 Annette Bruun Jensen
Annette Bruun Jensen David Copplestone2
David Copplestone2 Roberto Guidetti
Roberto Guidetti Paola Pittia
Paola Pittia Åsa Berggren
Åsa Berggren- 1Department of Plant and Environmental Sciences, University of Copenhagen, Frederiksberg, Denmark
- 2Biological and Environmental Sciences, University of Stirling, Stirling, United Kingdom
- 3Department of Life Sciences, University of Modena and Reggio Emilia, Modena, Italy
- 4IU International University of Applied Sciences, Bad Reichenhall, Germany
- 5Institute for Nutritional and Food Sciences, University of Bonn, Bonn, Germany
- 6Department of Bioscience and Technology for Food Agriculture and Environment, University of Teramo, Teramo, Italy
- 7Department of Ecology, Swedish University of Agricultural Sciences, Uppsala, Sweden
As space agencies progress toward long-duration missions and extraterrestrial colonisation, Bioregenerative Life Support Systems (BLSS) have become central to achieving closed-loop sustainability. Edible insects offer a highly efficient protein source suited for BLSS integration, yet the unique stressors of spaceflight, microgravity, ionising radiation, and limited microbial exposure, pose significant risks to insect immunity and pathogen dynamics. This review synthesises current research on insect immune function, microbiome stability, and disease susceptibility under space-relevant conditions, highlighting vulnerabilities introduced by physical, nutritional and behavioural stressors. We emphasise species-specific immune traits, life stage- and sex-dependent responses, and the contribution of natural behaviours and transgenerational immunity to colony resilience. Further, we examine the synergistic effects of the space environment and high-density rearing on pathogen transmission and virulence evolution. Mitigation strategies, including environmental controls, probiotic interventions and biosensor-based health monitoring, are discussed. By identifying critical knowledge gaps, particularly concerning immune suppression under microgravity and radiation, density-driven pathogen evolution, and the stability of behavioural immunity, we propose system-level responses to support robust insect health. Our synthesis advances the framework for designing resilient, health-optimised insect rearing systems for future space missions and terrestrial applications. Ensuring insect immune competence will be essential for ecological stability and food security in extraterrestrial environments.
1 Introduction
As humanity prepares for long-duration space missions and potential colonisation of extraterrestrial environments, developing closed-loop life support systems that can sustain human life independently from Earth is critical (ESA, 2022; NASA, 2023). Bioregenerative Life Support Systems (BLSS) are designed to integrate biological components such as plants, animals and microorganisms to recycle air, water, and waste while producing food and maintaining environmental stability (Berggren et al., 2025). Among candidate organisms for space-based protein production, insects stand out due to their high feed conversion efficiency, minimal space and water requirements, rapid growth, and nutritional richness (Berggren et al., 2019). Among insects, yellow mealworms (Tenebrio molitor) and house crickets (Acheta domesticus) are currently two of the primary species considered for BLSS, due to amongst other things their well-documented nutritional profiles, appreciated flavour and the established rearing knowledge of the species. The role of insects in BLSS has gained increasing scientific interest due to the targeted efforts in space exploration (ESA, 2022; Berggren et al., 2025). Successfully rearing viable insect populations in space will require both fundamental research and targeted development. For many candidate species, key thresholds and optimal conditions remain unknown. However, terrestrial data provide a starting point. Most edible insect species can be reared in controlled modules maintained at approximately 27 °C–30 °C and 50%–70% relative humidity, using plant-based feed (van Huis et al., 2013; Berggren et al., 2019). Ground-based experiments show that silkworm larvae (Bombyx mori) can be reared in controlled and enclosed modules (Tong et al., 2011). Under comparable Earth-based conditions, house crickets can reach harvest size within 8 weeks while requiring less than 2 L of water per kilogram of protein—an important logistical advantage in BLSS. Insects currently approved for consumption within the EU provide 53%–70% digestible protein along with essential micronutrients, making them a valuable complement to plant-based dietary staples (Rumpold and Schlüter, 2013).
However, space introduces novel stressors that challenge the viability of insect rearing. Key environmental variables such as microgravity and ionising radiation can significantly affect insect physiology, especially immune function (Iyer et al., 2022). For instance, space-flown Drosophila melanogaster have demonstrated suppressed innate immune responses and reduced expression of antimicrobial peptides, increasing their vulnerability to pathogens (Marcu et al., 2011; Taylor et al., 2014). Compounding this risk, studies show that pathogenic bacteria such as Serratia marcescens exhibit increased virulence when cultured in microgravity environments, potentially overwhelming weakened insect immune systems (Gilbert et al., 2020; Gilbert et al., 2022). Furthermore, sterile or microbially reduced space habitats may inadvertently contribute to immune dysregulation by limiting natural microbial exposure, a phenomenon also observed in human astronauts (Mermel, 2013). These findings underscore the importance of understanding host-pathogen-environment interactions under space conditions. The resilience of insects in BLSS, and by extension the success of extraterrestrial missions, depends heavily on our ability to maintain insect health, stable populations and productivity. This review aims to synthesise current knowledge on insect immunity and pathogen interactions in the context of spaceflight, with a particular focus on edible insect species relevant to BLSS. We identify critical research gaps and outline future strategies for monitoring, mitigation, and system design to ensure robust insect health in long-duration missions. This review synthesizes current knowledge on how five major spaceflight-relevant stressors: microgravity, radiation, nutrition, rearing density, and behavioural constraints, interact to affect insect health, with implications for immune competence, pathogen risks, and system resilience in BLSS (Figure 1).
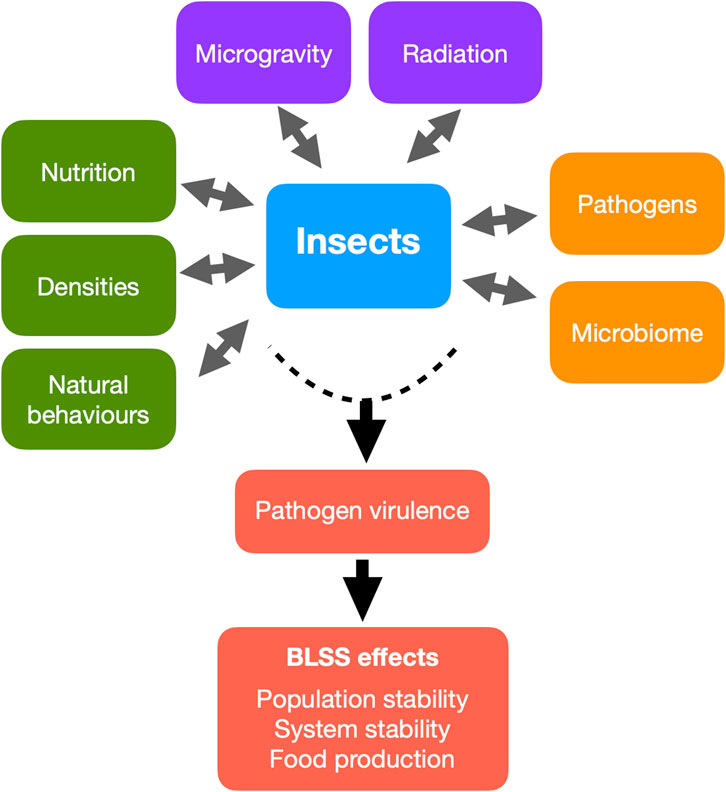
Figure 1. Main factors influencing the health of insects reared in Bioregenerative Life Support Systems in space. These factors can act individually or in interaction with one another to affect immune competence, pathogen risks and system resilience.
