 Milán Szabó
Milán Szabó Zoltán Szabó1,2
Zoltán Szabó1,2 Krisztina Nagy
Krisztina Nagy Imre Vass
Imre Vass- 1Institute of Plant Biology, HUN-REN Biological Research Centre, Szeged, Hungary
- 2Department of Biotechnology and Microbiology, Faculty of Science and Informatics, University of Szeged, Szeged, Hungary
- 3Institute of Biophysics, HUN-REN Biological Research Centre, Szeged, Hungary
- 4Department of Experimental Physics, Institute of Physics, University of Szeged, Szeged, Hungary
The dinoflagellate algae Symbiodiniaceae live in endosymbiosis with Anthozoa, which is essential for the existence of coral reefs. The cells of Symbiodiniaceae exist in two distinct forms: the flagellated, motile form and the non-flagellated, coccoid form, which play specific roles in the life cycle of these algae. The regulation and distribution of the different cell forms have been intensively studied in the past, however, the behavior and changes of the cell cycle are less characterized under conditions that mimic the coral tissue versus the free living environment, e.g., by manipulating the viscosity of the medium and thereby altering cell motility. In this work, we applied precisely controlled microfluidic tools to manipulate the viscosity of the medium, using the polysucrose Ficoll. We found that by the application of Ficoll the diurnal cycle of the different cell forms undergoes remarkable changes, the motility of the cells decreases, and the motile phase of the cell cycle becomes significantly shorter compared to the absence of Ficoll. The slowed motile cells are also amenable for single-cell analysis of the activity of PSII (Fv/Fm). We therefore propose that the method developed here could serve as a sensitive monitoring system of the cell cycle changes and manipulation of cell motility mimicking the coral host environment, with concomitant single-cell photosynthetic activity analysis of Symbiodiniaceae.
1 Introduction
Symbiodinium is a genus of dinoflagellate algae (in the family of Symbiodiniaceae). These are unicellular microalgae, commonly found in endosymbiotic relationship with cnidarians such as corals, sea anemones, jellyfish. They perform the essential photosynthetic energy generation process and thus play a crucial role in supporting the whole symbiosis (Roth, 2014; Davy et al., 2012). Certain environmental stress conditions (e.g., heat and light) cause the expulsion of these zooxanthellae from the host, which is manifested in coral bleaching (Weis, 2008). Symbiodiniaceae has two main cell types: the motile flagellated cells (referred to as mastigotes or gymnodinioid forms), and the non-motile coccoid form. The motile cells have longitudinal and traversal flagella, which move the cells in helical or cyclic patterns (Freduenthal, 1962; Lee et al., 2015). It has been shown that the motility of Symbiodiniaceae cells exhibits a diel rhythm, and the motility typically starts before or at the beginning of the light period, and peaks within a couple of hours from the start of the light period (Fitt et al., 1981; Yacobovitch et al., 2004), although the diel pattern of motility varies in Symbiodiniaceae species (Yamashita and Koike, 2016). The appearance of motile cells is preceded by cell division, which thus typically achieves its maximum rate before the onset of light, and therefore, the cell division and increase in motile cell forms are correlated (Yamashita and Koike, 2016). The motility of Symbiodiniaceae is controlled by an endogenous rhythm, which determines the alternation between motile and non-motile states (Fitt et al., 1981). These different cell forms play an important role in making contact with the host and can be significantly modified in free-living and symbiotic lifestyles (Fitt et al., 1981). The host cells impose significant regulations on the growth, metabolism and cell cycle of the endosymbiont algae (Koike et al., 2004; Stat et al., 2006), and the algal symbiont exerts a diverse and complex diurnal regulation of their cellular environment, indicating that the symbiosis between the host and algae requires synchronization and coordination of several parallel processes, which are not completely understood (reviewed in Sorek et al., 2014).
The cell cycle dynamics of Symbiodiniaceae has been intensively studied in the past decades using various microscopic tools (Fitt et al., 1981; Fitt and Trench, 1983; Cui et al., 2022; Yamashita and Koike, 2016; Nitschke et al., 2015). However, in these studies repetitive manual sampling was needed, which may be suboptimal to capture fine changes in cell cycle, and it was not possible to modify or manipulate the growth medium or environment of the cells in a non-disruptive manner. High-throughput precisely controlled environment is therefore imperative to monitor cell morphology without potential disturbances or alteration of the ambient conditions. These can be achieved using microfluidic methods, in which a precisely controlled environment with real-time monitoring of physiological events (with a couple of seconds time resolution) is possible, even for a couple of days (Bashir et al., 2022; Patil et al., 2023). Microfluidics has gained increasing interest due to its advantages in controlled microenvironments. This technology offers custom design and solutions to manipulate fluids in microscopic dimensions, particularly tailored for single cells or cell colonies, with widespread applications for microbiology and microbial physiology (Nagy et al., 2018; Weibel et al., 2007). The greatest advantage of microfluidics is the capability of high precision geometric control along with liquid manipulation, which enables laminar liquid flow and thereby precise regulation of flow with minimal shear stress to the cells (Nagy et al., 2018). By this means, several ecologically relevant factors (nutrients, chemical signals, pH, etc.) can be regulated or valuable interactions between organisms can be revealed (Ábrahám et al., 2024; Greipel et al., 2024). Application of microfluidics has attracted considerable interest to study physiological processes of photosynthetic microalgae by the combination of single cell morphology with microscopic scale chlorophyll (Chl) fluorescence imaging (Széles et al., 2022; Széles et al., 2023; Patil et al., 2023). Single-cell Chl fluorescence imaging is a particularly useful tool to reveal photosynthetic light energy utilization and excess energy dissipation, providing therefore valuable insights about the phenotypic responses and heterogeneity of photosynthetic activity in a range of single-celled algae (Oxborough et al., 2000; Oxborough and Baker, 1997; Komárek et al., 2010), reviewed in (Schreiber, 2004; Baker, 2008). Investigation of the photobiology of Symbiodiniaceae has progressed significantly in the past years, and precisely controlled microfluidics-based experimental setups combined with single-cell chlorophyll fluorometry allowed the multiparametric monitoring of thermal responses of Symbiodiniaceae to reveal species specific phenotypic differences (Behrendt et al., 2020; Andersson et al., 2021; Xiao et al., 2022) or monitoring the physiological changes of Symbiodiniaceae cells during protoplast formation and regeneration (Bashir et al., 2022).
