 Meredith K. Loth
Meredith K. Loth Kendall T. Mesch
Kendall T. Mesch Celine Herrera-Garcia1
Celine Herrera-Garcia1 Zoe R. Donaldson
Zoe R. Donaldson- 1Department of Molecular, Cellular, and Developmental Biology, University of Colorado Boulder, Boulder, CO, United States
- 2Department of Psychology and Neuroscience, University of Colorado Boulder, Boulder, CO, United States
Prairie voles (Microtus ochrogaster) are a powerful model for studying the neurobiology of social bonding, yet tools for region- and cell type-specific gene regulation remain underdeveloped in this species. Here, we present a lentivirus-mediated CRISPR activation and interference (CRISPRa/i) platform for somatic gene modulation in the prairie vole brain. This system enables non-mutagenic, titratable regulation of gene expression in the adult brain without germline modification. Our dual-vector system includes one construct expressing dCas9-VPR (VP64-p65-Rta) referred to as CRISPRa or dCas9-KRAB-MeCP2 (Kruppel-associated box-methyl CpG binding protein 2), referred to as CRISPRi under a neuron-specific promoter, and a second construct delivering a U6-driven sgRNA (single guide RNA) alongside an elongation factor 1 alpha (EF1α)-driven mCherry reporter. We detail the design, production, and stereotaxic delivery of these tools and demonstrate their application by targeting four genes implicated in social behavior (Oxtr, Avpr1a, Drd1, and Drd2) across two mesolimbic brain regions: the nucleus accumbens and ventral pallidum. Gene expression analyses confirmed robust, bidirectional transcriptional modulation for selected targets, establishing a proof of concept for CRISPRa/i in this non-traditional model. The dual-vector design is readily adaptable to other gene targets, cell types, and brain regions, and can be multiplexed to provide a flexible and scalable framework for investigating gene function in behaviorally relevant circuits. These advances represent the first successful implementation of somatic CRISPRa/i in prairie voles and expand the genetic toolkit available for this species.
1 Introduction
Bonding and affiliation are fundamental components of social behavior, and their study has implications for understanding both typical and disordered social functioning. Prairie voles (Microtus ochrogaster) have emerged as a powerful model species for investigating the neurobiology of social attachment (Sue Carter, Courtney Devries and Getz, 1995; Insel and Young, 2001; Young and Wang, 2004). Unlike traditional laboratory rodents such as mice and rats, but like humans, prairie voles have a naturally monogamous mating system, and exhibit pair bonding behavior and biparental care. This makes them uniquely suited for studying the molecular and neural mechanisms underlying social bonding, partner preference, and parental care. Extensive behavioral characterization, combined with region-specific receptor mapping, have revealed critical roles for oxytocin, vasopressin, and dopamine signaling in the formation and maintenance of social bonds in this species (Young et al., 2001; Lim et al., 2004; Aragona et al., 2006; Lim and Young, 2006; Donaldson and Young, 2008; Gobrogge and Wang, 2016).
Despite their value as a behavioral model, gene manipulation approaches in voles have historically been more limited compared to other established model organisms. Transgenic prairie voles were first developed over a decade ago (Donaldson et al., 2009) and more recent studies have used CRISPR/Cas9 to generate germline knockouts of the oxytocin receptor (Horie et al., 2019; Berendzen et al., 2023) or to deliver gene-editing tools via AAV to the brain (Boender et al., 2023). However, these CRISPR-based approaches have relied on active nuclease strategies, which introduce irreparable genomic damage and are limited to gene disruption.
In contrast, CRISPRa/i systems use catalytically inactive Cas9 (dCas9) fused to transcriptional activators or repressors, enabling potentially reversible, non-mutagenic regulation of endogenous gene expression. While multiple strategies exist for gene manipulation in the brain, few offer the combined advantages of spatial precision, temporal control, and the ability to incrementally adjust (titrate) expression levels in somatic tissues. As demonstrated by Savell et al. (2019b), such systems allow for dose-dependent changes in gene expression that better reflect physiological variability. As summarized in Supplementary Table S1, CRISPRa/i uniquely supports flexible, scalable, and region-specific gene up- or downregulation without permanent genomic alterations, making it particularly well-suited for probing gene function in behaviorally relevant brain circuits.
The broader application of CRISPRa/i across species is needed to open new directions for comparative neuroscience, behavioral genetics, and evolutionary biology. It will enable targeted, cell-specific manipulation of genes in organisms with ecologically and socially relevant behaviors. Thus, to advance genetic tool development in prairie voles, we established a lentivirus-mediated CRISPRa/i platform for somatic gene regulation in the brain. This system enables activation or interference of target genes in a spatially and cell type-specific manner without requiring germline manipulation. Our protocol involves co-injection of two lentiviral constructs: one effector encoding the dCas9-VPR (VP64-p65-Rta) referred to as CRISPRa or dCas9-KRAB-MeCP2 (Kruppel-associated box-methyl CpG binding protein 2), referred to as CRISPRi under the neuron-specific synapsin (SYN) promoter, and a second effector carrying a U6-driven sgRNA (single guide RNA) targeted to the gene of interest with a mCherry reporter under the elongation factor 1 alpha (EF1α) promoter. We selected lentivirus for delivery due to its packaging capacity, which accommodates the large size of dCas9-VPR/KRAB-MeCP2 fusion proteins and neuron-specific regulatory elements. In addition, the dual-vector design allows for modular exchange of sgRNA constructs without the need to repackage dCas9, supporting flexible and iterative gene targeting in vivo. We validated this approach by modulating expression of oxytocin, vasopressin, and dopamine receptor genes in the nucleus accumbens or ventral pallidum. This platform is adaptable for a range of gene targets, brain regions, and experimental timelines, and holds promise for investigating the molecular basis of social behavior, mapping gene function in specific circuits, and evaluating gene-environment interactions in a behaviorally relevant mammalian model.
2 Methods
2.1 Animals and housing
All procedures were approved under the University of Colorado’s Institute of Animal Care and Use Committee (IACUC) and performed in the light phase. All authors complied with the Animal Research: Reporting of In Vivo Experiments (ARRIVE) guidelines.
Prairie voles were bred in-house from colonies originating from Emory University and the University of California Davis, both of which maintain lines descended from wild-caught animals in Illinois (Sue Carter, Courtney Devries and Getz, 1995). These colonies have been used extensively in studies of social behavior and are genetically outbred to preserve natural variation. Voles were weaned at postnatal day 21 and were then housed in standard static rodent cages (17.5L x 9.0w. x 6.0h. in.) in groups of 2–4 with either same sex siblings or same sex voles from similar weaning time frames. Animals were given ad libitum access to water and rabbit chow (5326–3 by PMI Lab Diet). Rabbit chow was supplemented with sunflower seeds, dehydrated fruit bits, and alfalfa cubes. Enriched cages consisted of cotton nestlets, a plastic igloo, and a PVC pipe. Animals were kept in a temperature (23°C–26°C) and humidity-controlled room with a 14:10 h light-dark cycle.
