 Lissette Cabrera1,2
Lissette Cabrera1,2 Andrea Saavedra1Susana Rojas1
Andrea Saavedra1Susana Rojas1 Marcela Cid3Cristina Valenzuela1
Marcela Cid3Cristina Valenzuela1 David Gallegos1Pamela Careaga1
David Gallegos1Pamela Careaga1 Emerita Basualto1Astrid Haensgen1Eduardo Peña4Coralia Rivas4Juan Carlos Vera4Victoria Gallardo4,5
Emerita Basualto1Astrid Haensgen1Eduardo Peña4Coralia Rivas4Juan Carlos Vera4Victoria Gallardo4,5 Leandro Zúñiga6
Leandro Zúñiga6 Carlos Escudero5,7
Carlos Escudero5,7 Luis Sobrevia8,9,10Mark Wareing11,12
Luis Sobrevia8,9,10Mark Wareing11,12 Marcelo González1,5*
Marcelo González1,5*- 1Vascular Physiology Laboratory, Department of Physiology, Faculty of Biological Sciences, Universidad de Concepción, Concepción, Chile
- 2Department of Morphophysiology, Faculty of Medicine, Universidad Diego Portales, Santiago, Chile
- 3Department of Obstetrics and Childcare, Faculty of Medicine, Universidad de Concepción, Concepción, Chile
- 4Department of Pathophysiology, Faculty of Biological Sciences, Universidad de Concepción, Concepción, Chile
- 5Group of Research and Innovation in Vascular Health (GRIVAS Health), Chillán, Chile
- 6Centro de Investigaciones Médicas (CIM), School of Medicine, Universidad de Talca, Talca, Chile
- 7Vascular Physiology Laboratory, Group of Investigation in Tumor Angiogenesis (GIANT), Department of Basic Sciences, Universidad del Biobío, Chillán, Chile
- 8Cellular and Molecular Physiology Laboratory (CMPL), Division of Obstetrics and Gynecology, School of Medicine, Faculty of Medicine, Pontificia Universidad Católica de Chile, Santiago, Chile
- 9Department of Physiology, Faculty of Pharmacy, Universidad de Sevilla, Seville, Spain
- 10University of Queensland Centre for Clinical Research (UQCCR), Faculty of Medicine and Biomedical Sciences, University of Queensland, Herston, QLD, Australia
- 11Maternal and Fetal Health Research Centre, Institute of Human Development, University of Manchester, Manchester, UK
- 12Maternal and Fetal Health Research Centre, St. Mary's Hospital, Central Manchester University Hospitals NHS Foundation Trust, Manchester Academic Health Science Centre, Manchester, UK
HIGHLIGHTS
• Short-term incubation with insulin increases the L-arginine transport in HUVECs.
• Short-term incubation with insulin increases the NO synthesis in HUVECs.
• Insulin induces relaxation in human placental vascular bed.
• Insulin attenuates the constriction induced by hydrogen peroxide in human placenta.
• The relaxation induced by insulin is dependent on BKCa channels activity in human placenta.
Insulin induces relaxation in umbilical veins, increasing the expression of human amino acid transporter 1 (hCAT-1) and nitric oxide synthesis (NO) in human umbilical vein endothelial cells (HUVECs). Short-term effects of insulin on vasculature have been reported in healthy subjects and cell cultures; however, its mechanisms remain unknown. The aim of this study was to characterize the effect of acute incubation with insulin on the regulation of vascular tone of placental vasculature. HUVECs and chorionic vein rings were isolated from normal pregnancies. The effect of insulin on NO synthesis, L-arginine transport, and hCAT-1 abundance was measured in HUVECs. Isometric tension induced by U46619 (thromboxane A2 analog) or hydrogen peroxide (H2O2) were measured in vessels previously incubated 30 min with insulin and/or the following pharmacological inhibitors: tetraethylammonium (KCa channels), iberiotoxin (BKCa channels), genistein (tyrosine kinases), and wortmannin (phosphatidylinositol 3-kinase). Insulin increases L-arginine transport and NO synthesis in HUVECs. In the placenta, this hormone caused relaxation of the chorionic vein, and reduced perfusion pressure in placental cotyledons. In vessels pre-incubated with insulin, the constriction evoked by H2O2 and U46619 was attenuated and the effect on H2O2-induced constriction was blocked with tetraethylammonium and iberiotoxin, but not with genistein, or wortmannin. Insulin rapidly dilates the placental vasculature through a mechanism involving activity of BKCa channels and L-arginine/NO pathway in endothelial cells. This phenomenon is related to quick increases of hCAT-1 abundance and higher capacity of endothelial cells to take up L-arginine and generate NO.
Introduction
In the human placenta, an organ lacking innervation, endothelial factors released in response to shear stress, oxygen level, paracrine, or endocrine signals, are the main mechanisms that maintain low resistance and blood flow in the placental vascular bed (Wareing, 2014). In this regard, insulin induces relaxation in human umbilical veins via a mechanism involving increased transcriptional activity of SLC7A1 (coding for human Cationic Amino Acid Transporter-1, hCAT-1) and L-arginine transport (González et al., 2011). In addition, insulin also increases nitric oxide (NO) synthesis through activation of phosphatidylinositol 3-kinase (PI3K) and endothelial NO synthase (eNOS) in HUVECs (González et al., 2004). We reported that insulin may also generate hyperpolarization in this cell type (González et al., 2004), which enhances vasomotor activity of this hormone in the placental macrocirculation and microcirculation. Despite this evidence, detailed mechanisms underlying the vasomotor activity of insulin are still unclear.
Regulation of the plasma membrane potential in both endothelial cells and vascular smooth muscle cells (VSMCs) involves potassium (K+) channel activity—dependent hyperpolarization (Durand and Gutterman, 2013). The human placenta expresses large conductance calcium-activated K+ channels (BKCa) (Sand et al., 2006; Wareing, 2014), mainly expressed in VSMCs, and small (SKCa) and intermediate (IKCa) conductance K+ channels, predominantly expressed in the endothelium (Sandow and Grayson, 2009; Kerr et al., 2012). In HUVECs, BKCa channel blocker iberiotoxin inhibits K+ currents, whereas sildenafil (Luedders et al., 2006) and insulin (Wiecha et al., 1998) activate BKCa. These last results suggest functional presence of BKCa channels in this endothelial cell type, and confirm insulin control of endothelial plasma membrane polarization. Interestingly, insulin increases L-arginine transport by modulating hCAT-1 expression and its availability at the plasma membrane (González et al., 2011), an effect that correlates with membrane hyperpolarization in HUVECs (González et al., 2004). Indeed, the bioavailability of NO and propagation of hyperpolarization from endothelial cells to VSMCs would be the main mechanisms involved in regulation of blood flow in macrocirculation and microcirculation (Figueroa and Duling, 2009). However, it is still unclear if NO generation depends on whether or not this gas induces the activity of KCa channels in the human placenta endothelium.
Other vascular tone regulators are reactive oxygen species (ROS) and nitrogen species (RNS), which constitute a family of radical and non-radical derivatives of molecular oxygen (O2) and nitrogen (N2), respectively (Klandorf and Van Dyke, 2012). Specifically, hydrogen peroxide (H2O2) and peroxynitrite (ONOO−) induce rapid and transient contraction or relaxation in human placental chorionic plate arteries (Mills et al., 2009). In addition, H2O2 and ONOO− have higher stability than their respective precursors (Beckman and Koppenol, 1996), and may cause deleterious effects in vascular beds when insufficiently buffered/neutralized (González et al., 2011, 2015). In this regard, several reports have shown that ROS reduces NO availability and vascular relaxation in human placenta vasculature, during either healthy or pathological conditions such as preeclampsia (PE; Bernardi et al., 2008; Catarino et al., 2012), intrauterine growth restriction (IUGR; Takagi et al., 2004), and gestational diabetes mellitus (GDM; Coughlan et al., 2004).
However, it is unknown whether H2O2 might control vascular tone in the chorionic plate veins or in the placental microcirculation. It is also unknown whether insulin can regulate the vascular tone in these placental vessels. Therefore, the present study aimed to determine whether insulin attenuates the vascular response induced by H2O2 in the placental vasculature, and elucidate whether BKCa channel activity, endothelial expression, and activity of hCAT-1 are involved in this process.
Methods
Ethics Statement
This investigation conforms to the principles outlined in the Declaration of Helsinki, and has received approval from the Ethics Committee of the Faculty of Biological Sciences of Universidad de Concepción, the Hospital Regional Guillermo Grant Benavente, Concepción Chile, and National Research Ethics System (NRES ref; 08/H1010/55), UK, and the Comisión Nacional de Investigación en Ciencia y Tecnología (CONICYT grant number 11100192, Chile). All women signed written informed consent. The maternal and newborns clinical parameters are summarized in Table 1.
