 Sergi Valverde1,2,3*
Sergi Valverde1,2,3*- 1ICREA-Complex Systems Lab, Universitat Pompeu Fabra, Barcelona, Spain
- 2Institute of Evolutionary Biology (UPF-CSIC), Barcelona, Spain
- 3European Centre for Living Technology, University Ca' Foscari, Venezia, Italy
The presence of modular organization is a common property of a wide range of complex systems, from cellular or brain networks to technological graphs. Modularity allows some degree of segregation between different parts of the network and has been suggested to be a prerequisite for the evolvability of biological systems. In technology, modularity defines a clear division of tasks and it is an explicit design target. However, many natural and artificial systems experience a breakdown in their modular pattern of connections, which has been associated with failures in hub nodes or the activation of global stress responses. In spite of its importance, no general theory of the breakdown of modularity and its implications has been advanced yet. Here we propose a new, simple model of network landscape where it is possible to exhaustively characterize the breakdown of modularity in a well-defined way. Specifically, by considering the space of minimal Boolean feed-forward networks implementing the 256 Boolean functions with 3 inputs, we were able to relate functional characteristics with the breakdown of modularity. We found that evolution cannot reach maximally modular networks under the presence of functional and cost constraints, implying the breakdown of modularity is an adaptive feature.
1. Introduction
Complex networks pervade the evolution and organization of a wide range of systems, from cellular or brain webs to technological graphs. Their structure has important consequences for their stability, resilience and fragility. Some particular properties of these networks are very common, such as the presence of modular organization (Bonner, 1988; Hartwell et al., 1999; Ravasz et al., 2002; Schlosser and Wagner, 2004). In modular webs, different subsets of nodes display a higher integration among them than with the rest of the system. This feature allows some degree of segregation between different parts of the network and has been suggested to be a prerequisite for the evolvability of biological webs (Gerhart and Kirschner, 1997). Within the context of technological evolution, modular structures have been often proposed as a target for engineering design.
Modules are also expected to play a key role in providing a source of specialization, while their proper interconnection guarantee integration at the system-level scale. Both are needed in order to sustain proper functionality and we need to understand both how modules are generated and how their disconnection leads to functional decay. An illustrative example is provided by brain network topology or the so called connectome (Sporns, 2010). Connectomics has been a major breakthrough in pushing forward a new approach to brain disease where both brain areas and their connectivity patterns become integrated in a single picture. Under this view, neurological disorders including Alzheimer's disease or schizophrenia to challenged healthy cognition, such as in sleep or awareness, can be understood in terms of faulty intermodule communication (David, 1994; Alexander-Bloch et al., 2010; Meunier et al., 2010; Bashan et al., 2012; Godwin et al., 2015). This failure can lead to the so called breakdown of modularity (BM) first proposed in Valverde and Solé (2007). It involves a transition from high modularity to low modularity. Similar patterns can be found in other areas, but no general theory of this phenomenon and its implications has been advanced yet.
In spite of its importance, BM has received little attention and it is not well-understood. One important reason of this is connected with the difficulties associated to understanding the mapping between structure (genotype) and function (phenotype) in evolved networks. This is specially difficult when dealing with a property as modular structure, and the need for understanding how and when modular networks are expected to evolve and how optimality is tied to modular architecture. For example, it has been suggested that networks evolved under “modularly varying goals” must be modular (Kashtan and Alon, 2005). Specifically, computational experiments showed that optimal networks are non-modular whenever the goal was kept fixed or under randomly varying goals (with no common subgoals). However, Clune and co-workers have shown that modular networks evolve even in the presence of fixed and modular input-output mappings (Clune et al., 2013). Here modular patterns would be the byproduct of a cost-dependent selection process. More generally, when dealing with evolving networks, an important question is the role played by modular structures in enhancing robust functionalities and how modular structures are associated to evolvable designs. In other words: is the landscape of modularity associated to Boolean functions smooth? Are optimal modular solutions always tied to efficient functions and evolvable architectures?
In order to address these limitations, a simple case study that can be systematically explored would be desirable. Here we propose such a toy model of network landscapes where it is possible to exhaustively characterize BM in a well-defined way. In this context, it is worth noting that BM seems to be a common feature of computational systems (Valverde and Solé, 2007; van den Berg et al., 2012). Because computation-related networks can be seen as instances of functional Boolean processes performed on well-defined circuits, a minimal case study can help to gain insight into the role played by modular architecture. Specifically, we consider the set of minimal Boolean feed-forward networks implementing the 28 = 256 Boolean functions fμ with 3 input variables, i. e., the mapping
where Σ = {0, 1} is the Boolean state space. Such kind of Boolean representation has been widely used in the study of evolved networks and in different contexts, including cellular circuits (Macia and Solé, 2009) or pattern-forming genetic circuits (Munteanu and Solé, 2008). A systematic exploration requires necessarily a limitation of the combinatorial space to be analyzed. However, relevant computational spaces and specific cases can be observed even in the simplest networks (Gates and Rocha, 2016). Our analysis suggests that the optimization of specific input-output mappings is not always compatible with highly modular structures and how the BM might be an adaptive feature.