2.2 HEK293T cell line
Human embryonic kidney (HEK)293T cells (ATCC, RRID:CVCL_0063) were obtained from the American Type Culture Collection. They were maintained in standard HEK media: DMEM (High Glucose, Pyruvate; Gibco 11995081) supplemented with 10% FBS (Qualified US Origin; BioFluid 200-500-Q) and 1 U/mL Penicillin-Streptomycin (Gibco 15140122). Cells were cultured in T75 or T225 tissue culture flasks and passaged at 70%–80% confluence, with a maximum of 25 passages.
2.3 Viral vector design and production
Gene-specific sgRNA targets were designed using Benchling (RRID:SCR_013955), the only platform containing the prairie vole genome (MicOch1.0) for identifying potential off-target effects. sgRNA’s were purchased as oligonucleotides with BbsI cleavage site compatible overhangs from Integrated DNA Technologies (IDT). Oligos comprising sgRNAs were annealed together and cloned into the Bbs1 sites in lenti U6-sgRNA/EF1α-mCherry vector (RRID: Addgene_114199). sgRNA’s for CRISPRa were restricted to −500 bp upstream of the target gene, and sgRNA’s for CRISPRi were restricted to +300 bp downstream of the target gene as per prior recommendations (Maeder et al., 2013; Mali et al., 2013; Konermann et al., 2015). sgRNAs specificity was assessed using National Center for Biotechnology Information’s (NCBI) Basic Local Alignment Search Tool (BLAST). Information related to sgRNA sequences and distance from gene transcription start site (TSS) can be found in Supplementary Table S2. Complete plasmid sequence was confirmed via Plasmidsaurus prior to lentivirus production.
2.4 Lentivirus generation
Lentivirus production was performed as specified in Savell et al. (2019a) with some modifications. Briefly, large scale viruses were produced in a sterile environment following Biosafety Level 2 (BSL-2) safety guidelines. HEK-293T cells were transfected with a corresponding CRISPR plasmid: (lenti SYN-FLAG-dCas9-VPR (RRID: Addgene_114196)); lenti SYN-dCas9-KRAB-MeCP2 (RRID: Addgene_155365); lenti U6-sgRNA/EF1a-mCherry (RRID: Addgene_114199)); and psPAX2 packaging plasmid (RRID: Addgene_12260) and the pCMV_VSV-G envelope plasmid (RRID: Addgene_8454) and FuGENE ® HD Transfection Reagent (Promega) in supplemented Ultraculture media (L-glutamine, sodium pyruvate, and sodium bicarbonate) in a T225 culture flask. Supernatant was passed through a 0.45 µm filter and centrifuged at 106,883 g (38,100 rpm) for 65 min at 4°C in a 70Ti fixed angle titanium rotor (Beckmann Coulter). The viral pellet was resuspended in 1/100th supernatant volume of sterile PBS and stored at −80°C. Physical viral titer was determined using Lenti-X qRT-PCR Titration kit (Takara) and only viruses with >1 × 109 GC/mL were used. Viruses were stored in sterile aliquots of PBS at −80°C.
2.5 Stereotactic surgeries and viral injection protocol
Sexually naïve adult prairie voles were anesthetized with isoflurane (4% induction and 1.5%–2.5% maintenance) at an oxygen flow rate of 1L/min and secured in a head-fixed stereotaxic frame (Kopf Instruments). Female reproductively intact animals were used for all experiments, except those targeting Avpr1a, for which males were used. In total, 65 animals were used across all experimental groups. Body temperature was maintained at 37°C using a closed loop heating pad with a rectal thermometer. Eyes were lubricated with ophthalmic ointment (Sterile Lubricant Eye Ointment), and depth of anesthesia was monitored by breathing and toe and tail pinch response. Using a shaver, fur was removed from the dorsal portion of the head and, a midline incision was made and disinfected with 70% isopropyl alcohol followed by 10% betadine. Briefly, the scalp and connective tissue were removed above the frontal skull plates and the head leveled in the anterior-posterior plane. Guide holes were drilled using stereotaxic coordinates (all coordinates in respect to bregma (Paxinos and Watson, 2007): nucleus accumbens core and shell: AP: +1.7; ML ±: 1.0; ventral pallidum: AP: +1.5; ML ±: 0.9; All infusions were made under anesthesia using a gastight 30-gauge stainless steel injection needle (Hamilton Syringes) that extended into the infusion site. Bilateral lentivirus microinfusions were made using a UMP3-T syringe pump (World Precision Instruments) at a rate of 2 nL/s at DV: −4.8/4.7/4.6 mm for the nucleus accumbens or DV: −5.8/5.7/5.6 mm for the ventral pallidum. At each DV coordinate, 700/700/600 nL were injected for a total of 2 uL per hemisphere (See Supplementary Figure S1 for virus volume validation). Injection needles remained in place for 10 min post infusion to allow time for diffusion. Prairie voles were infused bilaterally with 2 uL of total lentivirus mix which comprised of a 1:10 ratio of sgRNA virus to dCas9-VPR or KRAB-MeCP2 virus in sterile PBS. Surgical incisions sites were closed with vicryl sutures. Animals received extended release meloxicam (4 mg/kg), lidocaine at the incision site, and saline (1 mL) for pain, and post-operative management. Animals recovered 72 h in a BSL-2 facility to minimize any exposure to animal shedding of lentivirus particles.
2.6 Tissue collection and microdissection
For samples used in qRT-PCR, animals were euthanized via rapid decapitation. Brains were immediately extracted and rinsed in sterile saline. Fluorescent guided dissections were performed manually using sterile RNase free razor blades and mCherry fluorescence was visualized with a fluorescent dissecting microscope Olympus MVX10 MacroZoom at the University of Colorado Boulder MCDB Light Microscopy Core Facility (RRID:SCR_018993). During dissection, tissue was kept on ice blocks and in cold saline. For each animal, the right and left hemispheres received different viral constructs (targeting sgRNA vs. LacZ control), and each hemisphere was dissected separately. No pooling was performed. Each data point in the qRT-PCR dataset represents tissue from a single hemisphere of an individual animal. Because viral constructs were injected into opposite hemispheres, each animal served as its own control, minimizing variability due to individual differences in gene expression or viral transduction. Dissected hemispheres were flash frozen on dry ice and stored at −80°C until RNA extraction.
For samples used for immunohistochemistry, animals were injected with 0.15–0.30 mL 1:2 ketamine/xylazine and transcardially perfused with 4% paraformaldehyde (PFA, Electron Microscopy Scienes) in phosphate buffered saline. Brains were kept in 4% PFA overnight and transferred to 30% sucrose. After brains sank, they were rinsed and flash frozen on dry ice and stored at −80°C for 24 h prior to sectioning on a freezing microtome (Leica June SM2000R, 40 µm/slice).
