We investigated the influence of hand posture in handedness recognition, while varying the spatial correspondence between stimulus and response in a modified Simon task. Drawings of the left and right hands were displayed either in a back or palm view while participants discriminated stimulus handedness by pressing either a left or right key with their hands resting either in a prone or supine posture. As a control, subjects performed a regular Simon task using simple geometric shapes as stimuli. Results showed that when hands were in a prone posture, the spatially corresponding trials (i.e., stimulus and response located on the same side) were faster than the non-corresponding trials (i.e., stimulus and response on opposite sides). In contrast, for the supine posture, there was no difference between corresponding and non-corresponding trials. Control experiments with the regular Simon task showed that the posture of the responding hand had no influence on performance. When the stimulus is the drawing of a hand, however, the posture of the responding hand affects the spatial correspondence effect because response location is coded based on multiple reference points, including the body of the hand.
In choice reaction time (RT) tasks, compatibility between elements of the stimulus and response sets influences performance, as measured by the speed and accuracy of the subject’s motor response (e.g., Kornblum et al., 1990
). For example, the manual response to a visual cue is usually faster and more accurate when the response key has the same relative location to the subject’s midline as the stimulus. The importance of the spatial location of the stimulus on stimulus-response (S-R) compatibility tasks is underscored by its influence on the subject’s performance even when it is not directly relevant to response selection, as shown by the Simon and spatial Stroop tasks (Umiltà and Nicoletti, 1990
; Lu and Proctor, 1995
).
The stimuli used in S-R compatibility tasks have been of a wide variety. Parts of the body, however, have never been used, even though they would make an interesting type of stimulus because they are processed differently by the brain. There is ample evidence showing that visual information about ordinary objects and visual information about parts of the human body are relatively segregated in different pathways, with specialized cortical areas devoted to the processing of information related to each category (Downing et al., 2001
; Parsons, 2003
; Zacks et al., 2003
; Urgesi et al., 2007
). In particular, information about the body is processed in areas of the mirror neuron system (MNS), whose neurons are active both during the execution and the observation of a movement (see Gallese et al., 1996
; Rizzolatti and Craighero, 2004
; Gawryszewski et al., 2007
).
To recognize visual objects, their perceptual characteristics must be compared to stored representations. Because visual objects are seen from different viewpoints, they must first be subjected to mental transformations aimed at reorienting them in space. Such strategy, in the case of body parts, is complicated by the fact that their internal representation is built from visual and somatosensory information (Schendan and Stern, 2007
). For instance, a simple task, such as judging the handedness of a drawing of the human hand, requires motor imagery that follows the same rules of the real movement, including compliance to biomechanical and physical constraints (Parsons, 1994
; Vargas et al., 2004
; De Lange et al., 2006
). More specifically, handedness judgment also involves a pre-attentive handedness-recognition process before the mental simulation of one’s own hand moving towards the stimulus (Parsons, 1987
, 1994
; Parsons and Fox, 1998
; Parsons et al., 1998
). It is important to stress the distinction between these two sequential phases: the automatic recognition of the hand (first phase) and the subsequent confirmatory motor imagery (second phase), even though they can share neural substrates (Gentilucci et al., 1998
).
The basic assumption that motivated the current study was that the confirmatory motor imagery process (second phase) necessary for handedness recognition might be influenced by proprioceptive inputs. Previous studies had already shown the basic influence of proprioceptive information on motor imagery. Parsons (1994)
, for instance, showed that mental rotation of the hand is faster when the subjects’ own hands assume a “canonical” posture (palms down on a table) instead of an “awkward” one (palms facing away from the subject’s midline). A similar result was obtained with children by Funk et al. (2005)
, while Ionta et al. (2007) and Ionta and Blanke (2009)
showed that proprioceptive influence on motor imagery depends on somatotopic correspondence. In a related study, Vargas et al. (2004)
investigated, by means of Transcranial Magnetic Stimulation (TMS), how corticospinal excitability, facilitated during the mental simulation of a hand movement, is affected by the actual posture of the hand. Participants were asked to imagine themselves joining the tips of two fingers while maintaining a hand posture either compatible or incompatible with the imagined movement. Results indicated that the actual limb posture exerted a modulatory effect upon the motor imagery process.
