 Srinivas Ayyadevara1,2
Srinivas Ayyadevara1,2 Çagdas Tazearslan3†
Çagdas Tazearslan3† Ramani Alla1,2James C. Jiang4
Ramani Alla1,2James C. Jiang4 S. Michal Jazwinski4
S. Michal Jazwinski4 Robert J. Shmookler Reis1,2,3*
Robert J. Shmookler Reis1,2,3*- 1Central Arkansas Veterans Healthcare System, VA Medical Center, Little Rock, AR, USA
- 2Department of Geriatrics, University of Arkansas for Medical Sciences, Little Rock, AR, USA
- 3Department of Biochemistry and Molecular Biology, University of Arkansas for Medical Sciences, Little Rock, AR, USA
- 4Tulane Center for Aging and Department of Medicine, Tulane University Health Sciences Center, New Orleans, LA, USA
A quantitative trait locus (QTL) in the nematode C. elegans, “lsq4,” was recently implicated by mapping longevity genes. QTLs for lifespan and three stress-resistance traits coincided within a span of <300 kbp, later narrowed to <200 kbp. A single gene in this interval is now shown to modulate all lsq4-associated traits. Full-genome analysis of transcript levels indicates that lsq4 contains a dimorphic gene governing the expression of many sperm-specific genes, suggesting an effect on spermatogenesis. Quantitative analysis of allele-specific transcripts encoded within the lsq4 interval revealed significant, 2- to 15-fold expression differences for 10 of 33 genes. Fourteen “dual-candidate” genes, implicated by both position and expression, were tested for RNA-interference effects on QTL-linked traits. In a strain carrying the shorter-lived allele, knockdown of rec-8 (encoding a meiotic cohesin) reduced its transcripts 4-fold, to a level similar to the longer-lived strain, while extending lifespan 25–26%, whether begun before fertilization or at maturity. The short-lived lsq4 allele also conferred sensitivity to oxidative and thermal stresses, and lower male frequency (reflecting X-chromosome non-disjunction), traits reversed uniquely by rec-8 knockdown. A strain bearing the longer-lived lsq4 allele, differing from the short-lived strain at <0.3% of its genome, derived no lifespan or stress-survival benefit from rec-8 knockdown. We consider two possible explanations: high rec-8 expression may include increased “leaky” expression in mitotic cells, leading to deleterious destabilization of somatic genomes; or REC-8 may act entirely in germ-line meiotic cells to reduce aberrations such as non-disjunction, thereby blunting a stress-resistance response mediated by innate immunity. Replicative lifespan was extended 20% in haploid S. cerevisiae (BY4741) by deletion of REC8, orthologous to nematode rec-8, implying that REC8 disruption of mitotic-cell survival is widespread, exemplifying antagonistic pleiotropy (opposing effects on lifespan vs. reproduction), and/or balancing selection wherein genomic disruption increases genetic variation under harsh conditions.
Introduction
C. elegans was the first complex animal (metazoan) in which genes that can mediate evolutionary changes in lifespan, through natural selection, were enumerated and characterized by linkage mapping (Ebert et al., 1993, 1996). At least 11 quantitative trait loci (QTLs) that modulate lifespan in C. elegans were defined by studying the progeny of several independent inter-strain crosses, most of which were identified in multiple crosses (Ebert et al., 1993, 1996; Ayyadevara et al., 2001, 2003; Shmookler Reis et al., 2007). One such QTL, lsq4 on chromosome IV, appeared in three different inter-strain crosses as highly significant linkage-mapping peaks for longevity (each P < 10−9) (Ebert et al., 1993, 1996; Ayyadevara et al., 2001, 2003; Shmookler Reis et al., 2007). Recombinant-congenic lines were identified and mapped, allowing the limits of this QTL to be defined absolutely rather than stochastically (as likelihood maxima). After the first round of recombination screening, the interval for the longevity QTL spanned 0.3 Mb, coinciding with QTL-mapping intervals that govern resistance to several stresses: acute temperature elevation, paraquat and ultraviolet irradiation (Vertino et al., 2011). A second round of recombinant screening narrowed the lsq4 interval to <0.20 Mb (Vertino et al., 2011), providing a manageable set of positional candidate genes to test for functional consequences of altered expression.
Further experiments, reported here, have led to identification of the gene responsible for lsq4-linked traits. Two strains representing diverged alleles of this QTL, differing only within a region that comprises ~0.3% of the genome (0.3 Mb), were first compared by transcriptional profiling on microarrays, either considering all genes (“genomewide” analysis) or restricting our attention to just those genes that fall within the implicated QTL interval. The genome-wide survey defined the underlying gene as one involved in spermatogenesis or sperm-specific expression. Results of the interval-restricted survey guided more quantitative comparisons by real-time PCR, which implicated 10 genes as differential-expression candidates. Finally, candidate and control genes were tested by RNA interference to ask whether knockdown of their expression in the shorter-lived, stress-susceptible strain could recapitulate any of the phenotypes associated with the more-robust QTL allele. These studies led to discovery of a gene for which reduction of transcript levels below a threshold is both necessary and sufficient to account for all observed functional consequences of QTL dimorphism. We note that this endpoint has only rarely been attained through pursuit of linkage mapping for a complex trait (Shmookler Reis, 2003; Parsons et al., 2005; Lai et al., 2007; Shmookler Reis et al., 2007; Vertino et al., 2011; Stumpferl et al., 2012).
Most of the genes and pathways currently considered to regulate longevity were initially discovered through mutagenesis screens, and were replicated in other species by targeted knockout or occasionally by introduction of a dominant-negative or overexpressing transgene. Many other genes and pathways were first implicated by RNA-interference screens, but share the weakness of mutational screens: both are limited to targets that extend lifespan when disrupted, absent or under-expressed. Studies that begin with discovery by gene mapping complement such screens, because QTL detection is inherently unbiased as to whether a gene (or a given allele) favors or opposes longevity. It is limited only by the evolutionary requirement that at least two common alleles (natural variants) affecting the trait must exist in wild-derived populations, implying that neither ensures a selective advantage across a range of environments. Given this fundamental difference in our approach from the prevalent sources of discovery for longevity genes, it is not surprising that rec-8 was previously unrecognized as a contributor to lifespan pathways, despite its present implication in longevity of taxa as diverse as nematodes, fungi and whales.
Materials and Methods
Nematode Strains
C. elegans strain CL2a (DR1345) is a wild-type strain isolated in California in 1972; we obtained DR1345 from the Caenorhabditis Genetics Center (St. Paul, MN). SR708 is a recombinant-inbred strain described in Vertino et al. (2011), derived from F2 progeny of an interstrain cross between DR1345 and Bergerac-BO (RW7000), followed by recurrent outcrossing to CL2a/DR1345 with marker-based selection for retention of the BO allele at lsq4.
General Methods
All worms were grown on plates of 1.7% agar in Nematode Growth Medium, “NGM” (Sulston and Hodgkin, 1988), at 20°C unless otherwise noted. Plates were spotted with a freshly grown central lawn of E. coli (Sulston and Hodgkin, 1988), either strain OP50, a leaky auxotroph for uracil, or strain HT115 for RNAi experiments. Other procedures are described in Vertino et al. (2011).
Transcript Quantitation
Gene expression at the transcript level was assessed in two ways—using full-genome microarrays and by real-time, reverse-transcription PCR (RT-PCR). cDNAs were reverse transcribed from total RNA preparations (RNeasy Mini kit, Qiagen), labeled by 5′ attachment of fluors (GeniSphere HS900), and hybridized (ArrayBooster, The Gel Company) to 22,490-feature, 60-mer oligonucleotide arrays printed on epoxy slides (Washington University Genome Sequencing Center, funded by NHGRI and HHMI). Slides were scanned on a ScanArray 5000 (Perkin-Elmer), data were normalized by array-median values, and analyzed using Significance Analysis of Microarrays [ver. 2.23B, Stanford University (Tusher et al., 2001)]. Quantitative RT-PCR was conducted on an MJ Research Opticon2 Thermal Cycler, using SYBR Green PCR Master Mix (Applied Biosystems) and normalizing transcript levels to the mean of three genes that did not vary significantly between strains (β-actin, T08G5.3, and Y71D11.3) (Ayyadevara et al., 2009).
RNA Interference and Functional Assays
Nematodes were picked at the L4 larval stage, and transferred to plates seeded with dsRNA-expressing bacteria [E. coli strain HT115 (DE3), transformed with expression plasmid L4440] and induced with IPTG, as previously described (Ayyadevara et al., 2009; Tazearslan et al., 2009). Progeny of those worms were maintained on the same dsRNA-expressing bacterial strain; or, where indicated, worms were studied after transfer at the L4/adult molt to plates seeded with dsRNA-expressing bacteria, on which they were subsequently maintained.
Longevity survivals were performed as described (Vertino et al., 2011), continuing dsRNA exposure throughout life. Worms, initially in groups of 35, were transferred every second day onto agar plates with fresh lawns of dsRNA-expressing bacteria, and scored for spontaneous motility, provoked movement, and pharyngeal pumping; worms displaying none of these traits were considered dead. For paraquat-resistance assays (Shmookler Reis et al., 2007), groups of 30 worms were maintained with RNAi exposure for 3.5 days, and transferred (as day-3 adults) to liquid survival medium containing 150 mM paraquat for variable periods of exposure, or to 0–80 mM paraquat at 20°C for 3 days (replacing paraquat medium daily). Deaths were assessed as above, at the times indicated. For thermotolerance assays, modified from Lithgow et al. (1995), groups of 30 worms were picked 24 h after the L4/adult molt onto 6-cm agar plates and immersed to a depth of 0.5 cm in a 35.5°C water bath. Plates were removed at the indicated times and worms were scored for survival in the same sequence. Male counts were performed 1 day after the L4/adult molt, for worms whose parents and grandparents (prior to egg-laying) had been exposed to dsRNA-expressing bacteria.
