 Mario A. J. Golding1
Mario A. J. Golding1 Nadia K. Khouri
Nadia K. Khouri Kathryn B. Anderson
Kathryn B. Anderson Troy D. Wood
Troy D. Wood Simone L. Sandiford
Simone L. Sandiford- 1Department of Basic Medical Sciences, Pharmacology and Pharmacy Section, Faculty of Medical Sciences, The University of the West Indies, Mona, Kingston, Jamaica
- 2Department of Microbiology and Immunology, The State University of New York (SUNY) Upstate Medical University, Syracuse, NY, United States
- 3Department of Chemistry, 417 Natural Sciences Complex, University at Buffalo, Buffalo, NY, United States
- 4Mosquito Control and Research Unit, The University of the West Indies, Mona, Kingston, Jamaica
Latin America and the Caribbean continue to be adversely affected by dengue with the disease being endemic in several countries in this region. As a result, the social and economic impacts of the disease have risen significantly. Currently, there are very few solutions available to limit the spread of the virus, with vector control being the most commonly used. However, due to increased insecticidal resistance, scientists in the region have actively been seeking new ways to limit the spread of the virus. This quest has led researchers to investigate the antiviral properties of natural products. While antiviral screening activities focused on preventing or treating infection of the human host remains a significant area of study, some scientists have now focused their attention on preventing infection or transmission in the mosquito vector. This review therefore aims to highlight the use of natural products in Latin America and the Caribbean for blocking viral transmission of dengue virus in mosquito cells. This novel approach is promising and could ultimately be used in conjunction with other methods to help significantly reduce dengue incidence in the region.
Introduction
Dengue virus (DENV) is a single positive-stranded RNA virus belonging to the Flaviviridae family (genus Flavivirus) which causes dengue infection. There are four antigenically distinct serotypes of the virus; DENV -1, -2, -3 and -4 (1, 2) all of which can cause infection of clinical significance. The virus is now endemic in over 100 countries worldwide (2, 3) where its economic impact is approximately US$39 billion annually (4). Dengue is responsible for the greatest burden of arboviral morbidity and mortality globally (2) and according to estimates, 390 million infections occur yearly of which 96 million manifests with symptoms (2, 5, 6).
Transmission of the virus is primarily accomplished through interactions between mosquitoes and humans, its natural hosts, and is successfully transmitted via alternate cycles of viral replication in each host (7). Primary dengue virus exposure may often result in asymptomatic infections or mild flu-like classical dengue but may occasionally cause the more severe dengue hemorrhagic fever/dengue shock syndrome (DHF/DSS) (2, 5, 8–10). Primary infection by one serotype is thought to result in lifelong immunity to that serotype but cross protection against other serotypes is partial and temporary (10). As such, secondary exposure to a different serotype increases the risks of the more severe forms of dengue (DHF/DSS) (11).
For decades, Latin America and the Caribbean (LAC) have grappled with dengue (12, 13) and now more than ever, it is a major public health concern with an estimated annual cost in the region of US$1-4billion owing to lost productivity, medical bills, and surveillance and vector control costs (4, 14, 15). With approximately 13.3 million dengue cases, LAC accounts for 14 percent of the world’s apparent infections (5). In 2019, for example, the region recorded nearly 3 million reported cases, the highest in its history (16) as illustrated in Table 1. Reasons for the explosive increase in the incidence of dengue have been attributed to rapid population growth, increased urbanization, globalization, international travel, changes in climate and poor and inadequate vector control measures (4, 11, 18).
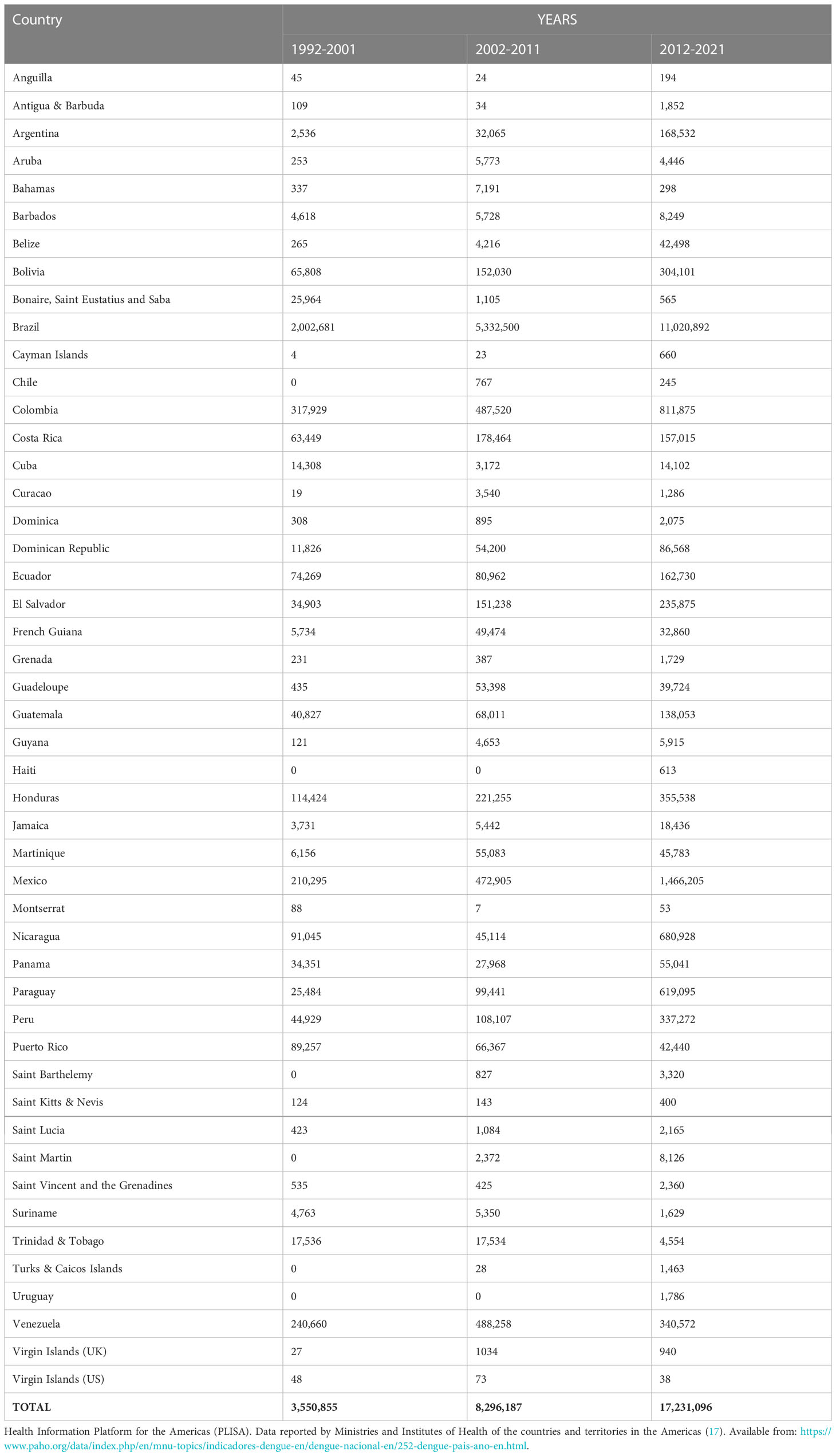
Table 1 Reported cases of dengue in the LAC region over the past 3 decades (1992-2021).
