 Cristiana Dimulescu
Cristiana Dimulescu Ronja Strömsdörfer
Ronja Strömsdörfer Agnes Flöel4,5‡
Agnes Flöel4,5‡ Klaus Obermayer
Klaus Obermayer- 1Neural Information Processing Group, Fakultät IV, Technische Universität Berlin, Berlin, Germany
- 2Bernstein Center for Computational Neuroscience, Berlin, Germany
- 3Einstein Center for Neuroscience, Berlin, Germany
- 4Department of Neurology, University Medicine, Greifswald, Germany
- 5German Center for Neurodegenerative Diseases, Greifswald, Germany
The human brain is a complex dynamical system which displays a wide range of macroscopic and mesoscopic patterns of neural activity, whose mechanistic origin remains poorly understood. Whole-brain modelling allows us to explore candidate mechanisms causing the observed patterns. However, it is not fully established how the choice of model type and the networks’ spatial resolution influence the simulation results, hence, it remains unclear, to which extent conclusions drawn from these results are limited by modelling artefacts. Here, we compare the dynamics of a biophysically realistic, linear-nonlinear cascade model of whole-brain activity with a phenomenological Wilson-Cowan model using three structural connectomes based on the Schaefer parcellation scheme with 100, 200, and 500 nodes. Both neural mass models implement the same mechanistic hypotheses, which specifically address the interaction between excitation, inhibition, and a slow adaptation current which affects the excitatory populations. We quantify the emerging dynamical states in detail and investigate how consistent results are across the different model variants. Then we apply both model types to the specific phenomenon of slow oscillations, which are a prevalent brain rhythm during deep sleep. We investigate the consistency of model predictions when exploring specific mechanistic hypotheses about the effects of both short- and long-range connections and of the antero-posterior structural connectivity gradient on key properties of these oscillations. Overall, our results demonstrate that the coarse-grained dynamics is robust to changes in both model type and network resolution. In some cases, however, model predictions do not generalize. Thus, some care must be taken when interpreting model results.
1 Introduction
The human brain is a complex dynamical system. It exhibits a rich variety of spatiotemporally organized activity, where different patterns correspond to different functionalities and mechanisms in human cognitive processes. Rasch and Born (2013) state that slow oscillations (SOs), that travel as plane waves in an anterior-posterior direction (Massimini et al., 2004), play a crucial role in memory consolidation during non-rapid eye movement (non-REM) sleep, and Muller et al. (2016) identified a dominant rotational temporal-parietal-frontal directionality of spindle oscillations that accompany SOs. Beyond spatiotemporal patterns during sleep, Das et al. (2024) showed that spatial modes that regulate plane waves are absent in navigational memory tasks in humans while in verbal memory tasks, they observed different clusters of traveling waves depending on the letters that appear in words. Hence, an indicator of the functionality of a rhythm is its spatiotemporal organization (see further, Breakspear et al. (2003); Mohan et al. (2024)). While reductionist approaches to the temporal dynamics of activity patterns have been widely researched to understand the functionality of the more local dynamics in the brain, most recently, neuroscientific research has shown an increasing interest to include the identification of the spatial dynamics, especially on a larger scale (see Pessoa, 2022; Sporns, 2022).
In-silico methods can support these investigations by computational modeling of specific brain activity for the evaluation of candidate mechanisms, as well as supporting clinical studies using personalized whole-brain models such as the Virtual Brain Twins (see Hashemi et al., 2025; Jirsa et al., 2023; Wang et al., 2024). Fousek et al. (2024) introduced a principled framework which provides a mechanistic description for resting state activity, suggesting a fundamental pathway for the generalisation of large-scale models. Additionally, in silico methods have been applied to surface EEG measurements (Sanchez-Vives et al., 2017; Cakan et al., 2022), and to intracranially recorded activity in humans (Deco et al., 2017; Das et al., 2024; Mohan et al., 2024; Muller et al., 2016), rodents (see Bhattacharya et al., 2022; Liang et al., 2023; Dasilva et al., 2021), and other species (Muller et al., 2014). Intracranial recording methods measure activity of higher spatial and temporal resolutions, hence, in silico methods require an adjustment to spatially denser models. On a smaller scale (i.e., not the whole brain), Capone et al. (2023) showed that different granularity of the recorded space changed the measured density of SO wave velocity in mice, where faster waves were neglected on a lower spatial resolution. On a larger scale, Popovych et al. (2021) found that the fit of simulated activity to empirical functional connectivity depends both on parcellation schemes and spatial resolution, and Proix et al. (2016) shows that the parcellation size affects the dynamics of a whole-brain model whereas it was challenging to identify a consistent type of change.
Key to the emergence of different types of spatiotemporal patterns is the dynamical landscape of a computational model that can be decomposed into different regions of interest by the different types of stability a dynamical system experiences. Sanchez-Vives et al. (2017) showed that bistability is required for the organization of neocortical SOs both in silico, as well as empirically. Cakan et al. (2022) identified a temporal destabilization of a stable high-activity state (up state) by a fatigue mechanism (spike-frequency adaptation) for transitioning into a low-activity state (down state) which is interrupted by noise to ultimately alternate at a low frequency
Different types of instability can also enable the formation of complex local patterns. Townsend and Gong (2018) applied methods from the analysis of turbulent flows to determine velocity vector fields over empirically recorded brain activity of mice. In those velocity vector fields, outward (sources) or inward (sinks) rotational waves emerge from unstable, or stable foci, respectively. Analogously for empirical data of humans, Das et al. (2024) investigated the organization of sinks and sources and their role for different memory tasks, showing that in spatial tasks more sources, in verbal memory tasks more sinks were detected. Deco et al. (2021a) found that large-scale models with regional heterogeneity of excitatory-inhibitory receptors are capable of reproducing the spatiotemporal structure of empirical functional connectivity reliably. Along the line of diversifying connectivity profiles (but without the inclusion of fine grained receptor-dynamics) Breakspear et al. (2003) emphasized the importance of balance between local short-range versus long-range connections1 for the transition from independent, locally appearing oscillations to chaotic synchronization to global patterns. Liang et al. (2023) supported this observation when investigating the spatiotemporal patterns in awake and anesthetized rodents. They not only emphasized the presence of complex local patterns during wakefulness but also showed, with computational modeling, that the coherence in low frequency bands is enhanced by stronger long-range connections between cortical areas further apart. Information processing has also been shown to be crucially affected by long-range connections by Deco et al. (2021b), where the authors compared two whole-brain models, one with connections which exponentially decayed with distance and one with additional sparse long-range connections that deviated from that rule. They investigated complex brain activity that is functionally beneficial for the transmission of information between cortical regions and found the information cascade, i.e., the flow of brain activity across different spatial scales, to be significantly improved by the presence of these long-range connections. Studies such as the above, where brain activity is simulated with networks equipped with empirically informed structure, have been shown to reliably predict empirically observed patterns. Cakan et al. (2022), for example, showed that the observed direction of SOs can be implicated by the antero-posterior structural connectivity gradient that decreases in connectivity strength from the anterior to the posterior direction.
Given the large number of computational modeling studies which investigate the spatiotemporal organization of neural activity on larger scales, we are left with the question in how far results generalize across the different whole-brain modeling approaches. Here, we specifically investigate whether, and how strongly, the specific choice of the dynamical system and of the spatial resolution changes the observed patterns, and how the connectivity profiles affect the emergent dynamics beyond empirically observed variability. We compare the emergent dynamics of whole-brain models based on the biophysically realistic adaptive linear-nonlinear cascade (aLN) model (Augustin et al., 2017; Cakan and Obermayer, 2020; Cakan et al., 2022) and the phenomenological Wilson-Cowan model (Wilson and Cowan, 1972), both equipped with spike-frequency adaptation as a fatigue mechanism. To identify the role of spatial density in the models, we show the results for three network parcellations based on the Schaefer local-global parcellation schemes (Schaefer et al., 2018) with 100, 200, and 500 nodes. We find that the coarse-grained dynamical landscape remains robust across models and network resolutions. However, results may not generalize when exploring specific dynamical states.
2 Materials and methods
2.1 Data
2.1.1 Participants
We used diffusion tensor imaging (DTI) data and anatomical T1 scans which were acquired at the Universitätsmedizin Greifswald from 27 participants (15 females; age range = 50–78 years, mean age = 63.55 years). Prior to participating in the study, all participants gave a written informed consent and were subsequently reimbursed for participation. The study was approved by the local ethics committee at the Universitätsmedizin Greifswald and was conducted in accordance with the Declaration of Helsinki.
2.1.2 Data acquisition and preprocessing
The acquisition parameters and preprocessing of the DTI and anatomical T1 scans were identical to those described in Cakan et al. (2022).
We defined the anatomical regions according to the Schaefer cortical parcellation scheme (Schaefer et al., 2018) with 100, 200, and 500 nodes, respectively. We employed the same probabilistic tractography algorithm with 5,000 randomly sampled streamlines per voxel, which yielded one structural connectivity matrix and one fiber length matrix per participant. One participant was excluded because the tractography procedure at the highest network resolution failed. Following probabilistic tractography, we normalized the resulting structural connectivity matrix for each participant by dividing the connection probability
In addition to the individual connectomes, we constructed average structural connectivity matrices C and average fiber length matrices D for each parcellation.