2. Feed-Forward Boolean Networks
The model used here is based on Boolean logic (Wegener, 1987), which has been used in the modeling and analysis of the flow of information in natural and artificial systems, such as gene regulatory networks (Kauffman, 1969; Shmulevich et al., 2005). A Boolean function can be represented using a truth table, functional forms and networks and it is worth noting that, despite the Boolean picture is a necessarily simple, neural and genetic networks display nonlinear functional responses that ultimately involve an almost all-or-none behavior. The interactions between these representations reveal the presence of functional constraints in the organization of complex systems (see below). Our truth tables give the value for the function fμ(a, b, c) ∈ {0, 1} for each possible combination of the inputs a, b, and c. The function is identified by its designation number
that is, the binary sequence of all function values (see Figure 1A). We can achieve more readable (but ambiguous) expressions using functional forms. The full disjunctive normal form (Figure 1A, top) is the sum of elementary products (terms) corresponding to input combinations on which fμ is true (minterms). For example, the minterm abc represents the combination 111, ab¬c represents 110 and so on.
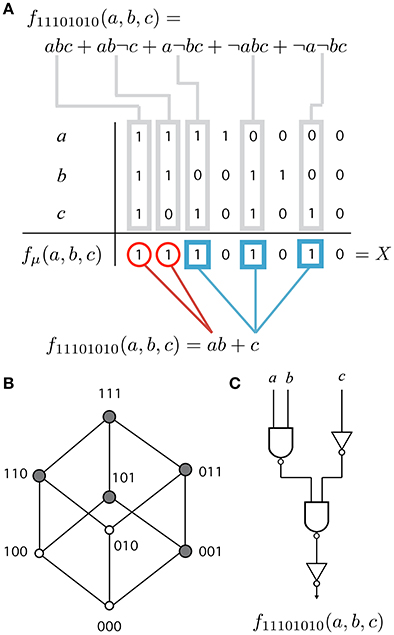
Figure 1. Different representations of a Boolean function. (A) Each function has ν = 3 variables named a, b and c. The truth table specifies the value fμ(a, b, c) ∈ {0, 1} for each of the 23 possible combinations of 3 binary inputs (columns). Combinations corresponding to fμ = 1 are called minterms. A function is identified by its designation number X = 11101010 (last row). More readable representations are the full (top) and minimized (bottom) disjunctive normal forms. (B) Hypercube consists of the set of terms (nodes), minterms (gray nodes) and the edges connecting the closest terms in the function. (C) A feed-forward Boolean network (FFBN) is a directed network made up of logic gates and wires that implement the function. The figure shows the FFBN of minimal cost.
A common network representation of a Boolean function is the simple feed-forward Boolean network (FFBN) without no feedback loops. We focus on a subset of the function space involving all the FFBNs that compute single-output Boolean functions with ν = 3 input variables and one output. The FFBN is a directed graph in which all the nodes carry the labels of negative-AND (NAND) gates while input nodes carry the labels of input variables.
Formally, a directed network G = (V, E) consists of a set of nodes vi ∈ V and a set of edges (vi, vj) ∈ E. The adjacency matrix A = [Aij] has elements such as Aij = 1 if there is link (i, j) ∈ E and Aij = 0 otherwise. The size of the network is the number of nodes N = |V| (logic gates) that it contains. The number of links is the sum,
In the following, we will study the undirected version of the FFBN. The network is undirected if for each edge (vi, vj) ∈ E there is another edge (vj, vi) ∈ E. An undirected network has m = L/2 edges. We also define the degree as the number of edges attached to the vi node. The average degree of the network
measures the overall connectivity in the system and it is one of the main network parameters.
We will also be interested in measuring the flow of information in the network. The path length is a measure of the distance between nodes in the network. The length l of a path is the number of edges traversed along the path. Let's define as the number of paths of length l that relate any pair of nodes vi and vj. Among all the alternative paths, we choose the path of minimal distance (or geodesic path), which defines the shortest path distance d(i, j) or the smallest value of l such that . The average path length of a graph (or network diameter) provides a measure of network efficiency and it is defined as follows:
where the normalization term discounts the influence of nodes on themselves.
