 Stephanie Juané Kennedy
Stephanie Juané Kennedy- 1Department of Haematology and Cell Biology, University of the Free State, Bloemfontein, South Africa
- 2Department of Haematology and Cell Biology, National Health Laboratory Service Universitas Academic Laboratory, Bloemfontein, South Africa
Pulmonary megakaryocytes were first identified over a century ago, but their origin is still debated. Two rare images of circulating megakaryocytes are presented to illustrate this discussion: The first shows an intact megakaryocyte with abundant cytoplasm and a hyperlobulated nucleus on a peripheral smear made from blood drawn through a Hickman line in a child with B-cell acute lymphoblastic leukaemia. The morphology of this megakaryocyte is contrasted with that of the second image, from a patient with primary myelofibrosis. This perspective focuses on the first image, which illustrates several hypotheses regarding the origin and function of pulmonary megakaryocytes. According to the most widely accepted theory, this megakaryocyte may have originated in the bone marrow (BM), entered the venous circulation, and migrated to the lungs, where it was aspirated from the superior vena cava before being trapped in the pulmonary vasculature to exert its incompletely understood functions. In this child with acute leukaemia, it could have been a role-player the cancer immune response. Alternatively, pulmonary megakaryocytes may have been compensating for the displacement of platelet-producing megakaryocytes by blasts in the BM. While a single anecdotal image cannot provide mechanistic evidence, this megakaryocyte, when interpreted in its clinical context, provides visual support for (1) the hypotheses that at least some pulmonary megakaryocytes have a BM origin, (2) the theory that lung megakaryocytes may augment platelet production during thrombocytopenic states, and (3) chemotherapy-induced megakaryocyte release from the BM. It serves as a reminder that we should continue to draw insights from clinical observations.
1 Introduction
A megakaryocyte is a rare finding on the peripheral blood (PB) smear (1, 2). Mature megakaryocytes with copious cytoplasm are particularly unusual, as circulating megakaryocytes are nearly always (99%) detected as bare nuclei devoid of cytoplasm (2). Since the concentration of circulating megakaryocytes, on average, is only between five and seven per millilitre, they are more likely to be seen in buffy coat preparations (2). Megakaryocyte numbers are increased in the blood of neonates and young infants, postpartum, postoperatively, and in patients with infection, inflammation, disseminated intravascular coagulation, and malignancy, particularly myeloproliferative neoplasms (2). The detection of intact megakaryocytes with plentiful cytoplasm is increased in the blood of infants and patients with primary myelofibrosis (PMF) (2).
Two rare images of circulating megakaryocytes are presented here: The first, from the Hickmann line of a young child with B-cell acute lymphoblastic leukaemia (B-ALL), and the second, from a man with PMF. This perspective article focuses on the first image, which is discussed in relation to the emerging body of literature on the origin and roles of pulmonary megakaryocytes.
Megakaryocytes are classically viewed as large bone marrow (BM)-resident, platelet-producing cells (3). However, the heterogeneous functions of megakaryocytes and platelets are increasingly recognized (3). Extramedullary (EM) megakaryocytes have been demonstrated in several organs, but most research has focused on megakaryocytes in the lungs (3). The developmental origin of pulmonary megakaryocytes has been debated for the last century (migration from the BM vs. in situ differentiation). Although it is most widely accepted that lung megakaryocytes are ‘seeded’ from the BM, this theory remains hypothetical (4). While rapid technological advances and emerging research tools are improving our understanding of the origin and function of pulmonary megakaryocytes, Malara and Balduini (1) emphasise that we should continue to draw insights from clinical observations.