2.7 RNA extraction and cDNA generation
Samples were processed for total purified RNA as described in Cunningham et al. (2019) using the Norgen Total RNA/gDNA kit (Norgen Biotek no. 48700). Briefly, frozen tissue was placed in 600 µL ice cold lysis buffer and homogenized using a Scilogex homogenizer. Homogenized tissue was kept on wet ice while all animals for the cohort were processed. The homogenizer tip was thoroughly cleaned in between each animal with DEPC-treated water and 70% ethanol to prevent cross contamination across samples. Homogenized samples were centrifuged at 4000 g for 10 min and supernatant was transferred to new pre-chilled tubes.
Total RNA (2 uL for each sample) was Nanodropped to determine RNA concentration (Agilent Technologies). RNA samples were all diluted to 25 ng/μL with RNAse-free water (Norgen). cDNA was generated using high Capacity cDNA Reverse Transcription kit (Applied Bio Systems). For each sample, cDNA was produced using a mixture of 2 μL of 10X RT buffer, 0.8 μL of dNTP, 2 μL of random primers, 1 μL of Transcriptase, 1 μL of RNA inhibitor, and 3.2 μL of ddH2O. RNA (250 ng) was then added, and cDNA was generated using reaction conditions 25°C for 10 min, 2 × (37°C for 120 min, 85°C for 5 min). Samples were stored at −20°C until processing.
2.8 qRT-PCR validation of gene expression
Prairie vole gene expression for Oxtr, Avpr1a, Drd1, Drd2, and Gapdh was quantified using qPCR. Primer sequences are provided in Supplementary Table S3. Samples and probes were processed in triplicate in MicroAmp Fast Optical 96 well Reaction Plate (Applied Bio Systems) with 0.66 µL of cDNA, 0.50 µL of probe, 5 µL of TaqMan Fast Advanced Master Mix (Applied Biosystems), and 3.84 µL of ddH2O per well for a total volume of 10 µL. Plates were covered with Optical Adhesive Film (Applied Biosystems), vortexed, and centrifuged prior to PCR amplification in Applied Biosystems QuantSTudio 3 qPCR machine (Applied Biosystems). DNA was amplified using the following cycling conditions: 50°C for 2 min, 95°C for 20 s, and 40 cycles of 95°C for 1 s and 60°C for 20 s.
The mean cycle threshold (Ct) value was calculated for each sample run in triplicate. Triplicates with a standard deviation greater than 0.30 were assessed for potential outliers. In cases where a clear outlier could be visually identified (e.g., one Ct value substantially deviating from the other two), that value was excluded and the remaining two values were averaged. If no clear outlier was apparent, the entire sample was excluded to avoid introducing bias. In total, 44 out of 864 wells (5.1%) were excluded from analysis based on this criterion, spanning multiple PCR plates and experiments. The exclusion rate is within the expected range for qPCR experiments and reflects a conservative quality control approach to ensure the accuracy and reliability of reported expression values. Gene expression quantification was performed using the ΔΔCt method (Livak and Schmittgen, 2001), with all samples normalized to Gapdh.
2.9 Immunohistochemistry and microscopy
Immunohistochemistry was performed to visualize the FLAG tag on dCas9-VPR and dCas9-KRAB-MeCP2 constructs. Free-floating brain slices from PFA-perfused prairie voles were stored at −20°C in cryoprotectant consisting of 50% glycerol in 0.05 M phosphate buffer. For immunofluorescent labeling of FLAG, sections were washed in 1X PBS, for 30 min, with fresh PBS every 5 min. Sections were blocked with 5% normal donkey serum (Jackson Laboratories) containing 0.2% Triton X-100 for 1 h followed by primary antibody incubation with mouse-anti-FLAG (Invitrogen, RRID:AB_1957945, 1:100) at 4°C for 40 h in 750 uL of solution. After washing, sections were incubated with AlexaFluor 488 secondary antibody (Life Technologies, 1:500) for 1 h. Slices underwent another series of washes and were then mounted on slides and coverslipped with Prolong Gold Antifade Mountant (Life Technologies) to preserve signal intensity and brightness.
Slices were imaged using a Nikon A1 Laser Scanning confocal Microscope at the University of Colorado Boulder MCDB Light Microscopy Core Facility (RRID:SCR_018993). Confocal images were taken with a 20x lens in two channels (green and red) of the nucleus accumbens and ventral pallidum. Confocal stacks were projected as single images using maximum fluorescence.
2.10 Data analysis and statistics
Data are shown as means ± standard error of the mean. Statistical significance α was set at 0.05. All n values represent number of animals. Statistical analyses were carried out using either GraphPad PRISM (version 10.4.1, Graphpad, San Diego, CA) or Python (v 3.12.4) in a reproducible computing environment. Python scripts were executed in Juypter Notebook (v 7.0.8) via Anaconda Navigator (v 2.6.4). Paired t-tests with error propagation were performed.
3 Results
3.1 CRISPR a/i constructs successfully expressed in the prairie vole brain
We employed a dual-vector system to enable gene activation or repression. One lentiviral construct encoded either dCas9-VPR (Figure 1A) or dCas9-KRAB-MeCP2 (Figure 1B) under the control of the neuron-specific synapsin (SYN) promoter. The second construct carried a U6-driven sgRNA targeting either Oxtr, Avpr1a, Drd1, Drd2, or a non-targeting lacZ control, and included a mCherry reporter driven by the EF1α promoter for visualization of sgRNA-expressing cells (Figures 1A,B). Following lentiviral injection, animals were given 21 days to allow for both recovery and robust expression of CRISPRa/i constructs (Figure 1C), after which immunohistochemistry (IHC) was performed to confirm expression in targeted brain regions, the nucleus accumbens (Figure 1D) or ventral pallidum (Figure 1E).