2 Insect immunity: a foundation for health in space
The first line of defence in insects is the exoskeleton, serving as a critical physical barrier. Once breached, insects initiate a cascade of cellular and humoral immune responses (Evans et al., 2006). The immune responses are of three different types. First, melanisation through the pro-phenoloxidase cascade generates microbe-toxic melanin (Cerenius and Söderhäll, 2004). Second, haemocytes drive encapsulation, isolating larger pathogens. Third, antimicrobial peptides (AMPs) such as defensins and attacins, lysate invading microbes (Bulet and Stöcklin, 2005). Unlike vertebrates, insects lack adaptive immunity and rely solely on the mechanisms encapsulation, melanisation and antimicrobial peptide production (Vilmos and Kurucz, 1998). Spaceflight has been shown to suppress all three responses in Drosophila, highlighting them as key targets for immune support strategies in BLSS environments (Marcu et al., 2011). The ability of an individual of a species to use these defences is a result of both genetic, evolutionary and environmental factors (Berggren, 2009; Berggren and Low, 2009). The immune functions are central to survival in terrestrial environments and become even more vital under the compounded stressors encountered in space-based systems.
2.1 The role of the insect microbiome in immune resilience
In terrestrial settings, the gut microbiota plays a key role in maintaining immune equilibrium, aiding in nutrient metabolism, immune priming, and pathogen exclusion (Douglas, 2015). This mutualistic relationship influences larval development, environmental responsiveness, and even behaviour. However, in sterilised, microbially constrained conditions of space habitats, these dynamics may be disrupted, leading to dysbiosis and increased susceptibility to opportunistic infections. Reduced microbial exposure and sanitised feed can compromise immune signalling pathways, a phenomenon observed in astronauts (Mermel, 2013) and likely relevant to insects. Though studies in edible insect species under space conditions remain sparse, findings from other systems suggest that gut microbes can affect how insects spread disease, compete with pathogens, and secrete antiviral agents (Engel and Moran, 2013). Experiments already shown that microgravity disrupts host–microbe relationships: it alters gene expression in symbiotic microbes and increases Serratia virulence in Drosophila (Casaburi et al., 2017; Gilbert et al., 2020). Future measures may include tailored probiotic supplementation and co-cultivation with beneficial microbial communities to restore microbial balance and reinforce immune function within BLSS. Probiotics have been seen to boost mealworm performance, although the gains are context dependent. In T. molitor, an indigenous Pediococcus pentosaceus promoted larval growth, whereas a Bacillus strain had the opposite effect (Lecocq et al., 2021), while the endogenous P. pentosaceus and an exogenous Enterococcus faecium strain enhanced metamorphosis and fungal resistance (Dahal et al., 2022). Before BLSS use, candidate strains should pass genome screens for transferable resistance or toxins (van der Fels-Klerx et al., 2018) and used in micro-g/radiation trials confirming they do not raise Salmonella or Listeria risk for humans (Gałęcki et al., 2023). Carrying out multigenerational tests would also verify whether any benefits persist without nutritional or reproductive costs.
2.2 Species-specific immune responses in edible insects
Species- and stage-specific differences in immune traits are critical to designing resilient BLSS systems. In A. domesticus, immune system development shows that phenoloxidase (that is needed to produce melanin) activity increases after metamorphosis, whereas encapsulation responses remain consistent, indicating that immune function varies across life stages (Piñera et al., 2013). Similarly, T. molitor displays cuticle colour-dependent immune investment, with darker morphs exhibiting greater phenoloxidase activity and pathogen resistance (Armitage and Siva-Jothy, 2005; Barnes and Siva-Jothy, 2000). These traits are modulated by environmental stressors such as diet, crowding and light. For example, food deprivation suppresses antimicrobial defences (Siva-Jothy and Thompson, 2002). Crowding during the final nymph stage increases phenoloxidase activity but has little effect on survival to adulthood (Piñera, 2012). In Piñera’s study, survival dropped only slightly in crowded groups (31%–33%) compared to isolated nymphs (41%). In contrast, constant illumination had a much stronger effect, reducing survival from 52% to 21%, despite little change in phenoloxidase levels (Lindberg et al., 2025; Piñera, 2012). These findings indicate that crowding and light stress can modulate immune responses and affect survival through distinct mechanisms. This sensitivity highlights the need to optimise rearing conditions in space, including feed quality and population density. Sex-based immune differences further complicate health dynamics in populations. Males of A. domesticus are more susceptible to the bacteria Serratia liquefaciens infection than females at equivalent doses (Gray, 1998), implying that sex ratios are relevant in BLSS. Additionally, immune activation is costly: elevated investment may impair female growth and reproduction (Bascuñán-García et al., 2010), suggesting trade-offs between immunity and reproduction. Moreover, immune support may extend beyond physiology. Behavioural immunity, including thermoregulatory “fever” responses observed in A. domesticus (Adamo, 1998), and chemical cues used in mate selection may contribute to pathogen avoidance and reproductive success. Chitin, a major exoskeletal component, can stimulate immune activity in insects and potentially also in human consumers and offer an increased gut immunity (Muzzarelli et al., 2012). Lastly, while foundational insights from D. melanogaster have informed our understanding of space-altered immunity (Taylor et al., 2014; Iyer et al., 2022), species-specific studies remain essential due to divergent ecological adaptations and immune systems.
3 Spaceflight physical stressors: microgravity and radiation
Spaceflight introduces two fundamental physical challenges - microgravity and radiation - that significantly alter insect immunity and pathogen dynamics. These stressors not only compromise the insect host’s immune capacity but may also enhance microbial virulence, creating a dual vulnerability that can jeopardise population stability in Bioregenerative Life Support Systems.
3.1 Microgravity-induced immune modulation
Microgravity presents a significant challenge to insect immunity, despite its robustness under terrestrial conditions (Berggren et al., 2025). In D. melanogaster, immune function is consistently suppressed following spaceflight. After 13 days in orbit, Iyer et al. (2022) reported broad downregulation of immune genes and impaired cellular responses, reducing both signalling capacity and effector function. Similarly, Marcu et al. (2011) found that although space-reared flies could clear Escherichia coli upon return to Earth, immune suppression during flight indicated a temporary vulnerability with potential relevance for BLSS conditions. Exposure to fungal pathogens further revealed a sharp decline in the production of key antifungal peptides, including Drosomycin and Metchnikowin (Taylor et al., 2014). Additional work showed that space-raised larvae had approximately 25% fewer actively phagocytosing plasmatocytes (Marcu et al., 2011), and those cells exhibited altered development and reduced pathogen-engulfing capacity after flight (Kaur et al., 2005). Compounding these immune deficits, altered gravity also increases intracellular reactive-oxygen species (ROS), contributing to oxidative stress (Bhattacharya and Hosamani, 2015). At the same time, elevated levels of heat-shock proteins suggest broader stress-induced modulation of immune regulation and inflammatory control (Shepard et al., 2018). In contrast to microgravity’s suppressive effects, brief exposure to hypergravity, at both 1 g and 4 g, enhanced fungal resistance and improved survival, indicating stress-induced immune priming (Taylor et al., 2014). Together, these findings highlight the critical role of gravitational forces in shaping insect immune function. Circadian disruption may compound immune stress. In A. domesticus, constant light nearly halved survival without increasing basal phenoloxidase levels, a pattern also observed under dim night-time lighting in Teleogryllus commodus (Piñera, 2012; Durrant et al., 2020). Space-flown Drosophila similarly become arrhythmic, downregulate clock genes and exhibit combined metabolic and immune costs (Iyer et al., 2022). Until clinostat data and more knowledge on edible species become available, BLSS colonies may benefit from maintaining a 12 h:12 h light–dark cycle and where feasible to help preserve circadian regulation. Additional immune deficits have been observed under real microgravity. In Drosophila spaceflight reduced the fraction of actively phagocytosing plasmatocytes and their phagocytic capacity over time and downregulated pattern-recognition receptors, opsonins, lysozymes and AMP/stress genes indicating coordinated cellular and humoral innate-immune impairments (Marcu et al., 2011).
