 Ashley Fischer
Ashley Fischer Jay P. Singh
Jay P. Singh Jonathan Van Hamme
Jonathan Van Hamme Eric Bottos
Eric Bottos Lauchlan H. Fraser
Lauchlan H. Fraser- Thompson Rivers University, Kamloops, BC, Canada
Mining activities are often severely disruptive to the landscape, and a significant barrier to reclamation following mining operations is the lack of quality topsoil. This project addresses knowledge gaps in the industry by exploring the compositional nature of topsoil stockpiles and their ability to facilitate post-mining revegetation after long-term storage. To do this, we conducted a microbial profiling of two topsoil stockpiles in the interior of British Columbia, Canada. Both stockpiles show depleted soil quality and significant changes compared to reference soils. Notably, there were declines in microbial diversity and significant shifts in community structure with increasing stockpile depths in one of the stockpiles. These results highlight the influence of topsoil-stockpile height on microbial communities in the soil, which ultimately influences the success of restoration. This research can help the industry to optimize restoration and expedite recovery in their mine-closure practices and provides insights into the general structure of the microbiome existing across a gradient in severely disturbed mining soils.
Introduction
The mining industry in British Columbia is an essential source of resources and is a key contributor to the economy. As of 2020, there were 16 major metal and coal mines, and during 2020, the mining industry produced approximately $7.3 billion and provided more than 30,000 jobs (Ministry of Energy, Mines and Low Carbon Innovation, 2020) mining activities typically include mineral exploration, mineral development, mine production, and mineral processing. These disturbance events cause lasting negative environmental impacts such as ecosystem degradation, habitat destruction, pollution, and loss of soil carbon (Tripathi et al., 2016). Federal and provincial regulations ensure that the restoration or reclamation of landscapes is conducted at mine sites to repair disturbances and return the land to a sustainable ecosystem with a historical level of productivity. Mining is particularly damaging to ecosystems because soils are stripped from landscapes and require reconstruction, which takes hundreds or thousands of years if left to natural processes (Bradshaw, 1997).
Topsoil is a valuable bioactive substrate hosting a wide variety of organisms, including plants, microorganisms, animals, viruses, and protists. Specifically, topsoil is defined as the uppermost part of the soil profile, typically ranging in depth from 7 to 25 cm, and contains the highest concentration of organic matter and microorganisms (Darmody et al., 2009). Preserving topsoil is valuable for mining operations because it could be used later for ecosystem rehabilitation. As such, stockpiles of topsoil are stored for future use. Stockpiling topsoil can be used post-mining to provide nutrients, structure, seeds, and to amend waste materials on site. However, long-term storage of topsoil has been shown to deteriorate soil health by altering its geochemical properties and microbial communities (Ghose and Kundu, 2004; Gorzelak et al., 2020; Harris et al., 1989; Mummey et al., 2002). For example, Boyer et al. (2011) observed that topsoil stockpiles contained compacted and anaerobic soil below one meter, resulting in a low abundance and diversity of earthworms. Moreover, a laboratory study found that stockpiling topsoil caused a decrease in soil organic matter degradation rate, implying diminished microbial activity (Felton and Taraba, 1994).
Soil health is key in restoration success and ecosystem functioning; soil organic matter, soil nutrients, and soil microorganisms play major roles in maintaining healthy, sustainable ecosystems. Currently, only a small proportion of microbial diversity has been identified (Rodrigues et al., 2025). Soil microorganisms provide many ecosystem functions, including roles as biological regulators, chemical cyclers, and ecosystem engineers (Saccá et al., 2017), but are extremely sensitive to disturbances and may take decades for recovery (Bastida et al., 2008; Mummey et al., 2002). Therefore, soil microorganisms are a key component to steering the recovery of disturbed ecosystems, and research on soil microbial functions and communities in various ecosystems and scales is essential.
Special attention should be paid to soil microbial communities in the revegetation of mine soil, where severe, large-scale disturbance occurs, particularly because soil microbes are large drivers in organic matter decomposition and nutrient cycling of macro- and micronutrients critical for plant growth and establishment (Gruber, 2015). Furthermore, soil bacteria and fungi form close relationships with plants, providing services such as nutrient acquisition and protection against environmental stress and pathogens. For example, Chen et al. (2007) found that mycorrhizal colonization significantly increased plant growth in copper (Cu) contaminated soils. This was thought to be a result of increased phosphorus (P) acquisition and decreased Cu concentrations in plant roots. Using their extended, root-like hyphae network, arbuscular mycorrhizal fungi (AMF) can acquire P in areas that are difficult to access. Additionally, AM fungi secrete organic acids, activating unavailable P sources, and can increase the expression of phosphate transporter genes in plants (Wang et al., 2017). Therefore, soil bacteria and fungi are paramount to the restoration process of a landscape and to maintaining a sustainable and resilient ecosystem. The inclusion of microbial community composition data when monitoring soil quality provides crucial insights and a more accurate understanding of how land management impacts the soil ecosystem.
Microorganisms cultured in the lab for study and characterization have been occurring since the 1870s. However, the use of sequencing in the 2000s highlighted that the number of observable, culturable bacteria and fungi are just a tiny fraction of those that are present but unseen (Nilsson et al., 2019). Therefore, molecular techniques, including high-throughput DNA sequencing, have been used as an alternative to culturing and characterization of microorganisms. Characterizing soil microbial communities in mining operations has the potential to aid our ability to restore degraded sites by providing insights and improving our understanding of how land management impacts the soil ecosystem, determining the responses of soils to disturbances, and assessing ecosystem sustainability.
Understanding how microbial communities respond to disturbance and how they recover is critical for optimizing restoration practices. Additionally, at a time when high-throughput sequencing technology has provided novel information about microbial taxa, we can characterize the microbial communities throughout the topsoil stockpile depths, which will provide new information to the field of ecological restoration. Although it is well understood that soil microbial communities are essential for many ecosystem functions, it is less well known how soil microbial activities and composition are impacted by mining disturbance. There is a growing body of literature that has examined the impact of stockpiling on microbial communities. Harris et al. (1989) found that a reduction in aerobic bacteria and biomass with increasing stockpile depth. Ezeokoli et al. (2019) report a decline in soil enzymatic activity and population of beneficial microbial taxa with depth. Similarly, Cabrera-Hernandez et al. (2025) and Hernandez et al. (2024) reveal a shift in bacterial and fungal community composition in stockpiles compared to reference sites. While these studies provide an insight into microbial communities in the shallower stockpiles, very little is known about microbial communities in deeper stockpiles (> 300 cm). Examining soil microbial community composition across a known environmental gradient from the top to the bottom of topsoil stockpiles has the potential to provide insights into factors that shape the microbiome in stored soils and address knowledge gaps. This study aims to improve our understanding of how bacterial and fungal communities in topsoil respond to soil disturbance from stripping, piling, and long-term storage on mine sites by providing soil microbial community composition across a depth gradient. Research investigating soil responses over varying disturbance events and across a range of ecosystem types is critical to developing a unifying theory of ecosystem resilience and recovery. This is especially critical at a time when anthropogenic forces, including climate change, are causing increasingly extreme changes to the environment.
Methods
Study sites
New Afton Mine
New Gold’s New Afton copper-gold mine (50.654442, −120.509320) was located approximately 10 kilometers west of Kamloops in British Columbia’s Southern Interior. It is situated within the historic Afton Mine, formerly owned by Afton operating Corp. New Afton began commercial production in July 2012 and is the largest underground hardrock mine in Canada. It comprises of underground workings, historic support facilities, a historic open pit, a concentrator, and a tailings facility. The end land use objective is to return the ecosystem to native grasslands that support wildlife and traditional hunting opportunities by First Nations. New Afton is located within the traditional territories of the Tk’emlúps and Skeetchestn Bands. These bands are part of the larger cultural group known as the Secwépemc or Shuswap First Nation. Additionally, New Afton is in the Bunchgrass (BGxw1) biogeoclimatic zone at approximately 700 m in elevation. The Biogeoclimatic Ecosystem Classification (BEC) system in British Columbia incorporates information on climate, soils, and vegetation to provide a framework for management practices (Meidinger and Pojar, 1991). The BGxw1, commonly known as the “middle grasslands”, is dominated by bluebunch wheatgrass, Junegrass, big sagebrush, and rabbit brush (Lloyd et al., 1991). The primary soil orders within the mine site include Tranquille, Trapp Lake, Godey, and Timber. These soils are moderately saline and calcareous and are dominated by Orthic Brown and Dark Brown Chernozems with one occurrence of an Eluviated Eutric Brunisol soil (Government of British Columbia, 2018). The tailings in the New Afton mine are alkaline (pH >8.5) and are high in copper and molybdenum.
