 Jimena Samper-Villarreal1,2*
Jimena Samper-Villarreal1,2* Joel C. Creed1,3
Joel C. Creed1,3 Duarte Frade1,4
Duarte Frade1,4 Bradley T. Furman1,5
Bradley T. Furman1,5 Salomão Bandeira1,6
Salomão Bandeira1,6 Hacen M. El-Hacen1,7A. Carolina Peralta1,8
Hacen M. El-Hacen1,7A. Carolina Peralta1,8 Ester A. Serrão1,4Beatriz E. Vera Vegas1,9
Ester A. Serrão1,4Beatriz E. Vera Vegas1,9 Brigitta I. van Tussenbroek1,10*
Brigitta I. van Tussenbroek1,10*- 1Seagrass Species Specialist Group (SSSG) International Union for Conservation of Nature (IUCN), Species Survival Commission (SSC), Gland, Switzerland
- 2Centro de Investigación en Ciencias del Mar y Limnología (CIMAR), Universidad de Costa Rica, San José, Costa Rica
- 3Departamento de Ecologia, Universidade do Estado do Rio de Janeiro, Rio de Janeiro, Brazil
- 4Centro de Ciências do Mar do Algarve, Universidade do Algarve, Faro, Portugal
- 5Florida Fish and Wildlife Research Institute, Florida Fish and Wildlife Conservation Commission, St Petersburg, FL, United States
- 6Departamento de Ciências Biológicas, Universidade Eduardo Mondlane, Maputo, Mozambique
- 7Parc National du Banc d’Arguin (PNBA), Mauritania & BirdEyes, Centre for Global Ecological Change, University of Groningen, Groningen, Netherlands
- 8Institute for Marine Remote Sensing, College of Marine Science, University of South Florida, St. Petersburg, FL, United States
- 9Centro de Botánica Tropical, Instituto de Biología Experimental, Facultad de Ciencias, Universidad Central de Venezuela, Caracas, Venezuela
- 10Reef Systems Unit, Instituto de Ciencias del Mar y Limnología, Universidad Nacional Autónoma de México, Puerto Morelos, Mexico
Seagrasses can form vast meadows in coastal areas and provide valuable ecosystem services. Despite their importance, seagrasses are threatened, and their spatial extent and ecological condition have declined worldwide. Globally, there are six seagrass bioregions based on ocean basin and species distribution. The Tropical Atlantic Bioregion encompasses seagrasses in the warm waters off the Atlantic American continent, the eastern Pacific and the west coast of Africa. Here, the extinction risk of the species was determined through the Categories and Criteria of the IUCN Red List of Threatened Species. The extinction risk for the 15 native seagrass species in the bioregion was assessed and summarized as an update to the previous IUCN effort, conducted 15 years ago. The updated regional assessments were based on expert opinion and compilation of revised data on species´ distribution, population status and trend. Of the 15 native species, nine were endemic to the bioregion; thus, the regional assessment was also a global one. Two other species were typical for the region but also occurred beyond bioregional boundaries (Halodule beaudettei and H. wrightii). The remaining species were either distributed circumglobally (2 species), or consisted of peripheral populations of species characteristic of neighboring bioregions (2 species). The detailed information of 15 assessments have been made freely accessible in an open repository. Nine species maintained their previous risk of extinction status, while one species (Thalassia testudinum) changed from the Least Concern to the Near Threatened category. Downgrading of the status of T. testudinum is worrying, as this is the dominant climax species in the wider Caribbean. Its replacement by less resistant and smaller but faster growing species, such as Syringodium filiforme and H. wrightii (both classified as Least Concern), could compromise the ecosystem services of seagrasses in this bioregion. An additional two species had been recently introduced, and are also briefly covered. The assessments included past and present taxonomical uncertainties of various assessed species. Issues concerning the current boundaries of the bioregions themselves were also encountered. To address this and better capture future and ongoing range expansion or reduction associated with climate change, we propose overlapping transitional boundaries with neighboring seagrass bioregions.
1 Introduction
Seagrasses form extensive meadows in the tropical Atlantic (Tropical Atlantic Bioregion, or Bioregion 2; Short et al., 2007; Figure 1). They have an essential ecological role as primary producers, in the provision of forage, nursery habitats and biodiversity, sediment stabilization, habitat connectivity within the larger seascape, etc (Hemminga and Duarte, 2000). In addition to their ecological importance, seagrasses also provide important ecosystem services to humans. These encompass enhanced water quality, carbon sequestration, coastal stabilization and erosion prevention, increased secondary production, and significant cultural importance (Nordlund et al., 2016; De los Santos et al., 2020; Creed et al., 2023).
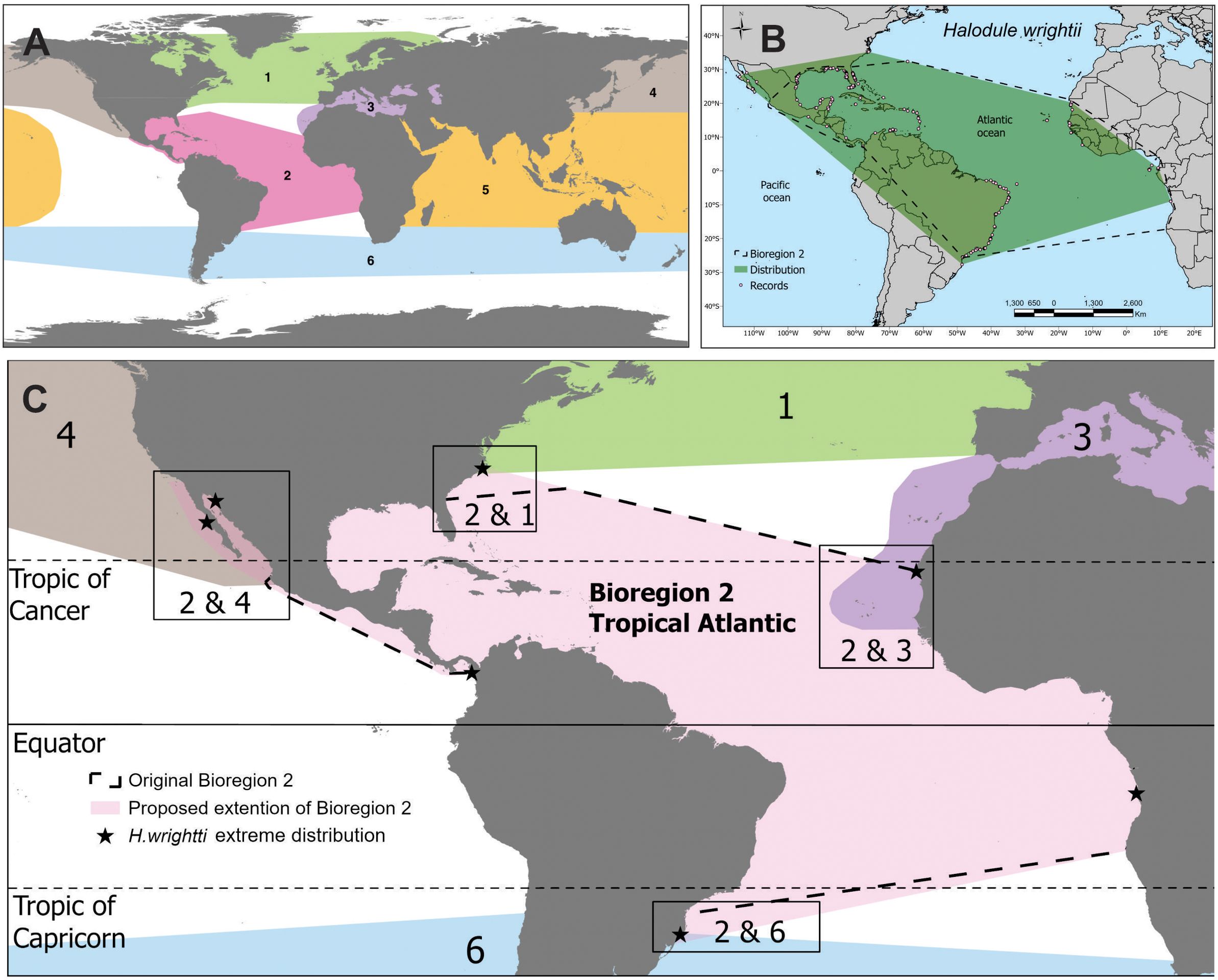
Figure 1. (A) Seagrass bioregions after Short et al. (2007). 1 Temperate North Atlantic, 2 Tropical Atlantic, 3 Mediterranean, 4 Temperate North Pacific, 5 Tropical Indo-Pacific, and 6 Temperate Southern Oceans, (B) Global distribution of Halodule wrightii extending beyond the Tropical Atlantic Bioregion, (C) Tropical Atlantic Bioregion, indicating suggested overlapping transition boundaries between neighboring bioregions indicated by shaded areas. The stars indicate the extreme reported distributions of H. wrightii.
Based on ocean basin and species distribution globally, there are six recognized seagrass bioregions (Figure 1). Two of them are tropical (the Tropical Atlantic and Tropical Indo-Pacific Bioregions), where seagrasses usually have spatial and functional connectivity with coral reefs and/or mangroves (Nagelkerken, 2009). Here they are frequented by large herbivores such as green turtles, dugongs and/or manatees (Short et al., 2007). Tidal ranges tend to be small in the tropical Atlantic, in contrast to the tropical Indo-Pacific, and thus large parts of the Tropical Atlantic Bioregion differ by lacking extensive intertidal flats. According to Short et al. (2007), the Tropical Atlantic Bioregion included 11 seagrass species, less than half of the 24 species found in the Indo-Pacific. The tropical Atlantic is among the bioregions that have reported the greatest loss in seagrass cover in recent years, largely due to degrading water quality and accelerating coastal development (1880-2013; Dunic et al., 2021). These include continued declines in Florida’s Big Bend Region (2001-2022; 12,160 ha loss; Yarbro et al., 2024) and Tampa Bay (2016-2022; 4,661 ha loss; Beck et al., 2024), as well as more acute mortality events in the Indian River Lagoon (2007-2019; 19,114 ha loss; Morris et al., 2022) and Florida Bay (2015; 8,777 ha loss; Hall et al., 2016). Recent International Union for Conservation of Nature (IUCN) biodiversity assessments showed that loss of wildlife biodiversity (in both land, freshwater, and marine systems) has been greatest in Latin America and the Caribbean. This is due to land-use change and cascading effects of global climate change (WWF, 2022), which makes the assessment of its key ecosystems, such as seagrass meadows, urgent.
At global or regional level, seagrass status can be assessed through various quantitative approaches. Waycott et al. (2009) and Dunic et al. (2021) derived the status of seagrass meadows from published losses and gains in area. Assessing areal seagrass meadow loss at landscape scale can also be done using remote sensing tools (Fonseca et al., 2002; Traganos et al., 2020; Lizcano-Sandoval et al., 2022; Yarbro et al., 2024). Another method is evaluating the conservation status of the seagrass species that comprise the meadows (Short et al., 2011). The first global assessment of seagrass species status using the approach of the IUCN Red List Categories and Criteria was published over a decade ago (IUCN, 2010; Short et al., 2011). It was designed to classify species according to their risk of global extinction, and thus provides additional information on the status of seagrass meadows beyond calculations of areal loss. This approach assesses the geographic range (distribution) of the species along with other key factors such as population trend, size and structure, habitat requirements and threats. It may also provide additional information regarding seagrass meadow mapping efforts. For example, less common species living in more marginal seagrass habitats, such as deeper or turbid waters, cannot be easily mapped and tend to be underrepresented in remote sensing surveys. The “seagrass meadows” or patches of these species are also usually small and discontinuous, as well as ephemeral (e.g., Samper-Villarreal et al., 2018). Furthermore, shifts in community composition may precede spatial loss and likewise be poorly captured by mapping approaches (e.g., Fourqurean et al., 2003; Van Tussenbroek et al., 2014). Therefore, an evaluation of the status of seagrass species, either globally or regionally, complements the more common areal extent assessments based on available mapping data.