Another factor contributing to the increase in dengue cases in the LAC region is geographic expansion of the vector responsible for its transmission (19). Aedes aegypti the main dengue vector is now widely distributed in various LAC countries and is highly domesticated and feeds preferentially on humans (20). This vector is of epidemic concern as the Ae. aegypti mosquito frequently take several blood meals from different individuals in a single gonotrophic cycle (21, 22). Additionally, Aedes albopictus, a known competent vector of dengue virus in Asia, is a highly invasive species (23) which has spread into the LAC region where it poses an additional challenge. While some studies have suggested that Ae. albopictus could potentially be helping to spread dengue virus in the region (24–27), its role in dengue transmission in LAC is still not yet well established (28).
In the absence of a suitable vaccine or effective antiviral agents, vector control remains crucial in limiting dengue infections in LAC. Some of the most common vector control strategies includes the use of insecticides against larvae and adults, source reduction of possible breeding sites and the use of insecticide-treated bed nets and curtains (29). Due to the increased development of resistance, the use of insecticides against the immature and adult stages of mosquitoes has become a major challenge in LAC. Insecticide resistance in the Caribbean, for example, dates to as far as 1955 when resistance to DDT (dichloro-diphenyl-trichloroethane) was observed (30). By 1974, there was widespread insecticide resistance in Ae. aegypti populations in several Caribbean countries to the organophosphate insecticides: malathion, fenthion and temephos, which had replaced DDT (30). A recent review by 31, noted that resistance in mosquito populations to DDT, temephos and deltamethrin is serious and widespread within the region. Furthermore, because synthetic insecticides are not specific, they may have harmful effects on other non-targeted organisms and the environment (32–34). Insecticides alone are ineffective at reducing mosquito populations as health education and community involvement are also necessary to sufficiently achieve vector reduction (29). Targeting the breeding sites of mosquitoes in one’s environs can help reduce mosquito populations. Removing old tires and containers that collect and hold water aids in the prevention of mosquito egg hatching (35). However, none of these approaches to vector control, even in combination, can eliminate dengue transmission.
As the search for solutions to the ongoing onslaught of dengue on the LAC region continues, researchers have begun to focus their attention on natural remedies. It has been estimated that up to 400 million people in LAC use herbal medicine to meet a primary health need (36). Generally, plants have been used for the treatment of a myriad of illnesses and health conditions such as colds, fever, flu, diabetes mellitus, hypertension (37) and treatment of dengue fever (38). One of the most widely used plants in the Caribbean for dengue fever is Carica papaya (Caricaceae) (39–41) where the juice is routinely extracted from the leaves and consumed by dengue patients to improve platelet count (42, 43). While scientific validation of the folkloric use of papaya for dengue treatment in the LAC region is lacking, several studies conducted in Asia have supported this use (44, 45). Other plants used by individuals in the Caribbean to boost overall immunity in the treatment of dengue include Trigonella foenum-graecum (fenugreek) seeds, Ocimum basilicum (basil) leaves, Azadirachta indica (neem) leaves, Psidium guajava (guava) juice, and Tinospora cordifolia (giloy) (41). (46). Tinospora cordifolia may also help to reduce fever during dengue infection (47).
In addition to boosting overall immunity and reducing fever during dengue infection, several plants and their derivatives have been shown to possess in vitro antiviral activity against dengue virus in mammalian cells in the LAC region (48–51). Generally, most antiviral studies for dengue only use one DENV serotype (typically DENV-2) or observe differential inhibition of the four serotypes. Identifying a plant/compound with significant activity against all four dengue serotypes would be ideal as multiple serotypes are now circulating within the region. Despite many plants showing positive in vitro anti-dengue activity in mammalian cells, none of these plants have progressed beyond investigational stages. The development of antivirals to treat the infection in humans has been difficult and remains elusive due to the need to identify safe fast-acting agents with equal efficacy against all DENV serotypes, a low likelihood of viral resistance and ability to reduce disease symptoms and severity (52–54).
In recent years, the search for effective antiviral agents for dengue infections has led scientists to the realization that targeting infection in the mosquito vector may be a beneficial approach to reducing virus transmission (55, 56). Like in mammalian cells, the arboviral infection cycle in mosquito cells undergo an initial interaction between viral surface proteins and receptor molecules on the host cell surface. This is followed by receptor-mediated endocytosis, which internalizes the viral particles within the cytoplasm, viral assembly in the endoplasmic reticulum (ER), and finally the secretion of mature virions (57). The arboviral cycle offers numerous potential targets for suppressing or interrupting viral production in mosquito cells. Ideally, antivirals to prevent dengue replication in the mosquito should have a high efficacy of infection prevention when delivered to the adult mosquito to completely block the virus’ development (56). These substances should also be safe in order to avoid negative consequences in humans (58). This review article is primarily focused on reviewing the literature on the use of natural products for blocking dengue viral infection/replication in mosquito cells in the LAC region.
Antiviral activity of natural products against dengue virus in insect cell lines
Currently, few studies conducted in the LAC region have sought to investigate natural products for their ability to inhibit viral infection or replication of dengue virus in mosquito cells, and none thus far have come from the Caribbean. These studies are routinely performed in the C6/36 insect cell line which is the most widely used and accepted model system of mosquito cells used for studying the antiviral potential of substances (7). For example, 59 demonstrated by immunofluorescence assay that the sulfated galactomannans BRS and LLS obtained from the seeds of Mimosa scabrella (Fabaceae) and Leucaena leucocephala (Fabaceae) respectively had significant inhibitory activity against DENV-1 in C6/36 cells. BRS and LLS reduced DENV-1 viral titre by 100-fold at concentrations of 347 mg L−1 and 37 mg L−1 respectively (Table 2). The half maximal cytotoxic concentration (CC50) for native galactomannans was >8.0 g L−1, and for derivative galactomannans it was 1.5 g L−1 for BRS and 2.0 g L−1 for LLS. The authors alluded to the fact that the sulfated galactomannans may be producing their antiviral activity either by inhibition of adsorption of the virus to the host cell and/or inhibition of the step that occurs after viral internalization but before the onset of late viral protein synthesis (59).
