Abstract
Phenolic compounds belong to a class of secondary metabolites and are implicated in a wide range of responsive mechanisms in plants triggered by both biotic and abiotic elicitors. In this study, we approached the combinational effects of ethylene and MeJA (methyl jasmonate) on phenolic compounds profiles and gene expressions in the medicinal plant Catharanthus roseus. In virtue of a widely non-targeted metabolomics method, we identified a total of 34 kinds of phenolic compounds in the leaves, composed by 7 C6C1-, 11 C6C3-, and 16 C6C3C6 compounds. In addition, 7 kinds of intermediates critical for the biosynthesis of phenolic compounds and alkaloids were identified and discussed with phenolic metabolism. The combinational actions of ethylene and MeJA effectively promoted the total phenolic compounds, especially the C6C1 compounds (such as salicylic acid, benzoic acid) and C6C3 ones (such as cinnamic acid, sinapic acid). In contrast, the C6C3C6 compounds displayed a notably inhibitory trend in this case. Subsequently, the gene-to-metabolite networks were drawn up by searching for correlations between the expression profiles of 5 gene tags and the accumulation profiles of 41 metabolite peaks. Generally, we provide an insight into the controlling mode of ethylene-MeJA combination on phenolic metabolism in C. roseus leaves.
Introduction
Catharanthus roseus (Madagascar periwinkle) is a medicinal plant producing about more than 150 different terpenoid indole alkaloids (TIAs). Among the many pharmaceutically important TIAs are vinblastine and vincristine, the well-known anticancer agents that have been long in clinical use (van Der Heijden et al., 2004). Nevertheless, report has also suggested that C. roseus contains a wide spectrum of phenolic compounds besides TIAs (Filippini et al., 2003; Mustafa and Verpoorte, 2007; Ferreres et al., 2008), such as flovonoids, caffeic acid, benzoic acid, and cinnamic acid derivatives. These natural products are often involved in signaling pathways triggered as a defense mechanism against biotic and abiotic stresses (Demkura et al., 2010). Furthermore, they contribute to human health by exerting various biological activities including antioxidant, antibacterial and anticancer (Valentao et al., 2001; Sousa et al., 2008).
Phenolics are the most widely distributed metabolites that are involved in the interactions between biology and environments (Garcia-Calderon et al., 2015). In plants, phenolic compounds are synthesized via the phenylpropanoid pathway that begins with the conversion of phenylalanine to cinnamic acid by phenylalanine ammonia lyase (PAL) and hence also compete with the indole alkaloid biosynthesis for common precursor chorismate (Ferreres et al., 2008). Based on the so far gathered knowledge on spatial and temporal complexities associated with phenolic pathway in C. roseus, the involvement of at least 20 coordinately regulated enzymatic steps occurring in different tissues types has been implicated, such as CM (chorismate mutase), ICS (isochorismate synthase), PAL (phenylalanine ammonia-lyase), and C4H (trans-cinnamate 4-monooxygenase) (Mustafa and Verpoorte, 2005; Ferreres et al., 2011). Phenolic compounds are referred to as cyclic compounds with exchangeable hydroxyl groups and are classified based on the number and binding position of convertible hydroxyl groups on the aromatic chain into three major groups: simple phenolic compounds (mass compounds have a C6C1 carbon skeleton, usually with a carboxyl group attached to the aromatic ring, such as benzoic acid), phenolic acid derivatives (derived from phenylalanine, having a C6C3 carbon skeleton, such as caffeic acid), and flavonoids (compounds having C6C3C6 carbon skeleton such as flavonoids and isoflavonoids; Dixon, 2001; Mustafa and Verpoorte, 2007). So far, around 5000 phenolic compounds have been identified and their contents fluctuate depending on environmental conditions and plant species (Mustafa and Verpoorte, 2007; Simirgiotis et al., 2015; Vos et al., 2015). The accumulation of phenolics may also affect other secondary metabolite pathways including alkaloid pathways, since plant defense is a complex system (Mustafa and Verpoorte, 2007; Ferreres et al., 2008; Figure 1).
Figure 1
Identification and structural characterization of phenolic compounds has been well developed and achieved using LC-MS/MS in different plant species (Ferreres et al., 2004; Lin and Harnly, 2007; Dong et al., 2014). The metabolic profiling analysis of phenolic compounds in some plant extracts showed that the C. roseus extracts contained the highest amount of a C6C3 hydroxytyrosol and a C6C1 gallic acid when compared to 26 other plant extracts analyzed. Other phenolics detected in this plant were ferulic acid and vanillic acid. No flavonoids were detected in this study (Proestos et al., 2005). Filippini et al. developed a stable callus of C. roseus producing anthocyanins by continuous cell-aggregate selection (Filippini et al., 2003). Similar anthocyanins were identified by LC-MS/MS metabolic profiling. They were also identified as 3-O-glucosides and 3-O-(6-O-p-coumaroyl) glucosides of petunidin, malvidin, and hirsutidin. Moreover, three caffeoylquinic acids and fifteen flavonol glycosides were identified by the same level of technology (Ferreres et al., 2008). As mentioned above, the potential of developing a platform for qualitative analysis of phenolic compounds has been highlighted in several studies. Therefore, we approached the comprehensively analysis of phenolic compounds located in C. roseus leaves using this method.
The phytohormones ET and methyl jasmonate (MeJA) were reported to elicit secondary metabolites such as alkaloids and phenolic compounds regulating plant growth and adaptation (Demkura et al., 2010; Vanstraelen and Benkova, 2012; Wasternack and Hause, 2013; Cocetta et al., 2015; Pozo et al., 2015). ET acts as an intermediate signaling molecule in elicitor-induced terpenoid indole alkaloids accumulation (Papon et al., 2005; Pan et al., 2015). ET is also able to induce flavonol accumulation alone or interact with auxin (Lewis et al., 2011; Watkins et al., 2014). The addition of MeJA to C. roseus hairy root cultures increased the yields of ajmalicine, serpentine, lochnericine, and hörhammericine (Rijhwani and Shanks, 1998). MeJA also tightly modulated phenolic metabolism and gene expression in blueberry (Cocetta et al., 2015), under solar ultraviolet B radiation (Demkura et al., 2010), or in broccoli sprouts (Carvacho et al., 2014). In some cases, concomitant activation of JA and ET response pathways is required for induction of plant defensive gene, such as PDF1.2 (Penninckx et al., 1998). These reports provided critical insight into the roles of ET or MeJA in the modulation of alkaloids and phenolic compounds, however, their combined effects on phenolic profiles remain into investigation. Here, we mainly focused on the combinational effects of ET and MeJA on phenolic compound profiles and gene expressions in C. roseus leaves in virtue of metabolomics approach.