A similar result was obtained by De Lange et al. (2006)
in an fMRI study which found that the posture of the participants’ own left and right arm influenced handedness judgments through modulation of a parietal-frontal network, whose activity increased with increasing biomechanical complexity of the imagined hand movements. That result indicates that motor imagery depends on the current configuration of the limbs, as determined by proprioceptive inputs.
Here, we further elucidated the role played by proprioceptive inputs on handedness discrimination by using a Simon-like S-R compatibility task, in which handedness of the drawings is the relevant feature for response selection while spatial location is the irrelevant one
1
. We investigated the effects of proprioceptive inputs on the task by asking participants to have their hands assume either a prone or a supine position. The spatial correspondence effect, characteristic of regular S-R compatibility tasks occurs because irrelevant information about the stimulus location interferes, at the response selection stage, with the learned association between the relevant stimulus feature and the correct response. In our modified task, there is also the possibility for the irrelevant proprioceptive information about the posture of the responding hand to conflict with the sensorimotor representation normally engaged by motor imagery. Thus, we asked how those two irrelevant sources of information interacted with each other and with the relevant feature to affect the dynamics of the response in a handedness judgment task.
Materials and Methods
We tested two groups, a handedness-task group and a regular Simon task group.
Participants
Handedness task. Twelve right-handed volunteers (eight male and four female, 18–22 years old, mean = 19.5 years) participated in the study.
Regular Simon task. Sixteen right-handed volunteers (eight male and eight female, 17–29 years old, mean = 21 years) participated in the experiment.
All had normal visual acuity and were naïve as to the purposes of the experiment. All participants provided written informed consent and the study was approved by our Institution’s Research Ethics Committee in accordance with the Helsinki Declaration (2008)
.
Stimuli
Handedness task. Drawings representing the human left and right hand in either back or palm views were presented randomly either to the left or to the right of the central fixation point. Left and right hands were mirror images of each other. Stimuli were about 15.5° high and 9.3° wide and had a black profile set against a gray background. Stimuli were located 7.5° either to the left or to the right of the central fixation point and were always presented with the fingers pointing upwards (see Figure 1
).
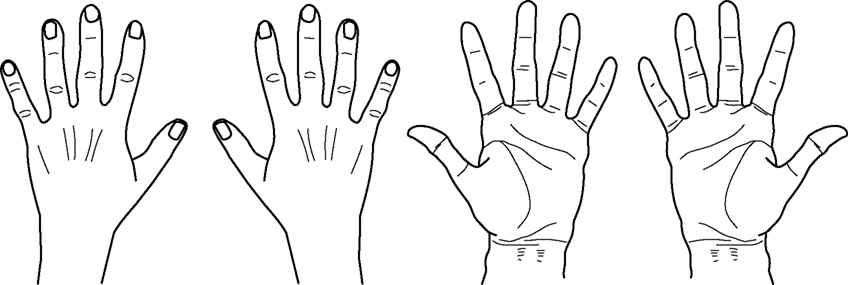
Figure 1. Drawings of left and right hands in back and palmar views (modified from Parsons, 1994
).
Regular Simon task. Drawings of geometric figures (squares and circles comprising 1° of visual angle) were presented on a 20-in VGA monitor screen. The geometric center of the stimuli were located 7.5° either to the left or to the right of the central fixation point.
Apparatus
The experiment was conducted in a quiet and dimly lit room. A personal computer (PC 486) was used both for stimulus presentation and recording the response. The head of the participants was positioned in a forehead-and-chin rest and the distance between the eyes and the screen was about 57 cm (see Figure 2
). The Micro Experimental Laboratory software (MEL, version 2.0) was used to determine events’ sequences and to measure response latency. The stimulus remained on the screen for 1000 ms and the subjects responded by pressing one of two micro-switches, located 25 cm to the left or to the right. Participants employed two hand postures (prone and supine) when responding. In both postures, they executed the same index finger movement (flexion) to press the response key, since the response apparatus allowed an 180° turn (see Figure 2
). We used an Eye Track System (Model 210 – Applied Science Laboratories) to control the subject’s eye fixation during the test.