Although research on edible insects remains limited, studies in A. domesticus suggest that both nerve cell development and neuroimmune interactions may be sensitive to gravity. Exposure of eggs and early-instar larvae to altered gravity disrupted the physiology of position-sensitive interneurons, indicating that early neural development can be gravity-dependent (Horn et al., 2002; Kirschnick et al., 2007). This is significant because insect immune responses are partially regulated through neuroendocrine signalling, linking neural development to immune competence (Goldsworthy et al., 2003; Urbański et al., 2022). Neurosecretory amines, tachykinin peptides (Urbański et al., 2022), and adipokinetic hormone (Goldsworthy et al., 2003) bind to haemocyte receptors and modulate key immune functions such as phagocytosis, nodulation, and melanisation. Gravity-induced changes in neurogenesis could therefore impair immunity by disrupting this neuro-immune axis, highlighting its integrative and bidirectional role in insect health. Collectively, these findings and the broader synthesis of Berggren et al. (2025), reveal that microgravity disrupts both immune balance and cellular maturation, potentially compromising insect population viability in space. Further research in BLSS-relevant species is urgently needed to determine species-specific thresholds and to guide the development for systems for long-duration missions.
3.2 Radiation-induced immune suppression and pathogen synergy
In parallel with microgravity, ionising radiation presents a second major environmental threat to insect health. Space-relevant radiation includes chronic exposure to galactic cosmic rays and solar particle events, both capable of inducing cellular damage and immunosuppression. Evidence from cotton leafworm Spodoptera littolaris larvae shows that exposure to gamma radiation at 50, 100, and 150 Gy leads to a dose-dependent decline in key immune enzymes, including phenoloxidase and lysozyme (Gabarty et al., 2013). These effects were significantly exacerbated in the presence of fungal infection, highlighting a synergistic interaction between physical stress and pathogen pressure. Radiation also affects the pathogens themselves. A single high-charge, high-energy particle reduced Bacillus subtilis spore germination by more than 90%, while kilogray-level gamma irradiation nearly inactivated Trichoplusia ni nucleopolyhedrovirus and greatly reduced the viability of Beauveria and Metarhizium (Jaques, 1968; Bücker and Horneck, 1975; Rodrigues et al., 2016). These findings suggest that cosmic rays and solar particles could lessen pathogen pressure in space-based insect farms. However, dose–response data remain scarce for most microbiome and pathogen taxa, making it difficult to predict ecological outcomes under long-term exposure. This dual impact, on both host immunity and pathogen viability, may be particularly consequential in BLSS, where containment of insect pathogens is essential.
Overall, radiation weakens insect immunity at both humoral and cellular levels, potentially increasing vulnerability to otherwise non-lethal infections. Species vary in their sensitivity to radiation, underscoring the need for targeted shielding strategies and careful selection of candidate species. While most current data derive from short-term exposures, chronic low-dose radiation in orbit or on planetary surfaces may have cumulative effects on immune function and transgenerational health. Future research should examine how space radiation effects physical and behavioural responses in edible insects as well as microbiome communities. Integrating shielding, dosimetry and biological monitoring into habitat design will be essential for maintaining insect health and ensuring the long-term viability of insect-based food systems.
4 Pathogen dynamics in edible insect production: interactions with environmental stressors
Edible insects offer a sustainable protein source for both Earth and space applications, but the rearing under high-density and controlled conditions mean that they have significant disease risks. Insects are vulnerable to a diverse array of pathogens: including viruses, bacteria, fungi, microsporidia, and gregarines, that can lead to population collapse and production loss (Eilenberg et al., 2015; Lecocq et al., 2019; Maciel-Vergara et al., 2021). Although these pathogens do not infect humans directly, their effects on population health and productivity mean a great need for rigorous disease management in both terrestrial and space-based systems.
4.1 Viral pathogens and host vulnerability
Viruses represent one of the most severe threats to insect rearing. Outbreaks of Acheta domesticus densovirus (AdDV) have caused catastrophic population losses (Maciel-Vergara and Ros, 2017), and recent studies continue to reveal novel viral agents, including iflaviruses in both wild and captive crickets (de Miranda et al., 2021a; de Miranda et al., 2021b). A novel densovirus have also caused high mortality in a commercially reared T. molitor (Armién et al., 2023). These viruses persist within dense rearing systems, often amplified through close contact and shared environments. In black soldier flies, Pienaar et al. (2022) documented both current and historic viral interactions, illustrating how rearing conditions shape virome evolution. These findings underscore the need for robust viral surveillance and biosecurity in insect rearing, especially where high density is a norm.
4.2 Bacterial threats and environmental entry routes
Bacteria infiltrate insect colonies via contaminated feed or environments and can induce systemic infections. Pseudomonas aeruginosa, for example, spreads effectively in dense populations, especially through cannibalism and contact with contaminated frass (Maciel-Vergara et al., 2018). Additionally, insects may carry zoonotic bacteria like Salmonella spp. and Listeria monocytogenes when raised on infected feed, posing indirect food safety risks (Osimani et al., 2018; van der Fels-Klerx et al., 2018). These outcomes highlight the importance of hygienic feed and controlled environment in the mitigation of pathogens.
4.3 Fungal and protozoan infections in confined systems
Entomopathogenic fungi, including Beauveria bassiana and Metarhizium anisopliae, are frequent in T. molitor rearing environments and exploit humid, unsanitised conditions and cause mortality via cuticle penetration (Eilenberg et al., 2015; Lecocq et al., 2019). Similarly, microsporidia and gregarines, though less frequently reported, infect tissues and disrupt reproduction, particularly under stress or poor sanitation (Maciel-Vergara et al., 2021). These parasites remain underdiagnosed due to limited detection tools, yet may become increasingly relevant in compact, resource-limited habitats like those in BLSS.
4.4 Density as a central driver of transmission and pathogen evolution
Among all controllable variables in insect farming, population density stands out as a dominant factor influencing disease emergence and spread. Crowded conditions elevate pathogen transmission through increased physical contact, coprophagy, and cannibalism. In A. domesticus, high-density rearing has been linked to repeated outbreaks of Acheta domesticus densovirus (AdDV), invertebrate iridovirus 6 (IIV-6), and novel iflaviruses, all of which can persist and spread via direct contact and contaminated substrates (de Miranda et al., 2021a; de Miranda et al., 2021b). Virome analyses also show divergence between wild and cultivated cricket virus strains, possibly reflecting evolutionary pressures in commercial systems that favour enhanced transmissibility (de Miranda et al., 2021b). Cannibalism in the giant mealworm Zophobas morio has proven an efficient transmission route for Pseudomonas aeruginosa and other pathogens under suboptimal hygiene (Maciel-Vergara et al., 2018). These findings reinforce the need for biosecurity protocols tailored to high-density conditions. When individuals are kept close together it may be harder to detect infections, especially when symptoms are mild, like in AdDV-infected crickets that show less movement (Low et al., 2022). While density’s role in pathogen dynamics is clear, its broader implications for insect health and behaviour extend beyond disease ecology.
