 Marie-Laure Specq1Mélisande Bourgoin-Heck1Nathalie Samson1
Marie-Laure Specq1Mélisande Bourgoin-Heck1Nathalie Samson1 François Corbin2
François Corbin2 Christian Gestreau3Maxime Richer4Hazim Kadhim5
Christian Gestreau3Maxime Richer4Hazim Kadhim5 Jean-Paul Praud1*
Jean-Paul Praud1*- 1Neonatal Respiratory Research Unit, Departments of Pediatrics and Pharmacology - Physiology, Université de Sherbrooke, Sherbrooke, QC, Canada
- 2Department of Biochemistry, Université de Sherbrooke, Sherbrooke, QC, Canada
- 3Aix-Marseille University, CNRS, CRN2M, Marseille, France
- 4Department of Pathology, Université de Sherbrooke, Sherbrooke, QC, Canada
- 5Neuropathology Unit and Reference Center for Neuro-Muscular Pathology, Brugmann University Hospital and Childrens' Hospital (CHU Brugmann - HUDERF), Université Libre de Bruxelles, Brussels, Belgium
Hyperbilirubinemia (HB) occurs in 90% of preterm newborns. Moderate HB can induce acute neurological disorders while severe HB has been linked to a higher incidence of apneas of prematurity. The present study aimed to test the hypothesis that even moderate HB disrupts cardiorespiratory control in preterm lambs. Two groups of preterm lambs (born 14 days prior to term), namely control (n = 6) and HB (n = 5), were studied. At day 5 of life, moderate HB (150–250 μmol/L) was induced during 17 h in the HB group after which cardiorespiratory control as well as laryngeal and pulmonary chemoreflexes were assessed during baseline recordings and during hypoxia. Recordings were repeated 72 h after HB induction, just before euthanasia. In addition, neuropathological studies were performed to investigate for cerebral bilirubin deposition as well as for signs of glial reactivity in brainstem structures involved in cardiorespiratory control. Results revealed that sustained and moderate HB: (i) decreased baseline respiratory rate and increased the time spent in apnea; (ii) blunted the cardiorespiratory inhibition normally observed during both laryngeal and pulmonary chemoreflexes; and (iii) increased heart rate in response to acute hypoxia. These acute physiological changes were concurrent with an activation of Alzheimer type II astrocytes throughout the brain, including the brainstem. Concomitantly, bilirubin deposits were observed in the leptomeninges, but not in brain parenchyma. While most cardiorespiratory alterations returned to normal 72 h after HB normalization, the expression of glial fibrillary acid protein (GFAP) and ionized calcium binding adaptor molecule 1 (Iba1) was still increased within the nucleus tractus solitarius. In conclusion, moderate and sustained HB in preterm lambs induced cardiorespiratory alterations, the latter of which were associated with neurohistopathological changes. These changes are indicative of an inflammatory response in the brainstem neuroanatomical substrates involved in cardiorespiratory control.
Introduction
Hyperbilirubinemia (HB) is one of the most common problems encountered in neonates, particularly preterms (Watchko, 2009). Neurotoxicity is the major consequence of neonatal HB and includes a host of neurological dysfunctions such as somnolence, abnormal muscle tone, feeding difficulties, and auditory dysfunction. Kernicterus, the most severe form of neurotoxicity, is responsible for long-term neurological sequelae, and although now very rare, milder forms of acute bilirubin-induced neurological dysfunction still remain frequent (Johnson and Bhutani, 2001; Mazeiras et al., 2012). A few studies have also suggested an association between HB and apneas and/or bradycardias in newborn infants (Johnson et al., 2009; Amin et al., 2014; Amin and Wang, 2015). Furthermore, a study in rat pups provided evidence that acute HB influences respiratory control; however, the degree of HB was much higher and more transient than levels clinically encountered in neonates (Mesner et al., 2008). More recently, we showed a decreased efficiency in bottle-feeding and an impaired swallowing-breathing coordination in a novel experimental model of sustained and moderate HB in preterm lambs (Bourgoin-Heck et al., 2015). Overall, the above studies suggest that HB induces respiratory alterations in the neonate. The main objective of the present study was to test the hypothesis that moderate HB, at a level similar to that commonly observed in newborn infants, induces acute and delayed alterations in cardiorespiratory control. In addition, we aimed to further elucidate the mechanisms involved in these alterations by assessing whether neurohistopathological alterations are present in the region of the nucleus tractus solitarius, which is devoted to central processing of cardiorespiratory afferent inputs (Kubin et al., 2006).
Materials and Methods
The study protocol was approved by the Ethics Committee for Animal Care and Experimentation of the Université de Sherbrooke (protocol # 260–10).
Preterm Lamb Model of Moderate Hyperbilirubinemia
Sixteen preterm lambs were born vaginally on gestational day 133 (term = 147 days) as previously described (Boudaa et al., 2013). Results on the effects of HB on bottle-feeding efficiency and nutritive swallowing-breathing coordination obtained in the same preterm lambs have previously been published (Bourgoin-Heck et al., 2015). Premature labor was induced by mifepristone (8 mg/kg), preceded by betamethasone (12 mg × 2). A unique preterm lamb model for moderate HB was designed for the purposes of the study (Bourgoin-Heck et al., 2015). Briefly at birth, lambs were randomly assigned to either the HB group or control group. On postnatal day 5, HB was induced by an intravenous infusion of 20 mg/kg of the bilirubin solution, stabilized with albumin and diluted in Ringer's Lactate buffer (Hansen et al., 1992; Mesner et al., 2008; Bourgoin-Heck et al., 2015), whereas control lambs received a bilirubin-free solution. Bilirubinemia was maintained between 150 and 250 μmol/L for a period of 17 h.
Neurological consequences of HB were characterized based on the “Lamb Acute Bilirubin Encephalopathy score” as well as on consistent electroencephalogram changes, as previously described (Bourgoin-Heck et al., 2015). Accordingly, the presence of acute bilirubin encephalopathy was assessed according to published guidelines for the detection of HB in newborns (American Academy of Pediatrics Subcommittee on Hyperbilirubinemia, 2004). Clinical examinations (consciousness, muscle tone, and audition) were repeated every 2 h throughout bilirubin infusion, then twice daily until the end of the experiment. Detailed results of the LABE score are presented for every lamb in Table 1 of the digital supplement of Bourgoin-Heck et al. (2015). In addition to the clinical detection of acute bilirubin encephalopathy, an electroencephalogram (EEG) was recorded for 6 h during bilirubin infusion and again 6 h at D3, in particular for detecting alterations in sleep organization or epileptiform discharges in HB lambs.
Finally, daily monitoring included vital signs (heart rate, respiratory rate, pulse oximetry, rectal temperature), weight and blood samples for albumin, glucose, bilirubin, and arterial blood gases.