2.2 Whole-brain network models
We used whole-brain networks that consist of
2.2.1 The aLN model
The adaptive linear-nonlinear (aLN) model is a mean-field neural mass model of a network of coupled adaptive exponential integrate-and-fire (AdEx) neurons. It was developed in Augustin et al. (2017) and validated against simulations of spiking neural networks in Cakan and Obermayer (2020). We used the neurolib framework introduced in Cakan et al. (2021) for the numerical simulations. The dynamics of each node (Cakan et al., 2022) is summarized by the following equations:
where
where
The mean adaptation current
All parameters not explained above are given and explained in Table 1. Values were chosen as in Cakan et al. (2022) with the global coupling strength
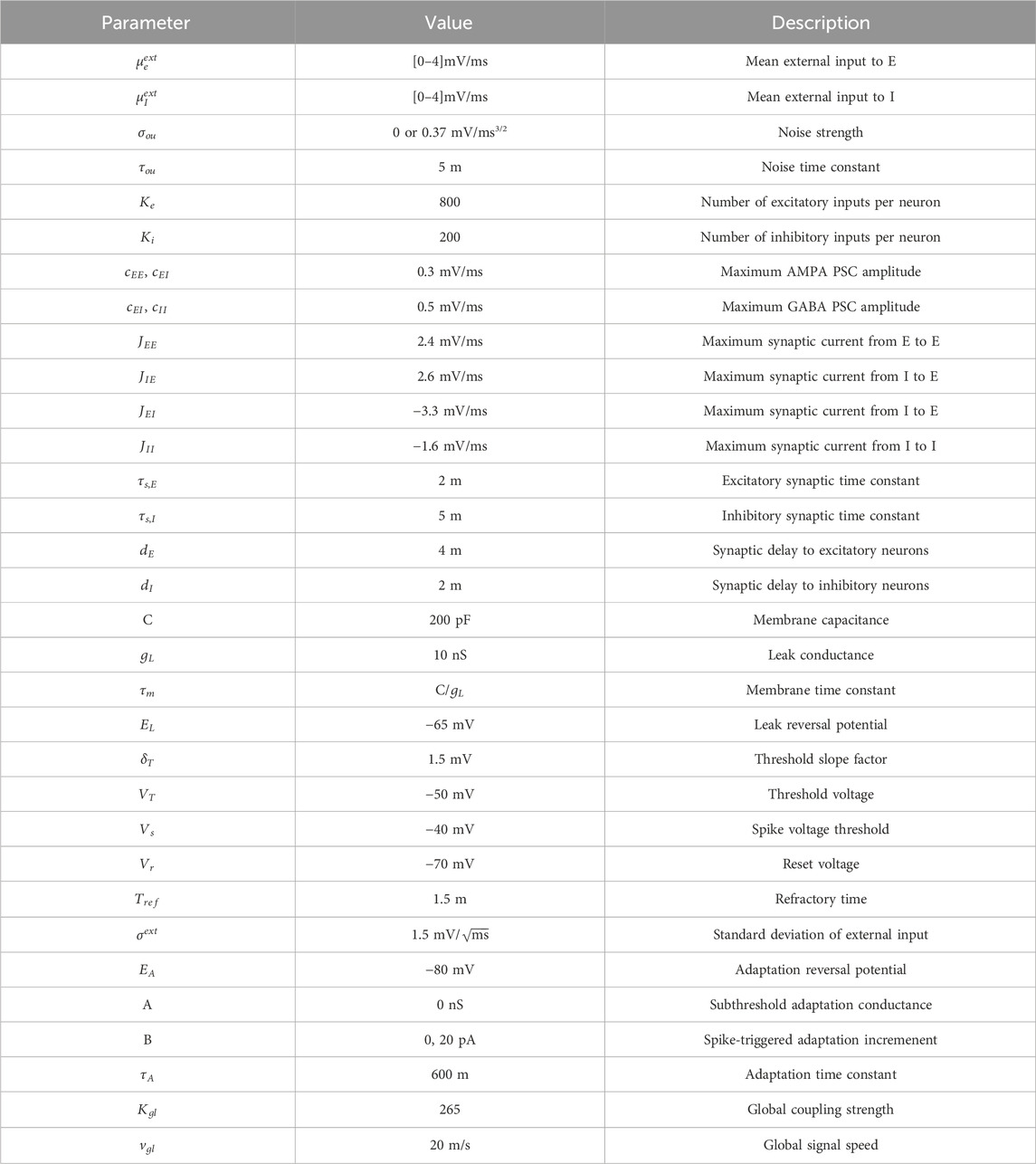
Table 1. Parameter values used for the aLN model. Values are taken from Cakan et al. (2022).
2.2.2 The Wilson-Cowan model
The Wilson-Cowan model (Wilson and Cowan, 1972) describes the dynamics of the proportions of excitatory
To simplify Equation 4, we consider a mean external input
A description for each parameter can be found in Table 2. These parameter values were chosen, because they give rise to a dynamical landscape which is similar to other systems that also reliably produce SOs (Cakan and Obermayer, 2020; Cakan et al., 2022). The parameter setting required minor adjustments compared to previous studies that used the Wilson-Cowan model to simulate various types of spatiotemporal patterns (Levenstein et al., 2019; Papadopoulos et al., 2020; Torao-Angosto et al., 2021).
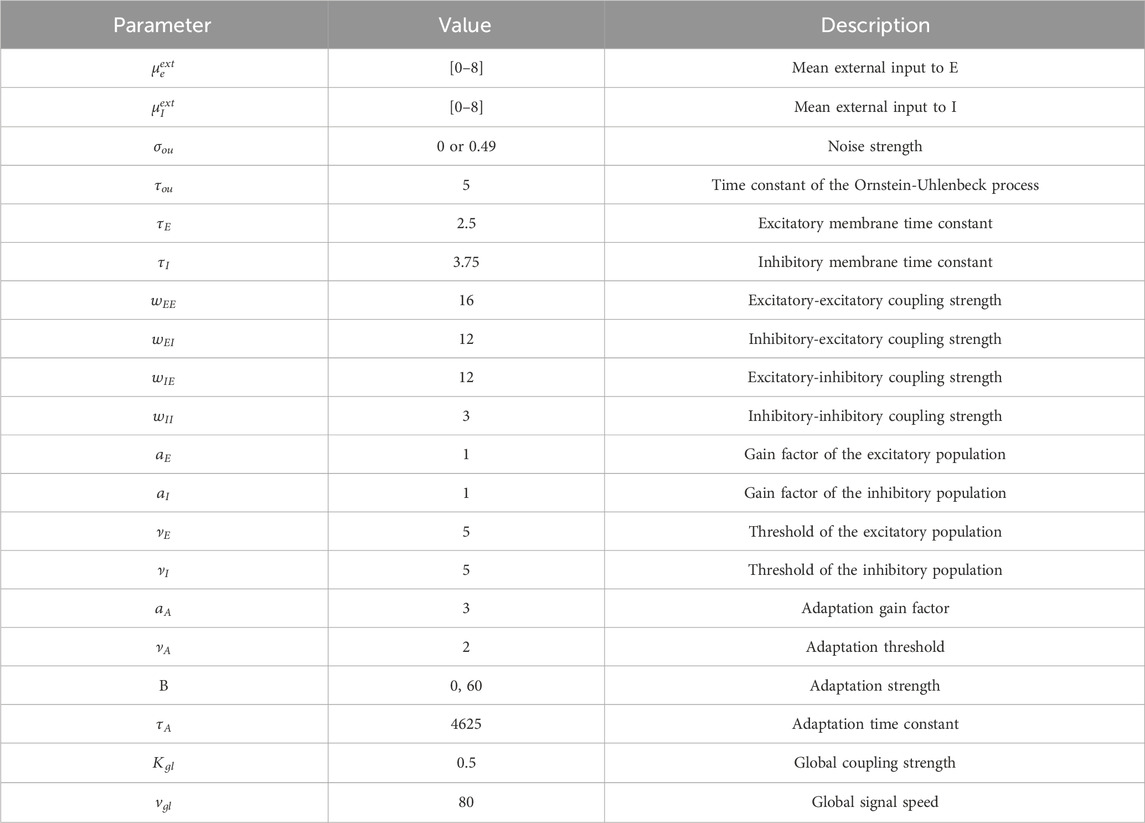
Table 2. Parameter values used for the Wilson-Cowan model.
2.2.3 Noise
For the investigation of simulated sleep SOs, noise input to each population
where
2.2.4 Numerical simulations
We use Euler-integration to conduct the numerical simulations. To compare computation times, we tracked the duration necessary to simulate both models at all resolutions on a single core of an AMD EPYC 7662 64-core processor. The simulation of 10 s (100.000 time steps, dt = 0.1 m) of activity with the aLN model (Wilson-Cowan model) takes 8.45 s (3.50 s) for 100, 16.65 s (8.32 s) for 200, and 95.46 s (44.91 s) for 500 nodes.
2.3 Analysis
2.3.1 State space analysis
The analysis of the state space was conducted numerically in the absence of noise. We randomly initialized and simulated the model for 101 x 101 parameter values (10,201 simulations in total) for the mean external inputs to the E and I populations for a duration of 30 s. The duration was extended to 1 min for the Wilson-Cowan model with adaptation, as in some cases
For every point in state space, we applied a negative, but increasing, followed by a positive, but decaying stimulus. Subsequently, we computed the difference between the average
Furthermore, we computed the difference between the maximum and minimum value of
Supplementary Figure SA1 shows the single-node bifurcation diagrams for both models with and without adaptation obtained using the procedures described above.