Results
The combined effects of ET and MeJA on the metabolic profiles of total phenolic compounds
Using LC-QTOF-MS/MS, we developed a novel widely non-targeted metabolomics method for the comprehensive profiling analysis of phenolic compounds in C. roseus. According to the identity check based on raw data and the features of peaks, the target masses of candidate metabolites identified in the profiling process were searched over a narrow + 5 ppm mass window in HMDB, METLIN, and KEGG databases. A total of 34 phenolics that belong to three categories (7 C6C1-, 11 C6C3-, and 16 C6C3C6 compounds; Table 1), and seven other metabolites were identified in our study (Table S2). The relatively level of total phenolic compounds significantly increased (p < 0.01) in plants treated with ET and MeJA compared with those under conditions of control or plus ET only (Figure 2A).
Table 1
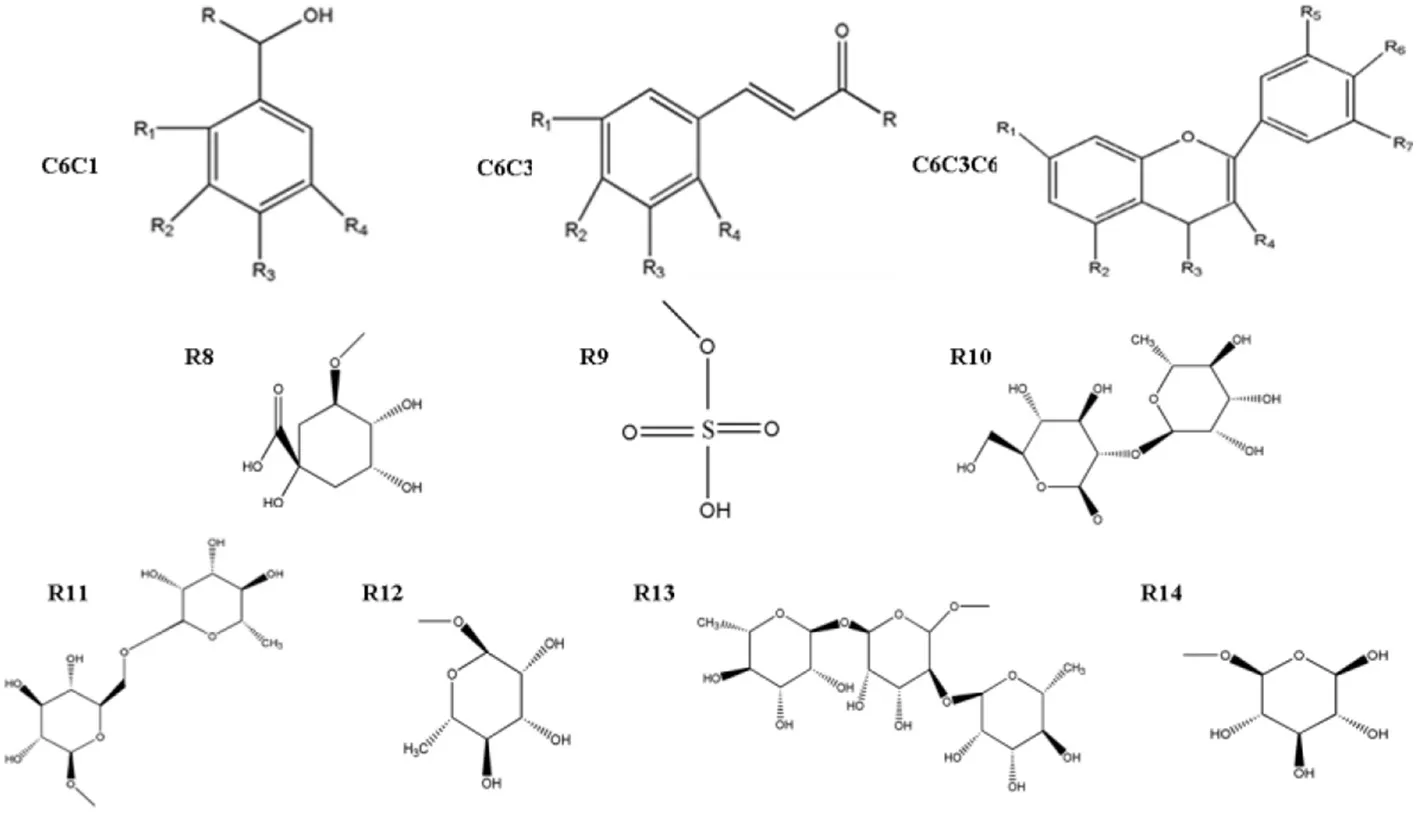 | |||||||||
|---|---|---|---|---|---|---|---|---|---|
| Structure | Derivates | R | R1 | R2 | R3 | R4 | R5 | R6 | R7 |
| C6C1 | Salicylic acid | = O | −OH | ||||||
| Vanillic acid | = O | −OCH3 | −OH | ||||||
| Vanillyl alcohol | −OCH3 | −OH | |||||||
| Syringic acid | = O | OCH3 | −OH | −OCH3 | |||||
| Gallic acid | = O | −OH | −OH | −OH | |||||
| Benzoic acid | = O | ||||||||
| C6C3 | Trans-cinnamric acid | −OH | |||||||
| Cinnamic acid | −OH | ||||||||
| p-Coumanic acid | −OH | −OH | |||||||
| o-Cumamic acid | −OH | −OH | |||||||
| Caffeic acid | −OH | −OH | −OH | ||||||
| Ferulic acid | −OH | −OCH3 | −OH | −OCH3 | |||||
| Sinapic acid | −OH | −OH | |||||||
| Chlorogenic acid | R8 | −OH | −OH | ||||||
| C6C3C6 | Catechin | −OH | −OH | −OH | −OH | −OH | |||
| Naringenin | −OH | −OH | −O | −OH | |||||
| Hirsutidin | −OCH3 | O− | −OH | −OCH3 | −OH | −OCH3 | |||
| Myricetin | −OH | −OH | O | −OH | −OH | −OH | −OH | ||
| Kaempferol | −OH | −OH | −OH | = O | −OH | ||||
| Isoscutellarein | −OH | −OH | −OH | = O | −OH | ||||
| Quercetin | −OH | −OH | = O | −OH | −OH | −OH | |||
| Quercetin-3-sulfate | −OH | −OH | = O | R9 | −OH | −OH | |||
| Malvidin | O− | −OH | – | OH | −OCH3 | −OH | −OCH3 | ||
| Naringin | R10 | −OH | = O | −OH | |||||
| Hesperidin | R11 | −OH | = O | −OH | −OCH3 | ||||
| Nicotiflorin | −OH | −OH | = O | R11 | −OH | ||||
| Petunidin | R13 | −OH | = O | R11 | −OH | −OH | |||
| Quercetin-3-o-rhamnoside | −OH | −OH | = O | R12 | −OH | −OH | |||
| Mauritianin | −OH | −OH | = O | R14 | −OH | ||||
Structure of identified phenolic compounds.
