 Önder Yıldırım1†
Önder Yıldırım1† Ümit Acar
Ümit Acar Rifat Tezel
Rifat Tezel Sercan Yapıcı
Sercan Yapıcı- 1Faculty of Fisheries, Muğla Sıtkı Koçman University, Muğla, Türkiye
- 2Bayramiç Vocational School, Çanakkale Onsekiz Mart University, Çanakkale, Türkiye
- 3Faculty of Science, Bartın University, Bartın, Türkiye
Flupyradifurone (FPF), a systemic butenolide insecticide introduced in 2014, is increasingly used as an alternative to neonicotinoids, yet its safety for non-target aquatic organisms remains poorly understood. This study evaluated the acute and sub-lethal toxicity of FPF in juvenile common carp (Cyprinus carpio). A 96-h static bioassay determined an LC50 of 140.47 mg/L. Fish were then exposed for 14 days to sub-lethal concentrations (1, 3, 5, 25, 75 and 125 mg/L) to assess hematological, biochemical, and genotoxic responses. Hematological analysis revealed significant, dose-dependent declines in red blood cells (1.71 × 106/μL in control vs. 1.12 × 106/μL at 125 mg/L), hemoglobin (8.34 vs. 3.34 g/dL), and hematocrit (26.08% vs. 13.73%), accompanied by reduced mean corpuscular volume, mean corpuscular hemoglobin, and mean corpuscular hemoglobin concentration at higher doses, indicating anemia and impaired oxygen transport. Biochemically, glucose increased sharply (102.21 mmol/L in control to 230.29 mmol/L at 125 mg/L), while triglycerides, cholesterol, total protein, and albumin declined significantly, suggesting metabolic disruption. Hepatic enzyme activities (alkaline phosphatase, serum glutamic oxaloacetic transaminase, serum glutamate pyruvate transaminase) increased markedly, with serum glutamic oxaloacetic transaminase rising from 36.47 U/L in controls to 144.02 U/L at 125 mg/L, indicative of hepatocellular damage. Comet assay confirmed pronounced DNA damage at ≥25 mg/L, with significant elevations in tail length, tail moment, and % DNA in tail. Collectively, these results demonstrate that FPF exposure compromises hematological health, disrupts metabolic balance, and induces genotoxicity in common carp, even at sub-lethal concentrations. Incorporating both physiological and genomic endpoints is essential for comprehensive ecological risk assessments of emerging insecticides.
1 Introduction
Pesticides are essential for managing pests that threaten agricultural production; nonetheless, their extensive use has generated considerable ecological and health issues. Residual pesticides may pollute surface water via runoff, endangering aquatic organisms and infiltrating the human food chain, potentially affecting human health (Campbell et al., 2016). These substances are associated with extensive environmental problems, including soil and water contamination, as well as persistent negative effects on ecosystem structure and function. Pesticide residues can accumulate over time, leading to alterations in biodiversity, disruption of trophic interactions, and impairment of ecosystem services such as nutrient cycling and water purification. Such ongoing disturbances may gradually undermine the resilience and stability of ecosystems. The toxicity of pesticides to non-target species is particularly concerning, since it can disrupt ecological balance and pose risks to food safety (Cheng et al., 2020). Escalating resistance to widely used pesticides, including neonicotinoids, has heightened the desire for novel pest management options (Bass et al., 2015), but they too entail hazards. Due to the interplay of environmental stresses such as climate change, urbanization, and increased agriculture, there is an imperative need for enhanced pesticide risk assessments that include both lethal and sub-lethal impacts on ecosystems and human health (Goulson et al., 2015).
Flupyradifurone (FPF) is a systemic butenolide insecticide launched in 2014 as a substitute for neonicotinoids, especially for controlling neonicotinoid-resistant pests (Nauen et al., 2015). It acts as a nicotinic acetylcholine receptor (nAChR) agonist, similar to neonicotinoids, although with a unique pharmacophore structure (Jeschke et al., 2015). FPF is used on several crops, including tomatoes, grapes, and citrus, to target pests such as aphids, whiteflies, and thrips (Bayer, 2013). Although FPF is often used as a replacement for imidacloprid in agriculture (Goodhue et al., 2020), its long-term ecological effects, especially on aquatic organisms, are inadequately researched (Siviter et al., 2023). Although the concentrations of flupyradifurone reported in natural waters are relatively low, the highest level recorded to date reached 0.16 μg/L in the Great Lakes basin (Metcalfe et al., 2019). However, due to its high solubility and persistence, there is concern that long-term agricultural use may lead to accumulation in freshwater ecosystems. Therefore, understanding both lethal and sublethal effects in fish is crucial to assess the ecological risks of FPF.
Sublethal exposure to pesticides often causes pronounced alterations in fish hematological and biochemical parameters, which are widely recognized as sensitive indicators of physiological stress and health status. Hematological indices such as red blood cell count, hemoglobin concentration, and hematocrit provide insight into oxygen transport and anemia induced by toxicants, while changes in leukocyte counts reflect immunological disturbances (Adhikari et al., 2004; Ramesh and Saravanan, 2009; Öz et al., 2024). Biochemical markers are equally important: elevated plasma glucose is a common response to stress, whereas alterations in total protein and albumin indicate impaired hepatic protein synthesis (Yilmaz et al., 2021). Hepatic enzymes including alanine aminotransferase (ALT), aspartate aminotransferase (AST), and alkaline phosphatase (ALP) are biomarkers of liver dysfunction, and their elevation signals hepatocellular damage under pesticide stress (Oruc and Üner, 1999). In addition, acetylcholinesterase (AChE) inhibition is a well-documented biomarker of neurotoxicity caused by several insecticides (Banaee et al., 2013). Collectively, these hematological and biochemical endpoints provide an integrated picture of how sublethal pesticide exposure disrupts multiple physiological systems in fish, making them highly relevant for assessing the toxic potential of novel insecticides such as FPF.
The common carp (Cyprinus carpio), belonging to the Cyprinidae family, is a resilient freshwater fish species native to Eurasia. This species has been extensively spread globally due to their adaptability and durability and it is now occurring in several countries outside its native range. Common carp thrive in a variety of aquatic environments, notably preferring slow-moving water bodies such as rivers with diminished flow, ponds, and lakes. They have a remarkable capacity to tolerate well different environmental conditions (Ji et al., 2012). Their resilience, together with their low maintenance needs and compatibility with many species, have contributed to rend common carp one of the favorite species to be used in scientific research especially those aiming to examine the impact of environmental pollutants on aquatic organisms. Their extensive distribution and accessibility further increase their appropriateness for such trials. In addition to their ecological relevance, common carp also hold substantial commercial value in many regions (Khoshnood, 2024).
Therefore, the objective of this study was to determine the dose-dependent hemato-biochemical alterations and genotoxic responses in common carp (C. carpio) exposed to different concentrations of flupyradifurone under controlled laboratory conditions. By establishing median lethal concentration (LC50) values and evaluating blood parameters, serum biochemistry, and comet assay results after sublethal exposure, this study aims to provide new insights into the potential ecological risks of this insecticide to freshwater fish species.