Instrumentation and Recording Equipment
Animals were chronically instrumented on postnatal day 5 and thereafter, and included pairs of needle-electrodes inserted subcutaneously: (i) above the occipital cortex for electroencephalogram (EEG); (ii) near to the right eye socket for electrooculogram (EOG); and (iii) on both sides of the thorax for electrocardiogram (ECG). In addition, the following were implanted: (iv) catheters into both jugular veins (IV infusion and injection; blood sampling); (v) a 5 Fr naso-pharyngeal catheter with its tip positioned 0.5 cm above the posterior border of the soft palate [induction of laryngeal chemoreflexes (LCR)]; (vi) elastic bands around the thorax and abdomen for respiratory inductance plethysmography (respiratory movements; NIMS, Miami Beach FL, USA); (vii) a Masimo pulse oximeter sensor at the tail base [arterial oxygen hemoglobin saturation (SpO2)]. Lastly, in order to assess the response to hypoxia, a custom-built nasal mask molded to fit the lamb's muzzle (Samson et al., 2005) was connected to a pneumotachograph (flow transducer 47304A Hewlett-Packard, Hewlett-Packard Company, Palo-Alto, CA, USA) and a 2-way system which provided either medical air or a gas mixture of 10% O2 in air. Arterial blood gases were also measured in two lambs from each group during hypoxia from an arterial brachial catheter surgically inserted under local anesthesia, prior to recordings.
Leads from all sensors were connected to our custom-built radio telemetry transmitters (Samson et al., 2011). All signals were continuously recorded using AcqKnowledge software (version 4.1, Biopac Systems Canada Inc., Montreal, Qc, Canada) and the lambs were filmed using a web-cam.
Design of the Study
All preterm lambs were housed and cared for with their dams in our animal quarters until the experimental day. Polysomnographic recordings were first performed on postnatal day 5, named experimental day 0 (D0), to assess the immediate effects of moderate HB on baseline cardiac and respiratory activity, as well as on cardiac and respiratory responses to several chemical stimuli, including laryngeal and pulmonary chemoreflexes and hypoxia (see below and Figure 1). Recordings were repeated 72 h after HB induction (D3) to assess the delayed effects of moderate HB.
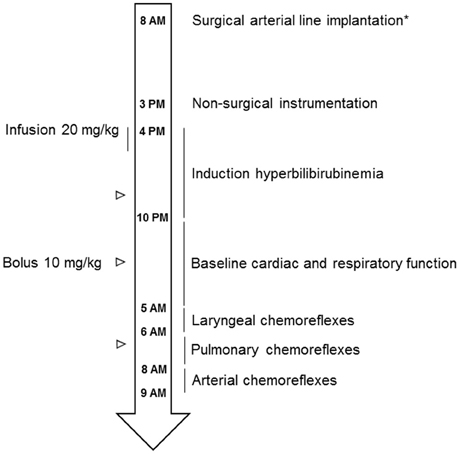
Figure 1. Schematic drawing of the experimental design. *Surgical arterial line implantation was performed in only 4 lambs (2 control and 2 HB lambs).
Baseline Cardiac and Respiratory Function
States of alertness, ECG, respiratory movements, hemoglobin O2 saturation, and pulse rate were continuously recorded for a period of 7 h from 10:00 p.m. to 5:00 a.m., immediately after reaching the bilirubinemia target. During these polysomnographic recordings, the non-sedated lambs were placed in a Plexiglas chamber (1.2 × 1.2 × 1 m, corresponding to guidelines from the Canadian Council for Animal Care) in which they could move freely. After 3 h of recording, they were temporarily returned to their mothers in order to feed before being brought back to the Plexiglass chamber for the remainder of the recording session.
Cardiorespiratory Responses to Chemical Challenges
Laryngeal chemoreflexes
Immediately following baseline cardiac and respiratory activity recordings, LCR were induced by two separate injections of 2 ml of hydrochloric acid (HCl, pH = 2, diluted in saline) through the naso-pharyngeal catheter during NREM sleep, after a 3 min baseline recording. Each lamb was given at least 30 min of recovery time between the two injections. Events such as agitation, cough, arousal and/or full awakening were noted by an observer present throughout the recordings, in addition to the continuous video recording.
Pulmonary chemoreflexes
Pulmonary chemoreflexes (Diaz et al., 1999) were induced during NREM sleep, 30 min after LCR assessment. Following a 3 min baseline recording, lambs were first given an IV bolus injection of 1 ml of vehicle (i.e., 10% Tween 80–10% ethanol—80% saline). After a five-min delay, two pulmonary chemoreflexes were induced at a 30 min interval by central IV injections of 25 μg/kg capsaicin (diluted in vehicle; Sigma Chemical, St-Louis, MO, USA).
Hypoxia
At the end of each experimental day, preterm lambs were exposed to 10% poïkilocapnic hypoxia. Following a 3 min baseline recording in normoxia (21% O2) with the nasal mask on, lambs were exposed to 10% O2 for 10 min. Thereafter, lambs were returned to air for a further 5 min recording. Rectal temperature and arterial blood samples were taken in order to assess pH, PaO2, and PaCO2 at the beginning and end of the 3-min baseline, as well as after 2, 6, and 10 min of hypoxia, and finally 1, 3, and 5 min after the return to normoxia.
Neurohistopathological Studies of the Brain
Following euthanasia at the end of experimental day 3 (D3), the brains of all 11 lambs were immediately perfused through the carotid arteries with cold 0.1 M phosphate buffered saline (pH 7.4) followed by 4% paraformaldehyde diluted in PBS. The brain was removed en bloc and post-fixed for 48 h. Selected tissue blocks were embedded in paraffin. Histopathological sections from all 11 brains were stained with hematoxylin-eosin and examined microscopically by experienced histomorphologists.
The same protocol was applied to three additional HB preterm lambs, which were euthanized at the peak of HB (D0) to assess bilirubin deposition and early brain inflammation at the acute phase of HB.
Immunohistochemical studies of glial cells in the nucleus tractus solitarius
The brainstems of the 11 lambs euthanized at D3 were sectioned at two-cm intervals, and blocks embedded in paraffin. The block encompassing the obex and the dorsomedial structures of the medulla immediately caudal and rostral to the area postrema was selected for special tissue processing. Series of 10 adjacent frontal sections (20 μm thick) were cut every 500 μm. Paraffin sections were mounted on slides, deparaffinized in xylene and rehydrated with washes in decreasing concentrations of ethanol. The first sections of each series were processed for hematoxylin and eosin staining for identification of structures and the four next slides were used for Iba1 (microglia activation; rabbit Wako 019-19741, 1:500) and GFAP (reactive astrocytes; mouse Sigma C9205, 1:100 000) staining.