Replicative Lifespan Assays in Saccharomyces Cerevisiae
Replicative lifespans were determined for budding yeast as described previously (Jiang et al., 2000). Briefly, YPK9 and BY4741 cells were grown in liquid YPG medium (2% peptone, 1% yeast extract, 3% glycerol) to eliminate petites unable to utilize the non-fermentable carbon source. They were then spotted on YPD plates (2% peptone, 1% yeast extract, 2% glucose, 2% agar). Budding cells were selected by micromanipulation and arranged in a row in an isolated region of the plate. The new buds were dissected to initiate the experiment, and the mother cells were discarded. The plates were incubated at 30°C and periodically examined under the microscope. The lifespan of a cell was recorded as the number of buds it produced (consecutively removed by micromanipulation and discarded), before it ceased dividing and lost refractility. Data are summarized by mean and standard deviation of the maximum generation (bud) number achieved in each of 40 mother cells.
Statistics
Transcript levels ascertained on microarrays were considered to display differential expression if their genome-wide false discovery rate q (the expected frequency of false positives for the entire microarray) was less than 5% (Tusher et al., 2001); in order to further restrict the gene list, a lower threshold (e.g., q < 1% or q = 0) was sometimes used. A separate cohort was expanded and its RNA extracted for each microarray, so that the biological N was equal to the number of assays.
In considering the significance of quantitative intergroup differences (e.g., for RT-PCR data), two strains were usually compared by two-tailed heteroscedastic t-tests (appropriate to samples of unknown or unequal variance), considering N to be the number of biological preparations used for each group (i.e., ignoring technical replicates). P-values less than 0.05 were considered to have attained nominal statistical significance despite multiple comparisons, so as not to inflate the Type II error rate in low-power assays (e.g., utilizing 2–4 biological samples of each strain).
In all C. elegans survival comparisons, significance was assessed by Cox-Mantel F-test, a conservative non-parametric statistic (Lee and Wang, 2003); similar P-values were obtained by the Gehans-Wilcoxon log-rank test. Comparisons of nematode proportions surviving in a dose-response experiment utilized the chi-squared test for each dose. Yeast replicative lifespan differences were evaluated by Mann–Whitney–Wilcoxon rank-sum tests.
Results
Effects of QTL Alleles on Genome-Wide Transcript Levels
We grew four independent cohorts each of strains CL2a and SR708, a near-congenic line differing from CL2a only by a Bergerac-BO-derived segment on chromosome 4, which contains the lsq4 QTL (Vertino et al., 2011) between snp_Y45F10C (13.33 Mb) and snp_W02A2 (13.65 Mb). Four microarrays (Washington Univ. Genome Sequencing Center) were hybridized with dual-label cDNA probes, reverse-transcribed from the RNA preparations obtained from these cohorts. Despite the very narrow lsq4-spanning region of Bergerac-BO DNA substituted in the CL2a genome to create SR708 (0.32 Mb, containing <50 genes), more than 150 genes were differentially expressed between these strains, at a false discovery rate (q) of ≤1%; a substantial majority were expressed at a lower level in SR708 than in CL2a. The 22 most significantly differential genes, those with q = 0, are listed in Table 1A. Of those genes, 12 are msp genes encoding Major Sperm Proteins, and another 3 encode proteins with MSP-like domains. The remaining 7 genes include a basic sperm nuclear protein unrelated to MSPs, two proteins with eggshell-protein domains, and two of the 12 NSPA-family members. The MSP family comprises >50 small basic proteins implicated in extracellular signaling and cytoskeletal functions, many of which are enriched in the amoeboid sperm cells. We used RT-PCR targeting two of these genes, msp-142 and msp-56, to confirm their differential expression as indicated on microarrays (Table 1B). To assess the generality of lsq4 allelic effects on msp expression, we also quantified transcripts for a conserved msp region (Table 1B). Overall, msp expression based on RT-PCR differed by nearly 5-fold between lsq4 alleles, a less extreme ratio than observed on microarrays.
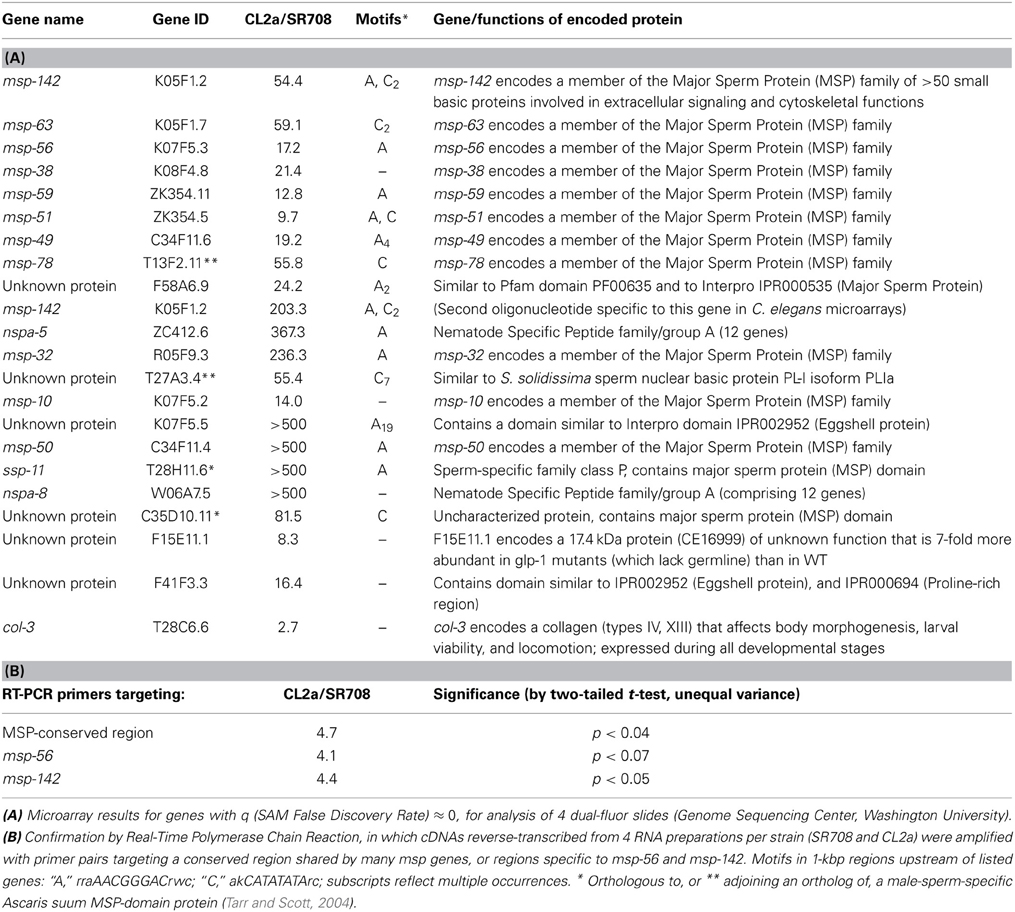
Table 1. Genes with highly differential expression dependent on the lsq4 allele.
In view of the striking enrichment observed for structurally or functionally related genes, and considering that none of the top 50 genes are located within the lsq4 interval, we inspected their upstream 1-kb regions with modules of RSAT (regulatory sequence analysis tools, http://rsat.ulb.ac.be/rsat/) to seek shared motifs. “Oligo-analysis” identified 3 over-represented motifs: AACGGGAC (enriched 32-fold, P < 2E−12), CATATATA (10-fold, P < 5E−7) and AAAGTTTC (4-fold, P < 2E−5). “Matrix scan” found extended versions of the first two motifs, rraAACGGGACrwc (P < 3E−33) and akCATATATArc (P < 2E-6). Although such enrichments were highly significant, lsq4-allele-specific binding of one or more transcription factors to these motifs could not account for the differential transcript abundances observed (see “Motifs” column, Table 1).
Differential-expression patterns can provide valuable clues as to the nature of the underlying gene that is dimorphic between QTL alleles. For example, altered expression of sperm-specific genes might reflect a change in the frequency of males, since male C. elegans not only produce more sperm than do hermaphrodites (>1000 vs. ~300 per worm), but also produce sperm that differ in many of their constituent proteins from hermaphroditic sperm (Tarr and Scott, 2004). On assessing the frequency of males, we noted a distinct “HIM” (high incidence of males) trait in strain CL2a/DR1345 [1.2 ± 0.2 (SEM) %, 1.4 ± 0.4% in two experiments, total N = 4100], well above the 0.1–0.2% seen in most other wild-type strains such as Bristol-N2, Bergerac-BO [RW7000], or a different CL2a subline [CB4857] (Hodgkin and Doniach, 1997). This trait was reverted in SR708, for which the male incidence was 0.2 ± 0.1% (total N = 4750), 6- to 7-fold less than CL2a (chi-squared P < 10−5 in each experiment), whereas none of the longer-lived recombinant-congenic lines (described in Vertino et al., 2011) differed from CL2a. Male incidence thus constitutes another allele-specific trait of lsq4, in addition to longevity, thermotolerance, and paraquat resistance (Vertino et al., 2011).