2 Materials and methods
2.1 Fish and experimental conditions
The research protocol received approval from the institutional ethics board (MSKU-HADYEK Local Ethics Commission of Animal Research, Reference number: 2024/10-1). Juvenile common carp (C. carpio) were sourced from the Central Fisheries Research Institute in Antalya, Türkiye. The fish were distributed among 50 L glass aquaria at a density of 10 fish per tank in triplicate and given a week to adapt to their new environment and fed twice daily at a rate of 2% their body weight with commercial pellets containing 28% protein and 3.5% lipid (Tetra, Germany).
Flupyradifurone (FPF) was purchased from Sigma-Aldrich (St. Louis, MO, United States) with a reported purity of 99.9% (Lot# BCCB1463). A stock solution was prepared at a concentration of 100 mg/mL using dechlorinated water, given the high-water solubility of the compound (3.2 g/L at 20 °C). The stock solution was stored in amber-colored vials at +4 °C in the dark. Prior to the experiment, it was appropriately diluted with water to achieve the desired exposure concentrations. To establish the LC50 value, common carp were first subjected to a preliminary range-finding test to identify appropriate concentration intervals of flupyradifurone. Based on the observed mortality after 96 h in the preliminary test, the definitive assay was conducted in triplicate at concentrations of 0, 1, 3, 5, 25, 75, 125, 175, 225, and 325 mg/L using static aquariums, with 10 fish per aquarium (30 aquariums in total; mean body weight 20.80 ± 0.73 g). Fish were not fed during the exposure period, in accordance with standard acute toxicity testing protocols. Daily mortality was recorded, and the median lethal concentration (LC50) and 95% confidence limits were calculated using Probit analysis (Finney, 1952), following the procedure described in AAT Bioquest (2025). During the 96 h-LC50 experiment, fish were left to starving. The mortality was checked twice daily, and the dead fish were taken out instantly. Based on the LC50 results, concentrations of 0, 1, 3, 5, 25, 75 and 125 mg/L were chosen to evaluate the fish’s physiological reactions to the pesticide. The specimens, with an average weight of 21.77 ± 0.64 g, were exposed to pesticide for a 2-week period. During the chronic exposure period, fish were fed a commercial pelleted diet (40% CP and 12% CL) suitable for C. carpio at a rate of approximately 2% of body weight per day. Feeding was conducted once daily, and any uneaten feed was removed to maintain water quality. Exposure solutions were renewed every 48 h with 100% replacement to maintain the stability of pesticide concentrations. Water renewal was conducted carefully to minimize handling stress, and all physicochemical parameters were matched to the previous conditions. This procedure follows semi-static test protocols recommended in OECD guidelines. Each treatment group consisted of three replicate tanks, each containing 10 fish, resulting in 30 fish per treatment concentration. After the chronic exposure period, blood samples were collected from fish sedated with 20 mg/L clove oil. For tissue sampling, fish were humanely euthanized using a higher clove oil concentration (200 mg/L), following the protocol described by Yılmaz and Ergün (2018).
Throughout the experiment water parameters were measured daily and water conditions were carefully regulated to suit carp requirements. The temperature was maintained at 24.68 °C ± 0.14 °C and 24.35 °C ± 0.31 °C for 96 h-LC50 and 14-day exposure treatments respectively. The oxygen content was observed as 6.44 ± 0.27 mg/L and 6.39 ± 0.32 mg/L for 96 h-LC50 and 14-day exposure treatments respectively.
2.2 Hematological and biochemical analysis
Prior to blood collection, the region between the caudal and anal fins was thoroughly sanitized with an alcohol solution to eliminate any potential contamination from mucus. A sterile plastic syringe was then used to extract blood from the caudal vein in this cleansed area. The collected blood was divided into two portions: one was transferred to tubes containing heparin for hematological analysis, while the other was distributed between K3EDTA tubes and gel serum tubes for biochemical and immunological evaluations, respectively.
To obtain serum, the gel tubes were subjected to centrifugation at 4 °C at 3,500 g for a duration of 15 min. Hematological parameters such as in red blood cells (RBC) count, hemoglobin (Hb) levels, and hematocrit (Hct) were evaluated using an automated blood analyzer (Mindray BC3000) that had been specifically calibrated for fish blood samples. The erythrocyte indices such as mean corpuscular volume (MCV), mean corpuscular hemoglobin (MCH), and mean corpuscular hemoglobin concentration (MCHC) were subsequently calculated based on these measurements use formulae below:
A range of biochemical markers were assessed using photometric techniques, employing test kits supplied by Bioanalytic Diagnostic Industry Co. These markers included: Glucose (GLU), Triglycerides (TRI), Cholesterol (CHOL), Total protein (TPROT), Albumin (ALB), Alkaline phosphatase (ALP), Serum glutamic oxaloacetic transaminase (SGOT); Serum glutamate pyruvate transaminase (SGPT).
2.3 Comet assay
Whole blood sample of 2 μL was mixed with 80 μL of low-melting-point agarose. This mixture was then carefully transferred onto microscope slides previously prepared with a coating of 1% high-melting-point agarose. The slides were covered with coverslips and kept in a dark environment at +4 °C for 10-15 min to allow the agarose to set. After removing the coverslips, the slides were submerged in a prepared lysis solution (2.5 M NaCl, 100 mM EDTA, 10 mM Tris, pH 10). Samples were then transferred to a freshly made working solution, created by adding 1% Triton X-100% and 10% DMSO to the lysis buffer. Following the lysis process, the slides were positioned in a horizontal electrophoresis apparatus (Cleaver Scientific, United Kingdom). The voltage was set to 25 V with a current of 300 mA, and electrophoresis was conducted for 30 min. Post-electrophoresis, the slides underwent three 5-min rinses with a neutralizing buffer (0.4 M Tris, pH 7.5) at +4 °C. For visualization, 50 μL of ethidium bromide solution (20 μg/mL) was applied to each slide, which was then covered with a coverslip. The prepared slides were examined under a fluorescence microscope (Carl Zeiss/Scope A1, Germany). Images of the comet test were taken for analysis and head diameter, tail length, and fluorescence intensity of the samples were calculated in the analysis program of the microscope (Zeiss ZEN blue edition software). At least 25 cells were evaluated for each control and experimental group. DNA damage was determined by measuring head and tail lengths, the ratio of tail length to total DNA length (% DNA tail), and tail moment length (Burlinson, 2012).
2.4 Statistical analysis
Data analysis was performed using SPSS 22.0 software (SPSS Inc., Chicago, United States) and reported as mean ± SD and median (min-max). Shapiro-Wilk normality tests and Levene’s Homogeneity test were used to analyze the normality of data and homogeneity of variances, respectively. One-way analysis of variance (ANOVA) was used to determine whether there was variation among treatments in the parameters where assumptions were met. Tukey and Tamhane’s T2 Post-Hoc tests were used to determine differences between groups. In cases where assumptions were not met, multiple comparisons were made with Kruskal–Wallis test, and pairwise comparisons were made with Dunn’s multiple comparisons test. The significance level for each analysis was set at 0.05.
3 Results
3.1 Acute toxicity
In the acute toxicity test, it was calculated that flupyradifurone had a 140.47 mg/L of 96-h LC50 for common carp (Figure 1). No mortality was observed at the sub-lethal concentrations of flupyradifurone and negative control (0.01% DMSO) during the 14 days.