Iba1 and GFAP immunochemical staining
After several washes in distilled water, sections were incubated in citric acid 10 mM (pH 6) and placed in a microwave for 11.5 min. After a 20 min wash in distilled water, sections were reacted for 30 min with an excess of peroxide (H2O2 0.03%) to abolish endogenous peroxidase activity. After three 10 min washes in PBS, sections were immersed for 1 h 30 in PBS containing 2% bovine serum albumin and 1.5% normal goat serum to block nonspecific binding sites. Thereafter, the sections were incubated for 1 h at room temperature with the primary antibody diluted in the same blocking reagent and directed against Iba1 or GFAP. After three 10 min washes in PBS, sections were incubated for 1 h at room temperature with the appropriate secondary antibody (goat anti-rabbit or anti-mouse) conjugated with an HRP labeled polymer (EnVision+ System K4011 or K4007, respectively, Dako Canada Inc, Mississauga, ON). Sections were then washed twice and reacted for 1 (Iba1) or 2 (GFAP) min with 3,3′-diaminobenzidine (DAB+ substrate-chromogen solution, Dako kit) under visual inspection. After several washes in distilled water, sections were mounted on slides, air dried, dehydrated, cleared and coverslipped. In the present study, controls were carried out by omitting the primary antibody in the reaction. Such a procedure failed to produce any positive staining for Iba1 or GFAP.
Data Analysis
States of Alertness and Cardiorespiratory Function
Standard electrophysiological and behavioral criteria were used to differentiate NREM sleep (stage N) from REM sleep (stage R) and wakefulness (Reix et al., 2004).
Spontaneous apneas
Spontaneous apneas were defined as at least two missed breaths. For each experimental day, the apnea index (number of apneas/hour), the time spent in apnea and the percentage of apneas coupled with desaturation or bradycardia were computed.
Laryngeal chemoreflexes
Analysis of LCR was performed as previously described within the first minute following each laryngeal injection (St-Hilaire et al., 2005) and included: (i) number of apneas and their total summed duration; (ii) number of bradycardias [defined by a decrease in heart rate (HR) >33% for >5 s] and their total summed duration; (iii) number of cardiac slowings (decrease in HR >33% for < 5 s); (iv) total summed duration of cardiac inhibition (total summed duration of both bradycardias and HR slowings); (v) % decrease in SpO2; and (vi) number of coughs (including laryngeal expiratory reflexes, which could not be discerned from coughs in our study).
Pulmonary chemoreflexes
Intravenous injection of capsaicin has previously been shown to trigger a biphasic response consisting of an apnea immediately followed by rapid shallow breathing (Diaz et al., 1999). Accordingly, in the first minute following IV capsaicin injection, analysis included: (i) total summed duration of respiratory and cardiac inhibition; (ii) number of bradycardias and cardiac slowings; (iii) % decrease in SpO2; (iv) respiratory rate (RR) during rapid shallow breathing, obtained between 30 and 60 s following the injection.
Hypoxia
Minute ventilation (VE), tidal volume (Vt), RR, HR as well as SpO2 were averaged on 20 s periods at each minute during baseline, hypoxic run, and return to normoxic condition.
Assessment of Glial Cell Activation in the Nucleus Tractus Solitarius
After completion of staining procedures, all sections were scanned (NanoZoomer-XR Digital slide scanner C12000, Hamamatsu Photonics, Boston MA, USA) for further observation with a computer and appropriate software (NDP.view2, Hamamatsu). The number of Iba1- and GFAP-positive cells was determined using a counting method similar to that detailed in previous immunohistochemical studies (Roda et al., 2004). Briefly, this method consists of densitometric and morphometric measurements in gray levels under x10 objective lens to discriminate stained particles from background in delimited regions of interest using ImageJ (developed by Wayne Rasband, NIH). Each stained particle with: (1) a size comprised between 11 and 100 consecutive pixels and (2) an optical density at least 1.5 times higher than that of the background (IJ-ISODATA thresholding method) was counted as an Iba1- or GFAP-positive cell. The investigator responsible for cell counting was blinded to the experimental conditions. Anatomical boundaries of structures retained for quantification [nucleus tractus solitarius (NTS) and area postrema (AP)] were first delimited on corresponding hematoxylin-eosin stained sections. Anatomical matching of sections across animals was respected. Bilateral counts of immunoreactive cells were performed for each structure clearly identified from at least 3 s. For the NTS, counts were performed within a 2 mm2 rectangular area centered either on the tractus solitarius in its intermediate part, or on the commissural subnucleus in its caudal part. The selected regions include the interstitial, lateral, and medial subnuclei of the NTS where cardiovascular but also laryngeal and some lung afferents terminate (Ciriello and Calaresu, 1981; Barraco et al., 1992), as well as ventrolateral, dorsal and commissural subnuclei where most of the bronchopulmonary afferents terminate (Kubin et al., 2006). For the AP, counts were determined on variable surface areas using manual selections adjusted to the size of this nucleus per side and section. These counts were used to calculate average numbers of immunoreactive cells per structure, section and animal.
Statistical Analysis
Results were first averaged in each lamb and then averaged for each group. Values are expressed as means and SD. Normality was first tested using the Kolmogorov-Smirnov test. Quantitative continuous variables with normal distribution (data for spontaneous apneas as well as for LCR and pulmonary chemoreflexes with the exception of count data) were analyzed using generalized estimating equations with repeated measures, with group (HB or control) and experimental day (D0 or D3) as independent variables. The Mann–Whitney U test was used for quantitative continuous variables with asymmetrical distribution and for repeated measurements over time (hypoxia and blood gas results). Count data (number of apneas and coughs during LCR and number of cardiac slowings and bradycardias during LCR and pulmonary chemoreflexes) were analyzed using generalized estimating equations with repeated measures, with a Poisson distribution. Lastly, results for Iba1 and GFAP expression were analyzed using Student's t-test. All statistical analyses were performed with SPSS. A value of p < 0.05 was considered statistically significant. Furthermore, given the relatively small number of studied animals (related to both the complexity of the ovine model and the ethical constraint of keeping the number of ewes and lambs to a minimum), a statistical trend, defined as p < 0.1, was considered in the discussion.
Results
The study was completed in six lambs (including 3 males) in the control group and five lambs (including 3 males) in the HB group. Accordingly, four ewes gave birth to twins, two to triplets and two to a single lamb. Five of the 16 lambs died from dystocia or neonatal respiratory distress (survival rate 69%). The mean birth weight was 2.38 (0.3) kg in the control group and 2.48 (0.5) kg in the HB group (p = 1).
A moderate HB was obtained for 17 h in all five HB lambs. Compared to control lambs, the “Lamb Acute Bilirubin Encephalopathy score” was significantly higher in HB (5 ± 1 vs. 0 ± 0.1) during bilirubin infusion, subsequently returning to near control levels after 24 h (1 ± 0.4) (Bourgoin-Heck et al., 2015; Figure 2C). In addition, a monotonous EEG resembling wakefulness was consistently observed in all HB lambs throughout the 7 h HB period, preventing differentiation of stage N from quiet W (Bourgoin-Heck et al., 2015; Figure 2B). Consequently, three stages of alertness were specifically defined for analysis purpose in the present study: agitated wakefulness, quiet state (= quiet wakefulness + N stage) and R stage. The latter was easily recognized in all lambs by the intense rapid eye movement activity characteristic of lambs as well as irregular breathing.