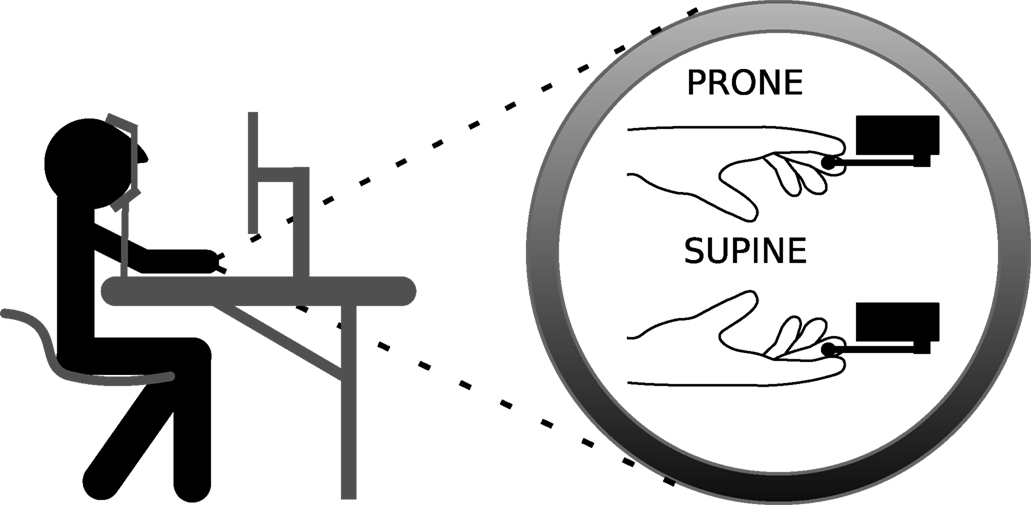
Figure 2. Experimental setup and the two hand postures assumed by the participants.
Task
Handedness task. The task was to press the right key to the appearance of a right-hand drawing and the left key to the appearance of a left-hand drawing. The relevant feature for response selection was stimulus’ handedness (left or right hand drawing) and the irrelevant feature was its location (left or right hemifield).
Regular Simon task. The task was to press a micro-switch (right or left) when a stimulus (circle or square) appeared. Usually, in this regular Simon task, the corresponding condition (stimulus ipsilateral to the response key) elicits faster responses than the non-corresponding condition (stimulus contralateral to the response key).
Procedure
Handedness task. Participants attended two sessions on different days. Each session was divided into four blocks of 68 trials, resulting in 272 trials per session. In two consecutive blocks, the participant responded with his/her palms face down (prone posture) and, in the other two blocks, with his/her palms face up (supine posture). The participants started with a posture in one session and with another posture in the other session. The order of the postures was counterbalanced across participants. Thus, the hand posture matched the view of the drawings on 50% of trials. The left or right hand drawings were presented randomly in either the back or palm view. We did not systematically ask the subjects about their strategy, only occasionally. Because hand views (palm or back) were randomly presented, it was not possible for the participant to use a gross asymmetric feature of the drawing, such as the relative position of the thumb, for handedness recognition.
Participants were instructed: (i) to maintain the gaze at the central fixation point, (ii) to avoid looking at the stimulus and (iii) to respond as fast as possible by pressing the right micro-switch to a right-hand stimulus and the left micro-switch to a left-hand stimulus.
Regular Simon task. There were three sessions on different days. The first session was considered practice and was not included in the analysis. Each session was subdivided into four blocks of 80 trials, resulting in 320 trials per session. Participants responded either with their palms face down (prone posture) in two blocks or with their palms face up (supine posture) in the other two. They were instructed to keep their gaze at the central fixation point until the stimulus appeared, avoid looking at the stimulus itself and to respond as fast as possible to its appearance. Half of the participants used the right micro-switch for the square and the left micro-switch for the circle. The others had the reverse assignment.
Experimental conditions were counterbalanced across subjects to account for practice or fatigue effects. Participants performed two consecutive blocks with the same posture.
