 Francina Boas
Francina Boas Mark Zvidzai*
Mark Zvidzai*- Department of Geography Geospatial Sciences and Earth Observation, University of Zimbabwe, Harare, Zimbabwe
Introduction: African elephants (Loxodonta Africana) are keystone species whose survival is increasingly threatened by habitat loss, climate change, and human-elephant conflict. Understanding their habitat preferences and how these may shift under future climate conditions is vital for effective conservation planning.
Methods: This study assessed the current and future distribution of suitable habitats for African elephant in Hwange National Park, Zimbabwe, using Species Distribution Models (SDMs), including MaxEnt, Random Forest, and an Ensemble model. Presence data derived from GPS-collared elephants (2015–2018) were combined with environmental variables such as NDVI, slope, elevation, distance to water and park boundary, and 19 bioclimatic variables for the current, 2050, and 2070 periods. Variable selection was guided by multicollinearity tests (VIF < 5), and models were evaluated using metrics such as AUC and TSS.
Results: Results showed that Random Forest and ensemble models outperformed MaxEnt in predictive performance. Seasonal modeling revealed minimal shifts between wet and dry seasons, while future projections indicated a decline and fragmentation of highly suitable habitat, particularly along the southern and eastern edges of the park. Key predictors included temperature and precipitation-related variables, especially maximum temperature of the warmest month and annual precipitation.
Discussion: These findings highlight the need to apply robust species distribution modelling tools to enhance actionable conservation and spatial planning strategies that incorporate climate projections to support long term survival of the threatened elephant population in the Hwange National Park ecosystem.
1 Introduction
African elephants (Loxodonta africana) are not only the largest land mammals but also essential keystone species that sustain savanna ecosystems (Sach et al., 2019). As mega-herbivores, their activities such as browsing, debarking, and uprooting trees, shape vegetation patterns and help maintain the open landscape, which supports diverse biodiversity (Feleha, 2018). Their dung is crucial for nutrient cycling and seed dispersal, aiding plant community regeneration (Razak et al., 2020). During dry periods, elephants often dig or restore waterholes, providing critical water sources for many species (Wool, 2019). Their long migrations connect different habitats, facilitating gene flow and strengthening ecosystem resilience (Lohay et al., 2020). Due to these extensive and vital influences, elephants are regarded as ecosystem engineers, with their conservation key to preserving broader ecological health.
Despite their ecological importance, African elephants face increasing threats that have caused significant population declines across their range. Habitat fragmentation caused by agriculture, human settlement, and infrastructure development continues to shrink and isolate suitable habitats. At the same time, illegal ivory poaching remains a major cause of death, especially in regions with weak law enforcement and high ivory demand (Hauenstein et al., 2019). Human-elephant conflict is also rising, particularly in areas bordering protected zones, where elephants often come into contact with human landscapes. In some regions, political instability and limited conservation funding make the challenges worse.
Adding to these challenges is the growing threat of climate change, which is increasingly influencing the landscapes that sustain elephant populations. Shifting temperature and rainfall patterns are altering the distribution of vegetation and water sources, key drivers of elephant movement and habitat use (Beirne et al., 2020). More frequent and intense droughts can force elephants to travel further in search of resources, increasing their exposure to poaching and human-elephant conflict (Lee et al., 2022). Recent studies (Mpakairi et al., 2020; Dejene et al., 2021) predicted major changes in habitat suitability for large herbivores like elephants in southern Africa due to future climate scenarios. Such changes could render some current protected areas inadequate, reinforcing the need for forward-looking conservation planning. Addressing these threats requires integrated and adaptive management strategies that are informed by spatially explicit knowledge of the current and future distribution of suitable habitats for the African elephants, especially in dynamic landscapes like Hwange National Park and the broader Kavango-Zambezi Transfrontier Conservation Area (KAZA-TFCA).
Species Distribution Models (SDMs) have emerged as powerful tools for predicting species’ potential ranges by combining environmental variables with occurrence records. For the African and Asian elephant species, SDMs have been widely applied to delineate suitable habitats, identify movement corridors, and forecast distributional shifts under scenarios of climate and land-use change (Robillard et al., 2015; Abir et al., 2025; Mpakairi et al., 2020). They have also been used to assess seasonal habitat use, inform conservation prioritisation, and guide strategies to mitigate human–elephant conflict (Thant et al., 2023; Yu et al., 2024). Methodological approaches such as MaxEnt (Elith et al., 2011), Random Forest (Searle et al., 2022), and ensemble modelling (Dejene et al., 2021; Black et al., 2024) have proven effective in capturing complex habitat–environment relationships across diverse landscapes. However, most applications have emphasised static or broad-scale predictions, with relatively limited attention to fine-scale temporal dynamics that shape elephant distribution. Understanding seasonal variability in suitable habitats is particularly important for African elephants, given their high mobility, dependence on shifting water and forage resources, and frequent interactions with human-dominated landscapes. Moreover, reliance on single algorithms often introduces methodological bias and underestimates predictive uncertainty. To address these limitations, there is a critical need for studies that integrate a suite of SDMs, which combine the complementary strengths of multiple algorithms, thereby improving predictive accuracy, robustness, and ecological interpretability. This study responds to this gap by applying an ensemble of SDMs to capture both the spatial and seasonal dynamics of African elephant distribution in the Hwange ecosystem, offering deeper insights into their adaptive responses to environmental variability and anthropogenic pressures.
Building on these insights, this study aims to advance our understanding of African elephant distribution by integrating a suite of SDMs to capture both spatial and seasonal dynamics of habitat suitability. Specifically, we (i) identify the environmental factors shaping elephant distribution, (ii) model the spatial distribution of suitable habitats across the study landscape, and (iii) examine how habitat suitability varies between wet and dry seasons and how the projected climate change scenarios are likely to impact on the distribution of suitable habitats. By employing multiple modelling algorithms within an ensemble framework, the study reduces methodological bias and enhances predictive reliability. We hypothesise that (1) elephant distribution is strongly influenced by the availability of water, vegetation productivity, and anthropogenic pressures, and (2) suitable habitats differ significantly between seasons, reflecting elephants’ adaptive responses to fluctuating resource availability and (3), elephant suitable habitats are expected to shrink in response to projected climate change scenarios. Together, these objectives aim to generate actionable and fine-scale insights that support long-term elephant conservation by anticipating habitat shifts and identifying priority areas for management.
2 Materials and methods
2.1 Study area
Hwange National Park (HNP), in north-western Zimbabwe (Figure 1), is the country’s largest protected area, spanning approximately 14,651 km² (Mpakairi et al., 2020). As part of the Kavango-Zambezi Transfrontier Conservation Area (KAZA TFCA), it plays a key role in supporting Zimbabwe’s elephant population. The park’s semi-arid savanna ecosystem experiences a wet season (November–April) with 570–650 mm of rainfall and a dry season (May–October) marked by scarce water availability (Mlambo et al., 2024). Due to its sandy soils and lack of permanent water sources, wildlife relies heavily on artificial waterholes, especially during the dry season. Situated in a climate-sensitive region, HNP is vulnerable to changing rainfall patterns and droughts, making it a critical area for assessing elephant habitat suitability under current and future climate scenarios.
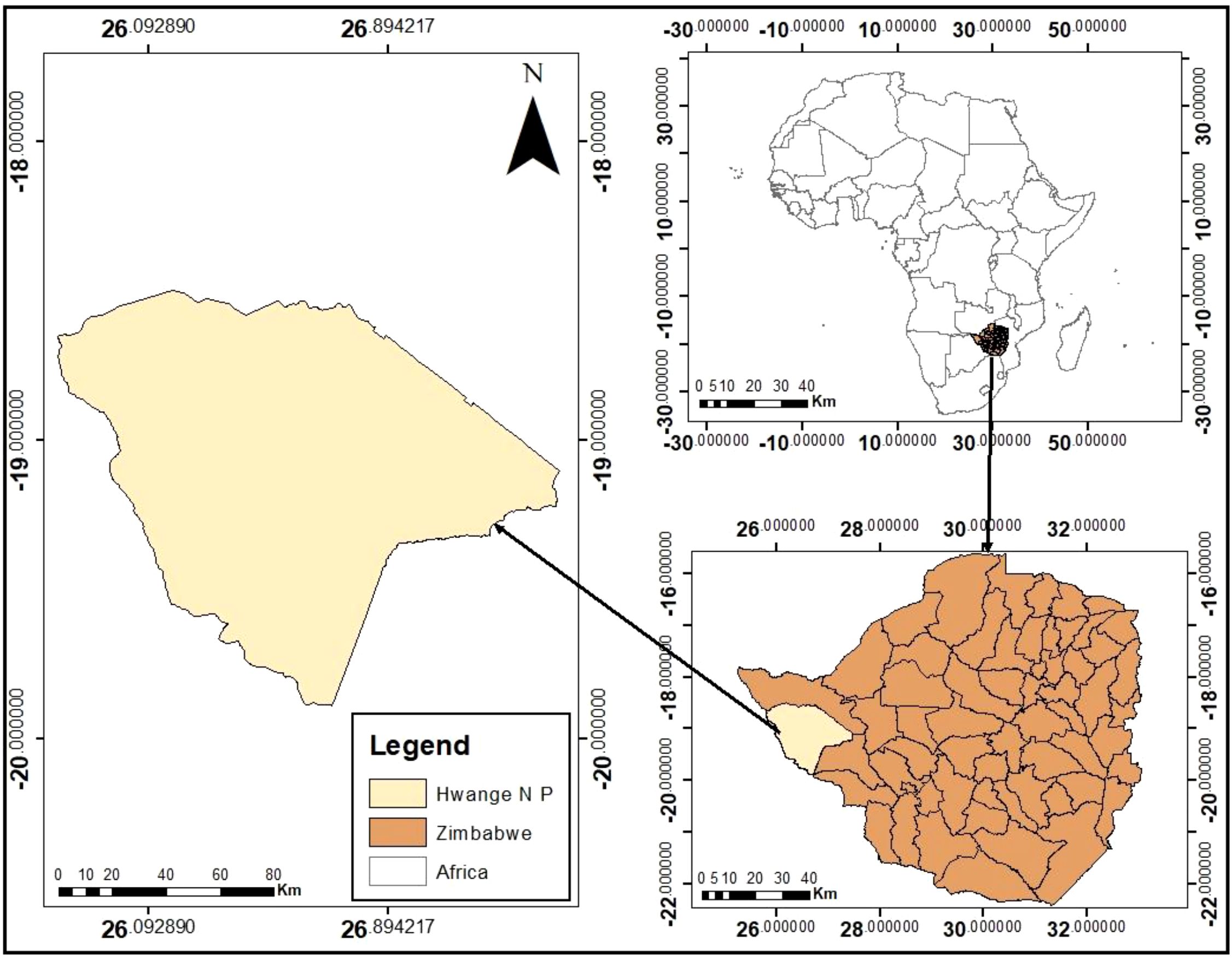
Figure 1. Location of hwange national park.
2.2 Elephant presence data
This study utilised GPS tracking data collected from African elephants (Loxodonta africana) in Hwange National Park between December 2015 and April 2018 (Mlambo et al., 2024). A total of 10 elephants (nine females and one male), (Table 1), were fitted with GPS collars, ensuring representation from different social herds to account for potential behavioral and spatial variability. The raw GPS dataset initially contained 47,500 presence records, which were subsequently cleaned and processed in ArcGIS 10.1 to remove errors, outliers, and duplicate entries, resulting in a refined dataset of 30,000 high-quality presence points.