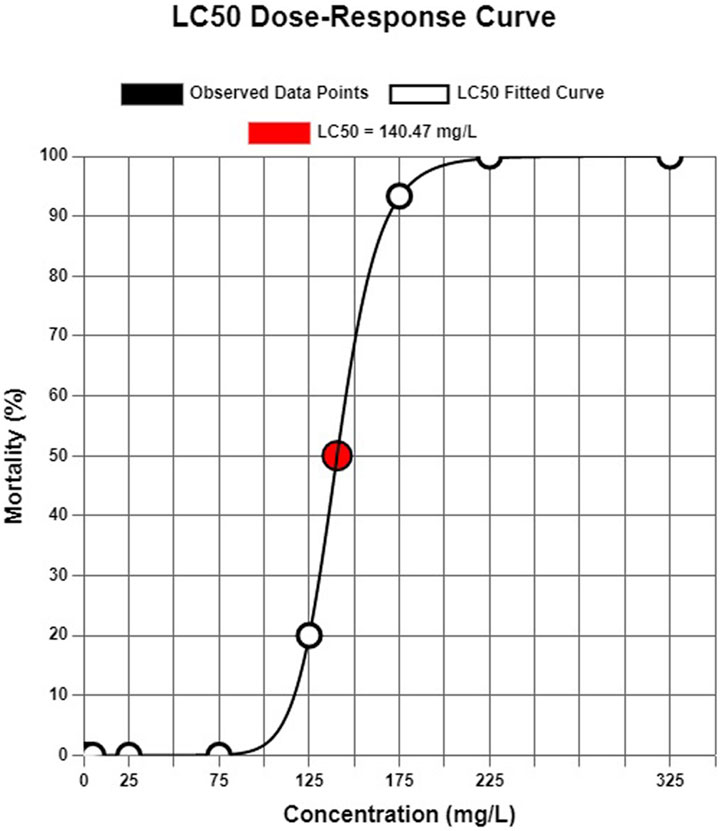
Figure 1. Probit model-based estimation of LC50 value for flypiradifurone on common carp (Cyprinus carpio).
3.2 Hematological and serum biochemical parameters
The changes in hematological parameters are summarized in Table 1. Increasing concentrations of the tested compound resulted in a significant reduction in RBC, Hb levels and Hct values across the experimental groups. In addition, dose-dependent alterations were observed in MCV, MCH, and MCHC. MCV, MCH and MCHC levels decreased significantly at higher doses. These findings indicate that the hematological parameters were adversely affected by escalating doses compared to the control group. Specifically, RBC, Hb, and Hct values declined significantly in a dose-dependent manner, indicating anemia and impaired oxygen transport, while erythrocyte indices (MCV, MCH, MCHC) were also markedly reduced at 75–125 mg/L.
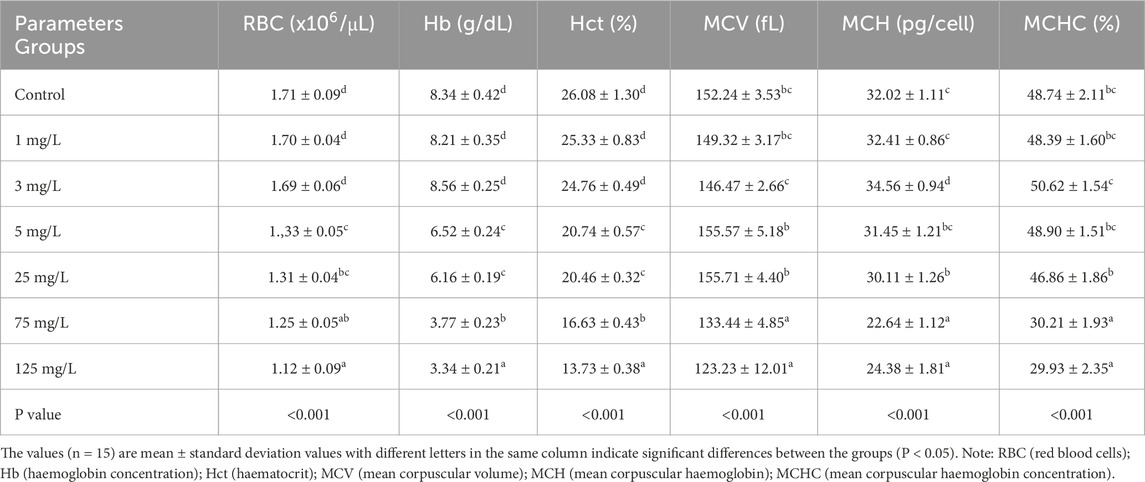
Table 1. Effects of different concentrations of flupyradifurone on the hematological parameters of common carp (Cyprinus carpio) after 14 days of exposure.
In this study, significant and dose-dependent effects of flupyradifurone exposure on the biochemical parameters of fish were observed. GLU levels showed a significant increase at concentrations of 25 mg/L and above compared to the control group. In contrast, TRIG and CHOL levels decreased as concentration increased, reaching their lowest values, particularly in groups exposed to 75 mg/L and higher. Liver function markers, including ALP, SGOT, and SGPT activities, exhibited notable increases (Table 2). Compared to the control group, SGOT activity increased approximately fourfold (P < 0.05) and SGPT activity nearly twofold (P < 0.05) in the 125 mg/L group. Additionally, TPROT (total protein) and ALB (albumin) levels showed a dose-dependent decrease with increasing flupyradifurone exposure (P < 0.05).
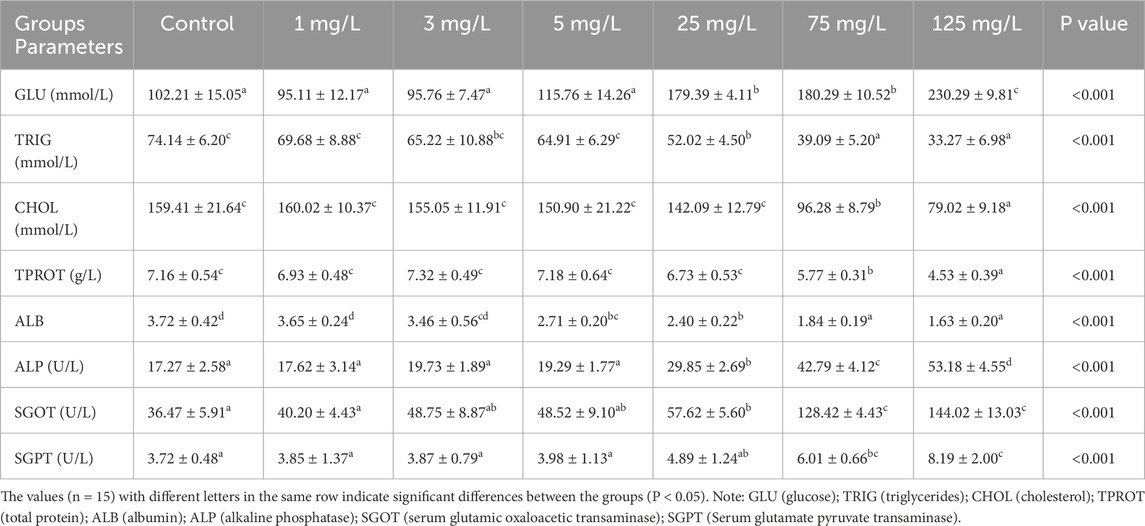
Table 2. Effects of different concentrations of flupyradifurone on the serum biochemical parameters of common carp (Cyprinus carpio) after 14 days of exposure.