3. Functional Modularity
The main goal of this paper is to characterize the landscape of Boolean functions associated to our system, and specifically the modularity of neighboring functions and how is the modularity of the circuit implementing each function with its one-bit neighbors. Given a decomposition of the network into a set of subgraphs Ci, the degree of modularity Q associated to this partition can be measured as follows (Girvan and Newman, 2002):
where is the number of connections attached to the i-th node (or node degree), Ci is the partition the i − th node belongs to and δ(x) = 1 is x >= 1 and δ(x) = 0 otherwise. Here, we use the Louvain method for community detection in order to find the optimal partition of the FFBN that maximizes the modularity value (Blondel et al., 2008).
Our hypothesis is that functional requirements constrain structural modularity, that is, evolution cannot reach maximally modular networks under the presence of functional constraints. In this case, we can define a functional (or phenotype) modularity or upper bound for the network (or genotype) modularity (2). However, several genotypes (FFBNs and functional forms) can be found for the same phenotype. When there is representation ambiguity, we often prefer shorter or minimized forms among all the alternatives. For example, a shorter functional form (Figure 1A, bottom) divides the support of fμ into groups (red circles and blue squares) that exploit functional symmetries (Quine, 1952; McCluskey, 1956).
Engineers are often concerned with the problem of obtaining the most economical design for electronic circuits. This is particularly relevant in the design of hardware systems. For example, the cost of VLSI (very large-scale integrated) circuits is determined by many factors including the chip area (Mead and Conway, 1980). When many of these factors are fixed, the minimization of cost can be solved by constructing networks which minimize the number of gates and the number of connections (Hellerman, 1963). We define functional modularity Q(X) as the modularity of the FFBN with minimal cost L + N or sum of number of logical gates and wires (see Figure 1C). This is equivalent to the minimization of Boolean circuits, which is a hard problem (in general) and there is no simple way to obtain the optimal solution (Wegener, 1987), although good approximate solutions exist (like “Espresso” Rudell and Sangiovanni-Vincentelli, 1987). Following (Hellerman, 1963), we perform an exhaustive search to obtain the optimal solution for each of the 256 Boolean functions of 3 variables (see SM for a detailed listing of the functions and their minimal FFBNs).
4. Phenotype Network
In order to uncover the relationship between modularity and functional requirements, we will make use of the concept of phenotypic network. A phenotypic network is a graph whose nodes represent (in our case) Boolean functions and where two functions are connected if they differ in only one minterm of the full disjunctive normal form. In this metagraph, each node maps a function onto its genotype (a FFBN). The notion of phenotypic network is derived from the conceptual framework associated to genotypic and phenotypic spaces proposed by several authors (Fontana and Schuster, 1987; Schuster et al., 1994; Huynen, 1996; Stadler et al., 2001; Crutchfield and van Nimwegen, 2002; Stadler, 2002). The hypercube is also related to the above definition. The hypercube Qν is a network of 2ν nodes represented by binary sequences where . The edges of the hypercube connect nodes whose sequences differ in exactly one bit, i.e., there is an edge (Si, Sj) ∈ Qn when dH(Si, Sj) = 1. Let's define the Hamming distance dH(Si, Sj) between any pair of sequences as follows:
In addition, every Boolean function can be represented as a subgraph of the hypercube (Harary, 1989) (see Figure 1B). Functions in the phenotype network are labeled with its designation number (a binary sequence) and thus, adjacent functions can be formally defined using the Hamming distance between the corresponding designation numbers. The only relevant difference between the standard hypercube and the phenotype network is that the same function can be represented with several nodes (genotypes).
Figure 2 maps the space of all reachable Boolean functions with 3 input variables. Even in this case, which is small and might not seem so relevant, several key classes of functions and circuit designs are involved (see below) either as single networks or as part of larger webs. These maps reveal several interesting features. First, a few functions appear several times in the network because they have more than one minimal FFBNs. For example, the function f10000110 accepts three different genotypes with the same minimal cost (they are displayed at the bottom left of Figure 2). Second, the network only considers 80 different functions out of the 256 logical functions of 3 variables. Two functions are equivalent and belong to the same class if one can be obtained from the other by a permutation of the input variables. We have discarded from further consideration functions that are equivalent to any class representative and have the same minimal FFBN (Hellerman, 1963).
