 Roman Kononov
Roman Kononov Vasilii Tiselko
Vasilii Tiselko  Oleg Maslennikov
Oleg Maslennikov  Vladimir Nekorkin
Vladimir Nekorkin - 1Nonlinear Dynamics Department, Gaponov-Grekhov Institute of Applied Physics of the Russian Academy of Sciences, Nizhny Novgorod, Russia
- 2Faculty of Radiophysics, Lobachevsky State University of Nizhny Novgorod, Nizhny Novgorod, Russia
- 3Center for Neurophysics and Neuromorphic Technologies, Moscow, Russia
- 4Phystech School of Applied Mathematics and Computer Science, Moscow Institute of Physics and Technology, Dolgoprudny, Moscow Region, Russia
Representing and integrating continuous variables is a fundamental capability of the brain, often relying on ring attractor circuits that maintain a persistent bump of activity. To investigate how such structures can self-organize, we trained a recurrent neural network (RNN) on a ring-based path integration task using population-coded velocity inputs. The network autonomously developed a modular architecture: one subpopulation formed a stable ring attractor to maintain the integrated position, while a second, distinct subpopulation organized into a dissipative control unit that translates velocity into directional signals. Furthermore, systematic perturbations revealed that the precise topological alignment between these modules is essential for reliable integration. Our findings illustrate how functional specialization and biologically plausible representations can emerge from a general learning objective, offering insights into neural self-organization and providing a framework for designing more interpretable and robust neuromorphic systems for navigation and control.
1 Introduction
A central challenge in physiology is to uncover how complex neural and physiological systems achieve robust, flexible information processing through the structured interaction of distributed components—a phenomenon deeply rooted in the principles of self-organization. In recent years, the rapidly growing field of network physiology has emphasized understanding the coordinated dynamics and functional connectivity within and across distinct subsystems, with the goal of elucidating mechanisms underlying adaptive behavior, resilience, and nonequilibrium phase transitions in living systems (Bartsch et al., 2015; Ivanov and Bartsch, 2014). Neural networks, in particular, serve as canonical models of such emergent dynamics, in which collective behaviors—ranging from oscillations to discrete or continuous attractor states—arise from recurring patterns of connectivity and population-level coding (Vladimirovich Maslennikov et al., 2022).
Within this framework, population coding and attractor dynamics have been recognized as fundamental organizing principles that underpin neural computation (Haken, 1983). Paradigmatic examples include bump and ring attractor networks, which enable the representation and integration of continuous variables. These networks are not confined to a single species or brain region but represent a species-agnostic neural motif for computation, found in contexts ranging from spatial orientation in mammals (Zhang, , 1996; Moser et al., 2008) to the internal compass of insects like the fruit fly (Kim et al., 2017), birds (Ben-Yishay et al., 2021), and even fish (Vinepinsky et al., 2020). This evolutionary convergence implies that by studying these motifs, we can gain insight into the general physiological principles of brain function across diverse species (Khona and Fiete, 2022; Basu et al., 2024). These distributed representations also exemplify physiological robustness, permitting reliable encoding in the presence of noise and fluctuating inputs (Averbeck et al., 2006; Pouget et al., 2000). In the hippocampal-entorhinal circuit, for instance, place and grid cells collectively generate a dynamic map of the environment (Hafting et al., 2005; McNaughton et al., 2006). Theoretically, such 2D spatial representations can be constructed by combining multiple 1D ring attractors, each encoding orientation along a different axis, highlighting the role of the ring attractor as a fundamental computational building block (Bush and Burgess, 2014).
Crucially, network-based models such as ring attractors not only capture these population-level coding schemes, but also provide a theoretical framework—rooted in the self-organization of collective variables—for understanding how internal states can be flexibly updated, maintained, and read out by downstream systems (Skaggs et al., 1994; Fiete et al., 2008; Seeholzer et al., 2019). In head direction systems and cortical integration circuits, the spontaneous emergence of ring attractors as solutions to path integration and spatial memory tasks exemplifies how nonequilibrium transitions and bifurcations in network structure give rise to functionally specialized modules (Georgopoulos et al., 1986; Salinas and Abbott, 1994; Ganguli and Sompolinsky, 2012; Knierim et al., 1995; Heinze et al., 2018). Such processes underscore a general principle, highlighted in the Synergetics tradition: physiological networks can self-organize connectivity patterns and explicit coding strategies to achieve both specialization and adaptive coordination among functional subunits.
Despite considerable progress, a central open question in both physiology and artificial intelligence remains: How can such modular, interpretable architectures—capable of robust continuous integration and flexibly dealing with circular variables—arise autonomously through learning mechanisms? Here, we use the term “autonomously” to describe the emergence of structured connectivity and dynamics driven by a high-level functional objective, rather than through handcrafted design. And how do the emergent patterns of organization constrain or enhance physiological function? Building bridges between biological plausibility and artificial network design is thus essential for advancing our theoretical understanding and for informing translational neuromorphic engineering (Banino et al., 2018; Izzo et al., 2023; Ganguli and Sompolinsky, 2012).