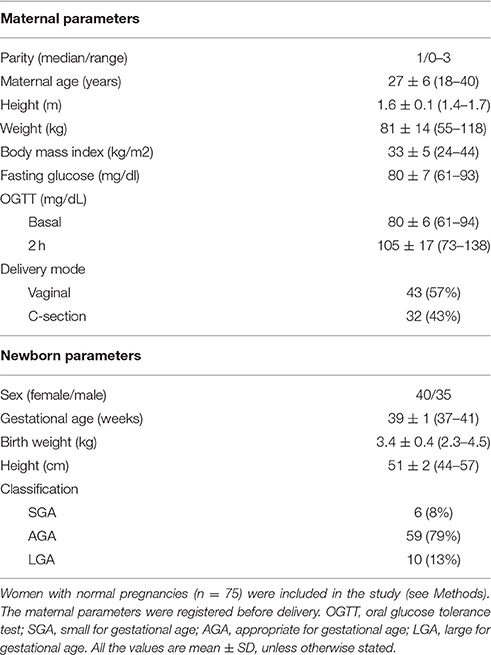
Table 1. Maternal and newborns clinical parameters.
Human Placenta and Umbilical Cords Collection
Placentas with their umbilical cords were collected after delivery from 75 full-term normal pregnancies from the Hospital Regional Guillermo Grant Benavente in Concepción (Chile) and St. Mary's Hospital in Manchester (UK). All pregnancies were single births. The pregnant women did not smoke or consume drugs or alcohol, had no intrauterine infection or any medical or obstetrical complications, were normotensive and exhibited a normal response to the oral glucose tolerance test. They were under a normal food regimen during the whole pregnancy period and newborns were at term, born by vaginal delivery or cesarean section. Placentas were transferred in a sterile container (4°C) to the laboratory. Sections of umbilical cords (10–20 cm length) were collected into sterile 200 ml phosphate-buffered saline (PBS) solution [(mM): 130 NaCl, 2.7 KCl, 0.8 Na2HPO4, 1.4 KH2PO4 (pH 7.4, 4°C)] and used for isolation of umbilical vein endothelial cells (HUVECs) between 6–12 h after delivery.
Cell Culture
HUVECs were isolated by collagenase digestion (0.25 mg/ml Collagenase Type I from Clostridium histolyticum; Gibco Life Technologies, Grand Island, NY, USA) as previously described (González et al., 2004). In brief, cells were cultured (37°C, 5% CO2) up to passage 3 in medium 199 (M199) (Gibco Life Technologies, Grand Island, NY, USA) containing 5 mM D-glucose, 10% newborn calf serum (NBCS), 10% fetal calf serum (FCS), 3.2 mM L-glutamine, and 100 U/ml penicillin-streptomycin (primary culture medium, PCM). Experiments were performed on cells incubated (0–30 min) in M199 in the absence or presence of insulin (1 nM). Cell viability estimated by Trypan blue exclusion was higher than 97% (not shown). Sixteen hours prior, the experimental incubation medium was changed to sera-free M199 (González et al., 2015).
L-Arginine Transport
Overall L-arginine transport (2 μCi/ml L-[3H]arginine (NEN, Dreieich, FRG), 0–250 μM L-arginine, 1 min, 37°C) was measured as previously described (González et al., 2015). Briefly, transport assays were performed in Krebs [in mM: 131 NaCl, 5.6 KCl, 25 NaHCO3, 1 NaH2PO4, 20 Hepes, 2.5 CaCl2, 1 MgCl2 (pH 7.4, 37°C)] in cells preincubated (12 h) with M199 in the absence (control) or presence (1–30 min) of insulin (1 nM). Cell monolayers were rinsed with ice-cold Krebs to terminate tracer uptake. Radioactivity in formic acid cell digests was determined by liquid scintillation counting, and uptake was corrected for D-[3H]mannitol (NEN) disintegrations per minute (d.p.m.) in the extracellular space. Overall transport at initial rates (i.e., linear uptake up to 1 min) was adjusted to the Michaelis-Menten hyperbola plus a nonsaturable, lineal component as described (Christensen, 1962). The maximal velocity (Vmax) and apparent Michaelis-Menten constant (Km) of saturable transport were calculated as described (Christensen, 1962) by this equation:
Immunofluorescence and Confocal Laser Scanning Microscopy
HUVECs were grown on microscope coverslips (106 cells/slide) (Marienfeld GmbH & Co. KG, Lauda Königshofen, Baden-Württemberg, Germany) in PCM. Cells were incubated for 30 min in M199 in the absence or presence of 1 nM insulin (see above). Cells were then fixed in 4% paraformaldehyde (15 min), rinsed (x3) with Hanks solution [(mM): CaCl2 1.26, KCl2 5.37, KH2PO4 0.44, MgSO4 8.11, NaCl 136.89, Na2HPO4 0.33, NaHCO3 4.16 (37°C, pH 7.4)], permeabilized (in some experiments cells were not permeabilized) with 0.1% Triton X-100 (20 min), and blocked (1 h) with 1% BSA. Monoclonal hCAT-1 antibody (1:100) (Sigma-Aldrich, St. Louis, MO, USA) was incubated (overnight at 4°C) in PBS containing 5% BSA. Cells were washed (x3) with Hanks solution followed by incubation (1 h) with the secondary antibody, fluorescein isothiocyanate (FITC) goat anti-mouse IgG (H+L) (λexc/λem:492/520 nm) (1:2000) (Thermo Fisher Scientific, Inc., Waltham, MA, USA) in PBS containing 5% BSA. Nuclei were counterstained with Vectashield mounting media stained with 4,′6-diamidino-2-phenylindole (DAPI) (Vector Laboratories, Burlingame, CA, USA). Samples were analyzed under an Olympus IX81 microscope with a DSU spinning disk confocal system (Olympus, Tokyo, Japan). Images were obtained with a Hamamatsu ORCA-R2 camera (Hamamatsu Photonics, Hamamatsu, Japan) controlled by the Olympus XcellenceR software using a Plan Apo N 60 × 1.42 NA objective. Each sample was examined through successive 0.2 μm optical slices along the z axis. Images were analyzed using Imaris software (Switzerland), considering the fluorescence of cell volume from optical slices.
DAF Fluorescence
HUVECs were grown on microscope coverslips and intracellular NO was determined in cells incubated with insulin (1 nM, 1–30 min) and exposed (45 min, 37°C) to 10 μM of 4-amino-5-methylamino-2′,7′-difluorofluorescein (DAF-FM) (Molecular Probes, Leiden, The Netherlands). The fluorescence was observed in fixed cells by fluorescence microscopy (Olympus IX81) and the signal density was analyzed by Image J software (Java-based imaging processing program, National Institute of Health, USA).
Isolated Cotyledon Perfusion
Techniques for perfusion of the placental cotyledon was that of Penfold et al. (1981), which was modified by perfusing only the fetal vascular compartment, instead of the dual perfusion model (Acevedo et al., 1995). After delivery (15–30 min), a fetal vein and artery pair on surface of chorionic plate, leading to peripheral cotyledon, was cannulated with plastic tubing. Each cotyledon was perfused with Krebs-Ringer solution at a constant flow rate (7 mL/min), maintained with oxygen levels similar to physiological conditions for placental vessels in situ. The perfusion pressure was continuously monitored and the viability of preparation was controlled as previously described (Acevedo et al., 1999).
Wire Myography
Chorionic plate veins, identified as branches of the umbilical vein, were dissected from biopsies and placed in an ice-cold physiological saline solution (PSS). Veins were mounted on a myograph (610 M; Danish Myotechnology, Aarhus, Denmark) and normalized to 0.9 L5.1kPaas described (Mills et al., 2009). Vessels were bathed in PSS and maintained with oxygen levels similar to physiological conditions for chorionic vessels in situ (Mills et al., 2009). After the vein rings were stabilized for isometric force measurements with optimal diameter (~310 μm), the maximal active response was determined with modified PSS containing 90 mM KCl. Two different protocols were designed to determine the effects of insulin consisting of incubation of pre-constricted (U46619) veins with the hormone (in the presence of inhibitors); or preincubation (30 min) with insulin (in the presence of inhibitors) prior to U46619 (10−10–10−5 M) or H2O2 (10−5–10−3 M) exposure.
Pharmacological Agents
General chemicals and pharmacological agents were purchased from Sigma-Aldrich, St. Louis, MO, USA, including; insulin (1–10 nM), H2O2(0.01–1 mM), tetraethylammonium (1 mM) (KCa channels inhibitor), iberiotoxin (100 nM) (BKCa channels inhibitor), genistein (50 μM) (tyrosine kinases inhibitor) and wortmannin (30 nM) (phosphatidylinositol 3-kinase inhibitor). U46619 (0.0001–1 μM; thromboxane A2 analog) was obtained from Tocris Bioscience, Bristol, UK.