QTL Allele Effects on Transcript Abundances for lsq4 Positional-Candidate Genes
We then restricted our expression survey to genes in the lsq4 interval (Table 2), by selecting data from the full-genome microarrays (see previous section). We identified 18 genes in the lsq4 interval, for which transcript levels differed between CL2a and the SR708 line, and retested these by quantitative real-time reverse-transcriptase PCR (qRT-PCR). Considering only the biological N of 2 (two independent grow-ups of each strain, distinct from those initially screened in microarrays), and not the technical duplication of assays for each of those, 10 of the 18 genes assessed showed allele-dependent changes that were nominally significant (P ≤ 0.05), whereas no more than one would be expected by chance (Table 3).
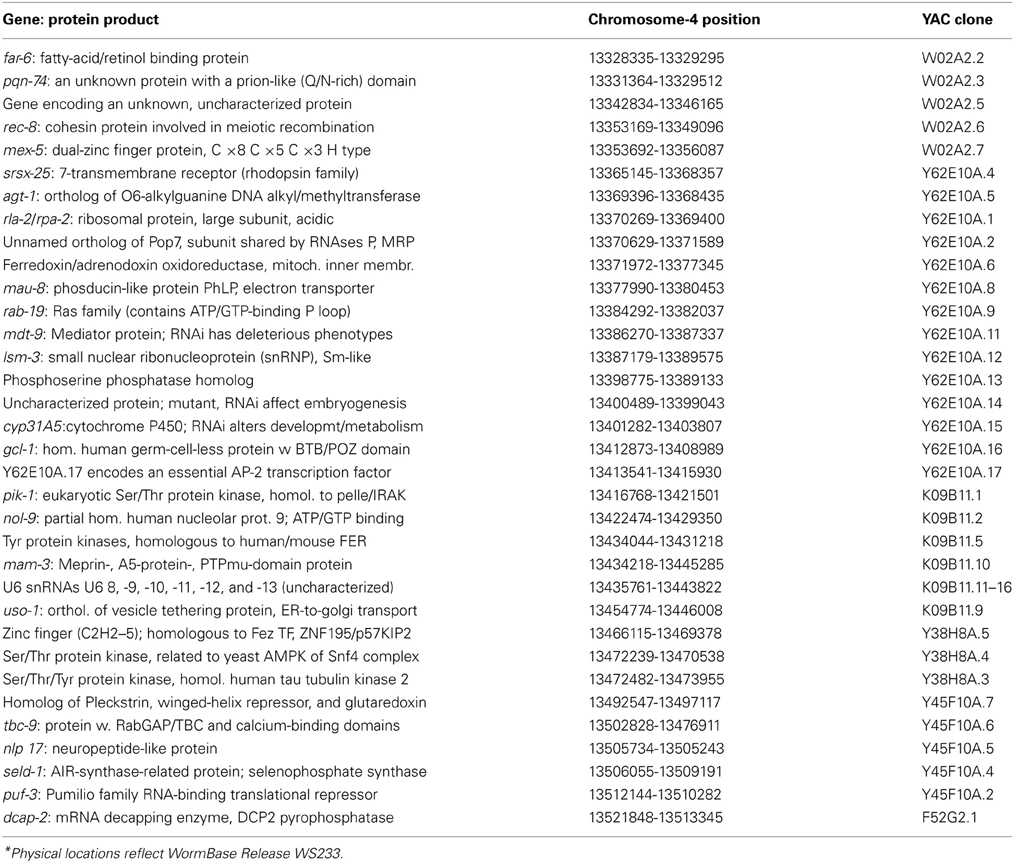
Table 2. Genes in the lsq4 interval—positional candidates for life-span determinants.
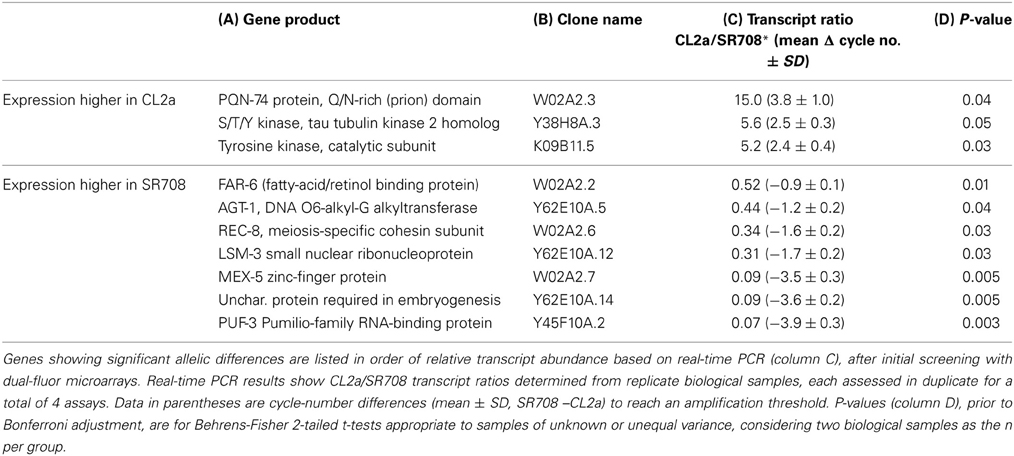
Table 3. Genes in the lsq4 interval for which transcript level depends on the lsq4 allele.
Functional Assessment of lsq4 Candidate Genes by RNA-Interference Knockdown
Although QTLs have their origins in DNA-sequence polymorphisms that could either alter protein sequence and function or have cis effects on transcription (or more rarely, both), allelic shifts in transcript level are relatively common and their functional importance can be readily assessed by RNA interference (RNAi). The 10 lsq4-interval genes confirmed to be differentially expressed dependent on the lsq4 allele present (listed in Table 3) were tested along with 4 control genes, to determine whether RNAi can shift the strain with higher transcript level toward the phenotypes of the lower-expression strain. Increased longevity in response to RNAi is a relatively exceptional outcome, reported for 1.2–1.4% of genes surveyed in high-throughput, single-point assays (Hamilton et al., 2005; Hansen et al., 2005), and thus permits stronger inference than the far more common result of reduced survival.
CL2a or SR708 worms were fed for 3 days beginning at the L4/adult molt, on bacteria expressing gene-specific double-stranded RNA (dsRNA) constructs from the Ahringer library (Kamath and Ahringer, 2003) if they were present there. We designed dsRNA-expression inserts for selected genes that were not represented in the Ahringer library, which were synthesized (IDT, San Diego CA), ligated into an expression plasmid, and transfected into bacteria, as reported previously (Ayyadevara et al., 2007, 2009). In the case of rec-8, encoding a cohesion protein (“cohesin”) required for alignment of homologous and sister chromatids during meiosis, we observed ~4-fold (73, 76% in two assays) suppression relative to worms fed the empty expression vector or an unrelated dsRNA; the range for all tested genes was 2- to 28-fold suppression (data not shown). We then assessed the treated worms for RNAi effects on established lsq4 traits: longevity, paraquat resistance, thermotolerance, and male frequency.
Longevity
In an initial screen for effects of RNA interference on lifespan, genes in the lsq4 interval were selected for RNAi suppression of expression in either parental strain CL2a or the recombinant line SR708 (differing from CL2a by insertion of the Bergerac-BO-derived lsq4 allele); positive or suggestive results were repeated multiple times (Table 4). Significant lifespan extension (P < 0.0006 to P < 4 × 10−5 prior to correction for multiple measures) was seen only for RNA interference targeting rec-8, and only in the SR708 line. Table 4 summarizes data from three independent survivals, in which rec-8 dsRNA treatment increased the longevity of SR708 worms to roughly the same level as CL2a (with or without dsRNA). Two of these survival experiments are illustrated in Figure 1. In Figure 1A, mean adult lifespan at 20°C was extended 26%, from 13.5 ± 0.7 d (mean ± s.e.m.) for SR708 adults fed bacteria carrying an empty dsRNA expression vector, to 17.0 ± 0.9 d for those exposed to rec-8 dsRNA (Cox-Mantel P < 0.0004). Note that inclusion of the period of larval development would add 2.5 days to the adult lifespan for each strain, to obtain the “total lifespan” more commonly shown. In contrast, the mean adult longevity of strain CL2a, 16.2 ± 0.9 d, was not shifted significantly (<3% change) as a result of any dsRNA treatment (Figure 1B and Table 4). In this experiment, RNA interference began at the L4/adult molt to avoid embryonic lethality as reported previously (Pasierbek et al., 2001). A very similar life extension relative to empty-vector control (25%, P < 4 × 10−5), however, was obtained in a second experiment wherein dsRNA treatment began in the parental generation (Figures 1C,D, and Table 4) to maximize the impact of knockdown. This resulted in substantial but transient embryonic lethality (Pasierbek et al., 2001), but no subsequent impairment in worms that completed embryogenesis. A third experiment also confirmed significant life extension by rec-8 (Table 4; P < 0.0006), but calculation of the mean age at death was complicated by censorship of worms lost due to burrowing in the agar. The other genes tested here (described briefly in Table 2 and Discussion) did not significantly affect longevity.