4.5 Nutritional modulation of pathogen susceptibility
Nutritional quality plays a subtle yet critical role in shaping insect susceptibility to infection. Feed composition influences immune function and microbial carriage: nutrient-poor diets in A. domesticus and T. molitor reduce protein levels, disrupt fat balance, and impair immunocompetence (Harsányi et al., 2020). In Hermetia illucens larvae, diet composition significantly alters gut microbiota, suggesting that tailored feed formulations can support microbial functions linked to digestion and larval health (Li et al., 2024). Protein availability also modulates defence. In Spodoptera littoralis challenged with nucleopolyhedrovirus, protein-biased diets improved survival and elevated constitutive immunity (lysozyme-like activity, encapsulation and phenoloxidase), and infected survivors actively selected more protein-rich foods - consistent with protein costs of resistance (Lee et al., 2006). By contrast, in adult Drosophila infected with Micrococcus luteus, flies reduced protein intake and survival improved on a low-protein, high-carbohydrate diet, with a simultaneous increase in the baseline production of antimicrobial peptides - showing that the best protein-to-carbohydrate ratio for immunity depends on context and life stage (Ponton et al., 2020) Among nutritional components, lipids play a particularly important immunomodulatory role. Balanced lipid profiles enhance immune performance by improving haemocyte membrane fluidity, which promotes receptor clustering and phenoloxidase-mediated pathogen clearance (Li et al., 2023). Specific fatty acids also serve as precursors for eicosanoids, lipid-derived signalling molecules essential for nodulation, haemocyte spreading and antimicrobial peptide production (Kim and Stanley, 2021). These effects are supported by diets with appropriate ratios of polyunsaturated to saturated fatty acids (Orkusz et al., 2024). Finally, feed hygiene also contributes to immune resilience: a recent systematic review found low Salmonella prevalence in farmed insects, linking pathogen suppression to clean feed and substrates (Marzoli et al., 2024). Together, these findings underscore the dual role of feed in supporting both growth and disease resistance.
4.6 Mitigation strategies: integrated controls for pathogen management
To safeguard insect production systems, especially in BLSS, integrated biosecurity protocols are essential. Core measures include sanitation of facilities, pathogen screening (particularly for viruses and zoonotic bacteria), and quarantining of new stock (Semberg et al., 2018; de Miranda et al., 2021a; de Miranda et al., 2021b). Environmental regulation, humidity, temperature, and especially density control is vital for stress reduction and disease suppression (van Huis, 2021). Furthermore, standardised protocols across feed, hygiene, and rearing environments are necessary to ensure resilience and reproducibility in the systems (Berggren et al., 2019). Emerging biotechnologies may offer additional layers of biosecurity for space-based insect farming. In Anopheles gambiae, CRISPR-mediated deletion of the FREP1 gene reduced Plasmodium infection by more than 95%, and when paired with a male-biasing gene drive, the trait reached fixation within approximately ten generations (Dong et al., 2018; Simoni et al., 2020). Similar contained genetic modifications could eventually be used to enhance pathogen resistance in edible insect colonies.
5 Rearing density and pathogen dynamics
In space-based insect production, rearing density is a central design parameter due to spatial constraints and the drive for high-efficiency biomass generation. However, elevated density introduces significant risks to insect health by amplifying pathogen transmission, altering behaviour, and intensifying stress. These effects are well-documented across terrestrial insect farming and are likely to be exacerbated in confined BLSS, where environmental buffering is limited. Beyond facilitating transmission, high-density rearing environments can shape pathogen evolution. The selection for faster-replicating or more virulent strains in dense populations poses long-term biosecurity concerns, particularly in BLSS where microbial escape and containment options are limited. In A. domesticus, producers define overcrowding not by biomass alone, but by physiological stress responses. Laboratory trials with approximately 30 nymphs per 0.5 L enclosure showed elevated phenoloxidase levels, indicating immune strain (Piñera, 2012). On European farms, outbreaks of densovirus and iflavirus have been linked to high-contact rearing conditions, where frass accumulation and cannibalism facilitate viral transmission (de Miranda et al., 2021a; de Miranda et al., 2021b). At comparable densities, T. molitor are prone to overheating and fungal epizootics when ventilation is inadequate (Eilenberg et al., 2015; Maciel-Vergara et al., 2021). Microgravity may exacerbate these risks by impairing air exchange. To mitigate potential effects, BLSS insect colonies may benefit from initiating stocking at no more than 80% of terrestrial density, with continuous monitoring of respiration and behaviour to detect early signs of distress (Low et al., 2022). However, the lack of studies in this area makes predictions difficult. Compounding these issues, behavioural changes linked to infection such as reduced activity or delayed emergence observed in AdDV-infected crickets, can reduce behavioural variability within colonies, making it more difficult to detect early signs of illness (Low et al., 2022). This phenomenon presents a diagnostic challenge, especially in systems lacking continuous monitoring. Moreover, density-related stress may suppress behavioural immunity, the natural defence mechanisms like grooming, avoidance, or thermoregulation that help insects mitigate pathogen exposure. Confinement reduces space for these behaviours to manifest, further increasing disease vulnerability. Effective density management, therefore, must go beyond avoiding overcrowding; it must actively support behavioural expression, hygiene, and population health surveillance. To mitigate these risks, a greater understanding of density affects for insect species based on their immune profiles, pathogen susceptibility, and behavioural ecology is needed. Environmental enrichment, optimised feed allocation, and harvesting methods may reduce stress and disease pressure. Integrating density-sensitive biosensors and behaviour-recognition systems may also increase the possibility to detect outbreaks early and enable proactive management.
6 Natural behaviours as immunological buffers
While insect immunity is often framed in physiological terms, behavioural and transgenerational adaptations play equally vital roles in sustaining health and resilience, particularly under the multiple stressors of space habitats. In confined and resource-limited systems like BLSS, these behaviours may serve as crucial front-line defences, complementing or even compensating for physiologically weakened immunity due to microgravity, radiation, or nutritional constraints.
6.1 Behavioural immunity and environmental navigation
Behavioural immunity encompasses pathogen avoidance behaviours such as grooming, thermoregulation, anorexia, and selective oviposition, all of which reduce pathogen exposure and infection risk (de Roode and Lefèvre, 2012). For example, A. domesticus displays fever-like thermoregulatory behaviour in response to intracellular infection, demonstrating a capacity for adaptive thermal regulation in immune defence (Adamo, 1998). Similarly, T. molitor larvae can detect and avoid faeces contaminated with pathogen-contaminated feed (Vigneron et al., 2019; Goerlinger et al., 2024). Beyond avoidance, insects can modify mating and foraging behaviours based on immune status or pathogen load. Male beetles may learn to avoid contaminated substrates, while females may exhibit selective mate choice based on chemical cues reflecting immune competence. Infection-driven behavioural shifts such as anorexia, lethargy, or altered movement patterns are not merely symptomatic but adaptive responses that reallocate energy toward immune functions (Adamo, 2006). These behaviours are regulated by hormones and connects the immune system to behaviours (Nunes et al., 2021). However, spaceflight conditions such as reduced gravity and confinement, may suppress these responses, limiting insects’ ability to perform protective behaviours and thereby elevate the infection risk.
6.2 Transgenerational immune priming
In addition to real-time behavioural defences, some insects exhibit transgenerational immune priming (TGIP), where parental immune experience enhances the immune function of offspring. In the greater wax moth Galleria mellonella, maternal ingestion of bacteria leads to the deposition of bacterial fragments into eggs, triggering upregulation of immune-related genes in the progeny (Freitak et al., 2014). TGIP appears to be underpinned by heritable epigenetic changes. In Manduca sexta, parental exposure to bacterial challenge altered DNA methylation and histone acetylation patterns in the offspring, leading to increased expression of immune-related genes (Gegner et al., 2019). Similarly, in bumblebees, primed queens produced daughters with constitutively upregulated antimicrobial and pattern-recognition genes, suggesting an epigenetically “prepared” immune state (Barribeau et al., 2016). Similarly, in T. molitor, both maternal and paternal pathogen exposure has been shown to confer elevated pathogen resistance to larvae (Zanchi et al., 2012). TGIP may prove particularly advantageous for multigenerational stability in BLSS, where immune traits acquired in one generation could buffer the next against endemic pathogens or environmental stress. Moreover, the heritability of behavioural immunity and immune investment suggests potential for selective breeding programs to enhance colony robustness over successive generations in space. However, the maintenance of TGIP and behavioural traits under space stressors remain uncertain. Confined conditions, limited mate choice, and altered sensory cues may impair behaviourally mediated disease avoidance. Likewise, chronic exposure to microgravity and radiation may disrupt the stability or effectiveness of inherited immune traits. Understanding how these factors interact with immune memory and behaviour is essential for engineering resilient insect colonies. Maintaining environments that support species-specific behaviours through spatial design, feed and temperature control, may help preserve these natural immune buffers. Further research into genetics and environmental constraints of behavioural and transgenerational immunity will be key to ensuring long-term insect health.