Motivated by these themes, and inspired by the pioneering insights of Hermann Haken, we here examine how a recurrent neural network trained with explicit population coding can self-organize into functional, physiologically congruent subpopulations that support robust, interpretable continuous variable integration. By dissecting the network’s emergent architecture, dynamical coding strategies, and the response to systematic perturbations, we aim to illuminate general organizing principles that underlie both neural computation and the broader dynamics of self-organized physiological networks.
2 Materials and methods
We address the problem of continuous navigation on a ring—a canonical task in theoretical and systems neuroscience—where an agent receives, at each timestep
where
To mimic the manner in which biological circuits represent continuous variables, we employ population coding for the input: the scalar velocity
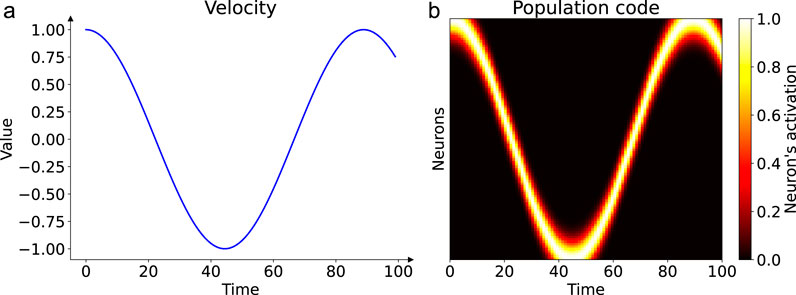
Figure 1. Population coding of a dynamic input signal. Panel (a) shows an example of a time-varying velocity signal, here one period of a sinusoid, presented to the network over time. Panel (b) illustrates how this scalar profile is encoded as a spatiotemporal activity pattern across the input neuron population. Each neuron (y-axis) is tuned to a preferred velocity. As the input velocity changes over time (x-axis), the peak of the activity bump shifts across the population, creating a dynamic representation of the signal shown in (a).
Such population codes enable robust integration and flexible transformation of noisy or ambiguous sensory signals, paralleling mechanisms observed in the brain’s sensory and motor systems. Notably, our input population encodes only velocity, not angular position, which differs from some biological head-direction systems where conjunctive coding is observed (Yoder et al., 2015). This choice allows us to specifically investigate how a network can autonomously learn to transform a pure velocity signal into a stable angular representation through its recurrent dynamics.
The target coordinate at time
In our discrete-time simulation, this integral is approximated using a second-order Euler method. During training, this serves as the supervisory signal. The “target neuron” refers to the output neuron whose preferred position
We arrange the neurons of the input and output populations to uniformly tile the relevant ranges,
The overall task and network architecture, which consists of functionally distinct input and output populations coupled through a recurrent weight matrix, is schematized in Figure 2. The artificial recurrent neural network we trained evolves according to:
where
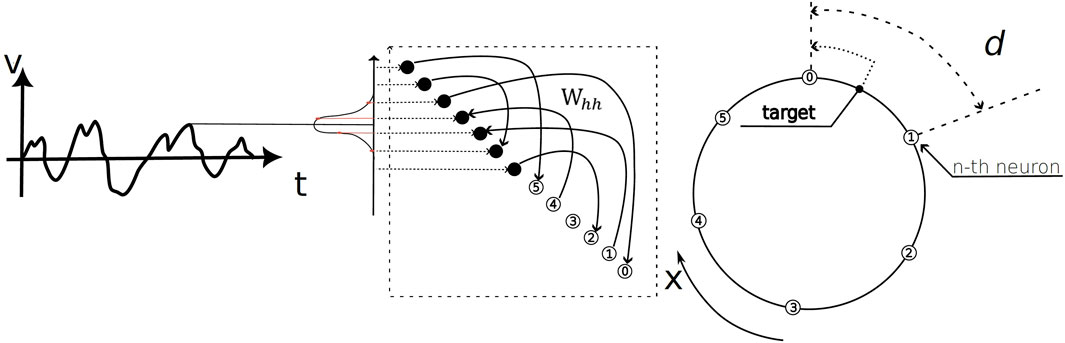
Figure 2. Schematic of the ring navigation task and network architecture. The model performs path integration by processing a time-varying velocity profile,
Our loss function is designed to reflect the graded, population-level readout present in biological codes. The neuron nearest to the target coordinate is treated as the “class” with the correct label, with adjacent neurons given smaller weights:
with
and
where
To prevent units in the output population from becoming permanently silent (a common issue in ReLU networks), we implemented a simple homeostatic mechanism. In addition to adding a small, loss-adaptive noise to the gradients during each training step, we also periodically reinitialized inactive neurons. At the end of several batches, any neuron whose activity remained below a small threshold
3 Results
We designed our network with a pre-defined modular structure to investigate functional specialization. The total population of N = 800 neurons was partitioned into two equally sized groups: the first 400 neurons were designated as the “output population” and received no direct external input, while the remaining 400 neurons were the “input population” and received the external velocity signal. Both populations were uniformly tiled over their respective domains, creating a structured basis for analysis of functional specialization and connectivity.