Figure 2
Subsequently, the visualization of the phenolic compounds profiles was performed by hierarchical cluster analysis (HCA; Figure 2B). The results showed that accumulation of phenolic metabolites displayed a clear variation in terms of their abundance upon different treatment. Co-treatment of both hormones contained the highest levels of most phenolics, followed by treatment with ET alone. Based on their responses to different treatment, phenolic profiles could be clearly grouped into three main clusters with six subclusters. Phenolics in cluster I were mainly represented by C6C3C6 compounds with lower levels detected in ET and MeJA, such as hesperidin, petunidin, quercetin, mauritianin, nicotiflorin, malvidin, 5,6-D-7,8,2,3,4-PF, myricetin, naringin, hirsutidin, catechin, isoscutellarein, and naringenin. Major C6C3-(chlorogenic acid, coumaric acid-hexose, sinapic acid, caffeic acid, and cinnamic acid) and C6C1-(vanillic acid and pro acid-O-hexoside) compounds were tightly grouped in subcluster 2. These phenolic displayed higher levels upon the combined action of ET and MeJA compared to the controls. In addition, those in subcluster 3 were mainly represented by compound C6C1-(benzoic acid and vanillyl alcohol) and C6C3C6-(Q-3-O-rhamnoside and kaempferol), also displaying highest levels under the combined condition. Three C6C3-(p-coumaric acid, ferulic acid, and trans-cinnamic acid) and one C6C1-(salicylic acid) in subcluster 6 showed ET-specific accumulation pattern. However, o-coumarica acid, quercetin 3-sulfate, and sinapic acid hexose in subcluster 4 and 5, respectively, showed little sensitivity to ET or/and MeJA, which subcluster 4, 5, and 6 belonging to the last cluster III. The results showed that ET and MeJA combination could mainly promote C6C1- and C6C3-metabolites accumulation, except for C6C3C6 compounds.
The combined effects of ET and MeJA on C6C1-, C6C3-, and C6C3C6-type compounds
Plant phenolic compounds are synthesized via the chorismate and can be majorly divided into three types, including C6C1, C6C3, and C6C3C6. Their specific responses were further clarified. For most of the C6C1- and C6C3-type compounds, there was a significant promotion by the combinational action of ET and MeJA compared with the treatments with control solution and plus ET alone (Figures 3A,B). Among these compounds, vanillyl alcohol in the leaves was found to be elevated 50 times in the presence of ET and MeJA compared with the control case. In contrast, the C6C3C6-type compounds displayed an inhibited accumulation upon ET plus MeJA compared with the cases upon ET alone or control solution (Figure 3C). Furthermore, parts of the intermediates in the phenolic or TIA biosynthetic pathway were identified here, including chorismic acid, phenylpyruvic acid, isochorismic acid, L-tyrosine, L-phenylalaine, tryptophan, and p-hydroxyphenylpyruvic acid. The actions of ET plus MeJA obviously promoted the first three compounds and inhibited the remaining four ones, respectively (Figure 3D). The further analysis of other major C6C3- and C6C3C6 compounds identified in our research showed that cinnamic acid (C6C3) was largely promoted and most of the C6C3C6 compounds were decreased by the combination of ET and MeJA (Figure S1).
Figure 3
The combined effects of ET and MeJA on the gene expression responsible for phenolic or alkaloid biosynthesis
Other than the metabolic analysis, the gene expressions responsible for phenolic biosynthesis (CM: chorismate mutase; ICS: isochorismate synthase; PAL: phenylalanine ammonia lyase; C4H: cytochrome P450 hydroxylation) were compared with that for indole alkaloid pathway (AS: anthranilate synthase). The treatment with ET precursor ACC alone enhanced the expression levels of ICS, AS, PAL, and C4H genes than the control treatment. The inclusion of MeJA plus ET further significantly improved the expression levels of PAL and C4H genes when compared with the treatment with ET alone (Figure 4). In this case, the CM, ICS, and AS gene expressions were not obviously changed.