2.3.2 State classification
To characterize the temporal dynamics of each point in the oscillatory regions (identified as described in Section 2.3.1), we used the procedure summarized in Figure 1. For each point in the slice of state space spanned by the external input currents to the excitatory and inhibitory populations,
where
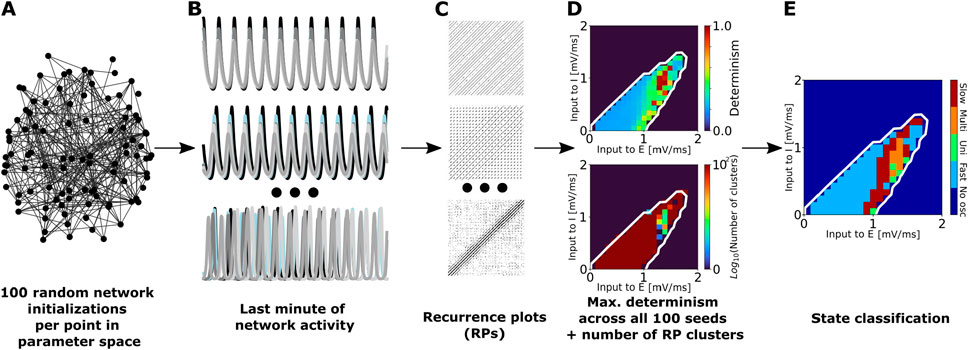
Figure 1. Summary of the procedure used to classify network states into unistable, multistable, fast metastable, and slow metastable. (A) For each model (aLN or Wilson-Cowan) and each parcellation (100, 200, or 500 nodes) we conducted 100 randomly initialized simulations of 2 min duration for each point in the slice of parameter space spanned by varying the external excitatory
For each parametrization, we clustered the resulting recurrence matrices using the DBSCAN algorithm (Ester et al., 1996). Additionally, we computed the determinism value DET,
for each initialization, where
We used the number of clusters to classify each state in the limit cycle as either unistable (if the number of clusters was equal to 1), multistable (if the number of clusters was
2.3.3 Kuramoto order parameter
Using the simulation data described in Section 2.3.1, we computed the Kuramoto order parameter R(t),
where
Subsequently, we summarized the results for each model and each network resolution using the mean and the standard deviation of
2.3.4 Interhemispheric cross-correlation
To investigate spatial properties of oscillatory states, we computed the sliding-window time-lagged cross-correlation as in Roberts et al. (2019). We calculated the intrahemispheric Kuramoto order parameter for each hemisphere. Subsequently, the windowed time-lagged cross-correlation between the two parameters was determined with a window of length W of 100 m and 90% overlap between consecutive windows, and with a lag
where
2.3.5 Singular value decomposition
To conduct a singular value decomposition (SVD), we firstly computed the velocity vector fields (cf. Roberts et al., 2019). For each node n, we used the instantaneous Hilbert transform of
The spatial derivative was calculated using the constrained natural element method (Illoul and Lorong, 2011), as described in Roberts et al. (2019). This method allows for the calculation of the components of the gradient vector without the need for interpolation to and from a 3D grid.
The SVD was then performed for the velocity vector fields
where the columns of U represent the left and the rows of
We then projected the spatial modes identified on the concatenated data onto the individual vector velocity fields of each parametrization and quantified the proportion of explained variance by each projected spatial mode
2.3.6 Structural gradient manipulation
To investigate the effect of the structural gradient on the propagation of SOs, we used the sleep model parametrization introduced in Cakan et al. (2022) for the aLN model, with minor adjustments of the adaptation parameters (see Supplementary Table SA4). The adjustment was necessary because the parcellations of higher resolution had stronger pairwise connectivity strengths compared to the 100 node case, which caused the model to be in the up state for prolonged intervals of time due to a shift in state boundaries. The manual increase of the adaptation parameters ensured that the model visually displayed SOs (Cakan et al., 2022). For the Wilson-Cowan model with 100 nodes, we conducted an evolutionary optimization in neurolib, with resting-state functional connectivity and functional connectivity dynamics and with power spectrum of EEG in sleep stage N3 as optimization objectives (full procedure described in Cakan et al. (2022)). As the evolutionary optimization was computationally not feasible for the networks with 200 and 500 nodes, we manually adjusted the adaptation parameters obtained for the network with 100 nodes (see Supplementary Table SA4) in the same manner as described above for the aLN model. To compare our results with previous work, compare dynamical landscapes across models and resolutions, we used the parameters given in Tables 1, and 2. For the sleep models, we modified a small number of parameters to place the model in a regime, where realistic SOs are produced.
The antero-posterior structural connectivity gradient defined as the slope of the linear regression between the node degree and its coordinate along the antero-posterior axis of the brain (Cakan et al., 2022) is shown in Supplementary Figure SA18 for the three parcellations. A more negative slope indicates that more anterior nodes have lower node degrees compared to more posterior nodes.
To manipulate the antero-posterior gradient, we introduced a parameter p, which represents the maximum percentage change in connection strengths applied to the most anterior node. We then computed N equidistant and increasing values
To isolate the effect of the gradient from other network properties, we constructed control models with gradient values similar to the networks described above (within a small tolerance limit). In these models, we preserved the total sum of connection strengths (i.e., the sum of all entries in the structural connectivity matrix), but destroyed the relationship between the connection strength and the corresponding fiber length. This was achieved by randomly permuting the entries of the structural connectivity matrix and calculating the antero-posterior gradient value as described above; this was repeated until the gradient was similar (within a tolerance) to the target value. Since the corresponding fiber length matrix was kept fixed, any relation between connection strength and distance was destroyed. Hence stronger connection strengths no longer corresponded to longer fibers on average.
To determine the direction of propagation of SO up/down state transitions along the antero-posterior axis, we first computed the proportion of regions in the down state as a function of time. The down states were identified by thresholding the excitatory
2.4 Manipulation of short- vs. long-range connection strengths
We collected all pairs
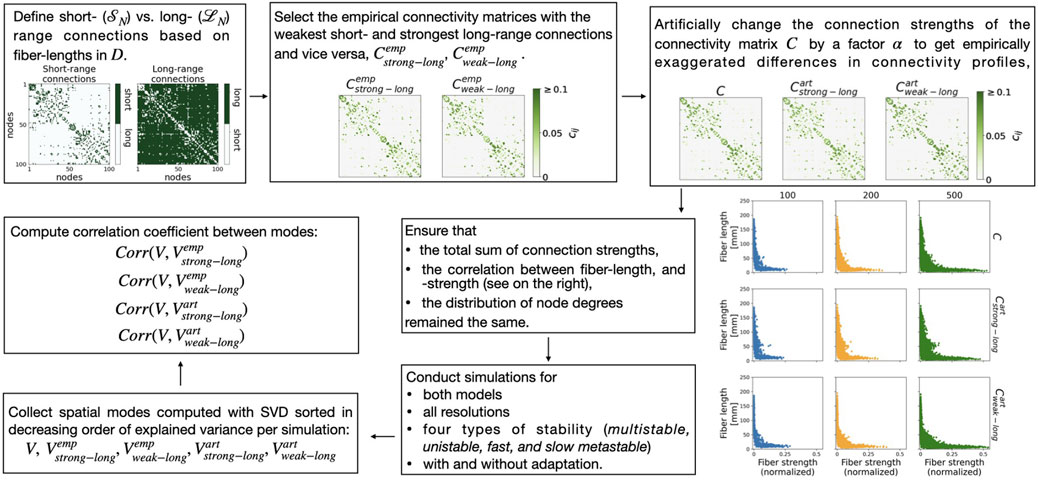
Figure 2. Summary of the procedure used to manipulate and investigate the effect of weaker versus stronger long-range connection strengths on the network dynamics. For an explanation, see text.