2 A megakaryocyte in the peripheral blood: evidence for pulmonary migration?
A nine-month-old girl was diagnosed with KMT2A-germ line infant B-ALL and chemotherapy was initiated chemotherapy. Her full blood count (FBC) at diagnosis showed severe thrombocytopenia (platelet count 17 x 109/L), anaemia (Hb 4.2 g/dL), and a leukocytosis (WCC 308.12 X 109/L) with 89% blasts. At the age of 17-months, she was receiving chemotherapy through a Hickmann line, when an FBC and smear were performed on a PB specimen that was drawn through the central venous catheter. The FBC showed a mild thrombocytosis (platelet count 463 x 109/L), mild anaemia (haemoglobin [Hb] of 10.7 g/dL), and leukopenia (white cell count [WCC] 0.76 x 109/L). The PB smear showed no blasts. Incidentally, a single megakaryocyte was observed at the longitudinal edge of the smear (Figure 1A). A buffy coat smear was subsequently prepared from the leukocyte-enriched layer after centrifugation, which yielded another megakaryocyte bare nucleus at the feathered edge. The child completed three years of chemotherapy and remains in complete remission two years later.
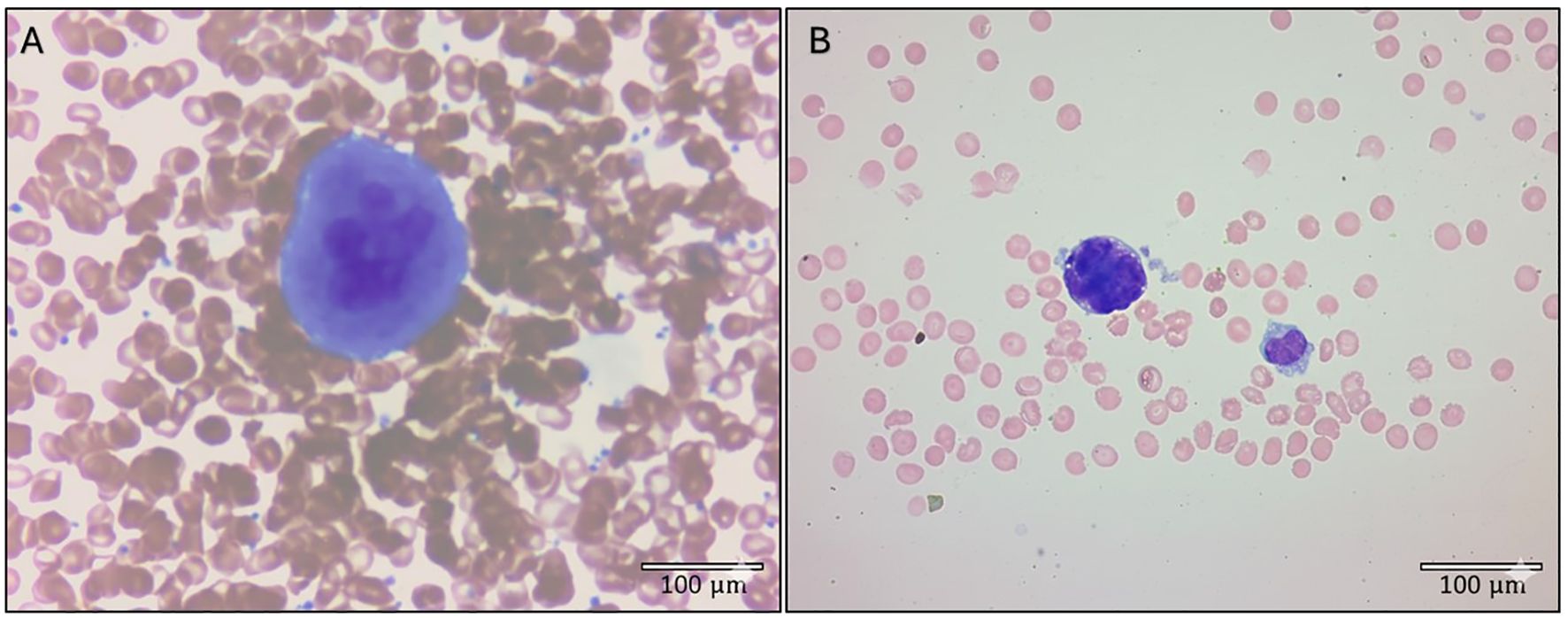
Figure 1. Rare circulating megakaryocytes detected on peripheral blood smears of two patients with haematological neoplasms. (A) Mature megakaryocyte from a child receiving chemotherapy for B-cell acute lymphoblastic leukaemia, exhibiting a characteristic lobulated nucleus, abundant cytoplasm, and large size. This smear was made from blood that was drawn through a Hickmann line. (B) Megakaryocyte from a man with primary myelofibrosis, displaying features of maturity. The nucleus is almost completely devoid of cytoplasm with only a few budding platelets seen. Both smears were stained with Wright’s stain. Photomicrographs were taken with 50× (A) and 100× (B) objectives. Images were adjusted for exposure and contrast; adjustments were applied uniformly across each image. Scale bars are included in each panel.