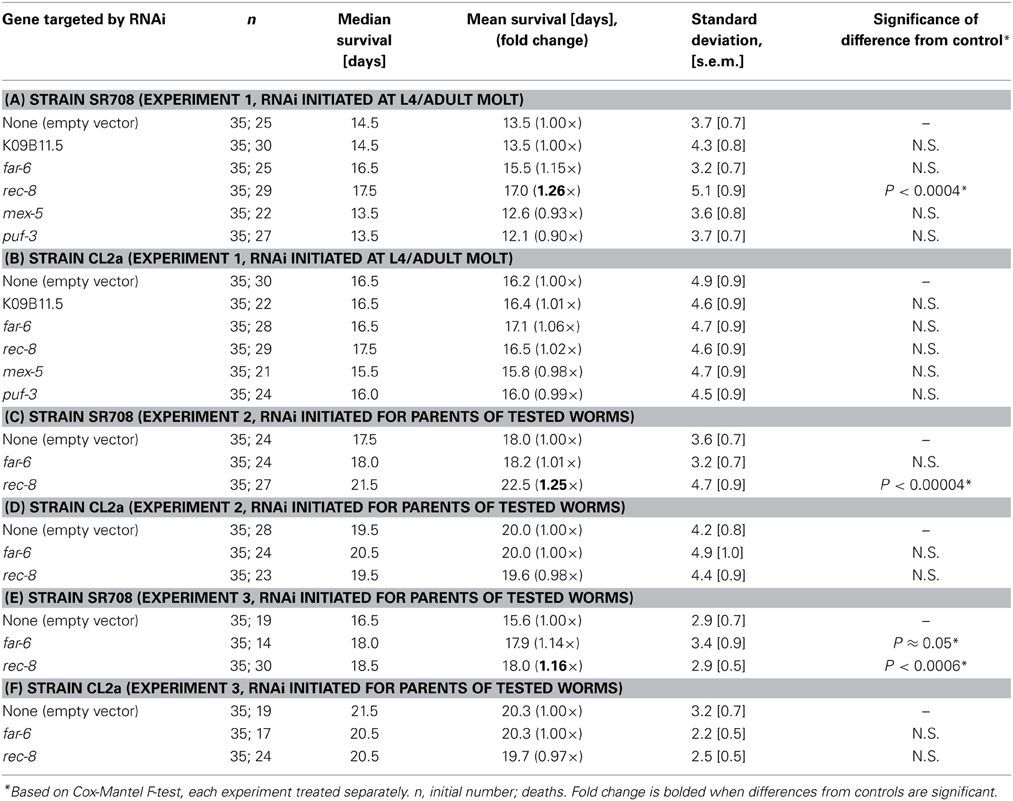
Table 4. Survival data for near-isogenic strains SR708 and CL2a, fed on E. coli expressing the indicated dsRNAs.
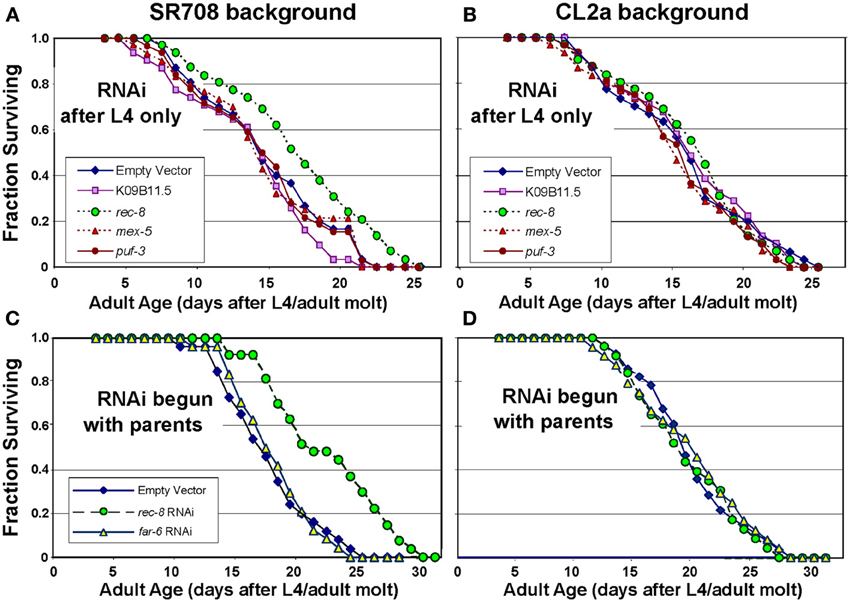
Figure 1. Rec-8 knockdown extends lifespan, dependent on genetic background in the lsq4 region. Groups of 35 worms were fed on bacteria induced to express the indicated dsRNA (or empty expression vector) continuously from the L4/adult molt (A,B) to avoid possible developmental effects, or initiated with the L4/adult molt of the parents of tested worms (C,D) to maximize knockdown. Lifespan survivals were monitored as described (see Methods). Of the indicated genes targeted by dsRNAs, only K09B11.5 had been found to be expressed at higher levels in CL2a, the longer-lived strain, and thus predicted to possibly reduce CL2a longevity; the other 4 genes were expressed at higher levels in SR708, and thus were predicted to extend survival of treated SR708, if causal to the QTL effect on longevity. (A,C) Lifespan survivals of control vs. dsRNA-treated SR708 adults; (B,D) Lifespan survivals of control and dsRNA-treated CL2a adults.
Paraquat survival
Paraquat exposure constitutes a toxic oxidative stress, through generation of superoxide radical (Ebert et al., 1993, 1996; Ayyadevara et al., 2001, 2003; Shmookler Reis et al., 2007). As illustrated in Figure 2A, siRNA targeting rec-8 was the only RNA-interference treatment that improved paraquat survival for strain SR708. This dsRNA-expressing construct extended survival by 17%, to 8.5 ± 0.3 [s.e.m.] h (Figure 2A; P < 0.01 by log-rank test), very close to that of mock-treated CL2a (8.3 ± 0.2 h; Figure 2B). In CL2a worms, however, rec-8 RNAi had no effect, as might be expected if the longer-lived lsq4 allele already expressed REC-8 at levels near optimal for adult survival. Conversely, RNAi to K09B11.5 (encoding a tyrosine protein kinase) reduced paraquat survival in the CL2a strain (Figure 2B; 25% decrease, P < 6E–5), to a greater extent than in SR708 (Figure 2A; 16%, P < 0.04). Although these data are consistent with a survival-curtailing role for K09B11.5 in view of its 5-fold higher expression in CL2a, they offer only weak support for a causal role because RNA interference often impairs robustness, presumably by disruption of diverse pathways.
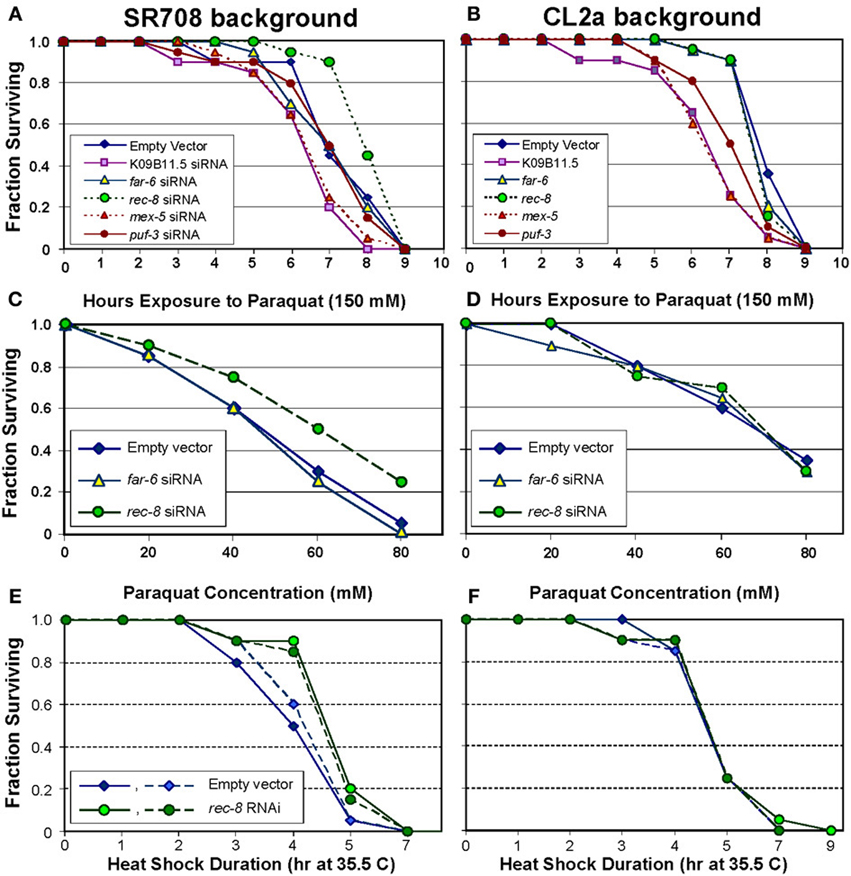
Figure 2. Rec-8 knockdown improves resistance to oxidative and thermal stresses, dependent on the lsq4–region background. Groups of 30 worms were fed on bacteria induced to express the indicated dsRNA (or empty expression vector) for 3 days from the L4/adult molt. They were then transferred to medium containing paraquat, and their subsequent survival monitored (see Methods). Of the indicated genes targeted by dsRNAs, only K09B11.5 had been found to be expressed at higher levels in CL2a, the longer-lived strain, and thus predicted to possibly reduce CL2a survival; the other 4 genes were expressed at higher levels in SR708, and thus were predicted to extend survival of treated SR708, if causal to the QTL effect on paraquat resistance. (A,B) Time courses of paraquat survival for control and dsRNA-treated adult worms, either (A) strain SR708, or (B) strain CL2a. (C,D). Paraquat dose-response curves for control vs. dsRNA-treated adults of strain (C) strain SR708, or (D) strain CL2a. (E,F) Thermotolerance survivals following abrupt transfer of standard survival plates from 20 to 35.5°C, containing control or dsRNA-treated adults of (E) strain SR708, or (F) strain CL2a. Each panel for heat-shock survival shows combined results from two independent experiments (solid and dashed lines indicate experiments 1 and 2, respectively). All panels show data representative of 2–4 experiments of each type.