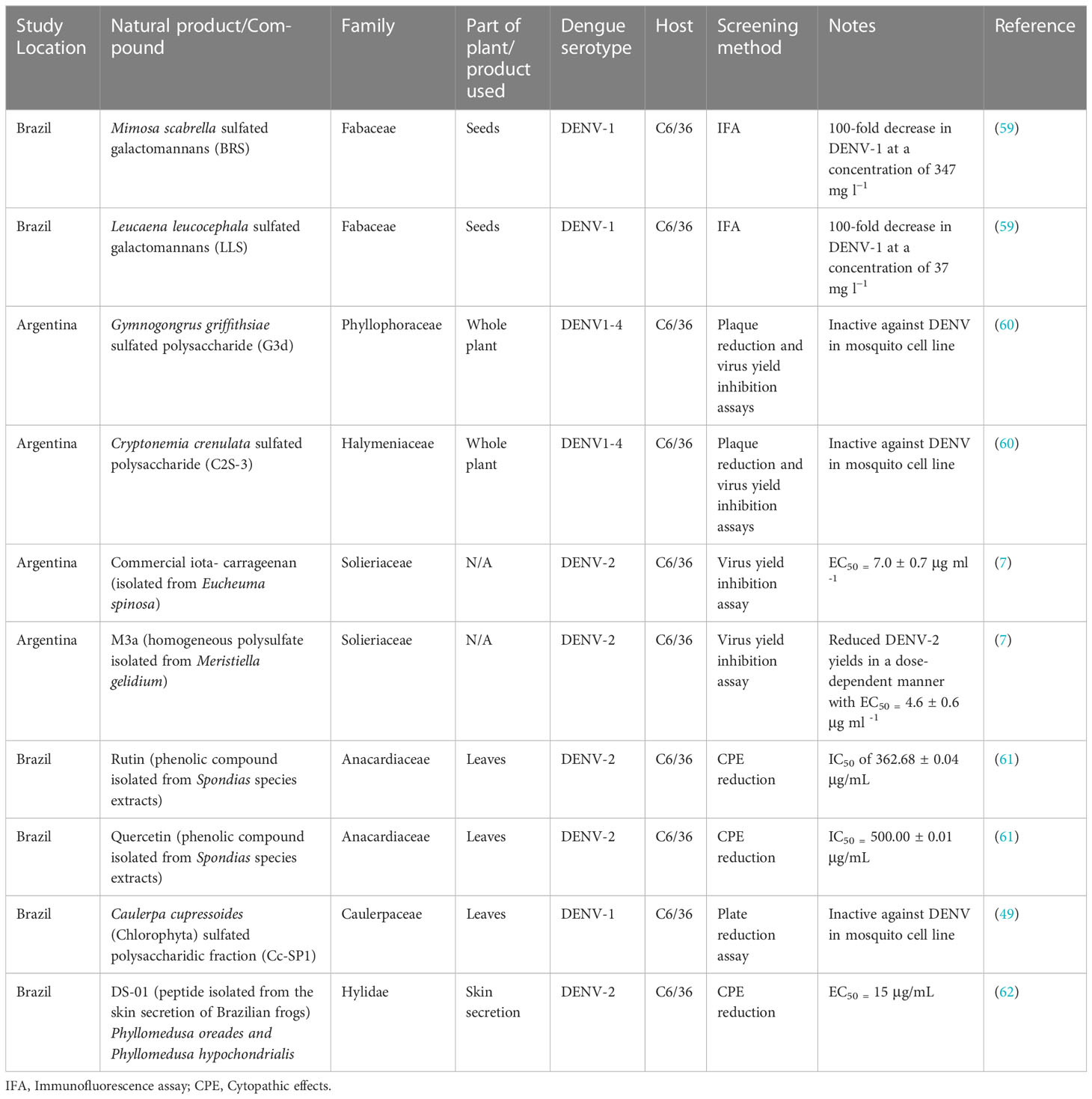
Table 2 Antiviral potential of natural products against dengue virus in mosquito cell lines.
In another study, the methanol:water (80:20 v/v) leaf extracts of Spondias mombin (hog plum) (Anacardiaceae) and Spondias tuberosa (Brazil plum) (Anacardiaceae) and their main phenolic compounds rutin, quercetin, and ellagic acid were investigated for their antiviral activity against DENV-2 by the 3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyl-2H-tetrazolium bromide (MTT) and cytopathic effect (CPE) reduction assays in C6/36 cells (Table 2). No cytotoxicity was observed with any of these compounds at concentrations up to 1000 µg/mL. Of the compounds evaluated, rutin was found to be most effective against DENV-2 with a half maximal inhibitory concentration (IC50) of 362.68 ± 0.04 µg/mL followed by quercetin with an IC50 of 500.00 ± 0.01 µg/mL (61). Based on these findings, the researchers concluded that rutin and quercetin have the potential to be developed into anti-dengue agents. They did underline, however, that more in vitro and in vivo investigations were needed to demonstrate the efficacy of these flavonoids against dengue virus.
Interestingly, several studies conducted in the region have observed differential susceptibility of vertebrate and invertebrate cells to antiviral screening against dengue. Firstly, researchers in Argentina demonstrated that the sulfated polysaccharides G3d and C2S-3 obtained from the red seaweeds Gymnogongrus griffithsiae and Cryptonemia crenulate respectively possess varying antiviral activity against all four dengue virus serotypes in different cell lines (61). They found that the polysaccharides were active inhibitors of DENV-2 and DENV-3 multiplication in Vero cells, HepG2 and foreskin PH cells but displayed no inhibitory activity in the mosquito C6/36 cells (Table 2). The compounds were less effective against DENV-4 and were totally inactive against DENV-1. Additionally, the researchers noted that since DENV multiplication levels in the vertebrate and invertebrate cell lines were similar, the observed differential susceptibility of virus serotype or host cell was not due to virus growth ability. Through mechanistic studies on DENV-2 in Vero cells, they demonstrated that G3d and C2S-3 were only effective inhibitors when administered in conjunction with the virus or shortly after infection. Their findings suggested that the primary mode of action of these compounds against DENV-2 was virus adsorption. However, virus internalization, was also thought to be affected (61).