Since the first global effort to document the conservation status of all known seagrass species up to 2007 (Short et al., 2011), climatic conditions have changed, species distributions have been altered and considerable seagrass-focused research has taken place, which warrants a full update on the status of seagrasses. As part of a larger IUCN effort to assess seagrass status at a global scale, we investigated current information for seagrass species in the Tropical Atlantic Bioregion (or Seagrass Bioregion 2, as described by Short et al., 2007). Regional assessments for species that are only found in this bioregion correspond to their global assessment. Our aim was to provide an updated assessment of the extinction risk for seagrass species for this bioregion according to the IUCN Red List guidelines (IUCN version 3.1, IUCN, 2012).
2 Methods
2.1 Tropical Atlantic Bioregion (seagrass bioregion 2)
The Tropical Atlantic Bioregion for seagrasses includes both tropical coasts of the Atlantic, with its boundaries approximating those of the Tropical Atlantic realm (Spalding et al., 2007), except that it also includes the tropical Pacific coasts of the American continent, also known as the Eastern Tropical Pacific (Short et al., 2007; Figure 1).
In the Atlantic, it encompasses the warm waters of the Atlantic Ocean (typically between 23 °C and 29 °C), primarily between the Tropic of Cancer and the Tropic of Capricorn. The seagrass meadows are distributed along the coastal areas, with denser meadows occurring in shallower waters (<10m deep). The coasts are characterized by diverse marine ecosystems, including coral reefs (hosting the >1000 km long Mesoamerican barrier/fringing reef system in the Caribbean), seagrass meadows, estuaries, mangroves and marsh lands. The major ocean currents in the tropical Atlantic are the warm Mauritia Current, North and South Equatorial Currents, the Caribbean Current and the Gulf Stream. Salinity levels are higher in the central Atlantic (34-35) due to high evaporation rates, and lower near the mouths of large rivers such as the Amazon, Orinoco and Congo. Large coastal sections of both the western Atlantic and Pacific of this bioregion are periodically impacted by tropical storms and hurricanes (cyclones). The extreme (northern and southern) limits of the Tropical Atlantic Bioregion are mainly determined by the distribution limits of characteristic typical tropical seagrass species (Short et al., 2007).
The western tropical Atlantic encompasses various subregions with typical characteristics. In the Gulf of Mexico, seagrasses are often the principal structured habitat type in its many bar-built and open water estuaries (Dawes et al., 2004; Herrera-Silveira et al., 2019). Seagrasses also colonize low- to moderate-energy coastlines, particularly in the Big Bend Region of Florida, and tens of miles onto the West Florida Shelf (Iverson and Bittaker, 1986; Bell et al., 2008), where seagrasses have been reported as deep as 90m (Zieman, 1982). The Mexican Campeche Bank has enormous extensions of meadows dominated by Syringodium filiforme (360 km2) between ~ 4 and 7m depth (Pérez-Espinosa et al., 2019). Typical seascape features of the Caribbean and southern Florida are shallow lagoons, bordered by mostly fringing reefs, with benthos dominated by well-developed seagrass meadows (Fourqurean and Rutten, 2004; Van Tussenbroek, 2011), though seagrasses are also common to estuaries and found in deeper offshore waters. Towards the south, seagrass meadows occur all along the Brazilian coast, but species distribution, abundance and dynamics are affected by physical drivers, particularly by coastal geomorphology, oceanography and regional climate and hydrology (Copertino et al., 2016).
In the eastern tropical Atlantic, in West Africa, seagrasses are mainly associated with shallow mudflats and sand banks, from the intertidal to ~4 m, due to high water turbidity. The West Africa seagrass distribution is likely affected by the prominent currents at the continental shelf in this region where, from north to south, the dominant currents are the Canary Current, the North Equatorial Current, the Guinea Current, and the Benguela Current (Freitas et al., 2019; Hutchings et al., 2009).
In the Eastern Tropical Pacific, the bioregion spans from the state of Jalisco in Mexico until the southern coast of Panama, where it also includes mangroves, coastal wetlands, and coral reefs. Reef lagoons formed by fringing reefs, typical of the Caribbean and southern Florida, are absent here, and seagrasses are mainly found in sheltered bays (Samper-Villarreal, 2024). The tropical Pacific coast experiences a warm and humid climate, where temperatures typically range from 24 °C to 30 °C with a distinct wet (May to October/December) and dry season (November/December to April). The region is influenced by the North and South Equatorial Currents and the Humboldt Current along the South American coast.
2.2 The IUCN assessment guidelines and regional assessment
The IUCN Red List Categories and Criteria (IUCN, 2012) serve to assess the extinction risk at the species level using pre-established criteria and extinction risk theory (Mace et al., 2008). The IUCN Red List Categories recognizes eight levels of risk: Extinct (E), Extinct in the Wild (EW), Critically Endangered (CR), Endangered (EN), Vulnerable (VU), Near Threatened (NT), Least Concern (LC) and Data Deficient (DD; IUCN, 2012).
An initiative to assess the global risk for seagrass species started in 2020, and developed a framework based on seagrass bioregional team leads. For the Tropical Atlantic Bioregion, the bioregional leader (first author) invited an international team of experts to jointly carry out the regional assessment, forming a team with nine members. The members were experts with ample seagrass field experience in the Tropical Atlantic Bioregion and members of the IUCN Species Survival Commission (SSC) Seagrass Species Specialist Group (SSSG). They were selected for their regional knowledge on seagrasses, covering the entire coastline of the Tropical Atlantic Bioregion, based in the United States, Mexico, Costa Rica, Venezuela, Brazil, Mauritania, Mozambique and Portugal.
The regional IUCN Red List Assessments for seagrass species in the Tropical Atlantic Bioregion were conducted between June 2020 and July 2021, based on available data for each seagrass species in the 2007 to 2020-time frame, using the IUCN standard formats for this purpose. Individual species were assigned for evaluation by two or more experts, determined by their regional expertise or knowledge on certain seagrass species. Species evaluations included information on taxonomy, distribution, population trends, ecology, life history, past, existing or future projected threats, and conservation actions for each seagrass. For each species, assessors reviewed the previous assessments published in 2010 (IUCN, 2010), and consulted both published and grey literature covering the period from 2007 to 2020-21. Only grey literature of recognized institutions, such as government agencies and thesis of academic institutions, was considered. If necessary, the assessors contacted other experts to verify information or to obtain additional data that was not available in the literature; in one case (Halodule bermudensis), an external expert to the assessment team was invited to co-author the assessment. Each species assessment was discussed in detail with the entire bioregional team during monthly virtual meetings until a consensus was reached. Quantitative species information, wherever available, or a consensus of expert opinion if needed, was used to determine if a species met the threshold for a threatened category under at least one IUCN Red List Criterion.
Preliminary species assessments were submitted for review to the IUCN, SSC SSSG Leadership Team between November 2020 and July 2021. Written feedback for each assessment was provided by the Leadership Team between June and September 2021, and modifications to assessments were made accordingly. Final assessments were submitted to the Leadership Team between July 2021 and February 2022, awaiting their publication on the IUCN Red List of Threatened Species Platform for the nine endemic seagrass species in the Tropical Atlantic Bioregion, and awaiting collation with other bioregional assessments for global assessment for species that also occur in other bioregions. The regional IUCN Red List assessments for all native seagrass species have been made publicly available in an open repository (Table 1).
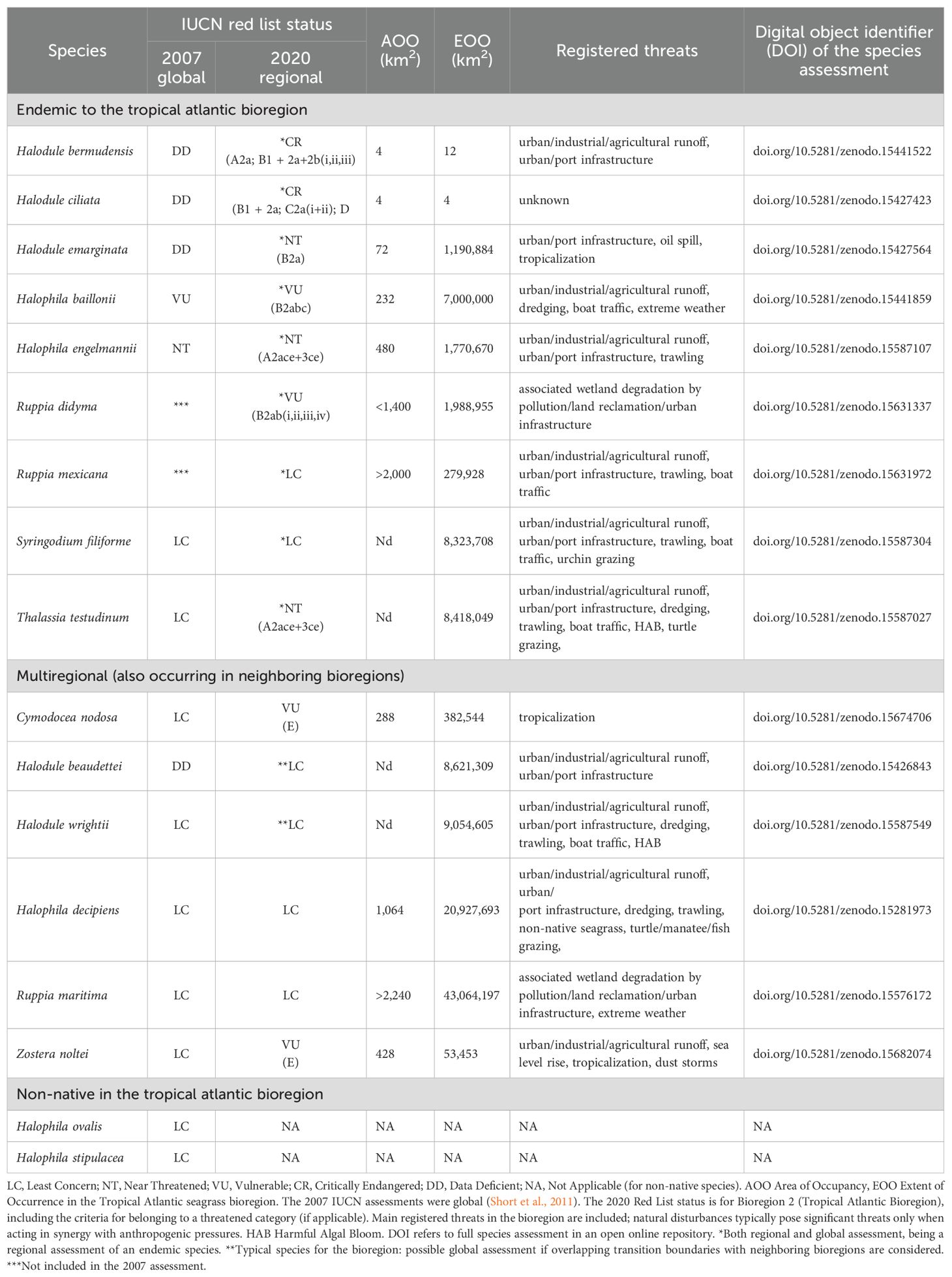
Table 1. IUCN Red List status.