7 Integrated system-level responses: monitoring and managing insect health
Maintaining insect health in Bioregenerative Life Support Systems requires continuous, adaptive monitoring capable of detecting physiological stress, disease outbreaks, and environmental imbalances before they escalate. Given the operational constraints of space missions including limited crew time, isolation, and system autonomy; biosensors, AI-driven diagnostics, and automation could be valuable components in resilient insect rearing systems. These tools would serve as an operational interface between insect health and environmental management, helping to detect deviations across all five key stress axes: microgravity, radiation, nutrition, density, and behavioural suppression. Biosensors capable of detecting volatile organic compounds, changes in humidity or temperature, or fluctuations in activity levels could provide early-warning signals of population distress or pathogen presence (Li et al., 2021; Asiri et al., 2025). For instance, disease progression in insects is often accompanied by changes in odour profiles, metabolic rates, or thermal regulation - all parameters detectable by electrochemical or optical sensors. When deployed in enclosed habitats, these non-invasive technologies could enable real-time surveillance. Hand-held qPCR units, flight-tested aboard the International Space Station, can detect Salmonella enterica and other pathogens directly from feed, water, or surface swabs in under an hour (Khodadad et al., 2021). Complementary CRISPR/Cas12a-based biosensors have also been developed to identify Listeria monocytogenes in insect powders, making them suitable for continuous monitoring of frass and feed (Li et al., 2021). When integrated with optical and AI-based surveillance systems, these lightweight diagnostic tools could form part of a fully automated early-warning system for BLSS insect rearing. Deep learning frameworks are already widely used in insect detection, as shown in a recent review of over 90 studies (Teixeira et al., 2023), and could be adapted to support health monitoring in space-optimised production systems. These platforms could automate alerts for density thresholds, behavioural suppression, or suboptimal environmental conditions that would allow for rapid corrective actions. While promising, the technological readiness of many sensor and AI systems remains variable. Space deployment requires miniaturization, low energy consumption, and autonomous functionality, which are not always compatible with current commercial platforms. Nonetheless, modular diagnostics embedded into rearing infrastructure represent a scalable solution for ensuring the real-time health of insect populations. To fully operationalise these technologies, sensor outputs must be tightly coupled with rearing protocols. For instance, behavioural deviations might trigger automated adjustments in light cycles, substrate distribution, or ventilation, while biosensor alerts could initiate diagnostic imaging or quarantine of affected units. As insect-based BLSS become more complex and multigenerational, system-level monitoring will become increasingly central not just to detect disease, but to ensure long-term viability, optimise productivity and support ecological processes. These technologies will likely form the backbone of precision bioengineering for insect health in space.
8 Research gaps and future directions
Despite advances in insect physiology, ecology, microbiology and space systems design, significant knowledge gaps remain across the five key stressors that shape insect health and disease dynamics in BLSS. Addressing these gaps is essential for ensuring the long-term viability of insect-based bioregenerative systems for space exploration.
8.1 Microgravity and immune competence
Microgravity-induced immune suppression is one of the most well-documented yet poorly understood challenges. D. melanogaster studies reveal downregulated immune gene expression and diminished cellular immunity following spaceflight (Iyer et al., 2022), but equivalent data in edible insect species is lacking. Comparative studies are needed to understand how microgravity alters immune system development, behaviour, and pathogen resistance across insect taxa. Investigating gravity-sensitive phases such as embryogenesis and moulting may yield insights into critical windows for immune disruption. Furthermore, the reversibility of space-induced immunosuppression, as suggested by Marcu et al. (2011), warrants systematic evaluation under BLSS conditions.
8.2 Radiation and pathogen interactions
Space radiation presents a cumulative threat to insect immune systems, with few studies addressing long-term, low-dose exposures relevant to orbital or planetary environments. Existing research in S. littolaris has shown gamma radiation suppresses phenoloxidase and lysozyme activity, especially when paired with fungal pathogens (Gabarty et al., 2013), but mechanisms remain unclear. Moreover, radiation’s impact on microbiome stability and transgenerational immune function is virtually unexplored. Future studies should simulate galactic cosmic rays and solar particle events to evaluate chronic impacts on insect health and virome evolution in enclosed systems.
8.3 Nutrition and microbial ecosystem design
Nutritional inputs affect not only insect growth and productivity but also immune robustness and microbiota composition. However, the link between diet, microbial balance, and immune outcomes in space remains speculative. While nutrient-rich feeds improve lipid profiles and pathogen resistance in T. molitor and A. domesticus (Harsányi et al., 2020; Orkusz et al., 2024), the effects of sterilised or pre-processed space feeds on gut health and immune development remain unknown. Research should include microbiome-informed feed and probiotic strategies tools to promote gut stability in sterile environments.
8.4 Density management and pathogen evolution
High-density rearing accelerates pathogen transmission and may promote virulence evolution. While several studies have shown increased viral loads and behavioural suppression under crowded conditions (de Miranda et al., 2021a; Low et al., 2022), population levels for density-induced stress and infection susceptibility remain undefined. Tools for real-time density diagnostics and automated behavioural monitoring could aid in establishing safe operational ranges. Longitudinal studies of pathogen adaptation under confinement are also needed to assess the risk of virulence escalation and to develop strain monitoring protocols.
8.5 Behavioural immunity and heritable resilience
Behavioural adaptations like thermoregulation, grooming, and selective oviposition reduce pathogen exposure, yet these behaviours may be suppressed in confined space habitats. The stability of these behaviours, and of transgenerational immune priming, under microgravity and chronic space stressors is still unknown. While TGIP has been documented in G. mellonella and T. molitor (Zanchi et al., 2012; Freitak et al., 2014), its persistence across multiple generations in BLSS has not been tested. Research into the genetic foundation for these and how the environmental can affect this is vital for long-term population resilience.
9 Conclusion
Insects offer a compelling solution for sustainable protein production and nutrient recycling in space, particularly within Bioregenerative Life Support Systems. However, their viability in closed-loop habitats hinges not only on productivity, but on the ability to maintain robust immune function and population resilience under a suite of spaceflight-specific stressors. This review identifies five key factors - microgravity, radiation, nutrition, rearing density, and behavioural constraints - that interact to shape insect immunity, pathogen dynamics, and microbial balance in space environments (Figure 1). Microgravity and ionising radiation disrupt immune gene expression, cellular maturation, and host-pathogen interactions, posing fundamental challenges to insect resilience. Nutritional inputs modulate immunity and microbiota composition, while high-density rearing conditions accelerate disease transmission and may select for more virulent pathogens. Compounding these risks, behavioural immunity and transgenerational priming that are two natural buffers against infection, may be suppressed in confined or simplified habitats and weaken long-term population stability. Ensuring the success of insect-based BLSS will require a systems-level approach that integrates environmental design, real-time health monitoring and adaptive management. Biosensors, AI diagnostics and automation are likely very useful tools to detect early signs of stress, optimise conditions, and manage pathogen outbreaks across these five axes. Moreover, the development of predictive models that simulate multi-generational effects and multi-factorial stress interactions will be essential for mission planning and habitat design. Ultimately, insect immune health should not be viewed in isolation, but as a central determinant of ecological stability and nutritional reliability in space. Through interdisciplinary research we can develop adaptive insect farming that are resilient not only to the known constraints of space travel, but to the evolving challenges of life beyond Earth.