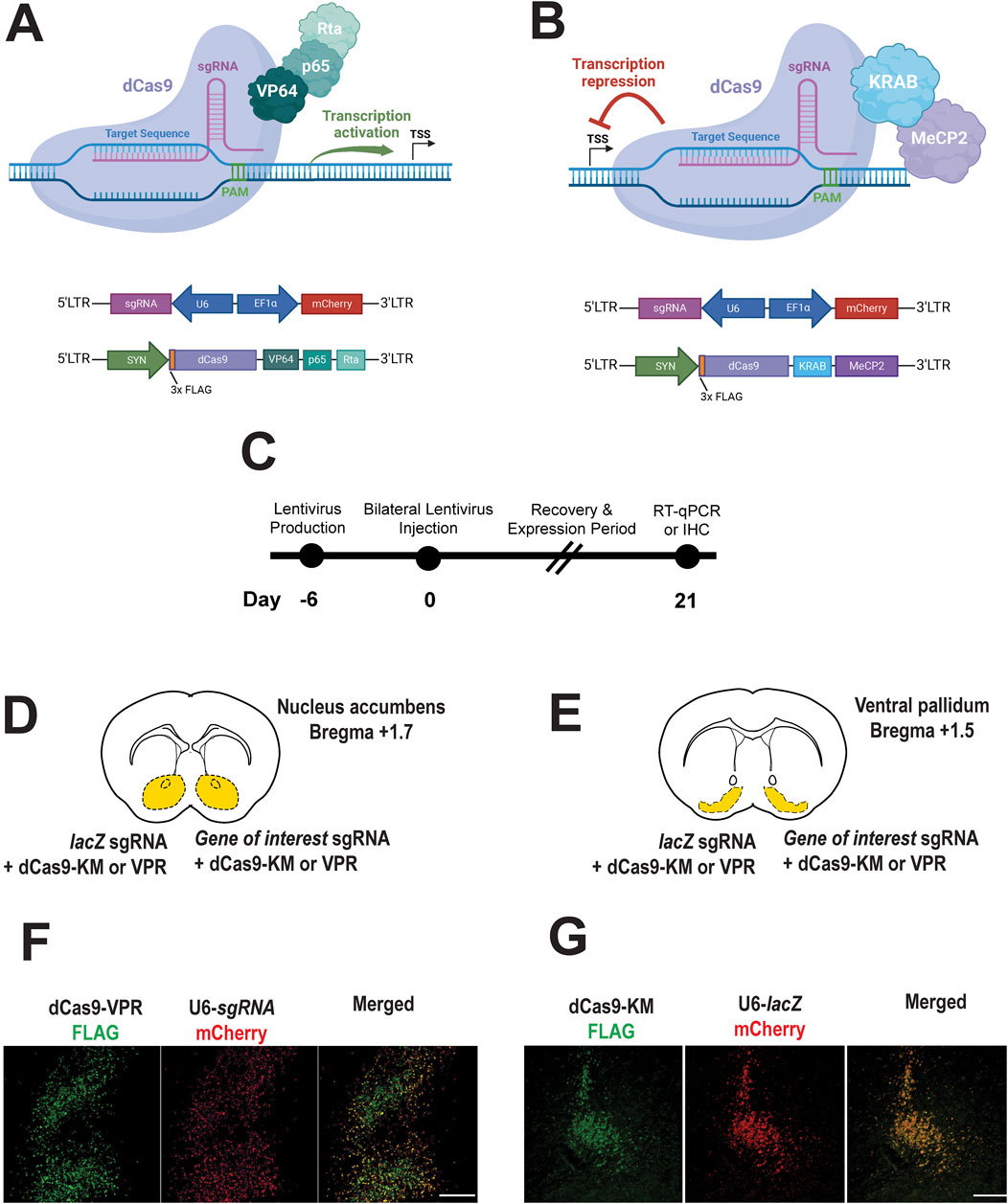
Figure 1. CRISPR a/i constructs successfully expressed in the prairie vole brain. (A,B) CRISPRa vector for expressing dCas9-VPR activator fusion (A), CRISPRi vector for expressing dCas9-KRAM-MeCP2 interference fusion (B), and sgRNA’s targeting either the bacterial lacZ gene (nontargeting control) or other genes of interest in the prairie vole (A,B). (C). Experimental timeline for CRISPRa or CRISPRi expression the prairie vole brain (nucleus accumbens or ventral pallidum). (D,E). Brain atlas diagrams of the nucleus accumbens (D) and the ventral pallidum (E) indicating dual bilateral injections of CRISPRa or CRISPi construct with sgRNA construct targeting lacZ (control) or gene of interest. (F,G). Immunohistochemistry images reveal successful transduction and show representative expression of CRISPRa (E) and CRISPRi (F) (FLAG, green) lentiviruses along with co-expression of U6-driven sgRNA’s (mCherry, red). Merged images confirm colocalization of CRISPRa/i with sgRNA viruses (scale bar = 100 µm).
Representative images demonstrate detection of constructs in the nucleus accumbens (Figure 1F) and the ventral pallidum (Figure 1G). Specifically, FLAG-tagged dCas9-VPR (CRISPRa) and FLAG-tagged dCas9-KM (CRISPRi) were detected in transduced neurons (Figures 1F,G, left panels, respectively). Co-expression of U6-driven sgRNA’s, co-labeled with a mCherry reporter under an EF1α, promoter was observed in the same regions (Figures 1F,G, middle panels). Merged images confirm colocalization of dCas9 constructs and sgRNA’s indicating successful neuronal expression of both components (Figures 1F,G, right panels).
These findings validate expression of lentiviral CRISPRa/i in a non-traditional mammalian model, demonstrating effective lentiviral-mediated delivery and neuronal expression of CRISPRa and CRISPRi constructs in the prairie vole brain across two brain regions.
3.2 CRISPR a/i modulation of behaviorally relevant genes
To evaluate the effectiveness of CRISPRa and CRISPRi in the prairie vole brain, we focused on four target genes implicated in social behavior: the nonapeptide receptors Oxtr and Avpr1a, and the two most abundant dopamine receptors, Drd1 and Drd2. We targeted these genes in mesolimbic brain regions where they are highly expressed and known to modulate pair bonding behaviors (Wang et al., 1999; Young, 1999; Gingrich et al., 2000; Young et al., 2001; Aragona et al., 2006; Gobrogge and Wang, 2016; Loth and Donaldson, 2021; Pierce et al., 2024). Specifically, Oxtr, Drd1, and Drd2 were targeted in the nucleus accumbens (NAc), while Avpr1a was targeted in the ventral pallidum (VP).
3.2.1 Oxtr mRNA expression
qPCR analysis revealed that CRISPRa significantly increased Oxtr expression, while CRISPRi led to a significant reduction (Figures 2A,B, respectively). Specifically, dCas9-VPR activation resulted in a 4.29-fold increase in Oxtr expression compared to lacZ control (paired t-test, p = 0.03). Conversely, dCas9-KRAB-MeCP2 repression significantly decreased Oxtr expression by 87% (p < 0.001). These findings confirm that CRISPRa/i tools can effectively regulate Oxtr transcription in the prairie vole brain.