Analyses
Central tendency measurements
The mean of correct Manual Reaction Time (MRT) values was entered into an ANOVA with one between-participant factor, task (Handedness task and regular Simon task) and two within-participant factors, hand posture (prone and supine) and correspondence (corresponding and non-corresponding conditions). A corresponding trial was the condition in which the stimulus appeared on the same side of the response key. For example, when the right-hand (left-hand) drawing appeared in the right (left) visual hemifield and the participant responded by pressing the right (left) key. A non-corresponding trial was the condition in which the stimulus appeared in the opposite side of the response key, for example, when the right-hand (left-hand) drawing appeared in the left (right) visual hemifield and the participant responded by pressing the right (left) key. We chose to use only right-handed subjects in order to have an homogeneous sample. Besides, we believe that the eventual existence of a general bias toward faster responses for right stimuli in this sample has no effect on our main goal of verifying the effect of hand posture on handedness recognition The Newman-Keuls method was used for post-hoc analyses, with α < 0.05. When interpreting statistics one should bear in mind, however, that they refer to two groups with different sizes.
Delta-plot analysis
The Delta-plot procedure (Ridderinkhof, 2002
) is based on correct MRT distributions for corresponding and non-corresponding trials in both prone and supine postures. These rank-ordered distributions were divided into five proportional bins (quintiles) such that each one contained the same number of trials (one-fifth). Mean MRTs for each bin was calculated and the size of the correspondence effect (mean MRT in non-corresponding trials minus mean MRT in corresponding trials) was plotted as a function of response speed for each hand posture. The correspondence effect amplitudes were entered into an ANOVA with task (Handedness task and Simon task) as a between-participant factor and hand posture (prone and supine) and bin (1st bin, 2nd bin, 3rd bin, 4th bin and 5th bin) as within-participant factors. The Newman-Keuls method was used for post-hoc analyses, with α < 0.05.
Results
Errors
Overall errors amounted to 5.81%, of which 4.17% were judgment errors, 1.57% were omission errors and 0.07% were anticipation errors. Errors were entered into an ANOVA with two within-participant factors, hand posture (prone and supine) and correspondence (corresponding and non-corresponding conditions) and no sources of variance were statistically significant.
Central tendency measures
The significant main effects were task (F1,26 = 283.75, p < 0.0025), posture (F1,26 = 6.18, p < 0.025) and correspondence (F1,26 = 7.11, p < 0.025). The average MRT for the regular Simon task (424 ± 4 ms) was significantly faster than MRT for the Handedness task (655 ± 7 ms). The MRT for the prone posture (515 ± 6 ms) was faster than for the supine posture (531 ± 6 ms). The MRT for the corresponding condition (517 ± 6 ms) was faster than non-corresponding condition (528 ± 6 ms). There was also a significant interaction between hand posture and correspondence (F1,26 = 10.35, p < 0.0035) and there was a three-way interaction between task, posture and correspondence (F1,26 = 5.15, p < 0.035).
The hand posture and correspondence interaction showed that, when the hands were in the prone posture, MRT for the corresponding condition (507 ± 6 ms) was significantly faster (p < 0.0002) than MRT for the non-corresponding condition (523 ± 5 ms). However, when hands were in the supine posture, there was no significant difference between corresponding and non-corresponding conditions (528 ± 6 ms vs. 534 ± 4 ms) (p > 0.08).
The three-way interaction showed that in the Handedness task the corresponding condition (636 ± 8 ms) was faster (p < 0.0048) than the non-corresponding (653 ± 8 ms) condition when the participants were in the prone posture, whereas, with the supine posture there was no difference between corresponding (668 ± 7 ms) and non-corresponding (665 ± 5 ms) conditions (see Figure 3
) (p > 0.05). For the Simon task, the corresponding condition was significantly faster (p < 0.035) than the non-corresponding condition in both postures (prone 410 ± 4 ms vs. 426 ± 3 ms and supine 423 ± 5 ms vs. 435 ± 4 ms) (p < 0.005).