3.3 DNA damage assessment by comet assay
The exposition to different concentrations of the pesticide flupyradifurone induces DNA damage in common carp erythrocytes. Microscopic images obtained using the comet assay technique clearly show the concentration-dependent genotoxic effects of flupyradifurone exposure (Figure 2). While the cells in the control group remain compact and intact, increasing pesticide concentrations lead to a noticeable increase in DNA migration, with “comet-like” formations becoming particularly prominent at high doses (≥25 mg/L).
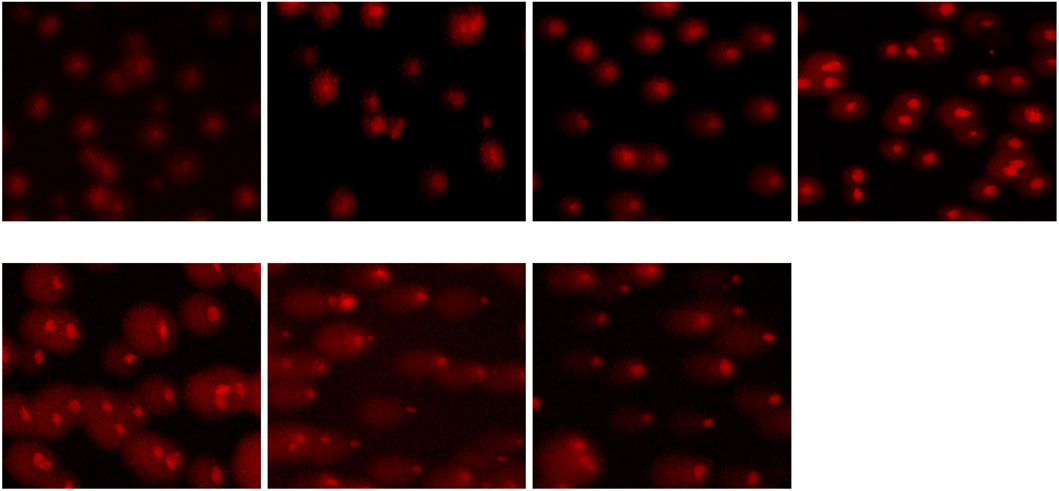
Figure 2. Comet Analysis Images of Increased DNA Damage in Common Carp Blood Cells Exposed to Different Concentrations of Flupyradifurone (Control, 1 mg/L, 3 mg/L, 5 mg/L, 25 mg/L, 75 mg/L, 125 mg/L Flupyradifurone).
The degree of DNA damage increased significantly with rising flupyradifurone concentrations. Tail length was nearly zero in the control group but showed a significant increase at 25 mg/L, 75 mg/L, and 125 mg/L, with the highest damage observed at approximately 55 μm in the 25 mg/L group (Figure 3). Head size decreased in all exposed groups compared to the control, dropping from approximately 20 μm to around 10-12 μm (Figure 3). Tail moment length followed a similar pattern, showing minimal values in the control group but significant increases at higher concentrations (25, 75, and 125 mg/L) (Figure 3). Notably, the percentage of DNA in the tail exhibited the most dramatic increase, rising from around 5% in the control to 40%-50% at 1 mg/L and reaching 75%-80% at the highest concentrations (Figure 3).
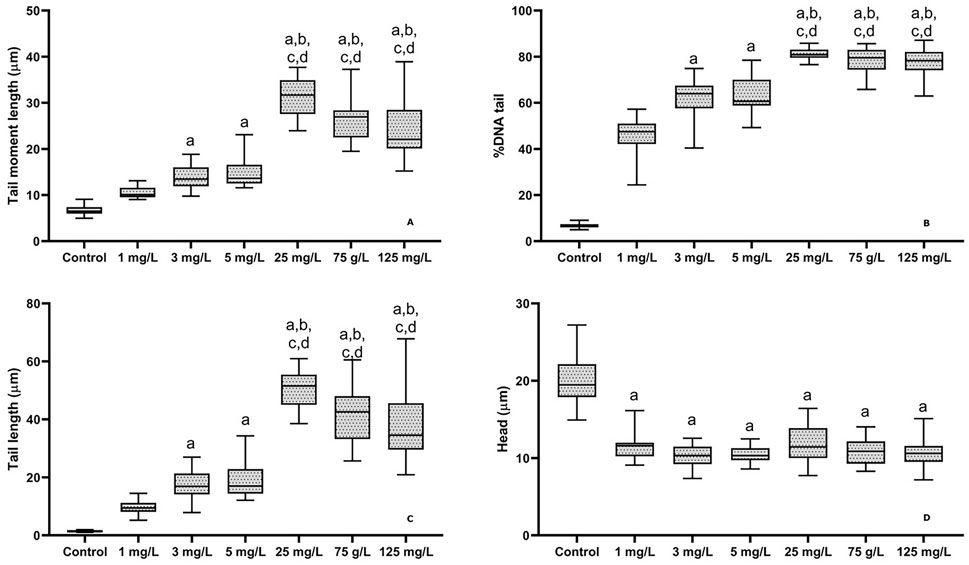
Figure 3. Genotoxic effect of flupyradifurone exposure in different doses common carp blood DNA after 14 days. Tail moment length, % tail DNA, Tail length and head. Results were expressed as median (min-max). Letters on the graph indicate significance (p < 0.05). compared to control, compared to 1 mg/L group, compared to 3 mg/L group, compared to 5 mg/L.
4 Discussion
The widespread use of agrochemicals, including novel insecticides such as flupyradifurone, has raised significant concerns regarding their environmental and ecological impacts, particularly in aquatic ecosystems. Flupyradifurone, a butenolide insecticide that targets nicotinic acetylcholine receptors (nAChRs), has gained substantial market interest due to its novel bioactive pharmacophore. However, its potential toxicity in aquatic organisms, including economically and ecologically important fish species like common carp, remains underexplored. In this study, we investigated the toxicological effects of flupyradifurone on common carp to better understand its implications for fish health including hematological, serum biochemical parameters and genotoxicity of blood. Acute toxicity assessment provides a critical step in understanding the environmental risks of emerging pollutants by providing rapid and reliable prediction of their toxic effects. It also allows the determination of safe concentration ranges and thresholds for the study of sublethal effects, providing a basis for long-term ecotoxicological studies (Majumder and Kaviraj, 2019).