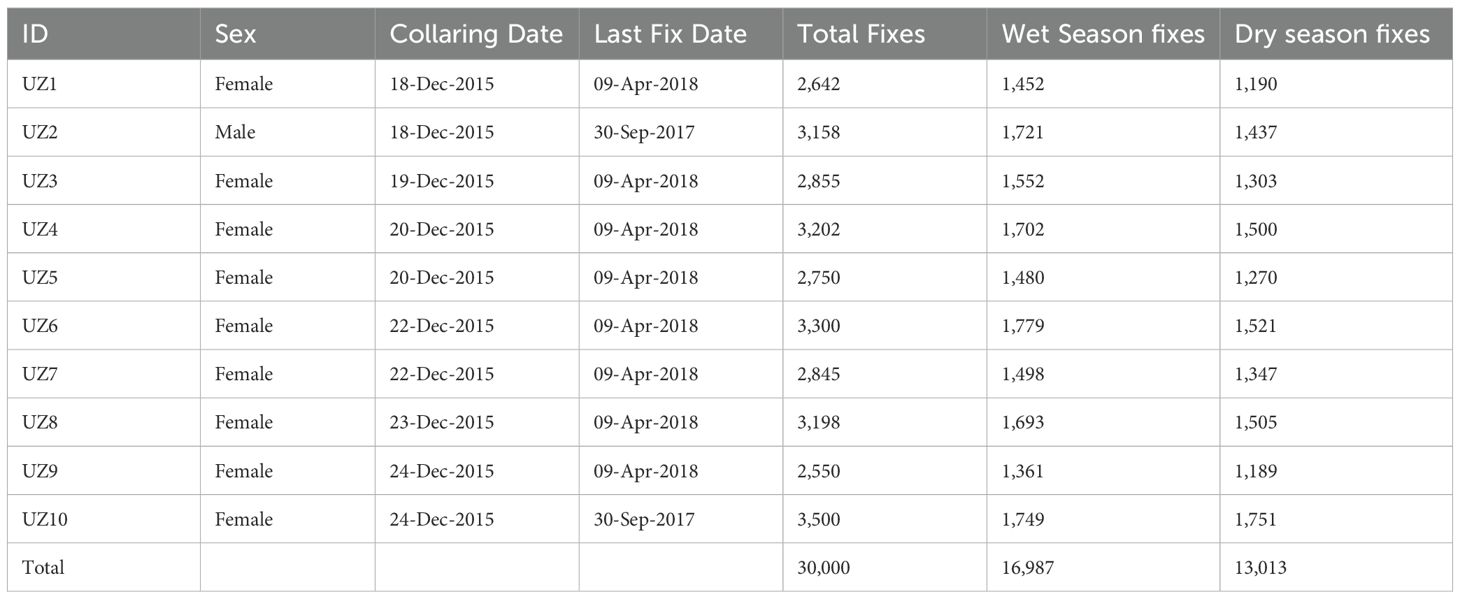
Table 1. Details of collared elephants showing elephant ID, sex, collaring and last fix dates, and the number of presence data points.
2.3 Environmental variables
To model elephant habitat suitability, a suite of environmental variables representing key ecological drivers (Table 2) were compiled. For seasonal analyses (wet and dry seasons), the study focused on seven biologically relevant predictors: mean temperature, precipitation, euclidean distance to artificial waterholes, NDVI as a proxy for vegetation productivity, elevation, slope, and distance to park boundary to assess edge effects (Dzinotizei, 2018; Ndaimani, 2018). These variables were selected based on their known influence on elephant movement and resource selection in semi-arid savannas (Dejene et al., 2021) For current distribution modelling using the combined dataset, the climate variables were expanded to include all 19 bioclimatic parameters from WorldClim while retaining the same topographic, vegetation and anthropogenic variables (Dejene et al., 2021). For the 2050 and 2070 climate projections, the study only focused on the 19 bioclimatic variables as they best capture climate-related impacts on elephant habitats. All environmental rasters were standardised and resampled to a common spatial resolution (500m) in QGIS.
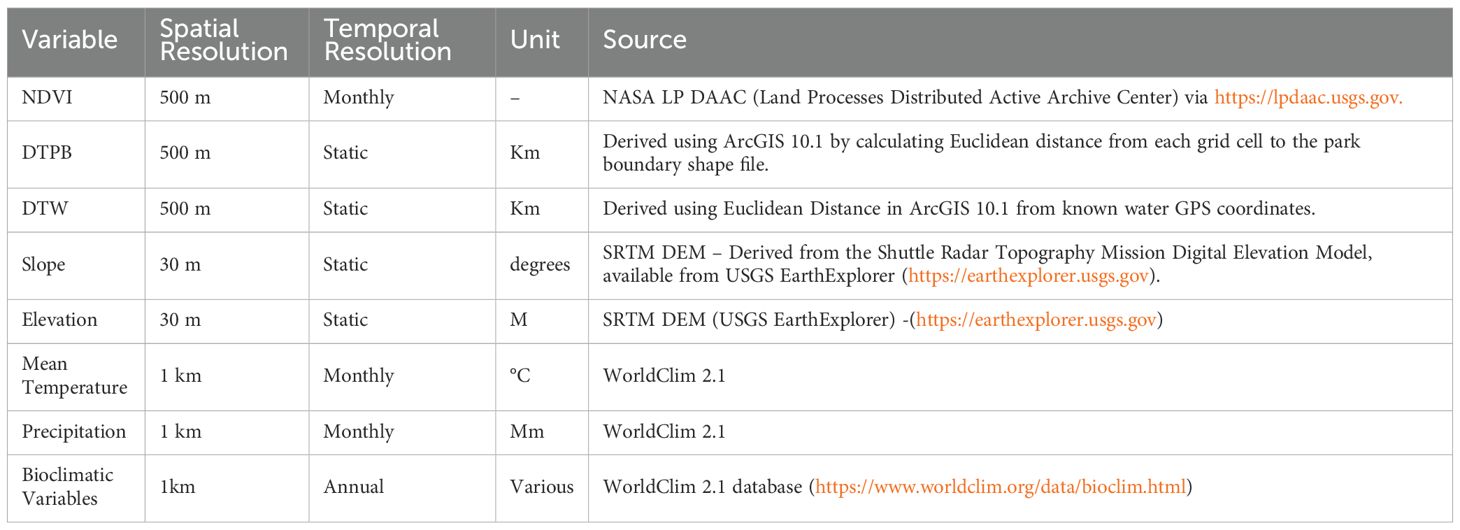
Table 2. Predictor variables used to model elephant distribution, showing spatial and temporal resolution, units of measurement and the sources.
2.3.1 Normalised difference vegetation index
NDVI served as a key indicator of vegetation productivity and forage availability for elephants in Hwange National Park (Mukomberanwa et al., 2024). Calculated from MODIS Terra (MOD13A1) satellite imagery at 500m resolution, NDVI values range from -0.1 (bare soil) to 0.7 (dense woodland), with elephants preferentially selecting areas with values >0.3 during wet seasons (Boult et al., 2018). During the dry season, NDVI values below 0.2 force elephants to shift from grazing to browsing (Koskey, 2016).
2.3.2 Mean temperature and precipitation
Mean monthly temperature (°C) and total precipitation (mm) were obtained from WorldClim 2.1 (Fick and Hijmans, 2017). Temperature extremes (>40 °C in October) significantly influence elephant water requirements (Chamaillé-Jammes et al., 2007), while precipitation patterns determine surface water availability. Mean temperature and precipitation were selected due to their strong influence on elephant ecology. Temperature affects thermoregulation and water needs, particularly during extreme heat (Valeix et al., 2008), while precipitation governs surface water availability and vegetation productivity, both critical for elephant survival and movement patterns (Valeix, 2011).
2.3.3 Slope and elevation
To analyse slope and elevation, a 30m Digital Elevation Model (DEM) was utilised, sourced from the Shuttle Radar Topography Mission (SRTM) data, which provides comprehensive elevation data. Slope was derived from elevation data using the slope function in QGIS. Elevation and slope were included as predictor variables due to their well-documented influence on habitat selection and movement patterns in African elephants. Elevation affects vegetation distribution and surface water runoff, while slope influences the energetic cost of movement, particularly in areas with rugged terrain (Mapheto, 2023).
2.3.4 Distance to park boundary and water
Distance to the park boundary was calculated using the Euclidean Distance tool in ArcGIS 10.1. Similarly, the Euclidean Distance tool was used to generate a raster representing the distance to the nearest water points. These distance layers were included as predictor variables due to their influence on elephant movement, habitat selection, and access to critical resources (De Knegt et al., 2011). Elephants tend to select areas closer to water sources and may avoid park boundaries due to potential human conflict or disturbance, making these variables ecologically relevant for modelling habitat suitability (Thant et al., 2023).
2.4 Bioclimatic variables
To incorporate climate change projections, 19 bioclimatic variables were sourced from the WorldClim database, employing the MIROC 8.5 General Circulation Model (GCM). This model, representing a high-emission trajectory, was selected to explore potential shifts in elephant habitat suitability under future climate conditions. These variables, which summarise annual trends, seasonality, and extreme climatic conditions, were included due to their known influence on species distribution and ecological niche dynamics (Dejene et al., 2021). We applied the Representative Concentration Pathway 8.5 (RCP8.5) that represents the worst case scenario because such scenarios are preferred for conservation planning as they prepare climate change managers for extreme outcomes (Beaumont et al., 2008).
2.5 Testing for multicollinearity
To ensure the robustness of the modelling approaches, the study carefully addressed multicollinearity among environmental predictors. Using the vifstep function from the usdm package in R (Naimi, 2016), variance inflation factors (VIFs) for all candidate variables were calculated, adopting a conservative threshold of 5 to identify and remove highly correlated predictors. This process minimised redundancy among variables while preserving ecologically meaningful predictors thereby enhancing model reliability and preventing overfitting (John et al., 2024). The final set of independent variables was then used to model elephant distribution patterns, ensuring both statistical validity and biological relevance in our seasonal assessments of elephant distribution as shown in Table 3.