Establishing a controlled microenvironment that allows the observation of individual cells in different life stages or forms of Symbiodiniaceae, which also allows the precise modulation of physico-chemical factors is essential. Precise adjustment of microenvironment and concomitant observation of single cells can be achieved by trapping the cells or increasing the viscosity of the medium to slow down rapidly motile cells without affecting the physiological state (i.e., photosynthetic capacity) of the cell population. In the current work, we applied an experimental strategy in which the Symbiodinium cells are not trapped or immobilized, but temporarily slowed down using the physiologically ‘inert’, non-toxic polysucrose Ficoll. Ficoll has been shown to be a useful tool to harvest phytoplankton cells or to improve the stability of photosynthetic activity measurements in microalgae, due to its non-osmotic features which largely preserve cell morphology and physiological parameters, although Ficoll certainly has an impact on viscosity (Joliot and Delosme, 1974; Price et al., 1974). Changing the viscosity using Ficoll has also been found to impact the motility of dinoflagellate Heterocapsa triquetra; increasing viscosity caused slowing down of swimming cells, although the response of the behavior of this alga to changes in viscosity has proven to be complex (Orchard et al., 2016).
The photosynthetic heterogeneity of the different cell forms of Symbiodiniaceae remained largely uncharacterized. This is to a large extent due to the motility of flagellated cells, which prevents accurate estimation of photosynthetic activity at the single-cell level. It was the aim of the current work to monitor the diel changes in motile and non-motile cells using combined morphological analysis and single-cell Chl fluorometry, under varying light-dark periods (normal 12 h: 12 h light: dark and continuous light), using microfluidic cultivating chambers. We aimed to establish conditions when fast-moving cells are transiently slowed down (but not completely immobilized) to mimic symbiotic conditions as opposed to non-symbiotic conditions and to reveal the ensuing changes in photosynthetic activity.
2 Materials and methods
2.1 Symbiodiniaceae cultivation
Symbiodinium microadriaticum Freudenthal culture strain CCMP2467 (former clade A1), originally isolated from the scleractinian coral Stylophora pistillata, was obtained from the National Center for Marine Algae and Microbiota (NCMA), Bigelow Laboratory for Ocean Sciences United States. Cells were grown at 24 °C under 50 µmol photons m-2 s-1 white light (12 h:12 h dark:light cycles) for a week until the mid-log growth phase in F/2 media.
2.2 Microfluidic methods
The microfluidic device was designed, by using the open-source KLayout software (ref: https://www.klayout.de), to allow culturing and observation of Symbiodinium cells and following their cell cycle characteristics. The observational chamber does not have an active flow inside, but the laminar flow near the perforated wall of the chamber allows diffusion of the culturing medium and nutrients through the chamber walls.
The devices were fabricated using soft lithography. Briefly, a master mold was created from SU-82015 photoresist (Microchem corp., Westborough MA, United States) on a silicon wafer substrate by laser direct writing using a Heidelberg μPG 101 micro pattern generator (Heidelberg Instruments GmbH, Heidelberg, Germany). The master mold was replicated by casting of Sylgard 184 polydimethylsiloxane (The Dow Chemical Company, Midland MI, United States). After punching access holes into the cured PDMS pieces, they were bound to glass coverslips by oxygen plasma treatment using a Harrick PDC-002 plasma cleaner (Harrick Plasma, Ithaca NY, United States). The assembled microfluidic devices were filled with distilled water for storage.
The microfluidic chamber used in the current study is shown below (Figure 1, for electron microscopic images of the devices refer to Supplementary Figure S1).
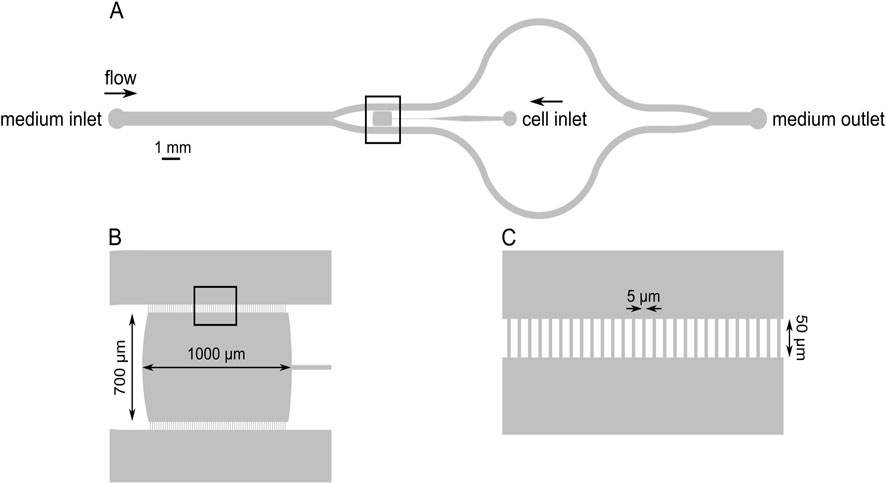
Figure 1. Schematic drawing of the microfluidic device indicating the cell loading port, the medium inlet and outlet, and the growth chamber (A). The growth chamber (the observation area) is marked with a square and shown enlarged in panel (B). The growth chamber was separated from the flow channels with comb-like borders, which allowed the diffusion of the medium into the cultivation chamber but largely prevented the escape of the cells (C).
The microfluidic platform is composed of three parts: the microfluidic device, a tubing system, and a syringe pump. The microfluidic device was mounted on the microscope stage, and a 60 cm long tube was inserted in the inlet hole, while a 40 cm long tube was inserted in the outlet hole (Figure 1). PFTE tubings with 1.6 mm outer diameter and 0.5 mm inner diameter were used. The inlet tube was attached to a syringe (Omnifix-F Solo Luer 5 mL, B. Braun) containing the F/2 medium with the indicated composition, mounted on a syringe pump (SyringeTwo-SKU 4000, New Era Pump Systems, Inc. United States). A constant flow of 40 μL/h was provided to the cells via the inlet port of the chamber using the syringe pump. The images were captured with a 10× objective (Hund Wetzlar, Helmut Hund GmbH, Wetzlar, Germany) by using a light microscope (H 600/12, Hund Wetzlar, Helmut Hund GmbH, Wetzlar, Germany) equipped with a microQ digital camera (UCMOS08000KPB, ToupTek Photonics Co., Ltd., Hangzhou, China). ToupView, a camera control software (ToupTek Photonics Co., Ltd., Hangzhou, China) was used to continuously capture the images at every 30 s, to monitor the changes over time (the microfluidic workstation with the accessories is shown in Supplementary Figure S2).