Figure 4
Integrated response analysis of genes—metabolites network
To facilitate access to the total metabolic profiles, the metabolites, genes and corresponding pathways were imported into Cytoscape for visualization of the network models. The intersection of the networks was done using the advanced network merge function in Cytoscape. Altered metabolites with KEGG from the merged data set were mapped to KEGG reference pathways, and interaction networks were generated in Cytoscape. As seen in Figure 5, the association network of differentially expressed metabolites using Cytoscape was constructed. Modes of action are shown in different colors and shapes. Networks are represented as graphs, where the green diamond, red polygon, and blue square nodes represent metabolites, genes, and related pathways detected, respectively. These closely connected and differentially expressed metabolites are regarded as the target signatures. The significantly overrepresented categories indicated that this emergent new and advanced network was composed by metabolites, genes, and related pathways. Fortunately, for the greater part of the identified metabolites and genes could be included in a network together through indirect interaction or only one intermediate partner. However, a small portion of phenolic derivatives were not found their compound identifiers (IDs) from KEGG, so these could not be included, such as protocatechuic acid-O-hexoside, coumaric acid-hexose, sinapic acid-hexose, quercetin-3-O-rhamnoside, and caffeic acid-hexose. The regulatory patterns of phenolic compounds presented here provide evidence that metabolites are actively involved in multifunctional pathways, and these insights help us to better understand the mechanisms underlying their responses to ET plus MeJA.
Figure 5
Discussions
In this study, we utilized the widely non-targeted metabolomics method for the direct chemical screening of phenolic compounds in the medicinal plants C. roseus. We identified a total of 34 phenolic compounds, mainly including C6C1-, C6C3-, and C6C3C6-type compounds. The result is consistent with previous reports that C. roseus contains the C6C1-, C6C3-, and C6C3C6 compounds (Dixon, 2001; Choi et al., 2004; Mustafa and Verpoorte, 2005, 2007). Our further analysis showed that there were 7, 11, and 16 kinds of compounds, respectively, belonging to C6C1-, C6C3-, and C6C3C6-type. MBRole performs enrichment analysis of a number of annotations of diverse nature coming from KEGG, which are annotated with their associated pathways, enzymes and chemical groups, as well as other interactions (Chagoyen and Pazos, 2011). However, a number of phenolic derivatives did not find these corresponding IDs, such as protocatechuic acid-O-hexoside, coumaric acid-hexose, sinapic acid-hexose, quercetin-3-O-rhamnoside, and caffeic acid-hexose. In addition, the results (Table S3) contain the list of annotations over-represented in the input set with respect to the background set and their associated P < 0.005 (Zhang et al., 2013). Every pathway listed in the MBRole data set with at least on metabolite identified by the MS associated with that pathway was noted, and the MS-identified metabolites were crossly listed with the pathway. Each metabolite could belong to multiple pathways (Figure 5).
The phenolic compounds play diverse and important functions in plant development and defense as reactive oxygen species scavengers, disease resistance inducer (Moreno et al., 1994; Murphy et al., 2000; Mustafa and Verpoorte, 2007; Watkins et al., 2014). In our study, C6C1-, C6C3-, and C6C3C6-compounds comprised 3.58, 51.81, and 44.60% of total phenolic compounds, respectively. It suggested that C6C3C6 compounds are mainly involved in plant flavonoid biosynthesis, functioning as filters against ultraviolet irradiation (Trantas et al., 2015). In contrast, the C6C1 compounds might mainly play an important role as signaling molecules, such as the BA (benzoic acid)-derivatives SA (salicylic acid; Cheynier et al., 2013). Because, the biosynthesis of C6C1 compounds are reported to be most strictly induced by biotic elicitors (Mustafa and Verpoorte, 2007) and these low-molecular weight phenolics occur universally in higher plants (Cheynier et al., 2013). We found that the elevated compounds in the leaves after the co-treatment mainly belonged to either the C6C1 or C6C3-compounds. Evidence is accumulating that components from SA-, JA-, and ET-dependent defense pathways can affect each other's signaling (van Wees et al., 2000). The synergistic cross-talk between JA and ET is known to occur preferentially for the response to necrotrophic pathogens (Wasternack and Hause, 2013) and activate JA-dependent plant defenses against herbivores via alkaloid biosynthesis (Onkokesung et al., 2010). However, SA and its functional analogs suppress JA-dependent defense gene expression, possibly through the inhibition of JA synthesis and action (van Wees et al., 2000; Vos et al., 2015). Conversely, other reports showed that ET and MeJA were shown to stimulate SA action (van Wees et al., 2000). We found that the exogenous ET plus MeJA slightly promoted accumulations of salicylic acid (SA) and its derivatives benzoic acid, isochorismic acid, whereas largely enhanced levels of vanillic acid, vanillyl alcohol, and syringic acid than did ET alone (Figures 3A,B). The compound syringic acid and its precursors (vanillic acid and vanillyl alcohol) compete with SA for benzoic acid as substrate (Mustafa and Verpoorte, 2007), thus, we proposed that ET-JA crosstalk mainly antagonized SA in vivo synthesis and bring carbon flux to vanillic acid accumulation. However, our further clustering analysis found that SA was induced in an ET-specific way, suggesting that SA interplays synergistically with ET.
Our clustering results showed the C6C3 branch pathway metabolites, namely p-coumaric acid, ferulic acid, and trans-cinnamic acid, were grouped together with SA. These phenylpropanoids are derived from phenylalanine catalyzed by PAL enzyme (Mustafa and Verpoorte, 2007; Cheynier et al., 2013). Besides the precursors of C6C1 compounds, the above phenylpropanoids are also precursors of other phenolics, which in many plants act as phytoalexiins or phytoanticipins, such as flavonoids, isoflavonoids (Dixon, 2001) or as physical barrier against pathogen attack, such as lignin (Mustafa and Verpoorte, 2007). Activation of PAL is often considered as a hallmark for elicitation of SAR (systemic acquired resistance) in plants and SA is a necessary and sufficient signal for SAR (Clarke et al., 2000; van Wees et al., 2000). We found that ET alone or plus MeJA increased PAL transcripts and effectively decreased L-phenylalanine level, suggesting SA-dependent SAR process was induced. ET and MeJA are also implicated in induced systemic resistance (ISR; van Wees et al., 2000). ISR pathway may result in induction of different pathways leading to the production of phenolic compounds and/or other secondary metabolites (Glazebrook et al., 2003; Mustafa and Verpoorte, 2007). In contrast to C6C1 and C6C3 compounds, almost all the C6C3C6-metabolites displayed inhibitory accumulation after treatment with ET plus MeJA (Figure 3C and Figure S1B). The biosynthetic pathway of C6C3C6 compounds lead to productions of flavonoids (flavonols) and isoflavonoids (Mustafa and Verpoorte, 2007; Mouradov and Spangenberg, 2014). In vegetative tissues, anthocyanins and other flavonoids usually accumulate transiently, as a plastic response to biotic or abiotic stressors (Del Valle et al., 2015). For example, IAA and ET were able to regulate flavonol biosynthesis through distinct signaling networks and quercetin is revealed to be the flavonol that modulates basipetal auxin transport (Lewis et al., 2011). Our result is consistent with this report, because the reduced quercetin was caused by ET plus MeJA, whilst not by ET alone.