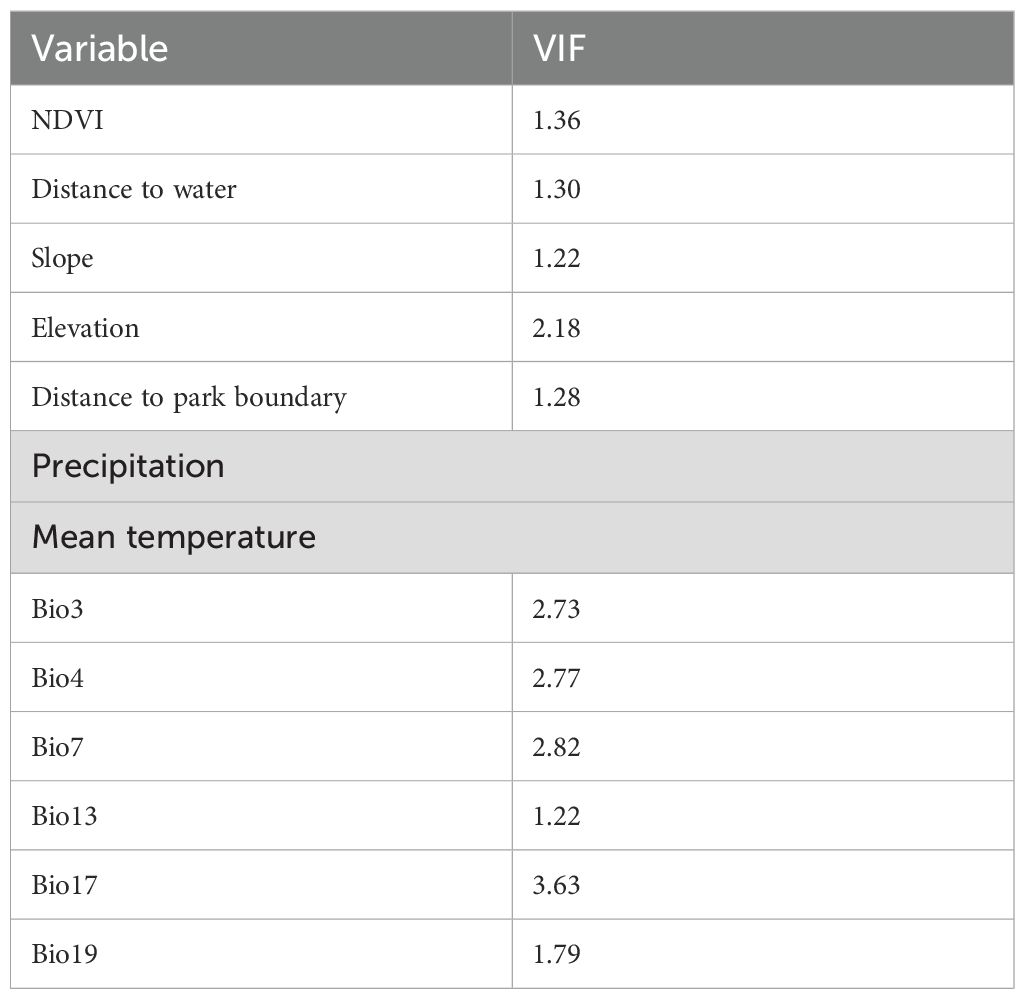
Table 3. Environmental variables that were used to estimate current elephant distribution after testing for multicollinearity.
Table 4 shows bioclimatic variables that were used to estimate potential distribution of suitable habitats under projected climate change scenarios.
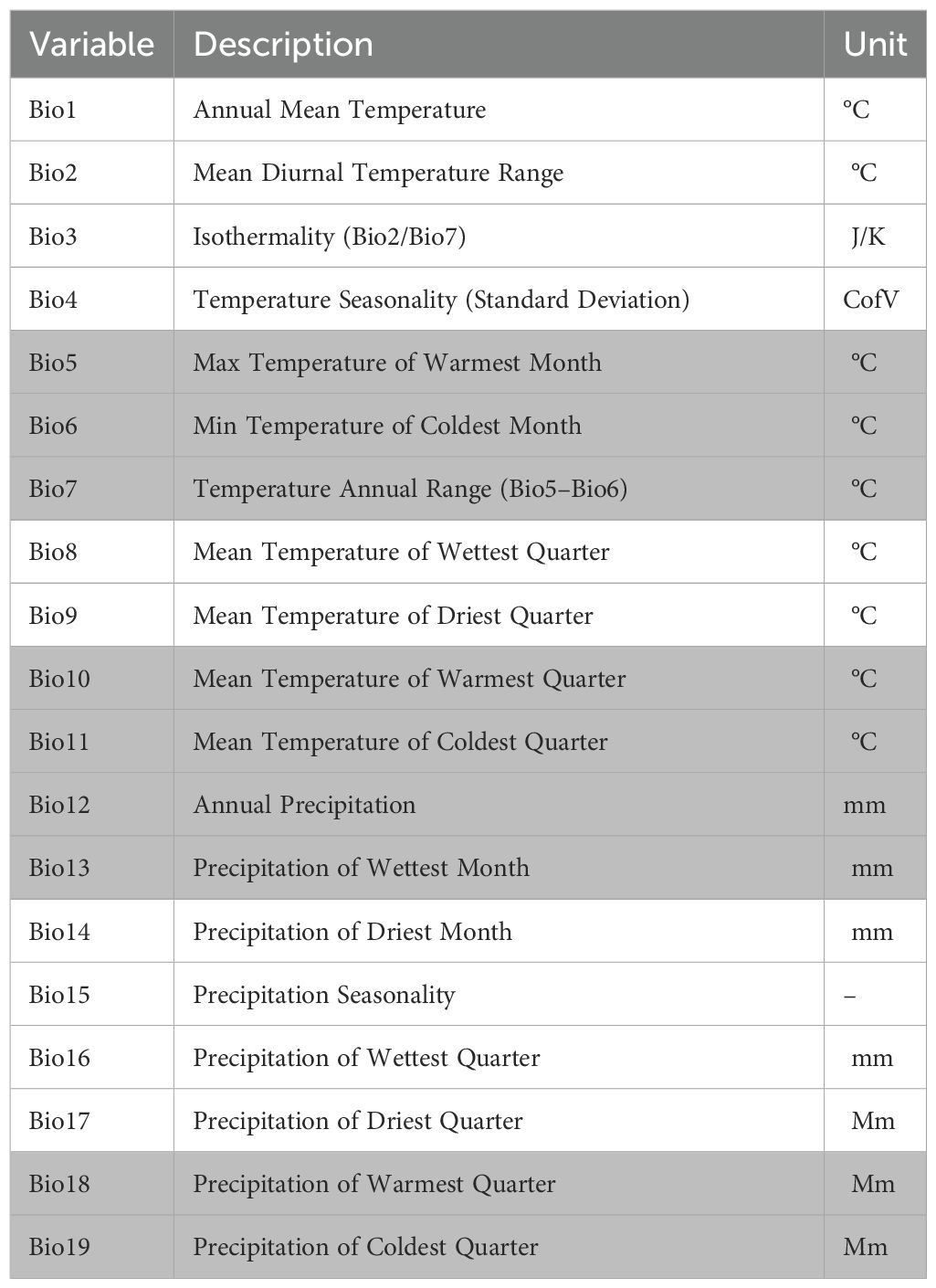
Table 4. Bioclimatic variables used to estimate elephant distribution under projected climate change scenarios. The variables shaded in gray are those used in the model after testing for multicollinearity.
2.6 Modelling approaches
This study implemented three complementary modelling approaches to predict elephant habitat suitability: Maximum Entropy (MaxEnt), Random Forest (RF), and their Ensemble, using the biomod2 package in RStudio. Each method was carefully selected to address different aspects of species distribution modelling while compensating for individual limitations.
Maximum Entropy Modelling (MaxEnt) was employed as the primary presence-background approach, particularly suitable for working with presence-only occurrence data. MaxEnt was implemented through the biomod2 interface, which provides optimised default parameters while allowing for necessary customisations. The model ran with randomly generated 10,000 background points from the study area polygon and 10 cross-validation replicates to ensure robust performance evaluation. Following recommendations from Phillips et al. (2006), hinge features were used to capture non-linear species-environment relationships while maintaining a regularisation multiplier of 1 to balance model complexity.
Random Forest (RF) modelling was implemented as the machine learning alternative, offering a different approach to capturing complex ecological relationships. Using the biomod2 framework, we configured the RF model with 1,000 trees (ntree) as recommended by Cutler et al. (2007) for ecological applications. The number of variables tried at each split was automatically determined as the square root of the total predictors, following Jin et al. (2020) original specifications.
The Ensemble model was developed to leverage the strengths of both individual modelling approaches while minimising their respective limitations. Within the biomod2 framework, predictions from MaxEnt and RF were combined using a weighted averaging approach based on model evaluation scores (Hao et al., 2020). This ensemble approach followed recommendations by Araújo and New (2007), who demonstrated that combined models typically outperform single-algorithm predictions in species distribution modelling. The ensemble process included evaluation of committee averaging and weighted mean combinations, with the final selection based on optimisation of True Skill Statistic (TSS) scores.
2.7 Model evaluation
All models were evaluated using a 70–30 split for training and testing datasets, with spatial stratification to ensure geographic representativeness. Model performance was assessed through Area Under the Curve (AUC) and True Skill Statistic (TSS), following established ecological modelling standards (Allouche et al., 2006). The Area Under the Curve (AUC) metric evaluated the models’ ability to distinguish suitable from unsuitable elephant habitats by analysing the Receiver Operating Characteristic (ROC) curve, which plots true positive rates against false positive rates across all classification thresholds. Ranging from 0 to 1, AUC values were interpreted as: 0.5 (random performance), 0.5-0.7 (poor to moderate), 0.7-0.9 (good), and ≥0.9 (excellent discrimination) (Pearce and Ferrier, 2000).
TSS values range from -1 to +1, where +1 indicates perfect agreement and values ≤0 suggest performance no better than random (Allouche et al., 2006; Sharma et al., 2020). TSS, unlike AUC, is threshold-dependent and therefore more directly relevant to conservation applications requiring binary distribution classifications. The ensemble model demonstrated superior predictive performance compared to individual models and was subsequently used for projecting habitat suitability under future climate scenarios, providing a comprehensive tool for elephant conservation planning under climate change.
2.8 Quantifying potential change in habitat suitability
After running the species distribution models, the continuous habitat suitability maps were reclassified into three distinct categories based on natural colour code breaks as our threshold for each suitability category: most suitable (0.7–1), less suitable (0.3–0.6), and unsuitable (0–0.2). The study adopted a similar approach used by Zvidzai et al. (2024), who applied categorical thresholds to interpret spatial habitat dynamics under climate change scenarios. QGIS version 3.36.3 was then used to calculate the total area (in km²) of each suitability class under current and projected climate conditions (2050 and 2070). This enabled the assessment of spatial trends in habitat suitability, including the expansion, contraction, or stability of highly suitable elephant habitats over time.
3 Results
3.1 Seasonal distribution (wet and dry season)
Results show that, during the dry season (Figure 2a), the highest concentrations in elephant movement data are observed in the northern east and eastern areas. In the wet season (Figure 2b), the northern east and eastern zones continue to act as primary elephant strongholds, while in the western and southern regions, the GPS movement data is sparse.