First, the assembled system was flushed with F/2 medium only. Then, Symbiodinium cells were loaded into the culture chamber through the cell inlet port using an automated pipette equipped with a 200 μL tip. After the assembly of the microfluidic station and loading the cells, the flow was initiated by starting the syringe pump, and the solutions that were flowed through the device exited through the outlet port of the chamber (the experimental flowchart is shown in Supplementary Figure S3). Our model calculations showed that it required about 30 min for the fresh medium to diffuse from the side channels to the center of the culture chambers. The Reynolds number is 0.0029 in F/2 medium without Ficoll and 9.44 × 10−4 in 7% Ficoll (the details are added in the Supplementary Material). The cells in the microfluidic chamber were illuminated with 100 μmol photons m–2 s–1 (12 h:12 h; light:dark diurnal cycle). The day and night cycle were adjusted manually by changing the light filters (blue-white filter for day and green filter for night). The reason for applying weak green light (<0.5 μmol photons m–2 s–1) during the night phase was to maintain weak background light for imaging, therefore, the entire diurnal cycle could be monitored for several days.
Ficoll® 400 was purchased from Sigma-Aldrich. Diluted Ficoll was prepared with F/2 medium at the indicated concentrations. Experiments were performed for the same duration either in the presence of F/2 only or F/2+Ficoll. For the Ficoll experiments, the Ficoll containing F/2 medium was flowed from the beginning of the experiment, and the cell cycle changes were monitored for the indicated time periods, until the diurnal cycles were consistently observed for several days. To test the reversibility of the effect of Ficoll, the F/2+Ficoll solution was replaced with F/2 only medium, and the monitoring was maintained on the same cell population for several days.
Each Ficoll treatment experiment and its reversibility was repeated 3 times, on different biological replicates (i.e., new culture was inoculated in new microfluidic chamber for each separate experiment).
2.3 Single-cell chlorophyll fluorescence imaging
Single-cell Chl fluorescence was determined by pulse-amplitude modulated imaging microfluorometry (Imaging-PAM M-Series chlorophyll fluorometer, microscopy version, with a IMAG-CG control unit and IMAG-L450 measuring head, Heinz Walz GmbH, Effeltrich, Germany) equipped with an IMAG-K6 CCD camera (Allied Vision Technologies GmbH, Ahrensburg, Germany), attached to the microscope. Single-cell photophysiology and morphology were monitored under identical conditions on the same cells. Areas of Interest (AOI) were selected on individual cells, in which the fluorescence of all pixels were averaged. F0 image was obtained using measuring light intensity = 6 (less than 0.5 μmol photons m−2 s−1), gain = 7, damping = 2, F0 averaging n = 3) after 3 min of dark adaptation (during dark adaptation, measuring light was off). A saturating pulse of blue light (460 nm, approx. 2000 μmol photons m−2 s−1, 0.8 s pulse width) was applied to obtain Fm images, and the Fv/Fm parameter, which reflects PSII activity, was calculated as Fv/Fm = (Fm − F0)/Fm (see also Bashir et al., 2022), where F0 is the minimal fluorescence yield when the reaction centres are open, Fm is the maximal fluorescence yield when the reaction centres are closed (in a dark-adapted state) (Schreiber, 2004).
2.4 Microplate experiments
To determine the effective concentration that initiated the slowing down of moving forms, various concentrations ranging from 1% to 10% Ficoll were prepared. Cells were centrifuged and resuspended in the respective Ficoll solution. The samples were transferred to 24-well plates (Vision Plate™ 24 Well, 4titude, Brooks Life Sciences, U.K.) each well representing different Ficoll % (including control without Ficoll). Cells were incubated for 1 h in the different solutions at growth conditions, then short (approx. 15 s) videos were obtained to demonstrate the cell motility in different Ficoll solutions.
2.5 Data analysis and statistical analysis
The number of motile and non-motile cells was determined with a custom-made Matlab software (MathWorks, Natick, Massachusetts, United States). The software segmented each cell based on the dislocation of round cells, and classified the motile and non-motile cells. With the adjustment of cell size and segmentation, the unwanted background objects, non-biological aggregates, and artefacts were excluded. With custom selection of the observation area, the objects of the microfluidic chambers (e.g., the perforated walls) were also excluded to exclusively focus on the biological changes. The software automatically segmented each image, and the number of cells in the different forms was plotted as a function of time for the entire period of the experiments. Statistical analysis was performed using OriginPro (OriginLab Corporation, Northampton, MA, United States). One-way analysis of variance (ANOVA) and Tukey’s post-hoc multiple comparison tests were performed on independent samples to detect statistically significant differences between treatments.
3 Results
Symbiodinium cells exhibited characteristic motile and non-motile features in the microfluidic chambers (Supplementary Table S1). Based on image segmentation, the motile cells were identified as circular objects due to their fast rotation speed (red objects in Figure 2).

Figure 2. Symbiodinium cells in the microfluidic chamber. Moving cells displaying a semi-circular movement are shown in red. Non-moving cells are depicted in blue.
The non-motile cells are separated from motile cells because of the lack of any motion or dislocation of these cells (blue objects in Figure 2). These results indicate that incubating Symbiodiniaceae cells in the cultivating microfluidic chamber maintained the general features of the different cell types observed previously in this alga and the different cell types could be identified and investigated in the microfluidic cultivation chambers.
To resolve the time-dependent changes in cell population and in the fractions of the different cell types, the image analysis was performed at each time point for the entire experimental period (Figure 3).
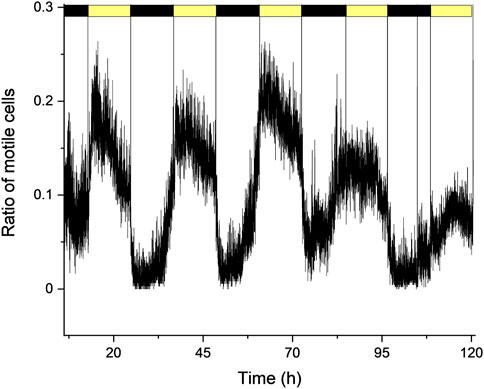
Figure 3. Changes in the ratio of motile cell forms (relative to total cells) under normal F/2 growth medium conditions, under periodic light: dark cycles (12 h: 12 h).
