 Mayuko Nomoto
Mayuko Nomoto Etienne François Akomo-Okoue
Etienne François Akomo-Okoue- 1Graduate School of Science, Kyoto University, Kyoto, Japan
- 2Institut de Recherche en Ecologie Tropicale (IRET/CENAREST), Libreville, Gabon
Introduction: With increasing human activities, the area affected by humans increases annually and many wild animals are under threat of extinction. On the other hand, there are serious conflicts between human and many wildlife. In order to mitigate conflicts, it is important to understand how animals perceive the balance of benefits and risks in human-inhabited areas and how they use the land. In this study, we investigated how African forest elephants (Loxodonta cyclotis), which are in serious conflict with humans, use anthropogenic environments by focusing on their trails.
Methods: This study targeted two parameters of anthropogenic environments: vegetation, particularly savannas, which have been maintained by human burning for many years, and population density, particularly in low-density seminatural land. Within Moukalaba-Doudou National Park, Gabon, and an adjacent village, we mapped elephant trails and compared their density and structure between vegetation (forest and savanna) and areas (inside and outside the national park).
Results: In the forest, only 31.4% of the elephant trails were well-established with exposed ground. In the savanna, the density of elephant trails was lower than that in the forest; however, the trails were narrower and 65% of the trails were well established. This suggests that elephants tend to follow specific routes in the savanna that may help them travel more easily, and that they are perceiving some risks. The density of elephant trails outside the national park was higher, and the trails were wider than those in the national park, suggesting frequent elephant use. This was probably due to the crop fields and mango trees attracting elephants.
Discussion: In the study area, available food may be a greater attractant to elephants relative to the risk of being threatened by humans. The findings show that elephant trail surveys can be carried out inexpensively over a short period of time. Moreover, they can clarify land use by elephant populations, including fine-scale heterogeneous movements.
1 Introduction
The extinction of animal species and populations and the degradation of their habitats due to human activities are pressing global issues (Dirzo et al., 2014; Ceballos et al., 2015). The impact of humans is not only on a direct level, such as overhunting, but also on an indirect level, such as deforestation, affecting many other animals (Brook et al., 2008; Haddad et al., 2015; Bogoni et al., 2022). Large terrestrial mammals are at an extremely high risk of extinction (Cardillo et al., 2005; Ripple et al., 2015; Atwood et al., 2020) and have a significant impact on the environment by creating heterogeneity in vegetation structure and small animal abundance (Pringle et al., 2023). Their disappearance has a cascading effect on other living organisms (Wilson, 1987; Terborgh, 1988; Keesing and Young, 2014; Bello et al., 2015), so it is important for the conservation of ecosystems to understand how such species are adapting to the human-influenced environments that have come to dominate in recent centuries (Graham et al., 2009; Ellis et al., 2020).
All terrestrial animals choose places to move to find water, food, mates, and to avoid dangers from predators. Many animals are generally forced to trade-off between the conflicting needs of foraging and avoiding predation. Herbivores perceive spatial variation in predation risk, which is known as the “landscape of fear” (Laundré et al., 2001; Riginos, 2015), and if the risk of predation is too great compared to the benefits of food, they will avoid certain areas or refrain from foraging (Riginos, 2015). This tradeoff between benefits and risks changes depending on the conditions and interacts dynamically (Riginos, 2015; Palmer et al., 2022). Humans are also predators, and their activities create and change new landscapes of fear for wild animals (Gaynor et al., 2019; Mills and Harris, 2020).
The movement of elephants, the largest land animals in the world today, is also affected by human activity (Buij et al., 2007; Granados et al., 2012; Breuer et al., 2016; Wall et al., 2021). They avoid human settlements and traffic roads (Buij et al., 2007; Blake et al., 2008), and their home ranges become smaller (Wall et al., 2021). They are more nocturnal in high-risk areas (Graham et al., 2009; Gaynor et al., 2018; Smit et al., 2023), and move faster with less tortuosity near farmlands (Troup et al., 2020). While there are many studies which show that elephants avoid anthropogenic environments and take risk-avoidance behavior, on the other hand, human–elephant conflicts caused by the overlap of elephant and human habitats are becoming a serious problem in various habitats with various levels of human activity (e.g. Songhurst et al., 2016; Matsuura et al., 2024). In order to mitigate conflicts, it is important to identify the factors in each area, including those that are specific to that area. Clarifying the local land use preferences of elephant populations in small-scale and inferring their behavior there will help to understand the extent to which the area is perceived by them as a risk and/or benefit.
Animal trails are the result of their movements. These trails are formed by terrestrial animals repeatedly passing the same route. As they are used repeatedly, the ground surface becomes compacted and the soil is exposed, creating trails that may remain for many years. Agnew (1966) distinguished two purposes for which animals use trails: to move from one area to another over a relatively long distance and to forage in a relatively small area. These two purposes are not always distinguishable; however according to Agnew (1966), trails for foraging are abundant but not very conspicuous, whereas trails for movement are rare but easy to observe. Each animal trail reflects specific behaviors and is a linear form of physical alteration by animals repeatedly trampling plants and compacting or scraping the ground. Investigating trails is better suited for understanding how elephants use local areas rather than assessing discrete traces such as feces.
The animal trails made by African forest elephants (Loxodonta cyclotis) are large. They can be tens of kilometers long and several meters wide (Blake and Inkamba-Nkulu, 2004). Vanleeuwe and Gautier-Hion (1998) classified elephant trails into three types: “boulevards,” which are straight and wide and extend over long distances; “foraging paths,” which are winding and have no consistent direction; and “clearing alleys,” which are a fine network of trails around open wetland called bai. Clearing alleys are thought to be the result of elephants monitoring the open environment before they enter bai (Vanleeuwe and Gautier-Hion, 1998). Therefore, although they are specific to a particular place, they are the result of risk-avoidance behavior. Boulevards are the largest trails and are mainly used for movement, connecting fruit trees, and water sources (Vanleeuwe and Gautier-Hion, 1998). This corresponds to the purpose of movement from one area to another according to Agnew’s classification. Foraging paths, as the name indicates, correspond to the purpose of foraging, according to Agnew’s classification. When passing through anthropogenic environments, elephants move in a directional manner along clearly defined trails (Von Gerhardt et al., 2014). Fields that are close to the elephant trails are more likely to be raided (Von Gerhardt et al., 2014).
Previous studies on elephant trails have focused on trails with exposed ground surfaces (Blake and Inkamba-Nkulu, 2004; Shannon et al., 2009; Von Gerhardt et al., 2014) and wide trails (Vanleeuwe and Gautier-Hion, 1998; Benitez and Queenborough, 2021). Such wide trails with exposed ground last for many years without disappearing. Elephants use trails other than the exposed and wide ones, but if they cease to use them before the trails become well-established, the trails will disappear. In addition, elephants also create new trails by passing through places that are not trails, but this cannot be discovered by investing only the wide, well-exposed trails. However, there has been limited research on the dynamics of the appearance and disappearance of elephant trails. In reality, trails must go through several stages of elephant use before they become well established. To understand the tendency of whether an elephant will follow an existing trail or create a new trail by passing through a place that is not a trail, it is necessary to clarify how many elephant trails exist and where they are located, not just for the wide, well-established trails, but for all types of trails.
Additionally, previous studies on forest elephant trails have often been conducted in forested habitats that the elephant species originally inhabited and where there is little human influence (Blake and Inkamba-Nkulu, 2004; Inogwabini et al., 2013; Vanleeuwe and Gautier-Hion, 1998). However, in addition to such a ‘pristine’ environments, it is necessary to consider the anthropogenic environments, which currently host a large proportion of elephants. A total of 57.4% of the current distribution of forest elephants is outside of protected areas (Wall et al., 2021). It has been suggested that there is a threshold for human population density or activity level that elephants and humans cannot inhabit (Hoare and Du Toit, 1999; Wall et al., 2021) and it is estimated to be 15 to 20 people/km2 for African savanna elephants (L. africana) in Zimbabwe (Hoare and Du Toit, 1999). The value for forest elephants is probably not very different from this. On the other hand, it is suggested that there is also a tipping point in human land use, and when the population density of a rural area reaches approximately 8 people/km2, the rate of deforestation increases, and the area seems to be converted into permanent farmland (Mayaux et al., 2013; Abernethy et al., 2016). In other words, it is thought that areas with a population density of up to about 8 people/km2, are the key to conservation and coexistence. Therefore, we will focus on two environmental parameters here. One is the vegetation, with a focus on savanna vegetation maintained by human fires. The other is human population density, with a focus on low-density seminatural lands.