3 Results
Fifteen native species were assessed for the Tropical Atlantic seagrass Bioregion (Table 1). Of these, 40% were threatened in 2020 (13% Critically Endangered and 27% Vulnerable), 20% were categorized as Near Threatened and the remaining 40% were considered to be of Least Concern (Table 1). Nine of the assessed species were endemic to the Tropical Atlantic Bioregion; thus, their assessment was also global. Six species also occurred in other bioregions, of which Halodule beaudettei and H. wrightii were mainly distributed in this bioregion (i.e., typical for the bioregion). The multiregional species (i.e., occurring in more than one seagrass bioregion) were evaluated on regional level only, awaiting final global assessments (but see Discussion for H. beaudettei and H. wrightii). The Tropical Atlantic bioregional IUCN Red List seagrass assessments for each species have been made publicly available in the open repository Zenodo, with digital object identifiers (DOI) assigned for each, and are briefly summarized here (in alphabetical order per subsection). Brief reports on the two non-native species are also included here.
3.1 Species endemic to the Tropical Atlantic Bioregion of seagrasses
3.1.1 Halodule bermudensis (CR, previously DD)
The taxonomic status of H. bermudensis is unclear and it was listed as Data Deficient (DD) in the previous assessment (Short et al., 2011). This species is endemic to Bermuda (Supplementary Figure S1), with the original description (Den Hartog, 1964) being the only published record of collected specimens of this morphological form within the genus (in 1913 at Gibbet Island and in 1922 at Walsingham Bay). It is possible that the genetic character of H. bermudensis could be determined from preserved type specimens (Ferrer-Gallego and Boisset, 2020). Inferred population reduction is likely ongoing, based on general decline of meadows across the Bermuda Platform (Fourqurean et al., 2019); therefore, it is listed as Critically Endangered (CR, Table 1).
3.1.2 Halodule ciliata (CR, previously DD)
The taxonomic status of H. ciliata is unclear and it was listed as Data Deficient (DD) in the previous assessment by Short et al. (2011). Plants with leaf tip morphology as described for H. ciliata have only been reported at Taboga Island, Panama (Den Hartog, 1970) (Supplementary Figure S1). No specimens of this seagrass have been seen for 20 years, despite concerted efforts to locate it and the only previously known location has been destroyed (F. Short and B. Sullivan personal communication). There have been no new reports of this species for the 2007–2020 time period. This species is listed as Critically Endangered (CR, Table 1). It may possibly be extinct because of its small extent of occurrence and area of occupancy (one location).
3.1.3 Halodule emarginata (NT, previously DD)
The taxonomic status of H. emarginata is unclear and it was listed as Data Deficient (DD) in the previous assessment (Short et al., 2011). Plants with leaf tips corresponding to H. emarginata have only been reported in Brazil, and stable populations have been found at 10 locations (Supplementary Figure S1). While new populations have been identified since the last assessment, these likely represent an increased effort and not a range expansion of the species. Presently, this species is found in a few fragmented populations and it is therefore classified as Near Threatened (NT, Table 1).
3.1.4 Halophila baillonii (VU, previously VU)
In its previous assessment, the clover grass H. baillonii was only known from approximately seven locations (Short et al., 2011). This is a small species that often grows in turbid and shallow waters with restricted visibility, and populations are often small and unstable, which is why it is potentially underreported. Its presence in shallow turbid estuaries, which are increasingly under human pressure, merit concern over its future distribution. Even though it has been reported at new sites since 2007 (Supplementary Figure S1), the area of occupancy (AOO) is still estimated to be less than 2,000 km² as reported in the previous assessment by Short et al. (2011), and in congruence with this assessment, it remains listed as Vulnerable (VU, Table 1).
3.1.5 Halophila engelmannii (NT, previously NT)
Commonly known as star grass, H. engelmannii, a small opportunistic seagrass, usually has a patchy distribution in either shallow habitats (< 10m), or in deeper habitats (10 - >20 m) as sparse to dense meadows in the Gulf of Mexico, and Atlantic coastal waters of Cuba and US (Supplementary Figure S1). In shallow waters, various populations have declined or disappeared, and the deeper waters remain understudied. Because of reported ongoing population decline in shallow near-shore waters, this species remained Near Threatened (NT, Table 1). More attention on status and trends for this diminutive, patchily distributed species will be required in the coming years to better understand its extinction risk, including renewed efforts to map and understand the role of deeper, offshore populations.
3.1.6 Ruppia didyma (VU, first time assessed)
This is the first assessment of this species, which was not included in Short et al. (2011), because at that time it was considered a synonym of R. maritima. However, its species status has since been recognized (Triest et al., 2018) and it was first reported for this bioregion in 1825. R. didyma was originally described from the Caribbean but is presently only known from a few locations in the Yucatan Peninsula, Mexico (Novelo, 1991) and Venezuela (Villarreal et al., 2014) (Supplementary Figure S1). It inhabits small waterbodies with clear, hypersaline water, generally near mangroves; and it has recently been found also in sinkholes with fresh to brackish water (Brigitta I. van Tussenbroek and Duarte Frade, unpublished data). R. didyma has a small area of occupancy, and occurs in 13–16 subpopulations that are likely to be severely fragmented given their disperse distribution and the large distance between them. The single known site in Puerto Rico, Guánica Lagoon, was drained in 1955 (Viqueira Rios et al., 2012), and there are no recent records from other Caribbean islands, where mangrove-dominated wetlands face continuing degradation, such as Saint Barthelemy and Guadeloupe (Ellison and Farnsworth, 1996; Jadot, 2016). In this first assessment the species is therefore categorized as Vulnerable (VU, Table 1).
3.1.7 Ruppia mexicana (LC, first time assessed)
This is the first assessment of R. mexicana as it was split from R. maritima based on chromosome counts and morphological evidence (Den Hartog et al., 2016), and is supported by recent molecular microsatellite and chloroplast haplotype data (Triest et al., 2018). Although at present it is currently known only from the Yucatan Peninsula and adjoining parts of Mexico (Supplementary Figure S1), it likely occurs more widely. R. mexicana is assessed here for the first time and occupies a wide range of habitats, apparently being persistent in eutrophic locations. The species is therefore assessed as Least Concern (LC, Table 1).
3.1.8 Syringodium filiforme (LC)
Manatee grass S. filiforme is a widespread and abundant species throughout its range (Supplementary Figure S1), often found mixed with other seagrass species. But it also forms dense and extensive monospecific meadows. Its overall spatial cover appears stable, and it is increasing in abundance in some places (Van Tussenbroek et al., 2014). Such gains may balance losses elsewhere. The species has a great capacity for recovery once conditions improve. In congruence with the previous assessment for this species (Short et al., 2011) it remains of Least Concern (LC, Table 1).
3.1.9 Thalassia testudinum (NT, previously LC)
Turtle grass T. testudinum, is the most robust species in the Tropical Atlantic Bioregion, where it forms extensive, dense and stable seagrass beds, and is considered to be the most important habitat-forming seagrass species throughout a large part of the bioregion (Supplementary Figure S1). The previous Red List Status LC (Least Concern, Short et al., 2011) of this species has been changed to NT (Near Threatened) due to localized threats throughout the region causing declines in condition and loss of many populations since the last assessment in 2007 (Table 1). Localized threats in the region included those driven by coastal urbanization and pressure from nutrient enrichment (Van Tussenbroek et al., 2014; Hall et al., 2016), algal blooms (e.g., holopelagic Sargassum spp., Van Tussenbroek et al., 2017), overgrazing by green sea turtles (Fourqurean et al., 2019) and competitive exclusion by the non-native seagrass H. stipulacea (Winters et al., 2020). These local pressures are all expected to increase, and in synergy they may result in unexpected collapse of stable T. testudinum dominated meadows (Van Tussenbroek et al., 2014). For these reasons the species has now changed category from Least Concern to Near Threatened (NT) under criterion A (population reduction, Table 1).
3.2 Species found in the Tropical Atlantic and neighboring bioregions
3.2.1 Cymodocea nodosa (regionally VU)
C. nodosa is found throughout the Mediterranean Sea, extending northwards into the Atlantic Ocean to mid-Portugal and southwards to Madeira, the Canary Islands, as well as the African continental coasts of Mauritania, Senegal, The Gambia and the archipelago of Cabo Verde (but see discussion in Creed et al., 2016). The Tropical Atlantic Bioregion contains the southernmost populations of the species (Supplementary Figure S1), which are very genetically distinct from the Mediterranean populations (Alberto et al., 2008). The overall population in this bioregion is thought to be presently stable, but ecological niche models predicted that the entire distribution area in the Tropical Atlantic Bioregion may be lost by 2050 under the RCP 2.6 and RCP 8.5 climate change scenarios and considering sea level rise (Chefaoui et al., 2018; 2021). The species is therefore listed as Vulnerable (VU) for this bioregion under criterion E (quantitative extinction analysis; Table 1).
3.2.2 Halodule beaudettei (regionally LC, previously DD)
The taxonomic status of H. beaudettei is unclear and it was listed as Data Deficient (DD) in the previous assessment (Short et al., 2011). Since then, the known range of the species has been expanded based on recent and previously published records (Supplementary Figure S1), mostly correcting past misidentifications. This is not a true species range expansion, yet illustrative of current taxonomic uncertainty. As this species has a wider distribution than reported in the previous assessment period (Short et al., 2011), and extending into Bioregion 1 (Temperate North Atlantic), it is now listed as Least Concern (LC, Table 1).
3.2.3 Halodule wrightii (regionally LC)
Commonly referred to as shoal grass, H. wrightii is a widespread and locally abundant species in the Caribbean Sea, Gulf of Mexico and Eastern Tropical Pacific (Supplementary Figure S1), where it is often found together with other seagrass species such as Thalassia testudinum, Syringodium filiforme, or Halophila spp., but it can also form dense and extensive monospecific meadows in habitats considered challenging for other major meadow forming species such as T. testudinum or S. filiforme. It is the dominant species in Brazil and the tropical coast of western Africa from Mauritania to Sierra Leone as well as Cabo Verde, São Tomé e Príncipe, and Angola. It extends into neighboring seagrass bioregions, being, Bioregion 1 (Temperate North Atlantic), Bioregion 4 (Temperate North Pacific) and Bioregion 6 (Temperate Southern Oceans) (Supplementary Figure S1). The overall population trend for this species is stable, and possibly increasing in some parts of its range, such as in Brazil, due to variations in water temperature and salinity (Copertino et al., 2016). Given its wide distribution and abundance, as well as its resilience to multiple stressors, in congruence with the previous assessment for this species (Short et al., 2011), in this regional assessment it remains of Least Concern (LC, Table 1).
3.2.4 Halophila decipiens (regionally LC)
The paddle grass H. decipiens has a tropical and subtropical, circumglobal distribution and can be locally abundant. In deeper waters, H. decipiens has few major threats; however, coastal development, reduced water quality (Short et al., 2011), invasive species, overgrazing, storms and hurricanes can affect seagrass beds in shallower and some deeper (>20m) areas. The population trend in the Tropical Atlantic Bioregion is stable, with range expansion such as in Brazil (Gorman et al., 2016) and eastern Florida, USA (Virnstein and Hall, 2009) (Supplementary Figure S1). In congruence with the previous assessment for this species (Short et al., 2011), H. decipiens remains of Least Concern (LC, Table 1) in this regional assessment.
3.2.5 Ruppia maritima (regionally LC)
R. maritima (sensu lato, see Discussion) is a colonizing seagrass that tolerates a wide range of environmental conditions, is capable of fast growth and reproduction (Strazisar et al., 2015), and has a wide distribution, extending into bioregions north and south of the tropical Atlantic (Short et al., 2007). The general population trend in the bioregion is stable or increasing (e.g. Martínez-Garrido et al., 2017), with R. maritima showing resilience to large disturbance events (hurricanes, oil spills), and occasionally replacing other seagrasses in the northern Gulf of Mexico following disturbance (Cho et al., 2009, 2017). Although parts of the region (West Africa and the Caribbean) remain poorly studied, the species is tolerant of disturbance, and is still widely distributed and locally common in the bioregion (Supplementary Figure S1). It is therefore listed as Least Concern in the Tropical Atlantic Bioregion (LC, Table 1).