Other than phenolic compounds, a serial of intermediates in the biosynthetic pathway of phenolic and alkaloids were identified in this study (Figure 3D). Among them, the chorismate is a critical substrate for multiple enzymes, such as CM, ICS, and AS, resulting in different secondary metabolic pathway and carbon flux (Mustafa and Verpoorte, 2007; Van Lanen et al., 2008). The restrained expression of CM gene in our transcriptional analysis paralleled with the decreased levels of L-phenylalanine and L-tyrosine. In contrast, the ICS and AS expressions were slightly activated in this case. This evidence suggests that ET and MeJA prefer to elicit benzoic acid-derived phenolic compounds (C6C1), for example, vanillyl alcohol (50-fold increase) in our study. Our subsequent correlation analysis showed that the C6C1 compounds correlated positively with their precursors including chorismate acid and isochorismic acid, but showed a negative correlation with the tryptophan (Figure S2). Tryptophan is a pivotal precursor for indole alkaloids in relation with AS enzyme function and found to be reduced in our research. It was interesting that AS expression, however, was activated in this case. We interpret this result that the combined action of ET and MeJA simultaneously promotes the indole alkaloid and phenolic pathways, whilst tryptophan was rapidly consumed for indole alkaloid synthesis (Choi et al., 2004; Chung et al., 2007). Phenylpyruvic acid and p-hydroxyphenylpyruvic acid are precursors of L-phenylalanine and L-tyrosine, respectively, essential for C6C3 and C6C3C6 biosynthesis (Mustafa and Verpoorte, 2007). The p-hydroxyphenylpyruvic acid largely decreased in our research under the combined action of ET and MeJA, interpreting globally inhibited levels of C6C3C6 compounds. The gene expression level of PAL was both elevated obviously in this study, leading to the conversion of phenylalanine into cinnamic acid, which is a precursor for several phenolic compounds such as cinnamic acid, p-coumaric acid, caffeic acid, naringenin, and syringlc acid. The expression of C4H responsible for the hydroxylation at the C-4 position of cinnamic acid to form p-coumaric acid significantly increased, thereby contributing to the C6C3 compounds synthesis (Hotze et al., 1995).
Conclusions
In conclusion, we report a comprehensive profiling analysis of phenolic compounds in response to the combinational action of ET and MeJA. Application of a widely non-targeted metabolomics method facilitated the identification of a total of 34 phenolic compounds and 7 intermediates for biosynthesis of phenolic compounds and indole terpenoid alkaloids (TIAs) using LC-MS/MS. Gene-to-metabolite networks were drawn up by searching for correlations between the expression profiles of 5 gene tags and the accumulation profiles of 41 metabolite peaks. The network revealed that the different branches of phenolic compounds biosynthesis and various other metabolic pathways are tightly regulated by ET-MeJA combination. Especially, the ET-MeJA cross-talk mainly promote C6C1- and C6C3-type, whilst inhibit C6C3C6-type phenolic compounds. Thus, this study provides an insight into the controlling mode of ET-MeJA combination on phenolic metabolism in C. roseus leaves. However, the underlying mechanisms in this induction process remain to be unraveled.
Materials and methods
Chemicals
All chemicals were of analytical reagent grade. Gradient grades of methanol, acetonitrile and acetic acid were purchased from Merck Company, German (http://www.merck-chemicals.com). Water was doubly deionized with a Milli-Q water purification system (Milford, MA, USA). Ethephon used to release ET and methyl jasmonate (MeJA) were both obtained from Sigma-Aldric (St. Louis, MO, USA).
Plant material, growth conditions, and sample preparation
C. roseus seeds were planted in pots containing perlite and kept moistened until the seeds had germinated, and then irrigated with 1/2 strength Hoagland's solution (Pan et al., 2015). On the basis of process conditions screened, the concentrations of ethephon and MeJA used for treatment were 30 μM and 150 μM, respectively. Seedlings were used for treatments 30 d after cultivation with roots subjected to the 1/2 strength Hoagland's solution (control, T1), or plus 30 μM ethephon alone (T2) or 30 μM ethephon + 150 μM MeJA (T3). The total 30 individual plants were randomly selected and equally subjected into the conditions of T1, T2, and T3. The experiments were biologically repeated three times. The leaves of plants were harvested, 3 d after treatment for analysis of phenolic compounds and 4 h after treatment for analysis of gene expressions.
LC-ESI-QTOF/MS analysis of phenolic compounds
For phenolic compound analysis, leaves were pulverized by grinding instrument (MM 400, Retsch, GmbH, Haan, Germany), and 50 mg tissue aliquots were extracted with 1.0 mL 70% aqueous methanol containing 0.1 mg/L lidocaine for water-soluble metabolites at 4°C overnight with three times vortexing. The extracts were clarified by centrifugation, combined, evaporated, and then filtered through 0.22 μm nylon membranes (SCAA 104; ANPEL http://www.anpel.com.cn/) before LC-MS analysis. Samples were analyzed using a liquid chromatography (LC) system coupled to a (QTOF) tandem mass spectrometer via electrospray ionization (ESI) interface (Agilent 6520) (Agilent Technologies, Santa Clara, CA, USA). Sample extracts were separated through a reversed phase on a Shim-pack LC column (VP-ODS C18 pore size 5.0 μm, 2*150 mm). The mobile phase consisted of solvent A and B. Solvent A contained 0.04% acetic acid in water, and solvent B 0.04% acetic acid in acetonitrile. The following gradient was used with a flow rate of 0.5 mL/min: 0–20 min, 5%B–95%B; 20–22.1 min: 95%B–5%B; 22.1–28 min: 5%B–5%B. Blank measurement with the initial solvent was carried out after each HPLC run. Injection volume and column temperature were set to 5 μL and 40°C, respectively. The optimized parameters for positive ion electrospray were as follows: capillary temperature of 350°C; curtain gas pressure of 40 psi; capillary voltage of 3500 V, fragmentation voltage of 135 V. The instrument was tuned prior to each batch run. A full-scan ranging between 50–1000 m/z was conducted with a scan time of 1 s and an interscan delay of 0.1 s in centered mode. The peak detection and matching were performed by Mass Hunter Qualitative (MHQ version B.03.01) and Mass Profiler Professional (MPP, version B.02.01) (Both from Agilent Technologies, Santa Clara, CA, USA). Metabolic features with mass, retention time, and abundance were obtained.