QR mill
Barkerville Gold Mines Ltd. (BGM) (formally International Wayside Gold Mines) (52.670306, −121.783556) is a Canadian company headquartered in Toronto, Ontario. Barkerville Gold Mines Ltd. BGM has various gold mines around the Barkerville-Wells area. Barkerville Gold Mines’ Quesnel River (QR) mill in Cariboo is located approximately 80 km east of the city of Quesnel in the southern interior of B.C. Ore, concentrate, and waste rock from the Bonanza Ledge Mine and Cariboo Gold Mine are transported, stored, and processed in the QR mill. The current reclamation goals set for QR mill are to restore the landscape so that it does not require further human intervention. QR mill is situated in the traditional territories of the Secwépemc or Shuswap First Nation and lies within the moist, warm Sub-Boreal Spruce (SBSmw) BEC zone based on the BC provincial BEC system. The SBSmw is part of the Canadian Boreal Forest Region and Quesnel Highland (mean annual precipitation ranges from 440–900 mm) and is dominated by Douglas-fir, red-stemmed feathermoss, knights plume, hybrid white spruce, subalpine fir, and electrified cat’s-tail moss (Annas and Coupé, 1979). The QR mill and surrounding area are dominated by Bedenesti, Deserters, and Dominion soil orders. These soils consist primarily of Brunisolic Gray Luvisols and Luvisolic Humo-Ferric Podzols that were deposited by glacial ice (Government of British Columbia, 2018).
Soil sampling
New Afton Mine
New Afton has a 6-year-old, 25-meter-deep topsoil stockpile with approximately 250,600 m3 of topsoil materials. Because additional topsoil materials have been added throughout the mine life, the oldest soil is at the bottom of the stockpile, and the youngest soil is at the surface. Four soil cores were extracted via solid stem auger drilling by Geotech Drilling Ltd, provided by New Afton. during September 26th and 27th of 2018, with each core being approximately 3 meters apart. The first 1.53 m was sampled in 0.3 m increments, then at 0.3 m intervals until the bottom at 13.7 m. Thus, sampling depths were at 0.3 m, 0.6 m, 0.9 m, 1.2 m, 1.5 m, 3.0 m, 4.6 m, 6.1 m, 7.6 m, 9.1 m, 10.7 m, 12.2 m, and 13.7 m. The outer 1 cm of the soil core was discarded to ensure that the upper layers did not contaminate the collected soil. Soil was placed into two 1 L Whirl-Pak® bags and Falcon® tubes as they were pulled up soil from the stockpile. Post-collection, samples were combined by depth intervals of the stockpile as follows: 0.0–0.6 m, 0.61–1.5 m, 1.5–6.1 m, and 6.11–13.72 m. The soil stockpile at the time of sampling was sparsely vegetated with grasses and weedy species, likely from natural regeneration. A nearby grassland site was sampled as a reference site, where approximately 6 kg of soil from the top 10 cm was collected using a trowel. The Whirl-Pak® samples were stored in a −20°C freezer at the Research Greenhouse, and the Falcon® tube samples were stored in a −80°C freezer in the TRUGen laboratory at Thompson Rivers University (TRU) until analysis.
QR mill
QR mill has a 20-year-old, 6-meter-deep topsoil stockpile. It is a combination of organic soil and general till soil stripped from the surface layers. The stockpile was intended to cover and re-contour during post-mining reclamation. Sampling at the QR mill of Barkerville Gold Mines Ltd. was completed in May 2019. Three soil pits approximately 100 m apart were dug using an excavator in May 2019 to access various layers of the stockpile from the surface to a depth of 575 cm. In the field, soil was placed into two 1 L Whirl-Pak® bags and Falcon® tubes as soil was removed from the stockpile. The stockpile at the time of sampling had vegetation cover, including cottonwood stands in the northwest corner. An adjacent undisturbed forested site was sampled as a reference site, where approximately 16 kg of soil from the top 10 cm was collected. The soil samples at QR mill were collected at different depths than New Afton due to operational constraints. As such, the samples were lumped differently. At QR mill, the samples were aggregated in groups: 0–0.6 m, 0.75–2.6 m, 3.5–3.9 m, and 5.0–5.75 m. After collections, the Whirl-Pak® samples were stored in a −20°C freezer at the TRU Research Greenhouse, and the Falcon® tube samples were stored in a −80°C freezer in the TRUGen laboratory at TRU until analysis.
Soil geochemistry
Soil samples were prepared for geochemical analyses by drying them at 70°C for 24 hours to constant weight. The samples were then sieved using a 1 mm sieve and ground to a fine powder using a mortar and pestle. The sieved samples were then sent to the Analytical Laboratory at the Ministry of Environment and Climate Change Strategy in Victoria, B.C., for analysis. The total soil Al, B, Ca, Cu, Fe, Mg, Mn, P, K, S, and Zn were determined using inductively coupled plasma–optical emission spectrometry (ICP-OES) after microwave digestion in an acid mixture (Sandroni, 2003). Available P was measured after extraction via Bray P-1 extraction and UV spectrophotometry (Bray and Kurtz, 1945). Available NH4-N and NO3-N were quantified in an autoanalyzer after extraction in 1M potassium chloride (Kachurina et al., 2000). Organic matter was measured in-house via loss-on-ignition (Singh et al., 2019b). Loss-on-ignition was calculated by weighing approximately 1.5 g of soil into pre-weighed tins and then heating at 500°C for 5 hours. pH and EC were determined using a Palintest® 800 meter. Total Cand N amounts were measured in-house with a ThermoScientific CHNS Elemental Analyzer.
DNA extraction and sequencing
The soil microbial community composition in each sample was characterized for both fungal and bacterial OTUs in the Applied Genomics Laboratory at TRU. Deoxyribose nucleic acids (DNA) from the soil samples were extracted using the MagAttract PowerSoil DNA Kit (Qiagen Inc.). Extractions were followed by DNA quantification using Qubit™ dsDNA HS Assay Kit (Thermo Scientific). The set of primers: 341F and 806R, was used to amplify the 16S rRNA gene (Ramirez et al., 2017), and the set of primers: ITS86F and ITS4R were used to amplify the internal transcribed spacer (ITS) region between the 5.86S rRNA gene and the 28S rRNA genes (Vancov and Keen, 2009). Samples were run through a second round of polymerase chain reaction (PCR), including the addition of barcoded primers. The thermocycler program for the first round bacterial and fungal amplicon generation consisted of: 95°C for 4 min followed by 25 cycles of 95°C for 30 seconds, 53.4°C for 45 seconds, and 72°C for 2 minutes, with a final extension at 72°C for 5 minutes. For generating sequencing barcode-adapted bacterial and fungal amplicons, the thermocycler program was the same except that 20 cycles were used with an annealing temperature of 65°C.
After the first and second rounds of PCR, the amplicons were purified using the Inovant and AgenCourt AMPure (Beckman Coulter Inc.) magnetic beads before sequencing. Barcoded amplicons were quantified using a Qubit dsDNA HS Assay Kit (Thermo Fisher Scientific, Mississauga, ON) and visualized on an agarose gel. The samples were extracted from the agarose gel using the E.Z.N.A Gel Extraction Kit (Omega Bio-Tek) and were pooled together in equimolar amounts. After pooling, the samples were sequenced on an Ion S5XL (ThermoFisher Scientific).
Sequence processing and statistical analysis
Raw sequences were demultiplexed in AMPtk 1.5.5 using the amptk ion script with default settings and –trim-len set to 350 bases. Demultiplexed data were imported into Qiime 2 with the qiime tools import script set with –input-format SingleEndFastqManifestPhred33V2 (Bolyen et al., 2019). Denoising was done with DADA2 denoise-single with max-ee set to 1.0 (p-trunc-len set to 0 because the reads were previously trimmed in AMPtk) (Callahan et al., 2016). The Greengenes2 database (version 2022.10) (McDonald et al., 2024) was used to classify the 16S rRNA gene sequences for taxonomic classification. For fungi, version 9 of the Qiime2-compatible UNITE reference database was used to classify the ITS sequences. After processing the sequencing data, any samples with fewer than 1000 reads were dropped. Rare ASVs (read less than 0.005%) were also dropped from the count table. After dropping low read samples and rare ASVs, New Afton had 1675 bacterial and archaeal ASVs and 3166 fungal ASVs. QR mills had 1478 bacterial and archaeal ASVs and 2565 fungal ASVs. Further, we built phylogenetic trees for 16S and ITS representative sequences for downstream analysis. The representative ASV sequences (both 16S and ITS) were first aligned with Mafft (version 7) (Katoh and Standley, 2013), and a maximum likelihood tree was generated using FastTree 2 (Price et al., 2010). The downstream statistical analyses for 16S and ITS data were the same for both sites and are outlined next. Species richness (SR) for 16S and ITS sequences for New Afton and QR mill was measured, and Faith’s phylogenetic diversity (PD) was measured using the count data and the phylogenetic trees. PD and SR of the samples were estimated using the picante R package (version 1.8.2) (Kembel et al., 2010). A linear regression model was used to examine the impact of stockpile depth on alpha-diversity (i.e., PD and SR). A post hoc multiple comparison was also conducted using the Benjamini-Hochberg correction to test the significance between stockpile depths. Moreover, the relative abundance of the most abundant phylum was visualized with a stacked bar plot. Phyla with relative proportions over 1% were shown, while phyla with less than 1% were categorized under the “other” group. Further, an NMDS analysis was used to visualize the differences in beta-diversity. The unweighted unifrac dissimilarity was estimated using the GUniFrac (version 1.8) package (Chen et al., 2021) and used for NMDS analysis. A PERMANOVA analysis was also conducted to test the difference in the microbial beta-diversity across stockpile depths. Additionally, a constrained ordination (using redundancy analysis) was conducted to examine the impact of soil geochemistry on the beta-diversity. The count data (Hellinger transformation) and the geochemical data (normalization) were transformed using the decostand function in vegan (version 2.7.1) (Oksanen et al., 2025). Further, a permutation test (permutations = 999) was used to identify the significant edaphic parameters from the geochemical data. These significant geochemical parameters were overlaid on the NMDS output to show their impact on the beta-diversity. A pairwise comparison between the groups was also conducted using the pairwiseAdonis package. Furthermore, a random forest classification model was used using the ranger package (version 0.17.0) to identify the top 10 ASVs explaining the variation across the stockpile depths. The stockpile depth was used as a response, and the ASV feature table was used as the explanatory variable. After running the random forest model, the top 10 ASVs were identified using a permutation test to determine significant (p < 0.05) ASVs. The significant ASVs were then extracted from the count table, and their relative abundances across the stockpile depths depth were investigated.