Savanna vegetation is defined by the dominance of grass rather than trees. In Gabon, forests intermingle with grassy and shrubby savannas, forming savanna mosaic forests. There, people usually live in villages in the savanna. In some African savanna mosaic forests, including Gabon, burning is important for maintaining savanna vegetation (Jeffery et al., 2014; Gray and Bond, 2015). Forest elephants move between the forest and savanna daily (Mills et al., 2018). Although fire can damage not only the savanna but also the surrounding forests, the presence of elephant trails on the savanna–forest border protects the forest from fire (Cardoso et al., 2019). Thus, the savanna mosaic forest was maintained through interaction with human and elephant activities.
Ellis and Ramankutty (2008) and Ellis et al. (2010, 2020) proposed the concept of an “Anthrome,” which incorporates the effects of human activity into biomes, which are classified based on factors such as climate. Ellis et al. (2020) distinguished seminatural lands from other types of human inhabited areas (urban, settlements, villages) based on a population density of less than 100 people/km2, and defined seminatural lands as “inhabited lands with minor use for permanent agriculture and settlements.” In other words, seminatural lands are areas where wild animals may encounter humans, but human pressure is relatively low. Gabon is home to more than half the forest elephants (Maisels et al., 2013; Thouless et al., 2016; Laguardia et al., 2021), and due to the low population density of 0.9 people/km2 in rural areas (Abernethy et al., 2016), the majority of the land is classified as seminatural lands with a population density of less than 10 people/km2 (Ellis et al., 2010, 2020).
Hence, the objectives of this study were: (1) to quantitatively measure the density and clarify the distribution of elephant trails, including narrow trails and trails with no exposed ground, in a Gabonese savanna mosaic forest; (2) to clarify whether the distribution and structure of elephant trails differ in heterogeneous environments, including savannas and seminatural environments, specifically, (2-a) compare the density and structure of the trails in forest and savanna environments, and (2-b) compare the density and structure of the trails in environments within national park and in environments around village outside national park; and (3) to consider whether a seminatural environment with a low population density is a risky place for elephants. The overall aim of this study was to understand how elephant habitat use patterns inferred from these results reflect the trade-off between foraging benefits and avoidance of human-induced risks.
2 Materials and methods
2.1 Study sites
Fieldwork was conducted in the northeastern part of the Moukalaba-Doudou National Park (MDNP), which is located in the southwest of the Republic of Gabon, and in the village of Doussala, on the opposite bank of the Moukalaba River, which forms the boundary of the MDNP (Figure 1). The annual rainfall in this area ranges from 1,176 to 2,043 mm (Hongo et al., 2018). The dry season is mainly from May to September, and the rainy season is from October to April (Takenoshita et al., 2008).
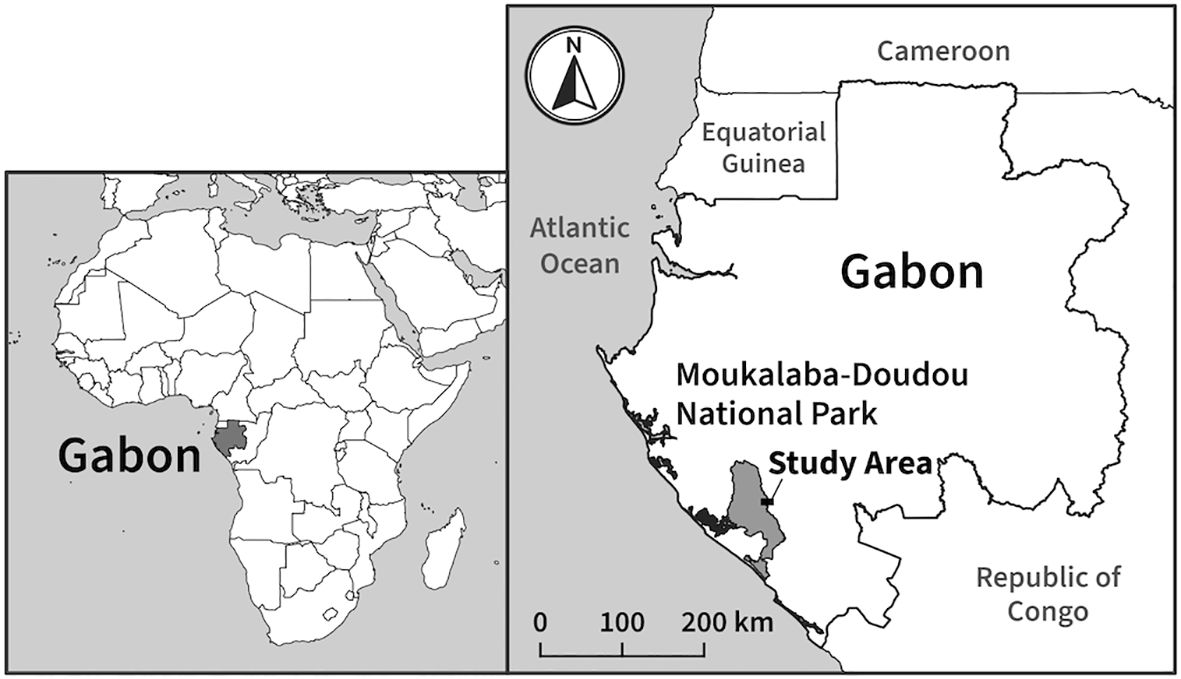
Figure 1. Study area.
The vegetation type in the Moukalaba area is savanna mosaic forest with a mixture of primary forest, secondary forest, riverine forest, mountain forest, and savanna (Ando et al., 2008; Hongo et al., 2018). Patchy savannas in the Moukalaba area are maintained by burning at the end of the dry season every year by the park and local people.
The fauna in this area is also rich in species, such as the western gorilla (Gorilla gorilla), chimpanzee (Pan troglodytes), leopard (Panthera pardus), African buffalo (Syncerus caffer), and waterbuck (Kobus ellipsiprymnus) (Nakashima, 2015). The density of forest elephants in this area is estimated to be 0.387 individuals/km2, which is slightly higher than the average density in Gabon (0.38 individuals/km2) (Laguardia et al., 2021). The only potential predators of elephants in this area are leopards and humans; however, the killing of forest elephants by leopards is extremely rare (Blake, 2004), and the impact of poaching is thought to be minimal because of the difficulty of access and strict control of illegal hunting (Terada et al., 2021).
The village of Doussala had a population of approximately 30–40 people (in 2019, Terada et al., 2021), and is located along an unpaved road in the savanna. The exact population density of this area is unknown; however, considering the population trends (Matsuura and Moussavou, 2015; Terada et al., 2021), it is thought that it does not differ greatly from the report from a neighboring area (0.8 people/km2, Thibault and Blaney, 2003). The surrounding forest is used for shifting cultivation. Elephant raiding of crops is a serious problem, and local people have no choice but to stay in the fields every night to scare them away (Matsuura et al., 2024). People use specific trails across the savanna on a daily basis to travel to and from their village and fields. In this area, livestock such as small goats and chickens are only kept in limited numbers around houses. There are 124 mango (Mangifera indica) trees around the village (Nomoto, unpublished data), and the fruit is an attractive food source for elephants. The elephants cross the Moukalaba River to travel between the national park and the village, and use the area around the village throughout the year. There are currently no permanent residents in the national park.
2.2 Elephant trail survey
Elephant trail data were collected between September and October 2017 in the MDNP and Doussala village. This period corresponds to the end of the dry season and beginning of the rainy season.
2.2.1 Establishment of survey plots
We drew grid lines on the map at 100 m intervals in both the north–south and east–west directions, and divided the survey area into two “area categories”: inside the MDNP (park) and outside the MDNP (village). We then selected several forest and savanna plots from each area category based on the vegetation map. Each plot covered an area of 1 ha. Forest and savanna are referred to as the “vegetation category.” For both categories, we selected plots that could be obtained by walking from the research base. We excluded plots that contained both vegetation types in a single plot, plots that included the Moukalaba River, and plots that contained roads that were sufficiently wide for vehicles to pass through. In addition, we excluded plots that contained crop fields and the trails frequently used by people, including trails to and from the fields. We selected 55 target plots on the map before the survey (Figure 2). The forest plots were mostly secondary forests [83.3%, calculated based on the vegetation map in Hongo et al. (2018)] and included a few riverine forests. Because of the small number of samples, we combined these two forest types into one category. The number of plots for each of the target vegetation categories (forest/savanna) and area categories (park/village) are listed in Table 1.

Figure 2. Vegetation and location of survey plots. The solid black line shows the boundary of MDNP (the western side). Doussala Village is located to the east of MDNP.