We identified the subjects with the weakest short- and the strongest long-range connections
and retained the corresponding connectivity matrices
To artificially manipulate two matrices beyond the empirically observed variability (see Figure 2, panel in the top right), we used the factors
Thus we ensured that the total sum of connections strengths remained constant, i.e.,
Furthermore, we individually inspected the total sum over the short- and long-range connections of
2.4.1 Correlation coefficient between spatial modes
We conducted numerical simulations for four locations in the state space covering unistability, multistability, fast and slow metastable patterns (see Supplementary Figure SA13). Simulations were performed for both models with and without adaptation, for the averaged connectivity matrix
This was done for each selected state, with and without adaptation, for the aLN and the Wilson-Cowan models, and for all three parcellations. The resulting correlation coefficient matrices have values ranging between
2.4.2 Coherence values
As in Section 2.3.6, we conducted numerical simulations of SOs for the aLN (parameters, see Supplementary Table SA4) and the Wilson-Cowan model (parameters, see Supplementary Table SA5) using the average connectivity matrix
For each numerical simulation, we computed, analogously to Liang et al. (2023), the magnitude-squared coherence
where
We separately consider the coherence between nodes connected with a short range (i.e., all pairs
3 Results
3.1 State space
Figures 3, 4 show the results of the state space analysis for the whole-brain aLN and Wilson-Cowan models. In line with previous results for the aLN model (Cakan et al., 2022), we identify several dynamical regimes: a down-state, where all network nodes display no or low activity; an up-state, characterized by constant high firing rate; an oscillatory region
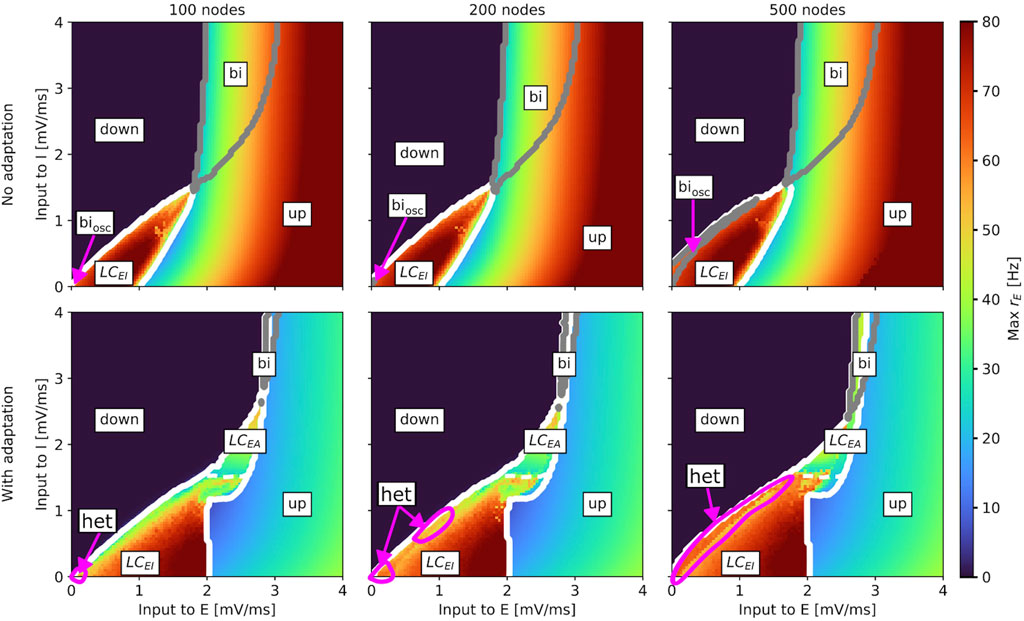
Figure 3. Slice of state space of the whole-brain aLN model without (b = 0 pA; top row) and with (b = 20 pA; bottom row) adaptation for a brain network with 100 (left column), 200 (middle column), and 500 (right column) nodes spanned by the external input currents to the E and I populations. In every panel, the horizontal axis shows the external input current to the excitatory population
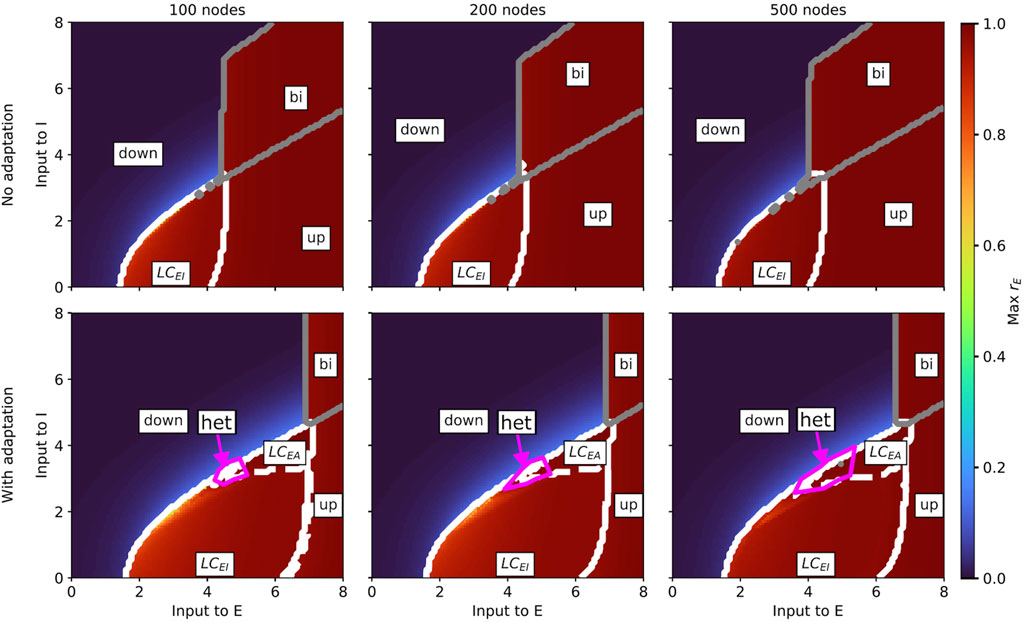
Figure 4. Slice of state space of the whole-brain Wilson-Cowan model without (b = 0; top row) and with (b = 60; bottom row) spike-triggered adaptation for a brain network with 100 (left column), 200 (middle column), and 500 (right column) nodes spanned by the external input currents to the E and I populations. In every panel, the horizontal axis shows the external input current to the excitatory population
Our results show that, for both models, the state space remains generally robust to changes in network resolution, but there are some differences between the aLN and the Wilson-Cowan implementations. For the aLN model, we observe a region of bistability between the down-state and the
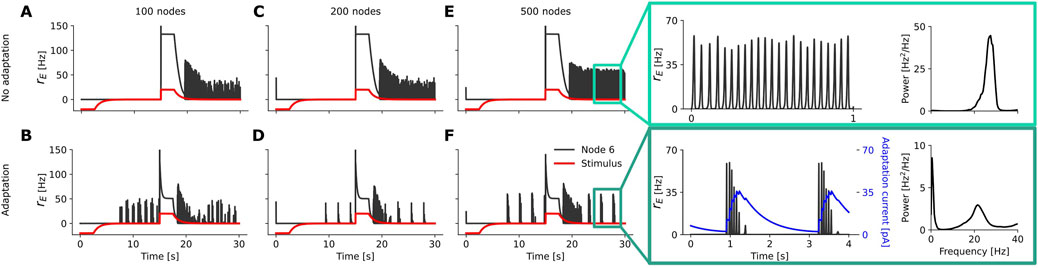
Figure 5. Example time series of the firing rate
For the Wilson-Cowan model, we also find a region of heterogeneous oscillations in the case with adaptation (example time series in Supplementary Figure SA2), which expands with increasing network resolution (bottom row in Supplementary Figures SA4, 6). However, in contrast to the aLN model, this region emerges at the border between the
3.2 State classification
The analysis methods presented in Section 2.3.2 allowed us to identify four types of states (unistable, multistable, fast metastable, and slow metastable) across the
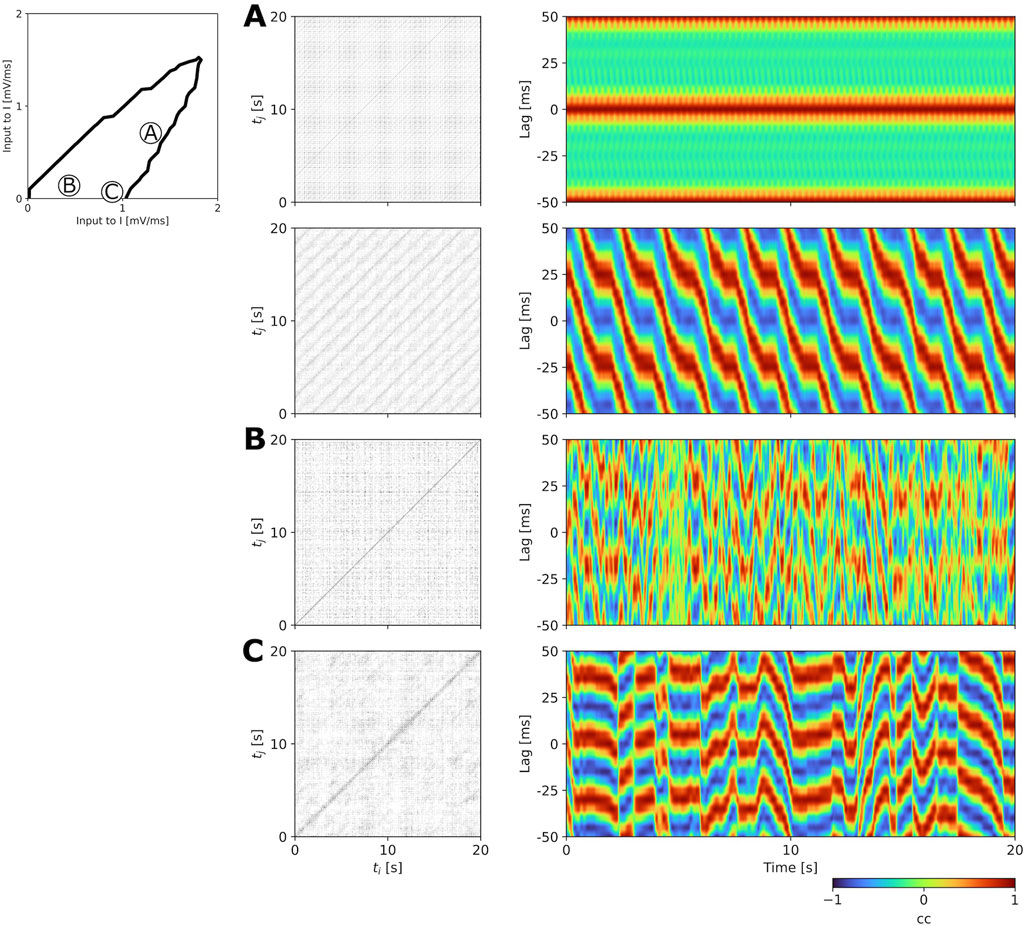
Figure 6. Examples of multistable (A), fast metastable (B), and slow metastable (C) states of the aLN model with 100 nodes and without adaptation (