3.2.6 Zostera noltei (regionally VU)
Worldwide, Z. noltei occurs in the eastern Atlantic as well as the Baltic, Mediterranean, Black, Caspian and Aral Seas. The Tropical Atlantic Bioregion contains populations of this species at their southernmost limits of distribution, with large areas in Mauritania and smaller sites (~ 40 km diameter) in Senegal Saloum Delta (Sidi Cheikh et al., 2023) (Supplementary Figure S1). There, it is predicted to be negatively affected in the future from ongoing climate change, especially extreme climate events and sea-level rise as the entire distribution in the Tropical Atlantic Bioregion may be lost by 2050 under the RCP 2.6 and RCP 8.5 climate change scenarios and considering sea-level rise (Chefaoui et al., 2021). Large-scale mortality events have already been reported in Mauritania (De Fouw et al., 2016). The species is therefore listed as Vulnerable (VU, Table 1) for this bioregion, under criterion E (quantitative extinction analysis).
3.3 Non-native species
3.3.1 Halophila ovalis (NA)
H. ovalis is wide-spread in the Indo-Pacific and Red Sea, and it is an introduced species in Florida and the Caribbean (Short et al., 2011) (Supplementary Figure S1). It is a rapidly growing opportunistic species with high shoot turnover rate. The Florida subpopulation was previously described as Halophila johnsonii (Eiseman and McMillan, 1980), an endemic species known only from portions of the Indian River Lagoon (Virnstein and Hall, 2009) south to Virginia Key (Gelber et al., 2000; Christian and Hall, 2012), but is now recognized as a synonym of H. ovalis subsp. ovalis (Waycott et al., 2021). Although H. ovalis populations are sparse and fragmented; the geographical range is gradually expanding in the Caribbean (Waycott et al., 2021). As a non-native species it is listed as Not Applicable (NA, Table 1) in this regional assessment.
3.3.2 Halophila stipulacea (NA)
H. stipulacea is native in the Indo-Pacific Ocean and Red Sea and is an introduced species in the Caribbean, where it is rapidly spreading, colonizing the Caribbean islands and mainland Venezuela (Vera et al., 2014; Willette et al., 2014) (Supplementary Figure S1). This small species has a high shoot turnover rate, and is therefore a rapidly growing species. At places it poses a threat to native species, but in others it does not (Winters et al., 2020). More recently in 2024, this species was also reported on the mainland of the United States (Key Biscayne, Florida) for the first time (Campbell et al., 2025). As a non-native species H. stipulacea in this regional assessment is listed as Not Applicable (NA, Table 1).
4 Discussion
4.1 Seagrass extinction risk in the bioregion, from an IUCN-red list viewpoint
Here we covered a total of 17 species (with 15 being native), whereas Short et al. (2007) only recognized 11 species for the Tropical Atlantic Bioregion. In this assessment we also included formerly DD or ignored species (Halodule beaudettei, H. bermudensis, H. ciliata, H. emarginata, Ruppia didyma), two temperate species (Cymodocea nodosa and Zostera noltei) that extend into the tropical waters of western Africa, and a newly described species (R. mexicana). Out of the 17 species present in the Tropical Atlantic Bioregion, nine are endemic to the bioregion (Halodule bermudensis, H. ciliata, H. emarginata, Halophila baillonii, H. engelmannii, Ruppia didyma, R. mexicana, Syringodium filiforme, Thalassia testudinum). In addition, two species (Halodule beaudettei and H. wrightii) are typical of this bioregion but extend beyond its boundaries. Others have circumglobal distributions (Halophila decipiens and Ruppia maritima), or are non-native (Halophila ovalis and H. stipulacea).
Most of the abundant or ubiquitous species in the Tropical Atlantic Bioregion (H. wrightii, R. maritima, R. mexicana, S. filiforme, and H decipiens) are classified as least concern (LC, except for T. testudinum-NT), even though the areal extents of seagrass meadows in this bioregion are in decline (Dunic et al., 2021). These species are not considered threatened, mostly because of their large geographical distribution. Species having more limited distribution or typically occurring in challenging habitats such as deep waters, turbid estuaries or saltpans (e.g., H. baillonii, H. engelmannii, R. didyma) received a threatened status based on these premises. Many of the populations of these species are ephemeral, occurring in marginal conditions for seagrass growth. Due to their inconspicuous and ephemeral nature, they may disappear from a geographical area without being noticed, which may be the case for H. engelmannii along the Yucatan coast of the Gulf of Mexico, although more thorough search actions are needed to verify this. More attention to such ephemeral seagrass species with limited distribution in monitoring programs and designing protected areas is recommended.
Most seagrass species are still widespread, and abundant throughout the region, which is to be expected from species that found ecosystems. Widespread and abundant founding species are often not threatened at the species level, even when the ecosystems are in decline, which for example also applies to mangroves. Therefore, conservation efforts of these species (such as T. testudinum, S. filiforme, Halodule spp., Ruppia spp.) can best be approached considering ecosystem vulnerabilities rather than the (sole) extinction risk of the individual species, following the Red List of Mangrove Ecosystems initiative (https://iucn.org/resources/conservation-tool/iucn-red-list-ecosystems/red-list-mangrove-ecosystems). From an ecosystem perspective it is worrying that T. testudinum, the most robust and iconic seagrass species in this region has been downgraded from Least Concern to Near Threatened linked to ongoing urban development, loss in water quality and algal blooms (Table 1). Even though the distribution of the species has not changed, the specific composition in the meadows has undergone changes which are increasingly visible (Van Tussenbroek et al., 2014), even with remote sensing tools (Ávila-Mosqueda et al., 2025). If typical non-climax seagrass species, such as H. wrightii or S. filiforme, were to eventually replace T. testudinum, there could be unanticipated effects on ecological functions, ecosystem services, and future resilience to physical and environmental disturbance (O’Brien et al., 2018). A similar shift has been projected for West Africa, where the existing tall, dense meadows of C. nodosa and Z. noltei are predicted to be replaced by the smaller, thinner H. wrightii under climate change and sea level rise scenarios (Chefaoui et al., 2021). Disturbingly little is known about the functionality and persistence of seagrass meadows under such system-wide changes of specific composition. Region-wide, and ongoing monitoring of such meadows by remote sensing and ground truthing could aid in observing future conservation state of these species and the ecosystem they found, and in developing site-specific management plans that address existing stressors or threats. Such monitoring programs could benefit from past and existing transnational networks within the region, e.g. CARICOMP in the Caribbean (Cortés et al., 2019) or RAMPAO in West Africa (Duval-Diop, 2016).
4.2 Assessment challenges
The first assessment of the conservation status of seagrasses following the IUCN procedure was a global evaluation conducted more than a decade ago (IUCN, 2010; Short et al., 2011). The current study sought to revisit that earlier effort, offering an updated assessment for species found in the Tropical Atlantic Bioregion. While investigating species’ status and trends we encountered several taxonomic and theoretical challenges. Here we describe the challenges, mention how we assessed species in the face of them, and propose how they may be addressed in the future.
4.2.1 Species identification
Various seagrass species share similar morphological vegetative characteristics, making it difficult to distinguish between them based on vegetative morphology alone, which is further hampered by reduced reproductive parts in adaptation to underwater fertilization (Kuo and Den Hartog, 2001). In addition, seagrasses exhibit significant phenotypic plasticity (the ability of an organism to change its characteristics in response to the environment; McDonald et al., 2016; Pazzaglia et al., 2021), and dimensions and morphologies of plant parts of related species can overlap, leading to confusion regarding species boundaries. Seagrasses are found in a wide range of environments, and species´ morphologies and dimensions may vary geographically (McDonald et al., 2016), which can lead to erroneous recognition of different (sub)species. In contrast, in some cases, what appears to be a single species may actually consist of two or more cryptic species that are genetically distinct but morphologically indistinguishable (Martínez-Garrido et al., 2016; Triest et al., 2018). Interbreeding between some congeneric seagrass species has also been reported (e.g. Ito and Tanaka, 2011; Sinclair et al., 2019; Liu et al., 2020), making it difficult to determine clear taxonomic distinctions. While genetic analysis is a powerful tool for distinguishing species, the genetic data available for seagrasses is still relatively limited (e.g., Dilipan et al., 2018; Tuya et al., 2024). In the past, clear seagrass identification guides or keys were not widely available, and during this assessment we were able to reveal various cases of species misidentification, requiring adjustments in geographical distributions.
Although all Halophila species in this bioregion have distinct morphologies, misidentifications have occurred in the past; possibly due to a combination of limited visibility and historical lack of seagrass identification guides. H. baillonii, for example, has been misidentified as H. decipiens, H. johnsonii, or H. engelmannii (Short et al., 2006; Samper-Villarreal et al., 2018; Creed and Samper-Villarreal, 2019). This species is often found in deep or turbid waters with low visibility, where it is covered with fine sediments, which hampers field identification. H. baillonii is currently not considered to be present in Florida and surrounding area, where it was previously reported and most likely wrongly identified. Similarly, H. engelmannii has been confused with H. baillonii, even though there is no taxonomic uncertainty regarding the identity of either one of these species (Kuo, 2020). Reports of H. engelmannii from the southern and southwestern Caribbean (Trinidad, Puerto Rico, Venezuela and Costa Rica) are also misidentifications of either H. baillonii or H. decipiens. In this assessment, we adjusted to the EOO (minimum convex polygon around all present native occurrences of a species) and AOO (Area of Occurrence) of both species.
Recent molecular advances have clarified several taxonomic uncertainties regarding Halophila species in the region, such as confirming that H. johnsonii is now considered a synonym of H. ovalis subs. ovalis. This species was likely first observed in Florida, USA in the 1950s, but at the time was not distinguished from H. decipiens and also misidentified as H. baillonii (Phillips, 1960). In 1972, specimens from the Indian River Lagoon, Florida, were considered to be H. johnsonii (Eiseman, 1973), and Johnson’s seagrass was formally described soon after (Eiseman and McMillan, 1980). Since then, the taxonomic status of H. johnsonii has been repeatedly called into question (Short et al., 2010; Fonseca and Olsen, 2016). Early molecular studies showed H. johnsonii was genetically similar to and closely allied with a number of its congeners (Short et al., 2010; Waycott et al., 2015) forming what was proposed as the “Halophila ovalis species complex” (sensu Den Hartog, 1970). Some argued that H. johnsonii was H. ovalis introduced from the Indo-Pacific (Short et al., 2010; Waycott et al., 2015, but see Kuo, 2020), an assertion supported by its observed asexual (York et al., 2008) and monoclonal (Waycott et al., 2015) population. Populations of this species were originally restricted to Florida (NOAA-NMFS, National Oceanic and Atmospheric Administration, National Marine Fisheries Service, 2007), but recently appeared in the broader Caribbean (Short et al., 2010; Waycott et al., 2015). DNA sequences, microsatellites, and ddRAD SNPs from H. johnsonii and H. ovalis throughout their respective ranges, revealed that the Florida and Antiguan populations of H. johnsonii were (1) identical, (2) entirely monoclonal, and (3) well resolved with east African subpopulations of H. ovalis (Waycott et al., 2021). In response, the United States’ National Marine Fisheries Service (NMFS) removed H. johnsonii from the List of Threatened and Endangered Species and eliminated its critical habitat designation in 2022 (Federal Register, 2022). It is now generally regarded as a synonym of H. ovalis subsp. ovalis (e.g., https://wfoplantlist.org, https://powo.science.kew.org, https://florida.plantatlas.usf.edu).