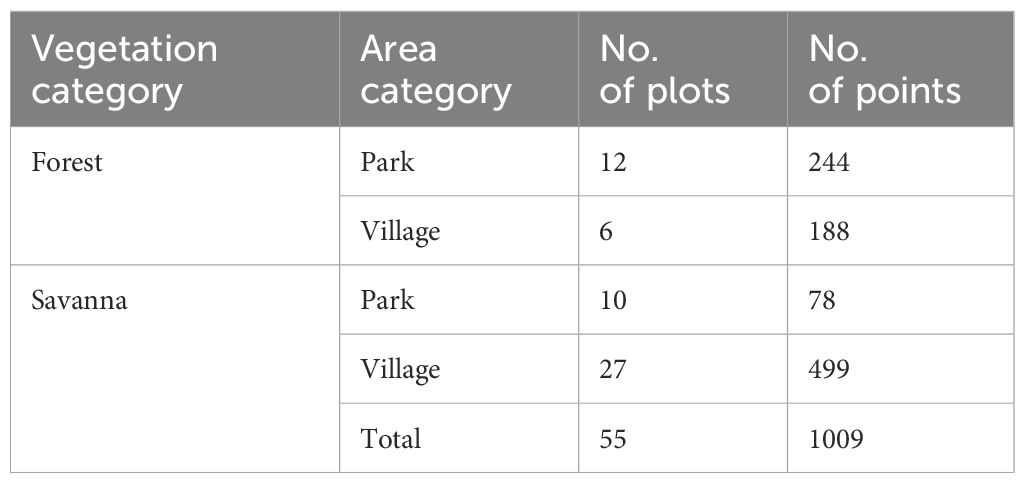
Table 1. Number of survey plots and total measuring points by vegetation and area categories.
2.2.2 Tracing and measuring of the elephant trails within the plots
We defined an animal trail as a linear area in which the ground and vegetation were trampled or uncovered by the passage of animals. Referring to the judgment of the research assistants, who were familiar with the local forest, we recorded the trails that elephants were thought to be able to pass through as elephant trails. They were at least 20 cm wide and with a space of at least 1.5 m high above them. We mapped all elephant trails in the survey plots by recording tracks every two seconds using a global positioning system (GPS) device (Garmin GPSMAP 64s) while walking along the trails. Along the elephant trails, we set up the measuring points every 20 m, which were measured based on the number of steps. One of the two research assistants who had practiced walking with a 1-meter stride in advance measured the steps. At these points, we measured the width of the trail using a non-sagging rigid measuring tape, and recorded the exposure level of the trail (Figure 3). We measured areas with exposed ground and/or bent plants as trail widths in 1 cm increments and then rounded them to the nearest 5 cm to account for measurement error. The exposure level was categorized into four levels (levels 1–4) depending on the degree of soil exposure and plant trampling. Levels 1 and 2 correspond to trails where the ground is not exposed, whereas levels 3 and 4 correspond to trails where the ground is exposed. The locations of the relatively fresh signs of the elephants, such as dung and clear footprints, were also recorded. The number of measuring points within the plot was used as an indicator of the linear density of the elephant trails for the analysis. This was because the accuracy of GPS tracking differs between closed-canopy forests and open savannas, so using linear density was considered inappropriate for comparison. The trail width and exposure level of the trail surface were used as indicators of the structure of the elephant trails.

Figure 3. Example of the exposure level of elephant trails. Trail width was measured in units of 1 cm as the part of the ground that has been trampled or exposed. Exposure level was defined as follows: Level 1 is where the ground is completely covered in plants, and the plants have not been compacted down; Level 2 is where the ground is completely covered in plants, and the plants have been compacted down; Level 3 is where vegetation such as plant sprouts can be seen, but the ground is not completely covered; Level 4 is where there are no plants on the ground at all.
The survey was conducted during a period when the savanna was gradually burned. Because we could not know in advance which areas had been burned, some survey plots were burned and some were not. As burning may have affected the results, we included whether the plot area had been burned in the model when analyzing elephant trail point density.
2.3 Statistical analysis
All analyses were performed using R version 4.3.3 (R Core Team, 2024). We examined whether elephant trail point density (number of measuring points in a survey plot), trail width, and exposure level differed depending on vegetation and area categories.
We modeled the elephant trail point density using a generalized linear model (GLM) with a negative binomial distribution and a log link function (glm.nb function in MASS package, Venables and Ripley, 2002). We included the vegetation category (forest/savanna), area category (park/village), their interaction, and burning (1/0) as explanatory variables. The trail width was modelled using a GLM with a t-distribution and identity link function (glmmTMB function in glmmTMB package, Brooks et al., 2017). We included the vegetation category, area category, and their interaction as explanatory variables. The generalized linear mixed model that includes the plot ID as a mixed effect did not converge. We modeled the exposure level using a cumulative link mixed model (CLMM) (clmm2 function in ordinal package, Christensen, 2023). We included vegetation category, area category, and their interaction as explanatory variables, and plot ID as a mixed effect. For the GLMs, we checked the models for overdispersion/underdispersion and residual distribution (simulateResiduals function in DHARMa package, Hartig, 2022). In addition, we performed Moran’s I test for spatial autocorrelation in the residuals for the GLMs (testSpatialAutocorrelation function in DHARMa package, Hartig, 2022).
In all models, we first tested whether the interaction between the vegetation category and area category had an effect using a likelihood ratio test. If the interaction did not have a significant effect, we adopted a model without the interaction, and then tested the effect of the interaction between the vegetation and area categories using the likelihood ratio test. If the interaction had a significant effect, we then tested a simple main effect (emmeans package, Lenth, 2024) to assess the effect of one factor at each level of the other factor.
3 Results
Elephant trails were found in 53 of the 55 plots surveyed, with 1009 measuring points (Table 1). No elephant trails were found in the two savanna plots in the park.
3.1 Elephant trail point density
The average point density of the elephant trails in the forest plots (both park and village) was 24.0 points per plot (1 ha) (n = 18, SD = 11.4, range: 4–45). Because the points were placed approximately every 20 m, there were, on average, a total of 480 m of elephant trails per plot. Elephant trails in the savanna had an average point density of 15.6 points per plot (n = 37, SD = 9.8, range: 0–38), which means that there were on average a total of 312 m elephant trails per plot.
The interaction between the vegetation category and area category was not significant (χ2 = 0.26, df = 1, p = 0.61). Elephant trail point density differed significantly between vegetation categories (Figure 4A; Table 2) and was higher in forest plots than in savanna plots. The density of elephant trails also differed significantly by area category (Figure 4A; Table 2) and was higher in the village plots than in the park plots. These differences were also significant if we only included measuring points with fresh signs nearby (elephant signs were found between the intersections before and after that point) (vegetation category: χ2 = 20.82, df = 1, p < 0.001; area category: χ2 = 9.10, df = 1, p = 0.003). The marginal means of elephant trail point density calculated from the model were 19.7 (95% confidence interval (CI): 14.9–26.0) points per plot in the park forest, 33.4 (95% CI: 23.8–46.9) points per plot in the village forest, 9.9 (95% CI: 7.1–13.6) points per plot in the park savanna, and 16.8 (95% CI: 13.7–20.5) points per plot in the village savanna (Figure 4A).