Results of the state classification for the entire slice of state space are summarized in Figure 7 (aLN model) and Figure 8 (Wilson-Cowan model). Qualitatively, the results are similar across models and resolutions, with all four regimes being present in all cases, and with the fast metastable regime occupying the largest portion of the
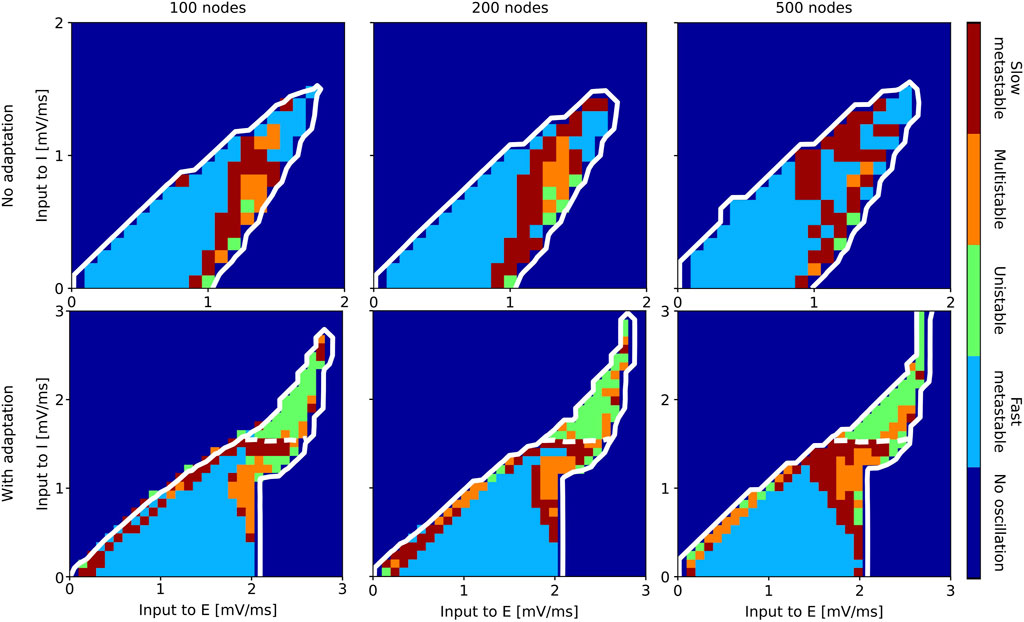
Figure 7. Classification of states inside the oscillatory regions for the aLN whole-brain model in the case without (b = 0 pA; top row) and with (b = 20 pA; bottom row) adaptation for a parcellation with 100 (left column), 200 (middle column), and 500 (right column) nodes. The slice of state space is spanned by the external input current to the E and I populations. The white solid contour marks the two oscillatory regions, and the white dashed lines indicate the approximate border between them.
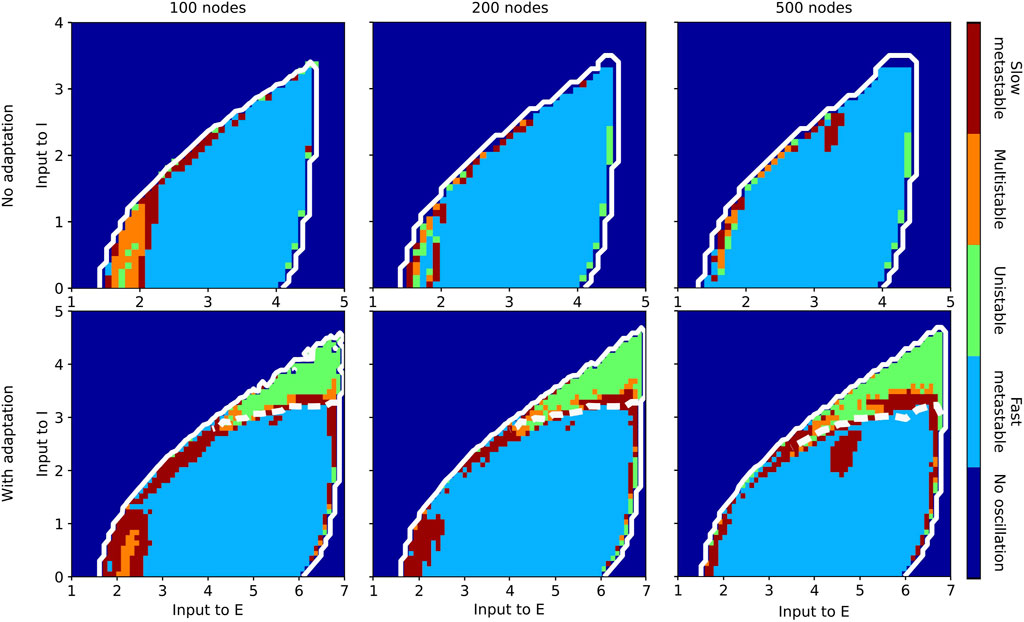
Figure 8. Classification of states inside the oscillatory regions for the Wilson-Cowan whole-brain model in the case without (b = 0; top row) and with (b = 60; bottom row) adaptation for a parcellation with 100 (left column), 200 (middle column), and 500 (right column) nodes. The slice of state space is spanned by the external input current to the E and I populations. The white solid contour marks the two oscillatory regions, and the white dashed lines indicate the approximate border between them.
As metastability is usually identified based on the mean and the standard deviation (SD) of the Kuramoto order parameter (metastability corresponds to a high standard deviation of the Kuramoto order parameter), we report these results for completeness in Supplementary Figures SA7, 8. The results show high synchrony (mean Kuramoto
3.3 Spatial modes of activity
Our analysis of the spatial modes of activity reveals that, in general, the modes which explain a larger proportion of variance of the activity (percentages given in Supplementary Tables SA1, 2) in the concatenated data (obtained by concatenating the velocity vector fields computed for each point in the oscillatory regions, with time steps in rows and nodes in columns) consist of large-scale waves traveling mainly along the horizontal and dorso-ventral axes. The results are summarized in Figures 8A,B for the aLN model and in Supplementary Figures SA12a, b in the appendix for the Wilson-Cowan model. For example, modes 1 and 4 in the aLN model (Figure 8A) and modes 2 and 4 in the Wilson-Cowan model (Supplementary Figure SA12a) exemplify large-scale waves with coherent horizontal and dorso-ventral directions of propagation encompassing approximately three-quarters of the brain. Another example of a large-scale wave pattern is represented by the hemispheric-segregated pattern present in the Wilson-Cowan model (mode 3 in Supplementary Figure SA12a) and in the aLN model (mode 9 in Figure 9). Interestingly, these modes explain similar proportions of variance (1.78% in the aLN vs. 1.44% in the Wilson-Cowan model). In contrast, modes explaining less variance within each model and each resolution usually capture more complex patterns of propagation. For example, in both models, mode 13 (Figure 9A, Supplementary Figure SA12b) displays smaller clusters of arrows with the same color and direction (i.e., same horizontal and dorso-ventral directions), as well as more neighboring arrows with different colors and directions compared to the large-scale modes indicated above. While we identify similar modes in both models (see above), the overall proportion of variance explained by the 15 first modes differs (30.28% for the aLN vs. 9.19% for the Wilson-Cowan model with adaptation). There is also a tendency towards decreased explained variance per mode with increasing model resolution, as well as differences in the percentages of variance explained by the dominant modes between the models with and without adaptation (Supplementary Tables SA1, 2).
