 Yichen Hu1†
Yichen Hu1† Qinghua Zou1†
Qinghua Zou1† Yanbo Sun1†Weiming Li1*Zhaochuan Yin1Yuanpei Zhao1Kaiwen Shi1Hongyuan Liu1Jiahui Wang2
Yanbo Sun1†Weiming Li1*Zhaochuan Yin1Yuanpei Zhao1Kaiwen Shi1Hongyuan Liu1Jiahui Wang2- 1Department of Gastrointestinal Surgery, Second Affiliated Hospital of Kunming Medical University, Kunming, Yunnan, China
- 2Department of Phase I Clinical Trial Ward, Yunnan Cancer Hospital, The Third Affiliated Hospital of Kunming Medical University, Kunming, Yunnan, China
Background: Intestinal ischemia-reperfusion (II/R) injury is a serious condition characterized by high morbidity and mortality rates. Research has shown that II/R injury is closely linked to autophagy and immune dysregulation. This study aims to investigate the potential correlations between autophagy-related genes and infiltrating immune cells in II/R injury.
Methods: GSE96733, GSE37013, and autophagy-related genes were obtained from the Gene Expression Omnibus (GEO) and the Human Autophagy Database, respectively. Subsequently, the biological functions of the differentially expressed genes (DEGs) were explored through DEGs analysis, Kyoto Encyclopedia of Genes and Genomes (KEGG) analysis, and Gene Ontology (GO) analysis. Using R software, human autophagy-related genes were converted to their mouse homologous autophagy-related genes (ARGs). The DEGs were then intersected with ARGs to obtain differentially expressed autophagy-related genes (DEARGs). To identify hub genes, protein-protein interaction (PPI) network analysis, Lasso regression, and random forest methods were employed. A nomogram model was constructed to assess its diagnostic value. Following this, immune infiltration analysis was performed to evaluate the potential correlation between Hub genes and immune cell infiltration. Additionally, a hub gene-related network was constructed, and potential drugs targeting hub genes for the treatment of II/R injury were predicted. Finally, the expression levels of hub genes in a mouse model of II/R injury were validated through dataset verification and quantitative real-time polymerase chain reaction (qRT-PCR).
Results: Our analysis identified 11 DEARGs. Among these, 5 DEARGs (Myc, Hif1a, Zfyve1, Sqstm1, and Gabarapl1) were identified as hub genes. The nomogram model demonstrated excellent diagnostic value. Immune cell infiltration analysis indicated that these 5 hub genes are closely associated with dendritic cells and M2.Macrophage. Furthermore, the regulatory network illustrated a complex relationship between microRNAs (miRNAs) and the hub genes. Additionally, trigonelline and niacinamide were predicted as potential therapeutic agents for II/R injury. In both dataset validation and qRT-PCR validation, the four hub genes (Myc, Hif1a, Sqstm1, and Gabarapl1) showed consistency with the results of the bioinformatics analysis.
Conclusion: Myc, Hif1a, Sqstm1, and Gabarapl1 have been identified as ARGs closely associated with immune infiltration in II/R injury. These hub genes may represent potential therapeutic targets for II/R injury.
1 Introduction
Intestinal ischemia-reperfusion (II/R) injury is a common and potentially fatal condition that arises in circumstances such as acute mesenteric ischemia, hemorrhagic or septic shock, severe burns, small intestine transplantation, and abdominal aortic surgery (Gonzalez et al., 2015). II/R injury not only induces local intestinal damage but also disrupts the intestinal mucosal barrier (Cai et al., 2025), permitting intestinal bacterial endotoxins to enter the bloodstream, which can lead to extraintestinal multiple organ dysfunction or even failure, accompanied by high morbidity and mortality rates (Pierro and Eaton, 2004). The pathophysiological mechanisms underlying II/R injury are complex and involve various factors, including oxidative stress (Jia et al., 2020), inflammatory responses, apoptosis (Cao et al., 2023), ferroptosis (Li et al., 2020), and autophagy (Chen et al., 2023). Oxidative stress is a key component in the pathogenesis of II/R injury, and the excessive production of reactive oxygen species (ROS) is one of the critical factors exacerbating II/R injury (Sasaki and Joh, 2007). Nuclear factor erythroid 2-related factor 2 (Nrf2) is a crucial transcription factor that regulates cellular antioxidant responses, playing a vital role in maintaining cellular redox balance and protecting against oxidative stress-related damage (Bae et al., 2024). Recent studies have reported that activating the Nrf2-mediated signaling pathway is one of the important mechanisms for alleviating II/R injury (Li et al., 2021). Bryostatin-1 mitigates intestinal barrier dysfunction and oxidative stress induced by II/R injury by activating the Nrf2/HO-1 signaling pathway, which supports this perspective (Liu M. et al., 2023). Apoptosis is another major contributor to cell death in II/R injury (Zu et al., 2016). By inhibiting apoptosis markers (Bax and caspase 3/9) or overexpressing anti-apoptotic markers (Bcl2), the severity of II/R injury can be reduced (Wen et al., 2019; Almoiliqy et al., 2020). Currently, certain therapeutic effects have been achieved in treating II/R injury through anti-inflammatory, antioxidant, and anti-apoptotic actions (Wang et al., 2021); however, diagnostic and treatment methods for II/R injury remain limited. Therefore, identifying key biomarkers and exploring new therapeutic strategies are crucial.
Autophagy is a highly conserved cellular process that plays a crucial role in maintaining cellular homeostasis by degrading and recycling damaged organelles, proteins, and other cellular components (Liu S. et al., 2023). Numerous diseases, particularly cancer, diabetes, heart disease, and muscle disorders, are associated with impaired autophagy (Klionsky et al., 2021). An increasing number of studies have demonstrated that mammalian target of rapamycin (mTOR), AMP activated protein kinase (AMPK), Nrf2, Beclin-1, and p62 are associated with II/R injury and represent promising therapeutic targets for this condition (Chen et al., 2020; Shen et al., 2023). As an important key regulator, mTOR affects autophagic activity and is a key target for the autophagic pathway (Zhenzhen et al., 2022). AMPK, by modulating autophagy, plays a pivotal role as an energy-sensing signaling molecule in II/R injury (Cai et al., 2022). Recent studies have shown that inflammation, oxidative stress, and apoptosis in I/R injury can be alleviated by activating AMPK/mTOR signaling and modulating autophagy (Shen et al., 2023). Nrf2 has been proven to be involved in autophagy, and research has demonstrated that enhancing autophagy through the Akt/GSK-3β/Nrf2 pathway can mitigate II/R injury (Pajares et al., 2016; Chen et al., 2020). These findings indicate that autophagy plays a crucial role in II/R injury. However, the autophagy-related genes involved in II/R injury remain largely unknown. Therefore, utilizing bioinformatics to deeply explore autophagy-related markers in II/R injury is instrumental in discovering new potential biomarkers for this condition. Furthermore, studies have shown that immune regulation plays a pivotal role in the pathogenesis of II/R injury (Hu et al., 2022; Zhang F. L. et al., 2023). Recent data demonstrates an important role for lymphocytes, particularly T cells but also B cells in II/R injury (Linfert et al., 2009). However, there have been few reports that comprehensively investigate the relationship between the expression of ARGs and immune infiltration in II/R injury, as well as the biological functions of ARGs. In recent years, non-coding RNAs, including but not limited to long non-coding RNA (lncRNA), circular RNAs (circRNA), and microRNA (miRNA), have garnered particular attention (Zhang J. et al., 2023). Research has demonstrated that the microRNA-122a inhibitor activates the EGFR-NLRP3 signaling pathway, significantly reducing pyroptosis and alleviating II/R injury, thereby highlighting the importance of miRNA in regulating II/R injury (Wang et al., 2022).
Although numerous studies have provided preliminary evidence for the regulatory role of autophagy dysregulation in II/R injury, most research has focused primarily on the biological functions of specific genes, neglecting their relationship with the immune microenvironment and the regulation of target gene expression by non-coding RNAs in II/R injury. Therefore, we obtained datasets of II/R injury from public databases and screened for hub genes associated with immune infiltration and autophagy through biological analysis. Additionally, we constructed a nomogram model to predict the diagnosis of II/R injury and developed a circRNA–miRNA-mRNA regulatory network for these hub genes. We also assessed the therapeutic potential of the hub genes through drug prediction. Finally, qRT-PCR validation confirmed the relevance of these hub genes in II/R injury, providing new insights into the pathogenesis of this condition.
2 Materials and methods
2.1 Data source and processing
The raw data of GSE96733 [species: Mus musculus; Platforms: GPL23038 (Clariom_S_Mouse) Affymetrix Clariom S Assay, Mouse (Includes Pico Assay)] was collected from the GEO website (http://www.ncbi.nlm.nih.gov/geo/). The GSE96733 (II/R 3 h) comprised four sham-operated samples and four II/R samples collected at 3 h (n = 4). The R software package “limma” was utilized to analyze the differentially expressed genes (DEGs) between sham-operated samples and those with II/R injury, with thresholds set at |log2FC| ≥ 1 and an adjusted P-value <0.05 for screening. Additionally, four sham-operated samples and four II/R samples at 6 h (n = 4) were included in GSE96733 (II/R 6 h) as a validation set. The human intestinal II/R injury expression profile microarray GSE37013 was also obtained from the GEO database to serve as a second validation set. From this dataset, we selected 21 samples, which included 7 human jejunum samples (sham), 7 human jejunum-reperfusion 30 min samples (II/R 30 min), and 7 human jejunum-reperfusion 120 min samples (II/R 120 min), and the platform used was GPL6947 Illumina HumanHT-12 V3.0 expression beadchip. The workflow of this research is shown in Figure 1.
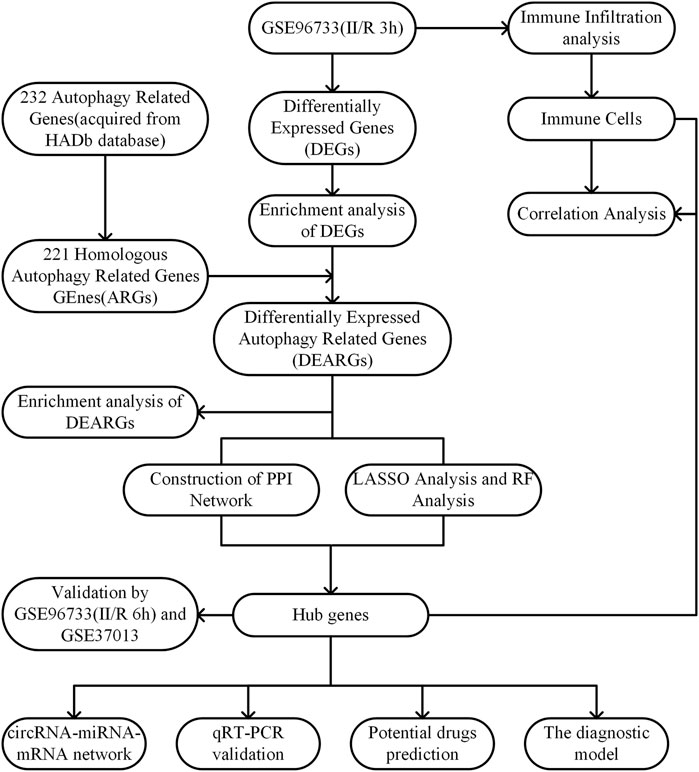
Figure 1. Experimental design roadmap.
2.2 Functional enrichment analysis of DEGs
The Gene Ontology (GO) analysis and Kyoto Encyclopedia of Genes and Genomes (KEGG) pathway enrichment of DEGs were performed using the R software (version 4.2.2) package “clusterProfiler” (Wu et al., 2021). The GO annotation encompasses three components: molecular function (MF), biological process (BP), and cellular component (CC). GO terms and signaling pathways exhibiting significant differences were identified under the criterion of adjusted P-value <0.05. Subsequently, the results were visualized using the R software packages “ggplot2″ and “GOplot” (Walter et al., 2015), respectively.