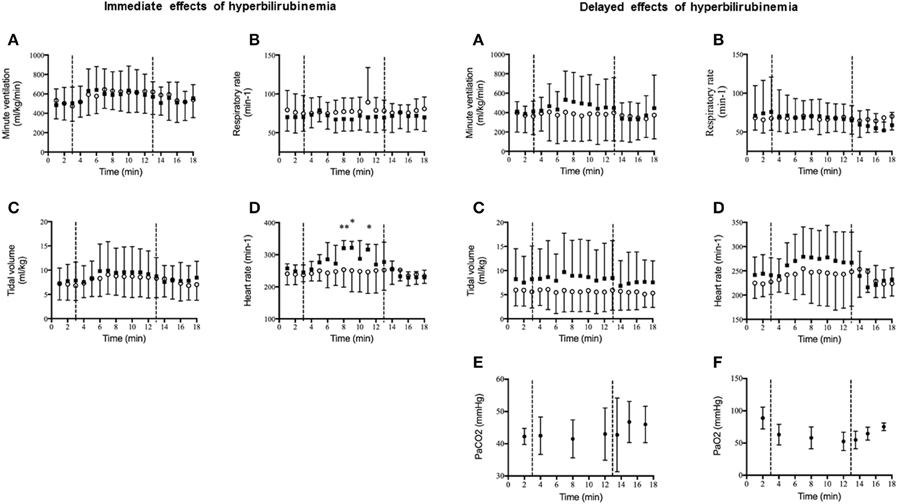
Figure 2. Immediate and delayed effects of moderate hyperbilirubinemia on cardiorespiratory response to hypoxia. (A) Minute ventilation (ml/kg/min); (B) respiratory rate (min−1); (C) tidal volume (ml/kg); (D) heart rate (min−1); (E) PaCO2 (mmHg); (F) PaO2 (mmHg). After a 3-min baseline recording in air, lambs were exposed to 10% O2 for 10 min (represented by the dashed lines), then returned to air for an additional 5-min recording. Values are reported as mean ± standard deviation. Control group (o, n = 6) and hyperbilirubinemia group (■, n = 5 immediate and n = 4 delayed). In order to minimize surgical intervention (surgical catheterization of the brachial artery), PaCO2 and PaO2 measurements were only obtained in 4 lambs (2 control and 2 HB lambs, and no statistical analysis were performed on these data). **p < 0.05 vs. control group and *p < 0.1 vs. control group.
Cardiac and Respiratory Function
During baseline respiration, breathing frequency of HB lambs was lower than controls on both D0 and D3 (Table 1). In addition, the time spent in apnea was significantly increased in the HB group on D0 (Table 1). No other differences were observed for baseline cardiac or respiratory activity.
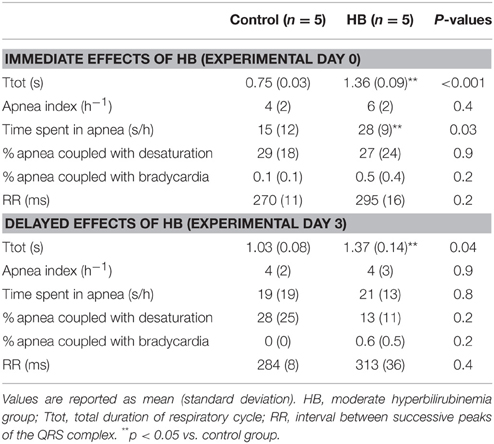
Table 1. Immediate and delayed effects of moderate hyperbilirubinemia on baseline cardiac and respiratory activity.
Laryngeal Chemoreflexes
Results obtained during LCR are summarized in Table 2. Overall, on D0, moderate HB significantly blunted the cardiorespiratory inhibition normally observed during LCR, including a decrease in the number and duration of apneas, lesser cardiac inhibition and a lesser decrease in SpO2. At D3, however, the LCR-induced cardiorespiratory inhibition was similar between both groups (Table 2).
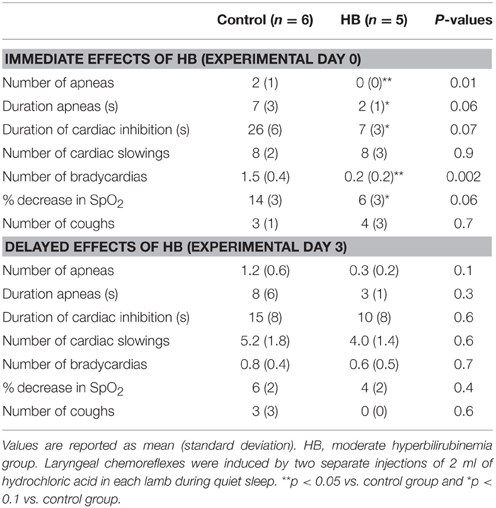
Table 2. Immediate and delayed effects of moderate hyperbilirubinemia on laryngeal chemoreflexes.
Pulmonary Chemoreflexes
Results obtained during pulmonary chemoreflexes are summarized in Table 3. Overall, at D0, moderate HB blunted the cardiorespiratory inhibition elicited by pulmonary chemoreflexes, as shown by a decrease in total duration of respiratory and cardiac inhibition and a decrease in the number of bradycardias. The rapid and shallow breathing following the cardiorespiratory inhibition phase was similar between the two groups. At D3, however, no significant differences were noted between groups for the cardiorespiratory inhibition phase, whereas the RR during the shallow breathing phase was significantly increased in HB lambs (Table 3).
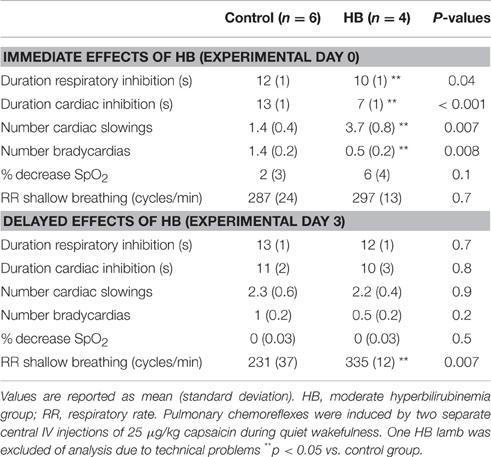
Table 3. Immediate and delayed effects of moderate hyperbilirubinemia on pulmonary chemoreflexes.
Hypoxia
Results are illustrated in Figure 2. At D0, in response to hypoxia, both tidal volume and minute ventilation increased in all lambs, without significant difference between groups. In contrast, hypoxia increased heart rate in HB lambs only (Figure 2D, left panel). After return to normoxia, no differences were noted between the two groups. At D3, cardiorespiratory responses were not significantly different between groups (Figure 2, right panel).