In normal F/2 growth medium, under a 12 h:12 h light: dark cycle, the cells showed a sustained daily oscillation between the motile and non-motile cell forms, in agreement with previous studies (Yamashita and Koike, 2016). In the dark phase, the motility was minimal as the non-motile coccoid form dominated the cell culture. Towards the end of the night cycle, cells started to divide typically into two moving forms (more detailed presentation of cell division in different conditions are also shown below). As the cell division intensified the number of motile cells started to increase, and the relative number of motile cells peaked 2–3 h after the onset of light. Afterwards, the number of motile cells started to decrease and at the onset of dark phase, the motility ceased nearly completely. This cycle could be repeatedly observed for several days, although it has to be noted that the peak amplitude (i.e., the maximal proportion of moving cells relative to total cell number) varied considerably at the different days and among the different replicates, in the range of moving cells/total cell ratio of 0.2–0.4. In the dark phase, this ratio decreased to 0–0.05 (Figure 3). These results indicate that the assessment of time-resolved changes of the cell types and the relative changes in motile cells of the Symbiodinium population during a diurnal cycle could be studied for extended time periods in the microfluidic chamber. It has to be noted that under constant light, the motile and non-motile cells also displayed a diurnal cycle, however the amplitude of the increase in motile cells decreased over time, as the cells tended to lose the diurnal rhythm due to the presence of constant light (Supplementary Figure S4), in agreement with previous observations in bulk cultures (Fitt and Trench, 1983).
In order to slow down the motile cells and create an environment with higher viscosity that mimics coral symbiotic conditions, the polysucrose Ficoll was applied. It was found based on the preliminary microplate experiment that the motility of the cells changed significantly in the Ficoll-containing solutions. In increasing Ficoll concentrations, the motility of the cells slowed down, cells showed a slow swimming or swinging pattern, or the circular motions were visually much slower at 7% Ficoll as compared to control condition (F/2 medium only), in which the cells displayed regular rapid circular movement (Supplementary Figure S5). Therefore, Ficoll provided an environment in which motile cells are remarkably slowed down but the complete immobilization of cells was avoided. In this manner the slowed flagellated cells could be observed and snap photographed in their characteristic spindle shape.
Ficoll (7%) caused a significant alteration in the cell motility in the cultures maintained in the microfluidic chambers (Figure 4A). In agreement with the microplate incubation experiment, the rotational movement of the flagellated cells slowed down as the circular movement pattern visually ceased or significantly decreased (Figures 4C,D). The diurnal cycle also displayed profound changes. At the onset of light, the number of moving forms increased, similarly to the normal F/2 conditions, the relative number of motile cells peaked 2–3 h after the onset of light. However, after the peak phase, the number of motile cells rapidly declined, and by the 6th h of the light phase, the motile cells disappeared nearly completely (Figure 4A). Therefore, the phase of the moving forms was much narrower as compared to the non-treated (Ficoll-free) cultures (c.f. Figures 4A,B). It seems that in the presence of Ficoll, the motile cells appear only temporarily as a spike after the light phase of the diurnal cycle sets on, therefore, the cultures in Ficoll conditions largely remained in non-motile form. However, this does not mean that Ficoll prevented the appearance of the moving forms, instead, the phase of the moving forms was restricted to a narrower time window. The motile cells can be distinguished by their characteristic spindle shape, which was much more apparent due to the slowing down of their rotation in high-viscosity medium (Figures 4C,D). These characteristic spindle-shaped cells could not be visually observed in normal F/2 medium, due to their fast rotation (Figures 4E,F).
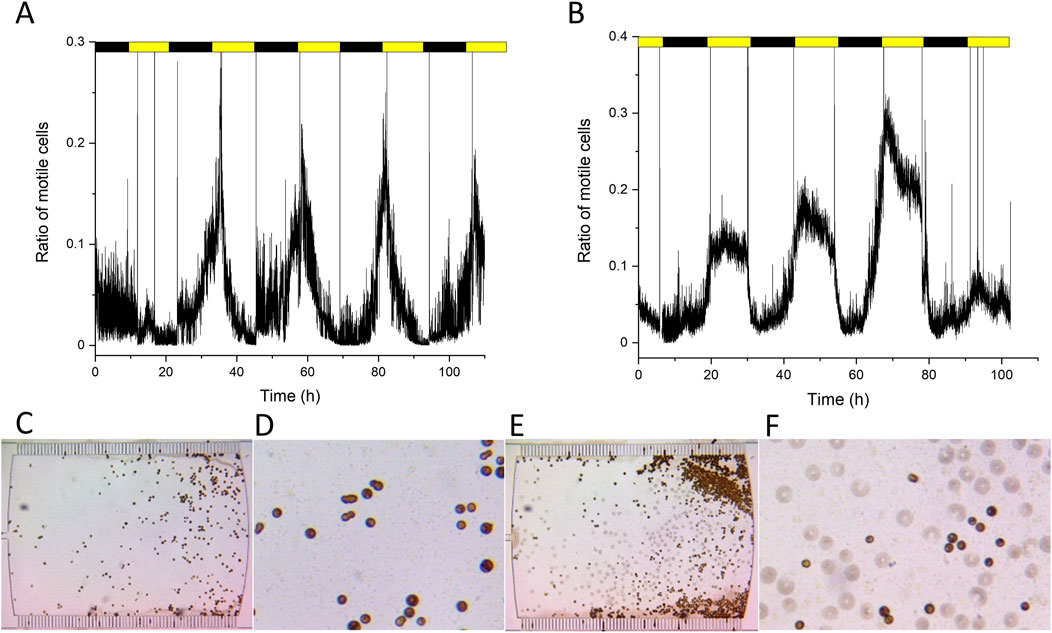
Figure 4. Motility cycles in Ficoll (A) and after washing out Ficoll (B). Bottom panels show the cells in microfluidic chambers in Ficoll treatment (C,D) and after washing out Ficoll (F/2 medium only, (E,F). Panels C and E, original images of the microfluidic chambers, (D,F), zoomed-in (4x) images of (C,E). Replicate experiments are shown in Supplementary Figure S6.