To dissect the computational dynamics, we express the network’s full state at time
with
corresponding to intra-population (diagonal blocks
The evolution of each sub-population is governed by:
which capture the continuous integration, stabilization, and transformation of sensory input in a modular, population-based format.
A successfully trained network exhibits clear functional specialization, as shown in Figure 3. While the input population activity directly mirrors the incoming velocity signal (Figure 3a), the output population integrates this signal to maintain a stable, localized bump of activity representing the agent’s angular position (Figure 3b). This functional division is a direct result of the self-organized recurrent connectivity matrix (Figure 3c), where the output-to-output block
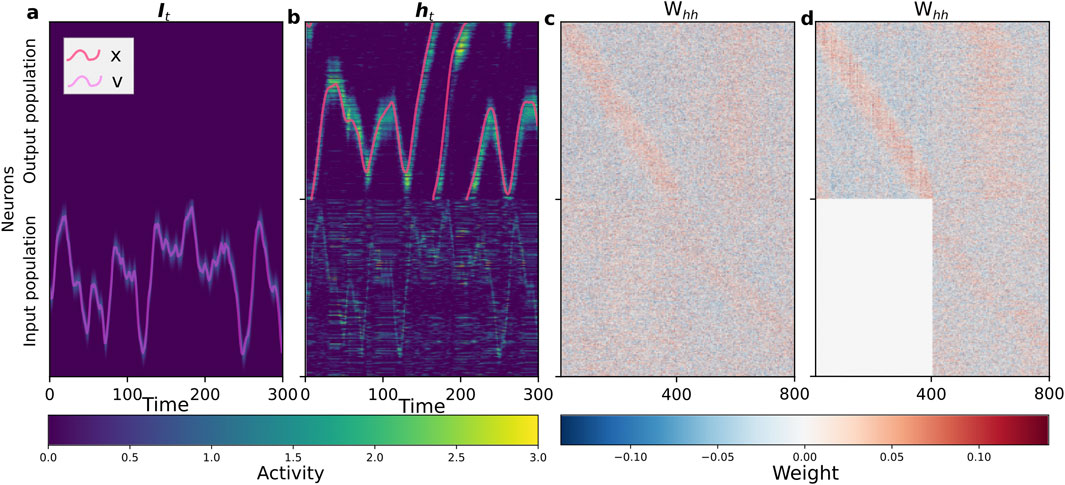
Figure 3. Self-organized network structure and dynamics during path integration. The network consists of an input population (neurons 1–400) and an output population (neurons 401–800). (a) Spatiotemporal activity of the population-coded input signal
To further probe the specialization of network modules, we simulated the autonomous activity of the input and output populations in the absence of inter-population coupling. Here, each population was initialized with Gaussian bumps at multiple, distinct positions. This setup allows us to differentiate persistent from transient attractor dynamics. In this isolated condition, the activity of the input
The results of these autonomous simulations, shown in Figure 4, confirm the functional roles of the two modules. The output population sustains the initial activity bumps for an extended period (Figure 4a), demonstrating a memory-like capability. By contrast, the input population shows rapid decay (Figure 4b), consistent with its role as a transient encoder. We note that the specific network instance visualized here shows some heterogeneity in its response, with stronger activity in one region of the ring. This is a feature of this particular trained network, as the effect varies across different training runs and is not a systematic bias of the model. The central finding demonstrated here—the clear functional contrast between the persistent output and dissipative input populations—is a robust result observed consistently across all successful networks.