Statistical Analysis
Values are mean ± S.E.M., where n indicates the number of different cell cultures (three to four replicates). Comparisons between two or more groups were performed by means of Student's unpaired t-test and analysis of variance (ANOVA), respectively. If the ANOVA demonstrated a significant interaction between variables, post-hoc analyses were performed by the multiple-comparison Bonferroni correction test. Values of p < 0.05 were considered statistically significant.
Results
Insulin Increases the L-Arginine/NO Pathway in HUVECs
Apparent Km was maintained in a range between 54 ± 16 to 153 ± 18 μM (Figure 1A, Table 2) in all experimental conditions. Insulin increased the Vmax of L-arginine transport with maximal effect (5.4 ± 0.9-fold) after 30 min of treatment and lower but significant (p < 0.005) increases after 3 min (2.2 ± 0.8-fold) and 5 min (2 ± 0.5-fold) of incubation (Figures 1A,B, Table 2). No changes in Vmax were detected after 10 or 20 min of treatment with insulin. The Vmax/Km was significantly increased 2.6 ± 0.8 and 2.5 ± 0.9-fold after 5 min and 30 min incubation with 1 nM insulin, respectively (Figure 1B, Table 2). Insulin (1 nM, 30 min) increased hCAT-1–associated fluorescence in permeabilized (1.8 ± 0.3-fold) and non-permeabilized (2.8 ± 0.3-fold) cells, compared to control (Figure 1C). The fold of increase induced by insulin in hCAT-1-associated fluorescence is 55% higher (2.8 vs. 1.8-fold) in non-permeabilized cells compared with permeabilized cells. From total fluorescence, in insulin-treated cells the 78% (27.8 vs. 35.6 arbitrary units of fluorescence) correspond to hCAT-1 expression on cell surface, meanwhile this percentage decreased until 50% (10 vs. 20 arbitrary units of fluorescence) in control cells (Figure 1D). Insulin also increased NO levels (Figure 2A) with maximal effect (2.5 ± 0.2-fold) (Figure 2B) after 30 min of incubation. No changes were detected in control cells incubated by 30 min in medium without insulin (Figure 2A). The basal level of DAF fluorescence in HUVECs was largely detected in a vesicular-like form; however, in cells incubated with insulin, the fluorescence was diffused throughout the cytoplasm (Figure 2C).
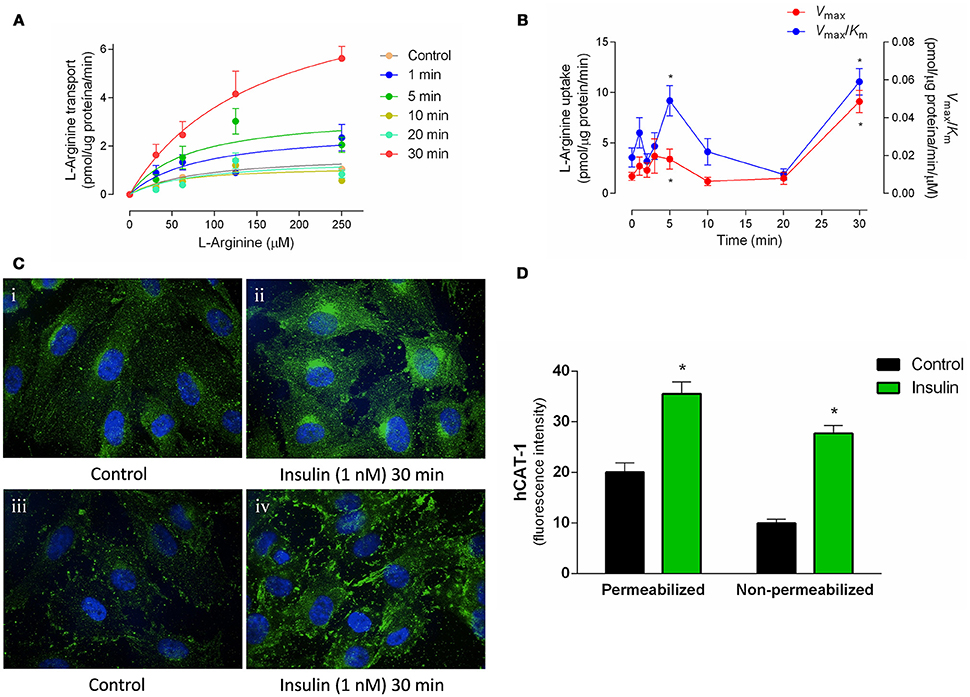
Figure 1. Insulin induces rapid increases in the hCAT-1 activity. L-Arginine transport (0–250 μmol/L L-arginine, 2 μCi/mL L-[3H]arginine, 1 min, 37°C) and hCAT-1 expression were determined in HUVECs pre-incubated (1–30 min) in medium 199 in absence (control) or presence of 1 nM insulin. Saturable transport was adjusted to Michaelis-Menten kinetic curve (A) and maximal velocity (Vmax) and maximal transport capacity (Vmax/Km) values were plotted and calculated from experimental data (B). hCAT-1 expression was determined through immunocytochemistry (green fluorescence) in permeabilized (Ci; Cii) or non-permeabilized (Ciii; Civ) cells. Control cells are in the absence of insulin. Images were obtained with 60x magnification in confocal microscopy. (D) Mean fluorescence intensity was determined based on cells volume of three different fields of each experiment and values in y-axis are presented as arbitrary units. *P < 0.05 vs. values in the absence of insulin. In (A,B), values are mean ± S.E.M. (n = 12–15). In (C), images are representative of three different cell cultures and graph (D) shows mean ± S.E.M. (n = 3).
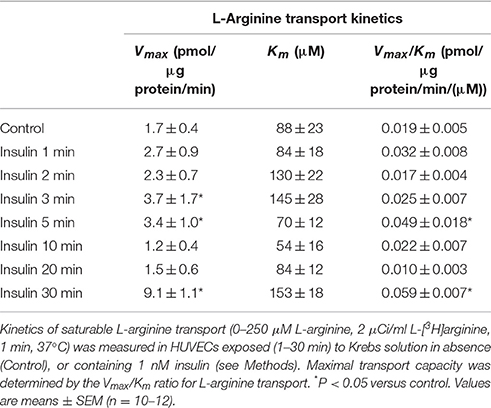
Table 2. Effect of insulin on L-arginine transport in HUVECs.
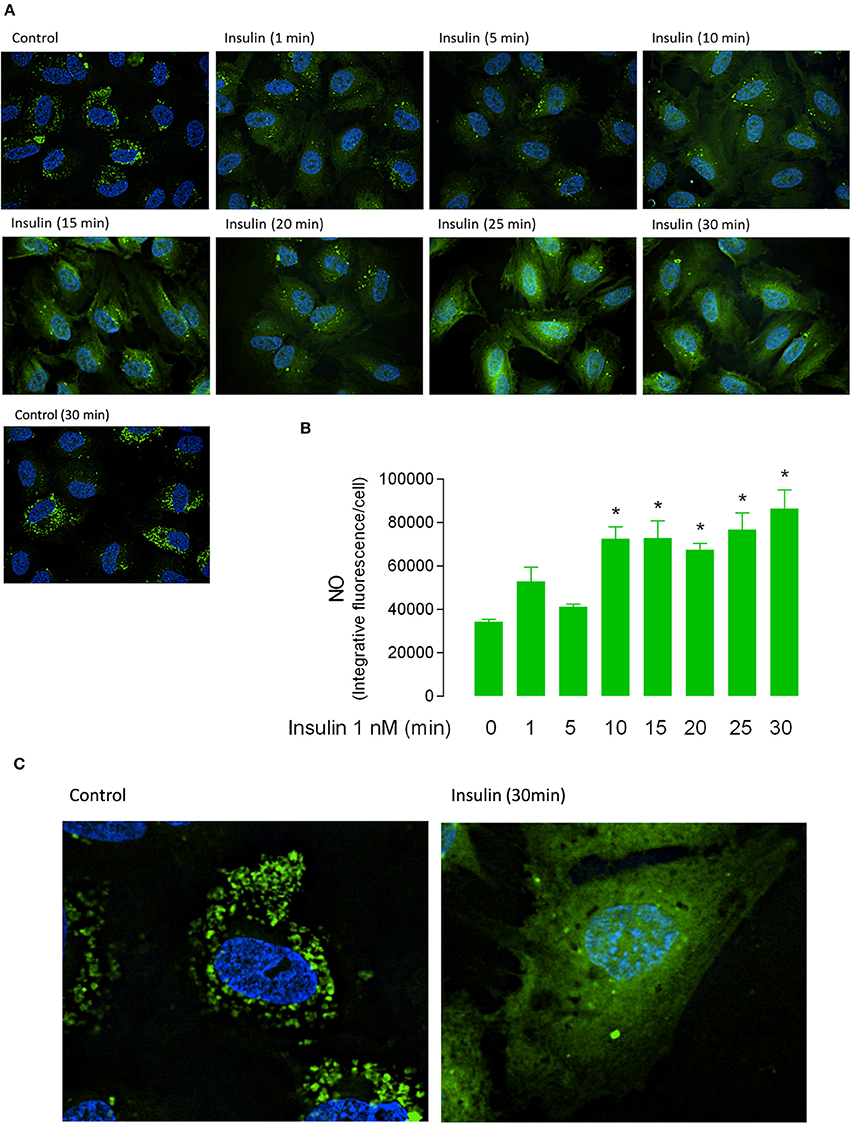
Figure 2. Short-term incubation with insulin increases the nitric oxide synthesis in fetal endothelium. Nitric oxide (NO) in HUVEC incubated in absence (Control 0, 30 min) or presence of 1 nM insulin (1–30 min). (A) Green fluorescence was observed through confocal microscopy in cells preloaded (30 min) with 4-amino-5-methylamino-2′,7′-difluorofluorescein (DAF-FM). (B) The integrative fluorescence per image was determined using Image J software and normalized by number of cells. (C) Magnification of fluorescence in control and insulin-treated cells, shown in (A). The images (A,C) are representative of triplicates from three different cell cultures.*P < 0.05 vs. values in the absence of insulin (n = 3).