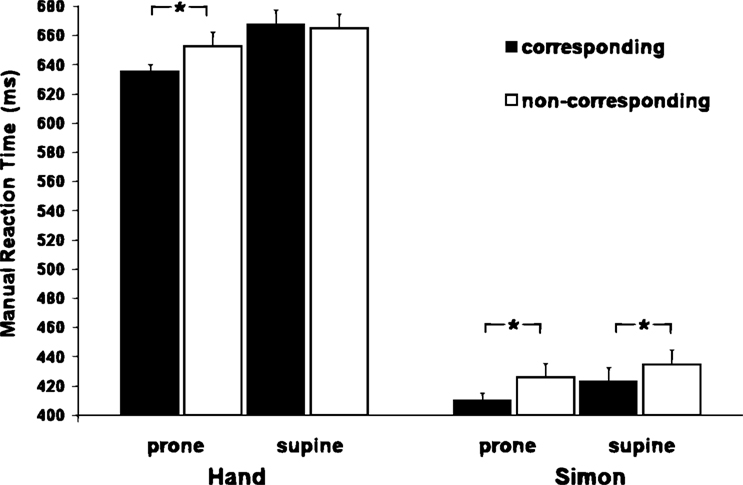
Figure 3. MRT as a function of hand posture and the spatial correspondence between stimulus and response in both, Handedness task and Simon task. Notice the influence of the posture on S-R compatibility only in the handedness recognition. This correspondence effect disappears when the hands assumed a supine posture.
Delta-plot analysis
Both hand posture (F1,26 = 9.71; p < 0.005) and bin (F4,104 = 18.32; p < 0.0001) were significant main effects in the Delta-plot analysis. There was also a significant interaction between task and posture (F1,26 = 7.21; p < 0.015). Overall, there was no significant difference between the correspondence effect amplitude in both prone (11 ± 3 ms) and supine (10 ± 2 ms) postures in the regular Simon task (p > 0.45). However, in the Handedness task, the amplitude of the correspondence effect in the prone posture (17 ± 2 ms) was significantly larger (p < 0.025) than in the supine posture (−3 ± 2 ms; see Figure 4
).
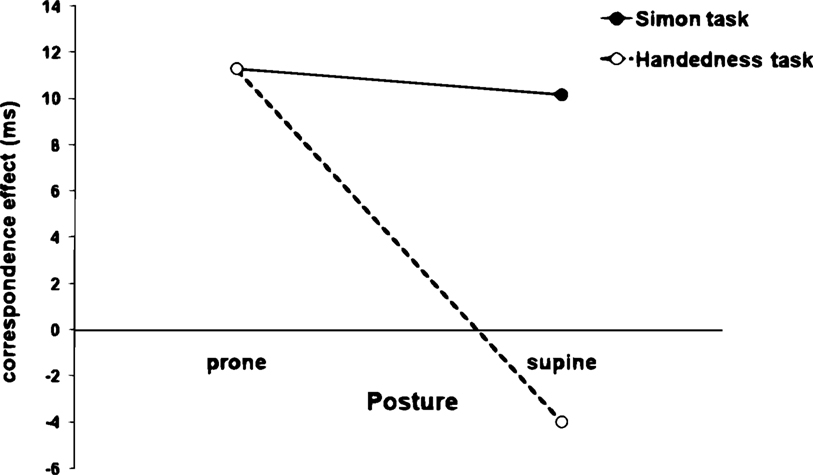
Figure 4. Influence of hand posture on amplitude of the correspondence effect for each task. There is no difference between the correspondence effect amplitude in both postures in the regular Simon task. However, in the Handedness task, the amplitude of the correspondence effect in the prone posture is larger than in the supine posture.
For both tasks and both postures, the correspondence effect decreased as a function of bin. For the handedness task, the correspondence effect for both postures was present and significant at the first and second bin, 26 (p < 0.0015) and 17 ms (p < 0.035), respectively, and practically disappeared at the slowest three bins (respectively, 3, −9 and 0 ms). For the regular Simon task, the correspondence effect for both postures was present and significant at the first three bins, 24 (p < 0.001), 20 (p < 0.0213) and 14 ms (p < 0.0263), respectively, and practically disappeared at the two slowest bins (respectively, 3 and −7 ms). The three-way interaction was not significant, indicating that posture did not modulate the trend of the correspondence effect as a function of bin in either task. The size of the correspondence effect for the prone posture was larger than for the supine posture at all bins.
The most important finding was no doubt the interaction between task (regular Simon task or Handedness task), hand posture (prone or supine) and correspondence (corresponding or non-corresponding conditions). In the Handedness task, when the hands assumed a prone posture, MRTs for the corresponding condition were faster than MRTs for the non-corresponding condition. This correspondence effect was no longer present, however, when the hands assumed a supine posture. In contrast, in the regular Simon task, for both postures, MRTs for the corresponding condition were faster than MRTs for the non-corresponding condition. The Delta-plot analysis confirmed this pattern and showed it was not affected by response speed.