2.3 Identification and functional enrichment of differentially expressed autophagy-related genes
We identified 232 genes associated with autophagy from the Human Autophagy Database (http://www.autophagy.lu/). Since our study focuses on mice, we utilized the R software package “homologene” through the online platform Xiantao Academic (https://www.xiantaozi.com/) to conduct homologous gene conversion and obtain mouse homologous autophagy-related genes (ARGs). Subsequently, we intersected the ARGs with DEGs to identify differentially expressed autophagy-related genes (DEARGs). The differential expression of DEARGs was visualized using the R software package “ggpubr”. The R software package “Corrplot” was utilized to examine the Spearman correlation among DEARGs. KEGG and GO analyses of DEARGs were conducted using the R software package “clusterProfiler” with a significance threshold of P < 0.05.
2.4 PPI network analysis of DEARGs
The STRING online database (https://cn.string-db.org/) was employed to perform protein–protein interaction (PPI) network analysis, investigating the relationships among DEARGs with an interaction score exceeding 0.4. The resulting network was then analyzed and visualized using Cytoscape (version 3.10.3). To identify hub genes associated with autophagy, CytoHubba was employed, which calculated hub genes based on three distinct algorithms: Degree, Closeness, and Betweenness. Degree quantifies the number of direct connections a gene node has with other nodes, and genes with high degree values are typically positioned at the core of the network, possessing extensive biological functions or regulatory roles (Eisele et al., 2021). Closeness measures a gene’s ability to integrate information across the entire network by calculating the average shortest path distance from the node to all other nodes in the network. Genes with high closeness are generally located at the core of the network and can rapidly respond to environmental changes or regulate other genes (Park and Kim, 2009). Betweenness measures the frequency at which a node appears on the shortest paths connecting other nodes. Genes with high betweenness centrality are usually situated at the hubs of biological networks and may be involved in core regulatory or signaling processes (Roy et al., 2013). Therefore, we selected the top five genes from each algorithm as autophagy-related hub genes. Finally, the differential expression of these hub genes was analyzed using the Xiantao Academic online software.
2.5 Machine learning and identifying hub genes
To identify the hub genes and establish a diagnostic model of II/R injury, the least absolute shrinkage and selection operator (Lasso) algorithm, a logistic regression method for filtering variables to enhance the predictive performance, was initially adopted in this work to screen the hub genes with the “glmnet” package (Friedman et al., 2010). Next, the random forest (RF) algorithm, integrating multiple trees through the idea of ensemble learning to gain better accuracy, was employed to narrow down the hub genes with the “randomForest” package as well (Liaw and Wiener, 2002). We selected genes with |standardized regression coefficient| ≥ 0.05 in Lasso and feture importance score ≥0.1 in RF. The hub genes were subsequently determined by taking the union of these selected genes and those identified through PPI screening.
2.6 Construction and verification of the diagnostic model
The nomogram was constructed based on the five hub genes using the “rms” package (Zhao and Li, 2023). The area under the receiver operating characteristic (ROC) curve was plotted with the “pROC” R package to evaluate the performance of the hub genes in diagnosing II/R injury (Hanley and McNeil, 1982). Subsequently, the expression of the five hub genes was validated using the GSE37013 validation dataset, and the accuracy of these hub genes was assessed using the ROC curve.
2.7 Validation of hub genes expression in other datasets
The expression levels of key II/R injury genes were extracted from the GSE96733 (II/R 6 h) and GSE37013 datasets and analyzed using the t-test. The expression of hub genes was assessed, and the results are presented as violin plots. The results were visualized using the R software package “ggplot2”.
2.8 Immune cell in filtration analysis
The CIBERSORT algorithm is employed to evaluate changes in immune cells during immune infiltration. This study applies the algorithm to investigate alterations in the relative proportions of immune cell infiltration in II/R injury. Correlation analysis is conducted on 25 types of infiltrating immune cells in mice, and the results are visualized using the “ggcorrplot” package (Chen et al., 2017).
2.9 Construction of the circRNA-miRNA-mRNA regulatory network
This study initially analyzed five hub genes using the TargetScan (https://www.targetscan.org/vert_80/) and miRDB (https://mirdb.org/) databases. Subsequently, we employed RNAhybrid software (https://bibiserv.cebitec.uni-bielefeld.de/rnahybrid) to predict the circRNAs targeted by these miRNAs. Finally, the interaction network among the aforementioned genes, miRNAs, and circRNAs was visualized and plotted using Cytoscape software.
2.10 Predicting potential drugs
Utilize the Comparative Toxicogenomics Database (CTD) (https://ctdbase.org/) to predict targeted drugs for five key genes. Subsequently, visualize the results using the “ggplot2” package in R software.
2.11 II/R injury model construction
Before constructing the mouse II/R injury model, all mice were fasted for 12 h with free access to water. The mice were anesthetized via intraperitoneal injection of pentobarbital at a dosage of 30 mg/kg. Upon exhibiting signs of limb weakness, the mice were placed flat on a sterile drape, secured, and the abdominal skin was prepared and disinfected. Following local infiltration with 2% lidocaine, a 1.5 cm incision was made along the midline of the abdomen, approximately 2 mm below the xiphoid process, using a scalpel. The abdominal cavity was then accessed layer by layer to expose the intestines, and the superior mesenteric artery (SMA) was identified and isolated. A non-traumatic microarterial clamp was employed to temporarily occlude the SMA and its branches, with timing initiated immediately. The cessation of arterial pulsation was observed, and the intestinal tract became edematous and pale, indicating successful ischemia. The intestine was gently returned to the abdominal cavity, covered with sterile gauze, and the intestinal lumen was kept moist. After 45 min, the microarterial clamp was removed to restore blood flow. The return of the intestine’s color to a healthy red and the gradual strengthening of arterial pulsations indicated successful reperfusion. Subsequently, 0.5 mL of warm saline was dripped into the abdominal cavity, and the mouse’s abdomen was sutured layer by layer using 3–0 silk thread. The mouse was then placed in a cage adjacent to a radiator for warmth. Following 3 h of perfusion, all mice were euthanized with an overdose of pentobarbital (intraperitoneal injection, 200 mg/kg). The small intestine tissue was subsequently excised and stored at −80°C. For the sham surgery group, only the SMA was separated without clamping, and the same procedures as described above were followed (Gubernatorova et al., 2016).
2.12 Quantitative reverse transcription-polymerase chain reaction
Total RNA was extracted from frozen intestinal tissue using an RNA extraction kit (Thermo Fisher, USA, product number 15596-018). Subsequently, reverse transcription was performed using a cDNA synthesis kit (TIANGEN, China, product number SB-Q204) (Watson et al., 2008). The relative mRNA expression levels were evaluated using the 2−ΔΔCq method, with β-actin serving as the internal control on the LightCycle 96 real-time fluorescence quantitative PCR system (Roche). Table 1 presents the primer sequences utilized.
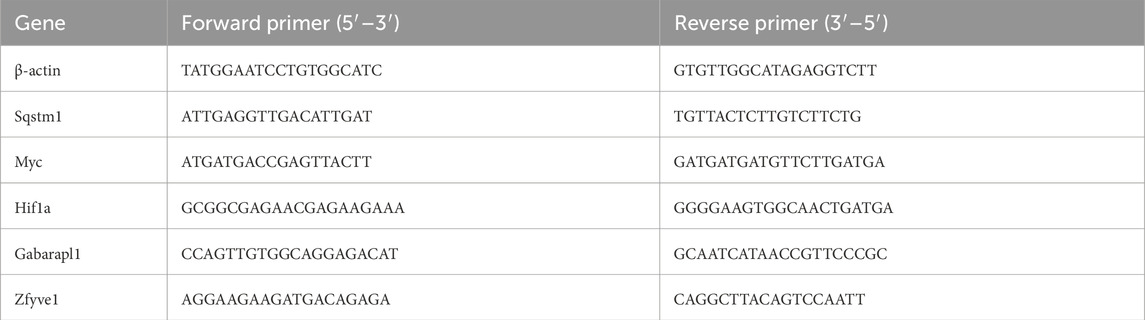
Table 1. Primer sequences used in the qRT-PCR experiments.
2.13 Statistical analysis
Statistical analyses were performed using R software (version 4.4.2) and GraphPad Prism (version 9.0.0.121). The comparison of gene expression in II/R injury between the two groups was conducted using an unpaired Student's t-test. A P-value of less than 0.05 was deemed statistically significant in all analyses.
3 Results
3.1 DEGs identification
Using the R software package “limma” for differential gene analysis, we identified a total of 1,027 DEGs from sham-operated samples and samples with II/R injury, comprising 605 upregulated genes and 422 downregulated genes (Figures 2A,B). See Supplementary Table S1 for details.
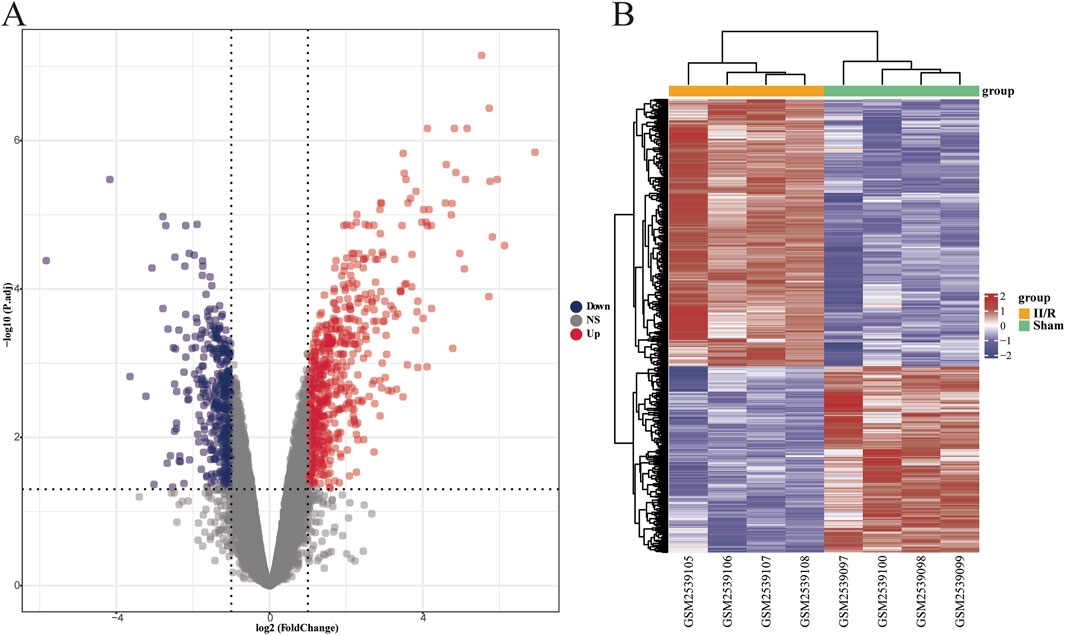
Figure 2. Identification of DEGs. (A) The volcano plot illustrates the differentially expressed genes between the Sham and II/R injury samples. Red dots represent upregulated genes, while dark blue dots indicate downregulated genes, with thresholds set at |log2FoldChange| ≥ 1 and an adjusted P-value <0.05. (B) The heatmap displays the differentially expressed genes between the Sham and III/R injury samples. Red signifies significantly upregulated DEGs, whereas dark blue indicates significantly downregulated DEGs in the samples.
3.2 Enrichment analysis of DEGs
To further investigate the potential biological changes associated with the screened DEGs, GO and KEGG pathway enrichment analyses were conducted using R software. The results from the GO biological process (BP) annotation indicated that the DEGs were predominantly enriched in immune response and autophagy (Figure 3A). Furthermore, the KEGG pathway enrichment analysis revealed that the DEGs were primarily enriched in the MAPK signaling pathway, PI3K−Akt signaling pathway, JAK−STAT signaling pathway, as well as pathways related to apoptosis and IL-17 signaling pathway (Figure 3B). These findings suggest a significant correlation between II/R injury and autophagy. See Supplementary Table S2 for details.