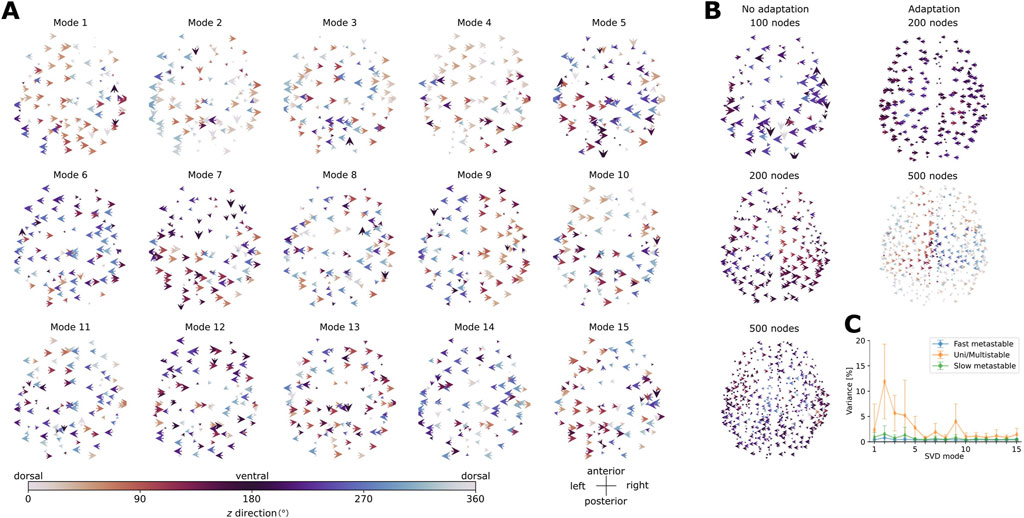
Figure 9. (A) First 15 modes obtained from the singular value decomposition of the velocity vector fields in the whole-brain aLN model with 100 nodes and spike-triggered adaptation (b = 20 pA). Modes are ordered in decreasing order of explained variance. (B) Left panels: Modes explaining the largest proportion of variance for the whole-brain aLN model without spike-triggered adaptation (b = 0 pA) with a parcellation of 100, 200 and 500 nodes. Right panels: Same as before, but with spike-triggered adaptation (b = 20 pA) and with a parcellation of 200 and 500 nodes. The arrows represent the orientation in the
To verify whether the modes obtained from the decomposition of the concatenated data can be reliably identified in the individual velocity vector fields computed for each parametrization in the oscillatory regions
As a further example, we examined the spatial modes of activity in the
3.4 Similarity of spatial modes of imbalanced short-versus long-range connection strengths
To identify the impact of the balance between short- and long-range connection strength, we compared the
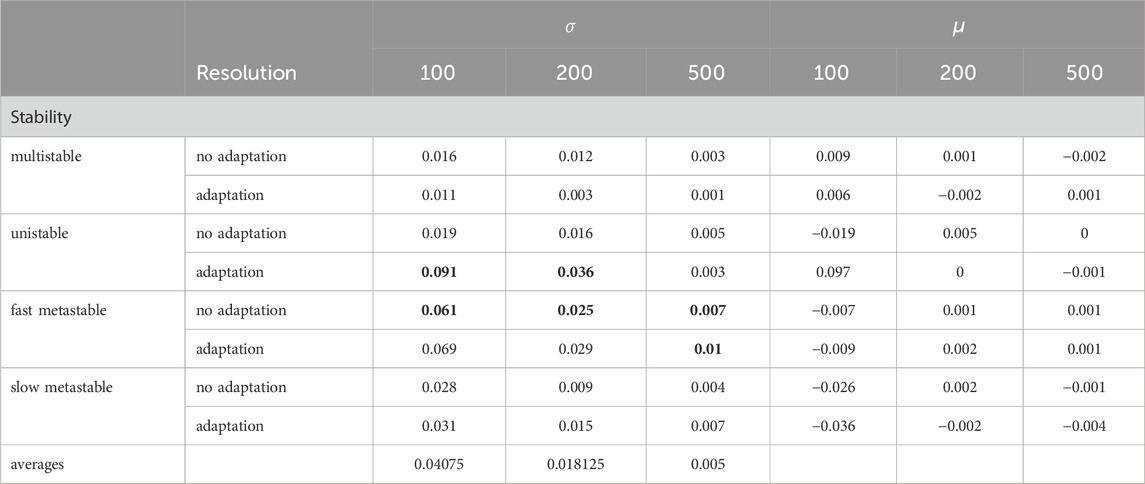
Table 3. Average standard deviation
All distributions are centered around a value of zero. However, we notice that the standard deviations (Table 3) for the fast metastable states are the largest (indicating higher similarity between spatial modes). Due to the model systems experiencing a higher amount of metastable attractors, inducing fast switching between activity states, they are inherently less stable compared to unistable or slow metastable states. Only for the cases of unistability with adaptation at resolutions
Nonetheless, we see a higher similarity of spatial organization in states of stability that promote more local, complex activity patterns rather than the global, synchronized patterns that appear in unistable or multistable states. While the states showing the broadest widths differ between both models (multistable states for the Wilson-Cowan model vs. unistable or fast metastable states for the aLN model), the overall low similarity in the spatial organization between activity patterns caused by the average connectivity matrix vs. by the connectivity matrices with weaker and stronger long-range connections generalizes across all resolutions, both model types and all settings. Finally, we see that the results of the comparison between the spatial organization of activity patterns induced by the different connectivity matrices are predominantly the same for the artificial versus empirical connectivity matrices for both models and all resolutions.
3.5 Effect of the antero-posterior gradient of structural connectivity strengths on sleep SO propagation
The results presented above show that for both the aLN and Wilson-Cowan models dynamical features remain generally robust to changes in the parcellation. Also, the phenomenological Wilson-Cowan model is capable of producing qualitatively similarly complex spatiotemporal dynamics as the biophysically realistic aLN model. In the current section, we explore whether this remains to be the case when both models are applied to the phenomenon of sleep SO propagation (Cakan et al., 2022). In particular, we examine whether the relation between the antero-posterior structural connectivity gradient and the propagation of sleep SOs as waves of silence from anterior to posterior brain areas remains present in both models and for all parcellations. Furthermore, we test whether changes in the strength of this connectivity gradient have a causal effect on the direction of propagation of SOs.
Figure 10 shows that the relation reported in Cakan et al. (2022) is present in both the aLN and Wilson-Cowan models for all three network resolutions. Furthermore, decreasing the gradient strength along the antero-posterior axis causes a reversal of the direction of SO propagation, with down states being initiated preferentially in posterior areas and traveling towards the front of the brain. Increasing the gradient strength increases this preference to propagate from anterior to posterior areas. In the Wilson-Cowan model, however, the relation between node degree and the transition phase decreases with the increase in resolution, as the magnitude of the correlation coefficients decreases at higher resolutions. This could potentially be caused by the fact that in the Wilson-Cowan model the adaptation strength
Figure 10. Correlation coefficient between the mean transition phases of the nodes from the up to the down state (blue) and vice versa (orange) and the node coordinates along the antero-posterior axis as a function of the percentage by which the connection strengths of the most anterior node were changed. The left (right) column shows results for the aLN (Wilson-Cowan) models with 100 (top row), 200 (middle row), and 500 nodes (bottom row). 0% corresponds to the unchanged structural antero-posterior gradient where the value of the
To ensure that the results presented in Figure 10 are not due to changes in the underlying network topology induced by the specific gradient manipulation method, we employ a control model in which we preserve the total sum of connection strengths in the network and destroy the relation between fiber length and connection strength (cf. Section 2.3.6). Supplementary Figure SA19 shows that the relationship described above remains present in the aLN model at all three network resolutions. In the Wilson-Cowan model, destroying the relation between the distance and connectivity strength destroys and even reverts the propagation direction of SOs, suggesting that the model is more sensitive to changes in the particular structure of the connectome.
3.6 Stronger long-range connections lead to an increase in coherence as observed empirically
Motivated by the findings that show that rare long-range connections play an effective role in the cascade of information processing (see Deco et al., 2021b) and that stronger long-range connections correlate with enhanced coherence between cortical regions over lower frequency ranges (Liang et al., 2023), we investigated how changes in the strength of long- versus short-range connections influence waves of SOs.
Since long-range connections are assumed to play a crucial role in the propagation of global patterns, we assume that the stronger the long-range connections, the higher the coherence over lower frequency values induced by slow oscillations. We therefore compared results obtained using the matrices
Figure 11; Supplementary Figure SA19 show the average power spectra and coherence values for the aLN and the Wilson-Cowan models for three different parcellations. In Figure 11A we see that for all parcellations the dominant temporal frequencies are
Figure 11. Power and coherence as a function of frequency for SO activity generated by the aLN model. Results are shown for the average connectivity matrix,
In the artificial case, we see that the change of coherence over frequency for the aLN (see Figure 11B) and for the Wilson-Cowan model (see Supplementary Figure SA20b) agrees with our expectation. The coherence over low frequencies is higher for SOs induced by
Table 4. Maximum coherence values for non-zero frequencies for the metastable states of the aLN model for all settings shown in Figure 10B. Both for the artificial and the empirical case, values in bold indicate the highest coherence values per parcellation, per set of nodes connected on a short-range (long-range). The corresponding frequencies were 0.5 Hz for all settings. Parameters are as for Figure 10.
The results of the Wilson-Cowan model agree mostly with the results of the aLN, however, the dominant temporal frequency varies more strongly depending on the parcellation and the used connectivity matrix, see Supplementary Table SA7. The coherence values are consistently larger for waves of SOs induced by
We observe the expected effect in neither model for the empirical matrices. We argue that this is due to the fitting process applied to the averaged matrix
Overall, models and resolutions agree with the expected increase in coherence values over low frequencies for the artificially manipulated matrices, but do not display the same effect for the empirically selected matrices.
4 Discussion
In this work, we investigated whether we can employ generalized whole-brain models for the study of complex brain dynamics or whether the latter are significantly influenced by the choice details of the dynamical system and the parcellation. To that end, we compared a biophysically realistic model (aLN) and a phenomenological model (Wilson-Cowan) with similar state spaces and bifurcations at three network resolutions (the Schaefer parcellation scheme with 100, 200, 500 nodes). Overall, we found that the results remain relatively robust to changes in both model and parcellation, but dynamics at detail appear sensitive to these changes, indicating the need for careful model adjustment depending on the application.
We started our analysis with the exploration of the coarse-grained structure of the dynamical landscape. We found that both the aLN and the Wilson-Cowan model display a down state of no or low activity, an up state of constant high activity, a fast limit cycle, where the activity oscillates between low and high values with frequencies
In a second step, we classified the oscillatory network states. We identified four types of states, namely, unistable, multistable, slow, and fast metastable states, in both models and at all resolutions, and observed quantitative changes with respect to the distribution of each type of state in the oscillatory regimes both across models and across resolutions (Figures 7, 8). Our detailed analysis of the types of oscillatory network states revealed that complex wave dynamics emerge even at low network resolutions and in relatively simple phenomenological models. Furthermore, the detailed mapping of the oscillatory regimes presented here can provide useful information for further studies aiming to explore induced state transitions, such as, for example, through the application of electrical stimulation (for example, see Ladenbauer et al. (2017); Ladenbauer et al. (2023)).