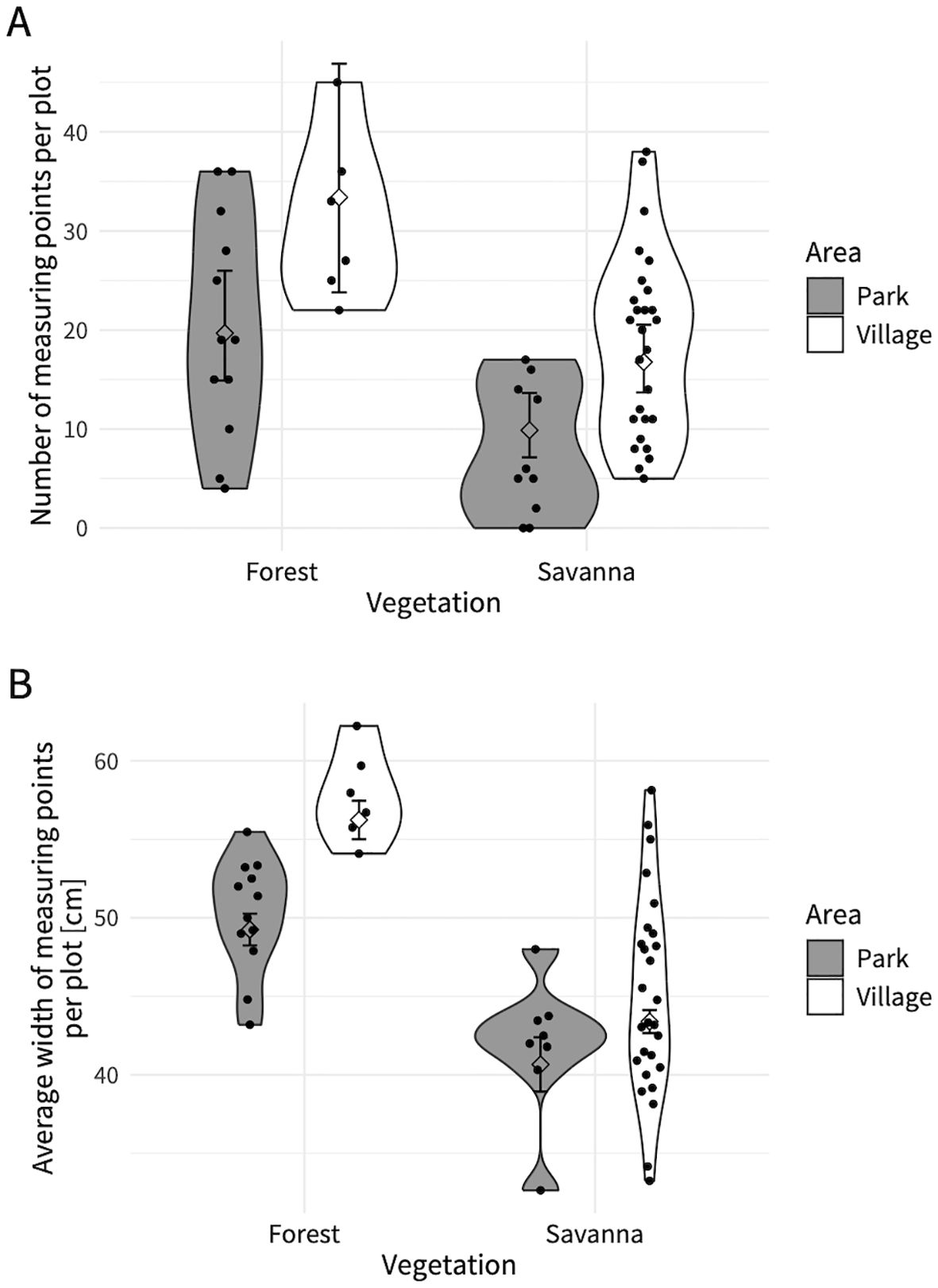
Figure 4. (A) Trail point density and (B) trail width on measuring points in different vegetation and area categories. Each dot shows (A) the number of measuring points on elephant trails in each plot and (B) the average trail width of measuring points per plot. Diamond-shaped dots and black line show the model’s predicted values and it’s 95% confidence interval. Violin plot shows the distribution of (A) trail point density and (B) trail width of each vegetation/area category.
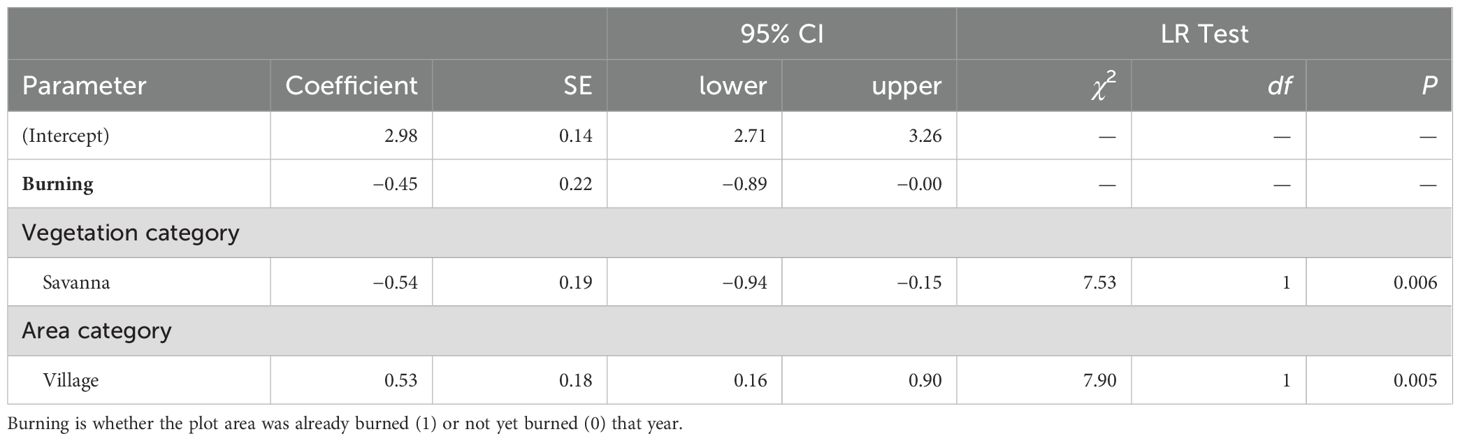
Table 2. Summary of the GLM for estimating elephant trail point density, and results of the likelihood ratio test.
3.2 Elephant trail structure: width
In the forest plots, the average elephant trail width was 53.2 cm (SD = 10.3, range: 30–130). In the savanna, the average width was 43.5 cm (SD = 8.8, range: 20–95). Of the elephant trails measured in the forest, trails with 50 cm width were the most common (31.3%), followed by those with 55 cm width (27.4%). For elephant trails measured in the savanna, trails with 40 cm width accounted for the largest proportion (30.9%), followed by those with 50 cm width (19.8%). If we only look at exposed trails, the average width of forest trails was 57.1 cm (SD = 13.0, range: 35–130) and the average width of savanna trails was 44.6 cm (SD = 8.9, range: 25–95).
The interaction between the vegetation category and area category was significant (χ2 = 11.4, df = 1, p < 0.001, Table 3A). There were significant differences in elephant trail widths between the vegetation categories in both the park and village plots (Figure 4B; Table 3B), with forest trails being wider than savanna trails. There were also differences in trail widths between the area categories in both forests and savannas (Figure 4B; Table 3B), with village trails being wider than park trails. The effect of the area category on the difference in trail width was greater in the forest than in the savanna (Figure 4B; Table 3A). The marginal means of the elephant trail widths calculated from the model were 49.3 cm (95% CI: 48.2–50.3) in the park forest, 56.2 cm (95% CI: 55.0–57.5) in the village forest, 40.7 cm (95% CI: 38.9–42.4) in the park savanna, and 43.4 cm (95% CI: 42.7–44.1) in the village savanna (Figure 4B).
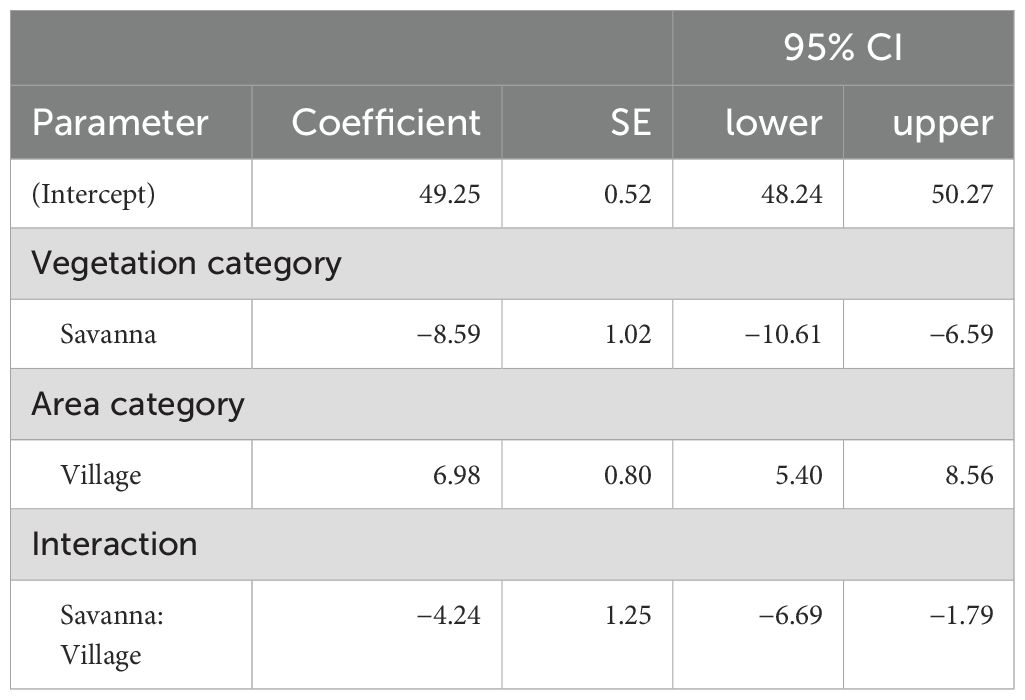
Table 3A. Summary of the GLM for estimating elephant trail width.