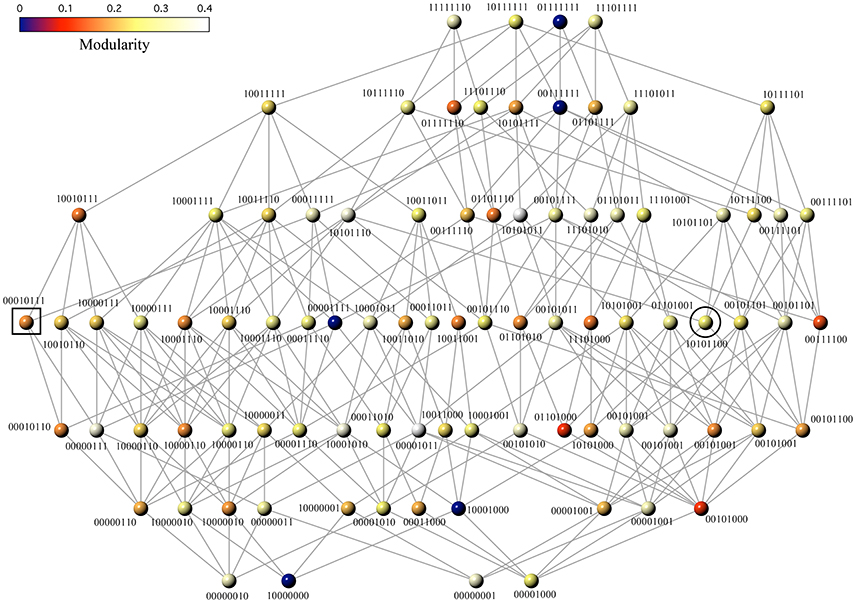
Figure 2. Phenotype network showing all reachable Boolean functions of 3-inputs. Each function fμ is labeled with its designation number X and mapped onto a minimal FFBN. Edges connect pairs of functions fμ and fY with Hamming distance dH(X, Y) = 1. Node color depicts functional modularity, i.e., the modularity value of the minimal FFBN (see text). The black square and circle mark the location of the majority and the multiplexor function, respectively.
5. Adaptation and the Breakdown of Modularity
The phenotype network maps the pathways to an eventual BM. Specifically, we can check if it is possible to evolve a less modular target function from any other source function. The distribution of modularity values P(Q) in this space has a well-defined peak with mean 〈Q〉 ≈ 0.2 (see Figure 3A). However, the variance displayed by this distribution suggests the possibility that BM is widespread. Specifically, we can find five functions with minimal modularity (Q(X) ≈ 0) that can be accessed from different neighborhoods of the phenotypic space (see yellow region in Figure 3B).
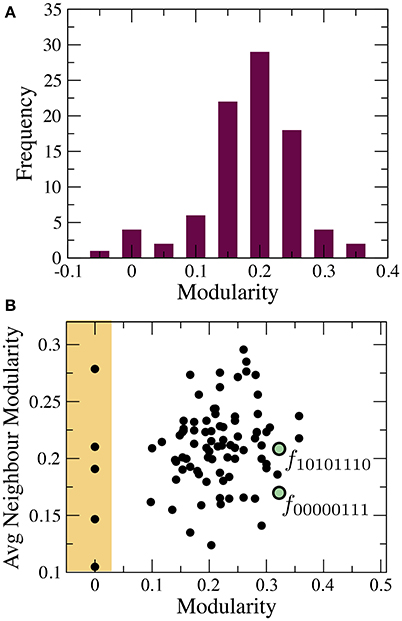
Figure 3. Statistical properties of the phenotype network. (A) distribution of modularity values and (B) correlation between node modularity and the average modularity of its nearest neighbors. The lack of correlation suggests the breakdown of modularity is not a specific property of some systems/environments. Green circles depict the location of two highly modular functions.
There are also highly modular functions surrounded by modular neighbors. To illustrate this behavior, we have chosen two important functions, namely the multiplexer (see Figure 4A) and the majority function (see Figure 4B). Both circuits have special relevance in both electronic designs and in synthetic biology (Macia and Solé, 2009). Each of these functions can be accessed in a few mutation steps from functions with higher modularity, i.e., Q(10101110) ≈ 0.3 and Q(00000111) ≈ 0.3, respectively. At least in these two cases, the evolution of useful functions is coupled to a reduction in modularity. These examples suggest how adaptation might lead to a BM. A decrease of functional modularity takes place when the evolutionary goal is non-separable, i.e., the computation of the output requires the interaction of several inputs. For example, the majority function is a global computation that combines all input variables to obtain the output. Similarly, consciously effortful tasks (like working memory) are expected to break the modularity of neural systems (Dehaene et al., 1998).