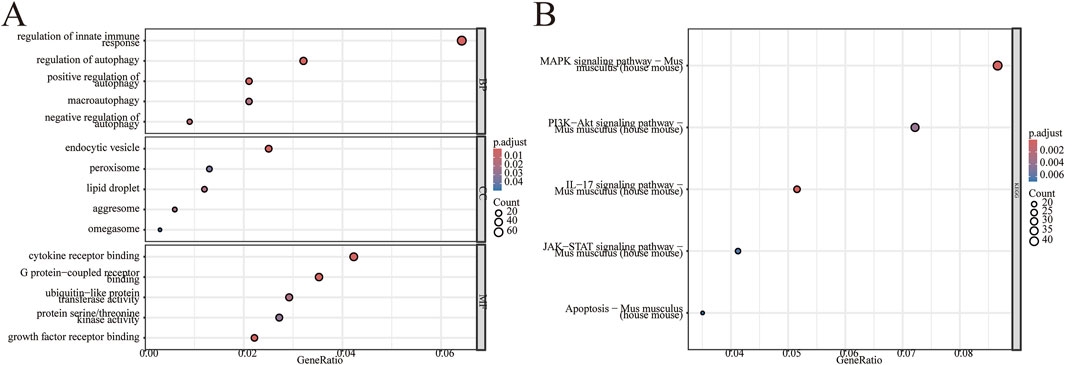
Figure 3. Enrichment analysis of DEGs. GO enrichment analysis was conducted on 1,027 differentially expressed genes, encompassing biological processes (BP), cellular components (CC), and molecular functions (MF). (A) The bubble plot illustrates the enriched GO terms. (B) The bubble plot displays the enriched KEGG pathways. GO, Gene Ontology; KEGG, Kyoto Encyclopedia of Genes and Genomes.
3.3 Identification of DEARGs
To further investigate autophagy-related genes in II/R injury, we retrieved 232 genes associated with autophagy from the Human Autophagy Database (http://www.autophagy.lu/). Given that our study focuses on mice, we utilized the R software package “homologene” on the Xiantao Academic online platform (https://www.xiantaozi.com/) to perform homologous gene conversion, thereby obtaining mouse homologous ARGs. By intersecting ARGs with DEGs, we identified 11 DEARGs (Figure 4A). A detailed correlation analysis was conducted on the expression levels of these 11 DEARGs within the dataset, revealing complex interactions among these genes (Figure 4B). Furthermore, utilizing the differential expression previously analyzed by the “limma” package, the R software package “ggpub” was employed to display the expression patterns of the 11 DEARGs in II/R injury compared to Sham samples (Figure 4C). The eight genes with increased expression were Sphk1, Myc, Hif1a, Sqstm1, Capn2, Bag3, Gabarapl1 and Hspb8, while the three genes with decreased expression were Tnfsf10, Rb1, Zfyve1 (Table 2). KEGG and GO analyses of the DEARGs were conducted with a significance threshold of P < 0.05, and the enrichment results were visualized using R software (Figures 5A,B). See Supplementary Table S3 for details.
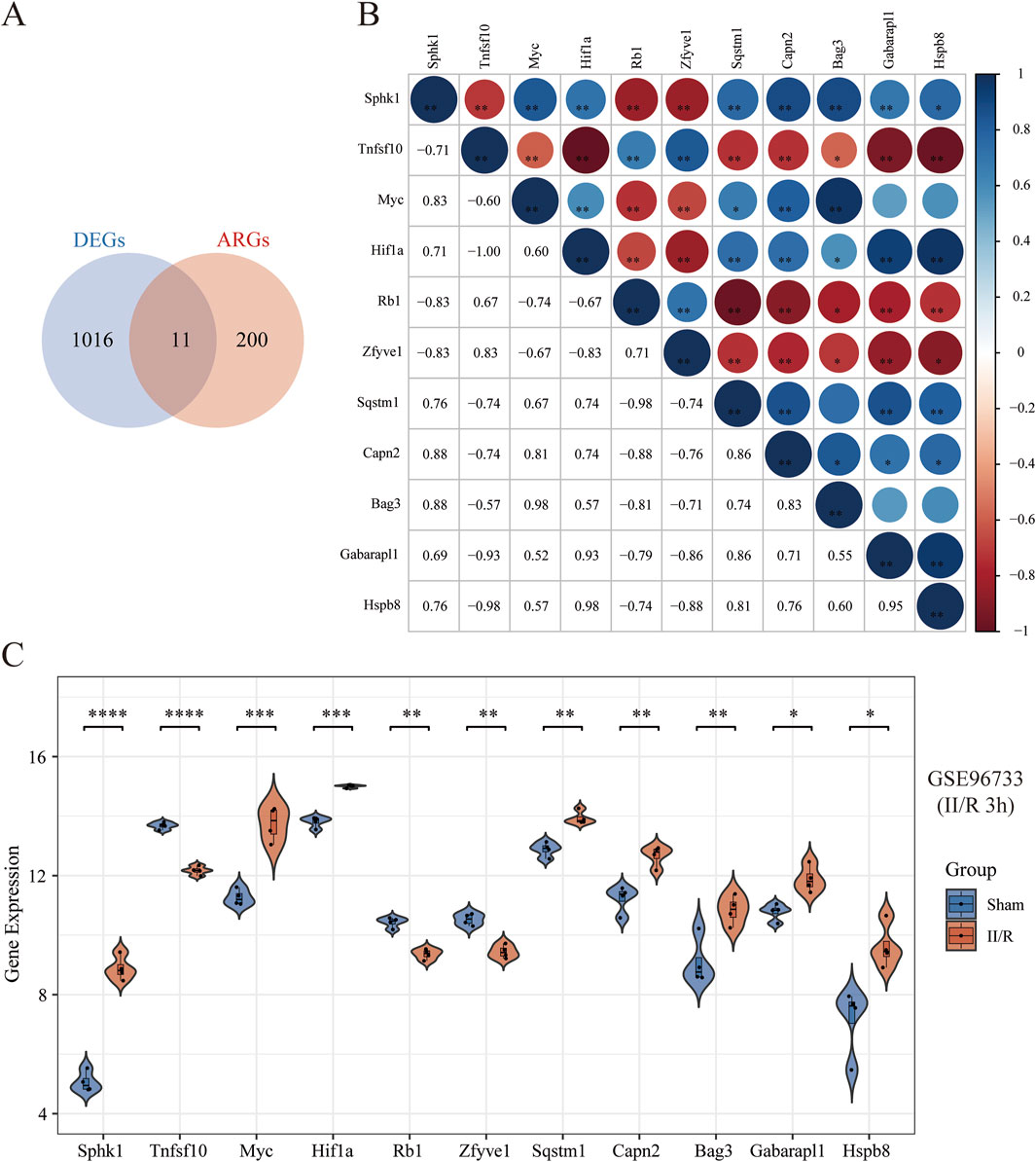
Figure 4. Identification of DEARGs. (A) Venn diagram of DEARGs between DEGs and ARGs. (B) Spearman’s correlation analysis of the 11 DEARGs. The abscissa and ordinate represent genes, different colors represent different correlation coefficients (blue represents positive correlation, and red represents negative correlation). The darker the color, the stronger the relation. Asterisks (*) stand for significance levels, *P < 0.05; **P < 0.01; ***P < 0.001. (C) Violin plot illustrate the differential expression of 11 autophagy-related genes in both II/R and Sham samples. The significance levels indicated as follows: *P < 0.05; **P < 0.01; ***P < 0.001; ****P < 0.0001. ns, not significant.
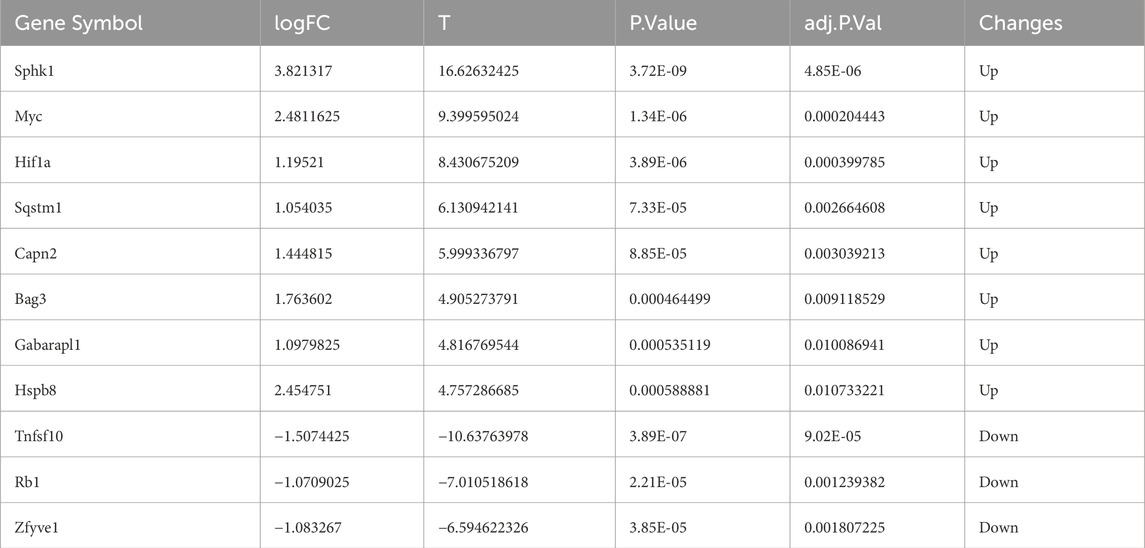
Table 2. The 11 DEARGs in II/R injury samples compared to Sham samples.
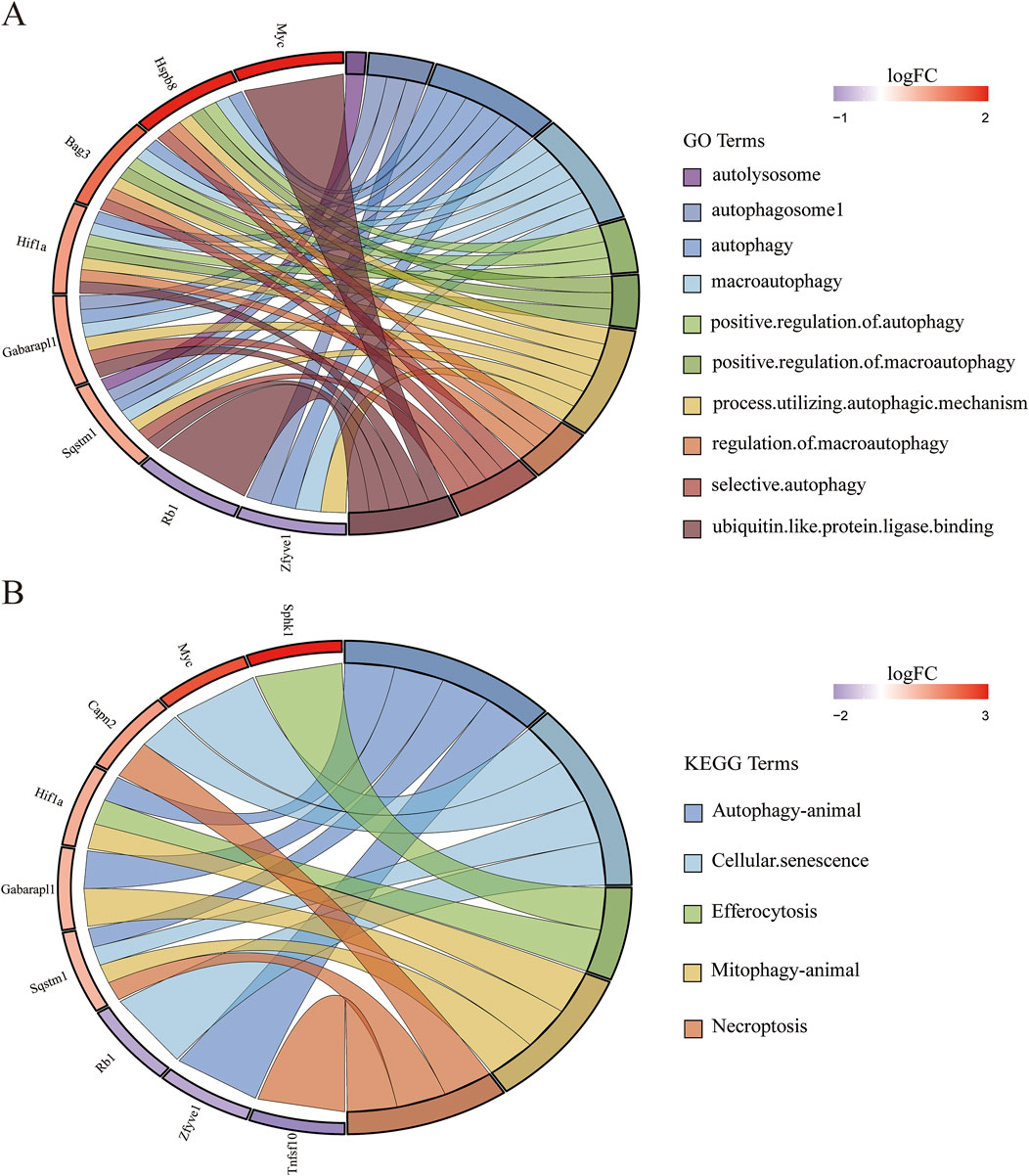
Figure 5. Gene-pathway association analysis in circos plots highlighting significant biological processes. (A) GO enrichment results of DEARGs. (B) KEGG enrichment results of DEARGs.