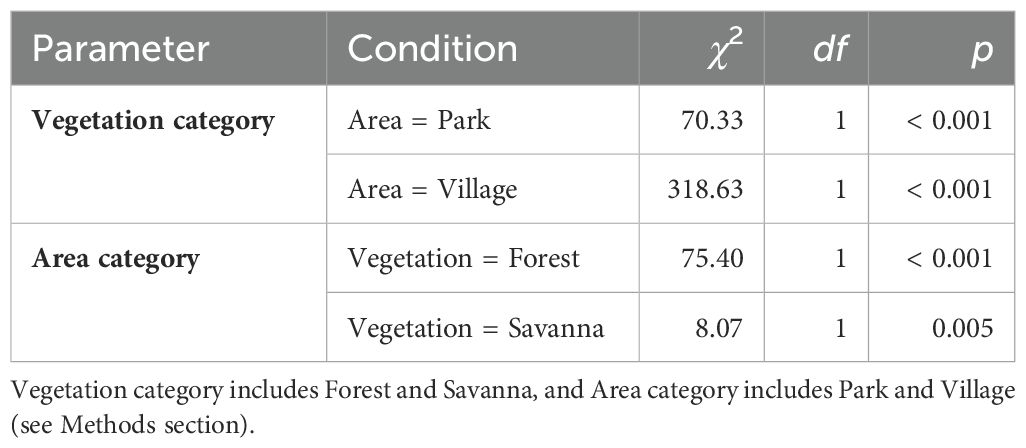
Table 3B. Results of the simple main effect test for fixed effects.
3.3 Elephant trail structure: exposure level
In the forest plots, the proportion of points where the ground was exposed was 31.4%. In contrast, 65.0% of the points were exposed in the savanna. The fitted model predicted that the proportion of level 2 points would be the highest in the forest plots (49.2% in the park forest and 49.8% in the village forest), whereas the proportion of level 3 points would be the highest in the savanna plots (53.1% in the park savanna and 52.0% in the village savanna; Figure 5).
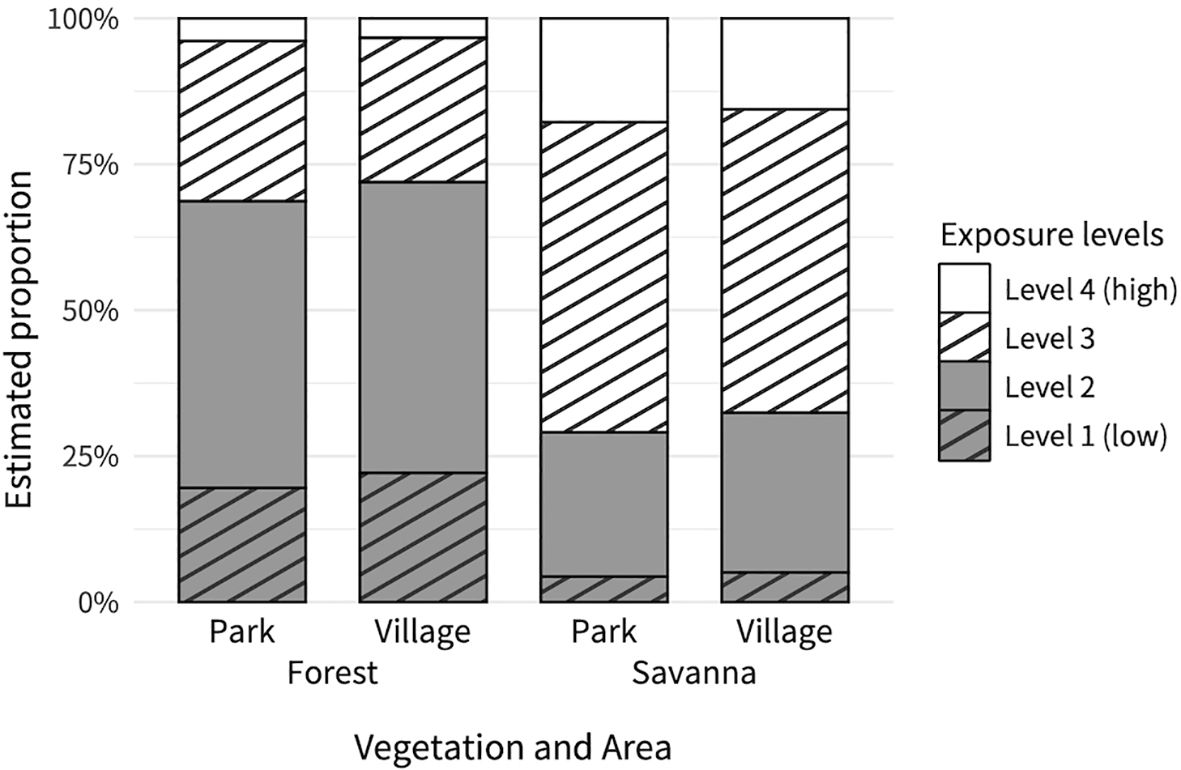
Figure 5. Relationship between exposure level on measuring points, and vegetation and area categories. Each bar shows the cumulative probabilities (proportion) of exposure level (1 to 4) on each vegetation and area category.
The interaction between the vegetation category and area category was not significant (χ2 = 0.20, df = 1, p = 0.66). The exposure level of the elephant trail surface at the measuring points differed depending on the vegetation category (Figure 5; Table 4), with the savanna exhibiting a higher exposure level than the forest. However, there was no difference in exposure levels according to the area category (Figure 5; Table 4).

Table 4. Summary of the CLMM for estimating the proportion of exposure level of elephant trails, and the results of likelihood ratio tests for fixed effects.
4 Discussion
4.1 Density and structure of elephant trails in forest plots
Our survey of elephant trails in Moukalaba found that there were an average of 480 m of elephant trails per plot (1 ha) in the forested areas. This is the first study to quantify the linear density and width of all elephant trails, including those that are not in permanent use. The trails that met the same criteria applied in previous research (i.e., those with exposed ground surface) only accounted for 31.4% of the total or 151 m per plot (31.4% of 480 m). This indicates that the majority of the trails used by the forest elephants were not firmly trampled and/or were not exposed.
The average width of the elephant trails in the forest was 53.2 cm and with 480 m trails per plot. This means that elephant trails cover 255.4 m2 of one hectare plot, or 2.6% of the area. Considering that the distribution of unexposed elephant trails is likely to change seasonally, it is likely that larger areas will be disturbed by elephants over the course of a year. Because large mammals are more likely to be captured by motion-triggered cameras on animal trails than at random locations (Cusack et al., 2015), changes in the distribution of elephant trails should also bring about changes in the locations of these animal species.
This study found that non-exposed trails accounted for a large proportion of elephant trails in forested areas. It was suggested that elephants also used the area outside the well-trodden trails to some extent. These trails were spread out in a network of many intersections. The distribution of well-exposed trails reveals the long-term movement patterns of the entire elephant population (Shannon et al., 2009). The distribution of trails, including non-exposed trails, can reveal the intensity of elephant land use at a certain point in time. In addition, by monitoring the seasonal changes in non-exposed trails, it is possible to understand short-term movement patterns and seasonal habitat use.
4.2 Elephant trails in savanna
This study revealed that there are many elephant trails in savannas, although the density of these trails is lower than that in forests. Forest elephants are generally frugivorous, consuming mainly fruits and leaves (Short, 1981; Tchamba and Seme, 1993; White et al., 1993), and the fruits preferred by elephants are often found in secondary forests (White et al., 1993). As most of the survey plots in this study were secondary forests, it is not surprising that many elephant trails were found in the forest plots. However, the savanna also had an average of 312 m of elephant trails per 1 ha, which was 65% of the trails in the forest. This may be related to the frequent use of savanna by forest elephants during the short dry season (Mills et al., 2018). The dominant fruit tree species in the savanna of Moukalaba was Nauclea latifolia; however, it was not ripe at that time. Psidium guineensis also grows in the savanna; however, its seeds have been found in very small numbers in elephant dungs collected during the same months of another year (Nomoto, unpublished data). The elephants probably came to the savanna not only to search for fruit trees but also to feed on grass shoots.