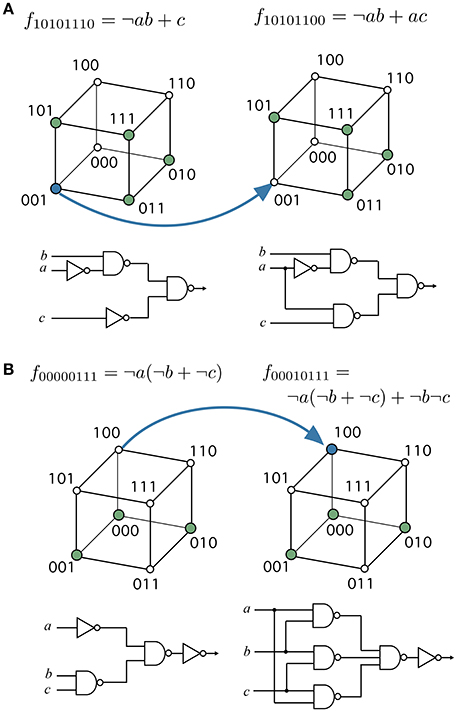
Figure 4. Evolving specific functions might involve a breakdown of modularity. (A) the multiplexer function has functional modularity Q(10101100) < Q(10101110), and (B) the majority function has Q(00010111) < Q(000000111). These target functions can be evolved from more modular ancestors, that is, functions with specific groups of inputs affecting different groups of outputs. Blue arrows represent edges in the phenotype network or the specific changes transforming the source function into the target function. Each function is represented with their minimal normal disjunctive form (top), hypercube (middle), and minimal FFBN (bottom).
How can we relate functional characteristics with the breaking of modularity? Figure 3B shows the absence of a clear correlation between the modularity of specific Boolean functions and the average modularity of its nearest neighbors in the phenotype network. Additional network analyses might be helpful to understand this pattern. In a previous study, we have suggested that the BM is related to the small-world behavior of complex networks (Valverde and Solé, 2007). Software projects have a natural tendency to become disordered structures (Peter, 1986; Eick et al., 2001). This degradation is caused by widespread software changes and indirect dependencies between unrelated pieces of code.
These changes in modular organization have important implications for both engineering and evolved circuit designs. In this context, it was early suggested that software design is an instance of a multi-objective optimization process (Valverde et al., 2002). When designing software, there is a tradeoff between efficient communication and separation of functional tasks (i.e., modularization). Indeed, small-sized software architectures are trees (as one should expect from optimization leading to hierarchical structures) but clustering emerges at larger sizes. As the number of components increases, conflicting constraints arise between different components that would prevent the reaching of an optimal solution. A need to exchange information between distant parts of a system can lead to a modularity reduction.
The minimization of FFBNs is a similar problem. The presence of functional constraints in the Boolean function creates a larger density of connections and a more regular or lattice-like topology (see Figure 5). On the other hand, the optimal circuit under a minimal number of functional contraint is a tree-like, modular architecture (Bagrow, 2012). Indeed, we can check there is a positive correlation between the average path length (Equation 1) and the modularity of minimal FFBNs (see Figure 5A). For example, the addition of a link between the two branches of the multiplexer (see Figure 4A) leads to a sharp decrease of its average path length.
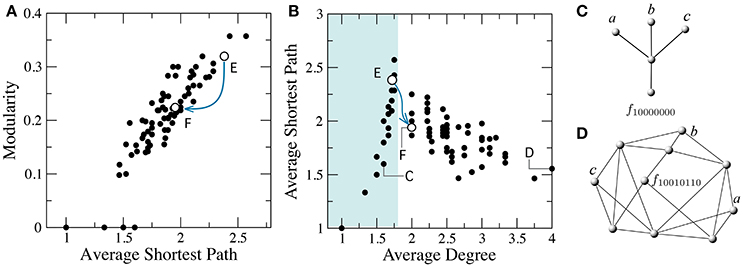
Figure 5. Breakdown of modularity involves a transition from tree-like to well-connected networks. (A) Correlation between average shortest path and modularity. (B) The average degree k* separates two main classes of minimal FFBNs: non-modular topologies with 〈k〉 > k* and tree-like networks with 〈k〉 ≤ k* ≈ 1.75 (cyan area). The minimal FFBNs for the functions f10000000 (C) and f10010110 (D) and the location of the multiplexer (F) and its ancestor function (E) are also shown. Blue lines denote the edge (f10101100, f10101110) in the phenotype network, which corresponds to a decrease in modularity and average shortest path.
We can look at these results from the perspective of spectral graph theory (Chung, 1997). Optimal characteristics of network structures can be mapped onto the spectrum of the graph Laplacian matrix. For example, the larger the second smallest eigenvalue λ2, the faster the information diffuses through the network. The magnitude of λ2 is inversely related to the mixing time for Markov chains, or the time required to achieve the steady state distribution (Mohar, 1997). In this context, the maximization of modularity is not compatible with low mixing time, i.e., rapid diffusion of information (Donetti et al., 2006).
In other words, the breakdown of modularity in minimal feed-forward Boolean circuits involves a transition from separable tasks (e.g., tree-like architectures) to rapid diffusion of information (well-connected networks). When functions can be separated into a set of disjoint tasks, we can design a modular network and different modules can process different tasks or different components of the inputs. On the other hand, we cannot simply obtain modularity and short path lengths when networks target non-separable tasks under cost pressures.