The lsq4 allelic difference in paraquat resistance, and its reversal by RNAi targeting rec-8, were confirmed by dose-response experiments in which worms were monitored for survival in varying concentrations of paraquat. As illustrated in Figures 2C,D, RNAi to rec-8 increased the paraquat LD50 by >30% (from 46 to 60 mM), for SR708 adults (chi-squared P < 0.005), but had no effect on CL2a.
Thermotolerance
Although C. elegans can tolerate a wide temperature range, sudden shifts of >14°C (termed “heat shock”) are lethal over the course of 3–7 h (Lithgow et al., 1995). We had observed an allelic difference in survival following abrupt transfer from 20 to 35.5°C, mapping coincidentally with the lsq4 longevity trait (Vertino et al., 2011). SR708 worms, when grown on E. coli expressing rec-8 dsRNA, acquired thermotolerance equivalent to CL2a (differing from SR708 only by the longer-lived lsq4 allele), whereas control worms consuming only vector-bearing bacteria did not (Figure 2E). The same rec-8 knockdown produced no improvement in survival of heat-shocked CL2a worms (Figure 2F). These plots (Figures 2E,F) show results of two independent experiments, indicated by solid vs. dashed lines, which were combined to assess significance of enhanced thermotolerance for SR708 worms fed rec-8 RNAi (P < 0.002 by Cox-Mantel test).
Male incidence
The longer-lived strain, CL2a, has a distinct HIM (high-incidence of males) trait with 1.8% males, whereas SR708 produced 0.2% males—a level typical of most wild-type strains (Figure 3). Continuous exposure of SR708 or CL2a hermaphrodites to rec-8 dsRNA from the initiation of development (i.e., beginning in the prior generation to precede embryogenesis) raised the incidence of their male progeny to 12.5 and 14.3% respectively, close to the 15% level reported previously for either rec-8 loss-of-function mutation or RNAi knock-down in an N2 background (Pasierbek et al., 2001). These high proportions of males greatly exceed those observed in SR708 or CL2a, whether untreated or exposed to a different dsRNA species, e.g., far-6. In several independent groups assessed together, for example, rec-8 RNAi increased the male incidence of SR708 by 120- to 180-fold (N = 1267–1920 per group, each chi-squared P < 10−39), while in the CL2a strain male incidence rose 8- to 15-fold (N = 1147–1542 per group, each chi-squared P < 10−28). Very similar results were seen in repeat experiments.
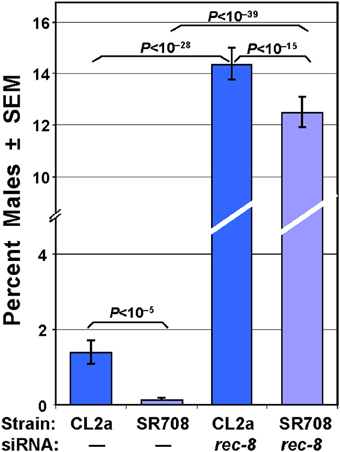
Figure 3. Male frequency in the two lsq4 isogenic strains. The longer-lived lsq4-isogenic strain, CL2a, has a significant HIM (high incidence of males) phenotype (two “–” bars on left). RNA interference targeting rec-8 greatly elevates the male frequency, but the difference between the two lsq4-allelic strains was preserved (two “rec-8” bars). N ranged from 1147–1920 progeny examined for male incidence per assessment. Data are shown for one of three experiments with similar results.
Deletion of REC8 can Extend the Replicative Lifespan of Yeast
The Rec8p cohesin-complex proteins of fission and budding yeast are functionally analogous to the nematode REC-8 protein, and are considered to be its orthologs (http://www.wormbase.org/, Pasierbek et al., 2001). We tested the effect of REC8 deletion on the number of mitotic progeny produced per mother cell, in the budding yeast Saccharomyces cerevisiae. This assay, performed as described previously (Jiang et al., 2000), was initially conducted for both the long-lived strain BY4741 (mean lifespan 29–30 bud generations) and a short-lived strain, YPK9 (20–21 bud generations). As summarized in Figure 4, replicative lifespan increased significantly in 4 of 5 of BY4741 rec8Δ clones, whereas it declined slightly but insignificantly in two YPK9 rec8Δ clones. The BY4741 rec8Δ clones increased in mean number of progeny, relative to their within-experiment controls, from 30.2 to 36.6 (21%, P < 0.03) and 34.4 (14%, not significant) in the first experiment. In a second experiment, 3 BY4741 rec8Δ clones gained 20, 20, and 25% in replicative potential (P < 0.05, 0.03, and 0.01 respectively). Combining the data for all five clones relative to their respective controls, the longevity extension upon deleting the REC8 gene was 20 ± 4% (SD) with an overall significance of P < 5 × 10−8. We also calculated the significance (Figure 4B) by a simple t-test comparison treating each mean value as a single data point, which resulted in a more conservative P-value of 0.001. Because strain YPK9 differed markedly from BY4741, we inquired whether this might relate to the relative abundance of REC8 transcripts in the two haploid strains (which undergo mitosis but not meiosis). REC8 expression was 20-fold greater in YPK9 (Figure 4C), suggesting that this strain must not be under strong selection against harmful effects of inappropriate Rec8p expression in mitotic cells.
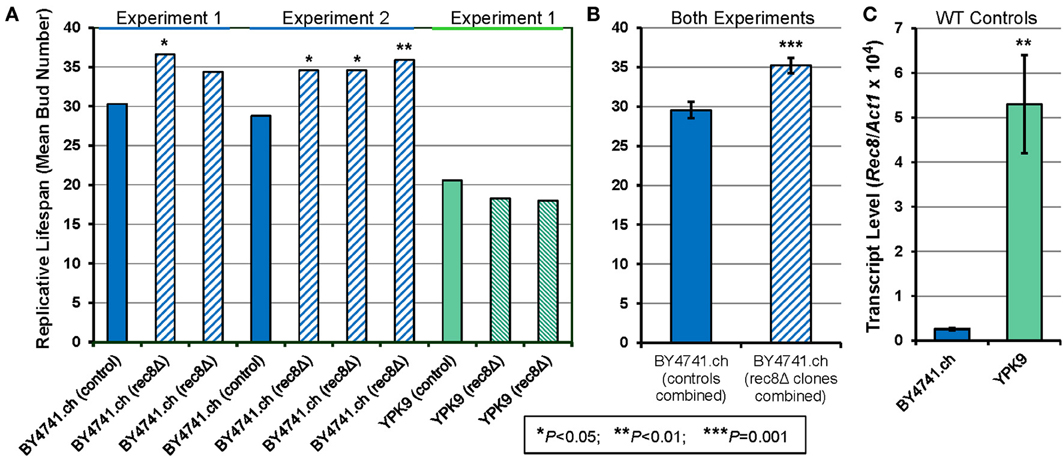
Figure 4. Deletion of REC8 increases replicative lifespan in a long-lived yeast strain that limits transcripts of this gene. (A) Replicative lifespan was measured for the BY4741.ch strain and 5 independent deletion mutations, all assessed as haploids, in two experiments. In the first experiment, strain YPK9 and two REC8 deletion mutants were also assessed. Means lifespans (in generations) are shown, with the significance of differences from the parental control strain indicated by asterisks. Four of the five BY4741 deletion clones were significantly longer lived than the within-experiment control, by P < 0.05; the combined significance of the 5 clonal life extensions was 4 × 10−8. (B) The group means and standard deviations are shown for the two genotype groups, treating each mean lifespan of a control or rec8Δ clone as a single point. Even by this conservative approach, the 5 rec8Δ clones differed from the two control assays at P = 0.001. (C) Transcript levels were assessed by quantitative real-time polymerase chain reaction (RT-PCR) for REC8 relative to ACT1 (actin), as described in Materials and Methods. Results are shown for triplicate cultures of the wild-type strains, with error bars indicating standard deviations. *P < 0.05; **P < 0.01; ***P = 0.001.
Discussion
Positional Candidate Genes for Life-Span Effects
Confirmed and putative genes in the lsq4 interval on chromosome IV are “positional candidates” that may underlie the traits associated with this QTL. These include 33 genes encoding known or likely proteins, and 6 sites templating small nuclear RNAs U6-8 through U6-13 (Table 2). The region contains a further 14 open reading frames (ORFs) that are unsupported either by homology to known genes or by RNA-derived clones, two of which are annotated as pseudogenes. Any of the candidate genes could be responsible for the allelic effects we demonstrated with respect to lifespan, resistance to heat and superoxide radical, and male incidence. Although different lsq4 traits could, in principle, arise from polymorphisms in different genes, a single gene that accounts for all of the observed QTL phenotypes would provide the most parsimonious explanation.
None of the identified candidates in the lsq4 interval were previously implicated in pathways modulating lifespan or stress resistance, although several could plausibly play such roles. They include genes encoding: (1) Four protein kinases and one phosphatase, which could be involved in signal transduction; K09B11.5 encodes a tyrosine protein kinase related to a yeast CDK and human/mouse Fer; (2) Two proteins with zinc-finger domains (e.g., MEX-5) and one with a winged-helix domain, characteristic of DNA-binding modulators of transcription; (3) The REC-8 protein, a cohesin required for meiosis and corresponding to SSC-1 in mitosis; (4) FAR-6, a fatty acid/retinol-binding protein; (5) AGT-1, an O6-alkyl-guanosine DNA-methyltransferase; and (6) Two RNA-binding proteins (PUF-3 and DCAP-2) that may affect mRNA stability.