Results
Community composition
Results showed that the topsoil stockpiles were dominated by the following bacterial phyla: Acidobacteriota, Actinobacteriota, Bacteroidota, Chloroflexota, Firmicutes, Gemmatimonadota, and Proteobacteria in New Afton (Figure 1A) and Acidobacteriota, Actinobacteriota, Bacteroidota, Chloroflexota, Desulfobacterota, Dormibacterota, Fibrobacterota, Firmicutes, Gemmatimonadota, Halobacteriota, Proteobacteria, and Spirochaetota in QR mill (Figure 1C). While the dominant bacterial and archaeal phyla were generally consistent between different stockpile depths (Figure 1A) at New Afton, the most significant difference was observed in Chloroflexota, which was significantly higher across all depths compared to the reference site (SI-6). Similarly, Firmicutes, despite its low relative abundance, was found to be significantly lower in reference compared to other stockpile depths (SI-6). In contrast, considerable differences were observed across all stockpile depths at the QR mill site (Figure 1C). The relative abundance of Acidobacteriota was lower across all depths compared to the reference (SI-7). Similarly, the relative proportion Actinobacteriota decreased gradually with stockpile depth (SI-7). The data also revealed that Desulfobacterota, Firmicutes, and Halobacteriota increased with stockpile depth (SI-7). In contrast, Proteobacteria showed a decline in relative proportions across stockpile depth (SI-7). Verrucomicrobiota, despite its lower proportion, displayed a further reduction in relative abundance with stockpile depth. The most considerable difference in Proteobacteria and Verrucomicrobiota was observed between the reference and stockpile depths below 60 cm (SI-7).
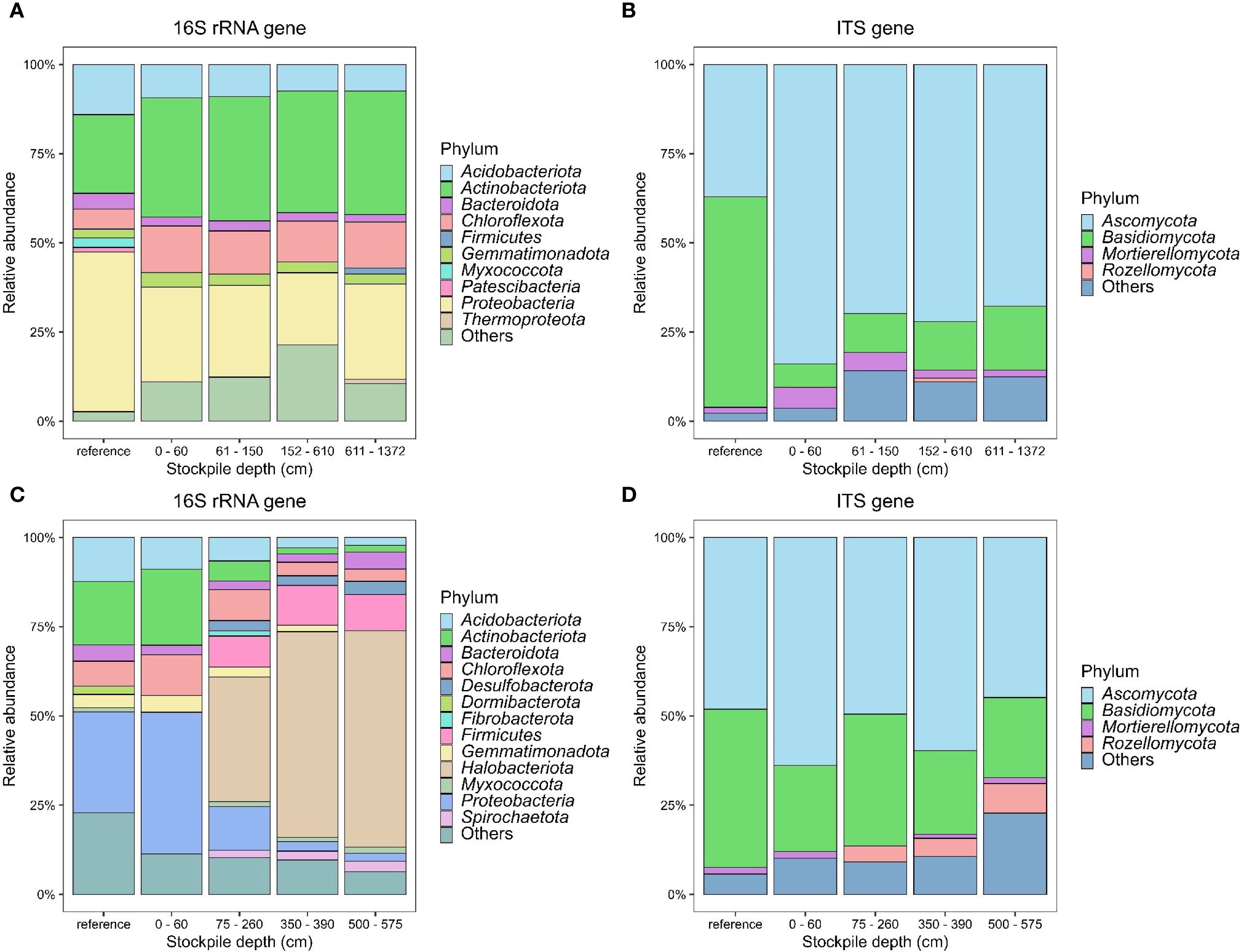
Figure 1. Stacked bar plots showing the relative abundance of dominant 16S and ITS phyla at New Afton (A, B) and QR mill (C, D). Unclassified phyla and phyla of abundance lower than 1% were lumped with “others”.
The dominant soil fungi were mainly composed of Ascomycota, Basidiomycota, Mortierellomycota, and Rozellomycota at New Afton and QR mill (Figures 1B, D). While Ascomycota abundance increased with stockpile depths, the relative proportions of Basidiomycota decreased at New Afton compared to the reference sites. Interestingly, the proportions of Glomeromycota and Kickxellomycota at New Afton reduced considerably compared to the reference site (SI-8). In contrast, Ascomycota abundance was consistent across all samples at the QR mill. Basidiomycota abundance reduced slightly at 0–60 cm depth compared to the reference sites.
The random forest model identified the top 10 ASVs from 16S and ITS sequences from New Afton and QR mill that explained the difference in microbial community across the stockpile depths (SI:1–4). At New Afton, ASV_90 was the most important 16S ASV (SI-5). It belonged to the Gemmatimonadota phylum. The relative proportion of ASV_90 was significantly lower in the reference sites compared to the stockpiles (SI-1). At QR mill, ASV_603 was the most important 16S ASV (SI-5) and belonged to the Acidobacteriota phylum under Vicinamibacteria class. Its relative abundance increased in the stockpiles but was not significantly different than the references (SI-2). Among the ITS ASVs, ASV_43 and ASV_126 were the most important sequences for New Afton and QR mill (SI-3 and -4), respectively. ASV_43 increased with stockpile depth compared to the reference sites and was significantly higher at 152–610 cm and 611–1372 cm depth. In contrast, ASV_126 showed a consistent relative proportion across all sites with a minor increase at 0–60 cm depth.
Alpha diversity
Our results showed no evidence of significant changes in alpha diversity measures for bacterial or fungal communities (observed species richness – Figures 2A, C; Faith’s phylogenetic diversity – Figures 2B, D) with stockpile depth in New Afton. Interestingly, the reference sites exhibited lower alpha diversity of 16S rRNA genes compared to the stockpiles. For the ITS genes, the reference sites displayed similar alpha diversity compared to the stockpiles (Figure 2). At QR mill, the alpha diversity of the 16S rRNA gene showed no significant relationship, indicating that it remained relatively stable across the stockpile depth. The reference site at QR for the 16S rRNA gene displayed slightly elevated levels of alpha diversity (Figures 3A, C). In contrast, the alpha diversity (both species richness and Faith’s phylogenetic diversity) shows a significant relationship with stockpile depth. Both species richness and Faith’s phylogenetic diversity were highest at 0–60 cm depths and gradually decreased with stockpile depths (Figures 3B, D).