Neurohistopathological Consequences of Moderate Hyperbilirubinemia
Hematoxylin-eosin stained brain sections from the three HB animals euthanized at the peak of hyperbilirubinemia (D0) showed notable bilirubin deposits in the leptomeninges throughout the entire brain, including the brainstem (Figure 3B). While minimal bilirubin deposition was present in brain parenchyma, Alzheimer type II astrocytes [characterized by enlarged, vesicular, and at times empty-looking, nuclei with marginated chromatin and scanty cytoplasm (Ellison et al., 2013)] were noted throughout the brain, including the brainstem (Figure 3A).
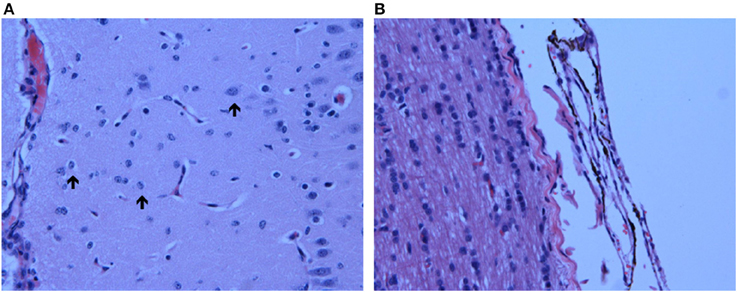
Figure 3. Neuropathological Observations in HB Lambs. (A) Presence of Alzheimer type II astrocytes, characterized by enlarged, pale nuclei with peripheral margination of chromatin (indicated by arrows), throughout the cortex of one HB preterm lamb euthanized at an HB level of 266 μmol/L; (B) bilirubin deposition in the pontine leptomeninges (indicated by dark brown coloration) in another HB preterm lamb with an HB level of 322 μmol/L at time of euthanasia. Images were taken at 40X.
Histopathological examination of the eosin-hematoxylin stained brain sections in the 11 preterm lambs euthanized at D3 showed no bilirubin deposition in any of the HB lambs. In addition, compared to controls, there were only a few foci of slight astrogliosis in the cerebral hemispheres and cerebellum in all HB lambs, as well as in the brainstem in two HB lambs.
Glial Cell Activation in the Nucleus Tractus Solitarius
Immunohistochemical staining showed that both Iba1- (Figure 4) and GFAP-positive cells were significantly increased in the NTS of HB lambs at D3 (Table 4). In addition, there was a modest increase in levels of GFAP expression in the area postrema (AP) in HB lambs when compared to controls (p = 0.07; Table 4). No significant difference was observed for Iba1 expression in the AP between HB and control lambs (Table 4).
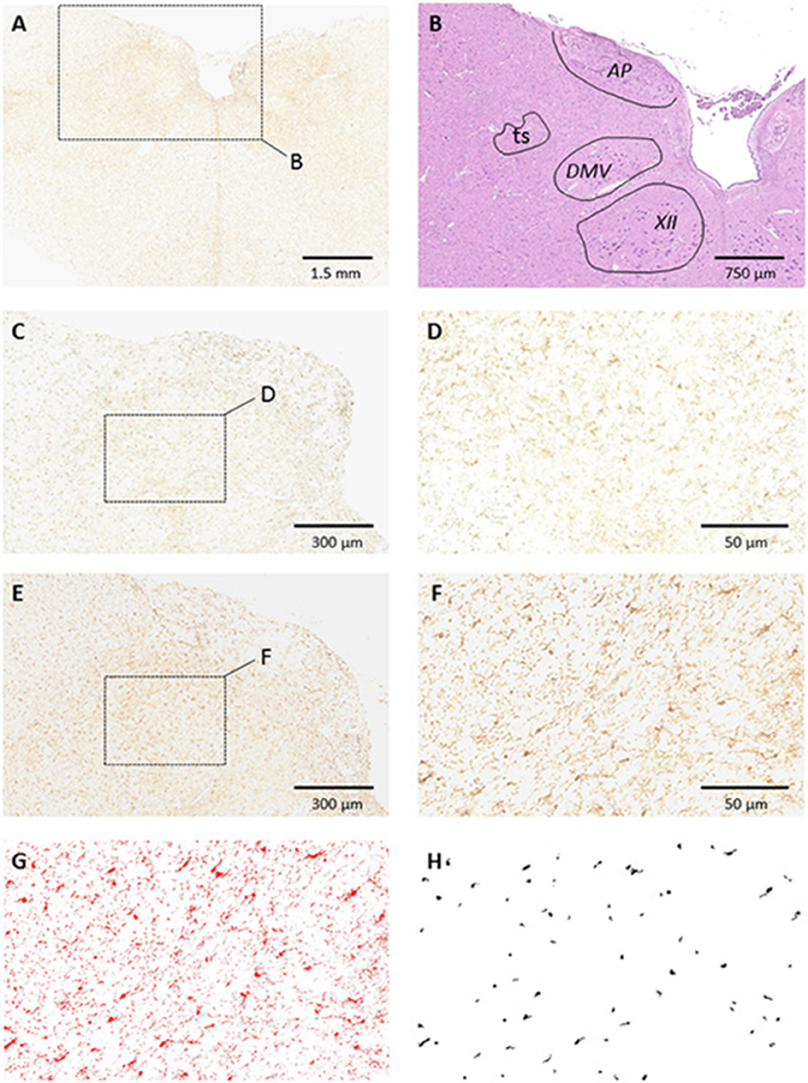
Figure 4. Glial Cell Activation of Iba1 in the Dorsal Medulla. (A) Low magnification microphotograph of a section showing Iba1 staining in the medulla. Note the higher density of Iba1 staining in the dorsomedial aspect in comparison with more ventral or lateral medullary structures; (B) counterstaining of the alternate section shown in (A) with hematoxylin-eosin illustrating the boundaries of dorsomedial nuclei; (C,D) example of Iba1 staining in a control animal at medium and high magnifications, respectively; (E,F) example of Iba1 staining in one HB preterm lamb at medium and high magnifications, respectively. Enlarged areas in (D,F) correspond to the medial part of the nucleus tractus solitarius (NTS). (G,H) Example of image processing of the section shown in (F). The image was converted to 8-bit and thresholded using Image J (G). Fixed morphometric and densitometric criteria were used for automatic counting of Iba1 particles (H). Note the increased number of Iba1-stained particles in NTS after HB. Abbreviations: AP, area postrema; ts, tractus solitarius; DMV, dorsal motor nucleus of the vagus nerve; XII, hypoglossal motor nucleus.
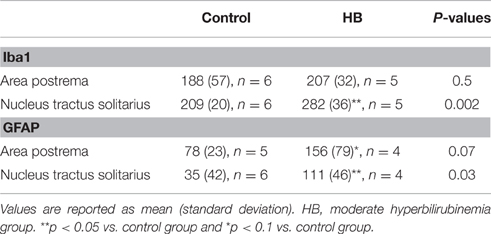
Table 4. Hyperbilirubinemia-induced changes in Iba1- and GFAP expression.