Differential Effects of Gene Allele on Transcript Levels
Genomewide microarray analysis can provide clues as to the nature of the gene underlying a QTL. Of the 22 genes for which expression depended most significantly on the lsq4 allele (each with false discovery rate q = 0), all were expressed at higher levels in CL2a than in SR708; 12 are members of the MSP (major sperm protein) family, and a further three are unknown proteins containing MSP-like domains (Table 1A). These results were corroborated and made more quantitative by RT-PCR targeting a region shared by many msp genes, or distinguishing two particular genes, msp-142 and msp-56 (Table 1B). For each target, CL2a showed between 4- and 5-fold more transcripts than SR708. Such a concerted shift in gene expression suggests that the key dimorphic gene within lsq4 may encode a factor modulating a differentiation state, or the transcription or stability of mRNAs, which could then affect transcript abundances of many other genes without regard to their chromosomal locations. Of the genes situated in the lsq4 interval, those most likely to alter transcript levels of other genes are mex-5 (encoding a protein with two zinc-finger domains, expressed in germ-line cells of adults), Y38H8A.5 (encoding a p57KIP2 zinc-finger protein), Y45F10A.7 (encoding a protein with a winged-helix transcriptional repressor domain), lsm-3 (encoding a small nuclear ribonucleoprotein), and puf-3 (encoding an RNA-binding translational suppressor, which might affect transcript levels indirectly). Although conserved motifs (potential binding sites for proteins or RNAs, which might affect rates of transcription or mRNA degradation) were enriched with high significance in the set of genes most decisively affected by the choice of lsq4 allele, the distribution of those motifs (Table 1, column 4) was not sufficient to explain the profile of differential gene transcription.
Dimorphism in rec-8 Accounts for All Observed lsq4 Traits
Multiple lines of evidence converge on rec-8 as the sole gene responsible for multiple phenotypes that depend on the allele of lsq4. First, rec-8 is the only gene in this interval that had been previously shown to influence male incidence: both rec-8-deficient mutants and wild-type worms exposed to rec-8 dsRNA have exceptionally high male frequencies, increasing nearly 100-fold following RNAi treatment (Pasierbek et al., 2001). We observed a higher proportion of males in the CL2a strain DR1345 (1.25 ± 0.29% s.e.m.) than has been seen in other wild-type isolates such as Bristol-N2, Bergerac-BO [RW7000], or a different CL2a subline [CB4857], each of which produces 0.1–0.2% males (Hodgkin and Doniach, 1997). Male frequency in the SR708 recombinant-congenic line, in which a narrow segment of Bergerac-BO DNA is embedded in chromosome IV of CL2a, was 0.2 ± 0.1%, 6- to 7-fold less than CL2a (chi-squared P < 10−5) in each of two experiments. CL2a has 4- to 5-fold higher transcript levels of genes encoding Major Sperm Proteins (Table 1B), as well as other genes expected to be expressed chiefly in sperm (Table 1A). Thus, allelic differences in the frequency of males, due to the “HIM” phenotype associated with low REC-8 activity, would markedly affect the abundance of male sperm, and of transcripts specific to male-produced sperm (Tarr and Scott, 2004), which include most MSP-domain genes (Tarr and Scott, 2005).
Only one previous report has linked male incidence to longevity. Mutation of him-6, the C. elegans ortholog of human BLM (encoding the RecQ helicase that underlies Bloom-syndrome progeria), in the nematode causes genomic instability leading to increased generation of males while reducing lifespan (Wicky et al., 2004; Grabowski et al., 2005). Based on such observations, neither the HIM trait nor genome destabilization per se would have been expected to improve longevity. Mutations that reduce C. elegans sperm number without altering male incidence can either modestly extend lifespan, like spe-10 (Cypser and Johnson, 1999) and spe-26 (Kim and Sun, 2007), or shorten it like folt-1 (Austin et al., 2010). It thus appears unlikely that an increase in total spermatogenesis could account for the greater longevity associated with the CL2a allele of lsq4; in fact just the opposite effect, a reduction in lifespan with increased hermaphrodite sperm production, has been described (Van Voorhies, 1992).
RNA interference has the potential to test genes for their functional roles, but it carries much stronger inferential power when a gene knockdown produces a positive effect, since detrimental effects can (and do) arise as artifacts. Of 14 lsq4-interval genes tested for an effect on lifespan when targeted by RNA interference, only rec-8 produced significant and reproducible effects, extending lifespan at least 25% in two of three independent experiments (Figure 1 and Table 4). This effect size accounts for most or all of the 24–30% life extension observed for the lsq4 interval as a whole (Vertino et al., 2011). Similarly, rec-8 was the only tested gene for which RNAi improved survival of paraquat (which causes oxidative stress by generation of superoxide), and one of only two that enhanced thermotolerance (Figure 2). These data create a compelling argument that the longevity and stress-resistance traits of lsq4 can be explained by differences in rec-8 expression, arising from allelic alterations in the gene sequence. We note that QTL alleles and mutations extending lifespan are often associated with resistance to one or more (but not all) stresses (Lithgow and Walker, 2002; Shmookler Reis et al., 2007).
We analyzed the 5 kilobase-pairs upstream of rec-8 (reading from mex-5 to rec-8), but found no sequence differences between these strains. At least one single-nucleotide polymorphism (SNP), pkP4058/WBvar00240602, distinguishes between strains CL2a and SR708 in the region just downstream of rec-8 (Vertino et al., 2011). Intronic and coding-sequence SNPs within the rec-8 gene were found for these strains at ~0.1% of nucleotides (i.e., averaging 1 SNP per kbp). Any of these could impact expression, but a systematic mutational scan would be required to determine which SNP is responsible for the QTL differences in traits and rec-8 transcript levels.
REC-8 Alters Survival through Effects on Adult Worms
Each of the distinctive traits associated with the CL2a allele at lsq4 was reproduced or exceeded in recombinant-congenic strain SR708 (differing from CL2a at only a few nucleotides, spanning less than 0.33% of the genome), by exposure of those worms to rec-8 dsRNA. Because germline cells begin to proliferate during the first larval (L1) stage, with germ-cell numbers amplifying progressively in L2–L4 larvae and young adults, RNAi treatment for male-incidence counts was initiated prior to embryogenesis of the tested parents, and maintained throughout their development and egg-laying. For other traits, however, we were able to vary the period of RNAi exposure, commencing either prior to oocyte maturation or at the onset of adulthood. Exposure of strain SR708 to rec-8 dsRNA only during adulthood extended adult lifespan by 26%, which was not inferior to treatment throughout development and aging (Figure 1 and Table 4). This implies that the survival benefits of reduced rec-8 expression primarily reflect diminished REC-8 activity in adults, with little or no involvement of differential effects during development. We cannot exclude a mechanism that entails altered rec-8 expression in germ cells (see the last section of Discussion), but note that this mode of action must be restricted to the germ cells of adult worms, since RNAi treatment was equally effective whether begun at the L4/adult molt or prior to larval development, when most germ cell proliferation occurs.
Deletion of REC8 can Extend the Replicative Lifespan of Haploid Budding Yeast
REC8 deletion from a long-lived S. cerevisiae strain, BY4741, reproducibly added about 20% to the replicative life span of haploid cultures. This was true whether the yeast were grown on glucose or raffinose, and on either carbon source it also applied to rho° mutants of BY4741 (data not shown). We are aware of only one previous study to assess effects of REC8 deletion on S. cerevisiae survival, which reported rec8Δ in an S288C background to have improved survival after “hibernation,” i.e., 58 months storage at 4°C (Postma et al., 2009). Why, then, was mitotic survival not increased (and perhaps reduced, although insignificantly) by the same deletion in the YPK9 strain? REC8 expression in mitotic yeast cells (in particular the haploid cultures studied here, since they do not normally undergo meiosis) is ectopic expression of a meiotic gene. We conjecture that such “leaky” and inappropriate expression is harmful because it aggravates the normal age-dependent destabilization of their genome (McMurray and Gottschling, 2004; Hachinohe et al., 2011). In this case, the YPK9 strain may owe its brief lifespan (30% less than BY4741; see Figure 4) to genomic instability from other sources, which overshadow any contribution from Rec8p, or to causes unrelated to genomic aberrations, leading to death before genome instability can play a role. The REC8 transcript level was roughly 20-fold higher (relative to actin mRNA) in the wild-type YPK9 strain than in BY4741.ch (Figure 4C), consistent with the absence of any selection against high REC8 expression in YPK9.
Possible Mechanisms and Evolutionary Implications
Since rec-8 was not previously recognized as a life-assurance gene or LAG (Tacutu et al., 2013; http://genomics.senescence.info/genes/index.html), nor is it known to interact with any nematode LAGs (Tacutu et al., 2010, 2012; http://netage-project.org/), it appears to employ a novel pathway not hitherto recognized to impact longevity. We infer that this pathway has been widely conserved, based on its effects in both nematodes and yeast. Indeed, rec8 was one of 6 genes with evidence of strong positive selection in the bowhead whale (lifespan >200 years), after its divergence from the Minke whale (lifespan <50 years). It is of interest that the 5 other genes are involved in cancer and/or stress responses (de Magalhaes, Pers. Commun.).