3.4 Construction of PPI and identification of hub genes
We explored the interactions among 11 DEARGs using the medium-confidence STRING database, resulting in a PPI network comprising 11 nodes and 11 edges (Figure 6A). Subsequently, we calculated the PPI network of these 11 DEARGs using three algorithms (Degree, Closeness, and Betweenness) with the CytoHubba plugin, selecting the top five genes as hub genes (Figures 6B–D). These hub genes include Sqstm1, Myc, Hif1a, Gabarapl1, and Zfyve1, which are represented in a heatmap format (Figure 6E). See Supplementary Table S4 for details.
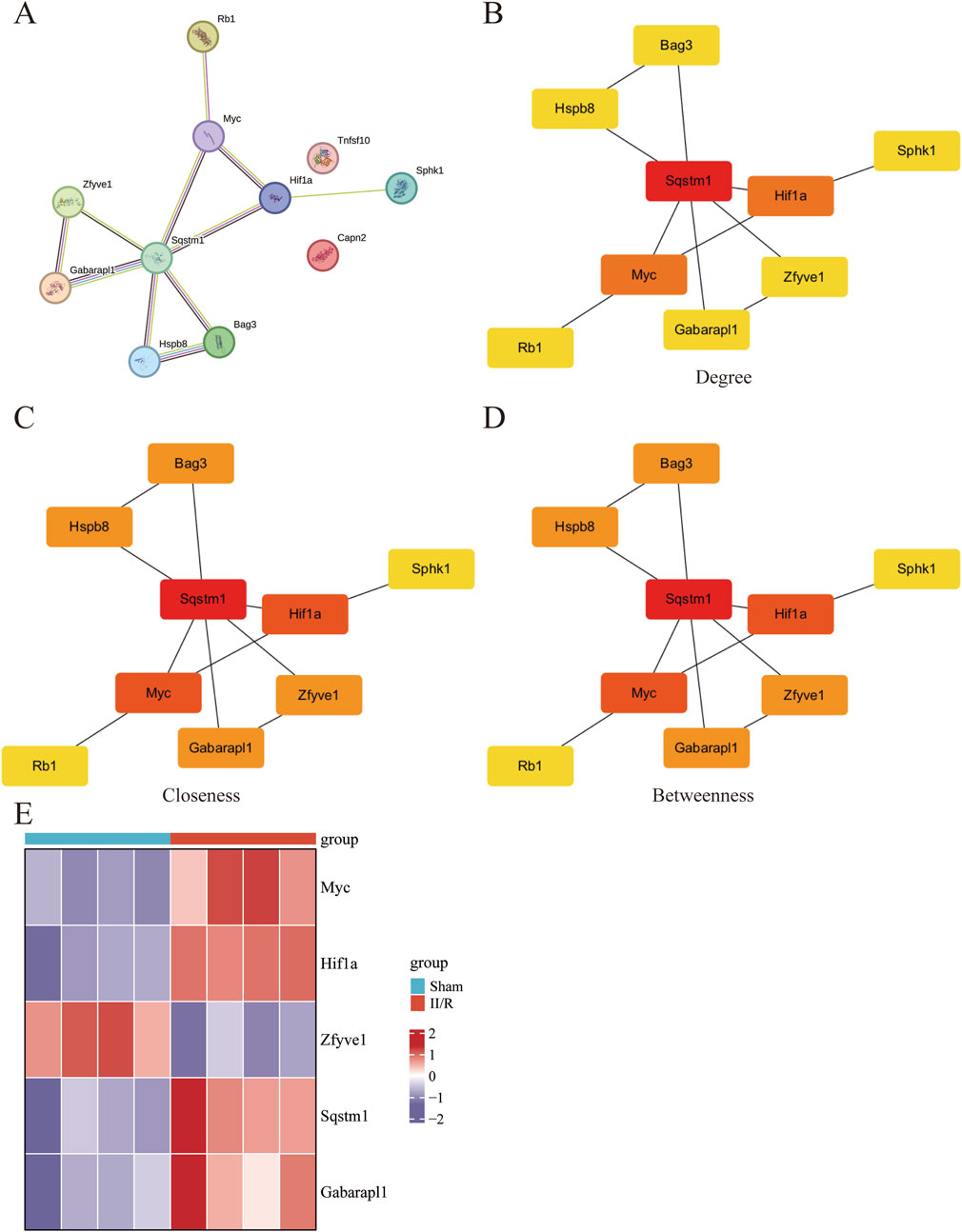
Figure 6. Screening of hub gene in DEARGs. (A) The protein–protein interaction network visualized by STRING. (B–D) The PPI network constructed from 11 DEARGs was analyzed using three algorithms: Degree, Closeness, and Betweenness. The top five genes were identified as hub genes. (E) The expression levels of Sqstm1, Myc, Hif1a, Gabarapl1, and Zfyve1 are illustrated using a heatmap.
3.5 Machine learning screened hub genes and confirmed the final hub genes
We employed machine learning algorithms to screen hub genes from DEARGs, initially utilizing the Lasso algorithm to identify four candidate hub genes based on |standardized regression coefficient| ≥ 0.05. Subsequently (Figures 7A–C). we applied the RF algorithm to select four candidate hub genes with feature importance score ≥0.1 (Figure 7D). The common gene Myc was identified through the intersection of a Venn diagram (Figure 7E). Finally, we took the union of the genes selected based on the PPI network and determined the final hub genes to be Sqstm1, Myc, Hif1a, Gabarapl1, and Zfyve1.
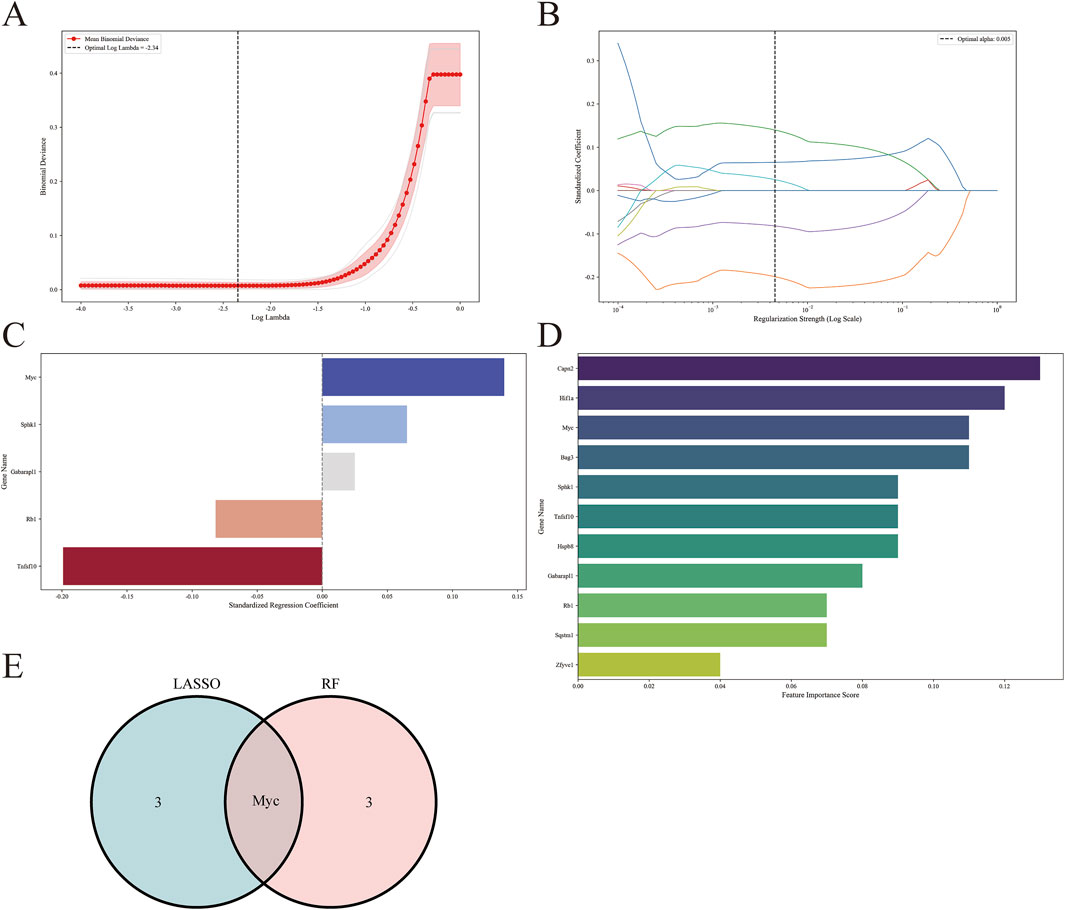
Figure 7. Screening for hub genes by machine learning algorithms. (A–C) Hub genes identified using the least absolute shrinkage and selection operator (Lasso) logistic regression algorithm. (D) Random forest (RF) algorithm to screen hub genes. (E) Venn diagram demonstrating overlapping hub gene screened by Lasso and RF.
3.6 Construction and verification of the diagnostic model
We utilized five identified hub genes (Sqstm1, Myc, Hif1a, Gabarapl1, and Zfyve1) as diagnostic markers for predicting II/R injury and constructed a nomogram (Figure 8A). The diagnostic performance was evaluated using ROC curves, achieving an area under the curve (AUC) value of 1 in the training set (Figure 8B). The efficiency of the predictive model was further validated using the GSE37013 dataset, which yielded an AUC value of 0.857 (95% CI = 0.695-1) (Figure 8C), indicating that the diagnostic model possesses good predictive value for II/R injury. However, the accuracy and reliability of this diagnostic model still require further investigation in future clinical trials. See Supplementary Table S5 for details.