In a similar way, (7) demonstrated a differential inhibition of DENV-2 in mammalian and mosquito cells when iota-carrageenan which was extracted from red seaweed was screened for antiviral activity (Table 2). No discernible decline in cell viability in either cell lines were observed following cytotoxicity screening of iota-carrageenan by MTT assay. In Vero cells, iota-carrageenan only inhibited virion binding at the initiation of the infectious cycle. In C6/36 cells, however, DENV-2 adsorption was unaffected possibly due to the presence of insufficient heparan sulfate (HS) on the surface of C6/36 cells (7). In the same study, the researchers also investigated the anti-dengue potential of M3a; a homogeneous polysulfate isolated from Meristiella gelidium. M3a was shown to decrease DENV-2 titre in a dose-dependent manner with half maximal effective concentration (EC50) values of 4.6 ± 0.6 and 0.93 ± 0.05 μg mL-1 in C6/36 and Vero cells, respectively (7). Furthermore, the sulfated polysaccharide fraction (Cc-SP1) from the green seaweed Caulerpa cupressoides (Chlorophyta) was investigated for its inhibitory activity against DENV-1 by plaque reduction assay (Table 2). The study which was conducted in Brazil found that Cc-SP1 was a potent inhibitor of DENV-1 (96%) at 0.35 μg mL-1 in Vero cell-line but was totally ineffective at inhibiting virus multiplication in the C6/36 mosquito cell-line (49). Cc-SP1 did not cause cytotoxicity up to 1000 μg mL-1. Like (7), the failure of Cc-SP1 to inhibit DENV infection in C6/36 could be due to inadequate heparin sulfate on the C6/36 cell surface.
On the contrary, researchers in Brazil demonstrated that Dermaseptin 01 (DS-01), a peptide isolated from the skin secretion of Brazilian frogs Phyllomedusa oreades and Phyllomedusa hypochondrialis possessed greater anti-dengue activity in insect cells than in mammalian cells (Table 2). DS-01 through MTT assay in LLCMK2 cells and CPE reduction assay in C6/36 cells displayed antiviral activity against DENV-2 with EC50 values of 60 μg/mL and 15 μg/mL respectively (62). The 50% cytotoxicity concentration of dermaseptin 01 was 105 μg/mL in insect cells (C6/36) and >1000 μg/mL in mammalian cells (Vero and LLCMK2). DS-01, when administered shortly after DENV-2 infection, is thought to potentially interfere with viral penetration and/or the release of viral particles from the endosomal vesicle (62). However, the researchers suggested that additional studies were needed to definitively determine the mechanism of DS-01’s antiviral activity on DENV-2 replication as well as its potential application as a therapeutic agent in the fight against dengue fever.
Approach and significance of targeting dengue infection in the mosquito vector
The studies highlighted above demonstrate the potential use of natural products for blocking dengue infection in mosquito cells. As the efficacy of these plant-based products can differ significantly between cell lines, the use of mosquito cells is critical to ensure that only viable candidates are further pursued in studies involving the mosquito vector. Researchers for example have previously demonstrated that while the antiviral drug favipiravir inhibits replication of chikungunya virus in mammalian and mouse models it does not do so in C6/36 cells as they lack an enzyme needed to convert the drug to its active form (63). Indeed, there is still much to be done as the studies discussed are all preliminary and the efficacy of these agents in vivo need to be investigated.
As the DENV transmission cycle involves mosquitoes and humans (64), targeting infection in either organism should potentially reduce dengue incidence. Over the years, antiviral research has primarily focused on human infection. However, due to a variety of impediments, research has only slowly progressed. Firstly, there is currently no ideal vertebrate animal model suited for anti-dengue screening because humans are the only known vertebrate hosts to develop the disease after DENV infection (64). Secondly, in the case of anti-dengue vaccines, there exists the risk of developing severe dengue disease upon recurrent infection by a heterologous virus serotype or immunization by a single DENV serotype (38). Lastly, dengue fever is complicated and multifaceted. Because DENV is an RNA virus with four unique serotypes that appear to be constantly evolving, tailoring antivirals to effectively target each serotype is difficult (65). Furthermore, there are usually significant delays between the time of infection after a mosquito bite, the onset of symptoms, and the time it takes to seek medical assistance (65). As a result, by the time dengue is diagnosed, it may be too late to treat it with antivirals.
Due to these ongoing challenges, preventing DENV infection/replication in—and—transmission by mosquitoes is now considered the most accessible and rational target for disease control (64). Dengue virus is transmitted to the midgut of mosquitoes following a blood meal from an infected host. There, epithelial cells are infected and the virus replicates before disseminating systemically through the hemocoel to secondary tissues. Following infection of the salivary glands and release of virions into the saliva, the virus can be transmitted to additional human hosts following subsequent feeding (66). To suppress dengue infection and replication in mosquitoes, antiviral compounds should ideally be administered to adult mosquitoes in a way that exposes them to the midgut tissue, which serves as both a point of entry and a key site for dengue replication (57). One way this could be achieved is by using attractive toxic sugar baits (ATSBs), a novel technique with recent implications for the management of adult mosquitoes in the field (57). The approach involves taking advantage of the mosquito’s propensity to feed on sugar and luring them into consuming artificial nectar laced with an insecticide. However, since synthetic insecticides are known to have deleterious effects on the environment and non-targeted organisms, the use of natural products in the baits could render them more environmentally safe.
Another approach is developing antivirals that hinder the completion of the pathogen’s life cycle in the mosquito (55). Antivirals which block viral transmission in the mosquito vector generally prevent pathogen transmission from infected to uninfected individuals by targeting molecules expressed on the surface of pathogens during their developmental phases within the insect vector or by targeting molecules expressed by the vectors (56). Several host factors in mosquitoes have recently been identified as playing important roles in dengue virus infection/replication in the mosquito vector. For example, Prohibitin, may act as a receptor protein to mediate DENV entry into mosquito cells (67) and mosquito galactose specific C-type lectins (mosGCTLs) act as susceptibility factors in Ae. aegypti mosquitoes to enhance DENV infection (68). Therefore, any substance that alters the expression or function of host factors in mosquitoes may ultimately reduce viral transmission from vector to uninfected hosts.
Added to this, the effect of these natural products and their metabolites on the mosquito’s capacity to spread pathogens must also be examined. Researchers have previously demonstrated for example that plant sugar meals influenced the ability of Anopheles coluzzi to transmit Plasmodium falciparum by affecting factors such as vector survival and fecundity, parasite development and infection prevalence and intensity (69). Likewise, several studies have illustrated that controlling the mosquito’s innate immune pathways reduces dengue virus production and infection in the mosquito (70–74). As such, how plant metabolites impact mosquito immunity must be explored. Recently, disruption of immune and oxidative stress pathways in addition to cuticle development was seen in Anopheles gambiae s.s. larvae exposed to phytochemicals from Murraya koenigii (75) and yeast-encapsulated orange oil increased expression of genes associated with apoptosis and innate immune responses in Ae. aegypti larvae (76).