Discussion
The present study in preterm lambs suggest that sustained and moderate HB can induce acute alterations in cardiorespiratory function, including (i) a decrease in baseline respiratory rate and an increase in time spent in apneas, (ii) a blunting of the cardiorespiratory inhibition normally observed during both laryngeal and pulmonary chemoreflexes, and (iii) an increased heart rate in response to acute hypoxia. These changes were associated with an activation of Alzheimer type II astrocytes throughout the brain, including the brainstem. While most cardiovascular responses returned to normal levels 72 h after HB normalization, increased microglial and astrocyte reactivity was still observed within the nucleus tractus solitarius.
Preterm Lamb Model of Moderate Hyperbilirubinemia
Compared to previous animal models of neonatal HB (Ahlfors et al., 1986; Brann et al., 1987; Park et al., 2001; Mesner et al., 2008; Ye et al., 2012), our preterm ovine model of moderate HB has several unique features. Indeed, in order to be clinically relevant, we induced a moderate (150–250 μmol/L) and sustained (17 h) HB in a (late) preterm animal. In contrast, previous studies reported data from more severe (from 220 to 1300 μmoles/L) and shorter duration HB (ranging from one injection to 4 h infusion) (Ahlfors et al., 1986; Brann et al., 1987; Park et al., 2001; Mesner et al., 2008; Ye et al., 2012). To our knowledge, there has been only one previous study on “moderate” HB (100–200 μmoles/L), but with considerably shorter HB duration (2 h) (Roger et al., 1995). The clinical relevance of our model is further shown by the obvious clinical manifestations (jaundice and neurological dysfunction), together with EEG signs of acute bilirubin-encephalopathy observed in all HB lambs. All the aforementioned abnormalities were reversed after bilirubin normalization, as reported in newborn infants during acute bilirubin-induced neurological dysfunction (American Academy of Pediatrics Subcommittee on Hyperbilirubinemia, 2004).
Moderate Hyperbilirubinemia and Cardiorespiratory Function
Observations of decreased baseline respiratory rate and increased time spent in apnea in HB lambs are in agreement with several clinical studies (Johnson et al., 2009; Amin et al., 2014; Amin and Wang, 2015). Such HB-related respiratory inhibition could potentially be mediated, at least in part, by prostaglandin E2 binding to its respective receptors in respiratory regions of the brainstem (Hofstetter et al., 2007).
The present results also reveal that moderate HB transiently blunted the cardiorespiratory inhibition normally observed during both laryngeal and pulmonary chemoreflexes. Such observations were somewhat unexpected, given that various postnatal stresses have been shown to increase apneas and bradycardias observed in the LCR (Lindgren et al., 1996; Frøen et al., 2000; Xia et al., 2008, 2010; St-Hilaire et al., 2010; Carreau et al., 2011). Nevertheless, the blunting of LCR observed in HB lambs in the present study may be non-beneficial, due to the heightened risk of aspiration as a result of the lack of efficient glottal closure. Unfortunately, this cannot be ascertained, for the design of the present study did not include the assessment of tracheal aspiration. The HB-related alteration in cardiorespiratory activity was further confirmed by the present observations in HB lambs, namely that (i) the cardiorespiratory inhibition normally triggered by pulmonary C-fiber stimulation was blunted, and (ii) the heart rate was increased in response to acute hypoxia. The above results expand previous data obtained with severe HB in rat pups showing an altered respiratory response to hypoxia and hypercapnia (Mesner et al., 2008).
Bilirubin Deposition and Brainstem Glial Activation
In the three preterm lambs euthanized at the peak of HB, bilirubin deposition was observed only within the leptomeninges overlying the entire brain. This is somewhat at odds with a previous report in rat pups with severe HB levels, in which bilirubin deposition was noted in the brainstem itself (Mesner et al., 2008). Such differences may be related to the lower HB level and/or to a lower permeability of the blood brain barrier in preterm lambs compared to rat pups. In addition, acute HB was associated herein with Alzheimer type II astrocytes throughout the entire brain of the three lambs, including in the brainstem. Alzheimer type II astrocytes have been reported in various acute central nervous system injuries, such as stroke or trauma. They may represent dysfunctional astrocytes (Adams and Parker, 2011) and are thought to represent glial cells in the process of transformation into reactive astrocytes (Panickar and Norenberg, 2005). Of interest, the presence of Alzheimer type II astrocytes in the cerebral cortex (Figure 3A), indicative of brain injury, was noted in concomitance with the monotonous EEG observed in all HB lambs.
In addition to the above, astrocyte and microglial activation in the NTS were detected 3 days following acute HB in preterm lambs. Cerebral glial activation, encompassing both astrocytes and microglia, has been previously reported in response to bilirubin (reviewed in Fernandes and Brites, 2009). The present observation, to the best of our knowledge, is the first report of glial activation in the NTS in response to HB. These findings provide further evidence that even a moderate HB in preterm lambs can induce an inflammatory response in the NTS region devoted to central processing of cardiorespiratory afferent inputs (Kubin et al., 2006). Both activated microglia and astrocytes could alter neuronal function such as synaptic transmission, including that involved in cardiovascular and respiratory reflexes in the NTS (Andresen and Paton, 2011). The results herein on HB-induced alterations in laryngeal and pulmonary chemoreflexes, as well as in the cardiac response to hypoxia, are in keeping with this hypothesis. Our findings are in agreement with a previous study showing abnormal ventilatory response to CO2 and hypoxia in rat pups with severe HB (Mesner et al., 2008). Of note, astrocytic damage in the NTS has previously been shown to interfere with cardiovascular reflex transmission by attenuating the arterial baroreflex, the arterial chemoreflex, and the von Bezold–Jarisch cardiopulmonary reflex (Lin et al., 2013). In addition, migroglial activation in the NTS was previously implicated in the impaired cardiovascular regulation observed in diabetic rats (Rana et al., 2014). Of note, the absence of a detectable bilirubin deposition in the brain parenchyma may suggest that HB in our preterm lambs exerts its effects on glial cells via the release of certain mediators by cells forming the blood brain barrier, including astrocytes (Sofroniew, 2015).
Limitation of the Study
As stated earlier, our ovine model provides unique characteristics (preterm birth, both moderate and sustained hyperbilirubinemia), which are of particular relevance for assessing the physiological consequences of hyperbilirubinemia in human newborns. One limitation may be that, contrary to humans, preterm newborn lambs (for yet unknown reasons) do not naturally develop hyperbilirubinemia.