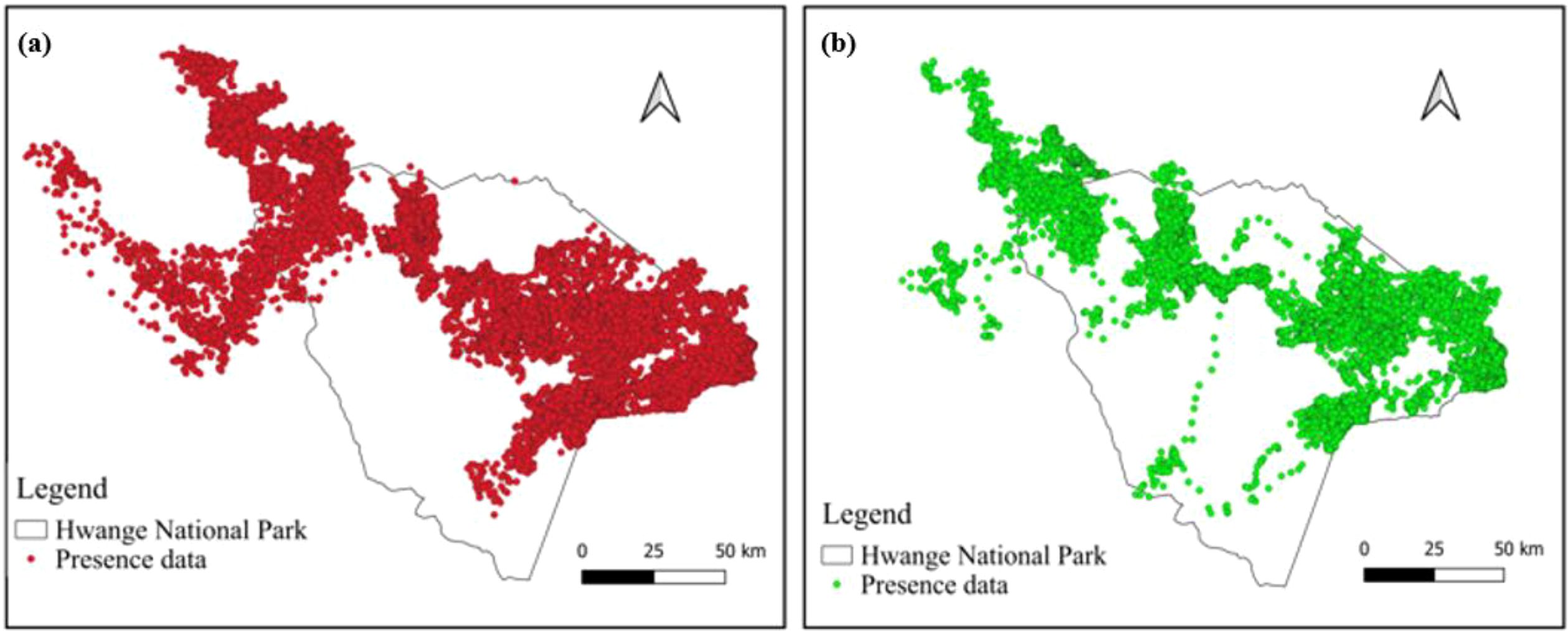
Figure 2. Spatial distribution of elephant movement data for (a) the dry season (May–October) and (b) wet season (November–April) in Hwange National Park.
3.2 Model performance comparison
In the Dry season, Ensemble (ENSE) achieved an AUC of 0.95, outperforming MaxEnt (MAX) (0.82) and Random Forest (RF) (0.91). For TSS, MaxEnt scored the highest at 0.87, followed by Ensemble (0.85) and Random Forest (0.80). Under wet season, the Ensemble model showed the best AUC performance (0.96), while Random Forest and MaxEnt scored 0.90 and 0.85, respectively. Overall, the Ensemble model demonstrates consistent and high performance across all conditions, particularly in AUC, while MaxEnt shows variability, excelling in TSS under Dry conditions but lagging in others as shown in Table 5.
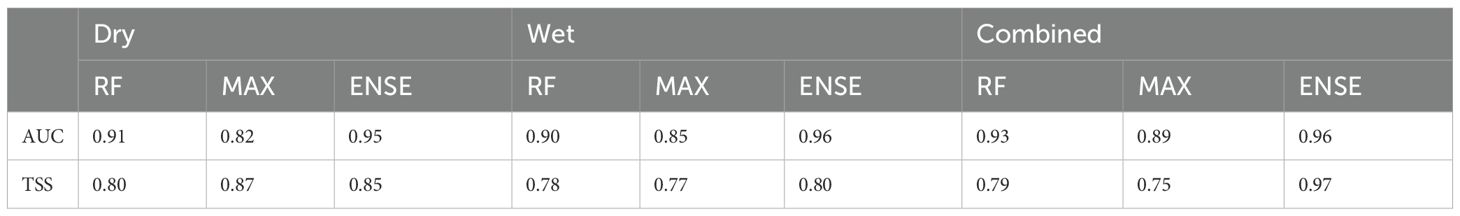
Table 5. Performance metrics for RF, MaxEnt, and Ensemble models based on the combined and wet and dry seasons elephant movement data .
3.3 Variable importance
In the wet season (Figure 3a), elevation and mean temperature are the most influential variables for all models, while slope and NDVI have the lowest impact. In the dry season (Figure 3b), mean temperature, precipitation and NDVI are the most influential with slope as the least important variable. When combining presence data from both seasons (c), NDVI continues to show the highest importance, with variables like precipitation or slope potentially ranking lower as shown in Figure 3. In the future bioclimatic variables chart (d), bioclimatic factors such as bio19, bio13, bio12 and bio5 emerged as the most significant predictors, while bio11 had the lowest influence (4%).
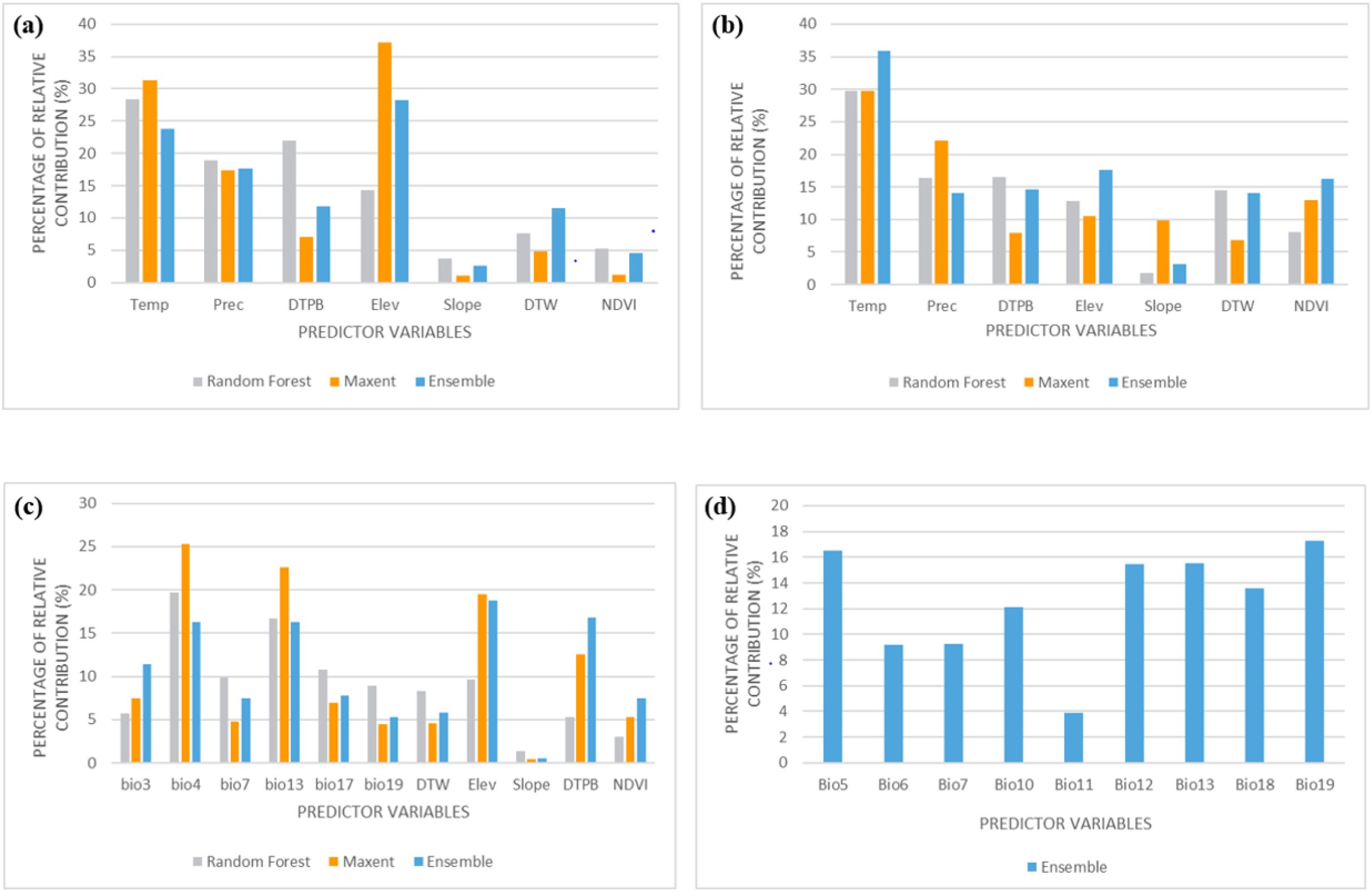
Figure 3. Variable importance for (a) wet season, (b) dry season, (c) combined presence data, and (d) bioclimatic variables used for future projections.
3.4 Modelling elephant distribution patterns based on combined elephant presence data
The MaxEnt model (Figure 4a) indicates areas that are highly suitable primarily in the northwest and central regions of Hwange National Park, with less suitable areas appearing in the southern part of the park. Similarly, the Random Forest model (Figure 4b) shows a comparable pattern but with a more compact and clearly defined distribution of less suitable habitat in the southern region. The ensemble model (Figure 4c) presents a smoother output, with extensive highly suitable areas in the northwest and central parts, while less suitable zones are mostly confined to the southern section of the park. The western part of Hwange National Park is consistently less suitable across all models.
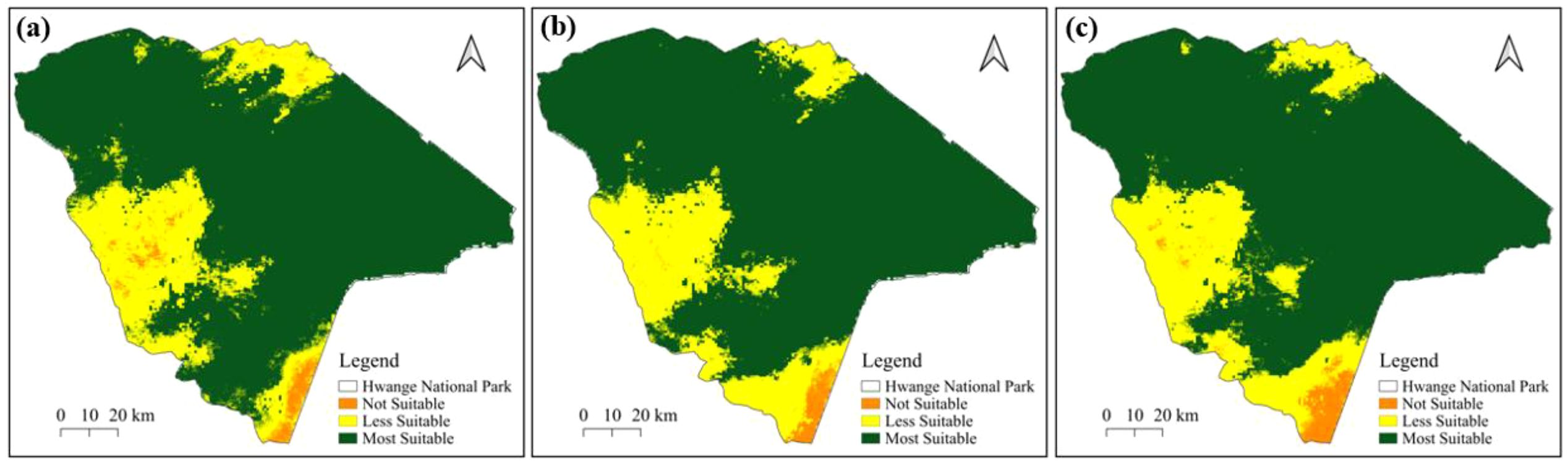
Figure 4. Potential distribution of suitable habitat for elephants in Hwange National Park for combined presence data based on (a) MaxEnt, (b), Random Forest, and (c) Ensemble models.