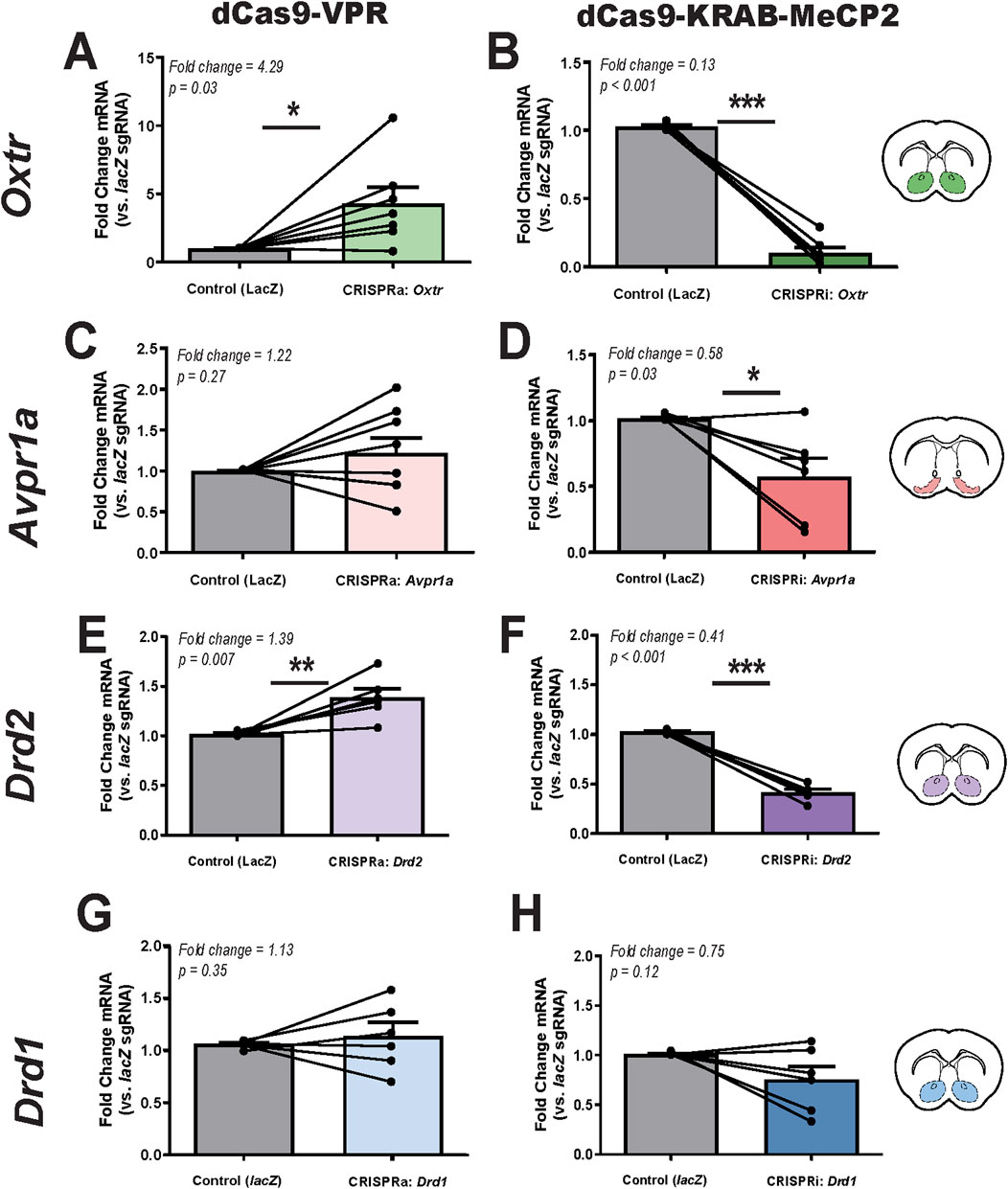
Figure 2. CRISPRa and CRISPRi modulation of target gene expression in the prairie vole brain. (A,B). Relative Oxtr mRNA expression following CRISPRa (dCas9-VPR, (A), n = 7) and CRISPRi (dCas9-KRAB-MeCP2, (B), n = 6) manipulation in the nucleus accumbens, as measured by qPCR. (C,D). Relative Avpr1a mRNA expression following CRISPRa (C), n = 8 and CRISPRi (D), n = 6 in the ventral pallidum. (E,F). Relative Drd2 mRNA expression following CRISPRa (E), n = 6 and CRISPRi (F), n = 6 in the nucleus accumbens. (G,H). Relative Drd1 mRNA expression following CRISPRa (G), n = 6 and CRISPRi (H), n = 6 in the nucleus accumbens. Injection sites are illustrated in coronal sections (inset), with colored markers indicating targeting of the nucleus accumbens (green, purple, blue) and ventral pallidum (pink). Each animal served as its own internal control, with lacZ control sgRNA injected into the left hemisphere and gene-specific sgRNA injected into the right hemisphere. Expression levels were normalized to Gapdh, and data are presented as fold change relative to lacZ control (mean ± SEM). Statistical significance was determined using paired t-tests, with p < 0.05 considered significant (*, p < 0.05; **, p < 0.01; ***, p < 0.001).
3.2.2 Avpr1a mRNA expression
qPCR analysis revealed greater variability in Avpr1a expression following CRISPRa and CRISPRi manipulation (Figures 2C,D). CRISPRa-driven activation led to a 1.22-fold increase in Avpr1a expression relative to the control, but this change was not statistically significant (p = 0.27). In contrast, CRISPRi significantly reduced expression by 42% (p = 0.03). Notably, Avpr1a expression levels exhibited high variability across animals and the magnitude of these effects were smaller than those observed for Oxtr. An additional CRISRPi sgRNA tested for Avpr1a yielded similar results with significantly reduced expression by 37% (p = 0.02) (Supplementary Figure S2). In sum, the overall trend of CRISPRa upregulation and CRISPRi repression was consistent.
3.2.3 Drd2 mRNA expression
Drd2 expression was modulated in a gene-specific manner, with CRISPRa inducing a significant 1.39-fold increase (p = 0.007) and CRISPRi leading to a 59% reduction (p < 0.001) (Figures 2G,H). Variability in Drd2 expression across animals was lower than Drd1 and Avpr1a, which may have contributed to its statistical significance. While Drd2 expression was significantly modulated by CRISPRa and CRISPRi, the magnitude of these changes was smaller compared to Oxtr, suggesting potential differences in baseline expression levels or regulatory mechanisms within the nucleus accumbens.
3.2.4 Drd1 mRNA expression
Drd1 mRNA expression was not significantly altered by either CRISPRa or CRISPRi manipulation. Similar to Avpr1a, Drd1 expression exhibited variability across animals following both CRISPRa and CRISPRi manipulation (Figures 2E,F). CRISPRa injection resulted in a 1.13-fold increase in Drd1 expression, though this change was not statistically significant (p = 0.35). Similarly, CRISPRi injection led to a 25% reduction in Drd1 expression, but this effect was also not significant (p = 0.12).
Taken together, these results confirm the feasibility of using CRISPRa/i to modulate Oxtr and Drd2 expression in prairie voles, while highlighting potential technical limitations in targeting Avpr1a and Drd1 with the current approach.
4 Discussion
Our findings establish lentivirus-mediated CRISPRa/i as a flexible and regionally precise tool for manipulating somatic gene expression in prairie voles. By delivering lentiviral constructs encoding dCas9-VPR or dCas9-KRAB-MeCP2 alongside U6-driven sgRNAs, we demonstrate the feasibility of bidirectional transcriptional control in neurons of the adult prairie vole brain. This system provides a valuable method for probing gene function in vivo—particularly for genes implicated in complex social behaviors—without requiring germline genetic manipulation. While in vivo CRISPRa/i systems have been validated in mice and rats (Savell et al., 2019a; 2020; Duke et al., 2020; Deng et al., 2022; Bendixen et al., 2023), to our knowledge, this is the first application in a non-traditional mammalian model. Expanding gene modulation tools to prairie voles opens new opportunities to study the molecular basis of social bonding in a species with affiliative behaviors more analogous to humans.