According to our findings, the 96-hour LC50 of flupyradifurone in common carp was determined to be 140.47 mg/L, which aligns with previously reported values for this species. U.S. Environmental Protection Agency (2014) documented LC50 values for common carp as >100 mg/L, consistent with our results and highlighting the moderate acute toxicity of flupyradifurone. Similarly, for other fish species, EFSA reported LC50 values of >74.2 mg/L for rainbow trout (Oncorhynchus mykiss) and >70.5 mg/L for fathead minnow (Pimephales promelas). Although Zhong et al. (2021) found a much lower 96-h LC50 value of 0.21 mg/mL for zebra fish embryos (D. rerio) also in the same study revealing that flupyradifurone is toxic to zebra fish embryos (Danio rerio), resulting in diminished heart rate, survival, and body length, alongside cardiac abnormalities and heightened oxidative stress. Notably, these adverse effects were observed even at relatively low concentrations. This discrepancy underscores the species-specific sensitivity to flupyradifurone and reinforces the importance of targeted toxicological evaluations for ecologically and economically significant fish species like common carp. Although the concentrations applied in this study (1–125 mg/L) are much higher than the currently reported environmental concentrations (up to 0.16 μg/L; Metcalfe et al., 2019), the observed hematological, biochemical, and genotoxic alterations provide valuable insight into potential risks. Importantly, the marked effects at lower sublethal concentrations (1–5 mg/L) suggest that even small increases above current environmental levels could pose ecological hazards, particularly under scenarios of pesticide accumulation and chronic exposure (Metcalfe et al., 2019). Given the physicochemical properties of FPF—namely, high water solubility, low volatility, and persistence in aquatic systems (Nauen et al., 2015) also in aqueous systems under both natural sunlight and artificial light, with hydrolysis half-lives exceeding 150 days at 15 °C, 25 °C, and 35 °C, and photolysis half-lives over 168 h in natural waters (Fang et al., 2022). There is a realistic potential for accumulation in freshwater environments via runoff, erosion, and leaching. Our findings, especially the observed hematological and genotoxic alterations in C. carpio, underscore the ecological risks associated with potential long-term and low-dose exposure scenarios.
Hemato-biochemical parameters are essential indicators for assessing fish health and understanding the impact of toxic substances and pollutants and general stress responses observed in fish exposed to various toxicants, where such exposures can lead to physiological adjustments and stress (Kumar et al., 2018; Öz et al., 2020; Çelik et al., 2024). Blood plays a critical role in transporting essential nutrients, gases, hormones, and waste products, making it a reliable source of information on the physiological and biochemical status of fish (Seibel et al., 2021; Ramya et al., 2023). The hematological parameters such RBC, Hct and Hb of common carp were decreased in the present study by flupyradifurone exposure at level of 5, 25, 75 and 125 mg/L meanwhile 1 and 3 mg/L did not show any differences compared to the control group. Labeo rohita exposed to sub-lethal concentrations of carbofuran (0.16–0.80 mg/mL) and cypermethrin (0.16–0.80 mg/mL) exhibited a significant, time- and dose-dependent decrease in red blood cell count and hemoglobin levels over a 28-day exposure period (Adhikari et al., 2004). Similar hematological effects have been observed in grass carp (Ctenopharyngodon idella) exposed to profenofos-based insecticides (El-bouhy et al., 2023) and in common carp, L. rohita and Nile tilapia (Oreochromis niloticus) exposed to esfenvalerate, azadirachtin, triflumezopyrim, and hexaflumuron, respectively (Murussi et al., 2016; Navruz et al., 2023; Ibrahim et al., 2024; Nayak et al., 2024). These results emphasize the consistent adverse effects of various insecticides on the hematological health of fish across different species. In this context, the observed decrease in RBC, Hb, and Hct levels in fish exposed to insecticides may be attributed to the destructive effects of these chemicals on cell membranes. According to Mostakim et al. (2015) exposure to toxicants can suppress RBC and Hb synthesis, resulting in reduced RBC counts and Hb levels. Additionally, another potential cause of anemia is the generation of reactive oxygen species (ROS) induced by insecticide exposure, which can damage RBCs and oxidize Hb molecules, thereby impairing their oxygen carrying capacity (Lutnicka et al., 2016). Furthermore, the reductions in RBC, Hb, and Hct levels, which are strongly associated with hematological parameters, are presumed to result from the inhibition of erythropoiesis, destruction of red blood cells, and damage to hematopoietic tissues in the kidney and spleen, and disruption of the hematopoietic process (Korkmaz et al., 2023).
Serum biochemical parameters are essential markers for assessing the toxic effects in fish, with changes in these values often reflecting metabolic imbalances or organ damage (Ma et al., 2018). In vertebrates, glucose (GLU) is the primary energy source, and any surplus is stored as glycogen in the liver and muscles. However, various stressors can alter serum GLU levels, potentially indicating metabolic disturbances or damage to vital organs (Yki-Järvinen, 1992). In our study, exposure to flupyradifurone led to a significant increase in serum GLU levels in common carp. Similar elevations in glucose have been observed in common carp exposed to imidacloprid and chlorpyrifos (Banaee et al., 2024). It is likely that under flupyradifurone induced stress, glycogen stored in the liver and muscles is mobilized via glycogenesis, thereby meeting the heightened energy demands and resulting in increased serum glucose levels. Studies on other contaminants like endosulfan have also reported significant changes in biochemical parameters, including stress biomarker enzymes and blood glucose levels, indicating a general response to toxic exposure (Kumar et al., 2016). Under stress conditions, TRIG plays a role in storing cellular energy and compensating for increased energy demands (Zhu et al., 2024). In the present study, serum TRIG, CHOL, and TPROT levels significantly decreased with increasing doses of flupyradifurone. This reduction in TRIG may result from fish exposed to flupyradifurone attempting to meet their heightened energy needs by performing gluconeogenesis from non-glycogen sources (Zhu et al., 2024). Cholesterol is primarily synthesized in the liver and serves as a crucial structural component of bile acids, plasma lipoproteins, and cell membranes, as well as acting as a precursor for the synthesis of all steroid hormones (Whalan, 2015). Therefore, the decreased CHOL levels may indicate impaired hepatic cholesterol synthesis (Yki-Järvinen, 1992). Meanwhile, reduced TPROT may be associated with compromised immune function and decreased resistance to diseases (Banaee et al., 2024). Similar findings were reported by Zhu et al. (2015) in common carp following fenpropathrin exposure. Furthermore, significant reductions in total serum protein were observed in L. rohita juveniles (Whalan, 2015) and C. catla juveniles (Simons and Ikonen, 2000) exposed to sub-lethal cypermethrin levels. Likewise, decreases in triglycerides and cholesterol were observed in O. mykiss (Orun et al., 2014) and Schizothorax esocinus (Akhtar et al., 2021) following cypermethrin exposure.