3.4.1 Wet season
During the wet season, results based on the MaxEnt model (Figure 5a) identify areas of high suitability primarily in the northwest and central regions of Hwange National Park, with less suitable habitats located in the southern area. The Random Forest model (Figure 5b) shows that much of the area is less suitable in the western region of the park. The ensemble model (Figure 5c), reveals extensively highly suitable areas in the northwest and eastern sections, while less suitable zones are predominantly found in the south. Notably, the eastern part of Hwange National Park is classified as suitable across all models as shown in Figure 5.
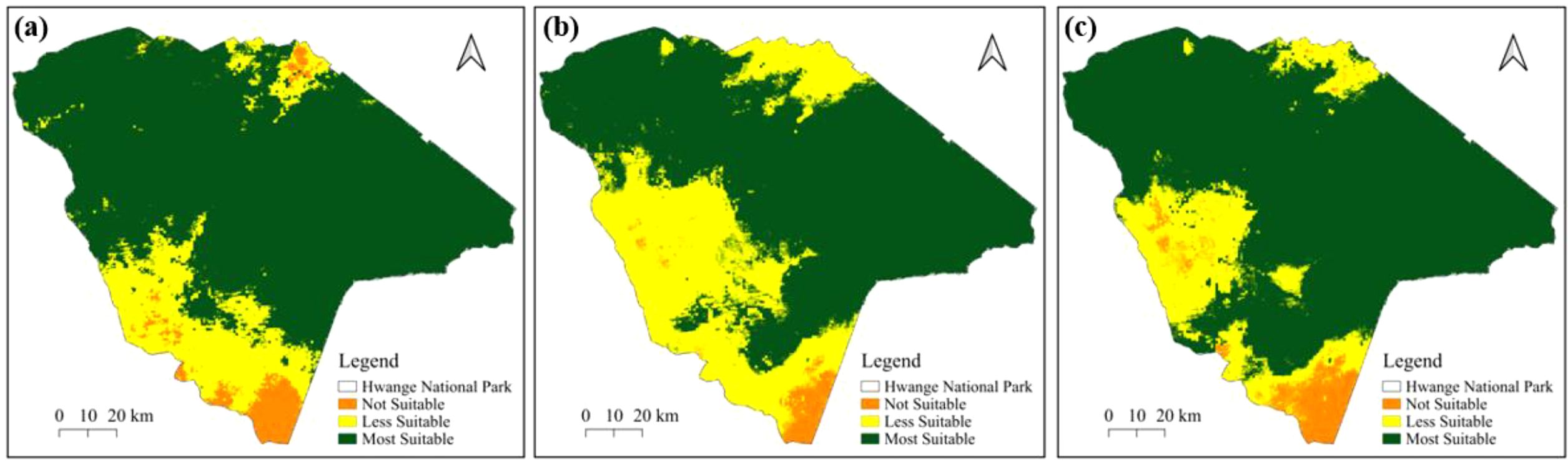
Figure 5. Potential distribution of suitable habitats for elephants in Hwange National Park for the wet season based on (a) the MaxEnt model, (b), Random Forest model and (c) the Ensemble model.
3.4.2 Dry season
For the dry season, the MaxEnt model (Figure 6a) identifies high suitability primarily in the northwest and central regions of Hwange National Park, with a larger area classified as unsuitable in the southern part. The Random Forest model (Figure 6b) reveals compact suitable areas in the eastern section of the park. Based on the ensemble model (Figure 6c), highly suitable areas are evident in the eastern part of the park, while there are patches of less suitable habitat in the northwest. Notably, the southern region of Hwange National Park shows a significantly larger area classified as unsuitable across all models as shown in Figure 6.
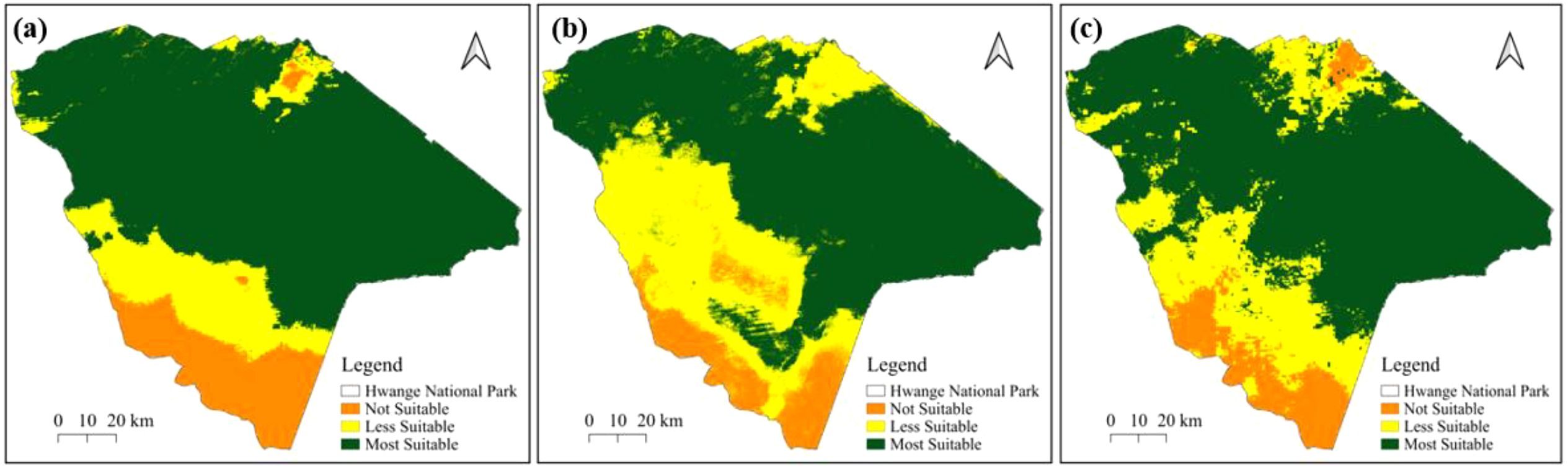
Figure 6. Potential distribution of suitable habitats of elephants in Hwange National Park for dry season based on (a) MaxEnt, (b) Random Forest and (c) Ensemble models.
3.5 Future distribution projections
Based on the current scenario, most of the landscape, especially within and around Hwange National Park, is classified as highly suitable, with less suitable and unsuitable zones mainly located in the southern and northeastern edges (Figure 7a). Projected suitable habitats for 2050 (Figure 7b), reveal a noticeable decline in suitable habitat, particularly along the park’s eastern and southern boundaries and near the KAZA edge, although core areas in Hwange remain intact. By 2070, unsuitable habitat expand further, particularly in the southern and eastern directions, while the most suitable areas become increasingly fragmented, indicating continued habitat decline (Figure 7c).
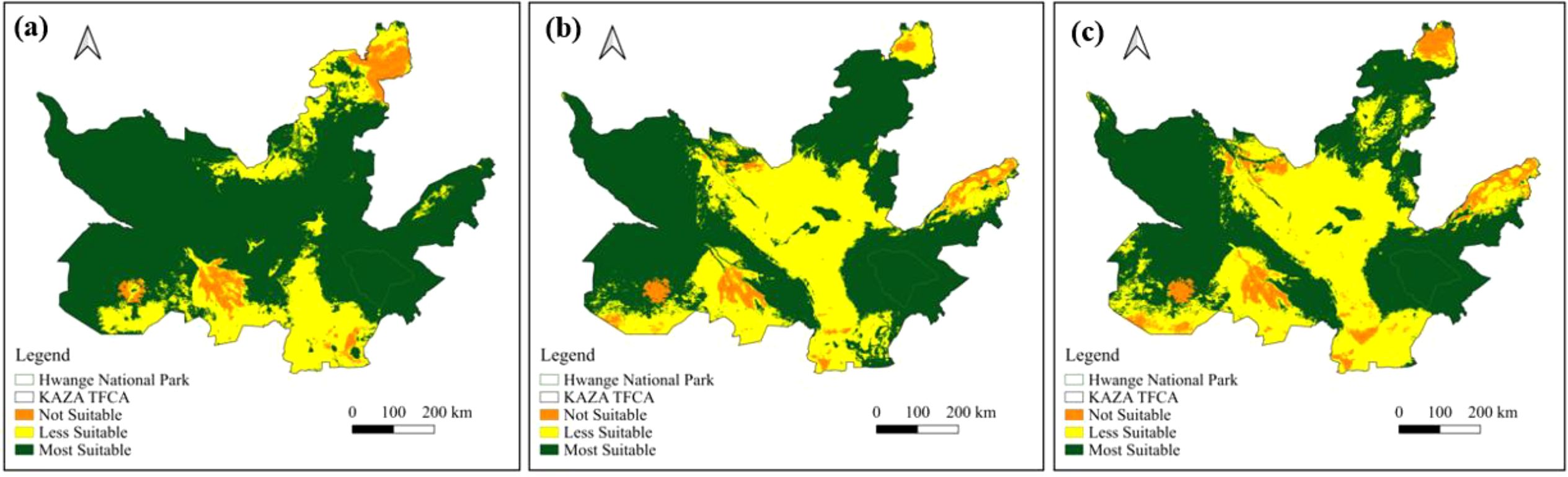
Figure 7. Potential distribution of suitable habitat for African elephants in the KAZA TFCA under different climate scenarios (a) current (2015-2018), (b) future (2050) and (c) future (2070).
3.6 Predicted range changes within the KAZA TFCA
Figure 8 shows that the most suitable elephant habitat is expected to decline from 388,489.9 km² (current) to 342,698.3 km² in 2050, and further to 282,470.3 km² by 2070. Less suitable areas increase from 107,609.7 km² to 153,210.0 km² in 2050 and reach 205,429.9 km² by 2070. Not suitable areas rise slightly from 23,900.4 km² to 24,101.7 km² in 2050 and 32,099.8 km² in 2070.