RNA isolation and quantitative real time PCR analysis
For gene expression analysis, the total leaf RNA was extracted by TRIZOL reagent (Invitrogen). DNA contamination was removed using Dnase I following the instructions provided by the manufacturer (TaKaRa, Japan). DNA and RNA purity were observed using 1% agarose gel electrophoresis and RNA concentration was determined using a Nanodrap spectrophotometer (Thermo). cDNA was synthesized from total RNA (2 μg) using ReverTra Ace QPCR RT Kit (TOYOBO, Japan) according to the manufacturer's instructions, using oligo (dT) as the primer. qRT- PCR analysis using cDNA as template and gene-specific primers was performed using a SYBR Premix Ex Taq (TaKaRa, Japan) with an initial denaturation at 95°C for 30 s, followed by 35 cycles at 94°C for 30 s, 56°C for 30 s and 72°C for 30 s. Amplification, detection, and data analysis were carried out on a Rotor-Gene 6000 real-time rotary analyzer (Corbett Life Science, Sydney, Australia). The primers used were 5′-GCG AAC ATT TGC AGA TCC AT-3′ and 5′-GGC CGA TTT GTT ATT GTT CC-3′ for AS; 5′-GGC CAC CAA GAT GAT CGA-3′ and 5′-CAA TGG CCA ATC TTG CAT TG-3′ for PAL; 5′-GCC GAT TCT CTG TAT CAC TAT C -3′ and 5′-ATG ATT AAA ATG ATC TTG GCT TT-3′ for C4H; 5′-CGA TTT GTT GAA ATT GCA GAC G-3′ and 5′-ATT GCA GAC GAT CGT TTA ACT C-3′ for CM; 5′-ATT GCA GAC GAT CGT TTA ACT C-3′ and 5′-TTC CTC GGT CAA ACA TTT CG-3′ for ICS; (from ExPlant Technologies B.V.) (Table S1). These were repeated three times for each sample to ensure the reproducibility of results. Ribosomal protein subunit 9 (Rsp9) 5′-GAG GGC CAA AAC AAA CTT GA-3′ and 5′-CCC TTA TGT GCC TTT GCC TA-3′ was used as an internal control to evaluate all C. roseus plants.
Statistical analysis
Metabolic pathways were performed in the Metaboanalyst web portal (http://www.metaboanalyst.ca) and MBRole (http://csbg.cnb.csic.es/mbrole). The pathways of metabolites were carried out on database sources including the KEGG (http://www.genome.jp/kegg/) to identify the top affected metabolic pathways and facilitate further metabolites interpretation. The metabolites and corresponding pathways were imported into Cytoscape software (v. 3.1.0) for visualization of the network models. The intersection of the networks was done using the advanced network merge function in Cytoscape. Pearson's correlation coefficients were calculated between metabolites and genes by SPSS 17.0. The Student's t-test and Tukey's test were used for mean value comparison. A total of 34 phenolic compounds were used for hierarchical clustering analysis by R (http://www.r-project.org/) to analyze phenolic profiles in response hormones.
Statements
Author contributions
Conceived and designed the experiments: ZT; Performed the experiments: YL and YW; Analyzed the data: ZZ and JL; Wrote the paper: TE and JL.
Acknowledgments
We sincerely thank Prof. Yu Fang for his constructive and critical comments on this manuscript. This study was financially supported by National Natural Science Foundation of China (31570520).
Conflict of interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Supplementary material
The Supplementary Material for this article can be found online at: http://journal.frontiersin.org/article/10.3389/fphys.2016.00217
- TIAs
terpenoid indole alkaloids
- ABA
abscisic acid
- ET
ethylene
- MeJA
methyl jasmonate
- PAL
phenylalanine ammonia lyase
- CM
chorismate mutase
- ICS
isochorismate synthase
- AS
anthranilate synthase
- C4H
cytochrome P450 hydroxylation
- 5.6-D-7,8,2,3,4-PF
5,6′-dihydroxy-7, 8,2′,3′, 4′-Pentamethoxyflavone
- Pro acid-O-hexoside
Protocatechuic acid-O-hexoside
- Q-3-O-rhamnoside
Quercetin-3-O-rhamnoside
- LC-MS/MS
liquid chromatography tandem mass spectrometry
- ESI
electrospray ionization.