We demonstrate effective region-specific modulation of key social behavior genes across two distinct brain regions: the nucleus accumbens (Oxtr, Drd1, Drd2) and the ventral pallidum (Avpr1a). Oxtr and Drd2 showed robust, consistent modulation following injection into the nucleus accumbens, confirming effective somatic gene regulation using CRISPRa/i. These results provide proof-of-concept for somatic, region-specific gene activation and interference in prairie voles. The ability to efficiently upregulate or downregulate key neuromodulatory receptors—without permanent genomic alterations—offers a powerful platform for studying the dynamics of oxytocinergic and dopaminergic signaling in behaviors ranging from pair bonding and biparental care.
However, not all targets exhibit equal efficiency of repression or activation. Avpr1a and Drd1 exhibited more variable modulation, highlighting areas for continued methodological refinement. For Avpr1a, CRISPRi led to a modest but statistically significant reduction in expression, while CRISPRa effects were inconsistent and did not reach statistical significance. The two CRISPRi sgRNAs, positioned at +115 and +257 bp relative to the transcription start site (TSS), yielded comparable outcomes, suggesting that TSS proximity alone does not explain the variability. Other factors—such as chromatin accessibility, local epigenetic context, or sequence-specific properties—may have influenced sgRNA efficacy at this locus. The presence of an Avpr1a pseudogene in prairie voles did not impact interpretation, as our qPCR primers were designed to exclude pseudogene amplification and the gRNAs targeted sequences specific to the functional gene (Young et al., 1997). Variability in Drd1 modulation may also reflect a combination of locus-specific and technical factors, such as lentiviral transduction variability, promoter interference, or subject-level biological differences. Additionally, sequence variation at or near sgRNA binding sites, such as naturally occurring SNPs (single nucleotide polymorphisms), could impact targeting efficiency in outbred populations and warrants future investigation. These findings highlight the need for systematic sgRNA validation and dose-response/multiplex testing as part of CRISPRa/i implementation in novel species.
Despite these gene-specific challenges, CRISPRa/i offers multiple advantages over nuclease-based editing. Because dCas9 lacks catalytic activity, this system avoids introducing DNA double-strand breaks, thereby preserving genomic integrity and enabling reversible gene regulation. This is particularly advantageous for somatic studies in the brain, where permanent edits may trigger developmental compensation or long-term side effects. In addition, CRISPRa/i allows for precise spatial and temporal control—gene expression can be manipulated after development, in specific brain regions, and within defined cell populations without altering the germline or affecting non-target tissues. CRISPRa/i also enables tunable gene expression, allowing researchers to modulate transcriptional output rather than merely switching genes on or off. Finally, the system supports multiplexed sgRNA delivery for simultaneous regulation of multiple genes within a circuit. While not the focus of the present study, these capabilities underscore the broader utility of the CRISPRa/i platform for dissecting complex gene networks and behaviorally relevant pathways.
Recent germline knockout studies have raised important questions about the necessity of oxytocin receptor signaling in pair bonding, with Berendzen et al. (2023) reporting that Oxtr deletion does not abolish partner preference in prairie voles. However, such findings may reflect developmental compensation or the inability of whole-animal knockouts to isolate spatially and temporally specific gene functions In contrast, the CRISPRa/i platform presented here enables spatially and temporally controlled modulation of endogenous gene expression in adulthood, offering a flexible approach for investigating gene function in specific brain regions during behaviorally relevant windows.
This flexibility may be particularly valuable for extending gene modulation to non-neuronal populations, where neuromodulatory receptors such as Oxtr, Drd1, and Drd2 may also be expressed. Our recent study in prairie voles revealed that these receptor transcripts are detectable in non-neuronal cell types within the nucleus accumbens (Loth et al., 2025), consistent with findings in other rodent species demonstrating Oxtr expression in astrocytes (Wei et al., 2020). In addition, transcriptional profiling from our lab has revealed glia-specific plasticity in response to social experience (Sadino et al., 2023; Brusman et al., 2024) and we have previously outlined conceptual models of glial involvement in pair bonding and neuromodulator signaling (Loth and Donaldson, 2021). Together, these findings highlight the importance of developing CRISPRa/i tools that can be adapted for cell-type–specific targeting, for example, using glial-specific promoters (e.g., GFAP, Iba1) or viral vectors with selective tropism. While the current study focused on neuronal manipulation, future iterations of this method may enable more refined interrogation of the non-neuronal contributions to pair bonding and other social behaviors.
Another key strength of CRISPRa/i is its gene-level specificity. Unlike pharmacological agents, which often target multiple receptor subtypes within a class (e.g., D1-like or D2-like dopamine receptors), sgRNAs can be designed to selectively target a single gene. This specificity is particularly valuable when dissecting the roles of closely related receptor isoforms in complex behaviors. For example, while commonly used D2 receptor antagonists can also bind D3 receptors (Scatton et al., 2001; Bock et al., 2004; Stahl, 2017), CRISPRi targeting of Drd2 allows for specific repression of Drd2 transcription without affecting Drd3. Similarly, pharmacological tools for oxytocin and vasopressin receptor systems frequently exhibit cross-reactivity due to high sequence homology, whereas CRISPRa/i enables selective modulation of Oxtr or Avpr1a independently. This specificity is crucial for resolving gene-specific contributions to behavior in systems where receptor subtypes have overlapping yet distinct roles.
There are a handful of limitations to our advance. While qPCR confirmed effective transcriptional modulation, we were unable to validate changes at the protein level due to limited antibody availability for our targets. Although autoradiography could potentially be used for receptor-level detection, it was not implemented here due to resource constraints. Additionally, we did not assess behavioral outcomes, and thus the functional impact of gene modulation remains to be tested in future work. Lentiviral vectors require BSL-2 containment and may not be optimal for all applications. Lastly, the long-term stability of lentiviral CRISPRa/i expression and the possibility of off-target effects warrant further investigation.
Despite these challenges, this work provides a critical proof-of-concept for somatic gene regulation using CRISPRa/i in prairie voles. Beyond validating a powerful new method, our results highlight key opportunities for future development:
• Refining sgRNA selection pipelines, particularly for Avpr1a inhibition, including exploration of sequence variation at sgRNA target sites as a potential contributor to guide efficiency.
• Exploring alternative viral platforms (e.g., AAV) using smaller dCas9 variants.
• Expanding this system to non-neuronal cell types via other cell-type specific promoters.
• Enhancing temporal precision through inducible dCas systems.
• Investigating potential off-target activity or compensatory gene expression in vivo.