Another important factor to consider is the effect of these natural products on the gut microbiome as studies have also highlighted its importance in arbovirus transmission (77). The mosquito gut microbiota is made up of viruses, bacteria, fungi, and protozoa, all of which have been demonstrated to play important roles in the mosquito’s physiological processes (77, 78). It has been previously shown that the microbiota can affect vector competence for infections in a variety of ways, including immune system activation or suppression, direct inhibition of the pathogen, or resource competition (77, 79). Researchers have noted the ability of various plant extracts to alter the gut microbiota of laboratory reared Ae. aegypti larvae and postulated that affecting these bacteria, will in turn, affect the vectorial competence and disease transmission by the mosquitoes (80).
Lastly, the concentration of the agents identified in vitro may be insufficient to inhibit viral infection in vivo depending on the route through which the mosquito obtains the antiviral agent. For instance, during blood-feeding, the female mosquito is known to excrete droplets of fluid in a process referred to as prediuresis (81, 82). Therefore, the possibility exists that a portion of the antiviral agent could be eliminated at this point. On the other hand, during a sugar meal, prediuresis is said to be rare or absent (82). Thus, incorporating the antiviral agent in sugar meals could be a better option to blood meals for evaluating anti-dengue activity of natural products in mosquitoes. The novel approach of blocking infection in the mosquito vector though not straightforward, could be the key to reducing the burden of dengue infection in the LAC region.
Summary and future perspectives
Currently, overall efforts to reduce dengue virus transmission have been insufficient to reduce the strain of dengue infection on the health and economic systems of LAC countries. During dengue epidemics, positive dengue cases frequently overburden hospitals and doctors’ offices. Likewise, businesses and workplaces are often impacted by employee absenteeism, resulting in millions of dollars in lost productivity. So far, the focus of DENV therapeutic efforts has been on the development of antivirals to target infection in the human host. However, with these efforts proving difficult to achieve and vector control strategies failing, targeting infection in the mosquito vector has emerged as a viable alternative approach. Some natural products have been shown to possess promising anti-dengue activity in mosquito cells as evident in this review. Yet, most of these studies are limited as they predominantly screen for anti-dengue activity in the C6/36 mosquito cell line using only one DENV serotype; typically, DENV-2 or DENV-1. Since these are not the only serotypes responsible for dengue infections, it is important for studies to routinely screen against all four serotypes. As evident by (61), not all serotypes may be equally susceptible to natural products when being screened. Studies in which natural products display favorable in vitro antiviral activity against DENV in C6/36 cells should ideally be followed up with in vivo assays such as screening in adult mosquitoes. This is because not all compounds that are active in vitro will produce similar results in vivo (83).
Notwithstanding the recent scientific advances in the fight against dengue virus, more work needs to be done in the LAC region especially in the Caribbean to combat dengue infections as severe dengue and its negative impacts remain a real threat. Despite the folkloric use of plants for many ailments in the Caribbean including dengue infection, to date, no study reporting the antiviral potential of plants has been documented in this region. This demonstrates emphatically the necessity for Caribbean researchers to capitalize on the untapped potential of plants in the region and seek to identify plants with antiviral activity against DENV. The approach of blocking dengue viral infection and or replication in the mosquito vector is worth further exploration as limiting the spread of the virus from vector to host is a promising avenue for ending the region’s war on dengue fever.
Author contributions
MAJG and SLS conceptualized the paper. MAJG wrote all drafts and SLS provided supervision. NKK wrote a section of the manuscript and KBA and TDW reviewed and edited the manuscript. All authors have read and agreed to the submitted version of the manuscript.
Acknowledgments
Mario Golding is a Global Infectious Diseases Scholar and received mentored research training in the development of this manuscript. This training was supported in part by the University at Buffalo Clinical and Translational Science Institute award UL1TR001412 and the Global Infectious Diseases Research Training Program award D43TW010919.
Conflict of interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Publisher’s note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
References
2. WHO. “Dengue and severe dengue” [Internet]. Geneva: World Health Organization (2022) [cited 2022 September 26]. Available from: https://www.who.int/news-room/fact-sheets/detail/dengue-and-severe-dengue.
3. San Martín JL, Solórzano JO, Guzmán MG, Brathwaite O, Bouckenooghe A, Zambrano B, et al. The epidemiology of dengue in the americas over the last three decades: a worrisome reality. Am J Trop Med Hygiene (2010) 82(1):128–35. doi: 10.4269/ajtmh.2010.09-0346
4. Ramos-Castañeda J, dos Santos FB, Martínez-Vega R, de Araujo JMG, Joint G, Sarti E. Dengue in Latin America: systematic review of molecular epidemiological trends. PloS Negl Trop Dis (2017) 11(1):e0005224. doi: 10.1371/journal.pntd.0005224
5. Bhatt S, Gething PW, Brady OJ, Messina JP, Farlow AW, Moyes CL, et al. The global distribution and burden of dengue. Nature (2013) 496(7446):504–7. doi: 10.1038/nature12060
6. Colón-González FJ, Harris I, Osborn TJ, Bernardo CSS, Peres CA, Hunter PR, et al. Limiting global-mean temperature increase to 1.5–2 °C could reduce the incidence and spatial spread of dengue fever in Latin america. Proc Natl Acad Sci (2018) 115(24):6243–48. doi: 10.1073/pnas.1718945115
7. Talarico LB, Noseda MD, Ducatti DR, Duarte ME, Damonte EB. Differential inhibition of dengue virus infection in mammalian and mosquito cells by iota-carrageenan. J Gen Virol (2011) 92(6):1332–42. doi: 10.1099/vir.0.028522-0
8. Cohen SN, Halstead SB. Shock associated with dengue infection. J Pediatr (1966) 68(3):448–56. doi: 10.1016/s0022-3476(66)80249-4
9. Halstead SB. "Mosquito-borne haemorrhagic fevers of south and south-East asia.". Bull World Health Organ (1966) 35(1):3.