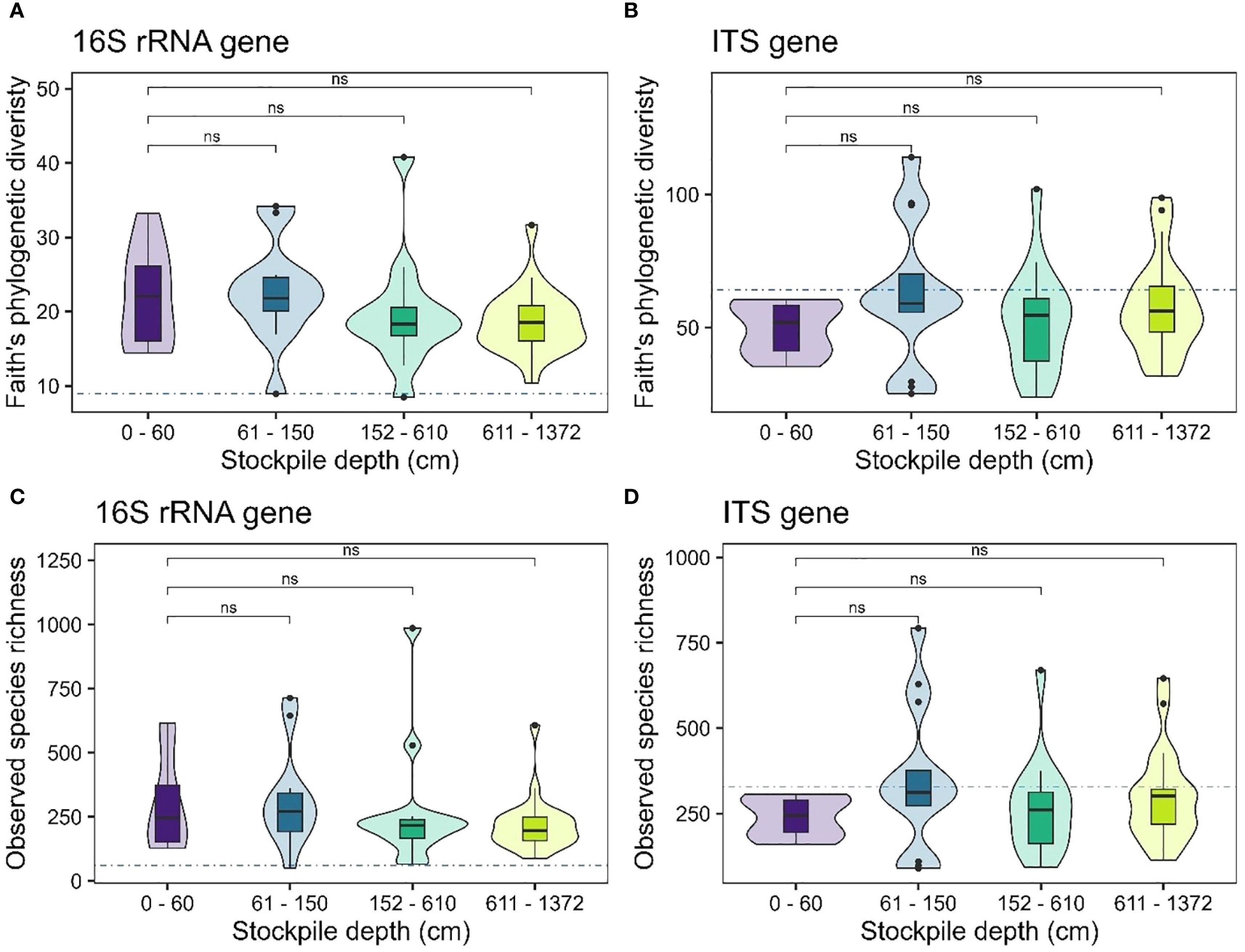
Figure 2. Alpha diversity of 16S and ITS gene across stockpile depths at New Afton. (A, B) show the Faith’s PD of New Afton for the 16S rRNA and the ITS gene, respectively. (C, D) show the species richness of 16S rRNA and the ITS gene, respectively. The horizontal blue dashed line indicates the mean diversity of the reference sites. The boxes in the boxplot show the interquartile range of the data, with the whiskers indicating the spread of the data, and the dots representing outliers. The violin plot shows the distribution of the data. “ns” denotes no significant differences between the comparisons.
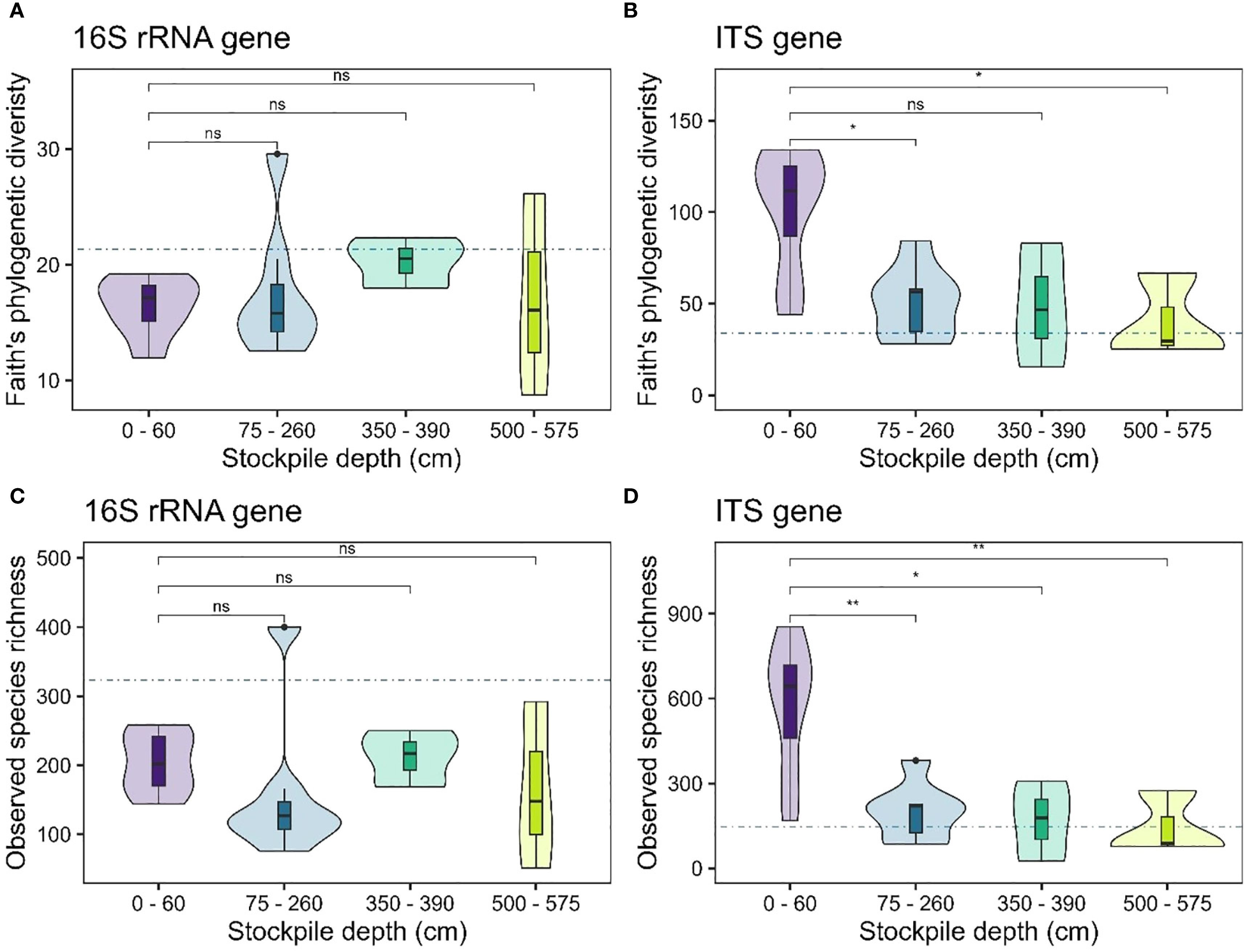
Figure 3. Alpha diversity of 16S and ITS gene across stockpile depths at QR mill. (A, B) show the Faith’s PD at QR mill for the 16S rRNA and the ITS gene, respectively. (C, D) show the species richness of 16S rRNA and the ITS gene, respectively. The horizontal blue dashed line indicates the mean diversity of the reference sites. The boxes in the boxplot show the interquartile range of the data, with the whiskers indicating the spread of the data, and the dots representing outliers. The violin plot shows the distribution of the data. “ns” denotes no significant differences between the comparisons. “*”, “**” indicate significance at α level 0.05 and 0.01, respectively.
Beta diversity and geochemical properties
The PERMANOVA analysis of the 16S and the ITS gene sequences at New Afton and QR mill reveals a significant relationship between beta-diversity and stockpile depth. At New Afton, the stockpile depth explained 14% (Figure 4A) and 16% (Figure 4B) of the variation in 16S and ITS beta-diversity, respectively (Figure 4). Additionally, a pairwise comparison between all groups was conducted to examine the differences between all groups (i.e., stockpile depth and references) for both the 16S and the ITS gene. The analysis revealed that the beta-diversity of reference sites was significantly different from samples of all stockpile depths, suggesting that the variation in beta-diversity could primarily be due to the differences between reference sites and stockpiles. Moreover, the reference sites occupied a distinct ordination space with no overlap with any of the stockpile samples. Despite the considerable differences between reference and stockpiles, some significant differences in beta-diversity between stockpile depths were observed. Most significant differences were observed between the topmost layer (e.g., 0–60) and bottommost layers (e.g., 152–610, 611–1372), suggesting significant changes in beta-diversity of 16S and ITS gene at New Afton as the stockpile depth increases. A redundancy analysis was also conducted to understand how geochemical parameters impacted the beta-diversity at New Afton. The results indicate that available P, soil organic matter (OM), and Mg were associated with 16S beta-diversity. The ITS beta-diversity was found to be associated with B, Ca, K, S, available P, NO3, C:N ratio, Al, Mg, Mn, and Na. Interestingly, available P influenced both 16S and ITS beta-diversity. Additionally, the available P was found to be associated with the reference sites, suggesting the changes in microbial community structure could be driven by phosphorus availability.
