. B.SvendsenJ. C.JensenL. F.SchulzC.Jäger−KleinickeT.SchwartenH. (2010). Effects of food deprivation on refuge use and dispersal in juvenile North Sea houting Coregonus oxyrinchus under experimental conditions. J. Fish Biol. 77, 1702–1708. 10.1111/j.1095-8649.2010.02772.x
70
PriedeI. G. (1985). Metabolic scope in fishes, in Fish Energetics: New Perspectives, eds TytlerP.CalowP. (London: Croom-Helm), 33–64.
71
PuckettK. J.DillL. M. (1984). Cost of sustained and burst swimming to juvenile coho salmon (Oncorhynchus kisutch). Can. J. Fish. Aquat. Sci. 41, 1546–1551. 10.1139/f84-192
72
R Development Core Team. (2014). R: A Language and Environment for Statistical Computing. Vienna: R Foundation for Statistical Computing. Available online at: http://www.R-project.org/
73
RichardsJ. G.MercadoA. J.ClaytonC. A.HeigenhauserG. J. F.WoodC. M. (2002). Substrate utilization during graded aerobic exercise in rainbow trout. J. Exp. Biol. 205, 2067–2077.
74
RocheD. G.BinningS. A.BosigerY.JohansenJ. L.RummerJ. L. (2013). Finding the best estimates of metabolic rates in a coral reef fish. J. Exp. Biol. 216, 2103–2110. 10.1242/jeb.082925
75
RønningB.JensenH.MoeB.BechC. (2007). Basal metabolic rate: heritability and genetic correlations with morphological traits in the zebra finch. J. Evol. Biol. 20, 1815–1822. 10.1111/j.1420-9101.2007.01384.x
76
RouleauS.GlémetH.MagnanP. (2010). Effects of morphology on swimming performance in wild and laboratory crosses of brook trout ecotypes. Funct. Ecol. 24, 310–321. 10.1111/j.1365-2435.2009.01636.x
77
ScantleburyD. M.MillsM. G. L.WilsonR. P.WilsonJ. W.MillsM. E. J.DurantS. M.et al. (2014). Flexible energetics of cheetah hunting strategies provide resistance against kleptoparasitism. Science346, 79–81. 10.1126/science.1256424
78
SchurmannH.SteffensenJ. F. (1997). Effects of temperature, hypoxia and activity on the metabolism of juvenile Atlantic cod. J. Fish Biol. 50, 1166–1180.
79
SeebacherF.WardA. J. W.WilsonR. S. (2013). Increased aggression during pregnancy comes at a higher metabolic cost. J. Exp. Biol. 216, 771–776. 10.1242/jeb.079756
80
SethH.GränsA.SandblomE.OlssonC.WiklanderK.JohnssonJ. I.et al. (2013). Metabolic scope and interspecific competition in sculpins of Greenland are influenced by increased temperatures due to climate change. PLoS ONE8:e62859. 10.1371/journal.pone.0062859
81
SinclairE. L. E.Noronha de SouzaC. R.WardA. J. W.SeebacherF. (2014). Exercise changes behaviour. Funct. Ecol. 28, 652–659. 10.1111/1365-2435.12198
82
SteffensenJ. F. (1989). Some errors in respirometry of aquatic breathers: how to avoid and correct for them. Fish Physiol. Biochem. 6, 49–59. 10.1007/BF02995809
83
SteinhausenM. F.Fleng SteffensenJ.Gerner AndersenN. (2010). The effects of swimming pattern on the energy use of gilthead seabream (Sparus aurata L.). Mar. Freshw. Behav. Physiol. 43, 227–241. 10.1080/10236244.2010.501135
84
SteinhausenM. F.SteffensenJ. F.AndersenN. G. (2005). Tail beat frequency as a predictor of swimming speed and oxygen consumption of saithe (Pollachius virens) and whiting (Merlangius merlangus) during forced swimming. Mar. Biol. 148, 197–204. 10.1007/s00227-005-0055-9
85
SvendsenJ. C.BanetA. I.ChristensenR. H. B.SteffensenJ. F.AarestrupK. (2013). Effects of intraspecific variation in reproductive traits, pectoral fin use and burst swimming on metabolic rates and swimming performance in the Trinidadian guppy (Poecilia reticulata). J. Exp. Biol. 216, 3564–3574. 10.1242/jeb.083089
86
SvendsenJ. C.GenzJ.AndersonW. G.StolJ. A.WatkinsonD. A.EndersE. C. (2014). Evidence of circadian rhythm, oxygen regulation capacity, metabolic repeatability and positive correlations between forced and spontaneous maximal metabolic rates in lake sturgeon Acipenser fulvescens. PLoS ONE9:e94693. 10.1371/journal.pone.0094693
87