The circulating megakaryocyte from this child in Figure 1A demonstrates abundant cytoplasm and a hyperlobulated nucleus. It resembles the typical cytologic appearance of a mature megakaryocyte that can be appreciated on a BM aspirate. However, it is highly unusual to see a megakaryocyte with these features on a PB smear (2). In contrast, Figure 1B shows a megakaryocyte on the longitudinal edge of a PB smear from a patient with primary myelofibrosis. It is almost completely bereft of cytoplasm with a few budding platelets seen. These features are more in keeping with the ‘bare nucleus’ morphology that is typically observed when megakaryocytes are seen on a PB smear (2).
The observation that a circulating megakaryocyte was detected in venous blood from a Hickmann line in this child, is noteworthy. The tip of this indwelling central venous catheter is located at the entry of the superior vena cava (SVC), proximal to the pulmonary vasculature. It raises the question if this megakaryocyte was perhaps ‘intercepted’ on its way to the lungs following its chemotherapy-induced release from the BM into the circulation?
3 Discussion
Our canonical understanding of megakaryocyte biology has long been dominated by the classical view of megakaryocytes as large polyploid cells that reside in the BM where they produce platelets. However, the heterogeneity of megakaryocytes is increasingly recognized (3). An emerging body of knowledge has shown that megakaryocytes and platelets have multifaceted roles beyond haemostasis; including inflammation, immunity, and cancer metastases (3–5). Functional subsets of megakaryocytes have subsequently been identified using single cell RNA sequencing techniques and transcriptomic analysis (6, 7). These subsets include active cycling-, hematopoietic stem cell (HSC) niche-, inflammatory response-, and thrombopoietic megakaryocytes (3, 6). Extramedullary (EM) megakaryocytes have been demonstrated in the lungs, spleen, liver, and blood, as well as the foetal liver and yolk sac (3, 4). Most research on EM megakaryocytes has focused on pulmonary megakaryocytes.
In their review on the origin of pulmonary megakaryocytes, Livada et al. (4) discussed the historical perspective (BM emigration and pulmonary vascular entrapment) as well as emerging paradigms (in situ differentiation from local haematopoietic progenitors) and concluded that the debate remains unsettled. Accordingly, three models for the origin of pulmonary megakaryocytes were proposed (4). The first assumes that they have a BM origin, while the second and third models suggest a BM-independent origin.
Until recently, the hypothesis that pulmonary megakaryocytes originate in the BM and migrate through the blood to the lungs, was assumed in the literature. It was first proposed by Aschoff (8) as early as 1893. However, direct evidence for this model is still lacking (4). This historical perspective was largely influenced by physical observations of these large cells within the lung’s vascular network, which led to the assumption that their presence was the result of passive mechanical entrapment in the narrow capillaries of the pulmonary circulation, and that this resulted in the release of platelets into the circulation (9–17). Although most recent experimental studies on pulmonary megakaryocytes have been performed in murine models, human evidence for the existence of lung megakaryocytes have existed for decades. In 1948, Brill and Halpern (10) consistently identified megakaryocytes in the lungs throughout 50 human necropsies. The megakaryocyte that was detected in a central venous line specimen from a live human, shown here, brings to mind the 1993 study by Levine et al. (13), who isolated megakaryocytes from the blood entering and leaving the lungs of cardiac catheterization patients, demonstrating a higher concentration of intact mature megakaryocytes in the pulmonary artery compared with a reduced number of megakaryocytes with partial cytoplasmic content in aortic specimens. These observations supported the theory that BM megakaryocytes are trapped in the pulmonary vasculature where platelets are produced.