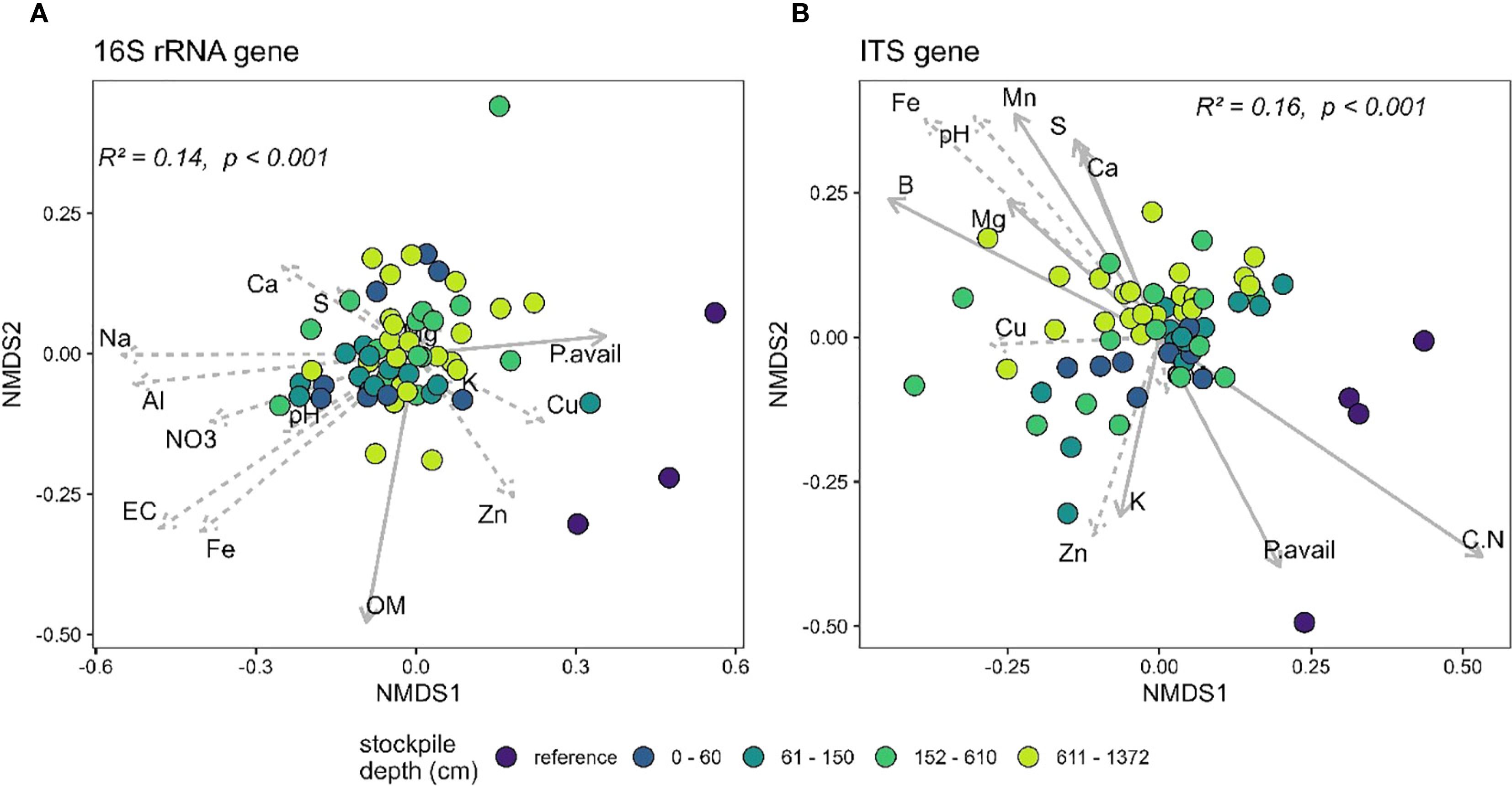
Figure 4. NMDS plot showing the beta-diversity of (A) 16S and (B) ITS gene at New Afton of reference and different stockpile depths. The NMDS was run on unweighted unifrac dissimilarity. The arrows denote the linear relationship between geochemical parameters and the samples. The arrows with a solid line indicate a significant relationship between the samples (reference site and stockpile depths) and the geochemical parameter, while the dashed arrows indicate a non-significant relationship.
At QR mill, stockpile depth explained 40% (Figure 5A) and 34% (Figure 5B) of the variation in the 16S rRNA gene and the ITS community. Similar to the results from New Afton, most of the variation could be driven by the difference in community composition between the reference and the stockpile depths. The pairwise PERMANOVA results demonstrate that stockpile samples from all depths are significantly different from the reference sites. However, there were community differences across stockpile depths for 16S and ITS at QR mills. Most of the differences, like New Afton, were observed between the topmost layer (0–60) and the bottommost layers (350–390, 500–575). The redundancy analysis revealed that B, Fe, Mg, and Mn impacted the 16S rRNA beta-diversity. The ITS community was found to be associated with Al, B, Fe, K, Mn, Na, P, Cu, and NH4. Interestingly, B was found to be associated with the 16S and ITS communities at both sites (New Afton and QR mill).
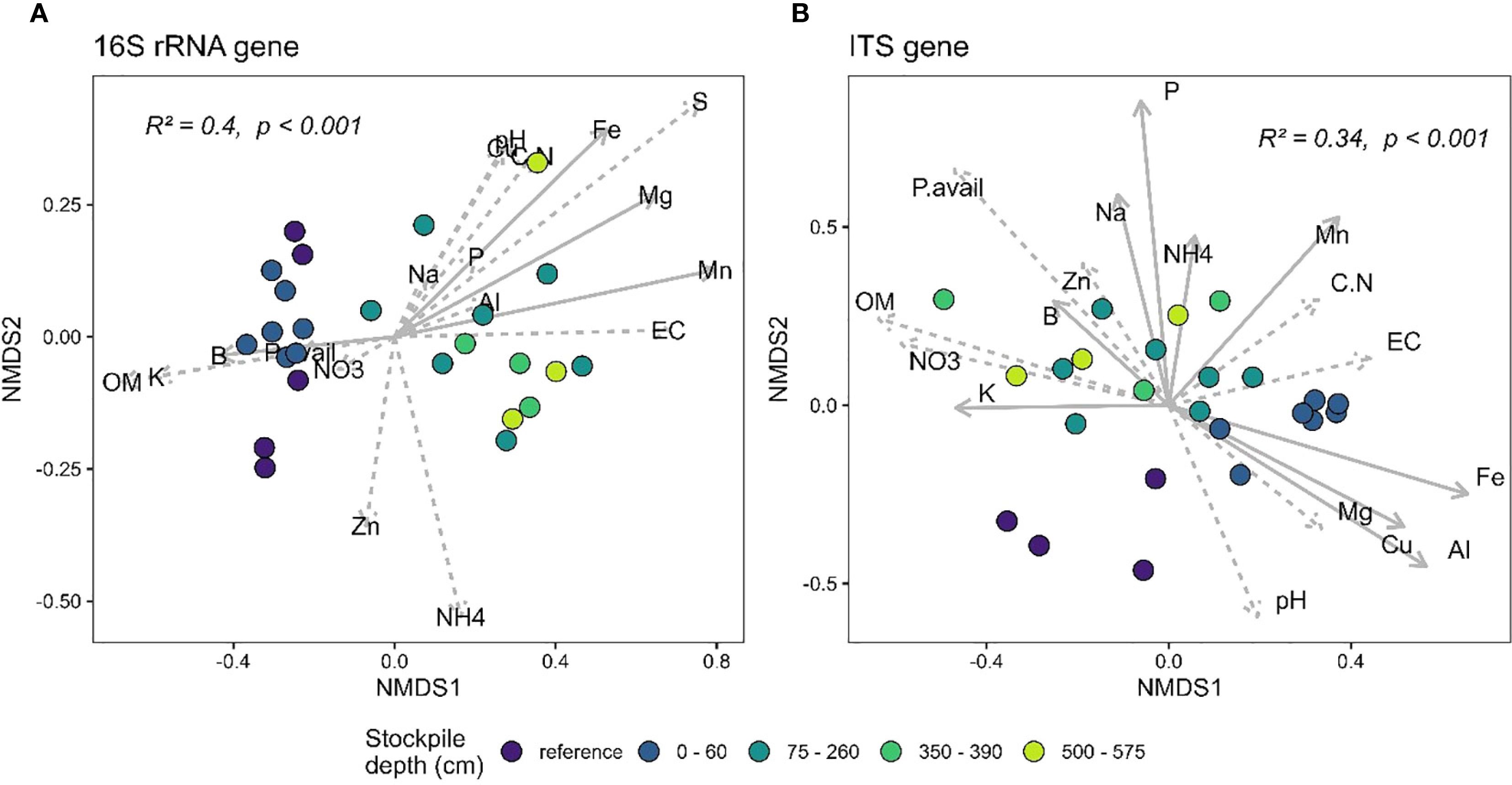
Figure 5. NMDS plot showing the beta-diversity of (A) 16S and (B) ITS gene at QR mill of reference and different stockpile depths. The NMDS was run on unweighted unifrac dissimilarity. The arrows denote the linear relationship between geochemical parameters and the samples. The arrows with a solid line indicate a significant relationship between the samples (reference site and stockpile depths) and the geochemical parameter, while the dashed arrows indicate a non-significant relationship.