To investigate whether the normal cell cycle could be regained from Ficoll treatment, the Ficoll solution was washed out and replaced with normal F/2 medium. After washing out Ficoll, the daily cell cycle regained its characteristics similar to the control F/2 condition within 30 h, i.e., the wider phase of motile cells during the light period (Figure 4B). The cells regained their fast rotation and motility and so they appeared as circular objects in the observing area (Figure 4F).
It also has to be noted that Ficoll did not impair the normal cell division capacity of the cultures. It was observed, both in the presence and absence of Ficoll, that in the dark phase largely coccoid cells existed, and towards the end of the dark phase, cell division increased. Under all conditions, the cultures maintained the typical behavior of one non-motile cell dividing into two motile daughter cells (Figure 5). Therefore remarkably, the Ficoll treatment did not impact the cell division capacity of Symbiodinium.

Figure 5. Cell division sequence in the presence or absence of Ficoll, before the onset of light (rows 1 and 3) and after the onset of light (rows 2 and 4). Arrows indicate the non-motile dividing cells and the motile daughter cells. The scale bar represents 10 μm.
The activity of PSII was measured under control and Ficoll-treated conditions (Figure 6). Non-slowed motile cells displayed incorrect Fv/Fm values, because cells visibly moved between the F0 (Figure 6A) and Fm (Figure 6B) images, thereby creating artificially high, unrealistic Fv/Fm of 0.7–0.9 (Figures 6C,D). Due to the rapid rotation of the cells in normal F/2 medium, it was not possible to account for the displacement of the cells during Fv/Fm measurement. Ficoll on the other hand, largely prevented the movement of cells, therefore, Fv/Fm could be measured more reliably (Figures 6G,H). However, it has to be noted that even in the case of Ficoll-slowed motile cells, care has to be taken to select the appropriate AOI separately for F0 and Fm (Figures 6E,F), otherwise artificially high Fv/Fm is calculated (blue-purple colored cells in the Ficoll treatment, showing unrealistic Fv/Fm = 0.8 or higher for Symbiodinium, Figures 6G,H).
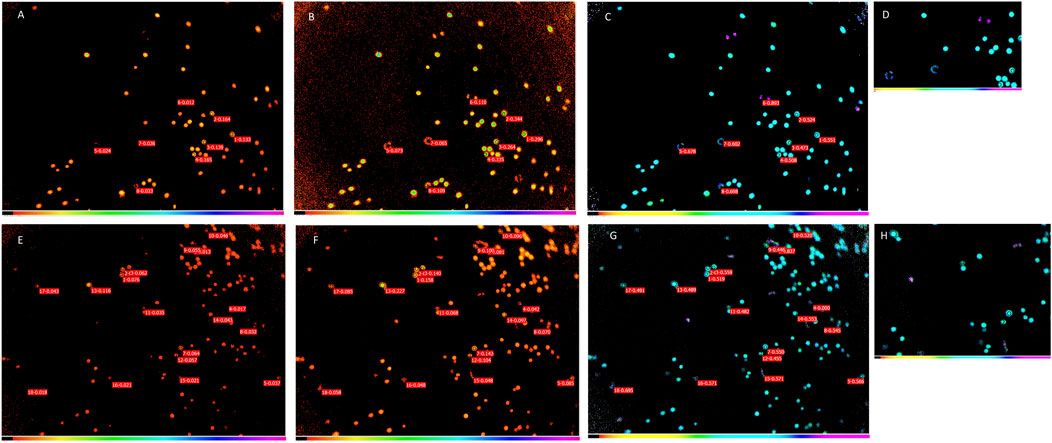
Figure 6. Chl fluorescence images of Symbiodinium cells in control (F/2) (A–D) and Ficoll-treated (E–H) cultures. F0 (minimal fluorescence yield) images (A,E), Fm (maximal fluorescence yield) images (B,F) and Fv/Fm (PSII activity) images (C,G). D and H are zoomed-in images of Fv/Fm images to show typical Fv/Fm of cells in control and Ficoll treatment, respectively.
In the case of slowed motile cells, the F0 and Fm were analyzed with different AOIs, to correct for the small movement of cells (decreased motility due to Ficoll environment), and the Fv/Fm was calculated with from the respective Fm and F0 AOI (similarly to the ‘nudging’ method, which corrects for small cell movements between F′ and Fm’ images, (Oxborough et al., 2000)). In the case of non-motile cells, the same AOI was taken for F0 and Fm, since the non-motile cells were not displaced. By comparing approx. 20 cells, the Fv/Fm values of the non-motile and motile cells were not significantly different (Figure 7).
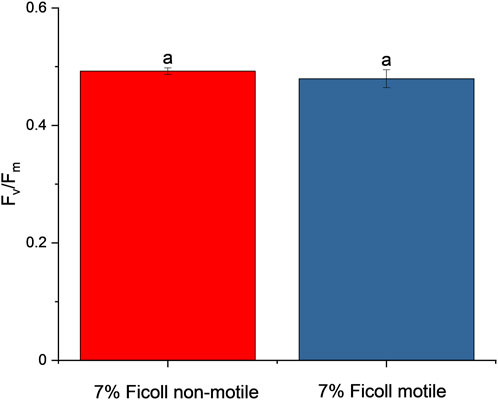
Figure 7. PSII activity (Fv/Fm) of non-motile and motile cells (mean ± S.D., n = 20). Values sharing common letters are not significantly different from one another (One-way ANOVA, p ≤ 0.05).
4 Discussion
An important outcome of the current work is that microfluidic chambers are suitable platforms for the cultivation of the coral endosymbiont algae, Symbiodiniaceae. The microfluidic method applied in the current study represents a large cultivation chamber (in microscopic terms, which is about 1000 × 700 μm) without cell traps. This allowed loading the initial cell culture, observing their morphology, growth, cell division as well as their single-cell photosynthetic activity, with a fixed cultivation setup and geometry for the entire period of the experiment, without the need of repetitive sampling from batch cultures. The cells maintained their motility and division capability and exhibited a normal diurnal cycle typical for Symbiodiniaceae (Figure 3), in agreement with previous findings, which were performed in bulk cultures (Fitt et al., 1981; Yacobovitch et al., 2004; Yamashita and Koike, 2016; Fitt and Trench, 1983).