SvendsenJ. C.SteffensenJ. F.AarestrupK.FriskM.EtzerodtA.JydeM. (2012). Excess posthypoxic oxygen consumption in rainbow trout (Oncorhynchus mykiss): recovery in normoxia and hypoxia. Can. J. Zool. 90, 1–11. 10.1139/z11-095
88
SvendsenJ. C.TudoracheC.JordanA. D.SteffensenJ. F.AarestrupK.DomeniciP. (2010). Partition of aerobic and anaerobic swimming costs related to gait transitions in a labriform swimmer. J. Exp. Biol. 213, 2177–2183. 10.1242/jeb.041368
89
TeulierL.OmlinT.WeberJ. M. (2013). Lactate kinetics of rainbow trout during graded exercise: do catheters affect the cost of transport?J. Exp. Biol. 216, 4549–4556. 10.1242/jeb.091058
90
TokićG.YueD. K. P. (2012). Optimal shape and motion of undulatory swimming organisms. Proc. Trans. R. Soc. Lond. B Biol. Sci. 279, 3065–3074. 10.1098/rspb.2012.0057
91
TudoracheC.O'KeefeR. A.BenfeyT. J. (2011). Optimal swimming speeds reflect preferred swimming speeds of brook charr (Salvelinus fontinalis Mitchill, 1874). Fish Physiol. Biochem. 37, 307–315. 10.1007/s10695-011-9498-8
92
TudoracheC.ViaeneP.BlustR.VereeckenH.De BoeckG. (2008). A comparison of swimming capacity and energy use in seven European freshwater fish species. Ecol. Freshw. Fish17, 284–291. 10.1111/j.1600-0633.2007.00280.x
93
VanhataloA.PooleD. C.DiMennaF. J.BaileyS. J.JonesA. M. (2011). Muscle fiber recruitment and the slow component of O2 uptake: constant work rate vs. all-out sprint exercise. Am. J. Physiol. Regul. Integr. Comp. Physiol. 300, R700–R707. 10.1152/ajpregu.00761.2010
94
VanhooydonckB.JamesR. S.TallisJ.AertsP.TadicZ.TolleyK. A.et al. (2014). Is the whole more than the sum of its parts?? Evolutionary trade-offs between burst and sustained locomotion in lacertid lizards. Proc. Trans. R. Soc. Lond. B Biol. Sci. 281:20132677. 10.1098/rspb.2013.2677
95
VidelerJ. J. (1993). Fish Swimming. London, UK: Chapman and Hall.
96
WalkerJ. A.GhalamborC. K.GrisetO. L.McKenneyD.ReznickD. N. (2005). Do faster starts increase the probability of evading predators?Funct. Ecol. 19, 808–815. 10.1111/j.1365-2435.2005.01033.x
97
WilsonA. M.LoweJ. C.RoskillyK.HudsonP. E.GolabekK. A.McNuttJ. W. (2013). Locomotion dynamics of hunting in wild cheetahs. Nature498, 185–192. 10.1038/nature12295
98
WilsonR. S.JamesR. S.Van DammeR. (2002). Trade-offs between speed and endurance in the frog Xenopus laevis: a multi-level approach. J. Exp. Biol. 205, 1145–1152.
Summary
Keywords
aerobic metabolic scope, anaerobic capacity, burst swimming, excess post exercise oxygen consumption, intraspecific variation and trade-off, locomotion, maximum sustained swimming speed, minimum cost of transport
Citation
Svendsen JC, Tirsgaard B, Cordero GA and Steffensen JF (2015) Intraspecific variation in aerobic and anaerobic locomotion: gilthead sea bream (Sparus aurata) and Trinidadian guppy (Poecilia reticulata) do not exhibit a trade-off between maximum sustained swimming speed and minimum cost of transport. Front. Physiol. 6:43. doi: 10.3389/fphys.2015.00043
Received
28 October 2014
Accepted
29 January 2015
Published
17 February 2015
Volume
6 - 2015
Edited by
Leonardo Julián Magnoni, Instituto de Investigaciones Biotecnológicas-Instituto Tecnológico de Chascomús (IIB-INTECH), Argentina
Reviewed by
William Davison, University of Canterbury, New Zealand; Hans Malte, Aarhus University, Denmark
Copyright
© 2015 Svendsen, Tirsgaard, Cordero and Steffensen.
This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) or licensor are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Jon C. Svendsen, Interdisciplinary Center of Marine and Environmental Research, University of Porto, 289 Rua dos Bragas, Porto 4050-123, Portugal e-mail: jos@aqua.dtu.dk
This article was submitted to Aquatic Physiology, a section of the journal Frontiers in Physiology.
Disclaimer
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article or claim that may be made by its manufacturer is not guaranteed or endorsed by the publisher.