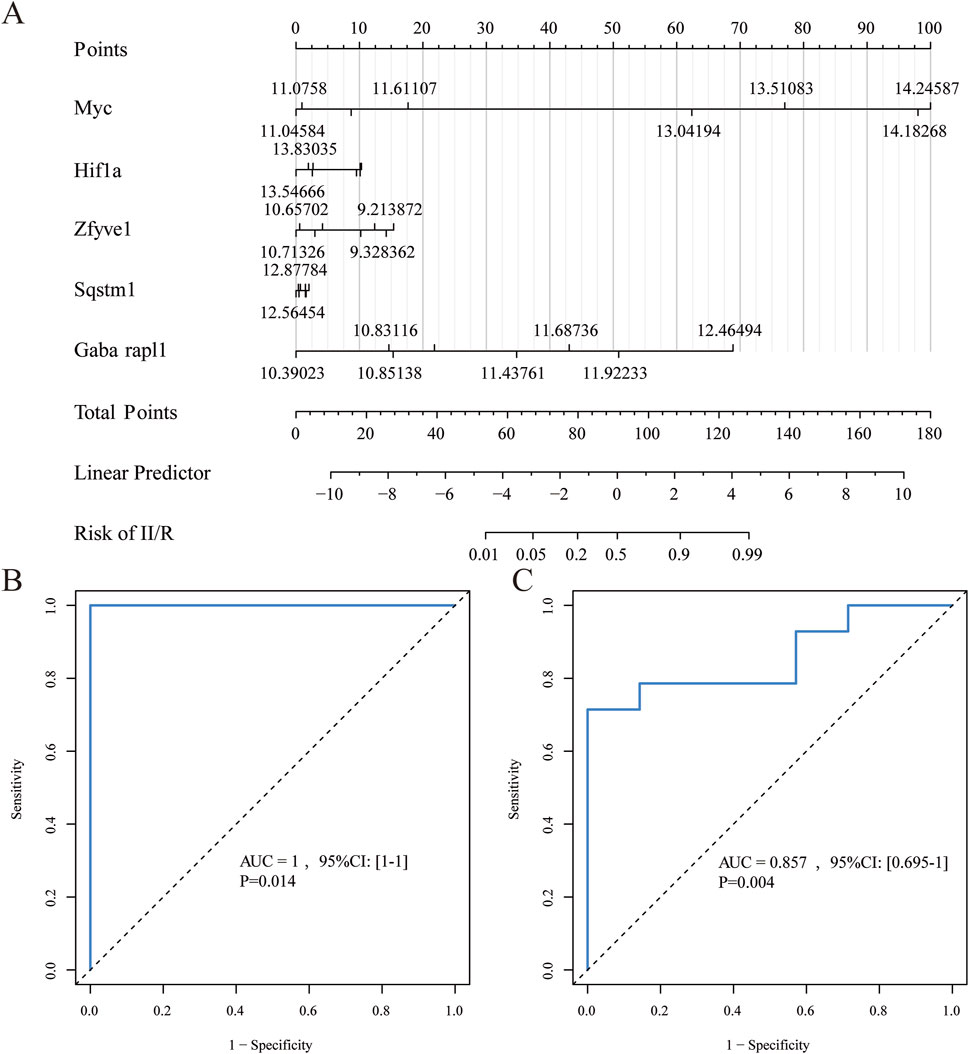
Figure 8. Diagnostic model construction. (A) Construction of a nomogram model with 5 hub genes. (B,C) ROC curve for evaluating and validating the diagnostic model’s performance.
3.7 Validation of hub genes expression in other datasets
We selected the data from GSE96733 (II/R 6 h) for validation and found that the expression of the five hub genes was consistent with the bioinformatics analysis of GSE96733 (II/R 3 h) (Figure 9A). Furthermore, upon validating with the GSE37013 dataset on human II/R injury, it was observed that the expression of SQSTM1 in the II/R group was lower than that in the sham group at both 30 min and 120 min of II/R injury (Figures 9B,C), indicating that autophagy plays a protective role in the early stages of II/R injury. See Supplementary Table S6 for details.
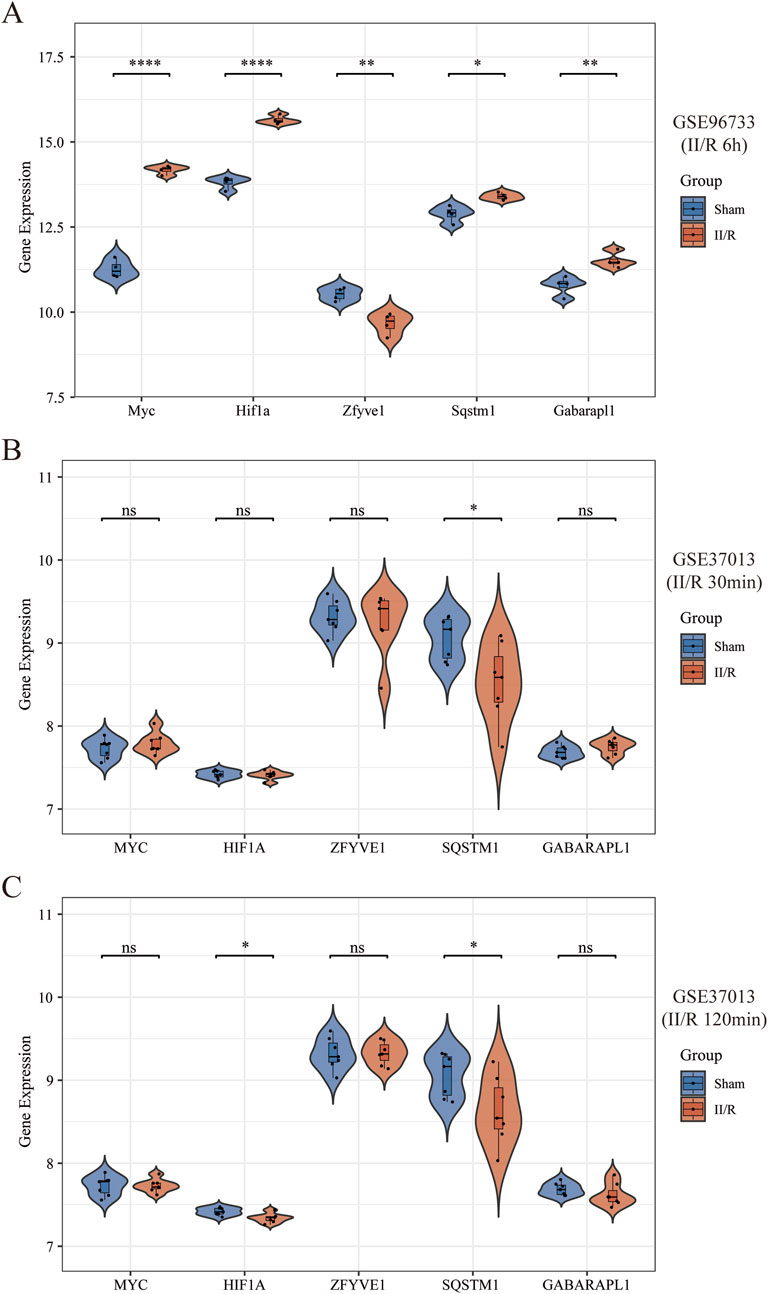
Figure 9. Validation of hub genes expression in other datasets. (A–C) The expression levels of Sqstm1, Myc, Hif1a, Gabarapl1, and Zfyve1 were verified by GSE96733 (II/R 6 h), GSE37013 (II/R 30 min), and GSE37013 (II/R 120 min) datasets, the results of which are presented as violin plot. The significance levels indicated as follows: *P < 0.05; **P < 0.01; ***P < 0.001; ****P < 0.0001. ns, not significant.
3.8 Immune cell analysis
We employed the CIBERSORT algorithm to assess the relative abundance of infiltrating immune cell subtypes in Sham samples and II/R injury samples. The bar graph illustrates the infiltration of various immune cell subtypes in each sample, while the box plot demonstrates the differences in the percentage of immune cells between the two sample groups. Compared to Sham, the II/R samples exhibited an increase in M2 macrophages (Figures 10A,B). Finally, the heatmap reveals the correlation between hub genes and immune cell infiltration, indicating that the expression of hub genes is significantly associated with immune cell infiltration among subtypes (Figure 10C). Notably, activated dendritic cells (DC.Actived) is negatively correlated with Zfyve1 and positively correlated with Sqstm1, Myc, Hif1a, and Gabarapl1. In contrast, the results for immature dendritic cells (DC.Immature) exhibit the opposite correlations. See Supplementary Table S7 for details.
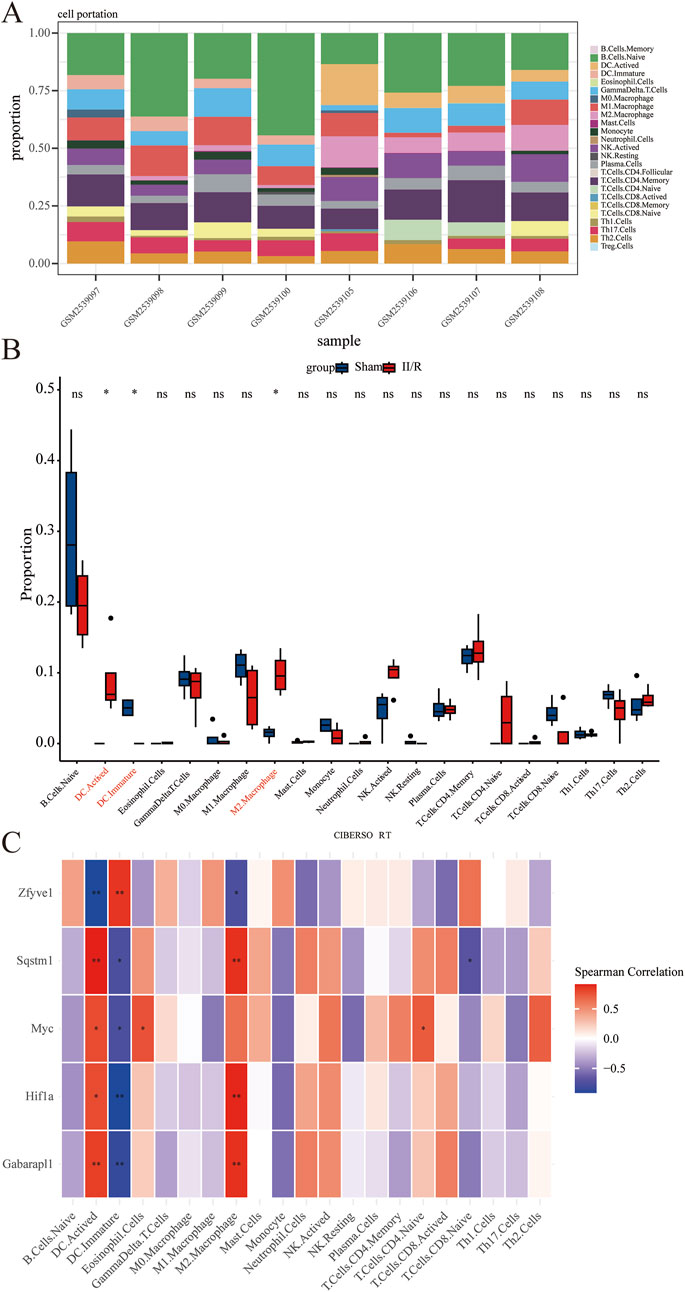
Figure 10. Results of immune cell analysis. (A) The histogram indicates the relative proportions of 25 immune cells. (B) Box plots illustrating the relative expression of each immune cell subtype between Sham and II/R injury samples. (C) Heatmap showing the correlation between immune cells and five hub genes. *P < 0.05; **P < 0.01.
3.9 Analysis of the circRNA-miRNA-mRNA regulatory network
The analysis results indicate that several miRNAs (including mmu-miR-6954-5p, mmu-miR-6919-3p, mmu-miR-7077-5p, mmu-miR-3099-5p, and mmu-miR-1956) demonstrate significantly higher node connectivity within the regulatory network. Notably, the Myc proto-oncogene interacts significantly with only one miRNA (mmu-miR-7077-5p), and a total of 14 circRNAs that interact with mmu-miR-7077-5p have been identified (Figure 11). See Supplementary Table S8 for details.