This study demonstrated that flupyradifurone exerts dose-dependent genotoxic effects on common carp blood DNA, indicating that this pesticide poses a potential hazard to aquatic organisms. While such genotoxic effects may be partially repaired by cellular mechanisms, repeated or chronic exposure could surpass repair capacity, leading to cumulative genetic alterations (Livingstone, 2001). Although individual fish may recover to some extent, persistent DNA damage could impair growth, reproductive success, and survival, potentially affecting population dynamics over time (Galloway and Goven, 2006). These findings underscore the importance of assessing long-term and multigenerational effects of Flupyradifurone to better understand its ecological risks. One primary mechanism of pesticide-induced DNA damage is oxidative stress, whereby exposure elevates reactive oxygen species (ROS) production in fish tissues, which in turn attack DNA molecules and cause strand breaks and base modifications (Das and Mukherjee, 2003; Tunçsoy, 2021). For example, cypermethrin exposure in L. rohita produced marked increases in ROS levels and lipid peroxidation in gill, muscle, brain, and liver tissues effects that were effectively mitigated by vitamin C supplementation (Das and Mukherjee, 2003). Another key mechanism involves the formation of DNA adducts certain pesticides or their metabolites can covalently bind to nucleophilic sites on DNA, disrupting its structure and impairing replication and transcription (Vani et al., 2012). Khisroon et al. (2021) showed that endosulfan exposure in C. idella not only elevated ROS production but also induced DNA adduct formation in a time and dose-dependent manner. Interestingly, the comet assay revealed a sharp increase in DNA damage even at the lowest tested concentration (1 mg/L), with tail DNA reaching 40%–50%. Such an early and pronounced response may be attributed to the high sensitivity of erythrocyte DNA to oxidative stress induced by flupyradifurone. Similar non-linear or threshold-type responses have been reported in fish exposed to other pesticides, where low doses were sufficient to trigger substantial genotoxic effects before dose-dependent increases were observed (Sharma and Jindal, 2022). Another possible explanation is inter-individual variability in DNA repair capacity, which can lead to disproportionate damage at lower concentrations. Although methodological factors inherent to the comet assay may also contribute to variability, the consistent trend of increasing damage with higher concentrations supports the biological relevance of this observation. Chromosomal aberrations have likewise been observed: acute exposure to the glyphosate-based herbicide Templo® induced erythrocyte morphological alterations and micronucleus formation in yellowtail tetra (Ullah et al., 2024), highlighting the clastogenic and cytotoxic potential of such formulations. Moreover, organophosphate insecticides can disrupt enzymatic processes that contribute to DNA damage; Alavinia et al. (2019) found that malathion exposure in O. mykiss caused DNA strand breaks concomitant with alterations in acetylcholinesterase activity, indicating genotoxic effects beyond neurotoxicity. The random amplified polymorphic DNA (RAPD) assay has also proven sensitive in detecting pesticide-induced genetic alterations: Pandey et al. (2018) documented RAPD profile changes in Channa punctatus following profenofos exposure, reflecting underlying DNA damage. Finally, the bioaccumulation of persistent pesticides in aquatic ecosystems can amplify genotoxic risk through trophic transfer and tissue accumulation (Ergenler and Turan, 2023).
5 Conclusion
In conclusion, the present study demonstrates that flupyradifurone exerts significant dose-dependent toxic effects on common carp, as evidenced by its moderate acute toxicity (96 h-LC50 = 140.47 mg/L) and clear sub-lethal impacts over 14 days of exposure. Hematological analyses revealed marked declines in RBC count, hemoglobin concentration, and hematocrit, alongside alterations in erythrocyte indices (MCV, MCH, MCHC), indicating anemia and impaired oxygen transport. Biochemical profiling showed elevated glucose and hepatic enzyme activities (ALP, SGOT, SGPT), together with reductions in triglycerides, cholesterol, total protein, and albumin, reflecting metabolic stress and potential liver dysfunction. The comet assay further confirmed pronounced genotoxicity, with significant increases in tail length, tail moment, and percentage tail DNA at concentrations ≥25 mg/L. Collectively, these findings underscore the ecological risk posed by flupyradifurone to freshwater fish and highlight the importance of incorporating both lethal and sub-lethal endpoints—including genotoxic and biochemical biomarkers—into pesticide risk assessments. Future work should extend to chronic exposures and trophic-transfer studies to fully elucidate long-term environmental implications.
Data availability statement
The original contributions presented in the study are included in the article/Supplementary Material, further inquiries can be directed to the corresponding author.
Ethics statement
The animal study was approved by MSKU-HADYEK Local Ethics Commission of Animal Research. The study was conducted in accordance with the local legislation and institutional requirements.
Author contributions
ÖY: Data curation, Validation, Conceptualization, Methodology, Writing – review and editing. ÜA: Data curation, Methodology, Validation, Writing – review and editing, Formal Analysis, Investigation. RT: Data curation, Formal Analysis, Investigation, Methodology, Resources, Validation, Visualization, Writing – original draft. YE: Validation, Writing – original draft. GB: Investigation, Resources, Visualization, Writing – original draft. SY: Data curation, Resources, Validation, Visualization, Writing – original draft.
Funding
The author(s) declare that no financial support was received for the research and/or publication of this article.
Acknowledgments
We would like to thank Serkan Erkan, director of the Mediterranean Fisheries Production Research and Training Institute (MEDFRI) for providing fish specimens.
Conflict of interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Generative AI statement
The author(s) declare that no Generative AI was used in the creation of this manuscript.
Any alternative text (alt text) provided alongside figures in this article has been generated by Frontiers with the support of artificial intelligence and reasonable efforts have been made to ensure accuracy, including review by the authors wherever possible. If you identify any issues, please contact us.
Publisher’s note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
Supplementary material
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fphys.2025.1676992/full#supplementary-material
References
AAT Bioquest (2025). Quest Graph™ IC50 calculator. Available online at: https://www.aatbio.com/tools/ec50-calculator (Accessed February 15, 2025).
Adhikari S., Sarkar B., Chatterjee A., Mahapatra C. T., Ayyappan S. (2004). Effects of cypermethrin and carbofuran on certain hematological parameters and prediction of their recovery in a freshwater teleost, Labeo rohita (Hamilton). Ecotoxicol. Environ. Saf. 58, 220–226. doi:10.1016/j.ecoenv.2003.12.003
Akhtar N., Khan M. F., Tabassum S., Ahmad M. S., Badshah K. D. (2021). Effects of cypermethrin on the hematological parameters, biochemical components of blood and histopathological changes in different organs of chirruh snow trout (Schizothorax esocinus). Pak. J. Zool. 53 (3), 943–953. doi:10.17582/JOURNAL.PJZ/20190410170431
Alavinia S. J., Mirvaghefi A., Farahmand H., Rafiee G., Alavinia S. J., Shiry N., et al. (2019). DNA damage, acetylcholinesterase activity, and hematological responses in rainbow trout exposed to the organophosphate malathion. Ecotoxicol. Environ. Saf. 182, 109311. doi:10.1016/j.ecoenv/2019.05.081
Banaee M., Sureda A., Mirvaghefi A. R., Ahmadi K. (2013). Biochemical and histological changes in the liver tissue of rainbow trout (Oncorhynchus mykiss) exposed to sub-lethal concentrations of diazinon. Fish physiology Biochem. 39 (3), 489–501. doi:10.1007/s10695-012-9714-1
Banaee M., Zeidi A., Haghi B. N., Beitsayah A. (2024). The toxicity effects of imidacloprid and chlorpyrifos on oxidative stress and blood biochemistry in Cyprinus carpio. Comp. Biochem. Physiol. C Toxicol. Pharmacol. 284, 109979. doi:10.1016/J.CBPC.2024.109979
Bass C., Denholm I., Williamson M. S., Nauen R. (2015). The global status of insect resistance to neonicotinoid insecticides. Pestic. Biochem. Physiol. 121, 78–87. doi:10.1016/j.pestbp.2015.04.004
Bayer (2013). Flupyradifurone technical information. Available online at: https://fluoridealert.org/wp-content/uploads/flupyradifurone.bayer-tech-info.sivanto.2013.pdf (Accessed February 11, 2025).
Burlinson B. (2012). “Genetic toxicology,” in The methods in molecular biology. Editors J. Parry, and E. Parry (New York, NY: Springer), 143–163.