6. Discussion
The interplay between fitness and system-level properties such as modularity has been investigated in natural and artificial designs. Simon proposed that nearly decomposable systems composed by independent modules allow faster adaptation to highly fluctuating environments (Simon, 1962, 1995). A modular architecture allows independent changes in different parts of the system without affecting the whole. Well-adapted modules are conserved and provide a robust infrastructure for future adaptation. This poses a puzzle because modular architectures cannot be always maintained or reached in artificial evolution. For example, evolutionary algorithms often yield designs that are not decomposable and it is difficult to understand the way these systems work. In software engineering, even if a modular design is provided as initial solution, development rapidly moves to entangled and monolithic solutions (Eick et al., 2001).
Here we have proposed that the breakdown of modularity takes place because there are changes in the fitness function, e.g., a shift from a well-known environment to a less predictable environment. There are functional constraints to network modularity. For example, tasks involving non-separable input-output mappings like learning a color naming task with interference, evolving a robust metabolic network in a highly fluctuating environment (Hintze and Adami, 2008) or developing software under constantly changing requirements do not seem to evolve modular networks. Everything is a novelty for a network exposed to a highly fluctuating environment and thus, it makes little sense to maintain costly memories for reusing past information. In a highly fluctuating environment, the only requirement for survival is to issue fast responses and quick adaptations. The lack of memory imposes a strong constraint on the complexity of evolved structures.
The reduction of modularity has to be contrasted with existing theories for the emergence of modularity (Wagner et al., 2007). For example, the breakdown to modularity does not require a change from a modularly changing environment (Kashtan and Alon, 2005) to another static environment. Introducing an additional, temporal dimension in the fitness function is likely to internally decouple the system (and thus creating the possibility of increasing modularity). Moreover, looking at a few case studies does not enable a full understanding of how and when modular networks are expected to evolve. Instead, we have proposed a simple model of network landscape where it is possible to exhaustively characterize the breakdown of modularity in a well-defined way and studied the influence of functional and cost constraints.
The study of biological networks requires the examination of the interactions between modularity, network diameter and function. The phenotype space can be classified in two types of functions depending if the minimal FFBNs (genotypes) is a sparse or a dense network. Sparse genotypes have treelike topologies and their computation involves minimal input interaction. On the other hand, the modularity of dense networks drops with the increasing number of distant interactions. Some of the sparse networks are also maximally modular in our system, contradicting the intuition that modularity depends on densely intra-connected communities. It can be shown that tree modularity is significant even when they are made of sparse modules (Bagrow, 2012). Nodes in sparse trees acting as bottlenecks are sufficient to achieve high modularity values. The above suggests that we have to extend our definitions of modularity to take into account different measures of internal network connectivity.
Finally, our results might be useful to understand the limits of the hypothesis put forward by Simon. The analysis of the phenotype network reveals how the breakdown of modularity is more likely to take place from regions of the landscape populated by highly modular functions. Although neutral models suggest that tinkering increases the possibilities to discover modular designs (Solé and Valverde, 2006; Wagner et al., 2007; Solé and Valverde, 2008; Tosh, 2016), the structure of the phenotype network suggests that it is not always possible to avoid the breaking of modularity. Sometimes modularity must be spared, as in the evolution of novelties (Wagner and Lynch, 2010). The road to innovation is not necessarily paved by modularity.
Author Contributions
The author confirms being the sole contributor of this work and approved it for publication.
Conflict of Interest Statement
The author declares that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Acknowledgments
The author thanks Ricard Solé for a careful review of this paper and useful comments and discussions. The author also wants to thank the suggestions of two referees. This work was supported by Botín Foundation by Banco Santander through its Santander Universities Global Division, FIS2016-77447-R from Spain Ministerio de Economía, Industria y Competitividad, AEI/MINEICO/FEDER and UE.
References
Alexander-Bloch, A. F., Gogtay, N., Meunier, D., Birn, R., Clasen, L., Lalonde, F., et al. (2010). Disrupted modularity and local connectivity of brain functional networks in childhood-onset schizophrenia. Front. Syst. Neurosci. 4:147. doi: 10.3389/fnsys.2010.00147
Bagrow, J. P. (2012). Communities and bottlenecks: trees and treelike networks have high modularity. Phys. Rev. E 85:066118. doi: 10.1103/PhysRevE.85.066118
Bashan, A., Bartsch, R. P., Kantelhardt, J. W., Havlin, S., and Ivanov, P. Ch. (2012). Network physiology reveals relations between network topology and physiological function. Nat. Commun. 3:702. doi: 10.1038/ncomms1705
Blondel, V. D., Guillaume, J.-L., Lambiotte, R., and Lefebvre, E. (2008). Fast unfolding of communities in large networks. J. Stat. Mech. Theory Exp. 2008:P10008. doi: 10.1088/1742-5468/2008/10/p10008
Chung, F. R. K. (1997). “Spectral graph theory,” in CBMS Regional Conference Series in Mathematics, Vol. 92 (Fresno: American Mathematical Society, California State University).