Because in the previous analyses the view of the hand stimulus was not taken into consideration, we decided to conduct a separate analysis of the Handedness task.
Central Tendency Measurements
Correct MRTs were entered into an ANOVA with the following factors: hand posture (prone or supine), match (matching or non-matching condition) and correspondence (corresponding or non-corresponding condition). A matching trial was the condition in which the participant’s hand was in the prone (supine) posture and the stimulus appeared in back (palm) view and vice-versa for the non-matching trial. The Newman-Keuls method was used for post-hoc analyses, with α < 0.05.
Results
Only the significant sources of variance involving the factor match will be reported here. The others were of course a replication of those already reported above. The main effect of match was significant (F1,11 = 15.12; p < 0.0025). The average MRT for the matching condition (641 ± 8 ms) was faster (p < 0.025) than the MRT for the non-matching condition (669 ± 6 ms).
There was also a significant interaction between hand posture and match (F1,11 = 13.04, p < 0.0045). When the hands were in the prone posture, the average MRT for the matching condition (back view – 613 ± 7 ms) was faster (p < 0.0045) than the average MRT for the non-matching condition (palm view 675 ± 5 ms). In contrast, when the hands were in the supine posture, there was no significant difference between matching and non-matching conditions (back view 670 ± 5 ms vs. palm view 663 ± 5 ms; see Figure 5
). Errors were also analyzed with the ANOVA, but no sources of variance were found to be significant.
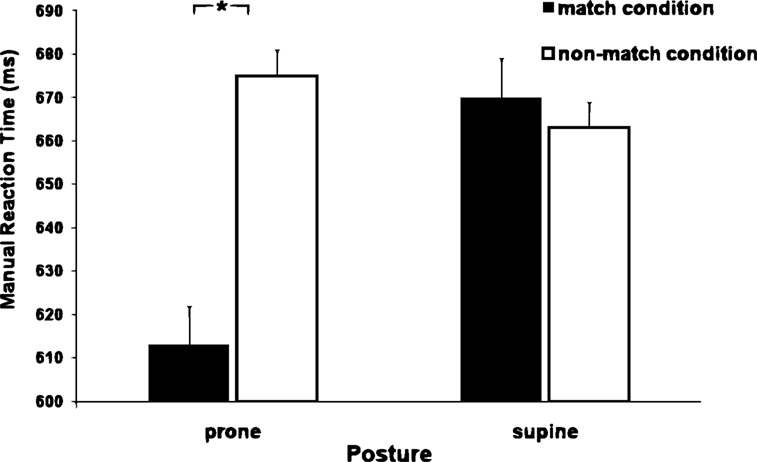
Figure 5. MRT as a function of hand posture and match conditions. Notice the interaction between hand posture and match condition. In the prone posture, MRT for the matching condition is faster than the MRT for the non-matching condition. However, in the supine posture there is no significant difference.
The novel finding with respect to the previous analyses was the interaction between hand posture (prone or supine) and stimulus views (back or palm view). With the hands in the prone posture and the stimulus in back view, MRTs were faster than when the stimulus was in palm view. In the supine posture, however, there was no difference between the back and palm views. Interestingly, as had happened with the correspondence effect, the match effect disappeared when the responses were executed with the hands in supine posture. Apparently, the supine posture has something “special” that contributes to annul effects that would otherwise manifest themselves.
In this paper, we have used a modified Simon task to investigate the influence of hand posture on handedness recognition of drawings of the human hand presented either on back or palm view. The participants had to ascertain stimulus handedness by pressing either left or right keys with their hands resting either in a prone or supine posture. As a control, subjects performed a regular Simon task using simple geometric shapes as stimuli.