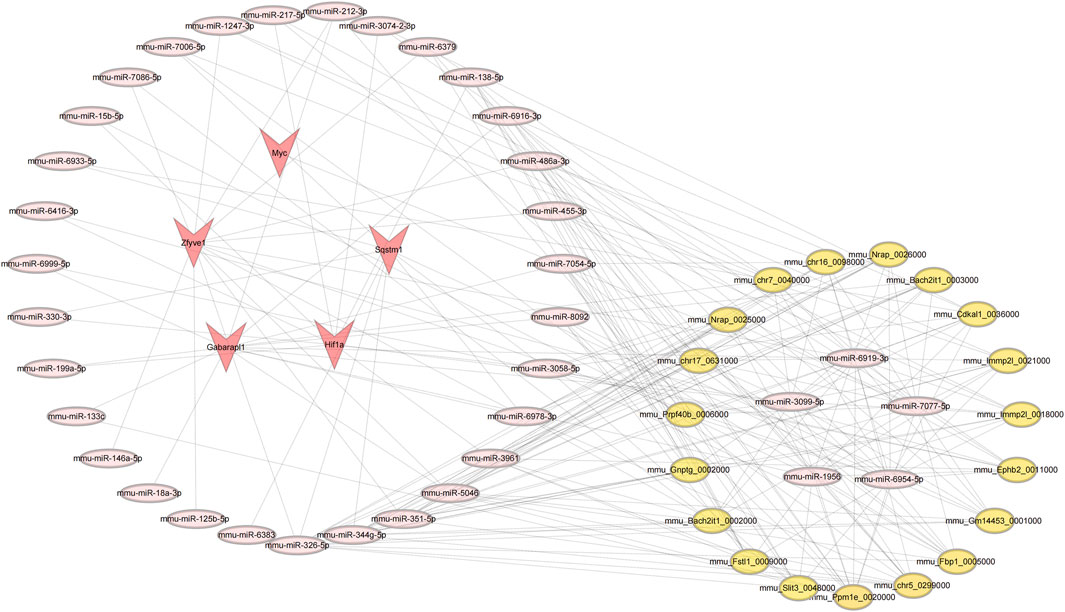
Figure 11. Results of the circRNA-miRNA-mRNA regulatory network. Red represents: mRNA; pink represents: miRNA; yellow represents: circRNA.
3.10 Potential drugs prediction
The study found that Trigonelline and Niacinamide were particularly notable, as they exhibited both the highest enrichment factors and the lowest false discovery rates (FDR values) (Figure 12). See Supplementary Table S9 for details.
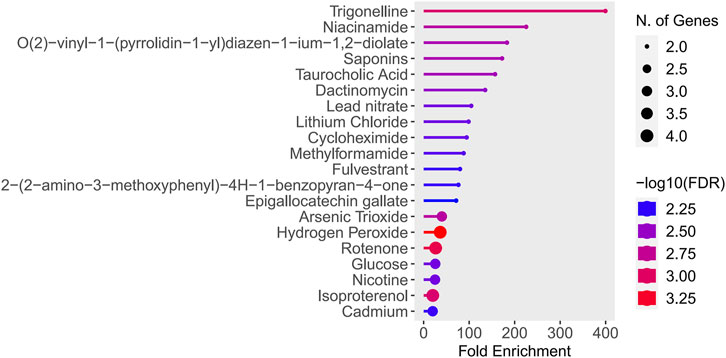
Figure 12. Results of drugs prediction.
3.11 RT-qPCR verification of hub genes
To validate the credibility of the GSE96733 dataset, we conducted qRT-PCR analysis on five autophagy-related genes. Compared to the Sham samples, the expression levels of Myc, Hif1a, Sqstm1, and Gabarapl1 were significantly increased in the II/R injury samples, aligning with the results from the bioinformatics analysis (Figure 13). See Supplementary Table S10 for details.
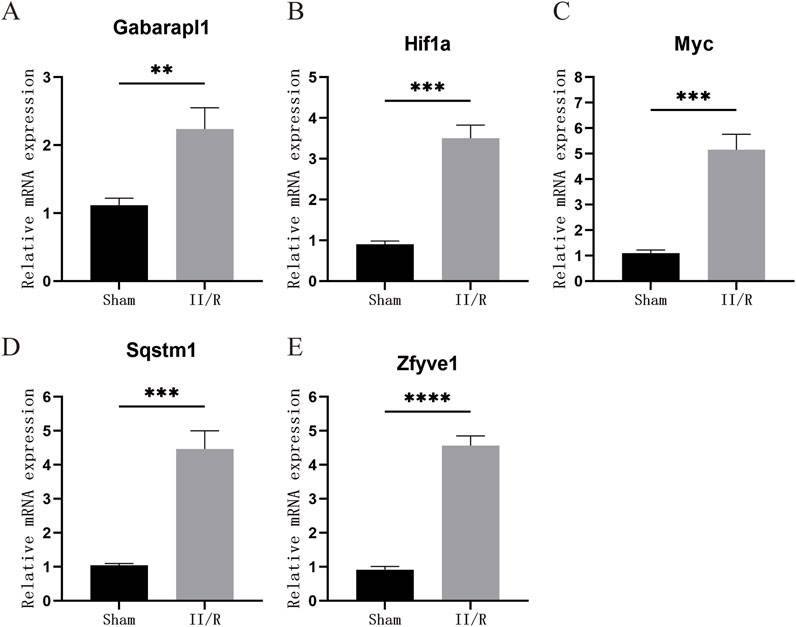
Figure 13. The expression levels of hub genes in the mouse II/R injury model. (A–E) The mRNA levels of Gabarapl1, Hif1a, Myc, Sqstm1, and Zfyve1 were assessed using qRT-PCR. *P < 0.05, **P < 0.01, ***P < 0.001, ****P < 0.0001. ns, not significant.
4 Discussion
II/R injury is a severe clinical condition, yet treatment options remain limited (Nadatani et al., 2018). Developing innovative therapeutic strategies is of significant importance in alleviating II/R injury. In previous studies, the autophagy marker Sqstm1/p62 was downregulated during a phase of 45 min of intestinal ischemia followed by 2 h of reperfusion. During this phase, the activation of autophagy plays a crucial protective role by clearing damaged mitochondria and misfolded proteins, with the mechanism involving the coordinated regulation of multiple molecular pathways (Chen et al., 2020). In the study of Corilagin’s effects on II/R injury, the degradation of Sqstm1 was significantly increased, while the expression of autophagy-related proteins (such as LC3II and Beclin1) was upregulated, suggesting that Corilagin activates autophagic flux through the AMPK/Sirt1 pathway, thereby alleviating oxidative stress and cell apoptosis to exert a protective effect (Li et al., 2023). At the stage of 45 min of intestinal ischemia followed by 4 h of reperfusion, autophagic flux is disrupted (e.g., the fusion of autophagosomes with lysosomes is blocked), leading to the accumulation of Sqstm1 within cells, which is manifested as an upregulation at the protein level. At this point, autophagy shifts from a protective mechanism to a damage-promoting factor (Wang et al., 2019; Wen et al., 2019). Therefore, we selected samples from GSE96733 with 45 min of intestinal ischemia followed by 3 h of reperfusion for our study, indicating that this period represents dynamic changes in autophagy. In the GSE37013 human dataset, whether subjected to 30 min or 120 min of intestinal ischemia-reperfusion, SQSTM1 levels in the II/R group were found to be lower than those in the sham group. This observation indirectly confirms the protective role of autophagy during this phase. It suggests that the 3-h mark may represent a critical turning point, where autophagy transitions from a protective mechanism to a pro-injury factor in II/R injury. However, the relationship between autophagy and II/R injury is not yet fully understood. Additionally, the application of bioinformatics analysis in the context of ARGs in II/R injury has not been thoroughly explored. Therefore, we conducted this study to investigate whether the expression of ARGs in II/R injury significantly differs from that in the sham group. Additionally, we explored the correlation between II/R injury and immune infiltration, which aids in understanding the potential immune mechanisms underlying II/R injury and in identifying potential molecular targets for its treatment.
In this study, we conducted KEGG and GO enrichment analyses using 1,027 DEGs obtained from the database. The KEGG results indicated a significant activation of immune-related disease pathways in II/R injury, particularly the IL-17 signaling pathway. Previous studies have suggested that regulating the production of IL-17 may be advantageous for mitigating tissue damage induced by II/R injury (Edgerton et al., 2009).
GO results indicated that autophagy plays a central role in the development of ischemia/reperfusion (II/R) injuryConsequently, based on these findings, we screened for autophagy-related genes among the DEGs. Subsequently, by utilizing the Lasso algorithm, RF algorithm, and PPI network, we ultimately identified five hub genes: Myc, Hif1a, Zfyve1, Sqstm1, and Gabarapl1. The expression levels of these five hub genes in the raw data showed significant differences between the sham-operated samples and the II/R injury samples. Furthermore, we validated these findings using GSE96733 (II/R 6 h), and the results were consistent with the original bioinformatics analysis. Additionally, we constructed a diagnostic model for II/R injury, which demonstrated good diagnostic efficacy when validated with GSE37013.
CIBERSORT is an inverse deconvolution analysis algorithm based on linear support vector regression that estimates the relative abundance of immune cells in mixed cell populations by analyzing gene expression data (Newman et al., 2015). Using this algorithm, we found that M2 macrophages and activated dendritic cells (DCs) were more abundant in II/R injury samples compared to sham-operated samples. Through correlation analysis of hub genes, we discovered that activated DCs were negatively correlated with Zfyve1 and positively correlated with Sqstm1, Myc, Hif1a, and Gabarapl1. In contrast, the results for immature DCs were opposite to those findings. For M2 macrophages, Zfyve1 was negatively correlated with them, while Sqstm1, Hif1a, and Gabarapl1 were positively correlated.
Macrophages, integral components of the innate immune system, exhibit significant plasticity, allowing them to adjust their phenotypes in response to environmental stimuli. Under homeostatic conditions, macrophages serve a protective role in maintaining the intestinal barrier (Ruder and Becker, 2020). Our earlier research indicated that promoting the transformation of M1 macrophages to M2 may mitigate II/R injury (Liu et al., 2015). Recent studies have demonstrated that enhancing the release of interleukin-10 by M2 macrophages through Toll-like receptor 2 signaling can alleviate II/R injury (Hu et al., 2022). It has been shown that Toll-like receptor (TLR) signaling activates the Nrf2 pathway and induces autophagy via p62, a downstream target of Nrf2. Furthermore, another study revealed that Nrf2 prevents acute lung injury induced by II/R injury by modulating TLR4 signaling and inducing autophagy (Yan et al., 2018). We can hypothesize that Nrf2 may ameliorate II/R injury by regulating TLR-induced autophagy and promoting M2 macrophages, a proposition that necessitates further research for validation. This provides a theoretical basis and new therapeutic strategies for the treatment of II/R injury. Dendritic cells (DCs) serve as crucial regulators of intestinal immunity. Previous studies have demonstrated that II/R injury increases the infiltration of DCs into the small intestine and is associated with the upregulation of TLR4 (Hagiwara et al., 2010). These findings suggest that TLR4 plays a pivotal role in regulating autophagy and immune cells during II/R injury.
Research on non-coding RNAs in II/R injury is becoming increasingly extensive. Studies have demonstrated that inhibiting miR-379-5p can enhance epithelial cell proliferation and improve barrier function following II/R injury (Jia et al., 2023). Suppressing miR-665-3p may represent a potential clinical approach for anti-inflammatory and anti-apoptotic treatment of II/R injury (Li et al., 2018). Additionally, miR-26b-5p may prevent II/R injury by targeting DAPK1 and inhibiting apoptosis in intestinal mucosal cells (Zhou et al., 2022). Our study revealed that multiple microRNAs (including mmu-miR-6954-5p, mmu-miR-6919-3p, mmu-miR-7077-5p, mmu-miR-3099-5p, and mmu-miR-1956) exhibit significantly higher node connectivity within the regulatory network. The Myc oncogene shows a significant interaction with only a single miRNA (mmu-miR-7077-5p). This specific regulatory relationship suggests that selecting Myc as a subsequent validation target offers a unique advantage: it is less influenced by the miRNA regulatory network, thereby facilitating the establishment of a clear molecular regulatory mechanism research system. Currently, no one has studied this RNA, providing a new perspective for research on II/R injury.