10. Torres JR, Orduna TA, Piña-Pozas M, Vázquez-Vega D, Sarti E. Epidemiological characteristics of dengue disease in Latin America and in the Caribbean: a systematic review of the literature. J Trop Med (2017) 2017):1–18. doi: 10.1155/2017/8045435
11. Wilder-Smith A, Ooi E-E, Horstick O, Wills B. Dengue. Lancet (2019) 393(1016):350–63. doi: 10.1016/s0140-6736(18)32560-1
12. Tapia-Conyer R, Betancourt-Cravioto M, Méndez-Galván J. Dengue: an escalating public health problem in Latin america. Paediatr Int Child Health (2012) 32(sup1):14–7. doi: 10.1179/2046904712z.00000000046
13. Cafferata ML, Bardach A, Rey-Ares L, Alcaraz A, Cormick G, Gibbons L, et al. Dengue epidemiology and burden of disease in Latin America and the Caribbean: a systematic review of the literature and meta-analysis. Value Health Regional Issues (2013) 2(3):347–56. doi: 10.1016/j.vhri.2013.10.002
14. Shepard DS, Halasa YA, Zambrano B, Dayan GH, Coudeville L. Economic impact of dengue illness in the americas. Am J Trop Med Hygiene (2011) 84(2):200–7. doi: 10.4269/ajtmh.2011.10-0503
15. Laserna A, Barahona-Correa J, Baquero L, Castañeda-Cardona C, Rosselli D. Economic impact of dengue fever in Latin America and the Caribbean: a systematic review. Rev Panamericana Salud Pública (2018) 42. doi: 10.26633/rpsp.2018.111
16. Pan American Health Organization/World Health Organization. Epidemiological update: dengue. Washington, D.C: PAHO/WHO (2020).
17. Health Information Platform for the Americas (PLISA). “Reported cases of dengue fever in the americas”. PAHO/WHO (2022). Available at: https://www3.paho.org/data/index.php/en/mnu-topics/indicadores-dengue-en/dengue-nacional-en/252-dengue-pais-ano-en.html?showall=1.
18. Lopez-Gatell H, Hernandez-Avila M, Hernández Avila JE, Alpuche-Aranda. CM. Dengue in Latin America: a persistent and growing public health challenge. Negl Trop Dis (2015), 203–24. doi: 10.1007/978-3-7091-1422-3_11
19. Torres JR, Castro J. The health and economic impact of dengue in Latin america. Cadernos Saúde Pública (2007) 23(suppl 1):S23–31. doi: 10.1590/s0102-311x2007001300004
20. van den Hurk AF, Hall-Mendelin S, Pyke AT, Frentiu FD, McElroy K, Day A, et al. Impact of wolbachia on infection with chikungunya and yellow fever viruses in the mosquito vector Aedes aegypti. PloS Negl Trop Dis (2012) 6(11):e1892. doi: 10.1371/journal.pntd.0001892
21. Mackenzie JS, Gubler DJ, Petersen LR. Emerging flaviviruses: the spread and resurgence of Japanese encephalitis, West Nile and dengue viruses. Nat Med (2004) 10(S12):S98–S109. doi: 10.1038/nm1144
22. Almire F, Terhzaz S, Terry S, McFarlane M, Gestuveo RJ, Szemiel AM, et al. Sugar feeding protects against arboviral infection by enhancing gut immunity in the mosquito vector Aedes aegypti. PloS Pathog (2021) 17(9):e1009870. doi: 10.1371/journal.ppat.1009870
23. Caminade C, Medlock JM, Ducheyne E, Marie McIntyre K, Leach S, Baylis M, et al. Suitability of European climate for the Asian tiger mosquito Aedes albopictus: recent trends and future scenarios. J R Soc Interface (2012) 9(75):2708–17. doi: 10.1098/rsif.2012.0138
24. Méndez F, Parra B, Rengifo G, Burbano ME, Arias JF, Muñoz J, et al. Human and mosquito infections by dengue viruses during and after epidemics in a dengue–endemic region of colombia. Am J Trop Med Hygiene (2006) 74(4):678–83. doi: 10.4269/ajtmh.2006.74.678
25. Calderón-Arguedas O, Troyo A, Moreira-Soto RD, Marín R, Taylor L. Dengue viruses in Aedes albopictus skuse from a pineapple plantation in Costa rica. J Vector Ecol (2015) 40(1):184–86. doi: 10.1111/jvec.12149
26. Ricas Rezende H, Malta Romano C, Morales Claro I, Santos Caleiro G, Cerdeira Sabino E, Felix AC, et al. First report of Aedes albopictus infected by dengue and zika virus in a rural outbreak in brazil. PloS One (2020) 15(3):e0229847. doi: 10.1371/journal.pone.0229847
27. Garcia-Rejon JE, Navarro J-C, Cigarroa-Toledo N, Baak-Baak. CM. An updated review of the invasive Aedes albopictus in the americas; geographical distribution, host feeding patterns, arbovirus infection, and the potential for vertical transmission of dengue virus. Insects (2021) 12(11):967. doi: 10.3390/insects12110967
28. Ali I, Mundle M, Joshua J. Anzinger, and Simone l. sandiford. “Tiger in the sun: a report of Aedes albopictus in jamaica. Acta Tropica (2019) 199:105112. doi: 10.1016/j.actatropica.2019.105112
29. Bardach AE, García-Perdomo HA, Alcaraz A, López ET, Gándara RA, Ruvinsky S, et al. Interventions for the control of Aedes aegypti in Latin America and the Caribbean: systematic review and meta-analysis. Trop Med Int Health (2019) 24(5):530–52. doi: 10.1111/tmi.13217
30. Brown AW. Insecticide resistance in mosquitoes: a pragmatic review. J Am Mosq Control Assoc (1986) 2(2):123–40.
31. Guedes RN, Beins K, Costa DN, Coelho GE, Bezerra HS. Patterns of insecticide resistance in Aedes aegypti: meta-analyses of surveys in Latin America and the caribbean. Pest Manage Sci (2020) 76(6):2144–57. doi: 10.1002/ps.5752
32. Saillenfait A-M, Ndiaye D, Sabaté J-P. Pyrethroids: exposure and health effects – an update. Int J Hygiene Environ Health (2015) 218(3):281–92. doi: 10.1016/j.ijheh.2015.01.002
33. Hamed A, El-Sherbini M, Abdeltawab M. Eco-friendly mosquito-control strategies: advantages and disadvantages. Egyptian Acad J Biol Sci E Med Entomol Parasitol (2022) 14(1):17–31. doi: 10.21608/eajbse.2022.221601
34. Ansari MS, Moraiet MA, Ahmad S. Insecticides: impact on the environment and human health. Environ Deterioration Hum Health (2013), 99–123. doi: 10.1007/978-94-007-7890-0_6
35. Gouge DH, Li S, Walker K, Sumner C, Nair S, Olson C. “Mosquitoes: biology and integrated mosquito management”. The University of Arizona: Tucson, AZ, USA: CALS Publications Archive (2016).
36. Caceres Guido ,P, Ribas A, Gaioli M, Quattrone F, Macchi A. The state of the integrative medicine in Latin America: the long road to include complementary, natural, and traditional practices in formal health systems. Eur J Integr Med (2015) 7(1):5–12. doi: 10.1016/j.eujim.2014.06.010
37. Clement Y, Sutherland P, Moodley P, Chevannes B. Herbal medicine practices in the Caribbean. Taylor & Francis: Routledge (2014).