We have shown that there is no influence of proprioceptive input from the responding hand on a Simon-like S-R compatibility task, with drawings of the human hand acting as the relevant stimulus, when the responding hand is in a supine posture. In contrast, with the responding hand in a prone position, not only there was a spatial correspondence effect, but proprioceptive information from the responding hand was also shown to interact with motor imagery used to perform the handedness recognition task. In control experiments, when geometric shapes were used as relevant stimuli, we found results typical of a regular Simon task, without interference from proprioceptive inputs: MRTs for the corresponding condition were faster than MRTs for the non-corresponding condition, regardless of the posture of the responding hand. The understanding of why and how stimulus properties affect performance in tasks like the present one is important because it may clarify how these properties are used for action selection.
A matching effect was observed in separate analysis of the handedness task (see the corresponding section). In our handedness recognition task, when the participant’s hands were in a prone posture, the MRTs for the back view of the stimulus (matching condition) were faster than for the palmar view (non-matching condition). That extends previous findings regarding the influence of proprioceptive information on motor imagery (Parsons, 1994
; Sirigu and Duhamel, 2001
; Vargas et al., 2004
; Funk et al., 2005
; De Lange et al., 2006
; Ionta and Blanke, 2009
). The fact that proprioceptive inputs from the hands in supine posture did not influence MRTs is in accordance with previous findings that the time needed to perform a handedness recognition task is longer when the responding hands are kept in an awkward posture (Parsons, 1994
; Sirigu and Duhamel, 2001
; Funk et al., 2005
; Ionta and Blanke, 2009
). The supine posture is not as “canonical” as the prone (people see their own hands in the latter posture more often) and this would hinder the MRTs for drawings of hands in this view (Parsons, 1994
; Sirigu and Duhamel, 2001
).
The distributional analysis (Ridderinkhof, 2002
), which has been shown elsewhere to be particularly useful in studying the role of activation and suppression in conflicting tasks (e.g., De Jong et al., 1994
), was used in the present work to reveal the temporal dynamics of the correspondence effect in the prone and supine postures. Our results showed that in the handedness task, the magnitude of the correspondence effect was larger in the prone than in the supine posture for every bin while in the classic Simon task there is no difference in the correspondence effect amplitude between the prone and supine postures. There is ample evidence indicating that the relative speed of processing either the irrelevant or relevant information influences the magnitude of the correspondence effect (e.g., Hommel, 1993
; De Jong et al., 1994
; Lu and Proctor, 1995
, 2001
). The irrelevant spatial code for stimulus location, which is presumed to be available instantly, has a greater chance of interacting with the relevant stimulus code and thus influencing response selection when both coincide temporally. Thus, any delay in identifying the relevant stimulus dimension has the potential to reduce or even eliminate the correspondence effect. This is confirmed by our observation that the correspondence effect decreases as a function of response speed (see the Delta-plot analysis). In theory, this fact could be used to explain why we found no difference in MRT for the corresponding and non-corresponding conditions for the supine posture. However, the difference between the prone and supine posture’s MRT (about 15 ms) does not seem to be large enough to justify this hypothesis. An alternative explanation takes into consideration that response location is coded according to multiple reference frames, including the body of the hand (Cho and Proctor, 2002
). Thus, the different effect the prone and supine postures have on the correspondence effect arises due to the fact that the response switch is located on different sides of the body of the hand in each case (Cho and Proctor, 2002
).
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
We are grateful to Dr. L.M. Parsons, who kindly gave permission to use the hand drawings. We also thank Professors Giacomo Rizzolatti and Giovanni Berlucchi for important discussions on the manuscript and all subjects who participated in the experiments.
This study was a part of APL MSc. Thesis and was supported by grants from CNPq/MCT, FAPERJ, CAPES, PROPP-UFF, UFF. CU was supported by a grant from MiUR (PRIN 2007).
- ^ In the present study, both the relevant and the irrelevant stimulus feature overlapped with the relevant response feature (see Kornblum et al., 1990 ). Therefore, the task resembles a spatial Stroop task (e.g., Lu and Proctor, 1995 ). However, the relevant right/left stimulus feature is not spatial in nature but rather concerns handedness. Therefore, the most appropriate term would be Simon-like S-R compatibility task. We decided to use, instead, the neutral term Handedness task.
W.M.A. (2008). World Medical Association Declaration of Helsinki – Ethical Principles for Medical Research Involving Human Subjects. In World Medical Association. Retrieved October 30, 2009, Available at: http://www.wma.net/en/30publications/10policies/b3/index.html
.