Clune, J., Mouret, J.-B., and Lipson, H. (2013). The evolutionary origins of modularity. Proc R. Soc B 280:20122863. doi: 10.1098/rspb.2012.2863
Crutchfield, J. P., and van Nimwegen, E. (2002). “The evolutionary unfolding of complexity,” in Evolution as Computation. Natural Computing Series, eds L. F. Landweber, and E. Winfree (Berlin; Heidelberg: Springer). doi: 10.1007/978-3-642-55606-7_4
David, A. S. (1994). Dysmodularity: a neurocognitive model for schizophrenia. Schizophr. Bull. 20, 249–255. doi: 10.1093/schbul/20.2.249
Dehaene, S., Kerszberg, M., and Changeux, J.-P. (1998). A neuronal model of a global workspace in effortful cognitive tasks. Proc. Natl. Acad. Sci. U.S.A. 95, 14529–14534. doi: 10.1073/pnas.95.24.14529
Donetti, L., Neri, F., and Muoz, M. A. (2006). Optimal network topologies: expanders, cages, Ramanujan graphs, entangled networks and all that. J. Stat. Mech. Theory Exp. 8:P08007. doi: 10.1088/1742-5468/2006/08/P08007
Eick, S. G., Graves, T. L., Karr, A. F., Marron, J. S., and Mockus, A. (2001). Does code decay? Assessing the evidence from change management data. IEEE Trans. Soft. Eng. 1, 1–12. doi: 10.1109/32.895984
Fontana, W., and Schuster, P. (1987). A computer model of evolutionary optimization. Biophys. Chem. 26, 123–147. doi: 10.1016/0301-4622(87)80017-0
Gates, A. J., and Rocha, L. M. (2016). Control of complex networks requires both structure and dynamics. Sci. Rep. 6:24456. doi: 10.1038/srep24456
Girvan, M., and Newman, M. E. (2002). Community structure in social and biological networks. Proc. Natl. Acad. Sci. U.S.A. 99, 7821–7826. doi: 10.1073/pnas.122653799
Godwin, D., Barry, R. L., and Marois, R. (2015). Breakdown of the brain's functional network modularity with awareness. Proc. Natl. Acad. Sci. U.S.A. 112, 3799–3804. doi: 10.1073/pnas.1414466112
Harary, F. (1989). The graph of a boolean function. J. Exp. Theor. Artif. Intell. 1, 163–169. doi: 10.1080/09528138908953698
Hartwell, L. H., Hopfield, J. J., Leibler, S., and Murray, A. W. (1999). From molecular to modular cell biology. Nature 402, C47–C52. doi: 10.1038/35011540
Hellerman, L. (1963). A catalog of three-variable or-invert and and-invert logical circuits. IEEE Trans. Electr. Comput. EC-12, 198–223. doi: 10.1109/PGEC.1963.263531
Hintze, A., and Adami, C. (2008). Evolution of complex modular biological networks. PLoS Comput. Biol. 4:e23. doi: 10.1371/journal.pcbi.0040023
Huynen, M. A. (1996). Exploring phenotype space through neutral evolution. J. Mol. Evol. 43, 165–169. doi: 10.1007/BF02338823
Kashtan, N., and Alon, U. (2005). Spontaneous evolution of modularity and network motifs. Proc. Natl. Acad. Sci. U.S.A. 102, 13773–13778. doi: 10.1073/pnas.0503610102
Kauffman, S. A. (1969). Metabolic stability and epigenesis in randomly constructed genetic nets. J. Theor. Biol. 22, 437–467. doi: 10.1016/0022-5193(69)90015-0
Macia, J., and Solé, R. V. (2009). Distributed robustness in cellular networks: insights from synthetic evolved circuits J. R. Soc. Interface 6, 393–400. doi: 10.1098/rsif.2008.0236
McCluskey, E. J. Jr. (1956). Minimization of boolean functions. Bell Syst. Tech. J. 35, 1417–1444. doi: 10.1002/j.1538-7305.1956.tb03835.x
Meunier, D., Lambiotte, R., and Bullmore, E. T. (2010). Modular and hierarchically modular organization of brain networks. Front. Neurosci. 4:200. doi: 10.3389/fnins.2010.00200
Mohar, B. (1997). “Some applications of Laplace eigenvalues of graphs,” in Graph Symmetry: Algebraic Methods and Applications. NATO Advanced Study Institute Series C, Vol. 497, eds G. Hahn and G. Sabidussi (Montreal, QC: Kluwer Academic Publishers), 225–275.