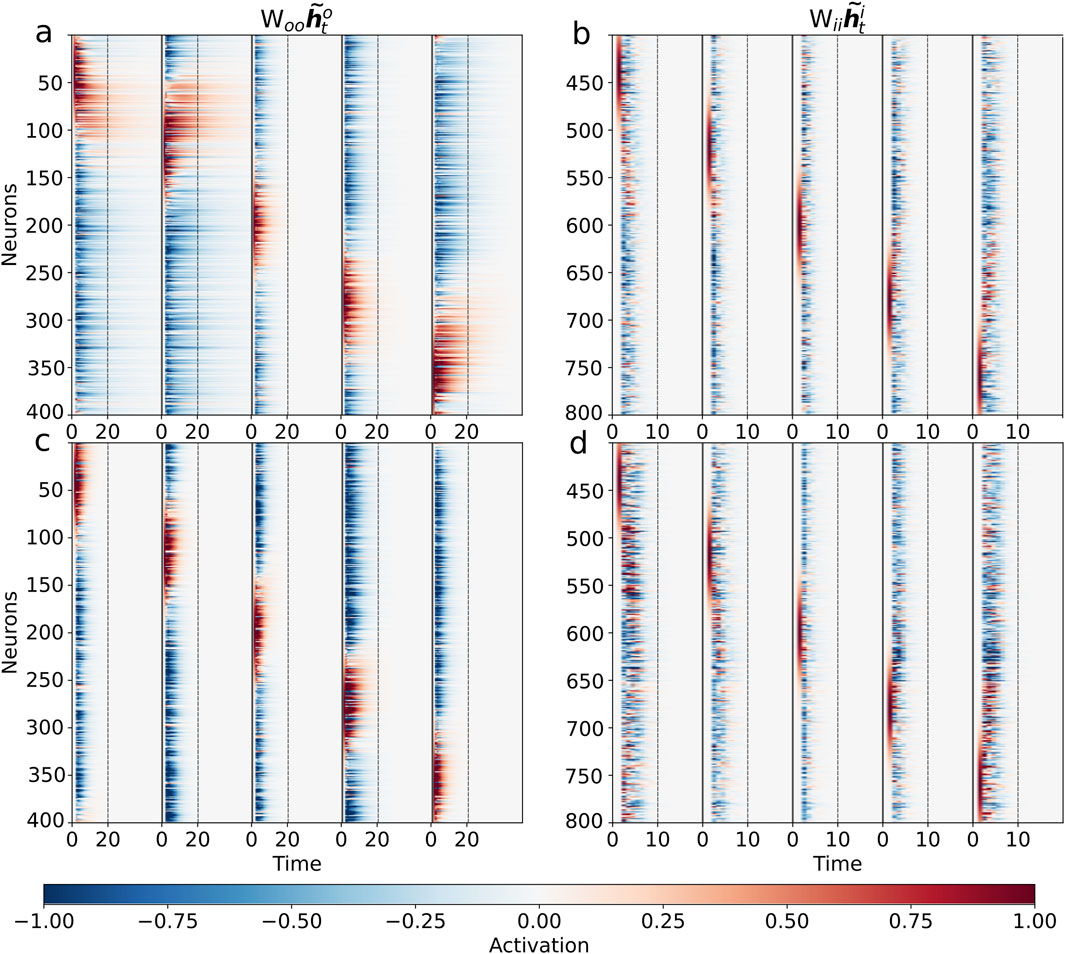
Figure 4. Autonomous dynamics reveal functional specialization of input and output populations. To test the intrinsic properties of the learned modules, we simulated their autonomous activity driven only by their internal recurrent connections (i.e., without external input or inter-population signals). Each population was initialized with five distinct Gaussian bumps of activity. (a,c) The output populations exhibit transiently persistent activity, maintaining the bumps for an extended period before eventually decaying to the zero state. This demonstrates a clear memory-like capability, essential for integration. Notably, the persistence in the fully connected network (a) is significantly more robust, sustaining the localized activity much longer than in the feedforward-only network (c). (b,d) In sharp contrast, the input populations from both the fully connected network (b) and the feedforward-only network (d) show dissipative dynamics, where the initial activity bumps decay rapidly. These simulations correspond to evolving the activity according to (Pouget et al., 2000; Hafting et al., 2005).
To investigate the role of modular connectivity, we selectively ablated the feedback pathway
The underlying reason for this instability is revealed by comparing the autonomous dynamics of the two networks in Figure 4. The output module from the network trained without feedback is intrinsically less stable, showing a much faster decay of activity (Figure 4c) compared to its counterpart from the fully connected network (Figure 4a). This highlights that the network learns to sustain activity for a prolonged but finite duration, a property we term transient persistence. This suggests that in the full network, the feedback pathway allows the input population to participate in a larger recurrent circuit that actively stabilizes the activity bump on the output ring. Without this top-down connection, the entire burden of maintaining persistent activity falls solely upon the internal recurrence of the output population
We then explored the mapping and transfer of control signals by delivering a velocity input that increased linearly from
As illustrated in Figure 5b, a properly trained network successfully integrates the velocity ramp (Figure 5a). However, network performance collapses when the population code-to-ring mapping is disrupted by permuting the output neurons (Figure 5c), confirming that the encoded structure and network topology must remain aligned for successful integration. The output population in the unperturbed circuit receives a smoothly propagating population control signal, but this signal can no longer synchronize correctly with a shuffled arrangement, revealing the codependency of circuitry and coding motif.