Trigonelline is a naturally occurring polar hydrophilic alkaloid found in various plants, including raw coffee beans and fenugreek seeds. It exhibits multiple effects, such as anti-diabetic, neuroprotective, and anti-cancer properties, and it modulates oxidative stress by inhibiting the detrimental Nrf2 pathway when autophagy is compromised (Nguyen et al., 2024). Furthermore, studies have demonstrated that trigonelline inhibits autophagy via the PI3K/AKT/mTOR signaling pathway, thereby mitigating cerebral ischemia-reperfusion pyroptosis (Qiu et al., 2025). Although there is currently no research on the effects of trigonelline on II/R injury, this opens new avenues for drug treatment strategies. The interaction between niacinamide and autophagy has increasingly become a focus of recent research. Oral administration of nicotinamide mononucleotide (NMN) combined with aerobic exercise can elevate the levels of nicotinamide adenine dinucleotide (NAD) in mice and enhance mitophagy (Wu et al., 2023). This also provides theoretical support for studying II/R injury. Further exploration of the specific effects of trigonelline and niacinamide on gene expression and their mechanisms of action is essential, as this will provide a scientific foundation for developing treatments for II/R injury.
Finally, we conducted in vitro experiments to verify whether the hub genes were differentially expressed in the II/R injury mouse model. Based on the results from reverse transcription RT-qPCR, we found that the four hub genes—Sqstm1, Myc, Hif1a, and Gabarapl1—were differentially expressed in the II/R injury group compared to the sham group, with expression trends consistent with the previous bioinformatics analysis results.
Sqstm1/p62 (sequestosome 1) is a classical autophagy receptor that plays a crucial role in selective autophagy, facilitating the elimination of abnormal intracellular components and the recycling of bioenergy substrates (Huang et al., 2024). The elevated RT-qPCR results indicate that autophagic flux is disrupted at 3 h post-reperfusion in II/R injury, transforming autophagy into a pro-damage factor. This finding provides a solid theoretical basis for the early treatment of II/R injury. Myc, a key member of the Myc proto-oncogene family, regulates nearly all physiological processes in cells, including the cell cycle, proliferation, metabolism, differentiation, and apoptosis (Jha et al., 2023). II/R injury can induce distal lung tissue damage by activating oxidative stress and inflammatory responses, while melatonin may exert a protective effect against such damage by upregulating the expression of N-myc downstream regulated gene 2 (NDRG2) (Yang et al., 2015). Although the protective role of Myc in II/R injury has not been directly investigated, this offers indirect theoretical support for its involvement. Studies have demonstrated that hypoxia and hypoxia-inducible factors (Hifs) 1 and 2 alpha are involved in tumor immune escape (Wu et al., 2022). However, the role of Hif1a in the context of II/R injury has not been directly investigated. Chronic hypoxia-induced Hif1a enhances myocardial ischemia tolerance by promoting mitophagy (Alanova et al., 2024). II/R injury is often accompanied by mitochondrial dysfunction and cell death, and Hif1a may help maintain intestinal epithelial cell homeostasis by activating the autophagy pathway. Gabarapl1 (Gamma-aminobutyric acid receptor-associated protein-like 1) is a crucial member of the autophagy-related ATG8 protein family, playing a significant role in various physiological and pathological processes through the regulation of autophagy (Poillet-Perez et al., 2017). Research on Gabarapl1 in the context of II/R injury is limited; however, it may influence the survival of intestinal epithelial cells by regulating the integrity of the autophagic flux, which warrants further investigation.
This study inevitably has several limitations. Firstly, the sample size of the gene expression profiles obtained from public databases is somewhat insufficient, and the individual differences among the samples may influence the generalizability of the analysis results. Furthermore, RT-qPCR only validated the mRNA levels of the hub genes, without confirming the protein levels of these genes. More relevant in vivo and in vitro experiments are necessary to elucidate the roles of these hub genes and their potential mechanisms in II/R injury.
5 Conclusion
In summary, we employed bioinformatics methods to analyze the relationship between autophagy-related genes and II/R injury. This analysis led to the identification of four hub genes—Sqstm1, Myc, Hif1a, and Gabarapl1—that may serve as potential biomarkers for the diagnosis, mechanistic research, and treatment of II/R injury. Furthermore, we conducted an immune cell infiltration analysis, constructed a circRNA-miRNA-mRNA network for the hub genes, and performed potential drug predictions. These findings provide promising indications for identifying targets for subsequent research and treatment.
Data availability statement
The datasets presented in this study can be found in online repositories. The names of the repository/repositories and accession number(s) can be found in the article/Supplementary Material.
Ethics statement
The animal study was approved by the Experimental Animal Ethics Committee of Yunnan Labreal Biotechnology Co., Ltd. The study was conducted in accordance with the local legislation and institutional requirements.
Author contributions
YH: Writing – original draft, Writing – review and editing, Formal Analysis, Data curation. QZ: Visualization, Writing – review and editing, Writing – original draft. YS: Writing – review and editing, Writing – original draft, Visualization. WL: Resources, Funding acquisition, Writing – review and editing. ZY: Writing – review and editing. YZ: Writing – review and editing. KS: Writing – review and editing. HL: Writing – review and editing. JW: Writing – review and editing.
Funding
The author(s) declare that financial support was received for the research and/or publication of this article. This work was funded by the National Natural Science Foundation of China (NSFC) (No. 82460114) and foreign cooperative research project of the Second Affiliated Hospital of Kunming Medical University (No. 2022dwhz09).
Conflict of interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
The reviewer D-LS declared a shared affiliation with the authors YH, QZ, YS, WL, ZY, YZ, KS and HL to the handling editor at the time of review.
Generative AI statement
The author(s) declare that no Generative AI was used in the creation of this manuscript.
Publisher’s note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
Supplementary material
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fphys.2025.1601968/full#supplementary-material
References
Alanova P., Alan L., Opletalova B., Bohuslavova R., Abaffy P., Matejkova K., et al. (2024). HIF-1α limits myocardial infarction by promoting mitophagy in mouse hearts adapted to chronic hypoxia. Acta Physiol. (Oxf) 240 (9), e14202. doi:10.1111/apha.14202
Almoiliqy M., Wen J., Xu B., Sun Y. C., Lian M. Q., Li Y. L., et al. (2020). Cinnamaldehyde protects against rat intestinal ischemia/reperfusion injuries by synergistic inhibition of NF-κB and p53. Acta Pharmacol. Sin. 41 (9), 1208–1222. doi:10.1038/s41401-020-0359-9
Bae T., Hallis S. P., Kwak M. K. (2024). Hypoxia, oxidative stress, and the interplay of HIFs and NRF2 signaling in cancer. Exp. Mol. Med. 56 (3), 501–514. doi:10.1038/s12276-024-01180-8
Cai J., Chen X., Liu X., Li Z., Shi A., Tang X., et al. (2022). AMPK: the key to ischemia-reperfusion injury. J. Cell Physiol. 237 (11), 4079–4096. doi:10.1002/jcp.30875
Cai Y., Wu Y., Guo Z., Ye Y., Zhu Y., Wen L., et al. (2025). Sevoflurane alleviates intestinal ischemia-reperfusion injury in aged mice. Med. Gas. Res. 15, 398–403. doi:10.4103/mgr.MEDGASRES-D-24-00033
Cao Z., Mu S., Wang M., Zhang Y., Zou G., Yuan X., et al. (2023). Succinate pretreatment attenuates intestinal ischemia-reperfusion injury by inhibiting necroptosis and inflammation via upregulating Klf4. Int. Immunopharmacol. 120, 110425. doi:10.1016/j.intimp.2023.110425
Chen R., Zhang Y. Y., Lan J. N., Liu H. M., Li W., Wu Y., et al. (2020). Ischemic postconditioning alleviates intestinal ischemia-reperfusion injury by enhancing autophagy and suppressing oxidative stress through the akt/GSK-3β/nrf2 pathway in mice. Oxid. Med. Cell Longev. 2020, 6954764. doi:10.1155/2020/6954764
Chen W., Ma L., Shao J., Bi C., Li J., Yang W. (2023). miR-185-5p/ATG101 axis alleviated intestinal barrier damage in intestinal ischemia reperfusion through autophagy. Heliyon 9 (7), e18325. doi:10.1016/j.heliyon.2023.e18325
Chen Z., Huang A., Sun J., Jiang T., Qin F. X., Wu A. (2017). Inference of immune cell composition on the expression profiles of mouse tissue. Sci. Rep. 7, 40508. doi:10.1038/srep40508
Edgerton C., Crispín J. C., Moratz C. M., Bettelli E., Oukka M., Simovic M., et al. (2009). IL-17 producing CD4+ T cells mediate accelerated ischemia/reperfusion-induced injury in autoimmunity-prone mice. Clin. Immunol. 130 (3), 313–321. doi:10.1016/j.clim.2008.09.019
Eisele B. S., Silva G. C. V., Bessow C., Donato R., Genro V. K., Cunha-Filho J. S. (2021). An in silico model using prognostic genetic factors for ovarian response in controlled ovarian stimulation: a systematic review. J. Assist. Reprod. Genet. 38 (8), 2007–2020. doi:10.1007/s10815-021-02141-0
Friedman J., Hastie T., Tibshirani R. (2010). Regularization paths for generalized linear models via coordinate descent. J. Stat. Softw. 33 (1), 1–22. doi:10.18637/jss.v033.i01
Gonzalez L. M., Moeser A. J., Blikslager A. T. (2015). Animal models of ischemia-reperfusion-induced intestinal injury: progress and promise for translational research. Am. J. Physiol. Gastrointest. Liver Physiol. 308 (2), G63–G75. doi:10.1152/ajpgi.00112.2013
Gubernatorova E. O., Perez-Chanona E., Koroleva E. P., Jobin C., Tumanov A. V. (2016). Murine model of intestinal ischemia-reperfusion injury. J. Vis. Exp. 111, 53881. doi:10.3791/53881
Hagiwara S., Iwasaka H., Hasegawa A., Asai N., Uchida T., Noguchi T. (2010). Dendritic cell activation in response to ischemia-reperfusion injury of the small intestine. Surg. Today 40 (2), 137–145. doi:10.1007/s00595-009-4033-6
Hanley J. A., McNeil B. J. (1982). The meaning and use of the area under a receiver operating characteristic (ROC) curve. Radiology 143 (1), 29–36. doi:10.1148/radiology.143.1.7063747