Placental Relaxation Induced by Insulin
The perfusion pressure in isolated cotyledon of placenta decreased from 64 ± 5 mmHg to 42 ± 5 mmHg and 33 ± 2 mmHg using 0.1 and 1 nM of insulin, respectively (Figure 3A); without changes in the flow (not shown). In the same preparation, both H2O2(2.5 ± 0.4-fold) and U46619 (3.4 ± 0.5-fold) increased perfusion pressure, effects attenuated by preincubation with insulin (1 nM; Figures 3B,C). In chorionic veins preconstricted with U46619, insulin (10 nM) caused 29 ± 2% of relaxation after 30 min of treatment (Figure 4). Coincubation with wortmannin (Figure 4A) or genistein (Figure 4B) did not block insulin's vasodilator effect. Indeed, vessels exposed to tyrosine kinases inhibitors showed relaxation of 61 ± 10 and 52 ± 5% with wortmannin and genistein, respectively (Figure 4D). In the presence of insulin, maximal relaxation caused by wortmannin or genistein was further increased (73 ± 15 and 71 ± 9%, respectively; Figure 4D). The effect of insulin on relaxation caused in preconstricted chorionic vein was blocked by coincubation with iberiotoxin (Figure 4C) meanwhile the BKCa inhibitor induced a maximal relaxation of 21 ± 2% in absence of insulin (Figure 4D). Using a different protocol (pre-incubation of 30 min with insulin), insulin (10 nM) decreased by 49 ± 5% the constriction induced by 100 μM H2O2(Figure 5A). Reduced vasoconstriction caused by insulin was abolished by coincubation with tetraethylammonium (TEA, Figure 5A) or IbTx (Figure 5B), but not with wortmannin (Figure 5C). Preincubation with wortmannin potentiated the effect of insulin on H2O2 constriction in 40 ± 8% (Figure 5C). Insulin, TEA, IbTx and wortmannin decreased the constriction caused by H2O2by 53 ± 8, 60 ± 11, 55 ± 10, and 74 ± 15%, respectively. Moreover, the combination of insulin and wortmannin exhibited the highest inhibition on H2O2 constriction (93 ± 16%; Figure 5D). In addition, the maximal contractile response to U46619 was reduced 56 ± 4% following preincubation with insulin. Meanwhile, the half-maximal effective concentration (EC50) of U46619 was reduced from 81 ± 12 nM to 61 ± 6 nM in insulin-treated veins (Figure 6). However, when vessels were constricted with U46619, insulin–associated relaxation was unaltered by TEA or IbTx (Figures 6A,B), but was blocked by wortmannin (Figure 6C).
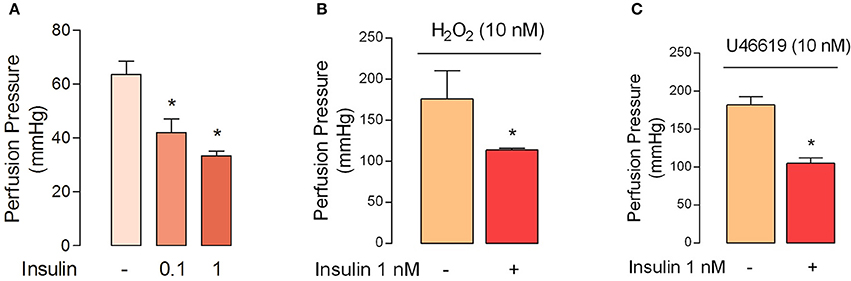
Figure 3. Insulin reduces the perfusion pressure in fetal-side circulation of isolated cotyledon from human placenta. (A) Perfusion pressure was measured in absence (−) or presence of insulin (30 min) in placentae, which have a mean basal perfusion pressure of 64 ± 5 mmHg. (B,C) Perfusion pressure was measured in placentae perfused (30 min) with Krebs solution in absence (−) or presence (+) of insulin and later perfused with H2O2 (B) or U46619 (C). *P < 0.05 vs. basal conditions in the absence of insulin. Values are mean ± S.E.M. (n = 5).
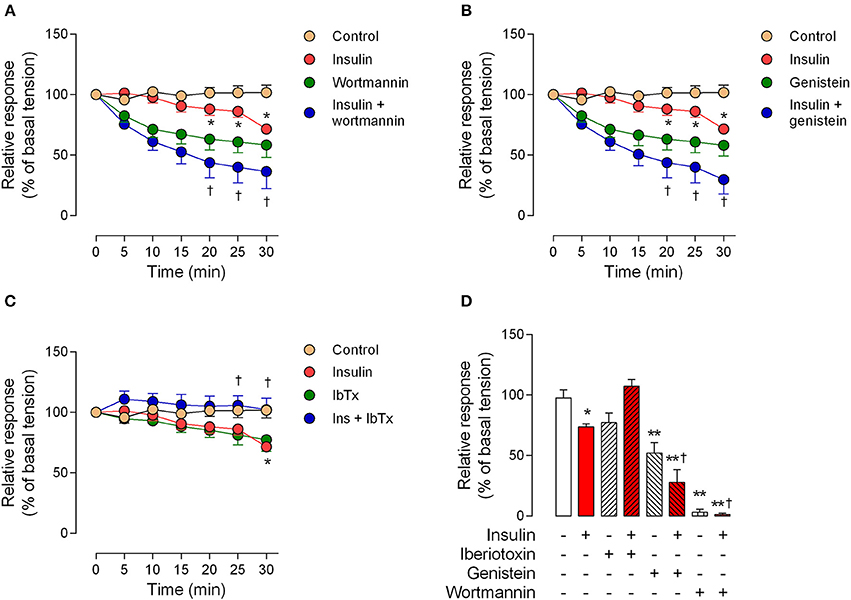
Figure 4. Relaxation induced by insulin is blocked by iberiotoxin. Response of human chorionic vein rings pre-constricted with U46619 and exposed to 10 nM insulin is showed in presence of 30 nM wortmannin (A), 50 μM genistein (B) or 100 nM iberiotoxin (C). Maximal responses are shown in (D), as a percentage fraction of the initial vessel response to KCl (see Methods). *P < 0.05 vs. non-treated (control) vessels. **P < 0.01 vs. non-treated (control) vessels. †P < 0.05 vs. values in insulin-treated vessels. Values are mean ± S.E.M. (n = 5–10).