We explored large-scale patterns through the spatial modes obtained from singular value decomposition. We found that results are qualitatively similar across models and resolutions, but that specific patterns emerge depending on either model or resolution. Given that recent work (Das et al., 2024; Mohan et al., 2024) investigating the relation between spatiotemporal wave patterns and cognitive function has shown an association between specific patterns and specific behavioral processes, future modeling work in this direction should take into account the variability introduced by model and parcellation when exploring such phenomena. In alignment with the variability introduced by the model components investigated in this study, Roberts et al. (2019) additionally emphasize the importance of delays for enabling the emergence of complex wave patterns in whole-brain models. It is important to note that the conduction velocities used in this study, while in agreement with the work of Cakan et al. (2022), are higher compared to those of Petkoski and Jirsa (2022). Given that axonal delays can play a significant role in the dynamics of complex oscillatory networks, future work should investigate the effects of decreasing the conduction velocity on the emergent wave patterns.
We showed that changes in the balance of connectivity strengths between short- and long-range connections alter the spatial organization in states exhibiting global patterns (multi- and unistable) as well as complex patterns (fast and slow metastable), a result which stays predominantly consistent across models, resolutions, parametrizations, and states (see Supplementary Figures SA16, 17). Artificially manipulating the long- versus short-range connection strengths beyond empirically observed variability had no significantly different effect to the loss of similarity between the spatial organization of activity patterns induced by the artificially manipulated and the empirical connectivity matrices. Furthermore, we noticed that the strongest similarity in the spatial modes collected from the activity patterns caused by the different connectivity matrices was observable in the fast and slow metastable states in which complex local activity patterns emerge (see Kelso, 2012).
Furthermore, in the specific case of sleep SOs, we have shown that the aLN model is robust to changes in network resolution and even in parcellation scheme (as we used the original parametrization introduced in Cakan et al. (2022) with only minimal parameter adjustments). In this case, we were also able to demonstrate that changes in the antero-posterior structural connectivity gradient have a causal effect on the propagation of SOs. In contrast, the Wilson-Cowan model required optimization for the Schaefer parcellation scheme with 100 nodes and an additional adjustment of its parameters for higher resolutions. Here, manipulating the antero-posterior gradient of node degrees showed a robust causal effect only in the case where the model parameters were explicitly fitted to data rather than adjusted to support SO activity. The model also displayed high sensitivity to the changes in of the relationship between connection strength and distance.
For understanding the impact of changes in the strength of short- vs. long-range connections on SOs, we investigated power spectra and coherence values (see Figure 11; Supplementary Figure SA20). For the case of artificially manipulated connectivity matrices we found the coherence in lower frequency bands
For the empirical connectivity matrices
We thus conclude that the deployment of whole-brain models for the investigation of the coarse-grained dynamics provides results which are fairly independent of model type and resolution. All model variants enable the same dynamical landscape with qualitatively similar changes in dynamical features with resolution and with the manipulation of the connectivity profiles. In the specific application to sleep SOs, both the phenomenological and the biophysically realistic model show similar changes in the temporal dynamics. While the antero-posterior directionality of simulated SOs by the aLN corresponds to the expected changes induced by the manipulation of the underlying antero-posterior structural connectivity gradient, the phenomenological Wilson-Cowan model requires a much more careful handling to demonstrate the empirically observed directionality. In total, this indicates that both model types are fairly robust to the simulation of empirically realistic temporal features, but not so for propagation dynamics. While computational costs increase (see Section 3.5) with resolution, the investigation of the fine-grained dynamics of wave propagations benefits from the higher resolution, for example, the higher resolved parcellations showed more robustness to the manipulation of connectivity profiles (see Section 3.4), and to the manipulation of structural gradients (see Section 3.5). In addition, if wave propagation across cortical space becomes the subject of investigation, a high resolution is mandatory. Nonetheless, for the investigation of quantitative features, detailed dynamics or specific application cases, the phenomenological Wilson-Cowan model requires a much more careful handling and finer tuning, while the biophysically realistic aLN model allows the investigation of specific features in a more reliable way.
Data availability statement
The original contributions presented in the study are included in the article/Supplementary Material, further inquiries can be directed to the corresponding author.
Ethics statement
The studies involving humans were approved by Local Ethics Committee at the Universitätsmedizin Greifswald. The studies were conducted in accordance with the local legislation and institutional requirements. Written informed consent for participation was obtained from the participants in accordance with the national legislation and institutional requirements.
Author contributions
CD: Conceptualization, Formal Analysis, Methodology, Visualization, Writing – original draft, Writing – review and editing. RS: Conceptualization, Formal Analysis, Methodology, Visualization, Writing – original draft, Writing – review and editing. AF: Investigation, Writing – review and editing. KO: Funding acquisition, Supervision, Writing – review and editing.
Funding
The author(s) declare that financial support was received for the research and/or publication of this article. This work was funded by the Deutsche Forschungsgemeinschaft (DFG, German Research Foundation) Project number 327654276, SFB1315, project number B03 (KO and AF) and project grants to AF: Research Unit 5429/1 (467143400), FL 379/34-1, FL 379/35-1.
Acknowledgments
A pre-print version of this paper was published at Dimulescu et al. (2025).
Conflict of interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Generative AI statement
The author(s) declare that no Generative AI was used in the creation of this manuscript.
Publisher’s note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
Supplementary material
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fnetp.2025.1589566/full#supplementary-material
Footnotes
1Breakspear et al. (2003) refer to excitatory couplings between cortical columns as long-range connections.
References
Augustin, M., Ladenbauer, J., Baumann, F., and Obermayer, K. (2017). Low-dimensional spike rate models derived from networks of adaptive integrate-and-fire neurons: comparison and implementation. PLoS Comput. Biol. 13, e1005545. doi:10.1371/journal.pcbi.1005545
Bhattacharya, S., Brincat, S. L., Lundqvist, M., and Miller, E. K. (2022). Traveling waves in the prefrontal cortex during working memory. PLoS Comput. Biol. 18, e1009827. doi:10.1371/journal.pcbi.1009827
Breakspear, M., Terry, J. R., and Friston, K. J. (2003). Modulation of excitatory synaptic coupling facilitates synchronization and complex dynamics in a biophysical model of neuronal dynamics. Netw. Comput. Neural Syst. 14, 703–732. doi:10.1088/0954-898X/14/4/305
Cabral, J., Kringelbach, M., and Deco, G. (2012). Functional graph alterations in schizophrenia: a result from a global anatomic decoupling? Pharmacopsychiatry 45, S57–S64. doi:10.1055/s-0032-1309001
Cakan, C., Dimulescu, C., Khakimova, L., Obst, D., Flöel, A., and Obermayer, K. (2022). Spatiotemporal patterns of adaptation-induced slow oscillations in a whole-brain model of slow-wave sleep. Front. Comput. Neurosci. 15, 800101. doi:10.3389/fncom.2021.800101
Cakan, C., Jajcay, N., and Obermayer, K. (2021). neurolib: a simulation framework for whole-brain neural mass modeling. Cogn. Comput. 15, 1132–1152. doi:10.1007/s12559-021-09931-9
Cakan, C., and Obermayer, K. (2020). Biophysically grounded mean-field models of neural populations under electrical stimulation. PLoS Comput. Biol. 16, e1007822. doi:10.1371/journal.pcbi.1007822
Capone, C., De Luca, C., De Bonis, G., Gutzen, R., Bernava, I., Pastorelli, E., et al. (2023). Simulations approaching data: cortical slow waves in inferred models of the whole hemisphere of mouse. Commun. Biol. 6, 266. doi:10.1038/s42003-023-04580-0
Chang, Y.-N., Halai, A. D., and Lambon Ralph, M. A. (2023). Distance-dependent distribution thresholding in probabilistic tractography. Hum. Brain Mapp. 44, 4064–4076. doi:10.1002/hbm.26330
Das, A., Zabeh, E., Ermentrout, B., and Jacobs, J. (2024). Planar, spiral, and concentric traveling waves distinguish cognitive states in human memory. bioRxiv, 2024–01doi. doi:10.1101/2024.01.26.577456
Dasilva, M., Camassa, A., Navarro-Guzman, A., Pazienti, A., Perez-Mendez, L., Zamora-López, G., et al. (2021). Modulation of cortical slow oscillations and complexity across anesthesia levels. NeuroImage 224, 117415. doi:10.1016/j.neuroimage.2020.117415
Deco, G., Kringelbach, M. L., Arnatkeviciute, A., Oldham, S., Sabaroedin, K., Rogasch, N. C., et al. (2021a). Dynamical consequences of regional heterogeneity in the brain's transcriptional landscape. Sci. Adv. 7, eabf4752. doi:10.1126/sciadv.abf4752
Deco, G., Kringelbach, M. L., Jirsa, V. K., and Ritter, P. (2017). The dynamics of resting fluctuations in the brain: metastability and its dynamical cortical core. Sci. Rep. 7, 3095. doi:10.1038/s41598-017-03073-5
Deco, G., Perl, Y. S., Vuust, P., Tagliazucchi, E., Kennedy, H., and Kringelbach, M. L. (2021b). Rare long-range cortical connections enhance human information processing. Curr. Biol. 31, 4436–4448.e5. doi:10.1016/j.cub.2021.07.064
Dimulescu, C., Strömsdörfer, R., Flöel, A., and Obermayer, K. (2025). On the robustness of the emergent spatiotemporal dynamics in biophysically realistic and phenomenological whole-brain models at multiple network resolutions
Ester, M., Kriegel, H.-P., Sander, J., and Xu, X. (1996). A density-based algorithm for discovering clusters in large spatial databases with noise. Knowl. Discov. Data Min. 96, 226–231.