Abbreviations
References
1
CarvachoH. B.PerezC.ZunigaG.MahnA. (2014). Effect of methyl jasmonate, sodium selenate and chitosan as exogenous elicitors on the phenolic compounds profile of broccoli sprouts. J. Sci. Food Agric.94, 2555–2561. 10.1002/jsfa.6596
2
ChagoyenM.PazosF. (2011). MBRole: enrichment analysis of metabolomic data. Bioinformatics27, 730–731. 10.1093/bioinformatics/btr001
3
CheynierV.ComteG.DaviesK. M.LattanzioV.MartensS. (2013). Plant phenolics: recent advances on their biosynthesis, genetics, and ecophysiology. Plant Physiol. Biochem.72, 1–20. 10.1016/j.plaphy.2013.05.009
4
ChoiY. H.TapiasE. C.KimH. K.LefeberA. W.ErkelensC.VerhoevenJ. T.et al. (2004). Metabolic discrimination of Catharanthus roseus leaves infected by phytoplasma using 1H-NMR spectroscopy and multivariate data analysis. Plant Physiol.135, 2398–2410. 10.1104/pp.104.041012
5
ChungI. M.HongS. B.PeeblesC. A.KimJ. A.SanK. Y. (2007). Effect of the engineered indole pathway on accumulation of phenolic compounds in Catharanthus roseus hairy roots. Biotechnol. Prog.23, 327–332. 10.1021/bp060258e
6
ClarkeJ. D.VolkoS. M.LedfordH.AusubelF. M.DongX. (2000). Roles of salicylic acid, jasmonic acid, and ethylene in cpr-induced resistance in arabidopsis. Plant Cell12, 2175–2190. 10.1105/tpc.12.11.2175
7
CocettaG.RossoniM.GardanaC.MignaniI.FerranteA.SpinardiA. (2015). Methyl jasmonate affects phenolic metabolism and gene expression in blueberry (Vaccinium corymbosum). Physiol. Plant153, 269–283. 10.1111/ppl.12243
8
Del ValleJ. C.BuideM. L.Casimiro-SoriguerI.WhittallJ. B.NarbonaE. (2015). On flavonoid accumulation in different plant parts: Variation patterns among individuals and populations in the shore campion (Silene littorea). Front. Plant Sci.6:939. 10.3389/fpls.2015.00939
9
DemkuraP. V.AbdalaG.BaldwinI. T.BallareC. L. (2010). Jasmonate-dependent and -independent pathways mediate specific effects of solar ultraviolet B radiation on leaf phenolics and antiherbivore defense. Plant Physiol.152, 1084–1095. 10.1104/pp.109.148999
10
DixonR. A. (2001). Natural products and plant disease resistance. Nature411, 843–847. 10.1038/35081178
11
DongX.ChenW.WangW.ZhangH.LiuX.LuoJ. (2014). Comprehensive profiling and natural variation of flavonoids in rice. J. Integr. Plant Biol.56, 876–886. 10.1111/jipb.12204
12
FerreresF.FigueiredoR.BettencourtS.CarqueijeiroI.OliveiraJ.Gil-IzquierdoA.et al. (2011). Identification of phenolic compounds in isolated vacuoles of the medicinal plant Catharanthus roseus and their interaction with vacuolar class III peroxidase: an H(2)O(2) affair?J. Exp. Bot.62, 2841–2854. 10.1093/jxb/erq458
13
FerreresF.LlorachR.Gil-IzquierdoA. (2004). Characterization of the interglycosidic linkage in di-, tri-, tetra- and pentaglycosylated flavonoids and differentiation of positional isomers by liquid chromatography/electrospray ionization tandem mass spectrometry. J. Mass Spectrom.39, 312–321. 10.1002/jms.586
14
FerreresF.PereiraD. M.ValentaoP.AndradeP. B.SeabraR. M.SottomayorM. (2008). New phenolic compounds and antioxidant potential of Catharanthus roseus. J. Agric. Food Chem.56, 9967–9974. 10.1021/jf8022723
15
FilippiniR.CaniatoR.PiovanA.CappellettiE. M. (2003). Production of anthocyanins by Catharanthus roseus. Fitoterapia74, 62–67. 10.1016/S0367-326X(02)00296-4
16
Garcia-CalderonM.Pons-FerrerT.MrazovaA.Pal'ove-BalangP.VilkovaM.Perez-DelgadoC. M.et al. (2015). Modulation of phenolic metabolism under stress conditions in a Lotus japonicus mutant lacking plastidic glutamine synthetase. Front. Plant Sci.6:760. 10.3389/fpls.2015.00760
17
GlazebrookJ.ChenW.EstesB.ChangH. S.NawrathC.MetrauxJ. P.et al. (2003). Topology of the network integrating salicylate and jasmonate signal transduction derived from global expression phenotyping. Plant J.34, 217–228. 10.1046/j.1365-313X.2003.01717.x
18
HotzeM.SchroderG.SchroderJ. (1995). Cinnamate 4-hydroxylase from Catharanthus roseus, and a strategy for the functional expression of plant cytochrome P450 proteins as translational fusions with P450 reductase in Escherichia coli. FEBS Lett.374, 345–350. 10.1016/0014-5793(95)01141-Z
19
LewisD. R.RamirezM. V.MillerN. D.VallabhaneniP.RayW. K.HelmR. F.et al. (2011). Auxin and ethylene induce flavonol accumulation through distinct transcriptional networks. Plant Physiol.156, 144–164. 10.1104/pp.111.172502
20
LinL. Z.HarnlyJ. M. (2007). A screening method for the identification of glycosylated flavonoids and other phenolic compounds using a standard analytical approach for all plant materials. J. Agric. Food Chem.55, 1084–1096. 10.1021/jf062431s
21
MorenoP. R.van der HeijdenR.VerpoorteR. (1994). Elicitor-mediated induction of isochorismate synthase and accumulation of 2,3-dihydroxy benzoic acid in Catharanthus roseus cell suspension and shoot cultures. Plant Cell Rep.14, 188–191. 10.1007/BF00233788
22
MouradovA.SpangenbergG. (2014). Flavonoids: a metabolic network mediating plants adaptation to their real estate. Front. Plant Sci.5:620. 10.3389/fpls.2014.00620
23
MurphyA.PeerW. A.TaizL. (2000). Regulation of auxin transport by aminopeptidases and endogenous flavonoids. Planta211, 315–324. 10.1007/s004250000300
24
MustafaN. R.VerpoorteR. (2005). Chorismate derived C6C1 compounds in plants. Planta222, 1–5. 10.1007/s00425-005-1554-0
25
MustafaN.VerpoorteR. (2007). Phenolic compounds in Catharanthus roseus. Phytochem. Rev.6, 243–258. 10.1007/s11101-006-9039-8
26