It has been shown earlier that elevated viscosity significantly alters the motility, swimming patterns, and swimming speed of dinoflagellates (Orchard et al., 2016; Sohn et al., 2013). Freduenthal (1962) showed that increasing the viscosity of the medium using methyl cellulose caused a significant hindrance of the movement of the motile cells of Symbiodiniaceae. Mucus excreted by the coral tissues is accompanied by changes in viscosity, which can therefore fulfil an important physiological role of adjusting the physicochemical and physiological barriers of corals, among other myriad of important functions (Bhagwat et al., 2024). Symbiodinium motility and division, on the other hand, are affected by many factors, including host factor-derived additives and organic compounds (Krueger and Gates, 2012). In the current study, only the viscosity of the medium was changed by the application of Ficoll, which could be precisely controlled by microfluidics. Increasing the viscosity of the medium using Ficoll allowed to reveal the changes in the cell cycle phases in Symbiodinium in a more viscous environment, such as in coral tissues.
The rationale of the application of a medium with elevated viscosity was that the motile cells can be slowed down transiently, and thereby their single-cell characteristics and physiology can be observed without the need for complete immobilization or forcing the cells permanently into non-motile forms. In other words, the establishment of an environment in which slowed motile cells could be observed is imperative.
The typical diurnal cell cycle was maintained in the presence of Ficoll, but with an altered time frame; from the onset to the peak of motile cell number, relative to total cell number, the timeframe was much shorter than in the normal F/2 medium. This could be due to the enhanced viscosity of the medium, which caused the cells to mainly remain in a microenvironment that simulates coral tissue conditions. Nonetheless, the cells retained their active capability for division and did not lose the ability for dividing into motile cells, and typically exhibited the spindle shaped form of mastigotes (Figure 5). However, the motile cells were slowed down, as the circular rotation, typical for motile cells (Nitschke et al., 2015) disappeared, and rather a sliding one-directional motion was observed (Figure 4). Therefore, it appears that a more viscous environment might mimic the coral tissue environment at least in a physico-chemical manner. Importantly, the diurnal alteration of motile and non-motile phases is not lost under these circumstances, only the period length changes. Furthermore, Ficoll did not cause a detrimental effect on cell physiology, as indicated by the fact that the cell cycle regained its properties comparable to control (F/2 only) conditions after washing out Ficoll.
Determination of photophysiological parameters of the motile Symbiodinium cells represent a significant challenge due to their very rapid circular motion in F/2 medium. The basic problem is that the rapid rotation resulted in the overestimation of Fv/Fm since the AOI selected for Fv/Fm calculation was not the same for F0 and Fm images (Figures 6C,D). Recently, software analysis developments allowed a more high-throughput semi-automated analysis of the PSII activity of Symbiodiniaceae cells, immobilized in microfluidic chambers (Pontén et al., 2024), however, these conditions are far from the physiological ones, as these do not allow the analysis of all cell forms of Symbiodiniaceae (i.e., the motile and non-motile forms). With the presented microfluidic setup, the determination of single-cell PSII activity of the motile form could be significantly improved in the presence of Ficoll. Although, the minor movement of the cells still resulted in somewhat altered AOI for Fv/Fm calculation, this slow displacement could be tracked and the AOI could be selected specifically for the F0 and Fm image, based on which the Fv/Fm could be accurately determined. By performing this analysis, the Fv/Fm of non-motile and motile cells, slowed down by Ficoll treatment, exhibited similar PSII activity under the investigated conditions. Although it has to be stressed that the Fv/Fm of motile cells cannot be measured in normal F/2 medium, with the method described above the Fv/Fm of slowed cells could be determined with higher precision, and the observed PSII activity was in the range of Fv/Fm = 0.45–0.5, in agreement with (Behrendt et al., 2020; Pontén et al., 2024).
By this means, employing transient slowing of motile cells, the photosynthetic activity (PSII activity) could be monitored more precisely at the single-cell level, along with the physiology of non-motile forms. It has to be noted that the photosynthetic activity measurements are restricted to Fv/Fm determination in the current study. Determination of Fv/Fm solely does not reveal the rate or the regulation of photosynthetic electron transport in the different cell types. For a detailed understanding of the photosynthetic performance of the different cell types, it would be necessary to perform single-cell fluorescence kinetic or light response curve analyses (e.g., Patil et al., 2023), however, this might be problematic even in the case of slowed-down motile cells, due to the long measurement time (at least several minutes). Due to its precisely controlled nature, microfluidics could be used to monitor important single-cell physiological features of coral host-symbiont interactions, bleaching responses under climate stress scenarios.
In conclusion, our results indicate that the application of Ficoll as an inert polysaccharide caused a slowdown of motile cells, which allowed the monitoring of photosynthetic activity of all cell types of Symbiodinium. Although with the presented microfluidic setup, the single-cell PSII activity can be measured, it is important to note that the PSII activity could be precisely determined only in non-motile or immobilized cells. Furthermore, as the viscosity of the medium increased with the application of Ficoll, this method might represent a physico-chemical environment that mimics coral tissues. However, it has to be noted that a microfluidic analysis of Symbiodinium cell cycle under free-living and in hospite conditions requires a thorough investigation taking into account several host factors, which was beyond the scope of the current study. The microfluidics method, as presented here, serves a high-precision-controlled environment, which might allow the investigation of host conditions and other stress factors. For example, co-culturing Symbiodiniaceae with coral host cells, or integrating thermal stress experimental scenarios are promising future applications of the microfluidics setup presented here.
Data availability statement
The original contributions presented in the study are included in the article/Supplementary Material, further inquiries can be directed to the corresponding author.
Author contributions
MS: Data curation, Conceptualization, Writing – original draft, Resources, Validation, Investigation, Supervision, Funding acquisition, Writing – review and editing. ZS: Formal Analysis, Investigation, Methodology, Writing – original draft, Visualization. ÁÁ: Methodology, Writing – review and editing, Investigation, Visualization, Formal Analysis. KN: Investigation, Writing – review and editing, Methodology, Validation, Formal Analysis, Data curation. LS: Methodology, Software, Writing – review and editing, Visualization. PG: Resources, Data curation, Investigation, Methodology, Writing – review and editing, Validation, Formal Analysis. IV: Writing – original draft, Supervision, Writing – review and editing, Conceptualization, Data curation, Investigation, Resources.