The view of the lung as a filter has overlooked any potential intrinsic haematopoietic or immunologic function of pulmonary megakaryocytes. Accordingly, Livada et al. (4) incorporated these emerging paradigms into their second and third models, and argued that future studies should focus on whether there is BM-independent megakaryopoieis in the lungs, either from a local HSC pool, or from a self-renewal mechanism analogous to tissue-resident haematopoietic cells (4). The second model is based on a pivotal study by Lefrancais et al. (16) which suggested that pulmonary megakaryocytes are derived from an HSC pool in the lungs. In addition to identifying a hematopoietic progenitor pool in the extravascular spaces of mouse lungs that was capable of megakaryocyte reconstitution in the BM following transplantation, Lefrancais et al. (16) identified over 700 genes that were differentially expressed between lung and BM megakaryocytes using RNA sequencing. These findings suggested that the lung is a dynamic hematopoietic tissue with its own developmental capacity, rather than merely a passive site for migrating MKs. The third hypothesis is that lung megakaryocytes are self-renewing and have an HSC-independent embryonic origin. This model, which suggests that self-renewing pulmonary megakaryocytes may be derived from the yolk sac during early embryonic development, relies on evidence for HSC-independent haematopoiesis within the mouse embryo, as well as various studies that identified tissue-resident haematopoietic cells; a concept that may deserve further study to identify HSC-independent tissue-resident megakaryocytes (4).
The quantitative contribution of pulmonary megakaryocytes to the circulating platelet pool is a central controversy in the field. Scattered reports over the last century have challenged the dogma that thrombopoiesis occurs primarily in the BM, suggesting that megakaryocytes in the lungs play an active role (9, 13–15, 18, 19). It was only in 2017 that Lefrancais et al. (16) provided the first direct visualization of platelet release by megakaryocytes in the pulmonary vasculature of mice and estimated that 50% of platelet biogenesis occurs in the lungs. More recent data showed this to be an overestimation and reasserted the BM as the primary site for thrombopoiesis (1, 5, 20). Asquith et al. (5) assessed the relative number, phenotype and ex-vivo proplatelet formation capacity of megakaryocytes from different tissues in mice. This study concluded that the contribution of pulmonary and splenic megakaryocytes to the circulating platelet mass was only marginal compared with BM megakaryocytes (5). In 2024, Livada et al. (20) went further to show that lung megakaryocytes may increase their platelet production capacity in response to thrombocytopenia. Using in vivo labelling and tracing models in mice, Livada et al. (20) demonstrated that lung megakaryocytes produced 10% of circulating platelets under homeostatic conditions, but in acute and chronic infection-induced models of thrombocytopenia, increased their output to 20%.
Defining megakaryocyte origin may aid in understanding how pulmonary megakaryocytes acquire an immune-modulatory phenotype, and its impacts in health and disease (4). The emerging knowledge of lung megakaryocyte biology opens numerous avenues for future clinical research, which is likely to be driven by technological advances. In vitro models like 3D organoids are increasingly used and may be capable of replicating the complex lung microenvironment to study pulmonary megakaryocyte behaviour outside the body. This will allow us to determine how recent pivotal findings on the origin and function of pulmonary megakaryocytes in animal models apply to human physiology and will lay the foundation for further exploration of these promising cellular targets for diagnostic and therapeutic interventions. Malara and Balduini (1), in a comment on the study by Asquith et al. (5), agree that rapid technologic advances and emerging tools are improving our understanding of the intricate mechanisms of pulmonary megakaryocyte biology, but emphasize the value of drawing insights from clinical observations. For example, from our assessment of PB smears, we know that circulating megakaryocytes are a rarity in health, but that their detection increases under pathologic conditions such as myeloproliferative neoplasms (1, 2). It should be noted that relying on morphologic observations may underestimate these numbers, and that phenotypic characterisation of circulating megakaryocytes using flow cytometry or immunostaining would provide a more accurate estimate. Puhm et al. (3) argue that the site of blood collection may also influence the rate at which megakaryocytes are observed; e.g. the likelihood of detecting a megakaryocyte in blood that was drawn from the vena cava (which drains BM-rich areas like the vertebrae and pelvis) may be higher than if the blood was drawn from a forearm vein (3).