In the previous assessments, H. beaudettei, H. bermudensis, H. ciliata, and H. emaginata were listed as data deficient (DD, Short et al., 2011) due to taxonomic uncertainties, which remain until today. Halodule wrightii is the only well-established species of this genus in the Tropical Atlantic Bioregion. The taxonomy of Halodule spp. is based primarily on leaf tip morphology, because observations of its sexual reproductive structures have been lacking or infrequent, and they are highly reduced (Kuo and Den Hartog, 2001). Furthermore, Halodule spp. are phenotypically plastic, and the use of leaf tip morphology as a diagnostic character has been repeatedly questioned (Phillips, 1967; Ito and Tanaka, 2011; Wagey and Calumpong, 2013). In some cases, the leaf tip shape, used to separate the species, has been shown to vary among shoots from the same rhizome (Phillips, 1967; Wheeler et al., 2020). Genetic studies have indicated that most Atlantic Halodule spp. populations are closely related, from the Caribbean and Brazil to all of Africa, with higher differentiation inside the Gulf of Mexico, and future synonymizing of several species with H. wrightii is possible (e.g. Tavares et al., 2023; Sousa et al., 2025).
The taxonomy of Ruppia remains problematic and is undergoing revision (Den Hartog and Triest, 2020). R. maritima has traditionally been considered cosmopolitan (Short et al., 2011), but recent molecular studies demonstrated that it is a complex of multiple species, including polyploids and recent and ancient hybrid lineages (Ito et al., 2010; 2013; 2015; Triest et al., 2018). R. maritima sensu stricto (i.e. the diploid species in this complex that retains the original name) was described from northern Europe (Den Hartog and Triest, 2020), and has a wide but patchy distribution throughout Europe and the Mediterranean Basin (Triest and Sierens, 2014). It also occurs in the Tropical Atlantic Bioregion as it is present in Cabo Verde (Martínez-Garrido et al., 2017). In other parts of the world, several populations previously treated as R. maritima have been described as distinct species (Novelo, 1991; Yu and Den Hartog, 2014; Den Hartog et al., 2016; Kurniawan et al., 2024). Further cryptic species may have been identified but await formal description (Ito et al., 2010; 2015; Martínez-Garrido et al., 2016; Triest et al., 2018). In the Tropical Atlantic Bioregion, considerable morphologically diversity has been reported (Fernald and Wiegand, 1914; Koch and Seeliger, 1988; Oliveira Filho et al., 1983), but the taxonomy and distribution of these potentially cryptic species remain insufficiently studied. Most records lack sufficient information to be assigned to the distinct species, and therefore they are collectively considered as R. maritima in the present assessment, pending further studies. Morphological, cytological, and molecular studies indicated that R. mexicana and R. didyma are considered distinct species (Novelo, 1991; Den Hartog et al., 2016; Triest et al., 2018), and their conservation status is determined for the first time in these assessments.
In the previous IUCN global assessment, both C. nodosa and Z. noltei were reported for Cabo Verde islands (IUCN, 2010). But recent large-scale surveys revealed only the presence of H. wrightii and R. maritima in these islands (Creed et al., 2016; Martínez-Garrido et al., 2017). Although not registered by latter authors, C. nodosa was recently documented in Santiago Island, confirming its presence in Cabo Verde (Associação Caboverdiana de Ecoturismo ECOCV, pers. com.). But, at present, there is insufficient evidence of occurrence of Z. noltei in Cabo Verde islands. On one hand, it is possible that H. wrightii and R. maritima were wrongly identified as Z. noltei. But, on the other hand, records for Cabo Verde islands may also stem from confusion with the similarly named Cape Vert, in Senegal which is an upwelling area and within the range of this species. The two places have previously been confused in the literature (e.g., records of Ecklonia sp. in Price et al., 1978). Thus, adjustments were made to Z. noltei EOO and AOO in the Tropical Atlantic Bioregion.
4.2.2 Defining boundaries of the bioregions
Short et al. (2007) established six seagrass bioregions based on species assemblages, their distribution ranges, and influences from tropical and temperate climates, along with physical barriers set by oceans and seas. These authors considered the distinction between temperate and tropical zones crucial, as well as the restriction of certain species assemblages to specific sea or ocean basins. The Tropical Atlantic Bioregion was characterized as having clear water with a high diversity of seagrasses on reefs and shallow banks, dominated by T. testudinum. The Eastern Tropical Pacific and the western tropical African coastline, included by these authors, do not entirely fit this characterization, but they share common species with the tropical western Atlantic, and their inclusion in this bioregion is therefore considered as justified.
However, the temperate-tropical divide is not clearly defined, and tropical species, such as H. wrightii, may spread into temperate waters up to the coasts of North Carolina in the western Atlantic (Stevenson et al., 2025). Typical temperate species, such as C. nodosa, Zostera marina and Z. noltei have marginal populations co-occurring with tropical species such as H. wrightii. Considering the diffuse divide between temperate and tropical species extreme distributions, we propose that the boundaries between adjacent bioregions could be overlapping. The overlapping transition boundaries between adjacent bioregions will maintain the original principal for the creation of the seagrass bioregions, based on temperate-tropical species assemblages. At the same time, they provide more flexibility in the assessments, reducing the overlap of regional assessments for species with marginal populations in a bioregion with main distribution in its neighboring bioregion(s). For the Tropical Atlantic Bioregion, for instance, such an overlapping transitional boundary includes the marginal populations at extreme distribution, such as Z. noltei and C. nodosa. These species have their main distribution in the Mediterranean Bioregion (C. nodosa) or Mediterranean and Temperate North Atlantic Bioregion (Z. noltei; Short et al., 2011). The establishment of an overlapping transition boundary, excludes the need for their regional assessments in the Tropical Atlantic Bioregion. H. beaudettei and H. wrightii have their main distributions in the Tropical Atlantic Bioregion, but they also extend into neighboring bioregions (Figures 1, S1), and their global assessments depend at present on regional evaluations of the marginal populations in the neighboring bioregions. In establishing overlapping transition boundaries, the assessment of these species would be global and final, with EOO of 10,827,677 km2 for H. beaudettei, and 50,655,025 km2 and H. wrightii.
We suggest to use of the global distribution of H. wrightii (rounded to half a degree) to define the transitional overlapping boundaries of the Tropical Atlantic Bioregion, and we propose the following overlapping transition zones between adjacent seagrass bioregions (Figure 1):
1. Northwestern Atlantic coast: overlapping with bioregion 1 (Temperate North Atlantic), from current boundary until North Carolina coast at 38°N. The extreme northern distribution of H. wrightii was at 37.70°N in 2023 (Stevenson et al., 2025), where it is co-occurring with Z. marina and R. maritima.
2. Southwestern Atlantic coast: overlapping with bioregion 6 (Temperate Southern Oceans), from current boundary until Brazilian coast at 28°N (Tavares et al., 2023).
3. Eastern Pacific coast: overlapping with bioregion 4 (Temperate North Pacific), from current boundary until 28°N on the Pacific coast at Guerrero Negro (27.97°N), where H. wrightii was found in 2019, coexisting with Z. marina and Ruppia sp. This transition overlapping zone also includes the Gulf of California with (annual populations of) Z. marina, Ruppia spp. and the tropical seagrass species H. wrightii and H. decipiens (Wong et al., 2022). The northern boundary of bioregion 2 in the area is currently unclear as it was not shown in the original map of the seagrass bioregions (Figure 1 in Short et al., 2007).
4. Northwestern African coast: overlapping with bioregion 3 (Mediterranean), corresponding to the area where the distribution of temperate Z. noltei or C. nodosa (whichever species extends further) overlaps with that of H. wrightii (Figure 1). This includes the Cabo Verde archipelago, where C. nodosa was recently reported (Associação Caboverdiana de Ecoturismo ECOCV, pers. com.). This region is influenced by both the cold-water Canary upwelling system and the warm-water Mauritania Current (Klenz et al., 2018), creating a unique transition area between temperate and tropical ecosystems (Spalding et al., 2007; Freitas et al., 2019).
Considering ongoing tropicalization (Hyndes et al., 2017; 2022), the overlapping transition boundaries of the neighboring bioregions may be of special interest, as tropical species may ingress into more temperate neighboring regions. Also, on the African coast, temperate species such as C. nodosa and Z. noltei (Hyndes et al., 2017; 2022), and (annual) populations of Z. marina may disappear or transgress towards more temperate waters due to tropicalization (Bartenfelder et al., 2022). Overlapping transition boundaries between tropical and temperate bioregions may thus be useful in future assessments of the conservation status of seagrass species, which may also apply to corals, mangroves or saltmarsh species. The proposed overlapping transition boundaries may need adjustments in the future, which may by itself also be an indicator of species shifts around the globe.
The original seagrass bioregions (Short et al., 2007, Figure 1) did not include large sections of the western African and eastern Pacific coasts. Along the west African coast, the southern limit of the Tropical Atlantic Bioregion extends beyond the southern extreme known distribution of H. wrightii in Angola (Figure 1); further studies on seagrass distribution may allow better definition of this boundary. Further south, the coast of Namibia is more temperate, characterized by low water temperatures due to the Benguela Current and strong coastal upwelling, harboring coastal lagoons with Ruppia sp. meadows (Clarke and Klaasen, 2001). With exception of Zostera capensis (Adams, 2016), seagrass meadows near the southern tip of Africa remain poorly studied. Along the eastern Pacific coast, the southern limit of the Tropical Atlantic Bioregion was established at the southern border of Panama (UNEP-WCMC (United Nations Environment Programme World Conservation Monitoring Centre) and Short, 2005). However, this region has been poorly studied (Samper-Villarreal, 2024). Nearby Panama, Colombia and Ecuador have potentially suitable sheltered estuarine habitats for seagrasses, meriting more studies. Further south, the coastline lacks sheltered embayments and is influenced by the cold Humboldt current. Although seagrasses have not been registered along these coastlines to date, the presence of sparse seagrass populations can´t be ruled out. We therefore recommend that unassigned coastal sections falling between current seagrass bioregions be absorbed during the pending updated global assessment of the seagrass conservation status.
5 Conclusions
Here, we have presented an update of the conservation status for the seagrass species in the Tropical Atlantic Bioregion following the previous worldwide assessment in 2007 (IUCN, 2010; Short et al., 2011), under the Categories and Criteria of the International Union for the Conservation of Nature (IUCN) Red List of Threatened Species.
Present conservation status assessments confirmed most of the previous ones (10 out of 11), with the exception of T. testudinum that passed from Least Concern (LC) to Near Threatened (NT) category (Criterium A. Population size reduction).
Whereas the last assessment covered 11 species for this bioregion, the current one covered fifteen native species, including the ones which were previously considered DD (Data Deficient), not yet recognized as species, or they were not included in the bioregion being peripheral populations typical for neighboring bioregions. In this manuscript, we also briefly covered the two non-native species, that were not assessed for conservation status, following the IUCN protocol (IUCN, 2012).
Issues concerning fixed boundaries of seagrass bioregions were identified. Therefore, some overlapping transition boundaries with neighboring bioregions are proposed, for practical assessment purposes, and considering changes in the geographical distribution of some seagrass species due to global climate change. Observations of species distribution (either expansion or retraction) in these overlapping zones of the bioregions may be of value to observe potential impacts of tropicalization.
We recommend the inclusion of unassigned coastal stretches to bioregions during the pending global assessment.
During the bioregional assessment, challenges concerning species identifications were identified and addressed, which included misidentifications, presence of cryptic species and ongoing uncertainty regarding the taxonomical status of several species.
Region-wide seagrass meadow monitoring programs, and future assessments of the species conservation status, are recommended to understand the conservation status and trends of these critical ecosystems in the Tropical Atlantic Bioregion.
We suggest implementation of targeted seagrass monitoring programs for the threatened and more marginal smaller seagrass species, which are often overlooked in monitoring programs of seagrass meadows.