On the basis of transcript profiles, the HIM (high incidence of males) trait, and RNAi results, dimorphic expression of rec-8 appears to be a sufficient explanation for all observed lsq4-associated traits. The molecular basis for those rec-8 traits remains unresolved. Why should lower rec-8 expression or activity be associated with greater longevity and resistance to stresses? Although reduction in REC-8 protein destabilizes the germ-line genome (generating males by X-chromosome non-disjunction), it might favor stability of somatic genomes if normal transcription levels permit “leaky” expression in soma, causing as-yet-unknown harmful effects. Despite evidence from immunocytology that the REC-8 cohesin is observed only in gonads (WormBase, www.wormbase.org), low-level expression in other tissues has not been excluded. Such ectopic expression could destabilize the somatic genome throughout life, potentially reducing lifespan—as occurs in C. elegans when a him-6 mutation produces genome instability (Grabowski et al., 2005). REC-8 has no known role in non-meiotic cells, but its ectopic presence in somatic cells might increase the potential for chromosome fragmentation during aging, consistent with the well-established association of DNA-repair gene expression with aging (Lee et al., 1999; Park et al., 1999; Doria et al., 2004; McCarroll et al., 2004) and reduced induction of such genes in very long-lived mutants (Ayyadevara et al., 2009). The fission yeast (S. pombe) ortholog of REC-8 protein, Rec8p, is also strongly induced in meiosis, but low-level expression in vegetative (mitotic) cells was unambiguously demonstrated for 3 epitope-tagged rec8 constructs (Krawchuck and Wahls, Pers. Commun.). An alternative hypothesis, which does not invoke REC-8 expression in somatic cells, is that genotoxicity in adult germ cells elicits a hermetic protective response in the soma. This possibility was raised by a recent report that DNA damage in germ cells elicits systemic stress resistance through an innate-immunity response (Ermolaeva et al., 2013). Although effects on lifespan were not demonstrated, they might be anticipated as a consequence of altered response to multiple stresses.
Nematode resistance to several stressors could arise from rec-8 suppression under either hypothesis. Thermal and oxidative stresses may in effect “add insult to injury” by increasing the normal burden on DNA repair machinery, which is exacerbated by somatic expression of rec-8. It is a common observation in nematode laboratories that male incidence (and thus X-chromosome non-disjunction) is increased after heat shock or other transient stresses (Ebert et al., 1993; Morran et al., 2009).
We note that rec-8 polymorphism, in our proposed model, constitutes an example of antagonistic pleiotropy in which the allele that is usually preferred under natural selection (i.e., the allele conferring higher rec-8 expression, because it helps to maintain the very low incidence of males that predominates in wild-derived strains), at the same time leads to a disadvantageous phenotype during aging of somatic tissues (shorter lifespan and reduced resistance to potentially genotoxic stresses). Antagonistic pleiotropy with respect to lifespan is thought to arise because long-term survival, and even reproduction later in life, are subject to a progressively declining force of natural selection (Kirkwood and Rose, 1991; Mueller and Rose, 1996). Dimorphic effects of rec-8 may also exemplify a special subclass of balanced polymorphisms wherein each allele confers a selective advantage in a particular context (Modiano et al., 2008; Seidel et al., 2008; Ramalho et al., 2010), since high male frequency (and hence increased mating) constitutes wasted reproductive effort under benign conditions but provides increased segregation of novel gene combinations, and hence greater opportunity for evolutionary adaptation, under harsh conditions in which the force of selection is high.
Funding
This work was supported by grants from the National Institute on Aging (National Institutes of Health), R01-AG091413 and P01-AG20641 to Robert J. Shmookler Reis, and R37-AG006168 to S. Michal Jazwinski. Additional support to Robert J. Shmookler Reis was provided by the Dept. of Veteran Affairs, including a Senior Research Career Scientist award; to Srinivas Ayyadevara through a pilot grant from the UAMS Pepper Center (NIH grant AG028718); and to Robert J. Shmookler Reis and Srinivas Ayyadevara from the Life Extension Foundation.
Conflict of Interest Statement
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Acknowledgments
We thank M. D. Krawchuck and W. P. Wahls for helpful discussions, and for kindly allowing us to cite their unpublished data on epitope-tagging of Rec8p in S. pombe.
Author Note
Because Srinivas Ayyadevara and Robert J. Shmookler Reis are employees of the Veterans Administration, this article is considered to be in the public domain; its contents, however, do not represent the views of the US government or any agency thereof.
Abbreviations
BO, C. elegans var. Bergerac-BO (here, strain RW7000 from R. Waterston); cDNA, complementary DNA; CL2a, C. elegans var. Cl2a (here strain DR1345 from D. Riddle); dsRNA, double-stranded RNA (produced via bidirectional T7 polymerase transcription); HIM, high incidence of males; LS, lifespan; MLS, maximum lifespan (i.e., the lifespan of the last-surviving worm in a cohort); MSP, major sperm protein; N2, C. elegans var. Bristol-N2 (here, subline N2-DRM from D. Riddle); NC, no change; NGM, nematode growth medium; NS, not significant; PCR, polymerase chain reaction; QTL, quantitative trait locus; RT-PCR, real-time, reverse-transcriptase polymerase chain reaction; RNAi, RNA interference; SAM, Significance Analysis of Microarrays; SD, standard deviation; SEM, standard error of the mean; SNP, single nucleotide polymorphism; UV, ultraviolet.
References
Austin, M. U., Liau, W. S., Balamurugan, K., Ashokkumar, B., Said, H. M., and LaMunyon, C. W. (2010). Knockout of the folate transporter folt-1 causes germline and somatic defects in C. elegans. BMC Dev. Biol. 10:46. doi: 10.1186/1471-213X-10-46
Ayyadevara, S., Ayyadevera, R., Hou, S., Thaden, J. J., and Shmookler Reis, R. J. (2001). Genetic mapping of quantitative trait loci governing longevity of Caenorhabditis elegans in recombinant-inbred progeny of a Bergerac-BO x RC301 interstrain cross. Genetics 157, 655–666.
Ayyadevara, S., Ayyadevera, R., Vertino, A., Galecki, A., Thaden, J. J., and Shmookler Reis, R. J. (2003). Genetic loci modulating fitness and life span in Caenorhabditis elegans: categorical trait interval mapping in CL2a x Bergerac-BO recombinant-inbred worms. Genetics 163, 557–570.
Ayyadevara, S., Dandapat, A., Singh, S. P., Siegel, E. R., Shmookler Reis, R. J., Zimniak, L., et al. (2007). Life span and stress resistance of Caenorhabditis elegans are differentially affected by glutathione transferases metabolizing 4-hydroxynon-2-enal. Mech. Ageing Dev. 128, 196–205. doi: 10.1016/j.mad.2006.11.025
Ayyadevara, S., Tazearslan, Ç., Alla, R., Bharill, P., Siegel, E. R., and Shmookler Reis, R. J. (2009). C. elegans PI3K mutants reveal novel genes underlying exceptional stress resistance and lifespan. Aging Cell 8, 706–725. doi: 10.1111/j.1474-9726.2009.00524.x
Cypser, J. R., and Johnson, T. E. (1999). The spe-10 mutant has longer life and increased stress resistance. Neurobiol. Aging 20, 503–512. doi: 10.1016/S0197-4580(99)00085-8
Doria, G., Barattini, P., Scarpaci, S., Puel, A., Guidi, L., and Frasca, D. (2004). Role of immune responsiveness and DNA repair capacity genes in ageing. Ageing Res. Rev. 3, 143–151. doi: 10.1016/j.arr.2003.04.001
Ebert, R. H., Cherkasova, V. A., Dennis, R. A., Wu, J. H., Ruggles, S., Perrin, T. E., et al. (1993). Longevity-determining genes in Caenorhabditis elegans: chromosomal mapping of multiple noninteractive loci. Genetics 135, 1003–1010.
Ebert, R. H., Shammas, M. A., Sohal, B. H., Sohal, R. S., Egilmez, N. K., Ruggles, S., et al. (1996). Defining genes that govern longevity in Caenorhabditis elegans. Dev. Genet. 18, 131–143.
Ermolaeva, M. A., Segref, A., Dakhovnik, A., Ou, H. L., Schneider, J. I., Utermöhlen, O., et al. (2013). DNA damage in germ cells induces an innate immune response that triggers systemic stress resistance. Nature 501, 416–420. doi: 10.1038/nature12452
Grabowski, M. M., Svrzikapa, N., and Tissenbaum, H. A. (2005). Bloom syndrome ortholog HIM-6 maintains genomic stability in C. elegans. Mech. Ageing Dev. 126, 1314–1321. doi: 10.1016/j.mad.2005.08.005
Hachinohe, M., Hanaoka, F., and Masumoto, H. (2011). Hst3 and Hst4 histone deacetylases regulate replicative lifespan by preventing genome instability in Saccharomyces cerevisiae. Genes Cells 16, 467–477. doi: 10.1111/j.1365-2443.2011.01493.x
Hamilton, B., Dong, Y., Shindo, M., Liu, W., Odell, I., Ruvkun, G., et al. (2005). A systematic RNAi screen for longevity genes in C. elegans. Genes Dev. 19, 1544–1555. doi: 10.1101/gad.1308205
Hansen, M., Hsu, A. L., Dillin, A., and Kenyon, C. (2005). New genes tied to endocrine, metabolic, and dietary regulation of lifespan from a Caenorhabditis elegans genomic RNAi screen. PLoS Genet. 1:119–128. doi: 10.1371/journal.pgen.0010017
Hodgkin, J., and Doniach, T. (1997). Natural variation and copulatory plug formation in Caenorhabditis elegans. Genetics 146, 149–164.
Jiang, J. C., Jaruga, E., Repnevskaya, M. V., and Jazwinski, S. M. (2000). An intervention resembling caloric restriction prolongs life span and retards aging in yeast. FASEB J. 14, 2135–2137.