Discussion
On a broad scale, our results are consistent with global microbial surveys (Delgado-Baquerizo et al., 2018; Tedersoo et al., 2022). Similar to Delgado-Baquerizo et al. (2018), our samples were dominated by Proteobacteria, Acidobacteriota, and Actinobacteriota among the 16S community. Among the ITS community, the samples had a high abundance of Ascomycota and Basidiomycota, which is similar to the observations made by Tedersoo et al. (2022). However, interesting trends are unraveled upon deeper inspection. At New Afton, the Chloroflexota emerged at a higher proportion, particularly in the soil stockpiles. In the reference sites near New Afton, the relative abundance of Chloroflexota remained below 10%, while it stayed above 10% for the rest of the stockpiles (SI-6). In some instances, Chloroflexota’s relative abundance was found to be over 30% among the stockpile samples (SI-6). The Chloroflexota phylum contains microorganisms that exhibit several metabolic pathways to harvest energy, including anoxygenic phototrophy (Pierson and Castenholz, 1974), chemoheterotrophy, and heterotrophy (Freches and Fradinho, 2024). These diverse metabolic pathways for survival and growth potentially allow Chloroflexota to thrive in disturbed conditions such as soil stockpiling. Similarly, Firmicutes’ relative abundance increased with soil depth at New Afton soil stockpiles. A similar observation was made by Bai et al. (2017); Zhang et al. (2017), who found that the relative proportion of Firmicutes increased with soil depth (Bai et al., 2017; Zhang et al., 2017). Firmicutes employ survival strategies such as endospore formation that enable them to grow under stressful conditions (Fajardo et al., 2019; Filippidou et al., 2016), which is likely the case in the New Afton stockpiles. Among the fungal phyla at New Afton, the relative abundance of Glomeromycota reduced with stockpile depth (SI-8). Glomeromycota contains arbuscular mycorrhizal fungi that are ubiquitous in soil ecosystems (Brundrett, 1991) and are essential for soil function as they support plant life and benefit biogeochemical cycles (Hawkins et al., 2023; Högberg and Högberg, 2002; Van Der Heijden et al., 2015). A decline in their abundance could severely impact the soil quality of the stockpiles at New Afton.
At QR mill, substantial changes in the 16S rRNA community composition were observed (Figure 1C). While the relative abundance of Acidobacteriota, Actinobacteriota, Verrucomicrobiota, and Proteobacteria declined, the proportion of Desulfobacterota and Halobacteriota increased with stockpile depth (SI-7). Several studies have reported similar results, where they found that Acidobacteriota, Actinobacteriota, Verrucomicrobiota, and Proteobacteria decreased with soil depths (Banerjee et al., 2021; Nayak and Mishra, 2020; Zhang et al., 2019; Zhao et al., 2021) likely due to slow growth (Goldman and Green, 2008) or stressful conditions like stockpiles. In contrast, Desulfobacterota and Halobacteriota proportions increased, likely due to their metabolic preferences (Kuever et al., 2015) and/or cellular ecophysiology (Dundas, 1977). While Desulfobacterota respire anaerobically (Thauer et al., 2007), Halobacteriota ecophysiology (Dundas, 1977) allow them to sustain disturbances such as soil stockpiling.
Analysis of alpha-diversity of 16S rRNA and ITS gene revealed no significant changes across stockpile depth at New Afton (Figure 2). However, the alpha-diversity of the 16S rRNA gene (both SR and PD) of the reference site was substantially lower than the stockpiles (Figures 2A, C). A plausible explanation is that soil is regularly added to the stockpiles, which results in frequent disturbances to the ecosystem, while the reference site did not receive such a disturbance. Disturbance is known to sometimes increase the diversity of the ecosystem, which is likely why the 16S rRNA alpha-diversity in stockpiles is higher than the reference sites at New Afton. Like New Afton, 16S rRNA alpha-diversity at QR mill did not show any significant relationship with stockpile depth, but the ITS gene alpha-diversity decreased substantially with depth (Figures 3B, D). Other studies have made similar observations, where they found a decline in alpha-diversity with depth (Cabrera-Hernandez et al., 2025; Hernandez et al., 2024). Decrease in microbial diversity with depth has also been observed due to depleted C levels (Eilers et al., 2012; Hansel et al., 2008). In support of this theory, the observed trend of microbial diversity depletion with stockpile depth in QR mill corresponded to the decrease in OM and increase in C:N ratio, especially when the soil was deeper than 60 cm.
The most significant change in diversity at New Afton and QR mill was observed in beta-diversity (Figures 4, 5). Expectedly, the reference sites were significantly different than stockpiles as disturbance is known to impact the beta-diversity (Myers et al., 2015; Vanschoenwinkel et al., 2013). Moreover, stockpiling can substantially change the soil’s abiotic properties (Gupta et al., 2019), which can significantly impact the beta-diversity and ecosystem functioning (Barbosa et al., 2023; Singh et al., 2019a; Xu et al., 2020). Interestingly, soil phosphorus (either available or total), along with Boron, impacted the 16S and ITS beta-diversity for both sites, suggesting their role in determining the community assembly. Phosphorus is a macronutrient and is essential for the survival of all organisms (Butusov and Jernelöv, 2013), and changes in its availability can have a substantial influence on microbial community composition (DeForest and Scott, 2010). Additionally, boron is known to impact not only on microbial community but also their biomass and activity (Vera et al., 2019; Xu et al., 2025). The results also indicate changes in beta-diversity across soil depths, particularly between the uppermost and lowermost depths (ST1 and 2), which is likely due to changes in the geochemistry of the stockpiles (Fischer et al., 2022).
Additionally, a substantial difference between the stockpile at New Afton and the QR mill is observed for both alpha and beta-diversity. The age of the stockpiles could explain this difference between New Afton and QR mills. The stockpile at New Afton is only six years old, while the age of the QR mill stockpile is 20 years. As such, the effects of stockpiling have accumulated for 20 years at the QR mill. Therefore, a significant change in alpha and beta-diversity was observed at the QR mill. Gorzelak et al. (2020) also report a similar observation, where the age of the stockpile changes microbial community composition considerably. It is likely that if the soils at the New Afton site were allowed to stockpile for 20 years, the stockpiles would exhibit similar results.
Conclusions
Soil stockpiles are a critical resource for the mining industry, as they are later used to reclaim sites at the end of mining operations. Studying soil stockpiling effects through a microbial perspective could provide valuable information about the quality of the soils because microbial communities (key ecosystem engineers) are sensitive to disturbances. Understanding how these disturbances (soil stockpiling) impact microbial communities could allow environmental managers to develop more effective reclamation strategies. To our knowledge, this is the first study that investigated the impact of soil stockpiling at a greater depth (i.e., > 300 cm), which demonstrates that the beta-diversity and in some instances alpha-diversity changes substantially with stockpile depth. The study also shows that stockpiling could potentially impact the quality of soil by reducing the abundance of arbuscular mycorrhizal fungi. Furthermore, the difference in results between the sites can be explained by the age of stockpiling, indicating that the duration of stockpiling has a significant effect on microbial community and must be taken into account for future studies. While this study provides an understanding of microbial community dynamics due to soil stockpiling, future research using transcriptomics and metabolomics can elucidate the functional roles of the microbes in the stockpiles.
Data availability statement
The original contributions presented in the study are publicly available. The processed data, code, and the figures are available on GitHub at https://github.com/jaymicro/Stockpile_microbiome_manuscript.
Author contributions
AF: Data curation, Validation, Methodology, Conceptualization, Investigation, Formal Analysis, Writing – original draft, Writing – review & editing. JS: Data curation, Formal Analysis, Writing – review & editing. JV: Writing – review & editing, Methodology, Data curation, Formal Analysis. EB: Writing – review & editing, Methodology, Formal Analysis, Data curation. LF: Writing – original draft, Investigation, Funding acquisition, Resources, Writing – review & editing, Methodology, Supervision, Conceptualization.
Funding
The author(s) declare financial support was received for the research and/or publication of this article. Natural Sciences and Engineering Research Council of Canada Industrial Research Chair in Ecosystem Reclamation.
Conflict of interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
The author(s) declared that they were an editorial board member of Frontiers, at the time of submission. This had no impact on the peer review process and the final decision.
Generative AI statement
The author(s) declare that no Generative AI was used in the creation of this manuscript.
Any alternative text (alt text) provided alongside figures in this article has been generated by Frontiers with the support of artificial intelligence and reasonable efforts have been made to ensure accuracy, including review by the authors wherever possible. If you identify any issues, please contact us.
Publisher’s note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
Supplementary material
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/frmbi.2025.1607677/full#supplementary-material
References
Annas R. and Coupé R. (1979). Biogeoclimatic zones and subzones of the Cariboo Forest Region (Williams Lake, BC, Canada: Ministry of Forests).