In addition to the Red List Species assessments, we suggest assessment of the seagrass meadows as an ecosystem functional group following the criteria and guidelines of IUCN Red List of Ecosystems (Bland et al., 2017).
Author contributions
JS-V: Validation, Data curation, Formal Analysis, Visualization, Conceptualization, Resources, Funding acquisition, Writing – review & editing, Supervision, Investigation, Project administration, Writing – original draft. JC: Conceptualization, Investigation, Supervision, Writing – review & editing, Formal Analysis, Writing – original draft, Validation, Data curation, Visualization. DF: Writing – original draft, Writing – review & editing, Conceptualization, Investigation, Validation, Visualization, Supervision, Formal Analysis, Data curation. BF: Data curation, Writing – review & editing, Investigation, Writing – original draft, Supervision, Visualization, Conceptualization, Validation, Formal Analysis. SB: Visualization, Data curation, Investigation, Validation, Conceptualization, Writing – review & editing, Writing – original draft, Formal Analysis. HE-H: Formal Analysis, Writing – original draft, Data curation, Investigation, Conceptualization, Visualization, Validation, Writing – review & editing. AP: Formal Analysis, Visualization, Writing – original draft, Data curation, Writing – review & editing, Conceptualization, Validation, Investigation. ES: Conceptualization, Visualization, Investigation, Writing – review & editing, Validation, Formal Analysis, Data curation, Writing – original draft. BVV: Conceptualization, Visualization, Investigation, Writing – review & editing, Validation, Formal Analysis, Data curation, Writing – original draft. BVT: Visualization, Validation, Conceptualization, Formal Analysis, Writing – review & editing, Investigation, Data curation, Writing – original draft, Supervision.
Funding
The author(s) declare financial support was received for the research and/or publication of this article. The Vicerectoría de Investigación of the Universidad de Costa Rica supported the work done by JS-V. Work by DF and ES (Atlantic Africa) was supported by FCT (Portugal) and the Aga-Khan Foundation program MARAFRICA, and EU-BiodivERsA BiodivRestore-253 -FCT DivRestore/0013/2020 and UIDP/04326/2020, LA/P/0101/2020, SFRH/BD/147343/2019.
Acknowledgments
We thank Kelly Kingdon (University of Trinidad and Tobago) and Hector Ruíz (University of Puerto Rico) for their feedback on our queries or verification of identification of Halophila spp. Valery Ávila Mosqueda helped with the elaboration of the distribution maps. Kathryn Coates contributed to the assessment of Halodule bermudensis.
Conflict of interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
The author(s) declared that they were an editorial board member of Frontiers, at the time of submission. This had no impact on the peer review process and the final decision.
Generative AI statement
The author(s) declare that no Generative AI was used in the creation of this manuscript.
Any alternative text (alt text) provided alongside figures in this article has been generated by Frontiers with the support of artificial intelligence and reasonable efforts have been made to ensure accuracy, including review by the authors wherever possible. If you identify any issues, please contact us.
Publisher’s note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
Supplementary material
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fcosc.2025.1656995/full#supplementary-material
References
Adams J. B. (2016). Distribution and status of Zostera capensis in South African estuaries—a review. South Afr. J. Bot. 107, 63–73. doi: 10.1016/j.sajb.2016.07.007
Alberto F., Massa S., Manent P., Diaz-Almela E., Arnaud-Haond S., Duarte C. M., et al. (2008). Genetic differentiation and secondary contact zone in the seagrass Cymodocea nodosa across the Mediterranean–Atlantic transition region. J. Biogeogr. 35, 1279–1294. doi: 10.1111/j.1365-2699.2007.01876.x
Ávila-Mosqueda S. V., Van Tussenbroek B. I., and Garza-Pérez J. R. (2025). Changes in seagrass landscape configuration in a Caribbean reef lagoon indicate an ecosystem shift after repeated disturbances. Coasts 5, 8. doi: 10.3390/coasts5010008
Bartenfelder A., Kenworthy W. J., Puckett B., Deaton C., and Jarvis J. C. (2022). The abundance and persistence of temperate and tropical seagrasses at their edge-of-range in the Western Atlantic Ocean. Front. Mar. Sci 9. doi: 10.3389/fmars.2022.917237
Beck M. W., Flaherty-Walia K., Scolaro S., Burke M. C., Furman B. T., Karlen D. J., et al. (2024). Hot and Fresh: Evidence of climate-related suboptimal water conditions for seagrass in a large Gulf coast estuary. Estuaries. Coasts. 47, 1475–1497. doi: 10.1007/s12237-024-01385-0
Bell S. S., Fonseca M. S., and Kenworthy W. J. (2008). Dynamics of a subtropical seagrass landscape: links between disturbance and mobile seed banks. Landscape Ecol. 23, 67–74. doi: 10.1007/s10980-007-9137-z
Bland L. M., Keith D. D. A., Miller R. M., Murray N. J., and Rodríguez J. P. (2017). Guidelines for the application of IUCN Red List of Ecosystems Categories and Criteria. Version 1.1 (Gland, Switzerland: IUCN).
Campbell J. E., Allen A. C., Sattelberger D. C., White M. D., and Fourqurean J. W. (2025). First record of the seagrass Halophila stipulacea (Forsskal) Ascherson in the waters of the continental United States (Key Biscayne, Florida). Aquat. Bot. 196, 103820. doi: 10.1016/j.aquabot.2024.103820
Chefaoui R. M., Duarte C. M., and Serrão E. A. (2018). Dramatic loss of seagrass habitat under projected climate change in the Mediterranean Sea. Global Change Biol. 24, 4919–4928. doi: 10.1111/gcb.14401
Chefaoui R. M., Duarte C. M., Tavares A. I., Frade D. G., Sidi Cheikh M. A., Ba M. A., et al. (2021). Predicted regime shift in the seagrass ecosystem of the Gulf of Arguin driven by climate change. Global Ecol. Conserv. 32, e01890. doi: 10.1016/j.gecco.2021.e01890
Cho H. J., Biber P. D., Darnell K. M., and Dunton K. H. (2017). Seasonal and annual dynamics in seagrass beds of the grand bay national estuarine research reserve, mississippi. Southeastern. Geogr. 57, 246–272. doi: 10.1353/sgo.2017.0024
Cho H. J., Biber P., and Nica C. (2009). “The rise of ruppia in seagrass beds: changes in coastal environment and research needs. Handbook on environmental quality,” in Handbook on Environmental Quality. Eds. Drury E. K. and Pridgen T. S. (New York, USA: Nova Science),New York 1–15.
Christian J. and Hall M. O. (2012). Overview of projects impacting seagrass habitat with potential for enhancement of Johnson’s seagrass (Halophila johnsonii). FWC-FMRI F2881-10-12-F (St Petersburg, FL, USA: National Oceanic and Atmospheric Administration, National Marine Fisheries Service).
Clarke N. V. and Klaasen E. S. (2001). Water plants of Namibia: an identification manual. Occasional contributions, 2 (Windhoek, Namibia: National Botanical Research Institute).
Copertino M. S., Creed J. C., Lanari M. O., Magalhães K., Barros K., Lana P. C., et al. (2016). Seagrass and submerged aquatic vegetation (VAS) habitats off the coast of Brazil: state of knowledge, conservation and main threats. Braz. J. Oceanogr. 64, 53–80. doi: 10.1590/S1679-875920161036064sp2
Cortés J., Oxenford H. A., van Tussenbroek B. I., Jordán-Dahlgren E., Cróquer A., Bastidas C., et al. (2019). The CARICOMP network of caribbean marine laboratories, (1985–2007): history, key findings, and lessons learned. Front. Mar. Sci. 5. doi: 10.3389/fmars.2018.00519
Creed J. C., Aranda L. S., de Sousa J. G., Brito do Bem C. B., Dutra B.S.A.V.M., Lanari M., et al. (2023). A synthesis of provision and impact in seagrass ecosystem services in the Brazilian Southwest Atlantic. Sustainability 15, 14722. doi: 10.3390/su152014722
Creed J. C., Engelen A., D’Oliveira E. C., Bandeira S., and Serrão E. A. (2016). First record of seagrass in Cape Verde, eastern Atlantic. Mar. Biodivers. Records. 9, 57. doi: 10.1186/s41200-016-0067-9
Creed J. C. and Samper-Villarreal J. (2019). Clarification of the nomenclature of the seagrass Halophila baillonii Ascherson. Aquat. Bot. 154, 42–44. doi: 10.1016/j.aquabot.2019.01.002
Dawes C. J., Phillips R. C., and Morrison G. (2004). Seagrass Communities of the Gulf Coast of Florida: Status and Ecology (St. Petersburg, FL: Florida Fish and Wildlife Conservation Commission Fish and Wildlife Research Institute and the Tampa Bay Estuary Program), iv + 74.
De Fouw J., Govers L. L., Van de Koppe J., Van Belzen J., Dorigo W., Sidi Cheikh M. A., et al. (2016). Drought, mutualism breakdown, and landscape-scale degradation of seagrass beds. Curr. Biol. 26, 1051–1056. doi: 10.1016/j.cub.2016.02.023
De los Santos C. B., Scott A., Arias-Ortiz A., Jones B., Kennedy H., Mazarrasa I., et al. (2020). “Seagrass ecosystem services: Assessment and scale of benefits,” in Out of the Blue: The Value of Seagrasses to the Environment and to People. Eds. Potouroglou M., Grimsditch G., Weatherdon L., and Lutz S. (United Nations Environment Programme, Nairobi), 19–21.
Den Hartog C. (1964). An approach to the taxonomy of the sea-grass genus Halodule Endl. (Potamogetonaceae). Blumea 12, 289–312.
Den Hartog C. (1970). The sea-grasses of the world (Amsterdam, London: North Holland Publishing Company).
Den Hartog C. and Triest L. (2020). A profound view and discourse on the typification and status of three confused taxa: Ruppia maritima, R. spiralis and R. cirrhosa. Botanica Marina. 63, 229–239. doi: 10.1515/bot-2019-0045
Den Hartog C., Van Tussenbroek B. I., Wong J. G. R., Ruaro P. M., and Guzmán J. M. (2016). A new Ruppia from Mexico: Ruppia mexicana n. sp. Aquat. Bot. 131, 38–44. doi: 10.1016/j.aquabot.2016.02.005
Dilipan E., Lucas C., Papenbrock J., and Thangaradjou T. (2018). Tracking the phylogeny of seagrasses: inferred from 18S rRNA gene and ancestral state reconstruction of morphological data. Proc. Natl. Acad. Sci. India. Sect. B.: Biol. Sci. 88, 497–504. doi: 10.1007/s40011-016-0780-5
Dunic J. C., Brown C. J., Connolly R. M., Turschwell M. P., and Côté I. M. (2021). Long-term declines and recovery of meadow area across the world’s seagrass bioregions. Global Change Biol. 27, 4096–4109. doi: 10.1111/gcb.15684
Duval-Diop D. (2016). “The West African regional network of marine protected areas: An effective tool for bridging the gap between two international environmental frameworks,” in Marine Transboundary Conservation and Protected Areas. Ed. Mackelworth P. (Routledge, Oxon), 180–197.
Eiseman N. J. (1973). Halophila johnsonii in the Indian River Lagoon, Florida, Technical Report (Inc: Harbor Branch Foundation).
Eiseman N. J. and McMillan C. (1980). A new species of seagrass, Halophila johnsonii, from the Atlantic coast of Florida. Aquat. Bot. 9, 15–19. doi: 10.1016/0304-3770(80)90003-0
Ellison A. M. and Farnsworth E. J. (1996). Anthropogenic disturbance of caribbean mangrove ecosystems: past impacts, present trends, and future predictions. Biotropica 28, 549–565. doi: 10.2307/2389096
Federal Register (2022). Endangered and threatened wildlife and plants; removal of Johnson’s seagrass from the Federal List of Threatened and Endangered Species including the corresponding designated critical habitat. Available online at: https://www.federalregister.gov/d/2022-08029 (Accessed June 30, 2025).