The proportion of exposed elephant trails was higher in the savanna than in the forest. The density of the exposed trails in the savanna was 203 m/ha, which was higher than that of the forest (151 m/ha). Several factors might have contributed to these differences. One possibility may be differences in the frequency and method of elephant use (whether they use existing trails or explore new routes), and another may be differences in the recovery speed of vegetation. Generally it takes more traffic to create a visible animal trail in a savanna than in a forest (Agnew, 1966). Therefore, elephants in this area tend to follow existing trails in the savanna. The fact that the width of the trails in the savanna was narrower than that in the forest also indicates that the elephants followed trails more faithfully in the savanna than in the forest. Additionally, elephant trails in the savanna are relatively straight (Nomoto, personal observation). Thus, elephant trails in savanna may be used mainly for passing through to other destinations, and the movement that is less tortuous and more faithful to the trails (Von Gerhardt et al., 2014; Troup et al., 2020) seems to reflect an awareness of the risks of open vegetation. In this study, no data were collected regarding the actual frequency of elephant use. Therefore, future studies need to compare the results with data from camera traps, as well as to compare them with data from various vegetation and topographical areas.
4.3 Elephant trails in the village
Compared with the elephant trails in the park, those around the village were denser and wider, but there was no significant difference in the exposure level of the trail surfaces. The values predicted by the model showed that trails around the village were approximately 7 cm wider than those in the park. This is thought to reflect differences in the frequency of use by elephants, at least to a certain extent. The village area is likely attractive to elephants owing to the presence of crop fields and mango trees (Ngama et al., 2019). Forest elephants avoid areas of intense industrial human activity (Barnes et al., 1991; Buij et al., 2007), and prefer secondary vegetation in abandoned villages and plantations (Barnes et al., 1991). The village covered by this study has a small population, and elephant poaching is relatively rare in this area (Terada et al., 2021); therefore the attractiveness of the available food may be greater than the risk of being threatened by humans. Consequently, it is possible that elephants frequently use areas around village. This is consistent with a report that elephant sounds were heard on 77.9% of the nights villagers stayed in the crop fields (Matsuura et al., 2024). To prevent crop damage, the local people stay overnight in the fields to scare them away (Terada et al., 2021; Matsuura et al., 2024). Even if they are chased away from the fields, they may be using the secondary vegetation in the forest outside without feeling much risk. Doussala Village is located on the bank of the Moukalaba River and is close to a water source. The survey period included the end of the dry season; therefore, the water from the river is likely to be valuable for elephants at this time of the year (Beirne et al., 2021).
This study showed that elephants frequently use seminatural lands outside the protected area, and that in such places, the pattern of vegetation disturbance is caused by a combination of human activity patterns and elephant land use patterns. In places where elephants and humans share the same habitat, it is believed that a unique ecosystem, different from a purely natural environment, is formed. Shifting cultivation is usually carried out over a cycle of approximately 1–2 years, changing the location of the fields. For savanna elephants, the area of elephant trails is expected to decrease by 79% when artificial waterholes are removed (Shannon et al., 2009). This means that if the place loses its attractiveness, the elephants will stop visiting and the trails will disappear. In the case of forest elephants, it is possible that changes in the distribution of elephant trails will occur as the location of the field shifts. As human activity continues to expand, human habitats are expected to occupy even larger areas in the future. The collection of ecological data in such anthropogenic areas will hopefully elucidate the local relationships between elephant and human land use and its transitions, and provide important insights into elephant–human coexistence.
4.4 Elephant trails as an indicator of the intensity of elephant land use
There has been a lot of research using GPS telemetry to study elephant migration patterns (Blake et al., 2008; Loarie et al., 2009; Mills et al., 2018). The advantage of elephant trail surveys is their relatively low cost compared to more expensive GPS telemetry surveys (Weinbaum et al., 2007). Additionally, GPS telemetry surveys can only collect data on individuals wearing the device, and the number of individuals that can be tracked is limited. In contrast, elephant trails are the result of the behavior of all individuals who use the area. Therefore, it is possible to clarify the environment-use patterns of the entire population by investigating trails (Newmark and Rickart, 2012), although it is difficult to determine the patterns for each individual. This information is useful for the quick and easy assessment of the intensity of elephant use in particular areas. These two methods are not mutually exclusive, and by combining them, it is possible to clarify the land use patterns of elephants in the area in more detail.
5 Conclusion
Elephant trails with various structures have been found not only in forests but also in anthropogenic areas, such as savannas and seminatural lands outside protected areas. Furthermore, in area where human–elephant conflict is serious, it was found that elephant trails were more densely distributed outside the protected area than within the protected area. The micro-level structure of the African savanna mosaic forests is strongly affected by both elephants and humans, and this is particularly noticeable in seminatural environment with low populations outside the protected area. Our study revealed that the majority of the trails used by forest elephants were not well-established and without exposed ground. Although this survey was conducted only during a specific season, from the end of the dry season to the beginning of the rainy season, we were able to clarify the land use by elephant populations, including the heterogeneity of such use on a fine scale. By tracking changes in different seasons and years, it will be possible to clarify elephants land use and impact on their environment in more detail. In order to mitigate human–elephant conflicts in long-term, it is necessary to consider the needs of both humans and elephants (Mumby and Plotnik, 2018). Assessing land use based on the development of elephant trails will be the first step toward understanding their needs.
Data availability statement
The raw data supporting the conclusions of this article will be made available by the authors, without undue reservation.
Ethics statement
Ethical approval was not required for the study involving animals in accordance with the local legislation and institutional requirements because this study is about animal traces and does not involve direct investigation of animals.
Author contributions
MN: Conceptualization, Data curation, Formal analysis, Funding acquisition, Investigation, Methodology, Project administration, Visualization, Writing – original draft, Writing – review & editing. EA-O: Writing – review & editing.
Funding
The author(s) declare that financial support was received for the research and/or publication of this article. This research was funded by the Leading Graduate Program in Primatology and Wildlife Science of Kyoto University (U04) for field research and crowdfunding on the academic crowdfunding platform "academist" (https://academist-cf.com/projects/352?lang=en) for publication.
Acknowledgments
We wish to thank the Centre National de la Recherche Scientifique et Technologique and Agence Nationale des Parcs Nationaux in Gabon for permission to conduct this research. We are most grateful to Prof. Alfred Ngomanda, Dr. Guy-Max Moussavou, Prof. Yuji Takenoshita, Dr. Naoki Matsuura, Ms. Keiko Tsubokawa, Dr. Mari Terakawa, members of the Institut de Recherche en Ecologie Tropicale, and the research group at the Moukalaba-Doudou National Park for support and advice on our fieldwork. This research owes much to the support and constructive advice on the fieldwork and statistical analysis from Dr. Shun Hongo. Mr. Jean-Pierre Doukaga and Mr. Rodrigue Nzaou Mapangou assisted us in collecting the data. We wish to thank Prof. Naofumi Nakagawa, Dr. Michio Nakamura, Dr. Lin Wang and five anonymous reviewers for constructive comments on the manuscript. We are indebted to Prof. Takakazu Yumoto, Prof. Tomoaki Nishihara, Dr. Kaori Mizuno, Dr. Masaya Tamura, Mr. Tsubasa Yamaguchi and colleagues in the Laboratory of Human Evolution Studies at Kyoto University for valuable comments on this study. We also thank the local people in Doussala for helping us in the field.
Conflict of interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Generative AI statement
The author(s) declare that no Generative AI was used in the creation of this manuscript.