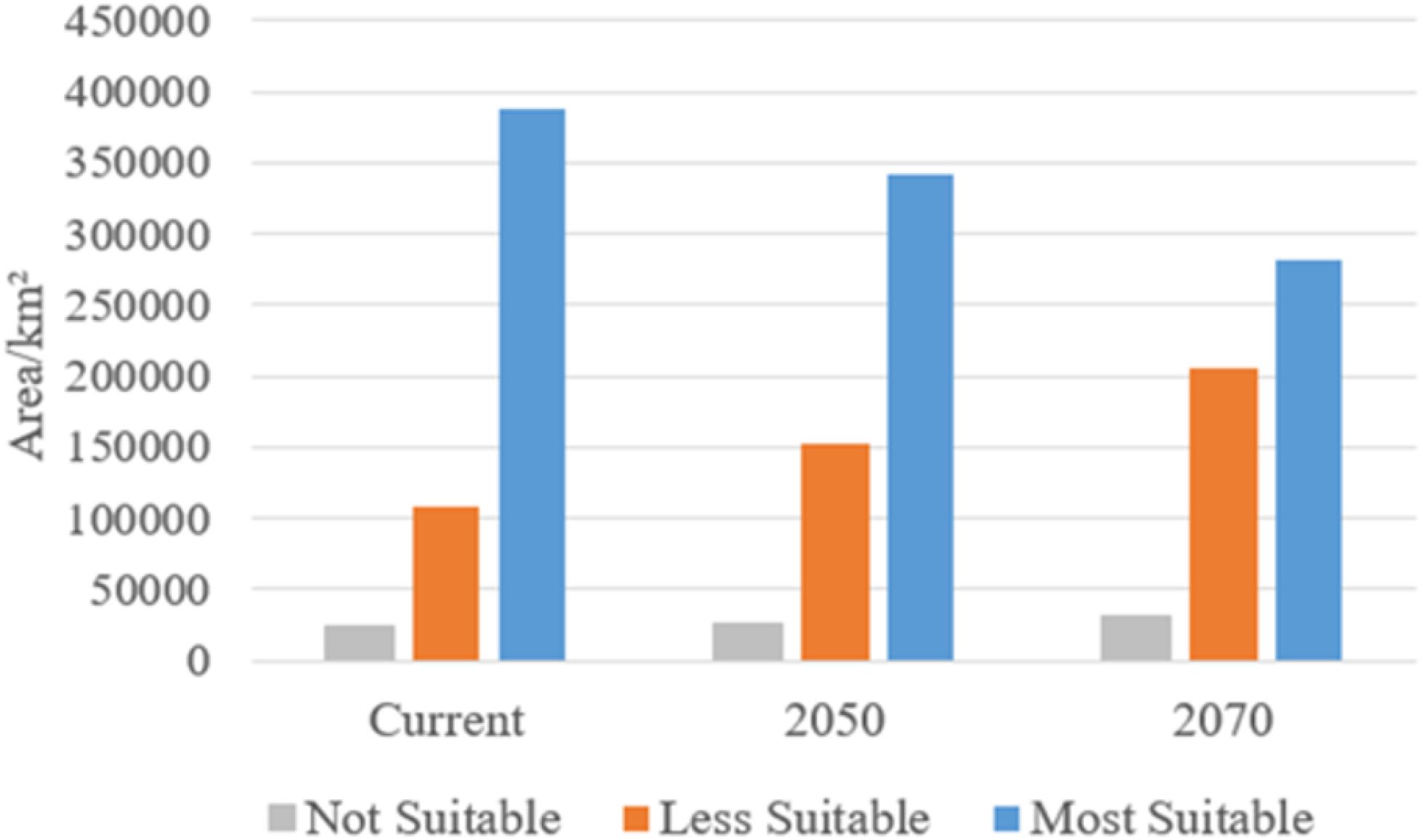
Figure 8. Predicted potential range changes for the African elephant under different climate change scenarios.
3.7 Response curves for the combined elephant presence data
Figure 9 shows how the probability of elephants presence responds to different environmental variables. Isothermality (bio3) shows an increase in African elephant presence, peaking at moderate values, after which the probability of elephant presence decreases. Precipitation patterns (bio17 and bio19) reveal distinct thresholds that indicate optimal moisture levels for the African elephant, while proximity to water significantly influences presence, with a clear decline in probability of African elephant presence as distance to water increases. Furthermore, both elevation and NDVI exhibit non-linear relationships, suggesting specific habitat preferences. The comparison of models highlights differences in predictions, with the Ensemble model often providing a more balanced response (Figure 9).
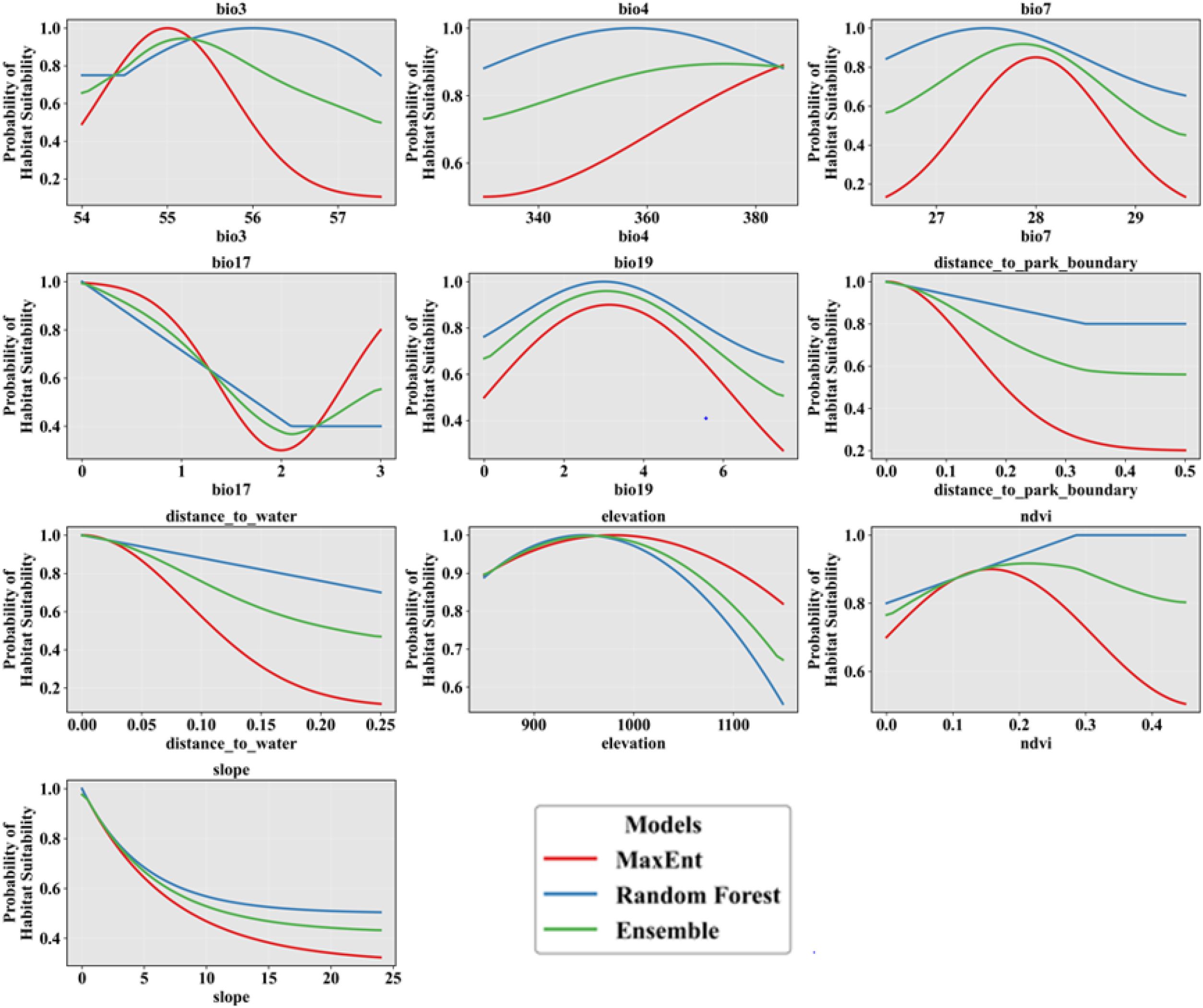
Figure 9. Response curves for combined elephant presence data.
3.8 Response curves of elephants to bioclimatic variables using ensemble modelling
Figure 10 illustrates how the probability of elephant presence response to bioclimatic variables based on the Ensemble model. The probability of African elephant presence was highest when the maximum temperature of the warmest month (bio5) was around 30 °C and declined at higher temperatures. Minimum temperature of the coldest month (bio6) and mean temperatures (bio10, bio11) showed a positive relationship, stabilising above 15 °C. The probability of elephant presence also increased as the Annual precipitation (bio12), and rainfall during the wettest month (bio13), and warmest quarter (bio18) increased. The probability of elephant presence also peaked at moderate values (70–90 mm) for precipitation of the coldest quarter (bio19), after which it started declining (Figure 10).
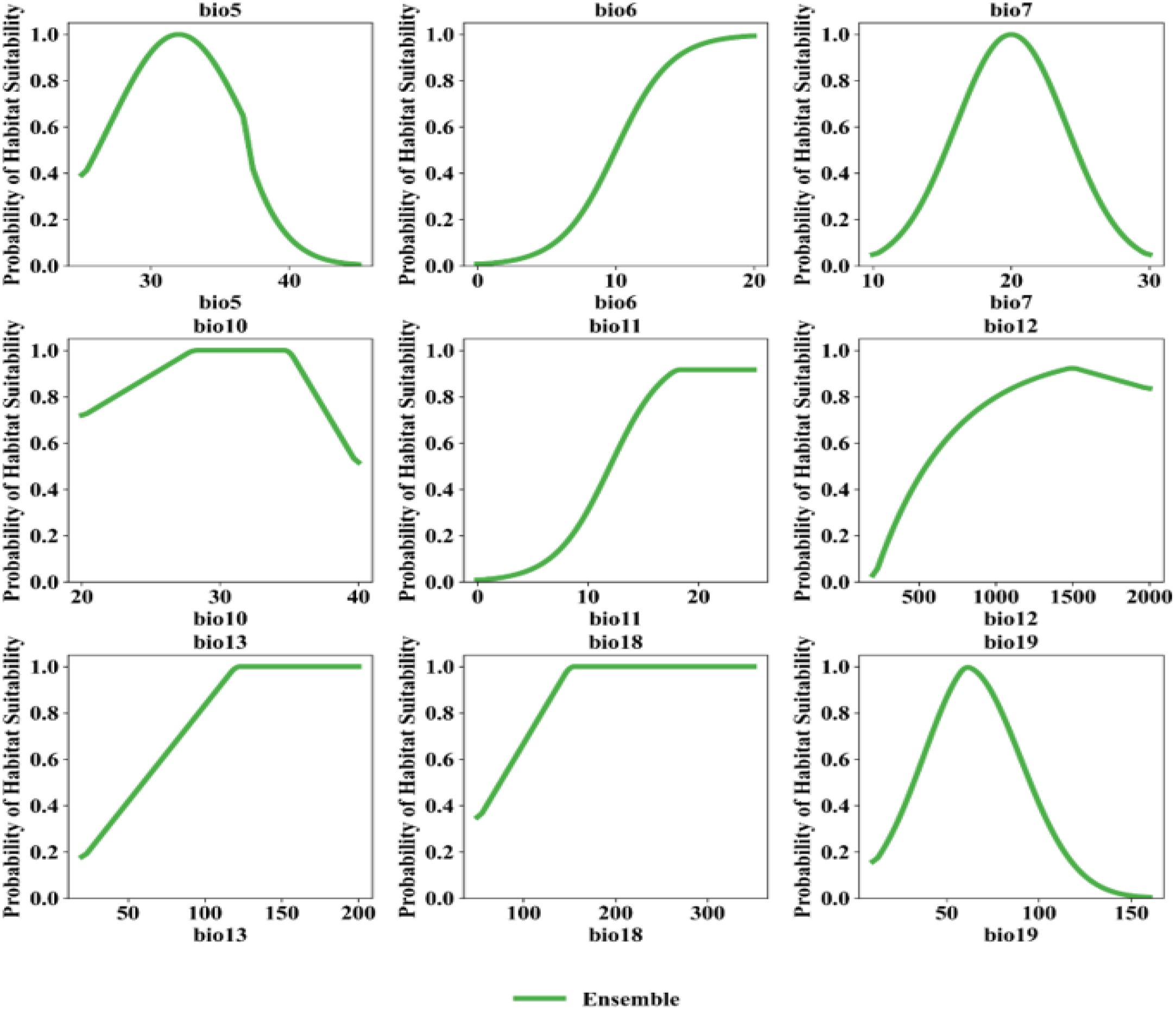
Figure 10. Response curves for African elephant for different bioclimatic variables based on the ensemble model.