Fousek, J., Rabuffo, G., Gudibanda, K., Sheheitli, H., Petkoski, S., and Jirsa, V. (2024). Symmetry breaking organizes the brain’s resting state manifold. Sci. Rep. 14, 31970. doi:10.1038/s41598-024-83542-w
Freyer, F., Aquino, K., Robinson, P. A., Ritter, P., and Breakspear, M. (2009). Bistability and non-Gaussian fluctuations in spontaneous cortical activity. J. Neurosci. 29, 8512–8524. doi:10.1523/JNEUROSCI.0754-09.2009
Freyer, F., Roberts, J. A., Becker, R., Robinson, P. A., Ritter, P., and Breakspear, M. (2011). Biophysical mechanisms of multistability in resting-state cortical rhythms. J. Neurosci. 31, 6353–6361. doi:10.1523/JNEUROSCI.6693-10.2011
Hashemi, M., Depannemaecker, D., Saggio, M., Triebkorn, P., Rabuffo, G., Fousek, J., et al. (2025). Principles and operation of virtual brain twins. IEEE Rev. Biomed. Eng. PP, 1–29. doi:10.1109/RBME.2025.3562951
Illoul, L., and Lorong, P. (2011). On some aspects of the cnem implementation in 3d in order to simulate high speed machining or shearing. Comput. & Struct. 89, 940–958. doi:10.1016/j.compstruc.2011.01.018
Jirsa, V., Wang, H., Triebkorn, P., Hashemi, M., Jha, J., Gonzalez-Martinez, J., et al. (2023). Personalised virtual brain models in epilepsy. Lancet Neurology 22, 443–454. doi:10.1016/S1474-4422(23)00008-X
Kelso, J. S. (2012). Multistability and metastability: understanding dynamic coordination in the brain. Philosophical Trans. R. Soc. B Biol. Sci. 367, 906–918. doi:10.1098/rstb.2011.0351
Kilpatrick, Z. P. (2013). Wilson-cowan model. New York, NY: Springer, 1–5. doi:10.1007/978-1-4614-7320-6_80-1
Ladenbauer, J., Khakimova, L., Malinowski, R., Obst, D., Tönnies, E., Antonenko, D., et al. (2023). Towards optimization of oscillatory stimulation during sleep. Neuromodulation Technol. A. T. Neural Interface 26, 1592–1601. doi:10.1016/j.neurom.2022.05.006
Ladenbauer, J., Ladenbauer, J., Külzow, N., de Boor, R., Avramova, E., Grittner, U., et al. (2017). Promoting sleep oscillations and their functional coupling by transcranial stimulation enhances memory consolidation in mild cognitive impairment. J. Neurosci. 37, 7111–7124. doi:10.1523/JNEUROSCI.0260-17.2017
Levenstein, D., Buzsáki, G., and Rinzel, J. (2019). Nrem sleep in the rodent neocortex and hippocampus reflects excitable dynamics. Nat. Commun. 10, 2478. doi:10.1038/s41467-019-10327-5
Liang, Y., Liang, J., Song, C., Liu, M., Knöpfel, T., Gong, P., et al. (2023). Complexity of cortical wave patterns of the wake mouse cortex. Nat. Commun. 14, 1434. doi:10.1038/s41467-023-37088-6
Massimini, M., Huber, R., Ferrarelli, F., Hill, S., and Tononi, G. (2004). The sleep slow oscillation as a traveling wave. J. Neurosci. 24, 6862–6870. doi:10.1523/JNEUROSCI.1318-04.2004
Mohan, U. R., Zhang, H., Ermentrout, B., and Jacobs, J. (2024). The direction of theta and alpha travelling waves modulates human memory processing. Nat. Hum. Behav. 8, 1124–1135. doi:10.1038/s41562-024-01838-3
Muller, L., Piantoni, G., Koller, D., Cash, S. S., Halgren, E., and Sejnowski, T. J. (2016). Rotating waves during human sleep spindles organize global patterns of activity that repeat precisely through the night. eLife 5, e17267. doi:10.7554/eLife.17267
Muller, L., Reynaud, A., Chavane, F., and Destexhe, A. (2014). The stimulus-evoked population response in visual cortex of awake monkey is a propagating wave. Nat. Commun. 5, 3675. doi:10.1038/ncomms4675
Papadopoulos, L., Lynn, C. W., Battaglia, D., and Bassett, D. S. (2020). Relations between large-scale brain connectivity and effects of regional stimulation depend on collective dynamical state. PLOS Comput. Biol. 16, e1008144–43. doi:10.1371/journal.pcbi.1008144
Pessoa, L. (2022). The entangled brain: how perception, cognition, and emotion are woven together. MIT Press.
Petkoski, S., and Jirsa, V. K. (2022). Normalizing the brain connectome for communication through synchronization. Netw. Neurosci. 6, 722–744. doi:10.1162/netn_a_00231
Pinto, D. J., Brumberg, J. C., Simons, D. J., Ermentrout, G. B., and Traub, R. (1996). A quantitative population model of whisker barrels: Re-examining the wilson-cowan equations. J. Comput. Neurosci. 3, 247–264. doi:10.1007/BF00161134
Popovych, O. V., Jung, K., Manos, T., Diaz-Pier, S., Hoffstaedter, F., Schreiber, J., et al. (2021). Inter-subject and inter-parcellation variability of resting-state whole-brain dynamical modeling. NeuroImage 236, 118201. doi:10.1016/j.neuroimage.2021.118201
Proix, T., Spiegler, A., Schirner, M., Rothmeier, S., Ritter, P., and Jirsa, V. K. (2016). How do parcellation size and short-range connectivity affect dynamics in large-scale brain network models? NeuroImage 142, 135–149. doi:10.1016/j.neuroimage.2016.06.016
Rasch, B., and Born, J. (2013). About sleep’s role in memory. Physiol. Rev. 93, 681–766. doi:10.1152/physrev.00032.2012
Richardson, M. J. E. (2007). Firing-rate response of linear and nonlinear integrate-and-fire neurons to modulated current-based and conductance-based synaptic drive. Phys. Rev. E - Stat. nonlinear, soft matter Phys. 76 (2 Pt 1), 021919. doi:10.1103/PhysRevE.76.021919
Roberts, J. A., Gollo, L. L., Abeysuriya, R. G., Roberts, G., Mitchell, P. B., Woolrich, M. W., et al. (2019). Metastable brain waves. Nat. Commun. 10, 1056. doi:10.1038/s41467-019-08999-0
Sanchez-Vives, M. V., Massimini, M., and Mattia, M. (2017). Shaping the default activity pattern of the cortical network. Neuron 94, 993–1001. doi:10.1016/j.neuron.2017.05.015
Schaefer, A., Kong, R., Gordon, E. M., Laumann, T. O., Zuo, X.-N., Holmes, A. J., et al. (2018). Local-global parcellation of the human cerebral cortex from intrinsic functional connectivity mri. Cereb. Cortex 28, 3095–3114. doi:10.1093/cercor/bhx179
Sporns, O. (2022). The complex brain: connectivity, dynamics, information. Trends Cognitive Sci. 26, 1066–1067. doi:10.1016/j.tics.2022.08.002
Torao-Angosto, M., Manasanch, A., Mattia, M., and Sanchez-Vives, M. V. (2021). Up and down states during slow oscillations in slow-wave sleep and different levels of anesthesia. Front. Syst. Neurosci. 15, 609645. doi:10.3389/fnsys.2021.609645
Townsend, R. G., and Gong, P. (2018). Detection and analysis of spatiotemporal patterns in brain activity. PLoS Comput. Biol. 14, e1006643. doi:10.1371/journal.pcbi.1006643
Wang, H. E., Triebkorn, P., Breyton, M., Dollomaja, B., Lemarechal, J.-D., Petkoski, S., et al. (2024). Virtual brain twins: from basic neuroscience to clinical use. Natl. Sci. Rev. 11, nwae079. doi:10.1093/nsr/nwae079
Wilson, H. R., and Cowan, J. D. (1972). Excitatory and inhibitory interactions in localized populations of model neurons. Biophysical J. 12, 1–24. doi:10.1016/S0006-3495(72)86068-5
Keywords: whole-brain modeling, network resolution, neural mass modeling, spatiotemporal dynamics, slow oscillations, network physiology
Citation: Dimulescu C, Strömsdörfer R, Flöel A and Obermayer K (2025) On the robustness of the emergent spatiotemporal dynamics in biophysically realistic and phenomenological whole-brain models at multiple network resolutions. Front. Netw. Physiol. 5:1589566. doi: 10.3389/fnetp.2025.1589566
Received: 07 March 2025; Accepted: 19 June 2025;
Published: 08 August 2025.
Edited by:
Viktor Jirsa, Aix-Marseille Université, FranceReviewed by:
Spase Petkoski, Aix Marseille Université, FranceYusuke Takeda, Advanced Telecommunications Research Institute International (ATR), Japan
Copyright © 2025 Dimulescu, Strömsdörfer, Flöel and Obermayer. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Ronja Strömsdörfer, c3Ryb2Vtc2RvZXJmZXJAdHUtYmVybGluLmRl
†These authors share first authorship
‡ORCID: Agnes Flöel, orcid.org/0000-0002-1475-5872