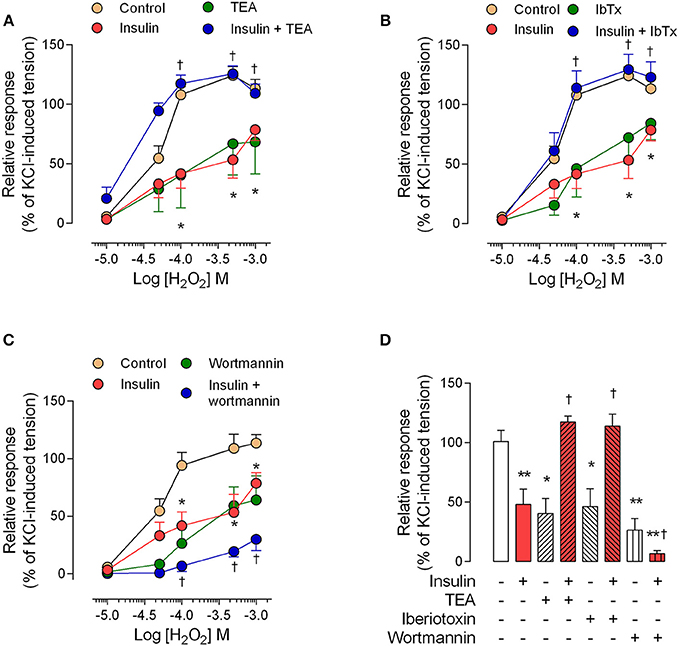
Figure 5. The effect of pre-incubation with insulin on H2O2 constriction is dependent on BKCa channels. Chorionic vein rings were pre-incubated (30 min, 5%CO2, 37°C) in absence (control) or presence of 10 nM insulin and/or 1 mM tetraethylammonium (TEA; A), 100 nM iberiotoxin (IbTx; B), or 30 nM wortmannin (C). After these treatments, vessels were exposed to hydrogen peroxide (H2O2). In (D) the effects are shown of treatments on maximal constriction induced by 100 μM (1 × 10−4 M) H2O2. *P < 0.05, **P < 0.01 vs. values in the absence of insulin. †P < 0.05 vs. values in insulin treated vessels. Values are mean ± S.E.M. (n = 5–7).
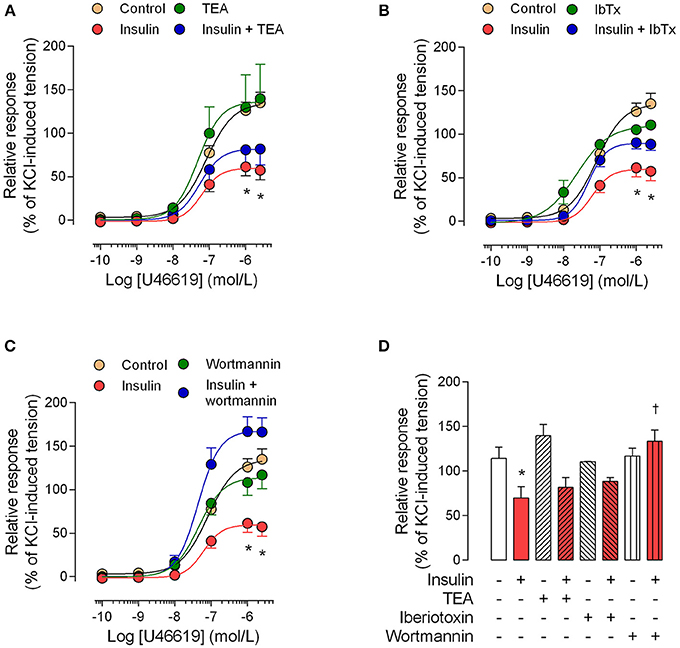
Figure 6. The effect of pre-incubation with insulin on U46619 constriction is dependent on PI3K. Chorionic vein rings were pre-incubated (30 min, 5%CO2, 37°C) in absence (control) or presence of 10 nM insulin and/or 1 mM tetraethylammonium (TEA; A), 100 nM iberiotoxin (IbTx; B) or 30 nM wortmannin (C). After these treatments, vessels were exposed to U46619. In (D), the effects are shown of treatments on maximal constriction induced by 1 μM (1 × 10−6 M) U46619. *P < 0.05 vs. values in the absence of insulin. †P < 0.05 vs. values in insulin-treated vessels. Values are mean ± S.E.M. (n = 5–7).
Discussion
The mechanism previously reported for vascular properties of insulin involves higher expression and activity of eNOS in HUVECs (Montagnani et al., 2002; González et al., 2004), human aortic coronary endothelial cells (HAECs; Federici et al., 2002) and bovine aortic endothelial cells (BAECs; Kuboki et al., 2000). The effect of long-term insulin incubation (several hours) on eNOS activity is known to be dependent on PI3K signaling (Montagnani et al., 2002; González et al., 2011). However, mechanisms underlying a rapid stimulation of vasodilatation induced by insulin (González et al., 2011) are still unclear, and have been associated with NO–mediated blood flow in healthy subjects (Scherrer et al., 1994; Steinberg et al., 1994; Lind et al., 2002). In this study, we demonstrated that insulin induces relaxation in pre-constricted vessels and attenuates constriction (when hormone is pre-incubated) in a mechanism mediated by BKCa channel activity and related to increased L-arginine transport and endothelial NO synthesis.
Mechanism of Relaxation Induced by Insulin
Insulin caused 42% relaxation in pre-constricted human umbilical vein (González et al., 2011). The EC50 of insulin was 1.8 ± 0.2 nM and the effect was abolished by preincubation with N-ethylmaleimide (NEM) and L-lysine, both competitive inhibitors of transport system y+ for cationic amino acid (González et al., 2011). Now, we confirmed that insulin induced relaxation in chorionic plate veins and in fetal-side of placental vascular bed, decreasing the vasoconstriction induced by U46619 and H2O2, stimuli that had previously been shown to have vasoconstrictor effects in chorionic plate arteries (Beckman and Koppenol, 1996; Hayward et al., 2013).
Relaxation induced by insulin in placental vessels was related to L-arginine/NO pathway in HUVECs. Previously, insulin has been noted to increase L-citrulline synthesis and L-arginine transport using concentrations between 0.1 and 10 nM in long-term (8 h) incubation assays, via a mechanism that involves high expression of hCAT-1 (Sobrevia et al., 1996; González et al., 2004). In isolated umbilical vein rings, insulin induced relaxation after 30 min of incubation, an effect disrupted by co-incubation with L-lysine, NEM or by endothelial denudation (González et al., 2011). This study confirms the effect of insulin in human placenta, and also suggests that this phenomenon is related to high NO synthesis, L-arginine transport and higher abundance of hCAT-1 in HUVECs. Regarding cell surface expression of hCAT-1, it has been determined that activation of protein kinase C (PKC) reduces cell surface expression of hCAT-1 and L-arginine transport in Xenopus laevis oocytes and U373MG glioblastoma cells (Rotmann et al., 2004a,b), reinforcing the idea that a rapid mobilization of hCAT-1 from plasma membrane to cytoplasm reduces L-arginine transport. Moreover, our work suggests that high Vmax of L-arginine transport mediated by insulin is associated with mobilization of hCAT-1 from cytoplasm to cell surface in HUVECs considering that insulin treatment results in a larger percentage of the total fluorescence from hCAT-1 expression on the surface of the cells. Further studies are necessary to determine the functional co-localization of hCAT-1 with relevant proteins for endothelial cell function like caveolin 1 or eNOS, as has been shown in porcine aortic endothelial cells (PAECs) (McDonald et al., 1997) or baby hamster kidney (BHK) cells (Lu and Silver, 2000).
One of the main mechanisms triggered by insulin is related to activation of PI3K in the human endothelium (González et al., 2004; Muniyappa et al., 2008). However, our results showed that inhibition of PI3K induces vessel relaxation; which was potentiated by insulin. Similarly, genistein induced relaxation in an endothelium-independent mechanism in rat aorta pre-constricted by sodium fluoride (NaF). In addition, when vessels were pre-treated with genistein, there was a decrease in constriction induced by U46619 (Je and Sohn, 2009) and an increase of eNOS phosphorylation in serine 1179 after 10–30 min of incubation via a protein kinase A (PKA)-dependent mechanism in BAEC (Liu et al., 2004). In the endothelial cell line ECV-304, the pre-incubation with genistein improved cell viability and reversed apoptosis induced by H2O2, which was associated with enhanced antioxidant capacity (Jin et al., 2015). On the other hand, the activity of protein kinase B/Akt plays a central role in the PI3K-dependent activation of eNOS catalyzing the phosphorylation of serine 1179 and serine 617, increasing the sensitivity of eNOS by Ca2+/calmodulin (CaM) complex (Tran et al., 2009). Previously, it has been shown that 100 nM insulin induces the phosphorylation of PKB/Akt after 2 or 10 min of treatment in HUVEC, increasing the eNOS activity in a mechanism dependent on insulin receptor substrate 1 (IRS1; Federici et al., 2004). With these evidences accounted for, our results show that inhibition of tyrosine kinases induces relaxation in pre-constricted placental vessels and, more importantly, the relaxation induced by insulin could be independent of PI3K pathway. It will still be important to explore the effects of insulin in PKB/Akt activity and CaM-dependent activation of eNOS, especially if the insulin-induced relaxation is associated with activity of K+ channels activated by Ca2+. In regard to this association, recently it has been shown that genistein, in combination with magnesium, induces relaxation in rat mesenteric arteries in a mechanism dependent on eNOS activity (blocked by L-NAME) and associated with high BKCa currents in rat mesenteric smooth muscle cells (Sun et al., 2015). These results suggest that genistein has a dual effect both in endothelial cells and VSMCs. In our experiments it is possible that the effects of genistein in the chorionic veins were not due to inhibition of protein kinases mainly, because we cannot discard the activation of BKCa by genistein directly in chorionic vein smooth muscle cells.