Conclusion
Our results show that moderate and sustained HB alters baseline respiration and reflex cardiorespiratory function. Compared to previous reports, the novelty of our study stems from the fact that (i) results were obtained with moderate and clinically relevant HB levels, (ii) in addition to respiratory impairment, cardiac function was also shown to be altered, and (iii) glial activation was revealed in the nucleus tractus solitarius, which is a key component in the control of cardiorespiratory function.
Author Contributions
M-LS, MB-H, NS, and J-PP conceived and designed the study. M-LS and FC conceived the bilirubin solution. M-LS, MB-H and NS performed the animal experiments. M-LS, CG, MR, HK analyzed the data. M-LS, NS, CG, MR, HK, and J-PP interpreted the results obtained. NS, CG, and J-PP drafted the manuscript. M-LS, MB-H, NS, FC, CG, MR, HK, and J-PP revised the manuscript. All authors read and approved the final version of the manuscript.
Funding
This study was supported by an operating grant from the Canadian Institutes of Health Research. J-PP is the holder of the Canada Research Chair in Neonatal Respiratory Physiology and a member of the Centre de recherche du Centre hospitalier universitaire de Sherbrooke. M-LS was the recipient of a Ph.D. scholarship from the Canadian Thoracic Society and the Fonds de recherche du Québec - Nature et technologies (FQRNT).
Conflict of Interest Statement
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Acknowledgments
The authors wish to thank Professor Guillaume Sébire (Neurology Division, Department of Pediatrics, McGill University, Montreal) for invaluable discussions on portions of this work. The authors also acknowledge the excellent technical assistance of Charlène Nadeau.
References
Adams, J. D., and Parker, K. K. (2011). Extracellular and Intracellular Signaling. Cambridge, UK: RSC Publishing.
Ahlfors, C. E., Bennett, S. H., Shoemaker, C. T., Ellis, W. G., Davis, S. L., Wennberg, R. P., et al. (1986). Changes in the auditory brainstem response associated with intravenous infusion of unconjugated bilirubin into infant rhesus monkeys. Pediatr. Res. 20, 511–515. doi: 10.1203/00006450-198606000-00005
American Academy of Pediatrics Subcommittee on Hyperbilirubinemia (2004). Management of hyperbilirubinemia in the newborn infant 35 or more weeks of gestation. Pediatrics 114, 297–316. doi: 10.5468/ogs.2016.59.4.319
Amin, S. B., and Wang, H. (2015). Unbound unconjugated hyperbilirubinemia is associated with central apnea in premature infants. J. Pediatr. 166, 571–575. doi: 10.1016/j.jpeds.2014.12.003
Amin, S. B., Bhutani, V. K., and Watchko, J. F. (2014). Apnea in acute bilirubin encephalopathy. Semin. Perinatol. 38, 407–411. doi: 10.1053/j.semperi.2014.08.003
Andresen, M. C., and Paton, J. F. R. (2011). “The nucleus of the solitary tract: processing information from viscerosensory afferents,” in Central Regulation of Autonomic Functions, eds I. J. Llewellyn-Smith and A. J. M. Verberne (New York, NY: Oxford University Press). 23–46.
Barraco, R., el-Ridi, M., Ergene, E., Parizon, M., and Bradley, D. (1992). An atlas of the rat subpostremal nucleus tractus solitarius. Brain Res. Bull. 29, 703–765. doi: 10.1016/0361-9230(92)90143-L
Boudaa, N., Samson, N., Carriere, V., Germim, P. S., Pasquier, J. C., Bairam, A., et al. (2013). Effects of caffeine and/or nasal CPAP treatment on laryngeal chemoreflexes in preterm lambs. J. Appl. Physiol. 114, 637–646. doi: 10.1007/s00421-014-2973-z
Bourgoin-Heck, M., Specq, M. L., Samson, N., Nadeau, C., Prodel, M., Corbin, F., et al. (2015). Effects of moderate hyperbilirubinemia on nutritive swallowing and swallowing-breathing coordination in preterm lambs. Neonatology 108, 42–48. doi: 10.1159/000381205
Brann, B. S. IV Stonestreet, B. S., Oh, W., and Cashore, W. J. (1987). The in vivo effect of bilirubin and sulfisoxazole on cerebral oxygen, glucose and lactate metabolism in newborn piglets. Pediatr. Res. 22, 135–140. doi: 10.1203/00006450-198708000-00006
Carreau, A. M., Patural, H., Samson, N., Doueik, A. A., Hamon, J., Fortier, P. H., et al. (2011). Effects of simulated reflux laryngitis on laryngeal chemoreflexes in newborn lambs. J. Appl. Physiol. 111, 400–406. doi: 10.1152/japplphysiol.00105.2011
Ciriello, J., and Calaresu, F. R. (1981). Projections from buffer nerves to the nucleus of the solitary tract: an anatomical and electrophysiological study in the cat. J. Auton. Nerv. Syst. 3, 299–310. doi: 10.1016/0165-1838(81)90071-0
Diaz, V., Dorion, D., Renolleau, S., Létourneau, P., Kianicka, I., and Praud, J. P. (1999). Effects of capsaicin pretreatment on expiratory laryngeal closure during pulmonary edema in lambs. J. Appl. Physiol. 86, 1570–1577.
Ellison, E., Love, S., Chimelli, L., Harding, B. N., Lowe, J. S., Vinters, et al. (2013). Neuropathology: A Reference Text of CNS Pathology. Edinburgh: Elsevier Mosby.