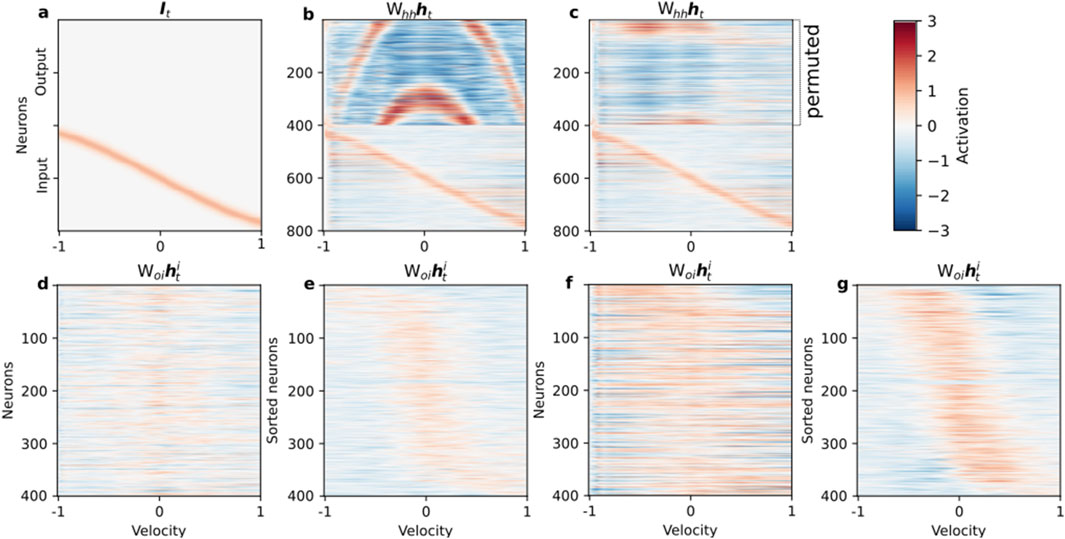
Figure 5. Topological alignment of control signals is critical for integration. We probed the network’s integration mechanism using a linearly increasing velocity input. (a) The input velocity ramp from −1 to 1 over the trial. (b) The activity of the fully trained network
Further analysis of the feedforward control signal
Taken together, these analyses show that population-coded recurrent networks can naturally self-organize into specialized modules for fast encoding and persistent memory. Faithful function relies not only on the learned synaptic weights, but also on the precise and consistent internal mapping between neural populations and their target representations. These features are hallmarks of modular, robust physiological computation as observed in biological navigation and memory systems.
4 Discussion and conclusion
In this study, we demonstrated that a recurrent neural network trained on a continuous integration task can autonomously self-organize into a modular architecture with functionally distinct and physiologically congruent subpopulations. Here, self-organization refers to the emergence of structured connectivity as a result of a supervised learning process, rather than arising from unsupervised, local update rules in the physical sense. Our findings contribute to bridging the gap between the dynamics of artificial neural networks and the principles of neural computation observed in biological systems.
Our central finding is the emergent division of labor within the network. The output subpopulation develops a ring attractor, a canonical structure for encoding circular variables like head direction and spatial orientation (Zhang, , 1996; Kim et al., 2017). This structure supports persistent, localized activity, enabling it to function as a robust memory module for the integrated position. Concurrently, the input subpopulation forms a dissipative, segment-like architecture that acts as a transient control unit, transforming velocity signals into directional commands that drive the movement of the activity bump on the ring. This modular separation—separating memory from control—is a key organizational principle in the brain, allowing for flexible and robust computation (Salinas and Abbott, 1994; Ganguli and Sompolinsky, 2012). Unlike models where ring connectivity is hardwired, here it emerges solely from the learning objective, suggesting that attractor dynamics are a natural and efficient solution for continuous variable integration.
A key distinction of our work, however, is the demonstration of the critical importance of topological alignment between these emergent modules. As shown in our permutation experiment (Figures 5a–c), the network’s function is not merely a product of its component parts but depends fundamentally on the learned, ordered mapping between the control signals from the input population and the spatial layout of the output ring attractor. This highlights that for distributed neural codes to be computationally effective, the wiring” must respect the coding”. Disruptions to this alignment, analogous to developmental disorders or brain injury, can lead to a catastrophic failure of function, even if the individual modules remain intact. Our perturbation analyses thus underscore the role of feedback and precise inter-module connectivity, echoing experimental findings where disrupting specific pathways compromises memory and integration (Seeholzer et al., 2019; McNaughton et al., 2006; Bonnevie et al., 2013).
Our results also inform the broader field of network physiology by providing a concrete computational example of how specialized subsystems can arise and coordinate within a larger, interconnected system. The balance between the persistent dynamics of the memory module and the dissipative dynamics of the control module illustrates how networks can achieve both stability and adaptability. This emergent coordination within a complex neural network serves as a key example of physiological resilience and the principles of system-level self-organization that are central to the study of network physiology (Bartsch et al., 2015; Ivanov and Bartsch, 2014; Ivanov, 2021).