Publisher’s note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
References
Abernethy K., Maisels F., White L. J. T. (2016). Environmental issues in Central Africa. Annu. Rev. Environ. Resour. 41, 1–33. doi: 10.1146/annurev-environ-110615-085415
Agnew A. D. Q. (1966). The use of game trails as a possible measure of habitat utilisation by larger mammals. East Afr. Wildl. J. 4, 38–46. doi: 10.1111/j.1365-2028.1966.tb00879.x
Ando C., Iwata Y., Yamagiwa J. (2008). Progress of habituation of western lowland gorillas and their reaction to observers in Moukalaba-Doudou National Park, Gabon. Afr. Study Monogr. Supplement. Issue. 39, 55–69. doi: 10.14989/66238
Atwood T. B., Valentine S. A., Hammill E., McCauley D. J., Madin E. M. P., Beard K. H., et al. (2020). Herbivores at the highest risk of extinction among mammals, birds, and reptiles. Sci. Adv. 6, eabb8458. doi: 10.1126/sciadv.abb8458
Barnes R. F. W., Barnes K. L., Alers M. P. T., Blom A. (1991). Man determines the distribution of elephants in the rain forests of northeastern Gabon. Afr. J. Ecol. 29, 54–63. doi: 10.1111/j.1365-2028.1991.tb00820.x
Beirne C., Houslay T. M., Morkel P., Clark C. J., Fay M., Okouyi J., et al. (2021). African forest elephant movements depend on time scale and individual behavior. Sci. Rep. 11, 12634. doi: 10.1038/s41598-021-91627-z
Bello C., Galetti M., Pizo M. A., Magnago L. F. S., Rocha M. F., Lima R. A. F., et al. (2015). Defaunation affects carbon storage in tropical forests. Sci. Adv. 1, e1501105. doi: 10.1126/sciadv.1501105
Benitez L., Queenborough S. A. (2021). Fruit trees drive small-scale movement of elephants in Kibale National Park, Uganda. Biotropica 53, 1620–1630. doi: 10.1111/btp.13010
Blake S. (2004). Do leopards kill forest elephants? Evidence from northern Congo. Mammalia 68, 225–228. doi: 10.1515/mamm.2004.021
Blake S., Deem S. L., Strindberg S., Maisels F., Momont L., Isia I.-B., et al. (2008). Roadless wilderness area determines forest elephant movements in the Congo Basin. PloS One 3, e3546. doi: 10.1371/journal.pone.0003546
Blake S., Inkamba-Nkulu C. (2004). Fruit, minerals, and forest elephant trails: Do all roads lead to Rome? Biotropica 36, 392–401. doi: 10.1111/j.1744-7429.2004.tb00332.x
Bogoni J. A., Ferraz K. M. P. M. B., Peres C. A. (2022). Continental-scale local extinctions in mammal assemblages are synergistically induced by habitat loss and hunting pressure. Biol. Conserv. 272, 109635. doi: 10.1016/j.biocon.2022.109635
Breuer T., Maisels F., Fishlock V. (2016). The consequences of poaching and anthropogenic change for forest elephants. Conserv. Biol. 30, 1019–1026. doi: 10.1111/cobi.12679
Brook B. W., Sodhi N. S., Bradshaw C. J. A. (2008). Synergies among extinction drivers under global change. Trends Ecol. Evol. 23, 453–460. doi: 10.1016/j.tree.2008.03.011
Brooks M. E., Kristensen K., van Benthem K. J., Magnusson A., Berg C. W., Nielsen A., et al. (2017). glmmTMB balances speed and flexibility among packages for zero-inflated generalized linear mixed modeling. R J. 9, 378–400. doi: 10.32614/RJ-2017-066
Buij R., McShea W. J., Campbell P., Lee M. E., Dallmeier F., Guimondou S., et al. (2007). Patchoccupancy models indicate human activity as major determinant of forest elephant Loxodonta cyclotis seasonal distribution in an industrial corridor in Gabon. Biol. Conserv. 135, 189–201. doi: 10.1016/j.biocon.2006.10.028
Cardillo M., Mace G. M., Jones K. E., Bielby J., Bininda-Emonds O. R. P., Sechrest W., et al. (2005). Multiple causes of high extinction risk in large mammal species. Science 309, 1239–1241. doi: 10.1126/science.1116030
Cardoso A. W., Malhi Y., Oliveras I., Lehmann D., Ndong J. E., Dimoto E., et al. (2019). The role of forest elephants in shaping tropical forest–Savanna coexistence. Ecosystems 23, 602–616. doi: 10.1007/s10021-019-00424-3
Ceballos G., Ehrlich P. R., Barnosky A. D., García A., Pringle R. M., Palmer T. M. (2015). Accelerated modern human-induced species losses: Entering the sixth mass extinction. Sci. Adv. 1, e1400253. doi: 10.1126/sciadv.1400253
Christensen R. H. B. (2023). ordinal—Regression Models for Ordinal Data. R package version 2023.12-4.1. Available online at: https://CRAN.R-project.org/package=ordinal.
Cusack J. J., Dickman A. J., Rowcliffe J. M., Carbone C., Macdonald D. W., Coulson T. (2015). Random versus game trail-based camera trap placement strategy for monitoring terrestrial mammal communities. PloS One 10, e0126373. doi: 10.1371/journal.pone.0126373
Dirzo R., Young H. S., Galetti M., Ceballos G., Isaac N. J. B., Collen B. (2014). Defaunation in the anthropocene. Science 345, 401–406. doi: 10.1126/science.1251817
Ellis E. C., Beusen A. H. W., Goldewijk K. K. (2020). Anthropogenic biomes: 10,000 BCE to 2015 CE. Land 9, 129. doi: 10.3390/land9050129
Ellis E. C., Klein Goldewijk K., Siebert S., Lightman D., Ramankutty N. (2010). Anthropogenic transformation of the biomes 1700 to 2000. Global Ecol. Biogeogr. 9, 589–606. doi: 10.1111/j.1466-8238.2010.00540.x
Ellis E. C., Ramankutty N. (2008). Putting people in the map: anthropogenic biomes of the world. Front. Ecol. Environ. 6, 439–447. doi: 10.1890/070062
Gaynor K. M., Branco P. S., Long R. A., Gonçalves D. D., Granli P. K., Poole J. H. (2018). Effects of human settlement and roads on diel activity patterns of elephants (Loxodonta africana). Afr. J. Ecol. 56, 872–881. doi: 10.1111/aje.12552
Gaynor K. M., Brown J. S., Middleton A. D., Power M. E., Brashares J. S. (2019). Landscapes of fear: Spatial patterns of risk perception and response. Trends Ecol. Evol. 34, 355–368. doi: 10.1016/j.tree.2019.01.004
Graham M. D., Douglas-Hamilton I., Adams W. M., Lee P. C. (2009). The movement of African elephants in a human-dominated land-use mosaic. Anim. Conserv. 12, 445–455. doi: 10.1111/j.1469-1795.2009.00272.x
Granados A., Weladji R. B., Loomis M. R. (2012). Movement and occurrence of two elephant herds in a human-dominated landscape, the Bénoué Wildlife Conservation Area, Cameroon. Trop. Conserv. Sci. 5, 150–162. doi: 10.1177/194008291200500205
Gray E. F., Bond W. J. (2015). Soil nutrients in an African forest/savanna mosaic: Drivers or driven? South Afr. J. Bot. 101, 66–72. doi: 10.1016/j.sajb.2015.06.003
Haddad N. M., Brudvig L. A., Clobert J., Davies K. F., Gonzalez A., Holt R. D., et al. (2015). Habitat fragmentation and its lasting impact on Earth’s ecosystems. Sci. Adv. 1, e1500052. doi: 10.1126/sciadv.1500052
Hartig F. (2022). DHARMa: Residual Diagnostics for Hierarchical (Multi-Level/Mixed) Regression Models. R package version 0.4.7. Available online at: https://CRAN.R-project.org/package=DHARMa.
Hoare R. E., Du Toit J. T. (1999). Coexistence between people and elephants in African savannas. Conserv. Biol. 13, 633–639. doi: 10.1046/j.1523-1739.1999.98035.x
Hongo S., Nakashima Y., Akomo-Okoue E. F., Mindonga-Nguelet F. L. (2018). Seasonal change in diet and habitat use in wild mandrills (Mandrillus sphinx). Int. J. Primatol. 39, 27–48. doi: 10.1007/s10764-017-0007-5
Inogwabini B.-I., Ngama-Nkosi M., Wema-Wema L., Longwango M. (2013). Elephant effect on forest physical structure and plant species composition in Salonga and Malebo (Lac Tumba landscape), Democratic Republic of Congo. Pachyderm 53, 28–37. doi: 10.69649/pachyderm.v53i.321
Jeffery K. J., Korte L., Palla F., Walters G. M., White L., Abernethy K. (2014). Fire management in a changing landscape: a case study from Lopé National Park, Gabon. Parks. Int. J. Protect. Areas Conserv. 20, 39–52. doi: 10.2305/IUCN.CH.2014.PARKS-20-1.KJJ.en
Keesing F., Young T. P. (2014). Cascading consequences of the loss of large mammals in an African Savanna. Bioscience 64, 487–495. doi: 10.1093/biosci/biu059
Laguardia A., Bourgeois S., Strindberg S., Gobush K. S., Abitsi G., Bikang Bi Ateme H. G., et al. (2021). Nationwide abundance and distribution of African forest elephants across Gabon using non-invasive SNP genotyping. Global Ecol. Conserv. 32, e0189. doi: 10.1016/j.gecco.2021.e01894
Laundré J. W., Hernández L., Altendorf K. B. (2001). Wolves, elk, and bison: reestablishing the “landscape of fear” in Yellowstone National Park, U.S.A. Can. J. Zool. 79, 1401–1409. doi: 10.1139/z01-094
Lenth R. V. (2024). emmeans: Estimated Marginal Means, aka Least-Squares Means. R package version 1.10.7. Available online at: https://CRAN.R-project.org/package=emmeans.