Hu J., Deng F., Zhao B., Lin Z., Sun Q., Yang X., et al. (2022). Lactobacillus murinus alleviate intestinal ischemia/reperfusion injury through promoting the release of interleukin-10 from M2 macrophages via Toll-like receptor 2 signaling. Microbiome 10 (1), 38. doi:10.1186/s40168-022-01227-w
Huang X., Liu L., Yao J., Lin C., Xiang T., Yang A. (2024). S-acylation regulates SQSTM1/p62-mediated selective autophagy. Autophagy 20 (6), 1467–1469. doi:10.1080/15548627.2023.2297623
Jha R. K., Kouzine F., Levens D. (2023). MYC function and regulation in physiological perspective. Front. Cell Dev. Biol. 11, 1268275. doi:10.3389/fcell.2023.1268275
Jia Y., Cui R., Wang C., Feng Y., Li Z., Tong Y., et al. (2020). Metformin protects against intestinal ischemia-reperfusion injury and cell pyroptosis via TXNIP-NLRP3-GSDMD pathway. Redox Biol. 32, 101534. doi:10.1016/j.redox.2020.101534
Jia Z., Wang Y., Gao J., Zheng M., Wang P., Zu G. (2023). miR-379-5P Inhibition enhances intestinal epithelial proliferation and barrier function recovery after ischemia/reperfusion by targeting EIF4G2. Shock 60 (4), 594–602. doi:10.1097/shk.0000000000002205
Klionsky D. J., Petroni G., Amaravadi R. K., Baehrecke E. H., Ballabio A., Boya P., et al. (2021). Autophagy in major human diseases. Embo J. 40 (19), e108863. doi:10.15252/embj.2021108863
Li B., Li W., Zheng M., Wang Y., Diao Y., Mou X., et al. (2023). Corilagin alleviates intestinal ischemia/reperfusion injury by relieving oxidative stress and apoptosis via AMPK/Sirt1-autophagy pathway. Exp. Biol. Med. (Maywood) 248 (4), 317–326. doi:10.1177/15353702221147560
Li G., Wang S., Fan Z. (2021). Oxidative stress in intestinal ischemia-reperfusion. Front. Med. (Lausanne) 8, 750731. doi:10.3389/fmed.2021.750731
Li Y., Cao Y., Xiao J., Shang J., Tan Q., Ping F., et al. (2020). Inhibitor of apoptosis-stimulating protein of p53 inhibits ferroptosis and alleviates intestinal ischemia/reperfusion-induced acute lung injury. Cell Death Differ. 27 (9), 2635–2650. doi:10.1038/s41418-020-0528-x
Li Z., Wang G., Feng D., Zu G., Li Y., Shi X., et al. (2018). Targeting the miR-665-3p-ATG4B-autophagy axis relieves inflammation and apoptosis in intestinal ischemia/reperfusion. Cell Death Dis. 9 (5), 483. doi:10.1038/s41419-018-0518-9
Linfert D., Chowdhry T., Rabb H. (2009). Lymphocytes and ischemia-reperfusion injury. Transpl. Rev. Orl. 23 (1), 1–10. doi:10.1016/j.trre.2008.08.003
Liu M., Wen H., Zuo L., Song X., Geng Z., Ge S., et al. (2023a). Bryostatin-1 attenuates intestinal ischemia/reperfusion-induced intestinal barrier dysfunction, inflammation, and oxidative stress via activation of Nrf2/HO-1 signaling. Faseb J. 37 (6), e22948. doi:10.1096/fj.202201540R
Liu S., Yao S., Yang H., Liu S., Wang Y. (2023b). Autophagy: regulator of cell death. Cell Death Dis. 14 (10), 648. doi:10.1038/s41419-023-06154-8
Liu W. F., Wen S. H., Zhan J. H., Li Y. S., Shen J. T., Yang W. J., et al. (2015). Treatment with recombinant Trichinella spiralis cathepsin B-like protein ameliorates intestinal ischemia/reperfusion injury in mice by promoting a switch from M1 to M2 macrophages. J. Immunol. 195 (1), 317–328. doi:10.4049/jimmunol.1401864
Nadatani Y., Watanabe T., Shimada S., Otani K., Tanigawa T., Fujiwara Y. (2018). Microbiome and intestinal ischemia/reperfusion injury. J. Clin. Biochem. Nutr. 63 (1), 26–32. doi:10.3164/jcbn.17-137
Newman A. M., Liu C. L., Green M. R., Gentles A. J., Feng W., Xu Y., et al. (2015). Robust enumeration of cell subsets from tissue expression profiles. Nat. Methods 12 (5), 453–457. doi:10.1038/nmeth.3337
Nguyen V., Taine E. G., Meng D., Cui T., Tan W. (2024). Pharmacological activities, therapeutic effects, and mechanistic actions of trigonelline. Int. J. Mol. Sci. 25 (6), 3385. doi:10.3390/ijms25063385
Pajares M., Jiménez-Moreno N., García-Yagüe Á J., Escoll M., de Ceballos M. L., Van Leuven F., et al. (2016). Transcription factor NFE2L2/NRF2 is a regulator of macroautophagy genes. Autophagy 12 (10), 1902–1916. doi:10.1080/15548627.2016.1208889
Park K., Kim D. (2009). Localized network centrality and essentiality in the yeast-protein interaction network. Proteomics 9 (22), 5143–5154. doi:10.1002/pmic.200900357
Pierro A., Eaton S. (2004). Intestinal ischemia reperfusion injury and multisystem organ failure. Semin. Pediatr. Surg. 13 (1), 11–17. doi:10.1053/j.sempedsurg.2003.09.003
Poillet-Perez L., Jacquet M., Hervouet E., Gauthier T., Fraichard A., Borg C., et al. (2017). GABARAPL1 tumor suppressive function is independent of its conjugation to autophagosomes in MCF-7 breast cancer cells. Oncotarget 8 (34), 55998–56020. doi:10.18632/oncotarget.19639
Qiu Z., Ma J., Zhang X., Jiao M., Zhi L. (2025). Electroacupuncture combined with trigonelline inhibits pyroptosis in cerebral ischemia-reperfusion by suppressing autophagy via the PI3K/AKT/mTOR signaling pathway. Brain Res. Bull. 221, 111200. doi:10.1016/j.brainresbull.2025.111200
Roy P., Kumar B., Shende A., Singh A., Meena A., Ghosal R., et al. (2013). A genome-wide screen indicates correlation between differentiation and expression of metabolism related genes. PLoS One 8 (5), e63670. doi:10.1371/journal.pone.0063670
Ruder B., Becker C. (2020). At the forefront of the mucosal barrier: the role of macrophages in the intestine. Cells 9 (10), 2162. doi:10.3390/cells9102162
Sasaki M., Joh T. (2007). Oxidative stress and ischemia-reperfusion injury in gastrointestinal tract and antioxidant, protective agents. J. Clin. Biochem. Nutr. 40 (1), 1–12. doi:10.3164/jcbn.40.1
Shen X., Shi H., Chen X., Han J., Liu H., Yang J., et al. (2023). Esculetin alleviates inflammation, oxidative stress and apoptosis in intestinal ischemia/reperfusion injury via targeting SIRT3/AMPK/mTOR signaling and regulating autophagy. J. Inflamm. Res. 16, 3655–3667. doi:10.2147/jir.S413941
Walter W., Sánchez-Cabo F., Ricote M. (2015). GOplot: an R package for visually combining expression data with functional analysis. Bioinformatics 31 (17), 2912–2914. doi:10.1093/bioinformatics/btv300
Wang F., Gu L., Wang Y., Sun D., Zhao Y., Meng Q., et al. (2022). MicroRNA-122a aggravates intestinal ischemia/reperfusion injury by promoting pyroptosis via targeting EGFR-NLRP3 signaling pathway. Life Sci. 307, 120863. doi:10.1016/j.lfs.2022.120863
Wang Y., Wen J., Almoiliqy M., Wang Y., Liu Z., Yang X., et al. (2021). Sesamin protects against and ameliorates rat intestinal ischemia/reperfusion injury with involvement of activating Nrf2/HO-1/NQO1 signaling pathway. Oxid. Med. Cell Longev. 2021, 5147069. doi:10.1155/2021/5147069
Wang Z., Li Z., Feng D., Zu G., Li Y., Zhao Y., et al. (2019). Autophagy induction ameliorates inflammatory responses in intestinal ischemia-reperfusion through inhibiting NLRP3 inflammasome activation. Shock 52 (3), 387–395. doi:10.1097/shk.0000000000001259
Watson M. J., Ke B., Shen X. D., Gao F., Busuttil R. W., Kupiec-Weglinski J. W., et al. (2008). Intestinal ischemia/reperfusion injury triggers activation of innate toll-like receptor 4 and adaptive chemokine programs. Transpl. Proc. 40 (10), 3339–3341. doi:10.1016/j.transproceed.2008.07.144
Wen J., Xu B., Sun Y., Lian M., Li Y., Lin Y., et al. (2019). Paeoniflorin protects against intestinal ischemia/reperfusion by activating LKB1/AMPK and promoting autophagy. Pharmacol. Res. 146, 104308. doi:10.1016/j.phrs.2019.104308
Wu Q., You L., Nepovimova E., Heger Z., Wu W., Kuca K., et al. (2022). Hypoxia-inducible factors: master regulators of hypoxic tumor immune escape. J. Hematol. Oncol. 15 (1), 77. doi:10.1186/s13045-022-01292-6
Wu T., Hu E., Xu S., Chen M., Guo P., Dai Z., et al. (2021). clusterProfiler 4.0: a universal enrichment tool for interpreting omics data. Innov. (Camb) 2 (3), 100141. doi:10.1016/j.xinn.2021.100141
Wu W., Yuan S., Tang Y., Meng X., Peng M., Hu Z., et al. (2023). Effect of exercise and oral niacinamide mononucleotide on improving mitochondrial autophagy in alzheimer's disease. Nutrients 15 (13), 2851. doi:10.3390/nu15132851
Yan J., Li J., Zhang L., Sun Y., Jiang J., Huang Y., et al. (2018). Nrf2 protects against acute lung injury and inflammation by modulating TLR4 and Akt signaling. Free Radic. Biol. Med. 121, 78–85. doi:10.1016/j.freeradbiomed.2018.04.557
Yang B., Ni Y. F., Wang W. C., Du H. Y., Zhang H., Zhang L., et al. (2015). Melatonin attenuates intestinal ischemia--reperfusion-induced lung injury in rats by upregulating N-myc downstream-regulated gene 2. J. Surg. Res. 194 (1), 273–280. doi:10.1016/j.jss.2014.11.018
Zhang F. L., Hu Z., Wang Y. F., Zhang W. J., Zhou B. W., Sun Q. S., et al. (2023a). Organoids transplantation attenuates intestinal ischemia/reperfusion injury in mice through L-Malic acid-mediated M2 macrophage polarization. Nat. Commun. 14 (1), 6779. doi:10.1038/s41467-023-42502-0
Zhang J., Liu Z., Liu Y., Shi Y., Chen F., Leng Y. (2023b). Role of non-coding RNA in the pathogenesis of intestinal ischemia- reperfusion injury. Curr. Med. Chem. 30 (36), 4130–4148. doi:10.2174/0929867330666221219094145
Zhao Y., Li L. (2023). The application of a topology optimization algorithm based on the kriging surrogate model in the mirror design and optimization of an aerial camera. Sensors (Basel) 23 (16), 7236. doi:10.3390/s23167236
Zhenzhen L., Wenting L., Jianmin Z., Guangru Z., Disheng L., Zhiyu Z., et al. (2022). miR-146a-5p/TXNIP axis attenuates intestinal ischemia-reperfusion injury by inhibiting autophagy via the PRKAA/mTOR signaling pathway. Biochem. Pharmacol. 197, 114839. doi:10.1016/j.bcp.2021.114839
Zhou B., Zhang W., Yan Z., Zhao B., Zhao J., Feng W., et al. (2022). MicroRNA-26b-5p targets DAPK1 to reduce intestinal ischemia/reperfusion injury via inhibition of intestinal mucosal cell apoptosis. Dig. Dis. Sci. 67 (5), 1794–1805. doi:10.1007/s10620-021-06975-7
Keywords: intestinal ischemia-reperfusion (II/R) injury, Autophagy, immune cell infiltration, bioinformatics analysis, hub genes, machine learning
Citation: Hu Y, Zou Q, Sun Y, Li W, Yin Z, Zhao Y, Shi K, Liu H and Wang J (2025) Identification of autophagy-related genes in intestinal ischemia-reperfusion injury and their role in immune infiltration. Front. Physiol. 16:1601968. doi: 10.3389/fphys.2025.1601968
Received: 28 March 2025; Accepted: 14 May 2025;
Published: 26 May 2025.
Edited by:
Gang Ye, Sichuan Agricultural University, ChinaReviewed by:
Jiawei Yang, Zunyi Medical University, ChinaDa-Li Sun, Kunming Medical University, China
Zixuan Zhong, Chongqing University of Education, China
Copyright © 2025 Hu, Zou, Sun, Li, Yin, Zhao, Shi, Liu and Wang. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Weiming Li, bGl3ZWltaW5nMTAzNzJAMTYzLmNvbQ==
†These authors have contributed equally to this work and share first authorship