4.1 Limitations and future directions
We acknowledge several limitations that open avenues for future research. First, our model employs non-spiking neurons and a supervised, gradient-based learning rule. While this framework is computationally powerful, future work should explore how these functional architectures could emerge using more biologically plausible spiking neurons and local, Hebbian-like learning rules (Song and Wang, 2005; Levenstein et al., 2024; Pugavko et al., 2023; Maslennikov et al., 2024). Second, the supervisory signal, while justifiable as an abstraction, could be replaced with reinforcement learning or unsupervised learning objectives to better model autonomous discovery in biological agents (Banino et al., 2018; Ivanov et al., 2025). Furthermore, training the network on more complex and physiologically grounded velocity profiles, such as those derived from animal tracking data (Sargolini et al., 2006), could reveal how network solutions are shaped by naturalistic input statistics. Exploring the network’s resilience to transient perturbations, such as temporary loss of connectivity between modules (Cooper and Mizumori, 2001), would also provide deeper insights into the robustness of these self-organized circuits.
On a translational level, our work illustrates how interpretable, modular architectures can be learned rather than handcrafted, offering a path toward more explainable AI and robust autonomous systems. This is particularly relevant for neuromorphic engineering and robotics, where many existing applications of ring attractors rely on hand-crafted weights (Rivero-Ortega et al., 2023). Our approach, where functional weights are learned, offers a promising route to developing more adaptive and flexible controllers. For mobile and field robotics, key considerations include not only robustness and interpretability but also low power consumption, a primary goal of neuromorphic systems (Izzo et al., 2023; Robin et al., 2022). Furthermore, the ring attractor motif is not limited to 1D orientation but serves as a foundational component for more complex spatial representations, such as modeling the 2D planar motion of a robot, linking back to the principles of grid cell computation (Knowles et al., 2023). The increasing availability of specialized neuromorphic hardware, such as Intel’s Loihi processors (Davies et al., 2018), and associated software frameworks like LAVA (Lava Framework Authors, 2022), makes these brain-inspired models increasingly viable for real-world, embedded applications where online learning and energy efficiency are paramount.
4.2 Conclusion
In summary, we have shown that explicit population coding guides a recurrent network to self-organize into a modular system comprising a ring attractor for memory and a dissipative controller for input processing. This emergent structure, highly reminiscent of biological circuits for navigation, depends critically on the precise topological alignment between its functional modules. Our findings underscore how general learning principles can give rise to specialized, interpretable, and physiologically plausible neural computations, advancing our understanding of both natural and artificial intelligence.
Data availability statement
The original contributions presented in the study are included in the article/supplementary material, further inquiries can be directed to the corresponding author.
Author contributions
RK: Formal Analysis, Investigation, Software, Visualization, Writing – original draft, Writing – review and editing. VT: Conceptualization, Methodology, Writing – review and editing, Investigation. OM: Conceptualization, Data curation, Funding acquisition, Investigation, Methodology, Project administration, Supervision, Writing – original draft, Writing – review and editing. VN: Conceptualization, Methodology, Project administration, Supervision, Writing – review and editing.
Funding
The author(s) declare that financial support was received for the research and/or publication of this article. The work was supported by the Russian Science Foundation, grant No 23-72-10088.
Conflict of interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Generative AI statement
The author(s) declare that no Generative AI was used in the creation of this manuscript.
Any alternative text (alt text) provided alongside figures in this article has been generated by Frontiers with the support of artificial intelligence and reasonable efforts have been made to ensure accuracy, including review by the authors wherever possible. If you identify any issues, please contact us.
Publisher’s note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
Footnotes
1The code used to generate the results in this study is available at https://github.com/p0mik13/PopulCoding_RingNavigationTask_F.