The rare image of a mature circulating megakaryocyte in Figure 1A may be the kind of the clinical observation that Malara and Balduini (1) were referring to. Here, the hypothesis is that this megakaryocyte originated in the BM, entered the venous circulation, and migrated to the lungs, where it was aspirated from the SVC before it could be trapped in the pulmonary vasculature to exert its incompletely understood functions. In a child with acute leukaemia, it may have been on its way to participate in the cancer immune response. Alternatively, pulmonary megakaryocytes may have been compensating for the displacement of platelet-producing megakaryocytes by blasts in the BM.
The anecdotal nature of this clinical observation limits the strength of these hypotheses. And while a single image cannot provide mechanistic evidence, this PB megakaryocyte, when interpreted in its clinical context, does provide visual support for (1) the hypothesis that at least some pulmonary megakaryocytes have a BM origin (4), (2) the recent theory that lung megakaryocytes may augment platelet production during thrombocytopenic states (9), and (3) the established role of chemotherapy-induced megakaryocyte release from the BM.
Morphologic observations such as these should prompt further research into the mechanisms by which pulmonary megakaryocytes could potentially compensate for failed BM thrombopoiesis in patients with haematologic neoplasms or those receiving chemotherapy. The knowledge gained may ultimately be translatable to the treatment of cancer-related thrombocytopenia. Targeting lung resident progenitor cells to augment platelet production could ultimately be a revolutionary approach to treat thrombocytopenia in both neoplastic and non-neoplastic diseases.
Regardless of its ontogeny or function, a megakaryocyte on the PB smear remains a rare finding.
Data availability statement
The original contributions presented in the study are included in the article. Further inquiries can be directed to the corresponding author.
Ethics statement
The studies involving humans were approved by Health Sciences Research Ethics Committee, University of the Free State. Ethical clearance no.:UFS-HSD2025/0153/2503. The studies were conducted in accordance with the local legislation and institutional requirements. The human samples used in this study were acquired from a by- product of routine care or industry. Written informed consent for participation was not required from the participants or the participants’ legal guardians/next of kin in accordance with the national legislation and institutional.
Author contributions
SK: Conceptualization, Writing – original draft, Writing – review & editing.
Funding
The author(s) declare financial support was received for the research and/or publication of this article. Funding for the publication of this article was received from the University of the Free State.
Conflict of interest
The author declares that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Generative AI statement
The author(s) declare that no Generative AI was used in the creation of this manuscript.
Any alternative text (alt text) provided alongside figures in this article has been generated by Frontiers with the support of artificial intelligence and reasonable efforts have been made to ensure accuracy, including review by the authors wherever possible. If you identify any issues, please contact us.
Publisher’s note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
References
1. Malara A and Balduini A. Megakaryocytes in the lung: guests or ghosts? Blood. (2024) 143:192–3. doi: 10.1182/blood.2023022897
2. Bain BJ and Leach M. Blood Cells (2025). Wiley. Available online at: https://www.perlego.com/book/5169472 (Accessed October 16, 2025).