Funding
The author(s) declare that financial support was received for the research and/or publication of this article. This work was supported by the National Research, Development and Innovation Office (NKFIH), grant: FK 146298 and the grant GINOP-2.3.2-15-2016-00026.
Acknowledgments
Sándor Kovács is acknowledged for the designing and testing the microfluidic chambers at the early phase of the work.
Conflict of interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Generative AI statement
The author(s) declare that no Generative AI was used in the creation of this manuscript.
Any alternative text (alt text) provided alongside figures in this article has been generated by Frontiers with the support of artificial intelligence and reasonable efforts have been made to ensure accuracy, including review by the authors wherever possible. If you identify any issues, please contact us.
Publisher’s note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
Supplementary material
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fphbi.2025.1645420/full#supplementary-material
References
Ábrahám, Á., Dér, L., Csákvári, E., Vizsnyiczai, G., Pap, I., Lukács, R., et al. (2024). Single-cell level LasR-mediated quorum sensing response of Pseudomonas aeruginosa to pulses of signal molecules. Sci. Rep. 14 (1), 16181. doi:10.1038/s41598-024-66706-6
Andersson, M., Johansson, S., Bergman, H., Xiao, L., Behrendt, L., and Tenje, M. (2021). A microscopy-compatible temperature regulation system for single-cell phenotype analysis – demonstrated by thermoresponse mapping of microalgae. Lab a Chip 21 (9), 1694–1705. doi:10.1039/D0LC01288B
Baker, N. R. (2008). Chlorophyll fluorescence: a probe of photosynthesis in vivo. Annu. Rev. Plant Biol. 59, 89–113. doi:10.1146/annurev.arplant.59.032607.092759
Bashir, F., Kovács, S., Ábrahám, Á., Nagy, K., Ayaydin, F., Valkony-Kelemen, I., et al. (2022). Viable protoplast formation of the coral endosymbiont alga symbiodinium spp. in a microfluidics platform. Lab a Chip 22, 2986–2999. doi:10.1039/D2LC00130F
Behrendt, L., Salek, M. M., Trampe, E. L., Fernandez, V. I., Lee, K. S., Kühl, M., et al. (2020). PhenoChip: a single-cell phenomic platform for high-throughput photophysiological analyses of microalgae. Sci. Adv. 6 (36), eabb2754. doi:10.1126/sciadv.abb2754
Bhagwat, P. V., Ravindran, C., and Irudayarajan, L. (2024). Beneficial properties of mucus in coral adaptations and ecological interactions. Mar. Biol. 171 (2), 46. doi:10.1007/s00227-023-04372-4
Cui, G., Liew, Y. J., Konciute, M. K., Zhan, Y., Hung, S.-H., Thistle, J., et al. (2022). Nutritional control regulates symbiont proliferation and life history in coral-dinoflagellate symbiosis. BMC Biol. 20 (1), 103. doi:10.1186/s12915-022-01306-2
Davy, S. K., Allemand, D., and Weis, V. M. (2012). Cell biology of cnidarian-dinoflagellate symbiosis. Microbiol. Mol. Biol. Rev. 76 (2), 229–261. doi:10.1128/mmbr.05014-11
Fitt, W. K., and Trench, R. K. (1983). The relation of diel patterns of cell division to diel patterns of motility in the symbiotic dinoflagellate Symbiodinium microadriaticum freudenthal in culture. New Phytol. 94 (3), 421–432. doi:10.1111/j.1469-8137.1983.tb03456.x
Fitt, W. K., Chang, S. S., and Trench, R. K. (1981). Motility patterns of different strains of the symbiotic dinoflagellate symbiodinium (=Gymnodinium) microadriaticum (freudenthal) in culture. Bull. Mar. Sci. 31 (2), 436–443.
Freduenthal, H. D. (1962). Symbiodinium gen. nov. and Symbiodinium microadriaticum sp. Nov., a zooxanthella: taxonomy, life cycle, and morphology.*. J. Protozoology 9 (1), 45–52. doi:10.1111/j.1550-7408.1962.tb02579.x
Greipel, E., Nagy, K., Csákvári, E., Dér, L., Galajda, P., and Kutasi, J. (2024). Chemotactic interactions of scenedesmus sp. and Azospirillum brasilense investigated by microfluidic methods. Microb. Ecol. 87 (1), 52. doi:10.1007/s00248-024-02366-3
Joliot, P., and Delosme, R. (1974). Flash-induced 519 nm absorption change in green algae. Biochim. Biophys. Acta 357 (2), 267–284. doi:10.1016/0005-2728(74)90066-8
Koike, K., Jimbo, M., Sakai, R., Kaeriyama, M., Muramoto, K., Ogata, T., et al. (2004). Octocoral chemical signaling selects and controls dinoflagellate symbionts. Biol. Bull. 207 (2), 80–86. doi:10.2307/1543582
Komárek, O., Felcmanová, K., Šetlíková, E., Kotabová, E., Trtílek, M., and Prášil, O. (2010). Microscopic measurements of the chlorophyll a fluorescence kinetics. In: D. J. Suggett, O. Prášil, and M. A. Borowitzka, editors. Chlorophyll a fluorescence in aquatic sciences: methods and applications. Netherlands, Dordrecht: Springer. p. 91–101. doi:10.1007/978-90-481-9268-7_5
Krueger, T., and Gates, R. D. (2012). Cultivating endosymbionts — host environmental mimics support the survival of symbiodinium C15 ex hospite. J. Exp. Mar. Biol. Ecol. 413, 169–176. doi:10.1016/j.jembe.2011.12.002
Lee, S. Y., Jeong, H. J., Kang, N. S., Jang, T. Y., Jang, S. H., and Lajeunesse, T. C. (2015). Symbiodinium tridacnidorum sp. Nov., a dinoflagellate common to indo-Pacific giant clams, and a revised morphological description of Symbiodinium microadriaticum freudenthal, emended trench and blank. Eur. J. Phycol. 50 (2), 155–172.