Campbell J. W., Cabrera A. R., Stanley-Stahr C., Ellis J. D. (2016). An evaluation of the honey bee (Hymenoptera: Apidae) safety profile of a new systemic insecticide, flupyradifurone, under field conditions in Florida. J. Econ. Entomol. 109 (5), 1967–1972. doi:10.1093/jee/tow186
Çelik M., Dikel S., Öz M. (2024). Investigation of the effect of water and feed sourced boron on the growth performance and blood parameters of Nile tilapia, Oreochromis niloticus. J. World Aquacult Soc. 55 (6), e13104. doi:10.1111/jwas.13104
Cheng B., Jiang F., Su M., Zhou L., Zhang H., Cao Z., et al. (2020). Effects of lincomycin hydrochloride on the neurotoxicity of zebrafish. Ecotoxicol. Environ. Saf. 201, 110725. doi:10.1016/j.ecoenv.2020.110725
Das B. K., Mukherjee S. C. (2003). Toxicity of cypermethrin in Labeo rohita fingerlings: biochemical, enzymatic and haematological consequences. Comp. Biochem. Physiol. C Toxicol. Pharmacol. 134, 109–121. doi:10.1016/S1532-0456(02)00219-3
El-bouhy Z. M., Mohamed F. A. S., Elashhab M. W. A., El-Houseiny W. (2023). Toxicity bioassay and sub-lethal effects of profenofos-based insecticide on behavior, biochemical, hematological, and histopathological responses in grass carp (Ctenopharyngodon idella). Ecotoxicol 32, 196–210. doi:10.1007/s10646-023-02628-9
Ergenler A., Turan F. (2023). DNA damage in fish due to pesticide pollution. NESciences 8, 195–201. doi:10.28978/nesciences.1405171
Fang N., Lu Z., Hou Z., Zhang C., Zhao X. (2022). Hydrolysis and photolysis of flupyradifurone in aqueous solution and natural water: degradation kinetics and pathway. Chemosphere 298, 134294. doi:10.1016/j.chemosphere.2022.134294
Finney D. J. (1952). Probit analysis: a statistical treatment of the sigmoid response curve. Ann. Entomol. Soc. Am. 45 (4), 686. doi:10.1093/aesa/45.4.686
Galloway T. S., Goven A. J. (2006). “Invertebrate immunotoxicology,” in The immunotoxicology and immunopharmacology. Editor R. Luebke (Boca Raton: CRC Press), 389–408.
Goodhue R., Mace K., Tolhurst T., Tregeagle D., Wei H., Rudder J., et al. (2020). Economic and pest management evaluation of nitroguanidine-substituted neonicotinoid insecticides: nine major California commodities; 2019, prepared for the department of pesticide regulation by the California department of food and agriculture’s office of pesticide consultation and analysis. University of California, and the University of California Cooperative Extension.
Goulson D., Nicholls E., Botías C., Rotheray E. L. (2015). Bee declines driven by combined stress from parasites, pesticides, and lack of flowers. Science 347 (6229), 1255957. doi:10.1126/science.1255957
Ibrahim R. E., Fouda M. M. S., Abdelwarith A. A., Younis E. M., Wagih E., Elshafey B. A., et al. (2024). Hexaflumuron insecticide exposure induces behavior alterations, hemato-biochemical disorders, antioxidant-immune dysfunction, and histopathological alterations in nile Tilapia (Oreochromis niloticus). Vet. Res. Commun. 48 (5), 3105–3120. doi:10.1007/s11259-024-10467-0
Jeschke P., Nauen R., Gutbrod O., Beck M. E., Matthiesen S., Haas M., et al. (2015). Flupyradifurone (Sivanto™) and its novel butenolide pharmacophore: structural considerations. Pestic. Biochem. Physiol. 121, 31–38. doi:10.1016/J.PESTBP.2014.10.011
Ji P., Liu G., Xu J., Wang X., Li J., Zhao Z., et al. (2012). Characterization of common carp transcriptome: sequencing, de novo assembly, annotation and comparative genomics. PLoS One 7 (4), e35152. doi:10.1371/journal.pone.0035152
Khisroon M., Hassan N., Khan A., Farooqi J. (2021). Assessment of DNA damage induced by endosulfan in grass carp (Ctenopharyngodon idella valenciennes, 1844). Environ. Sci. Pollut. Res. 28, 15551–15555. doi:10.1007/s11356-021-12727-x
Khoshnood Z. A. (2024). Review on toxic effects of pesticides in zebrafish, Danio rerio and common carp, Cyprinus carpio, emphasising atrazine herbicide. Toxicol. Rep. 13, 101694. doi:10.1016/J.TOXREP.2024.101694
Korkmaz N., Uğurer O., Örün İ. (2023). Toxic effects of the synthetic pyrethroid permethrin on the hematological parameters and antioxidant enzyme systems of the freshwater fish Cyprinus carpio L. Ecotoxicol 32, 646–655. doi:10.1007/s10646-023-02675-2
Kumar N., Ambasankar K., Krishnani K. K., Gupta S. K., Bhushan S., Minhas P. S. (2016). Acute toxicity, biochemical and histopathological responses of endosulfan in Chanos chanos. Ecotoxicol. Environ. Saf. 131, 79–88. doi:10.1016/j.ecoenv.2016.05.013
Kumar N., Krishnani K. K., Singh N. P. (2018). Comparative study of selenium and selenium nanoparticles with reference to acute toxicity, biochemical attributes, and histopathological response in fish. Environ. Sci. Pollut. Res. Int. 25 (9), 8914–8927. doi:10.1007/s11356-017-1165-x
Livingstone D. R. (2001). Contaminant-stimulated reactive oxygen species production and oxidative damage in aquatic organisms. Mar. Pollut. Bull. 42 (8), 656–666. doi:10.1016/s0025-326x(01)00060-1
Lutnicka H., Bojarski B., Ludwikowska A., Wrońska D., Kamińska T., Szczygieł J., et al. (2016). Hematological alterations as a response to exposure to selected fungicides in common carp (Cyprinus carpio L.). Folia Biol. 64, 235–244. doi:10.3409/fb64_4.235
Ma J., Li Y., Wu M., Zhang C., Che Y., Li W., et al. (2018). Serum immune responses in common carp (Cyprinus carpio L.) to paraquat exposure: the traditional parameters and circulating MicroRNAs. Fish. Shellfish Immunol. 76, 133–142. doi:10.1016/J.FSI.2018.02.046
Majumder R., Kaviraj A. (2019). Acute and sublethal effects of organophosphate insecticide chlorpyrifos on freshwater fish Oreochromis niloticus. Drug Chem. Toxicol. 42, 487–495. doi:10.1080/01480545.2018.1425425
Metcalfe C. D., Helm P., Paterson G., Kaltenecker G., Murray C., Nowierski M., et al. (2019). Pesticides related to land use in watersheds of the Great Lakes basin. Sci. Total Environ. 648, 681–692. doi:10.1016/j.scitotenv.2018.08.169
Mostakim G. M., Zahangir M. M., Mishu M. M., Rahman M. K., Islam M. S. (2015). Alteration of blood parameters and histoarchitecture of liver and kidney of silver barb after chronic exposure to quinalphos. J. Toxicol. 2015, 415984. doi:10.1155/2015/415984
Murussi C. R., Menezes C. C., Nunes M. E. M., Araújo M. D. C. S., Quadros V. A., Rosemberg D. B., et al. (2016). Azadirachtin, a neem-Derived Biopesticide, impairs Behavioral and hematological parameters in carp (Cyprinus carpio). Environ. Toxicol. 31, 1381–1388. doi:10.1002/tox.22143
Nauen R., Jeschke P., Velten R., Beck M. E., Ebbinghaus-Kintscher U., Thielert W., et al. (2015). Flupyradifurone: a Brief profile of a new butenolide insecticide. Pest Manag. Sci. 71, 850–862. doi:10.1002/ps.3932
Navruz F. Z., Acar Ü., Yılmaz S., Kesbiç O. S. (2023). Dose-dependent stress response of esfenvalerate insecticide on common carp (Cyprinus carpio): evaluating blood parameters and gene expression. Comp. Biochem. Physiol. C Toxicol. Pharmacol. 272, 109711. doi:10.1016/J.CBPC.2023.109711
Nayak S., Das S., Das I. I., Kumar R., Mohanty A. K., Sahoo L., et al. (2024). Impact of triflumezopyrim (insecticide) on blood and serum biochemistry of freshwater fish, subadult Labeo rohita (Hamilton, 1822). Chemosphere 367, 143554. doi:10.1016/J.CHEMOSPHERE.2024.143554
Oruc E. O., Üner N. (1999). Effects of 2, 4-Diamin on some parameters of protein and carbohydrate metabolisms in the serum, muscle and liver of Cyprinus carpio. Environ. Pollut. 105 (2), 267–272. doi:10.1016/S0269-7491(98)00206-1
Orun I., Selamoglu Z., Gulhan M. F., Erdogan K. (2014). Role of propolis on biochemical and hematological parameters of Oncorhynchus mykiss exposed to cypermethrin. J. Surv. Fish. Sci. 1, 21–35. doi:10.18331/sfs2014.1.1.3
Öz M., Karaşahin T., Aksoy N. H., İnanan B. E. (2020). Harmful effects of dietary supplementation of boron on blood parameters of rainbow trout (Oncorhynchus mykiss). J. Hell. Vet. Med. Soc. 71 (2), 2227–2234.