Role of Potassium Channels Activity
Using hippocampal neurons, O'Malley and Harvey (2004) showed that insulin (10 nM) increased (~3.8-fold) the mean channel activity (NfPo) of BKCa after 15 min incubation in the bath solution. Meanwhile, in similar experiments but via patch pipette solution, the effect was faster (2–8 min post-insulin). Authors also showed that insulin increased the mean open time (τo) of BKCa from ~0.76 ms (at 2–4 min) to ~2.01 ms (at 15–17 min). Similar effects were observed in HEK293 cells expressing hSlo (pore-forming α subunit of BKCa channel) and direct activation of BKCa channels through application of its selective channel opener, NS-1619 (O'Malley and Harvey, 2004). Similar to our results, this last study showed that the effect of insulin on BKCa activity is mediated by a mechanism independent of PI3K (but dependent of Ras/Raf/MEK/ERK pathway). Therefore, our study shows that the effects of insulin on placental vasculature are independent of PI3K activity, but mainly dependent of BKCa activity. We acknowledge that further experiments are required in order to elucidate underling intracellular pathway linked with this effect of insulin on BKCa channels, including potential participation of MAPK pathway.
Regarding the KCa expression and activity in human placenta, in 2006 Wareing et al. reported mRNA expression of BKCa in placental arteries and veins, showing high basal expression in arteries, suggesting a role of BKCa in fetoplacental relaxation induced by NO (Wareing et al., 2006). More recently, in chorionic plate artery smooth muscle cells (CPASMCs), the incubation with TEA and iberiotoxin demonstrated that BKCa are the main channels responsible for outward currents in CPASMCs (Brereton et al., 2013). Importantly, immunohistochemistry assays in placental tissue, have demonstrated a strong expression of BKCa in endothelium with similar localization of endothelial cell marker CD31 (Sand et al., 2006). In our study, we cannot discern if BKCa expression/activity is more important in endothelium or VSMCs, but the relaxation induced by TEA and iberiotoxin reveals a role of BKCa in the vascular tone regulation of placenta. In rabbit basilar artery, similar pre-incubation (30 min) with iberiotoxin and TEA reduced the vascular tone induced by sodium acetate (Cho et al., 2007). This result is similar to our finding about the reduction of vascular tone induced by H2O2; in both cases the mechanism of constriction is not via a receptor-mediated signaling pathway. The mechanism of vascular tone modulation by H2O2 in placenta is still unclear, but previous report of Mills showed a transient constriction induced by H2O2 in chorionic plate arteries, reversed by catalase (Mills et al., 2009). In umbilical artery, similar concentration (10–100 μM) of H2O2 enhanced the tension induced by prostanglandin F2α without change in sensitivity to calcium chloride (Okatani et al., 1997). The short half-life and rapid conversion of H2O2 to other reactive species, like hydroxyl radical (•OH) through Fenton reaction in presence of Fe2+ or Haber-Weiss reaction in presence of (MacFarlane et al., 2008), suggest a potential role of •OH in H2O2-induced constriction in placental vessels. At moment, we still cannot explain with direct evidence the effect of TEA or iberiotoxin on H2O2-induced constriction, but we speculate that the alteration in vascular homeostasis after 30 min of BKCa inhibition could reduce the constriction associated with oxidative stress. Although in our study we did not determine the changes in plasma membrane potential, is possible that a change of resting potential (both in endothelium and VSMCs) induced by iberiotoxin alters the vascular response to oxidative stress.
Related to the connection between the ex vivo and in vitro results, previously it has been demonstrated that HUVECs express Ca2+-activated potassium currents blocked by iberiotoxin (Wiecha et al., 1998; Watanapa et al., 2012). More importantly, 0.6 nM (100 μUI/ml) insulin increased the open-state probability (NPo) of BKCa after 3 min incubation in this cell type (Wiecha et al., 1998), in a similar fashion that 10 nM insulin increased the activity of BKCa in O'Malley and Harvey study. A different stimulus, quercetin, induces hyperpolarization, high concentration of intracellular Ca2+, cGMP synthesis and reduction of proliferation of HUVECs, and each of these effects are blocked when the cells are incubated with iberiotoxin (Kuhlmann et al., 2005). In addition, HUVECs treated with plasma samples from preeclamptic pregnancies exhibit a higher fraction of cells expressing outward currents associated with KCa channels, showing a compensatory mechanism attributed to some factors secreted in preeclampsia (Watanapa et al., 2012). These evidences allow us to propose that the insulin signaling induces the activation of KCa channels, changing the plasma membrane polarity for activation of hCAT-1 reflected in higher Vmax for L-arginine transport and, finally, higher NO synthesis. In this regard, a study published by Kavanaugh showed that the influx of L-arginine is increased by membrane hyperpolarization in Xenopus laevis oocytes expressing CAT-1 (Kavanaugh, 1993). Oppositely, the depolarization induced by increased extracellular concentration of K+ reduced the L-arginine transport in HUVECs (Sobrevia et al., 1995). Also in Xenopus laevis oocytes expressing hCAT-1, the incubation (6 h) with high concentration of K+ reduced the intracellular accumulation of L-[3H]arginine (Rotmann et al., 2004a). These findings show the dependency between hCAT-1 activity and voltage of the plasma membrane, but further studies are necessary to establish direct evidence of regulation of hCAT-1 activity through membrane hyperpolarization in placental endothelial cells.
Also, it is important to note that previously it has been shown that insulin evokes hyperpolarization (from −65.5 ± 0.4 to −82.3 ± 0.4 mV) and high intracellular concentration of Ca2+ (from 40 ± 3 nM to 372 ± 29 nM) in HUVECs (González et al., 2004). Although the mechanism of intracellular Ca2+ regulation by insulin in endothelium still is not clear, other agonists or stimuli that activate eNOS have been studied. VEGF-A and shear stress increase the activity of eNOS and NO synthesis in a mechanism dependent of enhancement of Ca2+ (Devika and Jaffar Ali, 2013). Importantly, Anaya et al., showed that the Ca2+ mobilization and NO availability (detected with DAF) were reduced in intact endothelium from umbilical vein isolated of GDM samples, without changes in the protein abundance of eNOS. Additionally, in HUVECs isolated from GDM, the response of Ca2+ mobilization to ATP is lower than control cells (Anaya et al., 2015). From these data, we suggest that the effects of insulin on L-arginine/NO pathway in HUVECs and BKCa-dependent relaxation in chorionic vein, reported in this study, could be significantly altered in GDM due to reduced capacity of endothelial cells for increased Ca2+ mobilization in response to some agonists.
In conclusion, we found that insulin induces a rapid relaxation in placental vascular bed through a mechanism associated with high activity of BKCa channels and L-arginine/NO pathways in endothelial cells.
Author Contributions
LC, AS, CV, DG, PC, EB, AH: Experimental work and first analysis of results. This article is a result of different undergraduate projects executed by these authors. SR: Experimental work, supervision and technical support in Vascular Physiology Laboratory. MC: Collection of informed consent and verification of clinical data of pregnants. EP: Acquisition of images from confocal microscopy. CR, JV: Collaboration with fluorescence results analysis. VG: Collaboration with discussion about the clinical implications of results. LZ: Collaboration in discussion related with calcium-activated potassium channels. CE, LS: Collaboration with discussion about the mollecular mechanisms involved. MW: Collaboration with the execution and analysis of wire myography experiments. MG: Supervision of experimental work, analysis of results, coordination and main discussion of the article.
Conflict of Interest Statement
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
The reviewer LM and handling Editor declared their shared affiliation, and the handling Editor states that the process nevertheless met the standards of a fair and objective review.
Acknowledgments
Authors thank the research staff at Vascular Physiology Laboratory and Department of Physiology from the Universidad de Concepción. Authors also thank the personnel of the Obstetrics and Gynaecology Department of the Hospital Guillermo Grant Benavente from Concepción labor ward for the supply of placentas. We appreciate the collaboration of Mr. Iann Scott as english grammar editor and Ms. Joanna Tereszczuk (CMA-Biobío) by image analysis of Figure 1. This study was supported by Fondo Nacional de Desarrollo Científico y Tecnológico (FONDECYT 11100192, Chile) and Vicerrectoría de Investigación y Desarrollo Universidad de Concepción (VRID-Asociativo 213.A84.014-1.0, Chile). CE is supported by Fondecyt 1140586 (Chile). LS is supported by Fondecyt 1150377. This article is dedicated to the memory of Professor Juan Carlos Vera.