Fernald M. L. and Wiegand K. M. (1914). The genus Ruppia in eastern North America. Rhodora 16, 119–127.
Ferrer-Gallego P. P. and Boisset F. (2020). A revision of the typification of some names in the seagrass genera Amphibolis, Cymodocea, Halodule and Syringodium (Cymodoceaceae). Willdenowia 50, 173–186. doi: 10.3372/wi.50.50203
Fonseca M. S. and Olsen J. L. (2016). “The enigmatic seagrass: the complex and emerging story of Johnson’s seagrass,” in Environment Coastal & Offshore Magazine (Technology Systems Corporation, Stuart, Florida), 20–33.
Fonseca M., Whitfield P. E., Kelly N. M., and Bell S. S. (2002). Modeling seagrass landscape pattern and associated ecological attributes. Ecol. Appl. 12, 218–237. doi: 10.1890/1051-0761(2002)012[0218:MSLPAA]2.0.CO
Fourqurean J. W., Boyer J. N., Durako M. J., Hefty L. N., and Peterson B. J. (2003). Forecasting responses of seagrass distributions to changing water quality using monitoring data. Ecol. Appl. 13, 474–489. doi: 10.1890/1051-0761
Fourqurean J. W., Manuel S. A., Coates K. A., Massey S. C., and Kenworthy W. J. (2019). Decadal monitoring in Bermuda shows a widespread loss of seagrasses attributable to overgrazing by the green sea turtle Chelonia mydas. Estuaries. Coasts. 42, 1524–1540. doi: 10.1007/s12237-019-00587-1
Fourqurean J. W. and Rutten L. M. (2004). The impact of Hurricane Georges on soft-bottom, back reef communities: site-and species-specific effects in south Florida seagrass beds. Bull. Mar. Sci 75, 239–257.
Freitas R., Romeiras M., Silva L., Cordeiro R., Madeira P., González J. A., et al. (2019). Restructuring of the ‘Macaronesia’ biogeographic unit: A marine multi-taxon biogeographical approach. Sci. Rep. 9, 15792. doi: 10.1029/2018JC014264
Gelber A., Deis D, and Precht W. F. (2000). The distributional ecology of the seagrass Halophila johnsonii (Tallahassee, FL: Florida Dept. Transportation, Environmental Management Office).
Gorman D., Turra A., Bergstrom E. R., and Horta P. A. (2016). Population expansion of a tropical seagrass (Halophila decipiens) in the southwest Atlantic (Brazil). Aquat. Bot. 132, 30–36. doi: 10.1016/j.aquabot.2016.04.002
Hall M. O., Furman B. T., Merello M., and Durako M. J. (2016). Recurrence of Thalassia testudinum seagrass die-off in Florida Bay, USA: initial observations. Mar. Ecol. Prog. Ser. 560, 243–249. doi: 10.3354/meps11923
Hemminga M. A. and Duarte C. M. (2000). Seagrass ecology (Cambridge, UK: Cambridge University Press).
Herrera-Silveira J., Morales-Ojeda S. M., Mendoza-Martínez J. E., Medina-Gómez I., Ramirez-Ramirez J., Sandoval-Gil J. M., et al. (2019). Pastos marinos. Programa mexicana del carbono: Estado del ciclo del carbono: Agenda azul y verde. Texcoco, Mexico. p. 150–177.
Hutchings L., van der Lingen C. D., Shannon L. J., Crawford R. J. M., Verheye H. M. S., Bartholomae C. H., et al. (2009). The Benguela Current: An ecosystem of four components. Prog. Oceanogr. 83, 15–32. doi: 10.1016/j.pocean.2009.07.046
Hyndes G. A., Heck K. L. Jr., Vergés A., Harvey E. S., Kendrick G. A., Lavery P. S., et al. (2017). Accelerating tropicalization and the transformation of temperate seagrass meadows. Bioscience 66, 938–948. doi: 10.1093/biosci/biw111
Hyndes G. A., Jarvis J. C., and Heck K. L. Jr. (2022). Tropicalization in seagrasses: Shifts in ecosystem function. Front. Mar. Sci 9. doi: 10.3389/fmars.2022.1091366
Ito Y., Ohi-Toma T., Murata J., and Tanaka N. (2010). Hybridization and polyploidy of an aquatic plant, Ruppia (Ruppiaceae), inferred from plastid and nuclear DNA phylogenies. Am. J. Bot. 97, 1156–1167. doi: 10.3732/ajb.0900168
Ito Y., Ohi-Toma T., Murata J., and Tanaka N. (2013). Comprehensive phylogenetic analyses of the Ruppia maritima complex focusing on taxa from the Mediterranean. J. Plant Res. 126, 753–762. doi: 10.1007/s10265-013-0570-6
Ito Y., Ohi-Toma T., Murata J., Tanaka N., and Muasya A. M. (2015). Phylogeny of ruppia (Ruppiaceae) revisited: molecular and morphological evidence for a new species from western cape, South Africa. Syst. Bot. 40, 942–949. doi: 10.1600/036364415X689988
Ito Y. and Tanaka N. (2011). Hybridisation in a tropical seagrass genus, Halodule (Cymodoceaceae), inferred from plastid and nuclear DNA phylogenies. Telopea 13, 219–231. doi: 10.7751/telopea20116015
IUCN (2010). Available online at: https://www.iucnredlist.org/search?query=seagrass&searchType=species (Accessed August 12, 2025).
IUCN (2012). IUCN Red List Categories and Criteria: Version 3.1. 2nd ed. (Gland, Switzerland and Cambridge, UK: IUCN).
Iverson R. L. and Bittaker H. F. (1986). Seagrass distribution and abundance in eastern Gulf of Mexico waters. Estuarine. Coast. Shelf. Sci 22, 577–602. doi: 10.1016/0272-7714(86)90015-6
Jadot C. (2016). Environmental Conservation in Saint Barthélemy — Current knowledge and research recommendations (Bronx, NY, USA: Wildlife Conservation Society).
Klenz T., Dengler M., and Brandt P. (2018). Seasonal variability of the Mauritania Current and hydrography at 18 N. J. Geophys. Res.: Oceans. 123, 8122–8137. doi: 10.1029/2018JC014264
Koch E. W. and Seeliger U. (1988). Germination ecology of two Ruppia maritima L. populations in Southern Brazil. Aquat. Bot. 31, 321–327. doi: 10.1016/0304-3770(88)90020-4
Kuo J. (2020). Taxonomy of the genus Halophila Thouars (Hydocharitaceae): a review. Plants 9, 1732. doi: 10.3390/plants9121732
Kuo J. and Den Hartog C. (2001). “Seagrass taxonomy and identification key,” in Global seagrass research methods. Eds. Short F. T. and Coles R. G. (Elsevier, Amsterdam, The Netherlands), 31–58.
Kurniawan F., Digdo A. A., Darus R. F., Anggraini N. P., Ismet M. S., Wicaksono P., et al. (2024). First record of Ruppia brevipedunculata in Indonesia. Aquat. Bot. 195, 103806. doi: 10.1016/j.aquabot.2024.103806
Liu S. Y. V., Kumara T. P., and Hsu C.-H. (2020). Genetic identification and hybridization in the seagrass genus Halophila (Hydrocharitaceae) in Sri Lankan waters. PeerJ 8, e10027. doi: 10.7717/peerj.10027
Lizcano-Sandoval L., Anastasiou C., Montes E., Raulerson G., Sherwood E., and Muller-Karger F. E. (2022). Seagrass distribution, areal cover, and changes, (1990–2021) in coastal waters off West-Central Florida, USA. Estuarine. Coast. Shelf. Sci 279, 108134. doi: 10.1016/j.ecss.2022.108134
Mace G. M., Collar N. J., Gaston K. J., Hilton-Taylor C. R. A. I. G., Akçakaya H. R., Leader-Williams N. I. G. E. L., et al. (2008). Quantification of extinction risk: IUCN’s system for classifying threatened species. Conserv. Biol. 22, 1424–1442. doi: 10.1111/j.1523-1739.2008.01044.x
Martínez-Garrido J., Creed J. C., Martins S., Almada C. H., and Serrão E. A. (2017). First record of Ruppia maritima in West Africa supported by morphological description and phylogenetic classification. Botanica Marina. 60, 583–589. doi: 10.1515/bot-2016-0128
Martínez-Garrido J., Serrão E. A., Engelen A. H., Cox C. J., García-Murillo P., and González-Wangüemert M. (2016). Multilocus genetic analyses provide insight into speciation and hybridization in aquatic grasses, genus Ruppia. Biol. J. Linn. Soc. 117, 177–191. doi: 10.1111/bij.12666
McDonald A. M., Prado P., Heck K. L. Jr., Fourqurean J. W., Frankovich T. A., Dunton K. H., et al. (2016). Seagrass growth, reproductive, and morphological plasticity across environmental gradients over a large spatial scale. Aquat. Bot. 134, 87–96. doi: 10.1016/j.aquabot.2016.07.007
Morris L. J., Hall L. M., Jacoby C. A., Chamberlain R. H., Hanisak M. D., Miller J. D., et al. (2022). Seagrass in a changing estuary, the Indian River Lagoon, Florida, United States. Front. Mar. Sci 8. doi: 10.3389/fmars.2021.789818
Nagelkerken I. (2009). Ecological connectivity among tropical coastal ecosystems Vol. 615 (Dordrecht: Springer).
NOAA-NMFS, National Oceanic and Atmospheric Administration, National Marine Fisheries Service (2007). Endangered Species Act 5-year review Johnson’s seagrass (Halophila johnsonii Eiseman) (St. Petersburg, USA: Status Review Team), 60 pp.
Nordlund L. M., Koch E. W., Barbier E. B., and Creed J. C. (2016). Seagrass ecosystem services and their variability across genera and geographical regions. PloS One 11, e0163091. doi: 10.1371/journal.pone.0163091
Novelo A. R. (1991). Ruppia didyma (Potamogetonaceae) in Mexico and the antilles. Anales. Inst. Biol. Univers. Nacional. Autónoma. México. Serie. Bot. 62, 173–180.
O’Brien K. R., Waycott M., Maxwell P., Kendrick G. A., Udy J. W., Ferguson A. J., et al. (2018). Seagrass ecosystem trajectory depends on the relative timescales of resistance, recovery and disturbance. Mar. pollut. Bull. 134, 166–176. doi: 10.1016/j.marpolbul.2017.09.006
Oliveira Filho E. C., Pirani J. R., and Giulietti A. M. (1983). The Brazilian seagrasses. Aquat. Bot. 16, 251–267. doi: 10.1016/0304-3770(83)90036-0
Pazzaglia J., Reusch T. B., Terlizzi A., Marín-Guirao L., and Procaccini G. (2021). Phenotypic plasticity under rapid global changes: The intrinsic force for future seagrasses survival. Evol. Appl. 14, 1181–1201. doi: 10.1111/eva.13212
Pérez-Espinosa I., Gallegos-Martínez M., Ressl R., Valderrama-Landeros L., and Hernández Cárdenas G. (2019). Spatial distribution of seagrasses and submerged aquatic vegetation of los Petenes, Campeche. Terra. Digitalis. 3, 1–11. doi: 10.22201/igg.25940694.2019.2.56
Phillips R. C. (1960). “Observations on the ecology and distribution of the Florida seagrasses,” in Florida Board of Conservation Professional Paper Series 2. St. Petersburg, USA. p. 1–72.
Phillips R. C. (1967). On species of the seagrass, Halodule, in Florida. Bull. Mar. Sci 17, 672–676.