• Assessing behavioral consequences of gene modulation in social bonding paradigms.
In summary, we establish lentivirus-mediated CRISPRa/i as a novel and effective method for somatic gene regulation in prairie voles, capable of modulating gene expression in a region- and cell type-specific manner across multiple behaviorally relevant brain regions. This system fills a critical methodological gap for genetic manipulation in this species, laying the foundation for future work on the molecular mechanisms of social behavior.
Data availability statement
The raw data supporting the conclusions of this article will be made available by the authors, without undue reservation.
Ethics statement
The animal study was approved by University of Colorado’s Institute of Animal Care and Use Committee (IACUC). The study was conducted in accordance with the local legislation and institutional requirements.
Author contributions
ML: Conceptualization, Data curation, Formal Analysis, Funding acquisition, Investigation, Methodology, Software, Supervision, Visualization, Writing – original draft, Writing – review and editing. KM: Formal Analysis, Investigation, Writing – review and editing. CH-G: Formal Analysis, Investigation, Writing – review and editing. LB: Investigation, Methodology, Writing – review and editing. ZD: Conceptualization, Formal Analysis, Funding acquisition, Methodology, Project administration, Resources, Software, Supervision, Writing – original draft, Writing – review and editing.
Funding
The author(s) declare that financial support was received for the research and/or publication of this article. This work was supported by awards from the Dana Foundation, the Whitehall Foundation, National Science Foundation (NSF) IOS-1827790, and National Institute of Health (NIH) DP2OD026143 to ZD and T32 DA 17637 support to ML.
Acknowledgments
We thank the voles for their sacrifice and contribution to research. We thank Jessica Abazaris and the rest of the animal care staff at the University of Colorado Boulder for their excellent care of the voles. Kelly Winther, Katie Gallagher, and Kresil Gordon managed the animal colony and provided experimental support. We acknowledge the Light Microscopy Core Facility, Porter B047, B049, B051 and B059 at the University of Colorado Boulder (RRID:SCR_018993) for help and advice with microscopy and thank Dr. James D. Orth for his assistance. We thank the Donaldson lab, Devanand Manoli’s lab, and Jessica Tollkuhn’s lab for their advice, feedback and support.
Conflict of interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Generative AI statement
The author(s) declare that no Generative AI was used in the creation of this manuscript.
Publisher’s note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
Supplementary material
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fgeed.2025.1602983/full#supplementary-material
References
Aragona, B. J., Liu, Y., Yu, Y. J., Curtis, J. T., Detwiler, J. M., Insel, T. R., et al. (2006). Nucleus accumbens dopamine differentially mediates the formation and maintenance of monogamous pair bonds. Nat. Neurosci. 9 (1), 133–139. doi:10.1038/nn1613
Bendixen, L., Jensen, T. I., and Bak, R. O. (2023). CRISPR-Cas-mediated transcriptional modulation: the therapeutic promises of CRISPRa and CRISPRi. Mol. Ther. 31 (7), 1920–1937. doi:10.1016/j.ymthe.2023.03.024
Berendzen, K. M., Sharma, R., Mandujano, M. A., Wei, Y., Rogers, F. D., Simmons, T. C., et al. (2023). Oxytocin receptor is not required for social attachment in prairie voles. Neuron 111 (6), 787–796.e4. doi:10.1016/j.neuron.2022.12.011
Bock, N., Moll, G. H., Wicker, M., Pilz, J., Rüther, E., Banaschewski, T., et al. (2004). Early administration of tiapride to Young rats without long-lasting changes in the development of the dopaminergic system. Pharmacopsychiatry 37 (4), 163–167. doi:10.1055/s-2004-827171
Boender, A. J., Boon, M., Albers, H. E., Eck, S. R., Fricker, B. A., Kelly, A. M., et al. (2023). An AAV-CRISPR/Cas9 strategy for gene editing across divergent rodent species: targeting neural oxytocin receptors as a proof of concept. Sci. Adv. 9 (22), eadf4950. doi:10.1126/sciadv.adf4950
Brusman, L. E., Sadino, J. M., Fultz, A. C., Kelberman, M. A., Dowell, R. D., Allen, M. A., et al. (2024). Single nucleus RNA-sequencing reveals transcriptional synchrony across different relationships. bioRxiv. doi:10.1101/2024.03.27.587112
Cunningham, A. M., Santos, T. L., Gutzeit, V. A., Hamilton, H., Hen, R., and Donaldson, Z. R. (2019). Functional interrogation of a depression-related serotonergic single nucleotide polymorphism, rs6295, using a humanized mouse model. ACS Chem. Neurosci. 10 (7), 3197–3206. doi:10.1021/acschemneuro.8b00638
Deng, Y., Diepstraten, S. T., Potts, M. A., Giner, G., Trezise, S., Ng, A. P., et al. (2022). Generation of a CRISPR activation mouse that enables modelling of aggressive lymphoma and interrogation of venetoclax resistance. Nat. Commun. 13 (1), 4739. doi:10.1038/s41467-022-32485-9
Donaldson, Z. R., Yang, S. H., Chan, A. W. S., and Young, L. J. (2009). Production of germline transgenic prairie voles (Microtus ochrogaster) using lentiviral vectors. Biol. Reproduction 81 (6), 1189–1195. doi:10.1095/biolreprod.109.077529
Donaldson, Z. R., and Young, L. J. (2008). Oxytocin, vasopressin, and the neurogenetics of sociality. Science 322 (5903), 900–904. doi:10.1126/science.1158668
Duke, C. G., Bach, S. V., Revanna, J. S., Sultan, F. A., Southern, N. T., Davis, M. N., et al. (2020). An improved CRISPR/dCas9 interference tool for neuronal gene suppression. Front. Genome Ed. 2, 9. doi:10.3389/fgeed.2020.00009
Gingrich, B., Liu, Y., Cascio, C., Wang, Z., and Insel, T. R. (2000). Dopamine D2 receptors in the nucleus accumbens are important for social attachment in female prairie voles (Microtus ochrogaster). Behav. Neurosci. 114 (1), 173–183. doi:10.1037/0735-7044.114.1.173
Gobrogge, K., and Wang, Z. (2016). The ties that bond: neurochemistry of attachment in voles. Curr. Opin. Neurobiol. 38, 80–88. doi:10.1016/j.conb.2016.04.011