Abbreviations
BAECs, Bovine aortic endothelial cells; BHK, Baby hamster kidney cells; BKCa, Large conductance calcium-activated potassium channels; eNOS, Endothelial nitric oxide synthase; GDM, Gestational diabetes mellitus; HAECs, Human aortic endothelial cells; hCAT-1, Human cationic amino acid transporter 1; HUVECs, Human umbilical vein endothelial cells; IbTx, Iberiotoxin; IUGR, Intrauterine growth restriction; KCa, Calcium-activated potassium channels; MAPK, Mitogen-activated protein kinases; NEM, N-ethylmaleimide; NO, Nitric oxide; PAECs, Porcine aortic endothelial cells; PE, Preeclampsia; PI3K, Phosphatidylinositol 3-phosphate; PKC, Protein kinase C; RNS, Reactive nitrogen species; ROS, Reactive oxygen species; SLC7A1, Solute carrier family 7 type 1; TEA, Tetraethylammonium; VSMCs, Vascular smooth muscle cells.
References
Acevedo, C. G., Rojas, S., and Bravo, I. (1999). L-arginine transport at the fetal side of human placenta: effect of aspirin in pregnancy. Exp. Physiol. 84, 1127–1136. doi: 10.1111/j.1469-445X.1999.01875.x
Acevedo, C. G., Rojas, S., Ramirez, M., and Bravo, I. (1995). Transport and metabolism of adenosine in the perfused human placenta. Placenta 16, 611–622. doi: 10.1016/0143-4004(95)90030-6
Anaya, H. A., Yi, F.-X., Boeldt, D. S., Krupp, J., Grummer, M. A., Shah, D. M., et al. (2015). Changes in Ca2+ signaling and nitric oxide output by human umbilical vein endothelium in diabetic and gestational diabetic pregnancies. Biol. Reprod. 93, 60. doi: 10.1095/biolreprod.115.128645
Beckman, J. S., and Koppenol, W. H. (1996). Nitric oxide, superoxide, and peroxynitrite: the good, the bad, and ugly. Am. J. Physiol. 271, C1424–C1437.
Bernardi, F., Guolo, F., Bortolin, T., Petronilho, F., and Dal-Pizzol, F. (2008). Oxidative stress and inflammatory markers in normal pregnancy and preeclampsia. J. Obstet. Gynaecol. Res. 34, 948–951. doi: 10.1111/j.1447-0756.2008.00803.x
Brereton, M. F., Wareing, M., Jones, R. L., and Greenwood, S. L. (2013). Characterization of K+ channels in human fetoplacental vascular smooth muscle cells. PLoS ONE 8:e57451. doi: 10.1371/journal.pone.0057451
Catarino, C., Santos-Silva, A., Belo, L., Rocha-Pereira, P., Rocha, S., Patrício, B., et al. (2012). Inflammatory disturbances in preeclampsia: relationship between maternal and umbilical cord blood. J. Pregnancy. 2012:684384. doi: 10.1155/2012/684384
Cho, Y. E., Ahn, D. S., Kim, Y. H., Taggart, M. J., and Lee, Y. H. (2007). Changes in stretch-induced tone induced by intracellular acidosis in rabbit basilar artery: effects on BKCa channel activity. Vascul. Pharmacol. 47, 74–82. doi: 10.1016/j.vph.2007.04.006
Coughlan, M. T., Vervaart, P. P., Permezel, M., Georgiou, H. M., and Rice, G. E. (2004). Altered placental oxidative stress status in gestational diabetes mellitus. Placenta 25, 78–84. doi: 10.1016/S0143-4004(03)00183-8
Devika, N. T., and Jaffar Ali, B. M. (2013). Analysing calcium dependent and independent regulation of eNOS in endothelium triggered by extracellular signalling events. Mol. Biosyst. 9, 2653–2664. doi: 10.1039/c3mb70258h
Durand, M. J., and Gutterman, D. D. (2013). Diversity in mechanisms of endothelium-dependent vasodilation in health and disease. Microcirculation 20, 239–247. doi: 10.1111/micc.12040
Federici, M., Menghini, R., Mauriello, A., Hribal, M. L., Ferrelli, F., Lauro, D., et al. (2002). Insulin-dependent activation of endothelial nitric oxide synthase is impaired by O-linked glycosylation modification of signaling proteins in human coronary endothelial cells. Circulation 106, 466–472. doi: 10.1161/01.CIR.0000023043.02648.51
Federici, M., Pandolfi, A., De Filippis, E. A., Pellegrini, G., Menghini, R., Lauro, D., et al. (2004). G972R IRS-1 variant impairs insulin regulation of endothelial nitric oxide synthase in cultured human endothelial cells. Circulation 109, 399–405. doi: 10.1161/01.CIR.0000109498.77895.6F
Figueroa, X. F., and Duling, B. R. (2009). Gap junctions in the control of vascular function. Antioxid. Redox. Signal. 11, 251–266. doi: 10.1089/ars.2008.2117
González, M., Flores, C., Pearson, J. D., Casanello, P., and Sobrevia, L. (2004). Cell signalling-mediating insulin increase of mRNA expression for cationic amino acid transporters-1 and -2 and membrane hyperpolarization in human umbilical vein endothelial cells. Pflugers. Arch. 448, 383–394. doi: 10.1007/s00424-004-1261-x
González, M., Gallardo, V., Rodríguez, N., Salomón, C., Westermeier, F., Guzmán-Gutiérrez, E., et al. (2011). Insulin-stimulated L-arginine transport requires SLC7A1 gene expression and is associated with human umbilical vein relaxation. J. Cell. Physiol. 226, 2916–2924. doi: 10.1002/jcp.22635
González, M., Rojas, S., Ávila, P., Cabrera, L., Villalobos, R., Palma, C., et al. (2015). Insulin reverses D-glucose-increased nitric oxide and reactive oxygen species generation in human umbilical vein endothelial cells. PLoS ONE 10:e0122398. doi: 10.1371/journal.pone.0122398
Hayward, C. E., Higgins, L., Cowley, E. J., Greenwood, S. L., Mills, T. A., Sibley, C. P., et al. (2013). Chorionic plate arterial function is altered in maternal obesity. Placenta 34, 281–287. doi: 10.1016/j.placenta.2013.01.001
Je, H. D., and Sohn, U. D. (2009). Inhibitory effect of genistein on agonist-induced modulation of vascular contractility. Mol. Cells 27, 191–198. doi: 10.1007/s10059-009-0052-9.
Jin, L., Zhao, X., Qin, Y., Zhu, W., Zhang, W., Liu, A., et al. (2015). Soy isoflavones protect against H2O2-induced injury in human umbilical vein endothelial cells. Mol. Med. Rep. 12, 4476–4482. doi: 10.3892/mmr.2015.3949
Kavanaugh, M. P. (1993). Voltage dependence of facilitated arginine flux mediated by the system y+ basic amino acid transporter. Biochemistry 32, 5781–5785. doi: 10.1021/bi00073a009
Kerr, P. M., Tam, R., Narang, D., Potts, K., McMillan, D., McMillan, K., et al. (2012). Endothelial calcium-activated potassium channels as therapeutic targets to enhance availability of nitric oxide. Can. J. Physiol. Pharmacol. 90, 739–752. doi: 10.1139/y2012-075
Klandorf, H., and Van Dyke, K. (2012). “Oxidative and nitrosative stresses: their role in health and disease in man and birds,” in Oxidative Stress-Molecular Mechanisms and Biological Effects, eds V. Lushchak and H. Semchyshyn (Croatia: InTech), 47–60.
Kuboki, K., Jiang, Z. Y., Takahara, N., Ha, S. W., Igarashi, M., Yamauchi, T., et al. (2000). Regulation of endothelial constitutive nitric oxide synthase gene expression in endothelial cells and in vivo: a specific vascular action of insulin. Circulation 101, 676–681. doi: 10.1161/01.CIR.101.6.676
Kuhlmann, C., Schaefer, C. A., Kosok, C., Abdallah, Y., Walther, S., Lüdders, D. W., et al. (2005). Quercetin-induced induction of the NO/cGMP pathway depends on Ca2+-activated K+ channel-induced hyperpolarization-mediated Ca2+-entry into cultured human endothelial cells. Planta. Med. 71, 520–524. doi: 10.1055/s-2005-864152
Lind, L., Fugmann, A., Millgård, J., Berne, C., and Lithell, H. (2002). Insulin-mediated vasodilatation, but not glucose uptake or endothelium-mediated vasodilatation, is enhanced in young females compared with males. Clin. Sci. 102, 241–246. doi: 10.1042/cs1020241
Liu, D., Homan, L. L., and Dillon, J. S. (2004). Genistein acutely stimulates nitric oxide synthesis in vascular endothelial cells by a cyclic adenosine 5′-monophosphate-dependent mechanism. Endocrinology 145, 5532–5539. doi: 10.1210/en.2004-0102
Lu, X., and Silver, J. (2000). Ecotropic murine leukemia virus receptor is physically associated with caveolin and membrane rafts. Virology 276, 251–258. doi: 10.1006/viro.2000.0555
Luedders, D. W., Muenz, B. M., Li, F., Rueckleben, S., Tillmanns, H., Waldecker, B., et al. (2006). Role of cGMP in sildenafil-induced activation of endothelial Ca2+-activated K+ channels. J. Cardiovasc. Pharmacol. 47, 365–370.