38. Kulkarni R. Antibody-dependent enhancement of viral infections. Dynamics Immune Activation Viral Dis (2020), 9–41. doi: 10.1007/978-981-15-1045-8_2
39. Scott R. Papaya leaf to the rescue - natural-medicine specialist suggests remedy for dengue fever. In: Papaya leaf to the rescue - natural-medicine specialist suggests remedy for dengue fever. Jamaica Gleaner: Lead Stories (2019). Available at: https://jamaica-gleaner.com/article/lead-stories/20191212/papaya-leaf-rescue-natural-medicine-specialist-suggests-remedy-dengue.
40. Wong M. “Vincentians turn to papaya leaf extract for dengue fever” [Internet]. Loop News (2020) [cited 2022 July 14]. Available from: https://caribbean.loopnews.com/content/vincentians-turn-papaya-leaf-extract-dengue-fever.
41. Sharine J. Dengue outbreak: 5 effective home remedies: loop st. Lucia. Loop News. Loop News (2020). Available at: https://stlucia.loopnews.com/content/5-effective-home-remedies-dengue-fever.
42. Fiscal RR. Ethnomedicinal plants used by traditional healers in Laguna, philippines. Asia Pacific J Multidiscip Res (2017) 5(4):132–37.
43. Teh BP, Ahmad NB, Mohamad SB, Tan TY, Razak MRMA, Afzan AB, et al. Carica papaya leaf juice for dengue: a scoping review. Nutrients (2022) 14(8):1584. doi: 10.3390/nu14081584
44. Yunita F, Hanani E, Kristianto J. "The effect of carica papaya l. leaves extract capsules on platelets count and hematocrit level in dengue fever patient.". Int J Med Aromat Plants (2012) 2(4):573–8.
45. Gadhwal AK, Ankit BS, Chahar C, Tantia P, Sirohi P, Agrawal RP. Effect of carica papaya leaf extract capsule on platelet count in patients of dengue fever with thrombocytopenia. J Assoc Physicians India (2016) 64(6):22–6.
46. WIC News. Dengue in Caribbean: how to treat the fever with home remedies? [Internet] WIC News (2019) [cited 2022 August 13]. Available from: https://wicnews.com/caribbean/dengue-caribbean-treat-fever-home-remedies-351723007/.
47. Parmar R. “10 health benefits of giloy - the ultimate immunity booster” [Internet]. PharmEasy Blog (2022) [cited 2022 November 17]. Available from: https://pharmeasy.in/blog/health-benefits-of-giloy-the-ultimate-immunity-booster/.
48. Leite FC, Mello CS, Fialho LL, Marinho CF, Lima ALA, Filho JM, et al. Cissampelos sympodialis has anti-viral effect inhibiting dengue non-structural viral protein-1 and pro-inflammatory mediators. Rev Bras Farmacognosia (2016) 26(4):502–6. doi: 10.1016/j.bjp.2016.03.013
49. Rodrigues JA, Eloy YR, Vanderlei ED, Cavalcante JF, Romanos MT, Benevides NM. An anti-dengue and anti-herpetic polysulfated fraction isolated from the coenocytic green seaweed caulerpa cupressoides inhibits thrombin generation in vitro. Acta Scientiarum Biol Sci (2017) 39(2):149. doi: 10.4025/actascibiolsci.v39i2.28081
50. Silva FC, Rodrigues VG, Duarte LP, Lula IS, Sinisterra RD, Vieira-Filho SA, et al. Antidiarrheal activity of extracts from maytenus gonoclada and inhibition of dengue virus by lupeol. Anais Da Acad Bras Ciências (2017) 89(3):1555–64. doi: 10.1590/0001-3765201720160046
51. Figueiredo GG, Coronel OA, Trabuco AC, Bazán DE, Russo RR, Alvarenga NL, et al. Steroidal saponins from the roots of solanum sisymbriifolium lam. (Solanaceae) have inhibitory activity against dengue virus and yellow fever virus. Braz J Med Biol Res (2021) 54(7):e10240. doi: 10.1590/1414-431x2020e10240
52. Gu F, Shi P-Y. The challenges of dengue drug discovery and development. Clin Invest (2014) 4(8):683–85. doi: 10.4155/cli.14.67
53. Lim SP. Dengue drug discovery: progress, challenges and outlook. Antiviral Res (2019) 163:156–78. doi: 10.1016/j.antiviral.2018.12.016
54. Troost B, Smit JM. Recent advances in antiviral drug development towards dengue virus. Curr Opin Virol (2020) 43:9–21. doi: 10.1016/j.coviro.2020.07.009
55. Londono-Renteria B, Troupin A, Colpitts TM. Arbovirosis and potential transmission blocking vaccines. Parasites Vectors (2016) 9(516). doi: 10.1186/s13071-016-1802-0
56. Bakhshi ,H, Failloux A-B, Sedigheh Z, Raz A, Djadid ND. Mosquito-borne viral diseases and potential transmission blocking vaccine candidates. Infect Genet Evol (2018) 63:195–203. doi: 10.1016/j.meegid.2018.05.023
57. Dong S, Dimopoulos G. Antiviral compounds for blocking arboviral transmission in mosquitoes. Viruses (2021) 13(1):108. doi: 10.3390/v13010108
58. Coutinho-Abreu IV, Ramalho-Ortigao M. Transmission blocking vaccines to control insect-borne diseases: a review. Memórias Do Instituto Oswaldo Cruz (2010) 105(1):1–12. doi: 10.1590/s0074-02762010000100001
59. Ono L, Wollinger W, Rocco IM, Coimbra TLM, Gorin PAJ, Sierakowski M-R. In Vitro And in vivo antiviral properties of sulfated galactomannans against yellow fever virus (BEH111 strain) and dengue 1 virus (Hawaii strain). Antiviral Res (2003) 60(3):201–8. doi: 10.1016/s0166-3542(03)00175-x
60. Talarico L, Pujol C, Zibetti R, Faria P, Noseda M, Duarte M, et al. The antiviral activity of sulfated polysaccharides against dengue virus is dependent on virus serotype and host cell. Antiviral Res (2005) 66(2-3):103–10. doi: 10.1016/j.antiviral.2005.02.001
61. Silva ARA, Morais SM, Marques MMM, Lima DM, Santos SCC, Almeida RR, et al. "Antiviral activities of extracts and phenolic components of two spondias species against dengue virus.". J Venomous Anim Toxins Including Trop Dis (2011) 17:406–4135. doi: 10.1590/S1678-91992011000400007
62. Cardoso JL, Soares MJ, Leite JR, Malaquias LC, Coelho LF. Antiviral activity of dermaseptin 01 against dengue virus type 2, herpes simplex virus type 1 and vaccinia virus. Sci Med (2013) 23(1):18. doi: 10.15448/1980-6108.2013.1.12751
63. Jacobs S, Wang L, Rosales Rosas AL, Van Berwaer R, Vanderlinden E, Failloux A-B, et al. Favipiravir does not inhibit chikungunya virus replication in mosquito cells and Aedes aegypti mosquitoes. Microorganisms (2021) 9(5):944. doi: 10.3390/microorganisms9050944
64. Blair CD, Olson KE. Targeting dengue virus replication in mosquitoes. Arthropod Vector: Controller Dis Transm (2017) 1:201–17. doi: 10.1016/b978-0-12-805350-8.00012-x
65. Newton W. “Dengue fever: the urgent hunt for antivirals and vaccines. Clinical Trials Arena (2022). Available at: https://www.clinicaltrialsarena.com/analysis/dengue-fever-antivirals-vaccines/#:~:text=Manufacturers%20have%20struggled%20to%20develop,potential%20deficits%20in%20preclinical%20data.