Kamath, R. S., and Ahringer, J. (2003). Genome-wide RNAi screening in Caenorhabditis elegans. Methods 30, 313–321. doi: 10.1016/S1046-2023(03)00050-1
Kim, Y., and Sun, H. (2007). Functional genomic approach to identify novel genes involved in the regulation of oxidative stress resistance and animal lifespan. Aging Cell 6, 489–503. doi: 10.1111/j.1474-9726.2007.00302.x
Kirkwood, T. B., and Rose, M. R. (1991). Evolution of senescence: late survival sacrificed for reproduction. Philos. Trans. R. Soc. Lond. B Biol. Sci. 332, 15–24. doi: 10.1098/rstb.1991.0028
Lai, C. Q., Parnell, L. D., Lyman, R. F., Ordovas, J. M., and Mackay, T. F. (2007). Candidate genes affecting Drosophila life span identified by integrating microarray gene expression analysis and QTL mapping. Mech. Ageing Dev. 128, 237–249. doi: 10.1016/j.mad.2006.12.003
Lee, C. K., Klopp, R. G., Weindruch, R., and Prolla, T. A. (1999). Gene expression profile of aging and its retardation by caloric restriction. Science 285, 1390–1393. doi: 10.1126/science.285.5432.1390
Lee, E. T., and Wang, J. W. (2003). Statistical Methods for Survival Analysis. New York, NY: John Wiley and Sons.
Lithgow, G. J., and Walker, G. A. (2002). Stress resistance as a determinate of C. elegans lifespan. Mech. Ageing Dev. 123, 765–771. doi: 10.1016/S0047-6374(01)00422-5
Lithgow, G. J., White, T. M., Melov, S., and Johnson, T. E. (1995). Thermotolerance and extended life-span conferred by single-gene mutations and induced by thermal stress. Proc. Natl. Acad. Sci. U.S.A. 92, 7540–7544. doi: 10.1073/pnas.92.16.7540
McCarroll, S. A., Murphy, C. T., Zou, S., Pletcher, S. D., Chin, C. S., Jan, Y. N., et al. (2004). Comparing genomic expression patterns across species identifies shared transcriptional profile in aging. Nat. Genet. 36, 197–204. doi: 10.1038/ng1291
McMurray, M. A., and Gottschling, D. E. (2004). Aging and genetic instability in yeast. Curr. Opin. Microbiol. 7, 673–679. doi: 10.1016/j.mib.2004.10.008
Modiano, D., Bancone, G., Ciminelli, B. M., Pompei, F., Blot, I., Simpore, J., et al. (2008). Haemoglobin S and haemoglobin C: ‘quick but costly’ versus ‘slow but gratis’ genetic adaptations to Plasmodium falciparum malaria. Hum. Mol. Genet. 17, 789–799. doi: 10.1093/hmg/ddm350
Morran, L. T., Cappy, B. J., Anderson, J. L., and Phillips, P. C. (2009). Sexual partners for the stressed: facultative outcrossing in the self-fertilizing nematode Caenorhabditis elegans. Evolution 63, 1473–1482. doi: 10.1111/j.1558-5646.2009.00652.x
Mueller, L. D., and Rose, M. R. (1996). Evolutionary theory predicts late-life mortality plateaus. Proc. Natl. Acad. Sci. U.S.A. 93, 15249–15253. doi: 10.1073/pnas.93.26.15249
Park, P. U., Defossez, P. A., and Guarente, L. (1999). Effects of mutations in DNA repair genes on formation of ribosomal DNA circles and life span in Saccharomyces cerevisiae. Mol. Cell. Biol. 19, 3848–3856.
Parsons, C. A., Mroczkowski, H. J., McGuigan, F. E., Albagha, O. M., Manolagas, S., Reid, D. M., et al. (2005). Interspecies synteny mapping identifies a quantitative trait locus for bone mineral density on human chromosome Xp22. Hum. Mol. Genet. 14, 3141–3148. doi: 10.1093/hmg/ddi346
Pasierbek, P., Jantsch, M., Melcher, M., Schleiffer, A., Schweizer, D., and Loidl, J. (2001). A Caenorhabditis elegans cohesion protein with functions in meiotic chromosome pairing and disjunction. Genes Dev. 15, 1349–1360. doi: 10.1101/gad.192701
Postma, L., Lehrach, H., and Ralser, M. (2009). Surviving in the cold: yeast mutants with extended hibernating lifespan are oxidant sensitive. Aging (Albany NY) 1, 957–960.
Ramalho, R. F., Santos, E. J., Guerreiro, J. F., and Meyer, D. (2010). Balanced polymorphism in bottlenecked populations: the case of the CCR5 5′ cis-regulatory region in Amazonian Amerindians. Hum. Immunol. 71, 922–928. doi: 10.1016/j.humimm.2010.05.022
Seidel, H. S., Rockman, M. V., and Kruglyak, L. (2008). Widespread genetic incompatibility in C. elegans maintained by balancing selection. Science 319, 589–594. doi: 10.1126/science.1151107
Shmookler Reis, R. J. (2003). From QTL mapping to genes: the long and winding road. J. Bone Miner. Res. 18, 186–189. doi: 10.1359/jbmr.2003.18.2.186
Shmookler Reis, R. J., Kang, P., and Ayyadevara, S. (2007). Quantitative trait loci define genes and pathways underlying genetic variation in longevity. Exp. Gerontol. 41, 1046–1054. doi: 10.1016/j.exger.2006.06.047
Stumpferl, S. W., Brand, S. E., Jiang, J. C., Korona, B., Tiwari, A., Dai, J., et al. (2012). Natural genetic variation in yeast longevity. Genome Res. 22, 1963–1973. doi: 10.1101/gr.136549.111
Sulston, J., and Hodgkin, J. (1988). “Methods,” in The Nematode Caenorhabditis Elegans, ed W. B. Wood (Cold Spring Harbor, NY: Cold Spring Harbor Laboratory Press), 587–606.
Tacutu, R., Budovsky, A., and Fraifeld, V. E. (2010). The NetAge database: a compendium of networks for longevity, age-related diseases and associated processes. Biogerontology 11, 513–522. doi: 10.1007/s10522-010-9265-8
Tacutu, R., Craig, T., Budovsky, A., Wuttke, D., Lehmann, G., Taranukha, D., et al. (2013). Human Ageing Genomic Resources: integrated databases and tools for the biology and genetics of ageing. Nucleic Acids Res. 41, D1027–D1033. doi: 10.1093/nar/gks1155
Tacutu, R., Shore, D. E., Budovsky, A., de Magalhães, J. P., Ruvkun, G., Fraifeld, V. E., et al. (2012). Prediction of C. elegans longevity genes by human and worm longevity networks. PLoS ONE 7:e48282. doi: 10.1371/journal.pone.0048282
Tarr, D. E., and Scott, A. L. (2004). MSP domain proteins show enhanced expression in male germ line cells. Mol. Biochem. Parasitol. 137, 87–98. doi: 10.1016/j.molbiopara.2004.05.002
Tarr, D. E., and Scott, A. L. (2005). MSP domain proteins. Trends Parasitol. 21, 224–231. doi: 10.1016/j.pt.2005.03.009
Tazearslan, Ç., Ayyadevara, S., Bharill, P., and Shmookler Reis, R. J. (2009). Positive feedback between transcriptional and kinase suppression in nematodes with extraordinary longevity and stress resistance. PLoS Genet. 5:e1000452. doi: 10.1371/journal.pgen.1000452
Tusher, V. G., Tibshirani, R., and Chu, G. (2001). Significance analysis of microarrays applied to the ionizing radiation response. Proc. Natl. Acad. Sci. U.S.A. 98, 5116–5121. doi: 10.1073/pnas.091062498
Van Voorhies, W. A. (1992). Production of sperm reduces nematode lifespan. Nature 360, 456–458. doi: 10.1038/360456a0
Vertino, A., Ayyadevara, S., Thaden, J. J., and Shmookler Reis, R. J. (2011). A narrow quantitative trait locus in C. elegans coordinately affects longevity, thermotolerance, and resistance to paraquat. Front. Genet. 2:63. doi: 10.3389/fgene.2011.00063
Wicky, C., Alpi, A., Passannante, M., Rose, A., Gartner, A., and Muller, F. (2004). Multiple genetic pathways involving the Caenorhabditis elegans Bloom's syndrome genes him-6, rad-51, and top-3 are needed to maintain genome stability in the germ line. Mol. Cell Biol. 24, 5016–5027. doi: 10.1128/MCB.24.11.5016-5027.2004
Keywords: longevity, lifespan, stress resistance, C. elegans, S. cerevisiae, genetics of aging, rec-8 or REC8, meiotic cohesin
Citation: Ayyadevara S, Tazearslan Ç, Alla R, Jiang JC, Jazwinski SM and Shmookler Reis RJ (2014) Rec-8 dimorphism affects longevity, stress resistance and X-chromosome nondisjunction in C. elegans, and replicative lifespan in S. cerevisiae. Front. Genet. 5:211. doi: 10.3389/fgene.2014.00211
Received: 28 March 2014; Accepted: 19 June 2014;
Published online: 04 August 2014.
Edited by:
Heidi A. Tissenbaum, University of Massachusetts Medical School, USAReviewed by:
Vadim Fraifeld, Ben Gurion University of the Negev, IsraelSeung-Jae Lee, Pohang University of Science and Technology, South Korea
Maria Markaki, Foundation for Research and Technology-Hellas, Greece
George A. Garinis, University of Crete, Greece
Copyright © 2014 Ayyadevara, Tazearslan, Alla, Jiang, Jazwinski and Shmookler Reis. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) or licensor are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Robert J. Shmookler Reis, Veterans Affairs Medical Center, GB-105, University of Arkansas for Medical Sciences, 4300 West 7th Street, Little Rock, AR 72205, USA e-mail:cmpzckB1YW1zLmVkdQ==
†Present address: Çagdas Tazearslan, Department of Genetics, Albert Einstein College of Medicine, Bronx, USA