4 Discussion
The main objective of this study was to apply and assess three species distribution models (Random Forest, MaxEnt, and Ensemble models) to understand current and future distribution of suitable habitats for the African elephant in Hwange National Park under climate change scenarios. Results demonstrated that all three models were capable of capturing elephant-environment relationships, with the ensemble model consistently producing more ecologically realistic spatial predictions. While Random Forest created compact and well-defined suitability areas, MaxEnt generated broader, more generalised surfaces. The ensemble model combined both strengths, resulting in maps and plots that are more balanced and aligned well with known elephant ecological behaviour. García-Callejas and Araújo (2016) found that ensemble models improve predictive accuracy and reflect ecological complexity more effectively than single algorithms. To this end, this study demonstrated the utility of integrating a suite of SDMs, which combine the complementary strengths of multiple algorithms, thereby improving predictive accuracy and conservation planning outcomes.
When elephant presence data from both seasons were combined, models consistently identified core habitat zones in the northeast, east-central, and northwest regions of the park. These zones likely represent year-round strongholds due to their association with artificial waterholes, high vegetation productivity (as indicated by NDVI), moderate elevation, and favourable topographic conditions. Previous studies found that elephants in semi-arid systems strongly prefer habitats with stable water access, moderate terrain, and productive vegetation, especially in protected areas (Chamaillé-Jammes et al., 2007; Muposhi et al., 2016). In contrast, the southwestern and far southeastern regions were consistently avoided in all models, indicating ecologically marginal or disturbed zones. This supports earlier findings by Gara et al. (2021) and Sach et al. (2019), who found that elephants tend to avoid areas with poor water and forage availability, human barriers, or landscape that are fragmented.
In the wet season, elephants expanded their range into areas typically avoided during the dry season, likely due to increased rainfall, surface water availability, and green-up of forage. Consistent with prior work, elephants were found to use ephemeral resources during wet periods, reducing their reliance on artificial waterholes (Muposhi et al., 2016; Naidoo et al., 2020). However, the northeast and east-central regions remained important core habitats even during wet months, highlighting the role of habitat fidelity and persistent environmental suitability. de Knegt (2010) found that elephants continue to revisit historically favourable areas regardless of season.
In the dry season, elephants showed a more restricted distribution, concentrating around northeastern and eastern Hwange, which are characterised by the highest densities of artificial waterholes. These locations are vital during the dry months (May–October), when natural water sources diminish. Previous studies found that artificial water provisioning determines dry-season elephant distribution in Hwange and other KAZA landscapes (Chamaillé-Jammes et al., 2007; Ochieng, 2015). Conversely, the southwestern and northwestern areas remained underused, showing low suitability. Chui et al. (2024), observed that dry season distributions become more clustered and selective around artificial water points, often resulting in local overuse and degradation.
A consistent observation across all seasons was the persistent avoidance of the southwestern part of the park. Mpakairi et al. (2020) found that elephants rarely use this region due to poor water access, low NDVI, human activity near boundaries, and historical rainfall deficits. These ecologically marginal zones may function as natural or anthropogenic boundaries limiting movement under normal conditions.
Variable importance analysis revealed season-specific drivers of habitat selection. In the wet season, elevation and mean temperature were the top predictors, while in the dry season, NDVI, mean temperature, and precipitation variables dominated. NDVI emerged as the strongest variable across all models. Previous studies found that NDVI is a reliable proxy for vegetation productivity and is often the best predictor of elephant forage availability and space use across savanna systems (Boult et al., 2018; Mpakairi et al., 2020; Mlambo et al., 2024). Also, Ndaimani (2018), demonstrated NDVI’s value in seasonal and long-term elephant habitat modelling. In general, the results reinforce that elephants respond strongly to environmental heterogeneity and seasonal availability of water and forage.
Future projections suggested a substantial contraction of suitable elephant habitat by 2050, worsening by 2070. This is consistent with one of our hypotheses. Core habitats in the north and center may persist, but the southern and eastern zones are predicted to become highly unsuitable due to increasing aridity and declining vegetation productivity. Previous studies (Gandiwa et al., 2016; Smit et al., 2020; Dejene et al., 2021) found that climate change is likely to reduce suitable ranges for large herbivores, including elephants, especially in southern Africa’s arid and semi-arid zones. This supports findings by Lu et al. (2025), who emphasised the need for landscape-level conservation strategies to maintain elephant connectivity in the face of climate extremes. These projected shifts imply that elephants may increasingly seek resources outside protected areas, potentially escalating human-elephant conflict.
Response curves further clarified key ecological thresholds. Suitability peaked at moderate maximum temperatures (15–25 °C), with steep declines at higher values, suggesting thermal sensitivity. Similarly, optimal precipitation values for habitat suitability clustered around 1250–1750 mm/year, with seasonal rainfall variables (Bio18, Bio19) playing key roles. Dunkin (2012) found that elephants are physiologically sensitive to high temperatures and require year-round water availability, making them vulnerable to climatic extremes. This shows that climate thresholds shape movement and distribution patterns for large herbivores across southern Africa’s savannas.
This study has some limitations. The GPS data were collected from only ten elephants (mostly female), which may limit the generalisability of movement patterns. Taylor et al. (2020) found that male elephants often display different ranging behaviour and habitat use compared to females. Furthermore, the monitoring period (2015–2018) was relatively short, possibly missing long-term ecological responses. This contrasts with long-term telemetry studies that found substantial interannual variation in elephant movement tied to multi-year rainfall cycles (Garstang et al., 2014). Additionally, the exclusion of anthropogenic features like roads, fences, and settlements is another limitation of the study as numerous studies found that human infrastructure strongly shapes elephant movement and habitat selection (Bastille-Rousseau et al., 2020; Gara et al., 2021; Hahn et al., 2024).
5 Conclusion
Understanding the interactions between species and their environment is essential for determining optimal habitat conditions and guiding conservation decisions. This study confirms the value of using GIS and remote sensing technologies in combination with a suit of SDMs to assess habitat suitability, especially in data-scarce regions like HNP. The integration of biophysical variables (NDVI, temperature, precipitation) with GPS data enabled the creation of ecologically meaningful seasonal and future distribution models.
Key findings revealed distinct dry and wet season spatial patterns influenced by resource availability. Ensemble SDMs provided high predictive accuracy, validating their utility in ecological forecasting. The use of multiple SDMs also provide expanded insights that can guide effective conservation planning. Crucially, future projections under the MIROC 8.5 climate scenario revealed potential contractions in suitable habitat by 2070, particularly in the southern part of HNP. This highlights the vulnerability of elephants to climate-induced habitat loss and the need for adaptive conservation planning.
The implications of these findings are significant. Management authorities may need to reconsider the spatial configuration of protected areas and artificial water provisioning to maintain ecological balance and minimise habitat degradation. The identification of future refugia and seasonal corridors is particularly urgent in the face of accelerating environmental change.
Overall, this study contributes to a growing body of work advocating for the integration of climate-smart spatial planning into biodiversity conservation. Continued research using expanded datasets, real-time satellite monitoring, and cross-border conservation frameworks will be key to ensuring the persistence of Africa’s elephants in the decades to come.
Data availability statement
The raw data supporting the conclusions of this article will be made available by the authors, without undue reservation.
Ethics statement
The animal study was approved by Zimbabwe Parks and Wildlife Management Authority. The study was conducted in accordance with the local legislation and institutional requirements.
Author contributions
MZ: Visualization, Validation, Conceptualization, Project administration, Formal Analysis, Writing – review & editing, Writing – original draft, Supervision. FB: Writing – review & editing, Formal Analysis, Writing – original draft, Methodology, Validation, Conceptualization, Investigation, Visualization, Software.
Funding
The author(s) declare that no financial support was received for the research, and/or publication of this article.
Conflict of interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Generative AI statement
The author(s) declare that no Generative AI was used in the creation of this manuscript.
Any alternative text (alt text) provided alongside figures in this article has been generated by Frontiers with the support of artificial intelligence and reasonable efforts have been made to ensure accuracy, including review by the authors wherever possible. If you identify any issues, please contact us.
Publisher’s note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
References
Abir K. A. M., Dey B., Redowan M., Haque A., and Ahmed R. (2025). Predicting suitable habitats for Asian elephant (Elephas maximus) in Tropical Asia under changing climatic scenarios. Geogr. Sustainabil. 6, 100279. doi: 10.1016/j.geosus.2025.100279
Allouche O., Tsoar A., and Kadmon R. (2006). Assessing the accuracy of species distribution models: Prevalence, kappa and the true skill statistic (TSS). J. Appl. Ecol. 43, 1223–1232. doi: 10.1111/j.1365-2664.2006.01214.x
Araújo M. B. and New M. (2007). Ensemble forecasting of species distributions. Trends Ecol. Evol. 22, 42–47. doi: 10.1016/j.tree.2006.09.010
Bastille-Rousseau G., Wall J., Douglas-Hamilton I., Lesowapir B., Loloju B., Mwangi N., et al. (2020). Landscape-scale habitat response of African elephants shows strong selection for foraging opportunities in a human dominated ecosystem. Ecography 43, 149–160. doi: 10.1111/ecog.04240
Beaumont L. J., Hughes L., and Pitman A. J. (2008). Why is the choice of future climate scenarios for species distribution modelling important? Ecol. Lett. 11, 1135–1146.
Beirne C., Meier A. C., Brumagin G., Jasperse-Sjolander L., Lewis M., Masseloux J., et al. (2020). Climatic and resource determinants of forest elephant movements. Front. Ecol. Evol. 8. doi: 10.3389/fevo.2020.00096
Black E., Boult V., Hirons L., and Woolnough S. (2024). The potential value of seasonal drought forecasts in the context of climate change: A case study of the African elephant conservation sector. Meteorol. Appl. 31, 1–15. doi: 10.1002/met.2190
Boult V. L., Quaife T., Fishlock V., Moss C. J., Lee P. C., and Sibly R. M. (2018). Individual-based modelling of elephant population dynamics using remote sensing to estimate food availability’. Ecol. Model. 21, 187–195. doi: 10.1016/j.ecolmodel.2018.09.010
Chamaillé-Jammes S., Valeix M., and Fritz H. (2007). Managing heterogeneity in elephant distribution: Interactions between elephant population density and surface-water availability. J. Appl. Ecol. 44, 625–633. doi: 10.1111/j.1365-2664.2007.01300.x
Chui S. Y. S., Getz W. M., Henley M., Seidel D. P., Keith M., Parrini F., et al. (2024). Habitat use strategies of African elephants under different seasonal and ecological constraints. Wildlife. Res. 51. doi: 10.1071/WR23139
Cutler D. R., Edwards T. C., Beard K. H., Cutler A., Hess K. T., Gibson J., et al. (2007). Random forests for classification in ecology. Ecology 88, 2783–2792. doi: 10.1890/07-0539.1
Dejene S. W., Mpakairi K. S., Kanagaraj R., Wato Y. A., and Mengistu S. (2021). Modelling Continental Range Shift of the African Elephant (Loxodonta africana) under a Changing Climate and Land Cover: Implications for Future Conservation of the Species. Afr. Zool. 56, 25–34. doi: 10.1080/15627020.2020.1846617
de Knegt H. J. (2010). Beyond the here and now: herbivore ecology in a spatial-temporal context, Doctoral thesis (Netherlands: Wageningen University and Research).
De Knegt H. J., Van Langevelde F., Skidmore A. K., Delsink A., Slotow R., Henley S., et al. (2011). The spatial scaling of habitat selection by African elephants. J. Anim. Ecol. 80, 270–281. doi: 10.1111/j.1365-2656.2010.01764.x
Dunkin R. C. (2012). From Tissues To Landscapes: How Thermal Physiology, Water Use, and Climate Influence Patterns of Landscape Use in Elephants (Santa Cruz: University of California).