Loarie S. R., van Aarde R. J., Pimm S. L. (2009). Elephant seasonal vegetation preferences across dry and wet savannas. Biol. Conserv. 142, 3099–3107. doi: 10.1016/j.biocon.2009.08.021
Maisels F., Strindberg S., Blake S., Wittemyer G., Hart J., Williamson E. A., et al. (2013). Devastating decline of forest elephants in Central Africa. PloS One 8, e59469. doi: 10.1371/journal.pone.0059469
Matsuura N., Moussavou G.-M. (2015). Analysis of local livelihoods around Moukalaba-Doudou National Park in Gabon. Tropics 23, 195–204. doi: 10.3759/tropics.23.195
Matsuura N., Nomoto M., Terada S., Yobo C. M., Memiaghe H. R., Moussavou G. M. (2024). Human-elephant conflict in the African rainforest landscape: crop-raiding situations and damage mitigation strategies in rural Gabon. Front. Conserv. Sci. 5. doi: 10.3389/fcosc.2024.1356174
Mayaux P., Pekel J.-F., Desclée B., Donnay F., Lupi A., Achard F., et al. (2013). State and evolution of the African rainforests between 1990 and 2010. Philos. Trans. R. Soc. Lond. B: Biol. Sci. 368, 20120300. doi: 10.1098/rstb.2012.0300
Mills K. L., Harris N. C. (2020). Humans disrupt access to prey for large African carnivores. eLife 9, e60690. doi: 10.7554/eLife.60690
Mills E. C., Poulsen J. R., Fay J. M., Morkel P., Clark C. J., Meier A., et al. (2018). Forest elephant movement and habitat use in a tropical forest-grassland mosaic in Gabon. PloS One 13, e0199387. doi: 10.1371/journal.pone.0199387
Mumby H. S., Plotnik J. M. (2018). Taking the elephants’ perspective: Remembering elephant behavior, cognition and ecology in human-elephant conflict mitigation. Front. Ecol. Evol. 6. doi: 10.3389/fevo.2018.00122/full
Nakashima Y. (2015). Inventorying medium- and large-sized mammals in the African lowland rainforest using camera trapping. Tropics 23, 151–164. doi: 10.3759/tropics.23.151
Newmark W. D., Rickart E. A. (2012). High-use movement pathways and habitat selection by ungulates. Mamm. Biol. = Z. Fur Saugetierkunde 77, 293–298. doi: 10.1016/j.mambio.2012.03.007
Ngama S., Bindelle J., Poulsen J. R., Hornick J.-L., Linden A., Korte L., et al. (2019). Do topography and fruit presence influence occurrence and intensity of crop-raiding by forest elephants (Loxodonta africana cyclotis)? PloS One 14, e0213971. doi: 10.1371/journal.pone.0213971
Palmer M. S., Gaynor K. M., Becker J. A., Abraham J. O., Mumma M. A., Pringle R. M. (2022). Dynamic landscapes of fear: understanding spatiotemporal risk. Trends Ecol. Evol. 37, 911–925. doi: 10.1016/j.tree.2022.06.007
Pringle R. M., Abraham J. O., Anderson T. M., Coverdale T. C., Davies A. B., Dutton C. L., et al. (2023). Impacts of large herbivores on terrestrial ecosystems. Curr. Biol. 33, R584–R610. doi: 10.1016/j.cub.2023.04.024
R Core Team (2024). R: A Language and Environment for Statistical Computing (Vienna, Austria: R Foundation for Statistical Computing). Available online at: https://www.R-project.org/.
Riginos C. (2015). Climate and the landscape of fear in an African savanna. J. Anim. Ecol. 84, 124–133. doi: 10.1111/1365-2656.12262
Ripple W. J., Newsome T. M., Wolf C., Dirzo R., Everatt K. T., Galetti M., et al. (2015). Collapse of the world’s largest herbivores. Sci. Adv. 1, e1400103. doi: 10.1126/sciadv.1400103
Shannon G., Matthews W. S., Page B. R., Parker G. E., Smith R. J. (2009). The affects of artificial water availability on large herbivore ranging patterns in savanna habitats: a new approach based on modelling elephant path distributions. Diversity Distrib. 15, 776–783. doi: 10.1111/j.1472-4642.2009.00581.x
Short J. (1981). Diet and feeding behaviour of the forest elephant. Mammalia 45, 177–186. doi: 10.1515/mamm.1981.45.2.177
Smit J. B., Searle C. E., Buchanan-Smith H. M., Strampelli P., Mkuburo L., Kakengi V. A., et al. (2023). Anthropogenic risk increases night-time activities and associations in African elephants (Loxodonta africana) in the Ruaha-Rungwa ecosystem, Tanzania. Afr. J. Ecol. 61, 64–76. doi: 10.1111/aje.13083
Songhurst A., McCulloch G., Coulson T. (2016). Finding pathways to human–elephant coexistence: a risky business. Oryx 50, 713–720. doi: 10.1017/s0030605315000344
Takenoshita Y., Ando C., Iwata Y., Yamagiwa J. (2008). Fruit phenology of the great ape habitat in the Moukalaba-Doudou National Park, Gabon. Afr. Study Monogr. Supplement. Issue. 39, 23–39. doi: 10.14989/66240
Tchamba M. N., Seme P. M. (1993). Diet and feeding behaviour of the forest elephant in the Santchou Reserve, Cameroon. Afr. J. Ecol. 31, 165–171. doi: 10.1111/j.1365-2028.1993.tb00529.x
Terada S., Yobo C. M., Moussavou G.-M., Matsuura N. (2021). Human-elephant conflict around Moukalaba-Doudou National Park in Gabon: socioeconomic changes and effects of conservation projects on local tolerance. Trop. Conserv. Sci. 14, 1–16. doi: 10.1177/19400829211026775
Terborgh J. (1988). The big things that run the world–A sequel to EO Wilson. Conserv. Biol. 2, 402–403. doi: 10.1111/j.1523-1739.1988.tb00207.x
Thibault M., Blaney S. (2003). The oil industry as an underlying factor in the bushmeat crisis in Central Africa. Conserv. Biol. 17, 1807–1813. doi: 10.1111/j.1523-1739.2003.00159.x
Thouless C., Dublin H. T., Blanc J., Skinner D. P., Daniel T. E., Taylor R., et al. (2016). African elephant status report 2016: an update from the African Elephant Database. Occasional Paper Series of the IUCN Species Survival Commission, No. 60 IUCN / SSC Africa Elephant Specialist Group. IUCN, Gland, Switzerland.
Troup G., Doran B., Au J., King L. E., Douglas-Hamilton I., Heinsohn R. (2020). Movement tortuosity and speed reveal the trade-offs of crop raiding for African elephants. Anim. Behav. 168, 97–108. doi: 10.1016/j.anbehav.2020.08.009
Vanleeuwe H., Gautier-Hion A. (1998). Forest elephant paths and movements at the Odzala National Park, Congo: the role of clearings and Marantaceae forests. Afr. J. Ecol. 36, 174–182. doi: 10.1046/j.1365-2028.1998.00123.x
Venables W. N., Ripley B. D. (2002). Modern Applied Statistics with S. 4th ed. (New York: Springer).
Von Gerhardt K., Van Niekerk A., Kidd M., Samways M., Hanks J. (2014). The role of elephant Loxodonta africana pathways as a spatial variable in crop-raiding location. Oryx 48, 436–444. doi: 10.1017/s003060531200138x
Wall J., Wittemyer G., Klinkenberg B., LeMay V., Blake S., Strindberg S., et al. (2021). Human footprint and protected areas shape elephant range across Africa. Curr. Biol. 31, 2437–2445. doi: 10.1016/j.cub.2021.03.042
Weinbaum K., Nzooh Z., Usongo L., Laituri M. (2007). Preliminary survey of forest elephant crossings in Sangha Trinational Park, Central Africa. Pachyderm 43, 52–62. doi: 10.69649/pachyderm.v43i.127
White L. J. T., Tutin C. E. G., Fernandez M. (1993). Group composition and diet of forest elephants, Loxodonta africana cyclotis Matschie 1900, in the Lopé Reserve, Gabon. Afr. J. Ecol. 31, 181–199. doi: 10.1111/j.1365-2028.1993.tb00532.x
Keywords: African forest elephant (Loxodonta cyclotis), animal trails, savanna-mosaic forest, seminatural lands, human-elephant conflict, land use pattern
Citation: Nomoto M and Akomo-Okoue EF (2025) Distribution and structure of African forest elephant trails in anthropogenic savanna-mosaic forest, Gabon. Front. Conserv. Sci. 6:1525065. doi: 10.3389/fcosc.2025.1525065
Received: 11 November 2024; Accepted: 24 February 2025;
Published: 25 April 2025.
Edited by:
Lin Wang, Chinese Academy of Sciences (CAS), ChinaReviewed by:
Dipanjan Naha, Cheetah Conservation Fund, NamibiaMingxia Zhang, Guangxi Normal University, China
Copyright © 2025 Nomoto and Akomo-Okoue. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Mayuko Nomoto, bm9tb3RvLnRnYWxiZkBnbWFpbC5jb20=