Author contributions
AJ: Writing – original draft, Validation, Conceptualization. DC: Writing – review and editing, Conceptualization. RG: Conceptualization, Writing – review and editing. MH: Writing – review and editing, Conceptualization. PP: Conceptualization, Writing – review and editing. ÅB: Writing – original draft, Visualization, Conceptualization.
Funding
The author(s) declare that financial support was received for the research and/or publication of this article. The European Space Agency (ESA) granted support to the Topical Team “Potential of insects as nutritional food in spaceflight (PINS)” no. 4000136088 that formed the base for this study.
Acknowledgments
We are thankful to ESA for granting support to the Topical team ‘Potential of insects as nutritional food in spaceflight (PINS) no. 4000136088 and to the International Society for Gravitational Physiology (ISGP) for supporting the scientific subcommission ‘Gravitational-related challenges for insect rearing and food production for spaceflight and colonisations’.
Conflict of interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Generative AI statement
The author(s) declare that Generative AI was used in the creation of this manuscript. Generative AI was used to locate specific published studies and for some of the language editing.
Any alternative text (alt text) provided alongside figures in this article has been generated by Frontiers with the support of artificial intelligence and reasonable efforts have been made to ensure accuracy, including review by the authors wherever possible. If you identify any issues, please contact us.
Publisher’s note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
References
Adamo S. A. (1998). The specificity of behavioral fever in the cricket Acheta domesticus. J. Parasitol. 84 (3), 529–533. doi:10.2307/3284717
Adamo S. A. (2006). Comparative psychoneuroimmunology: evidence from the insects. Behav. Cogn. Neurosci. Rev. 5 (3), 128–140. doi:10.1177/1534582306289580
Armién A. G., Polon R., Rejmanek D., Moeller R. B., Crossley B. M. (2023). Outbreak of densovirus with high mortality in a commercial mealworm (Tenebrio molitor) farm: a molecular, bright-field, and electron microscopic characterization. Vet. Pathol. 60, 689–703. doi:10.1177/03009858231180488
Armitage S. A. O., Siva-Jothy M. T. (2005). Immune function responds to selection for cuticular colour in Tenebrio molitor. Heredity 94, 650–656. doi:10.1038/sj.hdy.6800675
Asiri A., Perkins S. E., Müller C. T. (2025). The smell of infection: disease surveillance in insects using volatile organic compounds. Agric. For. Entomol. 27 (1), 81–89. doi:10.1111/afe.12651
Barnes A. I., Siva-Jothy M. T. (2000). Density-dependent prophylaxis in the mealworm beetle Tenebrio molitor L. (Coleoptera: tenebrionidae): cuticular melanization is an indicator of investment in immunity. Proc. R. Soc. B 267, 177–182. doi:10.1098/rspb.2000.0984
Barribeau S. M., Schmid-Hempel P., Sadd B. M. (2016). Royal decree: gene expression in trans-generationally immune primed bumblebee workers mimics a primary immune response. PLoS ONE 11 (7), e0159635. doi:10.1371/journal.pone.0159635
Bascuñán-García A. P., Lara C., Córdoba-Aguilar A. (2010). Immune investment impairs growth, female reproduction and survival in the house cricket, Acheta domesticus. J. Insect Physiol. 56 (2), 204–211. doi:10.1016/j.jinsphys.2009.10.005
Berggren Å. (2009). Effect of landscape and population variables on immune response in experimentally introduced bush-cricket populations. Landsc. Ecol. 24, 749–757. doi:10.1007/s10980-009-9348-6
Berggren Å., Low M. (2009). The relationship between morphological symmetry and immune response in wild-caught adult bush-crickets. Symmetry 1, 106–114. doi:10.3390/sym1020106
Berggren Å., Jansson A., Low M. (2019). Approaching ecological sustainability in the emerging insects-as-food industry. Trends Ecol. Evol. 34 (2), 132–138. doi:10.1016/j.tree.2018.11.005
Berggren Å., Jensen A. B., Copplestone D., Guidetti R., Heer M., Pittia P., et al. (2025). Insects in outer space: assessing the effects of microgravity on edible and model insect species for spaceflight food system. Front. Physiol. 16, 1622401. (in press). doi:10.3389/fphys.2025.1622401
Bhattacharya S., Hosamani R. (2015). “Altered gravity induces oxidative stress in Drosophila melanogaster (NASA Technical Report No. ARC-E-DAA-TN27793),” in Presented at the Annual Meeting of the American Society for Gravitational and Space Research, Alexandria, VA
Bücker H., Horneck G. (1975). The biological effectiveness of HZE-particles of cosmic radiation studied in the apollo 16 and 17 Biostack experiments. Acta Astronaut. 2 (3–4), 247–264. doi:10.1016/0094-5765(75)90095-8
Bulet P., Stöcklin R. (2005). Insect antimicrobial peptides: structures, properties and gene regulation. Protein & Peptide Lett. 12 (1), 3–11. doi:10.2174/0929866053406011
Casaburi G., Goncharenko-Foster I., Duscher A. A., Foster J. S. (2017). Transcriptomic changes in an animal-bacterial symbiosis under modeled microgravity conditions. Sci. Rep. 7, 46318. doi:10.1038/srep46318
Cerenius L., Söderhäll K. (2004). The prophenoloxidase-activating system in invertebrates. Immunol. Rev. 198 (1), 116–126. doi:10.1111/j.0105-2896.2004.00116.x
Dahal S., Jensen A. B., Lecocq A. (2022). Effect of probiotics on Tenebrio molitor larval development and resistance against the fungal pathogen Metarhizium brunneum. Insects 13, 1114. doi:10.3390/insects13121114
de Miranda J. R., Granberg F., Low M., Onorati P., Semberg E., Jansson A., et al. (2021a). Virus diversity and loads in crickets reared for feed: implications for husbandry. Front. Vet. Sci. 8, 642085. doi:10.3389/fvets.2021.642085
de Miranda J. R., Granberg F., Onorati P., Jansson A., Berggren Å. (2021b). Virus prospecting in crickets—discovery and strain divergence of a novel iflavirus in wild and cultivated Acheta domesticus. Viruses 13, 364. doi:10.3390/v13030364
de Roode J. C., Lefèvre T. (2012). Behavioral immunity in insects. Insects 3, 789–820. doi:10.3390/insects3030789
Dong Y., Simões M. L., Marois E., Dimopoulos G. (2018). CRISPR/Cas9-mediated gene knockout of Anopheles gambiae FREP1 suppresses malaria parasite infection. PLoS Pathog. 14 (3), e1006898. doi:10.1371/journal.ppat.1006898
Douglas A. E. (2015). Multiorganismal insects: diversity and function of resident microorganisms. Annu. Rev. Entomol. 60, 17–34. doi:10.1146/annurev-ento-010814-020822
Durrant J., Green M. P., Jones T. M. (2020). Dim artificial light at night reduces the cellular immune response of the black field cricket, Teleogryllus commodus. Insect Sci. 27 (3), 571–582. doi:10.1111/1744-7917.12665
Eilenberg J., Vlak J. M., Nielsen-LeRoux C., Cappellozza S., Jensen A. B. (2015). Diseases in insects produced for food and feed. Insects Food Feed 1, 87–102. doi:10.3920/JIFF2014.0022