OnkokesungN.BaldwinI. T.GalisI. (2010). The role of jasmonic acid and ethylene crosstalk in direct defense of Nicotiana attenuata plants against chewing herbivores. Plant Signal. Behav.5, 1305–1307. 10.4161/psb.5.10.13124
27
PanY. J.LiuJ.GuoX. R.ZuY. G.TangZ. H. (2015). Gene transcript profiles of the TIA biosynthetic pathway in response to ethylene and copper reveal their interactive role in modulating TIA biosynthesis in Catharanthus roseus. Protoplasma252, 813–824. 10.1007/s00709-014-0718-9
28
PaponN.BremerJ.VansiriA.AndreuF.RideauM.CrecheJ. (2005). Cytokinin and ethylene control indole alkaloid production at the level of the MEP/terpenoid pathway in Catharanthus roseus suspension cells. Planta Med.71, 572–574. 10.1055/s-2005-864163
29
PenninckxI. A.ThommaB. P.BuchalaA.MétrauxJ. P.BroekaertW. F. (1998). Concomitant activation of jasmonate and ethylene response pathways is required for induction of a plant defensin gene in Arabidopsis. Plant Cell10, 2103–2113. 10.1105/tpc.10.12.2103
30
PozoM. J.López-RáezJ. A.Azcón-AguilarC.García-GarridoJ. M. (2015). Phytohormones as integrators of environmental signals in the regulation of mycorrhizal symbioses. New Phytol.205, 1431–1436. 10.1111/nph.13252
31
ProestosC.ChorianopoulosN.NychasG. J.KomaitisM. (2005). RP-HPLC analysis of the phenolic compounds of plant extracts. investigation of their antioxidant capacity and antimicrobial activity. J. Agric. Food Chem.53, 1190–1195. 10.1021/jf040083t
32
RijhwaniS. K.ShanksJ. V. (1998). Effect of elicitor dosage and exposure time on biosynthesis of indole alkaloids by Catharanthus roseus hairy root cultures. Biotechnol. Prog.14, 442–449. 10.1021/bp980029v
33
SimirgiotisM. J.BenitesJ.ArecheC.SepulvedaB. (2015). Antioxidant capacities and analysis of phenolic compounds in three endemic nolana species by HPLC-PDA-ESI-MS. Molecules20, 11490–11507. 10.3390/molecules200611490
34
SousaC.ValentaoP.FerreresF.SeabraR. M.AndradeP. B. (2008). Tronchuda cabbage (Brassica oleracea L. var. costata DC): scavenger of reactive nitrogen species. J. Agric. Food Chem.56, 4205–4211. 10.1021/jf072740y
35
TrantasE. A.KoffasM.XuP.VerveridisF. (2015). When plants produce not enough or at all: metabolic engineering of flavonoids in microbial hosts. Front. Plant Sci.6:7. 10.3389/fpls.2015.00007
36
ValentaoP.FernandesE.CarvalhoF.AndradeP. B.SeabraR. M.BastosM. L. (2001). Antioxidant activity of Centaurium erythraea infusion evidenced by its superoxide radical scavenging and xanthine oxidase inhibitory activity. J. Agric. Food Chem.49, 3476–3479. 10.1021/jf001145s
37
van Der HeijdenR.JacobsD. I.SnoeijerW.HallardD.VerpoorteR. (2004). The Catharanthus alkaloids: pharmacognosy and biotechnology. Curr. Med. Chem.11, 607–628. 10.2174/0929867043455846
38
Van LanenS. G.LinS.ShenB. (2008). Biosynthesis of the enediyne antitumor antibiotic C-1027 involves a new branching point in chorismate metabolism. Proc. Natl. Acad. Sci. U.S.A.105, 494–499. 10.1073/pnas.0708750105
39
VanstraelenM.BenkovaE. (2012). Hormonal interactions in the regulation of plant development. Annu. Rev. Cell Dev. Biol.28, 463–487. 10.1146/annurev-cellbio-101011-155741
40
van WeesS. C.de SwartE. A.van PeltJ. A.van LoonL. C.PieterseC. M. (2000). Enhancement of induced disease resistance by simultaneous activation of salicylate- and jasmonate-dependent defense pathways in Arabidopsis thaliana. Proc. Natl. Acad. Sci. U.S.A.97, 8711–8716. 10.1073/pnas.130425197
41
VosI. A.MoritzL.PieterseC. M.Van WeesS. C. M. (2015). Impact of hormonal crosstalk on plant resistance and fitness under multi-attacker conditions. Front. Plant Sci.6:639. 10.3389/fpls.2015.00639
42
WasternackC.HauseB. (2013). Jasmonates: biosynthesis, perception, signal transduction and action in plant stress response, growth and development. An update to the 2007 review in Annals of Botany. Ann. Bot.111, 1021–1058. 10.1093/aob/mct067
43
WatkinsJ. M.HechlerP. J.MudayG. K. (2014). Ethylene-induced flavonol accumulation in guard cells suppresses reactive oxygen species and moderates stomatal aperture. Plant Physiol.164, 1707–1717. 10.1104/pp.113.233528
44
ZhangA. H.WangP.SunH.YanG. L.HanY.WangX. J. (2013). High-throughput ultra-performance liquid chromatography-mass spectrometry characterization of metabolites guided by a bioinformatics program. Mol. Biosyst.9, 2259–2265. 10.1039/c3mb70171a
Summary
Keywords
Catharanthus roseus, phenolic compounds, ethylene, methy jasmonate, non-targeted metabolomics
Citation
Liu J, Liu Y, Wang Y, Zhang Z-H, Zu Y-G, Efferth T and Tang Z-H (2016) The Combined Effects of Ethylene and MeJA on Metabolic Profiling of Phenolic Compounds in Catharanthus roseus Revealed by Metabolomics Analysis. Front. Physiol. 7:217. doi: 10.3389/fphys.2016.00217
Received
02 December 2015
Accepted
24 May 2016
Published
07 June 2016
Volume
7 - 2016
Edited by
M. Iqbal R. Khan, International Rice Research Institute, Philippines
Reviewed by
Akira Oikawa, Yamagata University, Japan; Péter Poór, University of Szeged, Hungary
Updates
Copyright
© 2016 Liu, Liu, Wang, Zhang, Zu, Efferth and Tang.
This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) or licensor are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Thomas Efferth efferth@uni-mainz.de;
This article was submitted to Plant Physiology, a section of the journal Frontiers in Physiology
Disclaimer
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article or claim that may be made by its manufacturer is not guaranteed or endorsed by the publisher.