Munteanu, A., and Solé, R. V. (2008). Neutrality and robustness in Evo-Devo: emergence of lateral inhibition. PLoS Comput. Biol. 4:e1000226. doi: 10.1371/journal.pcbi.1000226
Quine, W. V. O. (1952). The problem of simplifying truth functions. Amer. Math. Monthly 59, 521–531. doi: 10.2307/2308219
Ravasz, E., Somera, A. L., Mongru, D. A., Oltvai, Z. N., and Barabási, A.-L. (2002). Hierarchical organization of modularity in metabolic networks. Science 297, 1551–1555. doi: 10.1126/science.1073374
Rudell, R., and Sangiovanni-Vincentelli, A. (1987). Multiple-valued minimization for pla optimization. IEEE Trans. Comput. Aided Design Integr. Circuits Syst. 6, 727–750. doi: 10.1109/TCAD.1987.1270318
Schuster, P., Fontana, W., Stadler, P. F., and Hofacker, I. L. (1994). From sequences to shapes and back: a case study in RNA secondary structures. Proc. Biol. Sci. 255, 279–284. doi: 10.1098/rspb.1994.0040
Schlosser, G., and Wagner, G. P. (2004). Modularity in Development and Evolution. Chicago, IL: University of Chicago Press.
Shmulevich, I., Kauffman, S. A., and Aldana, M. (2005). Eukaryotic cells and dynamically ordered or critical but not chaotic. Proc. Natl. Acad. Sci. U.S.A. 38, 13439–13444. doi: 10.1073/pnas.0506771102
Simon, H. A. (1995). “Near-decomposability and complexity: how a mind resides in a brain,” in The Mind, the Brain, and Complex Adaptive Systems, eds H. Morowitz and J. Singer (Reading, MA: Addison-Wesley), 25–43.
Solé, R. V., and Valverde, S. (2006). Are network motifs the spandrels of cellular complexity? Trends Ecol. Evol. 21, 419–422. doi: 10.1016/j.tree.2006.05.013
Solé, R. V., and Valverde, S. (2008). Spontaneous emergence of modularity in cellular networks. J. R. Soc. Interface 5, 129–133. doi: 10.1098/rsif.2007.1108
Stadler, B. M., Stadler, P. F., Wagner, G. P., and Fontana, W. (2001). The topology of the possible: formal spaces underlying patterns of evolutionary change. J. Theor. Biol. 213, 241–274. doi: 10.1006/jtbi.2001.2423
Stadler, P. F. (2002). “Fitness landscapes,” in Biological Evolution and Statistical Physics, eds M. Lässig and A. Valleriani (Berlin; Springer-Verlag), 187–207.
Tosh, C. R. (2016). Can computational efficiency alone drive the evolution of modularity in neural networks? Sci. Rep. 6:31982. doi: 10.1038/srep31982
Valverde, S., Ferrer-Cancho, R., and Solé, R. V. (2002). Scale-free networks from optimal design. Europhys. Lett. 60, 512–518. doi: 10.1209/epl/i2002-00248-2
Valverde, S., and Solé, R. V. (2007). “Hierarchical small-worlds in software architecture,” in Special Issue on Software Engineering and Complex Networks, DCDIS Series B, Vol. 14, eds K. L. Teo and X. Liu (Watam Press), 1–11. Available online at: http://www.watsci.org/main.htm; http://www.watsci.org/DCDIS_supp/2007b6.pdf
van den Berg, D., Gong, P., Breakspear, M., and van Leeuwen, C. (2012). Fragmentation: loss of global coherence or breakdown of modularity in functional brain architecture? Front. Syst. Neurosci. 6:20. doi: 10.3389/fnsys.2012.00020
Wagner, G. P., and Lynch, V. J. (2010). Evolutionary novelties. Curr. Biol. 20, R48–R52. doi: 10.1016/j.cub.2009.11.010
Wagner, G. P., Pavlicecv, M., and Cheverud, J. M. (2007). The road to modularity. Nat. Rev. Gen. 8:921. doi: 10.1038/nrg2267
Keywords: modularity, phenotype network, Boolean function, evolution
Citation: Valverde S (2017) Breakdown of Modularity in Complex Networks. Front. Physiol. 8:497. doi: 10.3389/fphys.2017.00497
Received: 30 April 2017; Accepted: 28 June 2017;
Published: 13 July 2017.
Edited by:
John G. Holden, University of Cincinnati, United StatesReviewed by:
Renaud Lambiotte, University of Namur, BelgiumXiang yun Gao, China University of Geosciences, China
Copyright © 2017 Valverde. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) or licensor are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Sergi Valverde, c2VyZ2kudmFsdmVyZGVAdXBmLmVkdQ==