Price J. H., John D. M., and Lawson G. M. (1978). Seaweeds of the western coast of tropical Africa and adjacent islands: a critical assessment. II. Phaeophyta. Bull. Br. Museum. Natural History. Bot. 6, 87–182.
Samper-Villarreal J. (2024). Seagrasses in the Eastern Tropical Pacific: species, distribution, ecology, blue carbon, and threats. Latin. Am. J. Aquat. Res. 52, 336–349. doi: 10.3856/vol52-issue3-fulltext-3167
Samper-Villarreal J., Van Tussenbroek B. I., and Cortés J. (2018). Seagrasses of Costa Rica: from the mighty Caribbean to the dynamic meadows of the Eastern Tropical Pacific. Rev. Biol. Trop. 66, S53–S65. doi: 10.15517/rbt.v66i1.33260
Short F., Carruthers T., Dennison W., and Waycott M. (2007). Global seagrass distribution and diversity: a bioregional model. J. Exp. Mar. Biol. Ecol. 350, 3–20. doi: 10.1016/j.jembe.2007.06.012
Short F. T., Fernandez E., Vernon A., and Gaeckle J. L. (2006). Occurrence of Halophila baillonii meadows in Belize, Central America. Aquat. Bot. 85, 249–251. doi: 10.1016/j.aquabot.2006.04.001
Short F. T., Moore G. E., and Peyton K. A. (2010). Halophila ovalis in the tropical Atlantic Ocean. Aquat. Bot. 93, 141–146. doi: 10.1016/j.aquabot.2010.05.001
Short F. T., Polidoro B., Livingstone S. R., Carpenter K. E., Bandeira S., Bujang J. S., et al. (2011). Extinction risk assessment of the world’s seagrass species. Biol. Conserv. 144, 1961–1971. doi: 10.1016/j.biocon.2011.04.010
Sidi Cheikh M. A., Bandeira S., Soumah S., Diouf G., Diouf E. M., Sanneh O., et al. (2023). Seagrasses of west Africa: new discoveries, distribution limits and prospects for management. Diversity 15, 5. doi: 10.3390/d15010005
Sinclair E. A., Cambridge M. L., and Kendrick G. A. (2019). First report of hybridization in the seagrass genus Posidonia (Posidoniaceae). Aquat. Bot. 156, 10–13. doi: 10.1016/j.aquabot.2019.03.004
Sousa V. E., Silva Cortinhas M. C. F., Creed J. C., Batista M. G. S., Proietti M. C., and Copertino M. (2025). Assessing morphological variations in the seagrass genus Halodule (Cymodoceaceae) along the Brazilian coast through genetic analyses. PeerJ 13, e19038. doi: 10.7717/peerj.19038
Spalding M. D., Fox H. E., Allen G. R., Davidson N., Ferdaña Z. A., Finlayson M. A. X., et al. (2007). Marine ecoregions of the world: A bioregionalization of coastal and shelf areas. Bioscience 57, 573–583. doi: 10.1641/B570707
Stevenson L. V., Kamel S. J., Darnell K. M., Furman B. T., and Jarvis J. C. (2025). Cold tolerance in halodule wrightii across latitudinally distinct populations in the western atlantic ocean. Estuaries. Coasts. 48, 3. doi: 10.1007/s12237-024-01444-6
Strazisar T., Koch M. S., and Madden C. J. (2015). Seagrass (Ruppia maritima L.) life history transitions in response to salinity dynamics along the everglades-florida bay ecotone. Estuaries. Coasts. 38, 337–352. doi: 10.1007/s12237-014-9807-4
Tavares A. I., Assis J., Larkin P. D., Creed J. C., Magalhães K., Horta P., et al. (2023). Long range gene flow beyond predictions from oceanographic transport in a tropical marine foundation species. Sci. Rep. 13, 9112. doi: 10.1038/s41598-023-36367-y
Traganos D., Terauchi G., Glavan J., Panyawai J., Creed J., Weatherdon L., et al. (2020). “Seagrass mapping and monitoring,” in Out of the blue: The value of seagrasses to the environment and to people. Ed. Potouroglou M. (United Nations Environment Programme, Nairobi), 48–55.
Triest L., Beirinckx L., and Sierens T. (2018). Lagoons and saltwater wetlands getting more diversity: A molecular approach reveals cryptic lineages of a euryhaline submerged macrophyte (Ruppia). Aquat. Conserv. Mar. Freshw. Ecosyst. 28, 370–382. doi: 10.1002/aqc.2863
Triest L. and Sierens T. (2014). Seagrass radiation after Messinian salinity crisis reflected by strong genetic structuring and Out-of-Africa scenario (Ruppiaceae). PloS One 9, e104264. doi: 10.1371/journal.pone.0104264
Tuya F., Martínez-Pérez J., Fueyo Á., and Bosch N. E. (2024). Strong phylogenetic signal and models of trait evolution evidence phylogenetic niche conservatism for seagrasses. J. Ecol. 112, 446–460. doi: 10.1111/1365-2745.14232
UNEP-WCMC (United Nations Environment Programme World Conservation Monitoring Centre) and Short F. T. (2005). Global distribution of seagrasses UNEP-WCMC (New York). Cambridge, UK. doi: 10.34892/X6R3-D211
Van Tussenbroek B. I., Arana H. A. H., Rodríguez-Martínez R. E., Espinoza-Avalos J., Canizales-Flores H. M., González-Godoy M. G., et al (2017). Severe impacts of brown tides caused by Sargassum spp. on near-shore Caribbean seagrass communities. Mar. Pollut. Bull. 122 (1-2), 272-281. doi: 10.1016/j.marpolbul.2017.06.057
Van Tussenbroek B. I. (2011). Dynamics of seagrasses and associated algae in coral reef lagoons. Hidrobiológica 21, 293–310.
Van Tussenbroek B. I., Cortés J., Collin R., Fonseca A. C., Gayle P. M., Guzmán H. M., et al. (2014). Caribbean-wide, long-term study of seagrass beds reveals local variations, shifts in community structure and occasional collapse. PloS One 9, e90600. doi: 10.1371/journal.pone.0090600
Vera B., Collado-Vides L., Moreno C., and van Tussenbroek B. I. (2014). Halophila stipulacea (Hydrocharitaceae): a recent introduction to the continental waters of Venezuela. Caribbean. J. Sci 48, 66–70. doi: 10.18475/cjos.v48i1.a11
Villarreal Á., Gil B., Bastidas D., Winfield R., and Rodríguez J. (2014). Composición florística, ambientes y vegetación de la isla Zapara, estado Zulia, Venezuela. Boletin. del Centro. Investig. Biológicas. 48, 1–20.
Viqueira Rios R., Meyer Comas L., and Sturm P. (2012). Lajas Valley Agricultural Reserve Farm Inventory of the Guánica Lagoon and El Anegado Areas, Guánica, Puerto Rico (Ellicott City, USA: Center for Watershed Protection).
Virnstein R. W. and Hall L. M. (2009). Northern range extension of the seagrasses Halophila johnsonii and Halophila decipiens along the east coast of Florida, USA. Aquat. Bot. 90, 89–92. doi: 10.1016/j.aquabot.2008.05.007
Wagey B. and Calumpong H. (2013). Genetic analysis of the seagrass Halodule in Central Visayas, Philippines. Asian J. Biodivers. 4, 223–236. doi: 10.7828/ajob.v4i.305
Waycott M., Duarte C. M., Carruthers T. J., Orth R. J., Dennison W. C., Olyarnik S., et al. (2009). Accelerating loss of seagrasses across the globe threatens coastal ecosystems. Proc. Natl. Acad. Sci. U.S.A. 106, 12377–12381. doi: 10.1073/pnas.0905620106
Waycott M., Van Dijk K. J., Calladine A., and Bricker E. (2015). Population genetics of Halophila johnsonii Eiseman. St Petersburg, FL, USA: FWC-FWRI F2793-09-15-F (St. Petersburg, USA: National Oceanic and Atmospheric Administration, National Marine Fisheries Service).
Waycott M., Van Dijk K. J., Calladine A., Bricker E., and Biffin E. (2021). Genomics-Based Phylogenetic and Population Genetic Analysis of Global Samples Confirms Halophila johnsonii Eiseman as Halophila ovalis (R. Br.) Hook. f. Aquat. Conserv. Marineand. Freshw. Ecosyst. 8. doi: 10.3389/fmars.2021.740958
Wheeler M. E., Furman B. T., and Hall M. O. (2020). Leaf-tip morphology does not support species status for the seagrass Halodule beaudettei in Florida, USA. Gulf. Caribbean. Res. 31, SC31–SC35. doi: 10.18785/gcr.3101.13
Willette D. A., Chalifour J., Debrot A. D., Engel M. S., Miller J., Oxenford H. A., et al. (2014). Continued expansion of the trans-Atlantic invasive marine angiosperm Halophila stipulacea in the Eastern Caribbean. Aquat. Bot. 112, 98–102. doi: 10.1016/j.aquabot.2013.10.001
Winters G., Beer S., Willette D. A., Viana I. G., Chiquillo K. L., Beca-Carretero P., et al. (2020). The tropical seagrass Halophila stipulacea: reviewing what we know from its native and invasive habitats, alongside identifying knowledge gaps. Front. Mar. Sci 7. doi: 10.3389/fmars.2020.00300
Wong J. R., Márquez-Guzmán G. J., Santa-María Gallegos N. A., Sandoval Gil J. M., Vivanco Bervich M., Barba Santos M. G., et al. (2022). Los pastos marinos mexicanos: las plantas invisibles. Universidad Nacional Autónoma de México, Facultad de Ciencias. Ciudad de México, Mexico. p. 110.
WWF (2022). Living Planet Report 2022 – Building a nature positive society. Eds. Almond R. E. A., Grooten M., Juffe Bignoli D., and Petersen T. (Gland, Switzerland: WWF).
Yarbro L. A., Carlson P. R., and Johnsey E. (2024). Extensive and continuing loss of seagrasses in Florida’s Big Bend (USA). Environ. Manage. 73, 876–894. doi: 10.1007/s00267-023-01920-y
York R. A., Durako M. J., Kenworthy W. J., and Freshwater D. W. (2008). Megagametogenesis in Halophila johnsonii, a threatened seagrass with no known seeds, and the seed-producing Halophila decipiens (Hydrocharitaceae). Aquat. Bot. 88, 277–282. doi: 10.1016/j.aquabot.2007.11.005
Yu S. and Den Hartog C. (2014). Taxonomy of the genus ruppia in China. Aquat. Bot. 119, 66–72. doi: 10.1016/j.aquabot.2014.08.003
Keywords: marine angiosperms, IUCN red list of threatened species, bioregional extinction assessment, Halodule, Halophila, Ruppia, Syringodium, Thalassia
Citation: Samper-Villarreal J, Creed JC, Frade D, Furman BT, Bandeira S, El-Hacen HM, Peralta AC, Serrão EA, Vera Vegas BE and van Tussenbroek BI (2025) Updated extinction risk of seagrass species in the Tropical Atlantic Bioregion. Front. Conserv. Sci. 6:1656995. doi: 10.3389/fcosc.2025.1656995
Received: 30 June 2025; Accepted: 23 September 2025;
Published: 08 October 2025.
Edited by:
Mike van Keulen, Murdoch University, AustraliaReviewed by:
Cristina Piñeiro-Corbeira, University of A Coruña, SpainMariana Lima, University of Salford, United Kingdom
Copyright © 2025 Samper-Villarreal, Creed, Frade, Furman, Bandeira, El-Hacen, Peralta, Serrão, Vera Vegas and van Tussenbroek. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Brigitta I. van Tussenbroek, dmFudHVzc0BjbWFybC51bmFtLm14; Jimena Samper-Villarreal, amltZW5hLnNhbXBlcnZpbGxhcnJlYWxAdWNyLmFjLmNy