References
Averbeck, B. B., Latham, P. E., and Pouget, A. (2006). Neural correlations, population coding and computation. Nat. Rev. Neurosci. 7 (5), 358–366. doi:10.1038/nrn1888
Banino, A., Barry, C., Uria, B., Blundell, C., Lillicrap, T., Mirowski, P., et al. (2018). Vector-based navigation using grid-like representations in artificial agents. Nature 557 (7705), 429–433. doi:10.1038/s41586-018-0102-6
Bartsch, R. P., Liu, K. K. L., Bashan, A., and Ivanov, P.Ch (2015). Network physiology: how organ systems dynamically interact. PloS One 10 (11), e0142143. doi:10.1371/journal.pone.0142143
Basu, J., and Nagel, K. (2024). Neural circuits for goal-directed navigation across species. Trends Neurosci. 47 (11), 904–917. doi:10.1016/j.tins.2024.09.005
Ben-Yishay, E., Krivoruchko, K., Ron, S., Ulanovsky, N., Derdikman, D., and Gutfreund, Y. (2021). Directional tuning in the hippocampal formation of birds. Curr. Biol. 31 (12), 2592–2602.e4. doi:10.1016/j.cub.2021.04.029
Bonnevie, T., Dunn, B., Fyhn, M., Hafting, T., Derdikman, D., Kubie, J. L., et al. (2013). Grid cells require excitatory drive from the hippocampus. Nat. Neurosci. 16 (3), 309–317. doi:10.1038/nn.3311
Bush, D., and Burgess, N. (2014). A hybrid oscillatory interference/continuous attractor network model of grid cell firing. J. Neurosci. 34 (14), 5065–5079. doi:10.1523/JNEUROSCI.4017-13.2014
Cooper, B. G., and Mizumori, S. J. Y. (2001). Temporary inactivation of the retrosplenial cortex causes a transient reorganization of spatial coding in the hippocampus. J. Neurosci. 21 (11), 3986–4001. doi:10.1523/JNEUROSCI.21-11-03986.2001
Cueva, C. J., and Wei, X.-X. (2018). Emergence of grid-like representations by training recurrent neural networks to perform spatial localization. arXiv Prepr. arXiv:1803.07770. 10.48550/arXiv.1803.07770
Davies, M., Srinivasa, N., Lin, T.-H., Chinya, G., Cao, Y., Choday, S. H., et al. (2018). Loihi: a neuromorphic manycore processor with on-chip learning. Ieee Micro 38 (1), 82–99. doi:10.1109/mm.2018.112130359
Fiete, I. R., Burak, Y., and Brookings, T. (2008). What grid cells convey about rat location. J. Neurosci. 28 (27), 6858–6871. doi:10.1523/JNEUROSCI.5684-07.2008
Ganguli, S., and Sompolinsky, H. (2012). Compressed sensing, sparsity, and dimensionality in neuronal information processing and data analysis. Annu. Rev. Neurosci. 35 (1), 485–508. doi:10.1146/annurev-neuro-062111-150410
Georgopoulos, A. P., Schwartz, A. B., and Kettner, R. E. (1986). Neuronal population coding of movement direction. Science 233 (4771), 1416–1419. doi:10.1126/science.3749885
Hafting, T., Fyhn, M., Molden, S., Moser, M.-B., and Moser, E. I. (2005). Microstructure of a spatial map in the entorhinal cortex. Nature 436 (7052), 801–806. doi:10.1038/nature03721
Heinze, S., Narendra, A., and Cheung, A. (2018). Principles of insect path integration. Curr. Biol. 28 (17), R1043–R1058. doi:10.1016/j.cub.2018.04.058
Ivanov, P.Ch. (2021). The new field of network physiology: building the human physiolome. Front. Netw. physiology 1, 711778. doi:10.3389/fnetp.2021.711778
Ivanov, P.Ch, and Bartsch, R. P. (2014). “Network physiology: mapping interactions between networks of physiologic networks,” in Networks of networks: the last frontier of complexity (Springer), 203–222.
Ivanov, D. A., Larionov, D. A., Maslennikov, O. V., and Voevodin, V. V. (2025). Neural network compression for reinforcement learning tasks. Sci. Rep. 15 (1), 9718. doi:10.1038/s41598-025-93955-w
Izzo, D., Hadjiivanov, A., Dold, D., Meoni, G., and Blazquez, E. (2023). “Neuromorphic computing and sensing in space,” in Artificial intelligence for space: AI4SPACE (Boca Raton, FL: CRC Press), 107–159. doi:10.48550/arXiv.2212.05236
Khona, M., and Fiete, I. R. (2022). Attractor and integrator networks in the brain. Nat. Rev. Neurosci. 23 (12), 744–766. doi:10.1038/s41583-022-00642-0
Kim, S. S., Rouault, H., Druckmann, S., and Jayaraman, V. (2017). Ring attractor dynamics in the drosophila central brain. Science 356 (6340), 849–853. doi:10.1126/science.aal4835
Knierim, J. J., Kudrimoti, H. S., and McNaughton, B. L. (1995). Place cells, head direction cells, and the learning of landmark stability. J. Neurosci. 15 (3), 1648–1659. doi:10.1523/JNEUROSCI.15-03-01648.1995
Knowles, T. C., Summerton, A. G., Whiting, J. G. H., and Pearson, M. J. (2023). Ring attractors as the basis of a biomimetic navigation system. Biomimetics 8 (5), 399. doi:10.3390/biomimetics8050399