MacFarlane, P. M., Wilkerson, J. E., Lovett-Barr, M. R., and Mitchell, G. S. (2008). Reactive oxygen species and respiratory plasticity following intermittent hypoxia. Respir. Physiol. Neurobiol. 164, 263–271. doi: 10.1016/j.resp.2008.07.008
McDonald, K. K., Zharikov, S., Block, E. R., and Kilberg, M. S. (1997). A caveolar complex between the cationic amino acid transporter 1 and endothelial nitric-oxide synthase may explain the “arginine paradox”. J. Biol. Chem. 272, 31213–31216. doi: 10.1074/jbc.272.50.31213
Mills, T. A., Wareing, M., Shennan, A. H., Poston, L., Baker, P. N., and Greenwood, S. L. (2009). Acute and chronic modulation of placental chorionic plate artery reactivity by reactive oxygen species. Free. Radic. Biol. Med. 47, 159–166. doi: 10.1016/j.freeradbiomed.2009.04.019
Montagnani, M., Ravichandran, L. V., Chen, H., Esposito, D. L., and Quon, M. J. (2002). Insulin receptor substrate-1 and phosphoinositide-dependent kinase-1 are required for insulin-stimulated production of nitric oxide in endothelial cells. Mol. Endocrinol. 16, 1931–1942. doi: 10.1210/me.2002-0074
Muniyappa, R., Iantorno, M., and Quon, M. J. (2008). An integrated view of insulin resistance and endothelial dysfunction. Endocrinol. Metab. Clin. North. Am. 37, 685–711. doi: 10.1016/j.ecl.2008.06.001
Okatani, Y., Watanabe, K., Wakatsuki, A., and Sagara, Y. (1997). Vasospastic action of hydrogen peroxide in human umbilical artery: relation to protein kinase C and calcium influx. Acta Obstet. Gynecol. Scand. 76, 897–902. doi: 10.3109/00016349709034898
O'Malley, D., and Harvey, J. (2004). Insulin activates native and recombinant large conductance Ca(2+)-activated potassium channels via a mitogen-activated protein kinase-dependent process. Mol. Pharmacol. 65, 1352–1363. doi: 10.1124/mol.65.6.1352
Penfold, P., Drury, L., Simmonds, R., and Hytten, F. E. (1981). Studies of a single placental cotyledon in vitro: I. The preparation and its viability. Placenta 2, 149–154. doi: 10.1016/S0143-4004(81)80018-5
Rotmann, A., Closs, E. I., Liewald, J. F., and Nawrath, H. (2004a). Intracellular accumulation of L-Arg, kinetics of transport, and potassium leak conductance in oocytes from Xenopus laevis expressing hCAT-1, hCAT-2A, and hCAT-2B. Biochim Biophys. Acta 1660, 138–143. doi: 10.1016/j.bbamem.2003.11.009
Rotmann, A., Strand, D., Martiné, U., and Closs, E. I. (2004b). Protein kinase C activation promotes the internalization of the human cationic amino acid transporter hCAT-1. A new regulatory mechanism for hCAT-1 activity. J. Biol. Chem. 279, 54185–54192. doi: 10.1074/jbc.M409556200
Sand, A., Andersson, E., and Fried, G. (2006). Nitric oxide donors mediate vasodilation in human placental arteries partly through a direct effect on potassium channels. Placenta 27, 181–190. doi: 10.1016/j.placenta.2004.12.013
Sandow, S. L., and Grayson, T. H. (2009). Limits of isolation and culture: intact vascular endothelium and BKCa. Am. J. Physiol. Heart. Circ. Physiol. 297, H1–H7. doi: 10.1152/ajpheart.00042.2009
Scherrer, U., Randin, D., Vollenweider, P., Vollenweider, L., and Nicod, P. (1994). Nitric oxide release accounts for insulin's vascular effects in humans. J. Clin. Invest. 94, 2511–2515. doi: 10.1172/JCI117621
Sobrevia, L., Cesare, P., Yudilevich, D. L., and Mann, G. E. (1995). Diabetes-induced activation of system y+ and nitric oxide synthase in human endothelial cells: association with membrane hyperpolarization. J. Physiol. 489, 183–192. doi: 10.1113/jphysiol.1995.sp021040
Sobrevia, L., Nadal, A., Yudilevich, D. L., and Mann, G. E. (1996). Activation of L-arginine transport (system y+) and nitric oxide synthase by elevated glucose and insulin in human endothelial cells. J. Physiol. 490, 775–781. doi: 10.1113/jphysiol.1996.sp021185
Steinberg, H. O., Brechtel, G., Johnson, A., Fineberg, N., and Baron, A. D. (1994). Insulin-mediated skeletal muscle vasodilation is nitric oxide dependent. A novel action of insulin to increase nitric oxide release. J. Clin. Invest. 94, 1172–1179. doi: 10.1172/JCI117433
Sun, L., Hou, Y., Zhao, T., Zhou, S., Wang, X., Zhang, L., et al. (2015). A combination of genistein and magnesium enhances the vasodilatory effect via an eNOS pathway and BKCa current amplification. Can. J. Physiol. Pharmacol. 93, 215–221. doi: 10.1139/cjpp-2014-0306
Takagi, Y., Nikaido, T., Toki, T., Kita, N., Kanai, M., Ashida, T., et al. (2004). Levels of oxidative stress and redox-related molecules in the placenta in preeclampsia and fetal growth restriction. Virchows. Arch. 444, 49–55. doi: 10.1007/s00428-003-0903-2
Tran, Q. K., Leonard, J., Black, D. J., Nadeau, O. W., Boulatnikov, I. G., and Persechini, A. (2009). Effects of combined phosphorylation at Ser-617 and Ser-1179 in endothelial nitric-oxide synthase on EC50(Ca2+) values for calmodulin binding and enzyme activation. J. Biol. Chem. 284, 11892–11899. doi: 10.1074/jbc.M806205200
Wareing, M. (2014). Oxygen sensitivity, potassium channels, and regulation of placental vascular tone. Microcirculation 21, 58–66. doi: 10.1111/micc.12069
Wareing, M., Bai, X., Seghier, F., Turner, C. M., Greenwood, S. L., Baker, P. N., et al. (2006). Expression and function of potassium channels in the human placental vasculature. Am. J. Physiol. Regul. Integr. Comp. Physiol. 291, R437–R446. doi: 10.1152/ajpregu.00040.2006
Watanapa, W. B., Theerathananon, W., Akarasereenont, P., and Techatraisak, K. (2012). Effects of preeclamptic plasma on potassium currents of human umbilical vein endothelial cells. Reprod. Sci. 19, 391–399. doi: 10.1177/1933719111424434
Keywords: insulin, L-arginine, nitric oxide, hCAT-1, BKCa channels, placenta
Citation: Cabrera L, Saavedra A, Rojas S, Cid M, Valenzuela C, Gallegos D, Careaga P, Basualto E, Haensgen A, Peña E, Rivas C, Vera JC, Gallardo V, Zúñiga L, Escudero C, Sobrevia L, Wareing M and González M (2016) Insulin Induces Relaxation and Decreases Hydrogen Peroxide-Induced Vasoconstriction in Human Placental Vascular Bed in a Mechanism Mediated by Calcium-Activated Potassium Channels and L-Arginine/Nitric Oxide Pathways. Front. Physiol. 7:529. doi: 10.3389/fphys.2016.00529
Received: 19 April 2016; Accepted: 25 October 2016;
Published: 22 November 2016.
Edited by:
Michael A. Hill, University of Missouri, USAReviewed by:
Luis A. Martinez-Lemus, University of Missouri, USACynthia J. Meininger, Texas A&M University, USA
Copyright © 2016 Cabrera, Saavedra, Rojas, Cid, Valenzuela, Gallegos, Careaga, Basualto, Haensgen, Peña, Rivas, Vera, Gallardo, Zúñiga, Escudero, Sobrevia, Wareing and González. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) or licensor are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Marcelo González, bWdvbnphbGV6b0B1ZGVjLmNs