Öz M., Inanan B. E., Üstüner E., Karagoz B., Dikel S. (2024). Effects of dietary garlic (Allium sativum) oil on growth performance, haemato-biochemical and histopathology of cypermethrin-intoxicated Nile tilapia (Oreochromis niloticus). Vet. Med. Sci. 10 (3), e1449. doi:10.1002/vms3.1449
Pandey A. K., Nagpure N. S., Trivedi S. P. (2018). Genotoxicity assessment of pesticide profenofos in freshwater fish Channa punctatus (bloch) using comet assay and random amplified polymorphic DNA (RAPD). Chemosphere 211, 316–323. doi:10.1016/J.CHEMOSPHERE.2018.07.182
Ramesh M., Saravanan M. (2009). Hematological and biochemical responses in a freshwater fish Cyprinus carpio exposed to chlorpyrifos. Int. J. Integr. Biol. 7 (1), 52–57.
Ramya S., Barathinivas A., Jayakumararaj R., Pothiraj C., Ali D., Piccione G., et al. (2023). Eco-toxicological insights: effects of pesticides on ionic metabolism regulation in freshwater catfish, Mystus keletius. Aquat. Toxicol. 265, 106764. doi:10.1016/J.AQUATOX.2023.106764
Seibel H., Baßmann B., Rebl A. (2021). Blood will tell: what hematological analyses can reveal about fish welfare. Front. Vet. Sci. 8, 616955. doi:10.3389/fvets.2021.616955
Sharma G., Jindal R. (2022). In vivo genotoxic effects of commercial grade cypermethrin on fish peripheral erythrocytes. Environ. Mol. Mutagen. 63 (4), 204–214. doi:10.1002/em.22484
Simons K., Ikonen E. (2000). How cells handle cholesterol. Science 290 (5497), 1721–1726. doi:10.1126/science.290.5497.1721
Siviter H., Linguadoca A., Ippolito A., Muth F. (2023). Pesticide licensing in the EU and protecting pollinators. Curr. Biol. 33 (2), R44–R48. doi:10.1016/j.cub.2022.12.002
Tunçsoy M. (2021). Impacts of titanium dioxide nanoparticles on serum parameters and enzyme activities of Clarias gariepinus. Bull. Environ. Contam. Toxicol. 106, 629–636. doi:10.1007/s00128-020-03100-8
Ullah S., Zuberi A., Ullah I., Azzam M. M. (2024). Ameliorative role of vitamin C against cypermethrin induced oxidative stress and DNA damage in Labeo rohita (Hamilton, 1822) using single cell gel electrophoresis. Toxics 12 (9), 664. doi:10.3390/toxics12090664
U.S. Environmental Protection Agency (2014). Environmental fate and ecological risk assessment for foliar, soil drench, and seed treatment Uses of the new insecticide flupyradifurone (BYI 02960).
Vani T., Saharan N., Roy S. D., Ranjan R., Pal A. K., Siddaiah G. M., et al. (2012). Alteration in Haematological and biochemical parameters of Catla catla exposed to sub-lethal concentration of cypermethrin. Fish Physiology Biochem. 38, 1577–1584. doi:10.1007/s10695-012-9650-0
Whalan J. E. (2015). “A toxicologist’s guide to clinical pathology in animals,” in A toxicologist's guide to clinical pathology in animals (Cham, Switzerland: Springer), 1–11.
Yılmaz S., Ergün S. (2018). Trans-cinnamic acid application for rainbow trout (Oncorhynchus mykiss): I. Effects on haematological, serum biochemical, non-specific immune and head kidney gene expression responses. Fish. Shellfish Immunol. 78, 140–157. doi:10.1016/J.FSI.2018.04.034
Yilmaz S., Ergün S., Çelik E. Ş., Banni M., Ahmadifar E., Dawood M. A. (2021). The impact of acute cold water stress on blood parameters, mortality rate and stress-related genes in Oreochromis niloticus, Oreochromis mossambicus and their hybrids. J. Therm. Biol. 100, 103049. doi:10.1016/j.jtherbio.2021.103049
Zhong K., Meng Y., Wu J., Wei Y., Huang Y., Ma J., et al. (2021). Effect of flupyradifurone on zebrafish embryonic development. Environ. Pollut. 285, 117323. doi:10.1016/j.envpol.2021.117323
Zhu J., He N., Wang Q., Yuan G., Wen D., Yu G., Jia Y. (2015). The composition, spatial patterns, and influencing factors of atmospheric wet nitrogen deposition in Chinese terrestrial ecosystems. Sci. Total Environ. 511, 777–785. doi:10.1016/j.scitotenv.2014.12.038
Keywords: comet assay, common carp, fish haematology, lethal concentration, pesticides
Citation: Yıldırım Ö, Acar Ü, Tezel R, Erden Y, Bilge G and Yapıcı S (2025) Dose-dependent hemato-biochemical and genotoxic responses of common carp (Cyprinus carpio) to flupyradifurone. Front. Physiol. 16:1676992. doi: 10.3389/fphys.2025.1676992
Received: 31 July 2025; Accepted: 12 September 2025;
Published: 02 October 2025.
Edited by:
Mustafa Canli, Çukurova University, TürkiyeReviewed by:
Suat Dikel, Cukurova University Faculty of Sports Science, TürkiyePınar Arslan Yüce, Cankiri Karatekin University, Türkiye
Copyright © 2025 Yıldırım, Acar, Tezel, Erden, Bilge and Yapıcı. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Gökçen Bilge, Z2JpbGdlQG11LmVkdS50cg==
†These authors have contributed equally to this work and share first authorship