Fernandes, A., and Brites, D. (2009). Contribution of inflammatory processes to nerve cell toxicity by bilirubin and efficacy of potential therapeutic agents. Curr. Pharm. Des. 15, 2915–2926. doi: 10.2174/138161209789058165
Frøen, J. F., Akre, H., Stray-Pedersen, B., and Saugstad, O. D. (2000). Adverse effects of nicotine and interleukin-1beta on autoresuscitation after apnea in piglets: implications for sudden infant death syndrome. Pediatrics 105:E52. doi: 10.1542/peds.105.4.e52
Hansen, T. W., Cashore, W. J., and Oh, W. (1992). Changes in piglet auditory brainstem response amplitudes without increase in serum or cerebrospinal fluid neuron-specific enolase. Pediatr. Res. 32, 524–529. doi: 10.3109/15622975.2014.952776
Hofstetter, A. O., Saha, S., Siljehav, V., Jakobsson, P. J., and Herlenius, E. (2007). The induced prostaglandin E2 pathway is a key regulator of the respiratory response to infection and hypoxia in neonates. Proc. Natl. Acad. Sci. U.S.A. 104, 9894–9899. doi: 10.1073/pnas.0611468104
Johnson, L., and Bhutani, V. K. (2001). The clinical syndrome of bilirubin-induced neurologic dysfunction. Semin. Perinatol. 35, 101–113. doi: 10.1053/j.semperi.2011.02.003
Johnson, L., Bhutani, V. K., Karp, K., Sivieri, E. M., and Shapiro, S. M. (2009). Clinical report from the pilot USA Kernicterus Registry (1992 to 2004). J. Perinatol. 29, S25–S45. doi: 10.1038/jp.2008.211
Kubin, L., Alheid, G. F., Zuperku, E. J., and McCrimmon, D. R. (2006). Central pathways of pulmonary and lower airway vagal afferents. J. Appl. Physiol. 101, 618–627. doi: 10.1152/japplphysiol.00252.2006
Lin, L. H., Moore, S. A., Jones, S. Y., McGlashon, J., and Talman, W. T. (2013). Astrocytes in the rat nucleus tractus solitarii are critical for cardiovascular reflex control. J. Neurosci. 33, 18608–18617. doi: 10.1523/JNEUROSCI.3257-13.2013
Lindgren, C., Lin, J., Graham, B. S., Gray, M. E., Parker, R. A., and Sundell, H. W. (1996). Respiratory syncytial virus infection enhances the response to laryngeal chemostimulation and inhibits arousal from sleep in young lambs. Acta Paediatr. 85, 789–797. doi: 10.1111/j.1651-2227.1996.tb14153.x
Mazeiras, G., Rozé, J. C., Ancel, P. Y., Caillaux, G., Frondas-Chauty, A., Denizot, S., et al. (2012). Hyperbilirubinemia and neurodevelopmental outcome of very low birthweight infants: results from the LIFT cohort. PLoS ONE 7:e30900. doi: 10.1371/journal.pone.0030900
Mesner, O., Miller, M. J., Iben, S. C., Prabha, K. C., Mayer, C. A., Haxhiu, M. A., et al. (2008). Hyperbilirubinemia diminishes respiratory drive in a rat pup model. Pediatr. Res. 64, 270–274. doi: 10.1203/PDR.0b013e31817d9c0c
Panickar, K. S., and Norenberg, M. D. (2005). Astrocytes in cerebral ischemic injury: morphological and general considerations. Glia 50, 287–298. doi: 10.1002/glia.20181
Park, W. S., Chang, Y. S., Chung, S. H., Seo, D. W., Hong, S. H., and Lee, M. (2001). Effect of hypothermia on bilirubin-induced alterations in brain cell membrane function and energy metabolism in newborn piglets. Brain Res. 922, 276–281. doi: 10.1016/S0006-8993(01)03186-9
Rana, I., Badoer, E., Alahmadi, E., Leo, C. H., Woodman, O. L., and Stebbing, M. J. (2014). Microglia are selectively activated in endocrine and cardiovascular control centers in streptozotocin-induced diabetic rats. J. Neuroendocrinol. 26, 413–425. doi: 10.1111/jne.12161
Reix, P., Arsenault, J., Langlois, C., Niyonsenga, T., and Praud, J. P. (2004). Nonnutritive swallowing and respiration relationships in preterm lambs. J. Appl. Physiol. 97, 1283–1290. doi: 10.1152/japplphysiol.00060.2004
Roda, F., Pio, J., Bianchi, A. L., and Gestreau, C. (2004). Effects of anesthetics on hypoglossal nerve discharge and c-Fos expression in the brainstem hypoglossal premotor neurons. J. Comp. Neurol. 468, 571–586. doi: 10.1002/cne.10974
Roger, C., Koziel, V., Vert, P., and Nehlig, A. (1995). Mapping of the consequences of bilirubin exposure in the immature rat: local cerebral metabolic rates for glucose during moderate and severe hyperbilirubinemia. Early Hum. Dev. 43, 133–144. doi: 10.1016/0378-3782(95)01668-6
Samson, N., Dumont, S., Specq, M. L., and Praud, J. P. (2011). Radio telemetry devices to monitor breathing in non-sedated animals. Respir. Physiol. Neurobiol. 179, 111–118. doi: 10.1016/j.resp.2011.09.008
Samson, N., St-Hilaire, M., Nsegbe, E., Reix, P., Moreau-Bussière, F., and Praud, J. P. (2005). Effect of nasal continuous or intermittent positive airway pressure on nonnutritive swallowing in the newborn lamb. J. Appl. Physiol. 98, 2197–2203. doi: 10.1152/japplphysiol.00464.2005
Sofroniew, M. V. (2015). Astrocyte barriers to neurotoxic inflammation. Nat. Rev. Neurosci. 16, 249–263. doi: 10.1038/nrn3898
St-Hilaire, M., Duvareille, C., Avoine, O., Carreau, A. M., Samson, N., Micheau, P., et al. (2010). Effects of postnatal smoke exposure on laryngeal chemoreflexes in newborn lambs. J. Appl. Physiol. 109, 1820–1826. doi: 10.1152/japplphysiol.01378.2009
St-Hilaire, M., Nsegbe, E., Gagnon-Gervais, K., Samson, N., Moreau-Bussière, F., Fortier, P. H., et al. (2005). Laryngeal chemoreflexes induced by acid, water, and saline in nonsedated newborn lambs during quiet sleep. J. Appl. Physiol. 98, 2197–2203. doi: 10.1152/japplphysiol.01346.2004
Watchko, J. F. (2009). Identification of neonates at risk for hazardous hyperbilirubinemia: emerging clinical insights. Pediatr. Clin. North Am. 56, 671–687. doi: 10.1016/j.pcl.2009.04.005
Xia, L., Damon, T., Leiter, J. C., and Bartlett, D. (2008). Elevated body temperature exaggerates laryngeal chemoreflex apnea in decerebrate piglets. Adv. Exp. Med. Biol. 605, 249–254. doi: 10.1007/978-0-387-73693-8_44
Xia, L., Leiter, J. C., and Bartlett, D. (2010). Gestational nicotine exposure exaggerates hyperthermic enhancement of laryngeal chemoreflex in rat pups. Respir. Physiol. Neurobiol. 171, 17–21. doi: 10.1016/j.resp.2010.01.011
Keywords: hyperbilirubinemia, prematurity, brain, pathophysiology, pulmonary and laryngeal chemoreflexes, hypoxia, nucleus tractus solitarius
Citation: Specq M-L, Bourgoin-Heck M, Samson N, Corbin F, Gestreau C, Richer M, Kadhim H and Praud J-P (2016) Moderate Hyperbilirubinemia Alters Neonatal Cardiorespiratory Control and Induces Inflammation in the Nucleus Tractus Solitarius. Front. Physiol. 7:437. doi: 10.3389/fphys.2016.00437
Received: 23 June 2016; Accepted: 14 September 2016;
Published: 30 September 2016.
Edited by:
Keith Russell Brunt, Dalhousie University, CanadaReviewed by:
Ching Jung Lai, Tzu Chi University, TaiwanShabih U. Hasan, University of Calgary, Canada
Copyright © 2016 Specq, Bourgoin-Heck, Samson, Corbin, Gestreau, Richer, Kadhim and Praud. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) or licensor are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Jean-Paul Praud, amVhbi1wYXVsLnByYXVkQHVzaGVyYnJvb2tlLmNh