66. Raquin V, Lambrechts L. Dengue virus replicates and accumulates in Aedes aegypti salivary glands. Virology (2017) 507:75–81. doi: 10.1016/j.virol.2017.04.009
67. Kuadkitkan A, Wikan N, Fongsaran C, Smith. DR. Identification and characterization of prohibitin as a receptor protein mediating DENV-2 entry into insect cells. Virology (2010) 406(1):149–61. doi: 10.1016/j.virol.2010.07.015
68. Liu Y, Zhang F, Liu J, Xiao X, Zhang S, Qin C, et al. Transmission-blocking antibodies against mosquito c-type lectins for dengue prevention. PloS Pathog (2014) 10(2):e1003931. doi: 10.1371/journal.ppat.1003931
69. Hien DF, Dabiré KR, Roche B, Diabaté A, Yerbanga RS, Cohuet A, et al. Plant-mediated effects on mosquito capacity to transmit human malaria. PloS Pathog (2016) 12(8):e1005773. doi: 10.1371/journal.ppat.1005773
70. Xi Z, Ramirez JL, Dimopoulos G. The Aedes aegypti toll pathway controls dengue virus infection. PloS Pathog (2008) 4(7):e1000098. doi: 10.1371/journal.ppat.1000098
71. Sánchez-Vargas I, Scott JC, Poole-Smith BK, Franz AW, Barbosa-Solomieu V, Wilusz J, et al. Dengue virus type 2 infections of Aedes aegypti are modulated by the mosquito's RNA interference pathway. PloS Pathog (2009) 5(2):e1000299. doi: 10.1371/journal.ppat.1000299
72. Souza-Neto JA, Sim S, Dimopoulos G. An evolutionary conserved function of the JAK-STAT pathway in anti-dengue defense. Proc Natl Acad Sci (2009) 106(42):17841–6. doi: 10.1073/pnas.0905006106
73. Jupatanakul N, Sim S, Angleró-Rodríguez YI, Souza-Neto J, Das S, Poti KE, et al. Engineered Aedes aegypti JAK/STAT pathway-mediated immunity to dengue virus. PloS Negl Trop Dis (2017) 11(1):e0005187. doi: 10.1371/journal.pntd.0005187
74. Lee W-S, Webster JA, Madzokere ET, Stephenson EB, Herrero LJ. Mosquito antiviral defense mechanisms: a delicate balance between innate immunity and persistent viral infection. Parasites Vectors (2019) 12(1). doi: 10.1186/s13071-019-3433-8
75. Mang’era CM, Khamis FM, Awuoche EO, Hassanali A, Ombura FL, Mireji PO. Transcriptomic response of Anopheles gambiae sensu stricto mosquito larvae to curry tree (Murraya koenigii) phytochemicals. Parasites Vectors (2021) 14(1). doi: 10.1186/s13071-020-04505-4
76. Kelly PH, Yingling AV, Ahmed A, Hurwitz I, Ramalho-Ortigao M. Defining the mechanisms of action and mosquito larva midgut response to a yeast-encapsulated orange oil larvicide. Parasites Vectors (2022) 15(1). doi: 10.1186/s13071-022-05307-6
77. Hegde S, Rasgon JL, Hughes GL. The microbiome modulates arbovirus transmission in mosquitoes. Curr Opin Virol (2015) 15:97–102. doi: 10.1016/j.coviro.2015.08.011
78. Huang W, Wang S, Jacobs-Lorena M. Use of microbiota to fight mosquito-borne disease. Front Genet (2020) 11:196. doi: 10.3389/fgene.2020.00196
79. van Tol S, Dimopoulos G. Influences of the mosquito microbiota on vector competence. Adv Insect Physiol (2016) 51:243–91. doi: 10.1016/bs.aiip.2016.04.006
80. Gupta KK, Prejneshu M, Aggarwal M, Keshvam CC, Dubey VK, Mishra MK, et al. A strategy to decrease vectorial competence of dengue mosquito Aedes aegypti by alteration of its gut microbiota using Indian traditional medicinal plants. Int J Trop Insect Sci (2021) 41(4):2947–56. doi: 10.1007/s42690-021-00479-7
81. Benoit JB, Denlinger DL. Meeting the challenges of on-host and off-host water balance in blood-feeding arthropods. J Insect Physiol (2010) 56(10):1366–76. doi: 10.1016/j.jinsphys.2010.02.014
82. Lahondère C, Lazzari CR. Thermal stress and thermoregulation during feeding in mosquitoes. Anopheles Mosquitoes New Insights Into Malaria Vectors (2013). doi: 10.5772/56288
Keywords: Caribbean, Latin America, natural products, dengue, antiviral
Citation: Golding MAJ, Khouri NK, Anderson KB, Wood TD and Sandiford SL (2023) The use of natural products in Latin America and the Caribbean for blocking dengue infection in mosquito cells. Front. Virol. 3:1130171. doi: 10.3389/fviro.2023.1130171
Received: 06 January 2023; Accepted: 12 April 2023;
Published: 28 April 2023.
Edited by:
Shengzhang Dong, Johns Hopkins University, United StatesReviewed by:
Yingjun Cui, Yale University, United StatesChaker El Kalamouni, Université de la Réunion, France
Sheng-Qun Deng, Anhui Medical University, China
Copyright © 2023 Golding, Khouri, Anderson, Wood and Sandiford. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Simone L. Sandiford, c2ltb25lLnNhbmRpZm9yZEB1d2ltb25hLmVkdS5qbQ==