Bai R., Wang J.-T., Deng Y., He J.-Z., Feng K., and Zhang L.-M. (2017). Microbial Community and Functional Structure Significantly Varied among Distinct Types of Paddy Soils But Responded Differently along Gradients of Soil Depth Layers. Front. Microbiol. 8. doi: 10.3389/fmicb.2017.00945
Banerjee S., Zhao C., Kirkby C. A., Coggins S., Zhao S., Bissett A., et al. (2021). Microbial interkingdom associations across soil depths reveal network connectivity and keystone taxa linked to soil fine-fraction carbon content. Agricult. Ecosyst. Environ. 320, 107559. doi: 10.1016/j.agee.2021.107559
Barbosa C., Tamayo-Leiva J., Alcorta J., Salgado O., Daniele L., Morata D., et al. (2023). Effects of hydrogeochemistry on the microbial ecology of terrestrial hot springs. Microbiol. Spectr. 11, e00249–e00223. doi: 10.1128/spectrum.00249-23
Bastida F., Zsolnay A., Hernández T., and García C. (2008). Past, present and future of soil quality indices: A biological perspective. Geoderma 147, 159–171. doi: 10.1016/j.geoderma.2008.08.007
Bolyen E., Rideout J. R., Dillon M. R., Bokulich N. A., Abnet C. C., Al-Ghalith G. A., et al. (2019). Reproducible, interactive, scalable and extensible microbiome data science using QIIME 2. Nat. Biotechnol. 37, 852–857. doi: 10.1038/s41587-019-0209-9
Boyer S., Wratten S., Pizey M., and Weber P. (2011). Impact of soil stockpiling and mining rehabilitation on earthworm communities. Pedobiologia 54, S99–S102. doi: 10.1016/j.pedobi.2011.09.006
Bradshaw A. (1997). Restoration of mined lands—using natural processes. Ecol. Eng. 8, 255–269. doi: 10.1016/S0925-8574(97)00022-0
Bray R. H. and Kurtz L. T. (1945). Determination of total, organic, and available forms of phosphorus in soils. Soil Science 59, 39–46. doi: 10.1097/00010694-194501000-00006
Brundrett M. (1991). “Mycorrhizas in natural ecosystems,” in Advances in Ecological Research (London, UK: London, UK Elsevier), 171–313. doi: 10.1016/S0065-2504(08)60099-9
Butusov M. and Jernelöv A. (2013). “The role of phosphorus in the origin of life and in evolution,” in Phosphorus: An Element That Could Have Been Called Lucifer (New York, USA: Springer), 1–12.
Cabrera-Hernandez J. A., Davidson H., MacKenzie M. D., and Lanoil B. D. (2025). Impact of stockpiling on soil fungal communities and their functions. Restor. Ecol. 33, e14333. doi: 10.1111/rec.14333
Callahan B. J., McMurdie P. J., Rosen M. J., Han A. W., Johnson A. J. A., and Holmes S. P. (2016). DADA2: High-resolution sample inference from Illumina amplicon data. Nat. Methods 13, 581–583. doi: 10.1038/nmeth.3869
Chen B. D., Zhu Y.-G., Xiao X. Y., and Smith S. E.. (2007). Effects of the arbuscular mycorrhizal fungus Glomus mosseae on growth and metal uptake by four plant species in copper mine tailings. Environ. Pollut. 147, 374–380. doi: 10.1016/j.envpol.2006.04.027
Chen J., Zhang X., and Yang L. (2021). GUniFrac: Generalized UniFrac Distances, Distance-Based Multivariate Methods and Feature-Based Univariate Methods for Microbiome Data Analysis_. R package version 1.8. doi: 10.32614/CRAN.package.GUniFrac
Darmody R. G., Daniels W. L., Marlin J. C., and Cremeens D. L. (2009). TOPSOIL: WHAT IS IT AND WHO CARES? JASMR 2009, 237–269. doi: 10.21000/JASMR09010237
DeForest J. L. and Scott L. G. (2010). Available organic soil phosphorus has an important influence on microbial community composition. Soil Sci. Soc. America J. 74, 2059–2066. doi: 10.2136/sssaj2009.0426
Delgado-Baquerizo M., Oliverio A. M., Brewer T. E., Benavent-González A., Eldridge D. J., Bardgett R. D., et al. (2018). A global atlas of the dominant bacteria found in soil. Science 359, 320–325. doi: 10.1126/science.aap9516
Dundas I. E. D. (1977). “Physiology of halobacteriaceae,” in Advances in Microbial Physiology (London, UK: Elsevier), 85–120. doi: 10.1016/S0065-2911(08)60315-X
Eilers K. G., Debenport S., Anderson S., and Fierer N. (2012). Digging deeper to find unique microbial communities: The strong effect of depth on the structure of bacterial and archaeal communities in soil. Soil Biol. Biochem. 50, 58–65. doi: 10.1016/j.soilbio.2012.03.011
Ezeokoli O. T., Mashigo S. K., Paterson D. G., Bezuidenhout C. C., and Adeleke R. A. (2019). Microbial community structure and relationship with physicochemical properties of soil stockpiles in selected South African opencast coal mines. Soil Sci. Plant Nutr. 65, 332–341. doi: 10.1080/00380768.2019.1621667
Fajardo C., Costa G., Nande M., Botías P., García-Cantalejo J., and Martín M. (2019). Pb, Cd, and Zn soil contamination: Monitoring functional and structural impacts on the microbiome. Appl. Soil Ecol. 135, 56–64. doi: 10.1016/j.apsoil.2018.10.022
Felton G. K. and Taraba J. L. (1994). A laboratory examination of organic matter degradation in a B horizon soil from post-mining reconstructed prime farmland soil. Int. J. Surface. Mining. Reclamation. Environ. 8, 153–157. doi: 10.1080/09208119408964778
Filippidou S., Wunderlin T., Junier T., Jeanneret N., Dorador C., Molina V., et al. (2016). A combination of extreme environmental conditions favor the prevalence of endospore-forming firmicutes. Front. Microbiol. 7. doi: 10.3389/fmicb.2016.01707
Fischer A. M., Van Hamme J. D., Gardner W. C., and Fraser L. H. (2022). Impacts from topsoil stockpile height on soil geochemical properties in two mining operations in british columbia: implications for restoration practices. Mining 2, 315–329. doi: 10.3390/mining2020017
Freches A. and Fradinho J. C. (2024). The biotechnological potential of the Chloroflexota phylum. Appl. Environ. Microbiol. 90, e01756–e01723. doi: 10.1128/aem.01756-23
Ghose M. K. and Kundu N. K. (2004). Deterioration of soil quality due to stockpiling in coal mining areas. Int. J. Environ. Stud. 61, 327–335. doi: 10.1080/0020723032000093991
Goldman E. and Green L. H. (Eds.) (2008). Practical Handbook of Microbiology (Boca Raton, FL, USA: CRC Press). doi: 10.1201/9781420009330
Gorzelak M., McAmmond B. M., Van Hamme J. D., Birnbaum C., Thomsen C., and Hart M. (2020). Soil microbial communities in long-term soil storage for sand mine reclamation. Ecol. Restor. 38, 13–23. doi: 10.3368/er.38.1.13
Government of British Columbia. (2018). BC Soil Information Finder Tool - Province of British Columbia[WWW Document]. Available online at: https://www2.gov.bc.ca/gov/content/environment/air-land-water/land/soil/soil-information-finder (Accessed September 3, 2025).
Gruber K. (2015). Deep influence of soil microbes. Nat. Plants 1, 2015.194. doi: 10.1038/nplants.2015.194
Gupta S. D., Kirby W., and Pinno B. D. (2019). Effects of stockpiling and organic matter addition on nutrient bioavailability in reclamation soils. Soil Sci. Soc. America J. 83, S27–S41. doi: 10.2136/sssaj2018.07.0273
Hansel C. M., Fendorf S., Jardine P. M., and Francis C. A. (2008). Changes in bacterial and archaeal community structure and functional diversity along a geochemically variable soil profile. Appl. Environ. Microbiol. 74, 1620–1633. doi: 10.1128/AEM.01787-07
Harris J. A., Birch P., and Short K. C. (1989). Changes in the microbial community and physico-chemical characteristics of topsoils stockpiled during opencast mining. Soil Use Manage. 5, 161–168. doi: 10.1111/j.1475-2743.1989.tb00778.x
Hawkins H.-J., Cargill R. I. M., Van Nuland M. E., Hagen S. C., Field K. J., Sheldrake M., et al. (2023). Mycorrhizal mycelium as a global carbon pool. Curr. Biol. 33, R560–R573. doi: 10.1016/j.cub.2023.02.027
Hernandez J. A. C., Ribeiro H. M., Bayne E., MacKenzie M. D., and Lanoil B. D. (2024). Impact of stockpile depth and storage time on soil microbial communities. Appl. Soil Ecol. 196, 105275. doi: 10.1016/j.apsoil.2024.105275
Högberg M. N. and Högberg P. (2002). Extramatrical ectomycorrhizal mycelium contributes one-third of microbial biomass and produces, together with associated roots, half the dissolved organic carbon in a forest soil. New Phytol. 154, 791–795. doi: 10.1046/j.1469-8137.2002.00417.x
Kachurina O. M., Zhang H., Raun W. R., and Krenzer E. G. (2000). Simultaneous determination of soil aluminum, ammonium- and nitrate-nitrogen using 1 M potassium chloride extraction. Commun. Soil Sci. Plant Anal. 31, 893–903. doi: 10.1080/00103620009370485
Katoh K. and Standley D. M. (2013). MAFFT multiple sequence alignment software version 7: improvements in performance and usability. Mol. Biol. Evol. 30, 772–780. doi: 10.1093/molbev/mst010
Kembel S. W., Cowan P. D., Helmus M. R., Cornwell W. K., Morlon H., Ackerly D. D., et al. (2010). Picante: R tools for integrating phylogenies and ecology. Bioinformatics 26, 1463–1464. doi: 10.1093/bioinformatics/btq166
Kuever J., Rainey F. A., and Widdel F. (2015). “Desulfobacter,” in Bergey’s manual of Systematics of Archaea and Bacteria. Ed. Whitman W. B. (New York, USA: Wiley), 1–7. doi: 10.1002/9781118960608.gbm01011
Lloyd D., Angove K., Hope G., and Thompson C. (eds.). (1991). Field Guide to Site Identification and Interpretation for the Kamloops Forest Region. (Victoria, B.C: British Columbia Ministry of Forests) Available online at: https://www.for.gov.bc.ca/hfd/pubs/Docs/Lmh/Lmh23.pdf (Accessed September 4, 2025).