Dzinotizei Z. (2018). Effects of elephant densities on landscape heterogeneity in relation to surface water availability: A GIS and remote sensing- based approach. (Harare: University of Zimbabwe), 1–76.
Elith J., Phillips S. J., Hastie T., Dudík M., Chee Y. E., and Yates C. J. (2011). A statistical explanation of MaxEnt for ecologists. Diversity Distrib. 17, 43–57. doi: 10.1111/j.1472-4642.2010.00725.x
Feleha D. D. (2018). Impact of African Elephant (Loxodonta africana) on Flora and Fauna Community and Options for Reducing the Undesirable Ecological Impacts. J. Biol. Agric. Healthcare. 8, 49–65.
Fick S. E. and Hijmans R. J. (2017). WorldClim 2: new 1-km spatial resolution climate surfaces for global land areas. Int. J. Climatol. 37, 4302–4315. doi: 10.1002/joc.5086
Gandiwa E., Heittkönig I. M. A., Eilers P. H. C., and Prins H. H. T. (2016). Rainfall variability and its impact on large mammal populations in a complex of semi-arid African savanna protected areas. Trop. Ecol. 57, 163–180.
Gara T. W., Wang T., Dube T., Ngene S. M., and Mpakairi K. S. (2021). African elephant (Loxodonta africana) select less fragmented landscapes to connect core habitats in human-dominated landscapes. Afr. J. Ecol. 59, 370–377. doi: 10.1111/aje.12839
García-Callejas D. and Araújo M. B. (2016). of model and data complexity on predictions from species distributions models. Ecol. Model. 326, 4–12. doi: 10.1016/j.ecolmodel.2015.06.002
Garstang M., Davis R. E., Leggett K., Frauenfeld O. W., Greco S., Zipser E., et al. (2014). Response of African elephants (Loxodonta africana) to seasonal changes in rainfall. PloS One 9. doi: 10.1371/journal.pone.0108736
Hahn N. R., Wall J., Deninger-Snyder K., Tiedeman K., Sairowua W., Goss M., et al. (2024). Crop use structures resource selection strategies for African elephants in a human-dominated landscape. Ecol. Evol. 14, 1–12. doi: 10.1002/ece3.11574
Hao T., Elith J., Lahoz-Monfort J. J., and Guillera-Arroita G. (2020). Testing whether ensemble modelling is advantageous for maximising predictive performance of species distribution models. Ecography 43, 549–558. doi: 10.1111/ecog.04890
Hauenstein S., Kshatriya M., Blanc J., Dormann C. F., and Beale C. M. (2019). African elephant poaching rates correlate with local poverty, national corruption and global ivory price. Nat. Commun. 10, 2242–2263. doi: 10.1038/s41467-019-09993-2
Jin Z., Shang J., Zhu Q., Ling C., Xie W., and Qiang B. (2020). “RFRSF: employee turnover prediction based on random forests and survival analysis,” in Lecture Notes in Computer Science (including subseries Lecture Notes in Artificial Intelligence and Lecture Notes in Bioinformatics), 12343 LNCS (Switzerland: Springer Nature), 503–515. doi: 10.1007/978-3-030-62008-0_35
John L., Shekede M. D., Gwitira I., Mazhindu A. N., Pfukenyi D. M., and Chikerema S. (2024). Modelling climate change impacts on the spatial distribution of anthrax in Zimbabwe. BMC Public Health 24, 1–14. doi: 10.1186/s12889-024-17856-9
Koskey J. (2016). Effects of variations in nutrient content of wild forage and raided crops on foraging behaviour of African elephants (Loxodonta africana) in Rimoi game reserve (Nairobi: University of Kenya).
Lee P. C., Moss C. J., Njiraini N., Poole J. H., Sayialel K., and Fishlock V. L. (2022). Cohort consequences of drought and family disruption for male and female African elephants. Behav. Ecol. 33, 408–418. doi: 10.1093/beheco/arab148
Lohay G. G., Weathers T. C., Estes A. B., McGrath B. C., and Cavener D. R. (2020). Genetic connectivity and population structure of African savanna elephants (Loxodonta africana) in Tanzania. Ecol. Evol. 10, 11069–11089. doi: 10.1002/ece3.6728
Lu X., Wang J., Huang Z., Fang Z., Ali M., Ashraf A., et al. (2025). Long-distance corridors facilitate asian elephant adaptation to climate change. Integr. Conserv. 4, 57–70. doi: 10.1002/inc3.70001
Mapheto T. J. (2023). Induced Thicket Degradation In Addo Elephant (South Africa: Nelson Mandela University).
Mlambo L., Adam E., Shekede M. D., and Odindi J. (2024). The influence of biophysical characteristics on elephant space use in an African savanna. Ecol. Inf. 82, 102724. doi: 10.1016/j.ecoinf.2024.102724
Mpakairi K. S., Ndaimani H., Tagwireyi P., Zvidzai M., and Madiri T. H. (2020). Futuristic climate change scenario predicts a shrinking habitat for the African elephant (Loxodonta africana): evidence from Hwange National Park, Zimbabwe. in European Journal of Wildlife Research. Eur. J. Wildlife. Res. 66, 1-27. doi: 10.1007/s10344-019-1327-x
Mukomberanwa N. T., Taru P., Utete B., and Madamombe H. K. (2024). Predicting the dry season habitat occupancy of african savannah elephant using vegetation indices and modelling landscape variability in a mesic protected area. Afr. J. Ecol. 62. doi: 10.1111/aje.13318
Muposhi V. K., Gandiwa E., Chemura A., Bartels P., Makuza S. M., and Madiri T. H. (2016). Habitat heterogeneity variably influences habitat selection by wild herbivores in a semi-arid tropical savanna ecosystem. PloS One 11. doi: 10.1371/journal.pone.0163084
Naidoo R., Brennan A., Shapiro A. C., Beytell P., Aschenborn O., Du Preez P., et al. (2020). Mapping and assessing the impact of small-scale ephemeral water sources on wildlife in an African seasonal savannah. Ecol. Appl. 30. doi: 10.1002/eap.2203
Naimi B. and Araújo M. B. (2016). sdm: a reproducible and extensible R platform for species distribution modelling. Ecography 39, 368–375. doi: 10.1111/ecog.01881
Ndaimani H. (2018). GIS and Remote Sensing Applications for Modelling the Distribution of Elephants and their Interaction with Vegetation (Harare: University of Zimbabwe).
Ochieng E. O. (2015). Characterizing the spatial distributions of elephants in Mpala, Kenya (Master's thesis, University of Twente, Netherlands).
Pearce J. and Ferrier S. (2000). Evaluating the predictive performance of habitat models developed using logistic regression. Ecol. Model. 133, 225–245. doi: 10.1016/S0304-3800(00)00322-7
Phillips S. B., Aneja V. P., Kang D., and Arya S. P. (2006). Modelling and analysis of the atmospheric nitrogen deposition in North Carolina. Int. J. Global Environ. Issues 6, 231–252. doi: 10.1016/j.ecolmodel.2005.03.026
Razak S. N. A., Ahmad W. J. W., Noh N. A. I., Nasir N. A. M., Shukor A. M., and Nor S. M. (2020). Regeneration of degraded lowland dipterocarp forest: elephants as seed dispersal agent. Water Resour. Dev. Manage. 25, 438–446. doi: 10.1007/978-981-15-1971-0_44
Robillard C. M., Coristine L. E., Soares R. N., and Kerr J. T. (2015). Facilitating climate-change-induced range shifts across continental land-use barriers. Conserv. Biol. 29, 1586–1595. doi: 10.1111/cobi.12556
Sach F., Dierenfeld E. S., Langley-Evans S. C., Watts M. J., and Yon L. (2019). African savanna elephants (Loxodonta africana) as an example of a herbivore making movement choices based on nutritional needs. PeerJ, 1–27. doi: 10.7717/peerj.6260
Searle C. E., Kaszta Ż., Bauer D. T., Kesch K., Hunt J. E., Mandisodza-Chikerema R., et al. (2022). Random forest modelling of multi-scale, multi-species habitat associations within KAZA transfrontier conservation area using spoor data. J. Appl. Ecol. 59, 2346–2359. doi: 10.1111/1365-2664.14234
Sharma P., Panthi S., Yadav S. K., Bhatta M., Karki A., Duncan T., et al. (2020). Suitable habitat of wild Asian elephant in Western Terai of Nepal. Ecol. Evol. 10, 6112–6119. doi: 10.1002/ece3.6356
Smit I. P. J., Peel M. J. S., Ferreira S. M., Greaver C., and Pienaar D. J. (2020). Megaherbivore response to droughts under different management regimes: lessons from a large African savanna. Afr. J. Range Forage. Sci. 37, 65–80. doi: 10.2989/10220119.2019.1700161
Taylor L. A., Vollrath F., Lambert B., Lunn D., Douglas-Hamilton I., and Wittemyer G. (2020). Movement reveals reproductive tactics in male elephants. J. Anim. Ecol. 89, 57–67. doi: 10.1111/1365-2656.13035
Thant Z. M., Leimgruber P., Williams A. C., Oo Z. M., Røskaft E., and May R. (2023). Factors influencing the habitat suitability of wild Asian elephants and their implications for human–elephant conflict in Myanmar. Global Ecol. Conserv. 43, 68–84. doi: 10.1016/j.gecco.2023.e02468
Valeix M. (2011). Temporal dynamics of dry-season water-hole use by large African herbivores in two years of contrasting rainfall in Hwange National Park, Zimbabwe. J. Trop. Ecol. 27, 163–170. doi: 10.1017/S0266467410000647
Valeix M., Fritz H., Matsika R., Matsvimbo F., and Madzikanda H. (2008). The role of water abundance, thermoregulation, perceived predation risk and interference competition in water access by African herbivores. Afr. J. Ecol. 46, 402–410. doi: 10.1111/j.1365-2028.2007.00874.x
Wool R. (2019). The distribution patterns of African elephant (Loxodonta africana) and white rhinoceros (Ceratotherium simum simum) in relation to waterhole availability in a South African savanna. Available online at: http://dspace.library.uu.nl/handle/1874/383485%0Ahttp://localhost/handle/1874/383485 (Accessed August 4, 2024).
Yu Q., Hu Z., Huang C., Xu T., Onditi K. O., Li X., et al. (2024). Suitable habitats shifting toward human-dominated landscapes of Asian elephants in China. Biodivers. Conserv. 33, 685–704. doi: 10.1007/S10531-023-02766-W
Keywords: African elephant, species distribution modelling, MaxEnt, random forest, ensemble, climate change
Citation: Boas F and Zvidzai M (2025) Estimating the spatial and temporal distribution of suitable habitats for the African elephant (Loxodonta africana) in Hwange National Park. Front. Conserv. Sci. 6:1699938. doi: 10.3389/fcosc.2025.1699938
Received: 05 September 2025; Accepted: 28 October 2025;
Published: 24 November 2025.
Edited by:
Antonio-Román Muñoz, Universidad de Málaga, SpainReviewed by:
Jihosuo Biswas, Primate Research Centre NE India, IndiaRomel Ahmed, Shahjalal University of Science and Technology, Bangladesh
Copyright © 2025 Boas and Zvidzai. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Mark Zvidzai, bWFyay56dmlkemFpQGdtYWlsLmNvbQ==