Horie, K., Inoue, K., Suzuki, S., Adachi, S., Yada, S., Hirayama, T., et al. (2019). Oxytocin receptor knockout prairie voles generated by CRISPR/Cas9 editing show reduced preference for social novelty and exaggerated repetitive behaviors. Hormones Behav. 111, 60–69. doi:10.1016/j.yhbeh.2018.10.011
Insel, T. R., and Young, L. J. (2001). The neurobiology of attachment. Nat. Rev. Neurosci. 2 (2), 129–136. doi:10.1038/35053579
Konermann, S., Brigham, M. D., Trevino, A. E., Joung, J., Abudayyeh, O. O., Barcena, C., et al. (2015). Genome-scale transcriptional activation by an engineered CRISPR-Cas9 complex. Nature 517 (7536), 583–588. doi:10.1038/nature14136
Lim, M. M., Murphy, A. Z., and Young, L. J. (2004). Ventral striato-pallidal oxytocin and vasopressin V1a receptors in the monogamous prairie vole (Microtus ochrogaster). J. Comp. Neurol. 468, 555–570. doi:10.1002/cne.10973
Lim, M. M., and Young, L. J. (2006). Neuropeptidergic regulation of affiliative behavior and social bonding in animals. Hormones Behav. 50 (4), 506–517. doi:10.1016/j.yhbeh.2006.06.028
Livak, K. J., and Schmittgen, T. D. (2001). Analysis of relative gene expression data using real-time quantitative PCR and the 2(-Delta Delta C(T)) Method. Methods 25 (4), 402–408. doi:10.1006/meth.2001.1262
Loth, M. K., and Donaldson, Z. R. (2021). Oxytocin, dopamine, and opioid interactions underlying pair bonding: highlighting a potential role for microglia. Endocrinology 162 (2), bqaa223. doi:10.1210/endocr/bqaa223
Loth, M. K., Schmidt, J. C., Gonzalez, C. A., Brusman, L. E., Sadino, J. M., Winther, K. E., et al. (2025). Oxytocin and dopamine receptor expression: cellular level implications for pair bonding. bioRxiv. doi:10.1101/2025.03.03.640889
Maeder, M. L., Linder, S. J., Cascio, V. M., Fu, Y., Ho, Q. H., and Joung, J. K. (2013). CRISPR RNA–guided activation of endogenous human genes. Nat. Methods 10 (10), 977–979. doi:10.1038/nmeth.2598
Mali, P., Aach, J., Stranges, P. B., Esvelt, K. M., Moosburner, M., Kosuri, S., et al. (2013). CAS9 transcriptional activators for target specificity screening and paired nickases for cooperative genome engineering. Nat. Biotechnol. 31 (9), 833–838. doi:10.1038/nbt.2675
Paxinos, G., and Watson, C. (2007). Paxinos and Watson’s the rat brain in stereotaxic coordinates. 7th edn.
Pierce, A. F., Protter, D. S. W., Watanabe, Y. L., Chapel, G. D., Cameron, R. T., and Donaldson, Z. R. (2024). Nucleus accumbens dopamine release reflects the selective nature of pair bonds. Curr. Biol. 34 (3), 519–530.e5. doi:10.1016/j.cub.2023.12.041
Sadino, J. M., Bradeen, X. G., Kelly, C. J., Brusman, L. E., Walker, D. M., and Donaldson, Z. R. (2023). Prolonged partner separation erodes nucleus accumbens transcriptional signatures of pair bonding in male prairie voles. eLife 12, e80517–NA. doi:10.7554/eLife.80517
Savell, K. E., Bach, S. V., Zipperly, M. E., Revanna, J. S., Goska, N. A., Tuscher, J. J., et al. (2019a). A neuron-optimized CRISPR/dCas9 activation system for robust and specific gene regulation. eNeuro 6 (1), 0495–518. doi:10.1523/ENEURO.0495-18.2019
Savell, K. E., Sultan, F. A., and Day, J. J. (2019b). A novel dual lentiviral CRISPR-based transcriptional activation system for gene expression regulation in neurons. Bio-Protocol 9 (17), e3348. doi:10.21769/BioProtoc.3348
Savell, K. E., Tuscher, J. J., Zipperly, M. E., Duke, C. G., Phillips, R. A., Bauman, A. J., et al. (2020). A dopamine-induced gene expression signature regulates neuronal function and cocaine response. Sci. Adv. 6 (26), eaba4221. doi:10.1126/sciadv.aba4221
Scatton, B., Cohen, C., Perrault, G., Oblin, A., Claustre, Y., Schoemaker, H., et al. (2001). The preclinical pharmacologic profile of tiapride. Eur. Psychiatry 16 (S1), 29s–34S. doi:10.1016/S0924-9338(00)00526-5
Stahl, S. M. (2017). Drugs for psychosis and mood: unique actions at D3, D2, and D1 dopamine receptor subtypes. CNS Spectrums 22 (5), 375–384. doi:10.1017/S1092852917000608
Sue Carter, C., Courtney Devries, A., and Getz, L. L. (1995). Physiological substrates of mammalian monogamy: the prairie vole model. Neurosci. and Biobehav. Rev. 19 (2), 303–314. doi:10.1016/0149-7634(94)00070-H
Wang, Z., Yu, G., Cascio, C., Liu, Y., Gingrich, B., and Insel, T. R. (1999). Dopamine D2 receptor-mediated regulation of partner preferences in female prairie voles (Microtus ochrogaster): a mechanism for pair bonding? Behav. Neurosci. 113 (3), 602–611. doi:10.1037/0735-7044.113.3.602
Wei, J., Ma, L., Ju, P., Yang, B., Wang, Y. X., and Chen, J. (2020). Involvement of oxytocin receptor/erk/MAPK signaling in the mPFC in early life stress-induced autistic-like behaviors. Front. Cell. Dev. Biol. 8, 564485. doi:10.3389/fcell.2020.564485
Young, L. J. (1999). Frank A. Beach Award. Oxytocin and vasopressin receptors and species-typical social behaviors. Hormones Behav. 36 (3), 212–221. doi:10.1006/hbeh.1999.1548
Young, L. J., Lim, M. M., Gingrich, B., and Insel, T. R. (2001). Cellular mechanisms of social attachment. Hormones Behav. 40 (2), 133–138. doi:10.1006/hbeh.2001.1691
Young, L. J., and Wang, Z. (2004). The neurobiology of pair bonding. Nat. Neurosci. 7 (10), 1048–1054. doi:10.1038/nn1327
Keywords: CRISPRa, CRISPRi, somatic gene regulation, prairie voles, Oxtr, Avpr1a, Drd1, Drd2
Citation: Loth MK, Mesch KT, Herrera-Garcia C, Brusman LE and Donaldson ZR (2025) Lentiviral CRISPRa/i in the adult prairie vole brain: modulating neuronal gene expression without DNA cleavage. Front. Genome Ed. 7:1602983. doi: 10.3389/fgeed.2025.1602983
Received: 30 March 2025; Accepted: 14 May 2025;
Published: 30 May 2025.
Edited by:
Guillaume Pavlovic, PHENOMIN, Institut Clinique de la Souris (ICS), FranceReviewed by:
Daniela Avila-González, Instituto Nacional de Perinatología (INPER), MexicoWendy Portillo, National Autonomous University of Mexico, Mexico
Copyright © 2025 Loth, Mesch, Herrera-Garcia, Brusman and Donaldson. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Zoe R. Donaldson, em9lLmRvbmFsZHNvbkBjb2xvcmFkby5lZHU=