Kononov, R. A., Maslennikov, O. V., and Nekorkin, V. I. (2025). Dynamics of recurrent neural networks with piecewise linear activation function in the context-dependent decision-making task. Izv. VUZ. Appl. Nonlinear Dyn. 33 (2), 249–265. doi:10.18500/0869-6632-003147
Levenstein, D., Efremov, A., Eyono, R. H., Peyrache, A., and Richards, B. (2024). Sequential predictive learning is a unifying theory for hippocampal representation and replay. bioRxiv, 2024–04. doi:10.1101/2024.04.28.591528
Maslennikov, O., Perc, M., and Nekorkin, V. (2024). Topological features of spike trains in recurrent spiking neural networks that are trained to generate spatiotemporal patterns. Front. Comput. Neurosci. 18, 1363514. doi:10.3389/fncom.2024.1363514
Maslennikov, O. V., Pugavko, M. M., Shchapin, D. S., and Nekorkin, V. I. (2022). Nonlinear dynamics and machine learning of recurrent spiking neural networks. Physics-Uspekhi 65 (10), 1020–1038. doi:10.3367/ufne.2021.08.039042
McNaughton, B. L., Battaglia, F. P., Jensen, O., Moser, E. I., and Moser, M.-B. (2006). Path integration and the neural basis of the’cognitive map. Nat. Rev. Neurosci. 7 (8), 663–678. doi:10.1038/nrn1932
Moser, E. I., Kropff, E., and Moser, M. B. (2008). Place cells, grid cells, and the brain's spatial representation system. Annu. Rev. Neurosci. 31, 69–89. doi:10.1146/annurev.neuro.31.061307.090723
Pouget, A., Dayan, P., and Zemel, R. S. (2000). Information processing with population codes. Nat. Rev. Neurosci. 1 (2), 125–132. doi:10.1038/35039062
Pugavko, M. M., Maslennikov, O. V., and Nekorkin, V. I. (2023). Multitask computation through dynamics in recurrent spiking neural networks. Sci. Rep. 13 (1), 3997. doi:10.1038/s41598-023-31110-z
Rivero-Ortega, J. D., Mosquera-Maturana, J. S., Pardo-Cabrera, J., Hurtado-López, J., Hernández, J. D., Romero-Cano, V., et al. (2023). Ring attractor bio-inspired neural network for social robot navigation. Front. Neurorobotics 17, 1211570. doi:10.3389/fnbot.2023.1211570
Robinson, B. S., Norman-Tenazas, R., Cervantes, M., Symonette, D., Johnson, E. C., Joyce, J., et al. (2022). Online learning for orientation estimation during translation in an insect ring attractor network. Sci. Rep. 12 (1), 3210. doi:10.1038/s41598-022-05798-4
Salinas, E., and Abbott, L. F. (1994). Vector reconstruction from firing rates. J. Comput. Neurosci. 1 (1), 89–107. doi:10.1007/BF00962720
Sargolini, F., Fyhn, M., Hafting, T., McNaughton, B. L., Witter, M. P., Moser, M.-B., et al. (2006). Conjunctive representation of position, direction, and velocity in entorhinal cortex. Science 312 (5774), 758–762. doi:10.1126/science.1125572
Seeholzer, A., Moritz, D., and Gerstner, W. (2019). Stability of working memory in continuous attractor networks under the control of short-term plasticity. PLoS Comput. Biol. 15 (4), e1006928. doi:10.1371/journal.pcbi.1006928
Skaggs, W., Knierim, J., Kudrimoti, H., and McNaughton, B. (1994). A model of the neural basis of the rat’s sense of direction. Adv. Neural Inf. Process. Syst. 7. Available online at: https://pubmed.ncbi.nlm.nih.gov/11539168/.
Song, P., and Wang, X.-J. (2005). Angular path integration by moving “hill of activity”: a spiking neuron model without recurrent excitation of the head-direction system. J. Neurosci. 25 (4), 1002–1014. doi:10.1523/JNEUROSCI.4172-04.2005
Vinepinsky, E., Cohen, L., Perchik, S., Ben-Shahar, O., Donchin, O., and Segev, R. (2020). Representation of edges, head direction, and swimming kinematics in the brain of freely-navigating fish. Sci. Rep. 10 (1), 14762. doi:10.1038/s41598-020-71217-1
Yoder, R. M., Peck, J. R., and Taube, J. S. (2015). Visual landmark information gains control of the head direction signal at the lateral mammillary nuclei. J. Neurosci. 35 (4), 1354–1367. doi:10.1523/JNEUROSCI.1418-14.2015
Keywords: recurrent neural networks, bump attractors, population coding, continuous variable integration, nonlinear dynamics, network physiology, neural representation
Citation: Kononov R, Tiselko V, Maslennikov O and Nekorkin V (2025) Population coding and self-organized ring attractors in recurrent neural networks for continuous variable integration. Front. Netw. Physiol. 5:1693772. doi: 10.3389/fnetp.2025.1693772
Received: 27 August 2025; Accepted: 14 October 2025;
Published: 31 October 2025.
Edited by:
Eckehard Schöll, Technical University of Berlin, GermanyReviewed by:
Sajad Jafari, Amirkabir University of Technology, IranThomas Knowles, Bristol Robotics Laboratory, United Kingdom
Copyright © 2025 Kononov, Tiselko , Maslennikov and Nekorkin . This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Oleg Maslennikov , b2xtYW92QGlwZnJhbi5ydQ==