Nagy, K., Ábrahám, Á., Keymer, J. E., and Galajda, P. (2018). Application of microfluidics in experimental ecology: the importance of being spatial. Front. Microbiol. 9 (496), 496. doi:10.3389/fmicb.2018.00496
Nitschke, M. R., Davy, S. K., Cribb, T. H., and Ward, S. (2015). The effect of elevated temperature and substrate on free-living symbiodinium cultures. Coral Reefs 34 (1), 161–171. doi:10.1007/s00338-014-1220-8
Orchard, M. J., Humphries, S., Schuech, R., and Menden-Deuer, S. (2016). The influence of viscosity on the motility and sensory ability of the dinoflagellate Heterocapsa triquetra. J. Plankton Res. 38 (4), 1062–1076. doi:10.1093/plankt/fbw004
Oxborough, K., and Baker, N. R. (1997). An instrument capable of imaging chlorophyll a fluorescence from intact leaves at very low irradiance and at cellular and subcellular levels of organization. Plant, Cell Environ. 20 (12), 1473–1483. doi:10.1046/j.1365-3040.1997.d01-42.x
Oxborough, K., Hanlon, A. R. M., Underwood, G. J. C., and Baker, N. R. (2000). In vivo estimation of the photosystem II photochemical efficiency of individual microphytobenthic cells using high-resolution imaging of chlorophyll a fluorescence. Limnol. Oceanogr. 45 (6), 1420–1425. doi:10.4319/lo.2000.45.6.1420
Patil, P. P., Nagy, K., Ábrahám, Á., Vass, I., and Szabó, M. (2023). Monitoring the photosynthetic activity at single-cell level in Haematococcus lacustris. Photosynthetica 61 (4), 473–482. doi:10.32615/ps.2023.042
Pontén, O., Xiao, L., Kutter, J., Cui, Y., Wählby, C., and Behrendt, L. (2024). PACMan: a software package for automated single-cell chlorophyll fluorometry. Cytom. Part A 105 (3), 203–213. doi:10.1002/cyto.a.24808
Price, C. A., Mendiola-Morgenthaler, L. R., Goldstein, M., Breden, E. N., and Guillard, R. R. L. (1974). Harvest of planktonic marine algae by centrifugation into gradients of silica in the CF-6 continuous-flow zonal rotor. Biol. Bull. 147 (1), 136–145. doi:10.2307/1540574
Roth, M. S. (2014). The engine of the reef: photobiology of the coral–algal symbiosis. Front. Microbiol. 5, 422. doi:10.3389/fmicb.2014.00422
Schreiber, U. (2004). Pulse-amplitude-modulation (PAM) fluorometry and saturation pulse method: an overview. In: G. C. Papageorgiou, and Govindjee, editors. Chlorophyll a fluorescence. Advances in photosynthesis and respiration. Dordrecht: Springer, Vol. 19. p. 279–319. doi:10.1007/978-1-4020-3218-9_11
Sohn, M. H., Lim, S., Seo, K. W., and Lee, S. J. (2013). Effect of ambient medium viscosity on the motility and flagella motion of Prorocentrum minimum (dinophyceae). J. Plankton Res. 35 (6), 1294–1304. doi:10.1093/plankt/fbt071
Sorek, M., Díaz-Almeyda, E. M., Medina, M., and Levy, O. (2014). Circadian clocks in symbiotic corals: the duet between Symbiodinium algae and their coral host. Mar. Genomics 14, 47–57. doi:10.1016/j.margen.2014.01.003
Stat, M., Carter, D., and Hoegh-Guldberg, O. (2006). The evolutionary history of symbiodinium and scleractinian hosts—Symbiosis, diversity, and the effect of climate change. Perspect. Plant Ecol. Evol. Syst. 8 (1), 23–43. doi:10.1016/j.ppees.2006.04.001
Széles, E., Nagy, K., Ábrahám, Á., Kovács, S., Podmaniczki, A., Nagy, V., et al. (2022). Microfluidic platforms designed for morphological and photosynthetic investigations of Chlamydomonas reinhardtii on a single-cell level. Cells 11 (2), 285. doi:10.3390/cells11020285
Széles, E., Kuntam, S., Vidal-Meireles, A., Nagy, V., Nagy, K., Ábrahám, Á., et al. (2023). Single-cell microfluidics in combination with chlorophyll a fluorescence measurements to assess the lifetime of the chlamydomonas PSBO protein. Photosynthetica 61 (SI), 417–424. doi:10.32615/ps.2023.028
Weibel, D. B., DiLuzio, W. R., and Whitesides, G. M. (2007). Microfabrication meets microbiology. Nat. Rev. Microbiol. 5 (3), 209–218. doi:10.1038/nrmicro1616
Weis, V. M. (2008). Cellular mechanisms of Cnidarian bleaching: stress causes the collapse of symbiosis. J. Exp. Biol. 211, 3059–3066. doi:10.1242/jeb.009597
Xiao, L., Johansson, S., Rughöft, S., Burki, F., Sandin, M. M., Tenje, M., et al. (2022). Photophysiological response of symbiodiniaceae single cells to temperature stress. ISME J. 16 (8), 2060–2064. doi:10.1038/s41396-022-01243-6
Yacobovitch, T., Benayahu, Y., and Weis, V. M. (2004). Motility of zooxanthellae isolated from the Red Sea soft coral Heteroxenia fuscescens (Cnidaria). J. Exp. Mar. Biol. Ecol. 298 (1), 35–48. doi:10.1016/j.jembe.2003.08.003
Keywords: coral symbiont algae, single-cell analysis, microfluidics, cell cycle, photosynthetic activity
Citation: Szabó M, Szabó Z, Ábrahám Á, Nagy K, Sass L, Galajda P and Vass I (2025) Assessment of the morphological features, physiological and photosynthetic activity of the different cell forms of Symbiodiniaceae using microfluidic methods. Front. Photobiol. 3:1645420. doi: 10.3389/fphbi.2025.1645420
Received: 11 June 2025; Accepted: 12 August 2025;
Published: 29 August 2025.
Edited by:
Radek Kana, Academy of Sciences of the Czech Republic (ASCR), CzechiaReviewed by:
Anthony William Larkum, University of Technology Sydney, AustraliaFaiza Bashir, Hanyang University, Republic of Korea
Copyright © 2025 Szabó, Szabó, Ábrahám, Nagy, Sass, Galajda and Vass. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Milán Szabó, c3phYm8ubWlsYW5AYnJjLmh1