3. Puhm F, Laroche A, and Boilard E. Diversity of megakaryocytes. Arterioscler Thromb Vasc Biol. (2023) 43:2088–98. doi: 10.1161/ATVBAHA.123.318782
4. Livada AC, Pariser DN, and Morrell CN. Megakaryocytes in the lung: History and future perspectives. Res Pract Thromb Haemost. (2023) 7:100053. doi: 10.1016/j.rpth.2023.100053
5. Asquith NL, Carminita E, Camacho V, Rodriguez-Romera A, Stegner D, Freire D, et al. The bone marrow is the primary site of thrombopoiesis. Blood. (2024) 143:272–8. doi: 10.1182/blood.2023020895
6. Sun S, Jin C, Si J, Lei Y, Chen K, Cui Y, et al. Single-cell analysis of ploidy and the transcriptome reveals functional and spatial divergency in murine megakaryopoiesis. Blood. (2021) 138:1211–24. doi: 10.1182/blood.2021010697
7. Liu C, Wu D, Xia M, Li M, Sun Z, Shen B, et al. Characterization of cellular heterogeneity and an immune subpopulation of human megakaryocytes. Adv Sci (Weinh). (2021) 8:e2100921. doi: 10.1002/advs.202100921
8. Aschoff L. Ueber capilläre Embolie von riesenkernhaltigen Zellen. Archiv f Pathol Anat. (1893) 134:11–25. doi: 10.1007/BF01924893
9. Howell WH and Donahue DD. The production of blood platelets in the lungs. J Exp Med. (1937) 65:177–203. doi: 10.1084/jem.65.2.177
10. Brill R and Halpern MM. The frequency of megakaryocytes in autopsy sections. Blood. (1948) 3:286–91. doi: 10.1182/blood.V3.3.286.286
11. Kaufman RM, Airo R, Pollack S, Crosby WH, and Doberbeck R. Origin of pulmonary megakaryocytes. Blood. (1965) 25:767–75. doi: 10.1182/blood.V25.5.767.767
12. Pedersen NT. Occurrence of megakaryocytes in various vessels and their retention in the pulmonary capillaries in man. Scand J Haematol. (1978) 21:369–75. doi: 10.1111/j.1600-0609.1978.tb00381.x
13. Levine RF, Eldor A, Shoff PK, Kirwin S, Tenza D, and Cramer EM. Circulating megakaryocytes: delivery of large numbers of intact, mature megakaryocytes to the lungs. Eur J Haematol. (1993) 51:233–46. doi: 10.1111/j.1600-0609.1993.tb00637.x
14. Zucker-Franklin D and Philipp CS. Platelet production in the pulmonary capillary bed: new ultrastructural evidence for an old concept. Am J Pathol. (2000) 157:69–74. doi: 10.1016/S0002-9440(10)64518-X
15. Fuentes R, Wang Y, Hirsch J, Wang C, Rauova L, Worthen GS, et al. Infusion of mature megakaryocytes into mice yields functional platelets. J Clin Invest. (2010) 120:3917–22. doi: 10.1172/JCI43326
16. Lefrançais E, Ortiz-Muñoz G, Caudrillier A, Mallavia B, Liu F, Sayah DM, et al. The lung is a site of platelet biogenesis and a reservoir for haematopoietic progenitors. Nature. (2017) 544:105–9. doi: 10.1038/nature21706
17. Dejima H, Nakanishi H, Kuroda H, Yoshimura M, Sakakura N, Ueda N, et al. Detection of abundant megakaryocytes in pulmonary artery blood in lung cancer patients using a microfluidic platform. Lung Cancer. (2018) 125:128–35. doi: 10.1016/j.lungcan.2018.09.011
18. Slater DN, Trowbridge EA, and Martin JF. The megakaryocyte in thrombocytopenia: a microscopic study which supports the theory that platelets are produced in the pulmonary circulation. Thromb Res. (1983) 31:163–76. doi: 10.1016/0049-3848(83)90017-8
19. Trowbridge EA, Martin JF, and Slater DN. Evidence for a theory of physical fragmentation of megakaryocytes, implying that all platelets are produced in the pulmonary circulation. Thromb Res. (1982) 28:461–75. doi: 10.1016/0049-3848(82)90163-3
Keywords: circulating megakaryocyte, peripheral blood megakaryocyte, pulmonary megakaryocytes, lung megakaryocytes, thrombopoiesis
Citation: Kennedy SJ (2025) A megakaryocyte in the peripheral blood: evidence for pulmonary migration? Front. Hematol. 4:1679951. doi: 10.3389/frhem.2025.1679951
Received: 05 August 2025; Accepted: 30 October 2025;
Published: 20 November 2025.
Edited by:
Ahmed Esmat Abdel Moneim, Helwan University, EgyptReviewed by:
Pei Xiong Liew, Augusta University, United StatesNikolaos Sousos, University of Oxford, United Kingdom
Craig Morrell, University of Rochester Medical Center, United States
Copyright © 2025 Kennedy. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Stephanie Juané Kennedy, S2VubmVkeVNKQHVmcy5hYy56YQ==; c3RlcGhhbmllLmtlbm5lZHlAbmhscy5hYy56YQ==