Engel P., Moran N. A. (2013). The gut microbiota of insects – diversity in structure and function. FEMS Microbiol. Rev. 37, 699–735. doi:10.1111/1574-6976.12025
ESA (European Space Agency) (2022). Terrae novae 2030+ strategy roadmap. Available online at: https://esamultimedia.esa.int/docs/Terrae_Novae_2030_strategy.pdf.doi:10.5270/esa-2022-tn2030
Evans J. D., Aronstein K., Chen Y. P., Hetru C., Imler J. L., Jiang H., et al. (2006). Immune pathways and defence mechanisms in honey bees apis mellifera. Insect Mol. Biol. 15, 645–656. doi:10.1111/j.1365-2583.2006.00682.x
Freitak D., Schmidtberg H., Dickel F., Lochnit G., Vogel H., Vilcinskas A. (2014). The maternal transfer of bacteria can mediate trans-generational immune priming in insects. Virulence 5 (4), 547–554. doi:10.4161/viru.28367
Gabarty A., El-Sonbaty S. M., Ibrahim A. A. (2013). Synergistic effect of gamma radiation and entomopathogenic fungi Beauveria bassiana and Metarhizium anisopliae on the humoral immune enzyme response in cotton leafworm Spodoptera littoralis (boisd). Egypt. Acad. J. Biol. Sci. A, Entomol. 6 (3), 1–10. doi:10.21608/eajbsa.2013.13221
Gałęcki R., Bakuła T., Gołaszewski J. (2023). Foodborne diseases in the edible insect industry in Europe—New challenges and old problems. Foods 12 (4), 770. doi:10.3390/foods12040770
Gegner J., Baudach A., Mukherjee K., Halitschke R., Vogel H., Vilcinskas A. (2019). Epigenetic mechanisms are involved in sex-specific trans-generational immune priming in the lepidopteran model host Manduca sexta. Front. Physiol. 10, 137. doi:10.3389/fphys.2019.00137
Gilbert R., Torres M., Clemens R., Hateley S., Hosamani R., Wade W., et al. (2020). Spaceflight and simulated microgravity conditions increase virulence of Serratia marcescens in the Drosophila melanogaster infection model. npj Microgravity 6, 4. doi:10.1038/s41526-019-0091-2
Gilbert R., Tanenbaum N., Bhattacharya S. (2022). Asparagine biosynthesis as a mechanism of increased host lethality induced by Serratia marcescens in simulated microgravity environments. Heliyon 8, e09379. doi:10.1016/j.heliyon.2022.e09379
Goerlinger A., Develay C., Balourdet A., Rigaud T., Moret Y. (2024). Infection risk by oral contamination does not induce immune priming in the mealworm beetle (Tenebrio molitor) but triggers behavioral and physiological responses. Front. Immunol. 15, 1354046. doi:10.3389/fimmu.2024.1354046
Goldsworthy G. J., Chandrakant S., Opoku-Ware K. (2003). Adipokinetic hormone enhances nodule formation and phenoloxidase activation in adult locusts injected with bacterial lipopolysaccharide. J. Insect Physiol. 49 (8), 795–803. doi:10.1016/S0022-1910(03)00118-5
Gray D. A. (1998). Sex differences in susceptibility of house crickets, Acheta domesticus, to experimental infection with Serratia liquefaciens. J. Invertebr. Pathol. 71, 288–289. doi:10.1006/jipa.1997.4742
Harsányi E., Juhász C., Kovács E., Huzsvai L., Pintér R., Fekete G., et al. (2020). Evaluation of organic wastes as substrates for rearing Zophobas morio, Tenebrio molitor, and Acheta domesticus larvae as alternative feed supplements. Insects 11, 604. doi:10.3390/insects11090604
Horn E., Agricola H., Boser S., Forster S., Kamper G., Riewe P., et al. (2002). Crickets in space: morphological, physiological and behavioral alterations induced by space flight and hypergravity. Adv. Space Res. 30 (4), 819–828. doi:10.1016/S0273-1177(01)00642-1
Iyer J., Mhatre S. D., Gilbert R., Bhattacharya S. (2022). Multi-system responses to altered gravity and spaceflight: insights from Drosophila melanogaster. Neurosci. Biobehav. Rev. 142, 104880. doi:10.1016/j.neubiorev.2022.104880
Jaques R. P. (1968). The inactivation of the nuclear polyhedrosis virus of trichoplusia ni by gamma and ultraviolet radiation. Can. J. Microbiol. 14, 1161–1163. doi:10.1139/m68-194
Kaur I., Simons E. R., Castro V. A., Ott C. M., Pierson D. L. (2005). Changes in monocyte functions of astronauts. Brain Behav. Immun. 19 (6), 547–554. doi:10.1016/j.bbi.2004.12.006
Khodadad C. L. M., Oubre C. M., Castro V. A., Flint S. M., Roman M. C., Ott C. M., et al. (2021). A microbial monitoring system demonstrated on the international space station provides a successful platform for detection of targeted microorganisms. Life 11 (6), 492. doi:10.3390/life11060492
Kim Y., Stanley D. (2021). Eicosanoid signaling in insect immunology: new genes and unresolved issues. Genes 12 (2), 211. doi:10.3390/genes12020211
Kirschnick U., Agricola H.-J., Horn E. (2007). Effects of altered gravity on identified peptidergic neurons of the cricket Acheta domesticus. Gravit. Space Res. 19 (2).
Lecocq A., Jensen A. B., Eilenberg J. (2019). Diseases of insects in European production systems: diagnosis, prevention and management. Berl. Münch. Tierärztl. Wochenschr. 132, 245–250. doi:10.2376/0005-9366-18061
Lecocq A., Natsopoulou M. E., Berggreen I. E., Eilenberg J., Heckmann L. H. L., Nielsen H. V., et al. (2021). Probiotic properties of an Indigenous Pediococcus pentosaceus strain on Tenebrio molitor larval growth and survival. J. Insects Food Feed 7, 975–986. doi:10.3920/JIFF2020.0156
Lee K. P., Cory J. S., Wilson K., Raubenheimer D., Simpson S. J. (2006). Flexible diet choice offsets protein costs of pathogen resistance in a caterpillar. Proc. R. Soc. B 273, 823–829. doi:10.1098/rspb.2005.3385
Li F., Ye Q., Chen M., Zhou B., Zhang J., Pang R., et al. (2021). An ultrasensitive CRISPR/Cas12a-based electrochemical biosensor for Listeria monocytogenes detection. Biosens. Bioelectron. 179, 113073. doi:10.1016/j.bios.2021.113073
Li J., Upadhyay S. K., Bajaj P., Tu W.-C., Liu L., Wang J., et al. (2023). Lipids as a key element of insect defense systems. Front. Genet. 14, 1183659. doi:10.3389/fgene.2023.1183659
Li X., Zhou S., Xiong Q. (2024). Metagenomics analysis of the gut microbiome structure and function in black soldier fly (Hermetia illucens) larvae reared with different types of diet. J. Entomol. Sci. 59. Article 42. doi:10.18474/jes24-42
Lindberg F. A., Waern I., Nilsson E., Jansson A., Holm L., Roman E. (2025). Effect of fasting and two different photoperiods on immune parameters in adult male and female house crickets (Acheta domesticus). Comp. Immunol. Rep. 8, 200210. doi:10.1016/j.cirep.2025.200210
Low M., Eksell I., Jansson A., Berggren Å. (2022). Viral infection changes the expression of personality traits in an insect species reared for consumption. Sci. Rep. 12, 9503. doi:10.1038/s41598-022-13735-8
Maciel-Vergara G., Ros V. I. D. (2017). Viruses of insects reared for food and feed. J. Invertebr. Pathol. 147, 60–75. doi:10.1016/j.jip.2017.01.013
Maciel-Vergara G., Jensen A. B., Eilenberg J. (2018). Cannibalism as a possible entry route for opportunistic pathogenic bacteria to insect hosts, exemplified by Pseudomonas aeruginosa, a pathogen of the giant mealworm Zophobas morio. Insects 9, 88. doi:10.3390/insects9030088