McDonald D., Jiang Y., Balaban M., Cantrell K., Zhu Q., Gonzalez A., et al. (2024). Greengenes2 unifies microbial data in a single reference tree. Nat. Biotechnol. 42, 715–718. doi: 10.1038/s41587-023-01845-1
Meidinger D. and Pojar J. eds. (1991). Ecosystems of British Columbia. (Victoria, B.C.: Special Report Series 6, British Columbia Ministry of Forests). Available online at: https://sherwoods-forests.com/Downloads/BGC-zones-of-British-Columbia.pdf (Accessed September 4, 2025).
Ministry of Energy, Mines and Low Carbon Innovation (2020). Chief Inspector’s annual reports - Province of British Columbia [WWW Document]. Available online at: https://www2.gov.bc.ca/gov/content/industry/mineral-exploration-mining/health-safety/chief-inspector-s-annual-reports (Accessed August 5, 2025).
Mummey D. L., Stahl P. D., and Buyer J. S. (2002). Soil microbiological properties 20 years after surface mine reclamation: spatial analysis of reclaimed and undisturbed sites. Soil Biol. Biochem. 34, 1717–1725. doi: 10.1016/S0038-0717(02)00158-X
Myers J. A., Chase J. M., Crandall R. M., and Jiménez I. (2015). Disturbance alters beta-diversity but not the relative importance of community assembly mechanisms. J. Ecol. 103, 1291–1299. doi: 10.1111/1365-2745.12436
Nayak S. K. and Mishra B. B. (2020). Frontiers in Soil and Environmental Microbiology. 1st ed (Boca Raton, FL, USA: CRC Press). doi: 10.1201/9780429485794
Nilsson R. H., Anslan S., Bahram M., Wurzbacher C., Baldrian P., and Tedersoo L. (2019). Mycobiome diversity: high-throughput sequencing and identification of fungi. Nat. Rev. Microbiol. 17, 95–109. doi: 10.1038/s41579-018-0116-y
Oksanen J., Simpson G., Blanchet F. G., Kindt R., Legendre P., Minchin R., et al. (2025). vegan: Community Ecology Package. R package version 2.7-1. doi: 10.32614/CRAN.package.vegan
Pierson B. K. and Castenholz R. W. (1974). A phototrophic gliding filamentous bacterium of hot springs, Chloroflexus aurantiacus, gen. and sp. nov. Arch. Microbiol. 100, 5–24. doi: 10.1007/BF00446302
Price M. N., Dehal P. S., and Arkin A. P. (2010). FastTree 2 – Approximately maximum-likelihood trees for large alignments. PloS One 5, e9490. doi: 10.1371/journal.pone.0009490
Ramirez K. S., Knight C. G., De Hollander M., Brearley F. Q., Constantinides B., Cotton A., et al. (2017). Detecting macroecological patterns in bacterial communities across independent studies of global soils. Nat. Microbiol. 3, 189–196. doi: 10.1038/s41564-017-0062-x
Rodrigues J. F. M., Tackmann J., Malfertheiner L., Patsch D., Perez Molphe Montoya E., Näpflin N., et al. (2025). The MicrobeAtlas database: Global trends and insights into Earth’s microbial ecosystems. bioRxiv, 2025–07. doi: 10.1101/2025.07.18.665519
Saccá M. L., Barra Caracciolo A., Di Lenola M., and Grenni P. (2017). “Ecosystem services provided by soil microorganisms,” in Soil Biological Communities and Ecosystem Resilience (Cham, Switzerland: Springer), 9–24.
Sandroni V. (2003). Microwave digestion of sediment, soils and urban particulate matter for trace metal analysis. Talanta 60, 715–723. doi: 10.1016/S0039-9140(03)00131-0
Singh J. P., Ojinnaka E. U., Krumins J. A., and Goodey N. M. (2019a). Abiotic factors determine functional outcomes of microbial inoculation of soils from a metal contaminated brownfield. Ecotoxicol. Environ. Saf. 168, 450–456. doi: 10.1016/j.ecoenv.2018.10.114
Singh J. P., Vaidya B. P., Goodey N. M., and Krumins J. A. (2019b). Soil microbial response to metal contamination in a vegetated and urban brownfield. J. Environ. Manage. 244, 313–319. doi: 10.1016/j.jenvman.2019.04.111
Tedersoo L., Mikryukov V., Zizka A., Bahram M., Hagh-Doust N., Anslan S., et al. (2022). Global patterns in endemicity and vulnerability of soil fungi. Global Change Biol. 28, 6696–6710. doi: 10.1111/gcb.16398
Thauer R. K., Stackebrandt E., and Hamilton W. A. (2007). “Energy metabolism and phylogenetic diversity of sulphate-reducing bacteria,” in Sulphate-reducing bacteria: Environmental and engineered systems, eds. Barton L. L. and Hamilton W. A.. (Cambridge, UK / New York, USA: Cambridge University Press) 1–37.
Tripathi N., Singh R. S., and Hills C. D. (2016). Reclamation of Mine-Impacted Land for Ecosystem Recovery. 1st ed (West Sussex, UK: Wiley). doi: 10.1002/9781119057925
Vancov T. and Keen B. (2009). Amplification of soil fungal community DNA using the ITS86F and ITS4 primers. FEMS Microbiol. Lett. 296, 91–96. doi: 10.1111/j.1574-6968.2009.01621.x
Van Der Heijden M. G. A., Martin F. M., Selosse M., and Sanders I. R. (2015). Mycorrhizal ecology and evolution: the past, the present, and the future. New Phytol. 205, 1406–1423. doi: 10.1111/nph.13288
Vanschoenwinkel B., Buschke F., and Brendonck L. (2013). Disturbance regime alters the impact of dispersal on alpha and beta diversity in a natural metacommunity. Ecology 94, 2547–2557. doi: 10.1890/12-1576.1
Vera A., Moreno J. L., García C., Morais D., and Bastida F. (2019). Boron in soil: The impacts on the biomass, composition and activity of the soil microbial community. Sci. Total. Environ. 685, 564–573. doi: 10.1016/j.scitotenv.2019.05.375
Wang D., Lv S., Jiang P., and Li Y. (2017). Roles, regulation, and agricultural application of plant phosphate transporters. Front. Plant Sci. 8, 817. doi: 10.3389/fpls.2017.00817
Xu X., Hao Y., Cai Z., Cao Y., Jia W., Zhao J., et al. (2025). Nanoscale−boron nitride positively alters rhizosphere microbial communities and subsequent cucumber (Cucumis sativa) growth: A metagenomic analysis. Sci. Total. Environ. 958, 178115. doi: 10.1016/j.scitotenv.2024.178115
Xu R., Li B., Xiao E., Young L. Y., Sun X., Kong T., et al. (2020). Uncovering microbial responses to sharp geochemical gradients in a terrace contaminated by acid mine drainage. Environ. pollut. 261, 114226. doi: 10.1016/j.envpol.2020.114226
Zhang C., Li J., Wang J., Liu G., Wang G., Guo L., et al. (2019). Decreased temporary turnover of bacterial communities along soil depth gradient during a 35-year grazing exclusion period in a semiarid grassland. Geoderma 351, 49–58. doi: 10.1016/j.geoderma.2019.05.010
Zhang B., Penton C. R., Xue C., Quensen J. F., Roley S. S., Guo J., et al. (2017). Soil depth and crop determinants of bacterial communities under ten biofuel cropping systems. Soil Biol. Biochem. 112, 140–152. doi: 10.1016/j.soilbio.2017.04.019
Keywords: mine reclamation, soil microbial ecology, ecosystem restoration, soil response to disturbance, high-throughput sequencing
Citation: Fischer A, Singh JP, Van Hamme J, Bottos E and Fraser LH (2025) Investigating impacts from topsoil stockpile height on soil microbial communities. Front. Microbiomes 4:1607677. doi: 10.3389/frmbi.2025.1607677
Received: 07 April 2025; Accepted: 27 August 2025;
Published: 18 September 2025.
Edited by:
Nancy Trun, Duquesne University, United StatesReviewed by:
Richard K. Tennant, University of Exeter, United KingdomQi Zhang, Nanjing University, China
Copyright © 2025 Fischer, Singh, Van Hamme, Bottos and Fraser. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Lauchlan H. Fraser, bGZyYXNlckB0cnUuY2E=