Maciel-Vergara G., Jensen A. B., Lecocq A., Eilenberg J. (2021). Diseases in edible insect rearing systems. J. Insects Food Feed 7, 621–638. doi:10.3920/JIFF2021.0024
Marcu O., Lera M. P., Sanchez M. E., Levic E., Higgins L. A., Shmygelska A., et al. (2011). Innate immune responses of Drosophila melanogaster are altered by spaceflight. PLoS ONE 6, e15361. doi:10.1371/journal.pone.0015361
Marzoli F., Bertola M., Fazion J. P., Cento G., Antonelli P., Dolzan B., et al. (2024). A systematic review on the occurrence of salmonella in farmed Tenebrio molitor and Acheta domesticus or their derived products. Int. J. Food Microbiol. 410, 110464. doi:10.1016/j.ijfoodmicro.2023.110464
Mermel L. A. (2013). Infection prevention and control during prolonged human space travel. Clin. Infect. Dis. 56 (1), 123–130. doi:10.1093/cid/cis861
Muzzarelli R. A. A., El Mehtedi M., Bottegoni C. (2012). Dietary chitin and chitosan: beneficial effects and practical applications. Mar. Drugs 10, 1673–1698. doi:10.3390/md10081673
NASA (National Aeronautics and Space Administration) (2023). NASA’s moon to Mars strategy and objectives development: a blueprint for sustained human presence and exploration throughout the solar system. Washington, DC: NASA. doi:10.25923/5hyb-5x34
Nunes C., Sucena É., Koyama T. (2021). Endocrine regulation of immunity in insects. FEBS J. 288, 3928–3947. doi:10.1111/febs.15581
Orkusz A., Dymińska L., Banaś K., Harasym J. (2024). Chemical and nutritional fat profile of Acheta domesticus, gryllus bimaculatus, Tenebrio molitor, and Rhynchophorus ferrugineus. Foods 13, 32. doi:10.3390/foods13010032
Osimani A., Milanović V., Cardinali F., Garofalo C., Clementi F., Ruschioni S., et al. (2018). The bacterial biota of laboratory-reared edible mealworms (Tenebrio molitor L.): from feed to frass. Int. J. Food Microbiol. 272, 49–60. doi:10.1016/j.ijfoodmicro.2018.03.001
Pienaar R. D., Gilbert C., Belliardo C., Herrero S., Herniou E. A. (2022). First evidence of past and present interactions between viruses and the black soldier fly, Hermetia illucens. Viruses 14, 1274. doi:10.3390/v14061274
Piñera A. (2012). The maturation of the immune system and the effects of crowding and light stress during development on the immune function of the adult house cricket Acheta domesticus. Available online at: http://rave.ohiolink.edu/etdc/view?acc_num=miami1344570343.
Piñera A., Adamo S. A., Johnson L., Killian K. A. (2013). Maturation of the immune system of the male house cricket, Acheta domesticus. J. Insect Physiol. 59, 752–760. doi:10.1016/j.jinsphys.2013.05.008
Ponton F., Morimoto J., Robinson K., Kumar S. S., Cotter S. C., Wilson K., et al. (2020). Macronutrients modulate survival to infection and immunity in drosophila. J. Anim. Ecol. 89, 460–470. doi:10.1111/1365-2656.13126
Rodrigues I. M. W., Forim M. R., da Silva M. F. G. F., Fernandes J. B., Batista Filho A. (2016). Effect of ultraviolet radiation on fungi Beauveria bassiana and Metarhizium anisopliae, pure and encapsulated, and bio-insecticide action on Diatraea saccharalis. Adv. Entomol. 4 (3), 151–162. doi:10.4236/ae.2016.43016
Rumpold B. A., Schlüter O. K. (2013). Nutritional composition and safety aspects of edible insects. Mol. Nutr. Food Res. 57, 802–823. doi:10.1002/mnfr.201200735
Semberg E., de Miranda J. R., Low M., Jansson A., Forsgren E., Berggren Å. (2018). Diagnostic protocols for the detection of Acheta domesticus densovirus (AdDV) in cricket frass. J. Virol. Methods 264, 61–64. doi:10.1016/j.jviromet.2018.12.003
Shepard J., Hosamani R., Bhattacharya S. (2018). “Stress-induced heat shock protein 40 and immune function in flies,” in Presented at the Annual Meeting of the American Society for Gravitational and Space Research (ASGSR), Bethesda, MD. NASA Technical Report 20180007482
Simoni A., Hammond A. M., Beaghton A. K., Galizi R., Taxiarchi C., Kyrou K., et al. (2020). A male-biased sex-distorter gene drive for the human malaria vector Anopheles gambiae. Nat. Biotechnol. 38 (9), 1054–1060. doi:10.1038/s41587-020-0508-1
Siva-Jothy M. T., Thomson J. J. W. (2002). Short-term nutrient deprivation affects immune function. Physiol. Entomol. 27, 206–212. doi:10.1046/j.1365-3032.2002.00286.x
Taylor K., Kleinhesselink K., George M. D., Morgan R., Smallwood T., Hammonds A. S., et al. (2014). Toll-mediated infection response is altered by gravity and spaceflight in drosophila. PLoS ONE 9, e86485. doi:10.1371/journal.pone.0086485
Teixeira A. C., Ribeiro J., Morais R., Sousa J. J., Cunha A. (2023). A systematic review on automatic insect detection using deep learning. Agriculture 13 (3), 713. doi:10.3390/agriculture13030713
Tong L., Yu X., Liu H. (2011). Insect food for astronauts: gas exchange in silkworms fed on mulberry and lettuce and the nutritional value of these insects for human consumption during deep space flights. Bull. Entomol. Res. 101, 613–622. doi:10.1017/S0007485311000228
Urbański A., Johnston P., Bittermann E., Keshavarz M., Paris V., Walkowiak-Nowicka K., et al. (2022). Tachykinin-related peptides modulate immune-gene expression in the mealworm beetle Tenebrio molitor L. L. Sci. Rep. 12, 17277. doi:10.1038/s41598-022-21605-6
van der Fels-Klerx H. J., Camenzuli L., Belluco S., Meijer N., Ricci A. (2018). Food safety issues related to uses of insects for feed and food. Compr. Rev. Food Sci. Food Saf. 17 (5), 1172–1183. doi:10.1111/1541-4337.12385
van Huis A. (2021). Prospects of insects as food and feed. Agric 11 (2), 301–308. doi:10.1007/s13165-020-00290-7
van Huis A., Van Itterbeeck J., Klunder H., Halloran A., Mertens E., Vantomme P. (2013). Edible insects: future prospects for food and feed security. Food Agric. Organ. U. N. FAO Forestry Paper 171).
Vigneron A., Jehan C., Rigaud T., Moret Y. (2019). Immune defenses of a beneficial pest: the mealworm beetle, Tenebrio molitor. Front. Physiol. 10, 138. doi:10.3389/fphys.2019.00138
Vilmos P., Kurucz E. (1998). Insect immunity: evolutionary roots of the mammalian innate immune system. Immunol. Lett. 62, 59–66. doi:10.1016/S0165-2478(98)00023-6
Keywords: edible insects, space agriculture, food, microbiology, pathogen virulence
Citation: Jensen AB, Copplestone D, Guidetti R, Heer M, Pittia P and Berggren Å (2025) Immune challenges and pathogen risks in edible insects: safeguarding health in space life-support systems. Front. Physiol. 16:1628696. doi: 10.3389/fphys.2025.1628696
Received: 14 May 2025; Accepted: 13 August 2025;
Published: 03 September 2025.
Edited by:
Marc-Antoine Custaud, Université d'Angers, FranceReviewed by:
Judith-Irina Buchheim, LMU Munich University Hospital, GermanyRafael Cardoso Maciel Costa Silva, Rio de Janeiro State University, Brazil
Copyright © 2025 Jensen, Copplestone, Guidetti, Heer, Pittia and Berggren. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Åsa Berggren, YXNhLmJlcmdncmVuQHNsdS5zZQ==