 Victor M. Hernandez-Escalante1†
Victor M. Hernandez-Escalante1† Edna J. Nava-Gonzalez2†
Edna J. Nava-Gonzalez2† V. Saroja Voruganti3
V. Saroja Voruganti3 Jack W. Kent4Karin Haack4
Jack W. Kent4Karin Haack4 Hugo A. Laviada-Molina5
Hugo A. Laviada-Molina5 Fernanda Molina-Segui5Esther C. Gallegos-Cabriales6Juan Carlos Lopez-Alvarenga4Shelley A. Cole4
Fernanda Molina-Segui5Esther C. Gallegos-Cabriales6Juan Carlos Lopez-Alvarenga4Shelley A. Cole4 Marguerite J. Mezzles4
Marguerite J. Mezzles4 Anthony G. Comuzzie4
Anthony G. Comuzzie4 Raul A. Bastarrachea4*
Raul A. Bastarrachea4*- 1Facultad de Medicina, Merida, Universidad Autonoma de Yucatan, Yucatan, Mexico
- 2Facultad de Salud Publica y Nutricion, Universidad Autonoma de Nuevo Leon, Nuevo Leon, Monterrey, Mexico
- 3Nutrition and UNC Nutrition Research Institute, University of North Carolina, Chapel Hill, NC, USA
- 4Department of Genetics, Texas Biomedical Research Institute, San Antonio, TX, USA
- 5Departamento de Investigación, Escuela de Ciencias de la Salud, Universidad Marista de Merida, Merida, Yucatan, Mexico
- 6Facultdad de Enfermeria, Universidad Autonoma de Nuevo Leon, Nuevo Leon, Monterrey, Mexico
The prevalence of type 2 diabetes (T2D) is rising rapidly and in Mexicans is ~19%. T2D is affected by both environmental and genetic factors. Although specific genes have been implicated in T2D risk few of these findings are confirmed in studies of Mexican subjects. Our aim was to replicate associations of 39 single nucleotide polymorphisms (SNPs) from 10 genes with T2D-related phenotypes in a community-based Mexican cohort. Unrelated individuals (n = 259) living in southeastern Mexico were enrolled in the study based at the University of Yucatan School of Medicine in Merida. Phenotypes measured included anthropometric measurements, circulating levels of adipose tissue endocrine factors (leptin, adiponectin, pro-inflammatory cytokines), and insulin, glucose, and blood pressure. Association analyses were conducted by measured genotype analysis implemented in SOLAR, adapted for unrelated individuals. SNP Minor allele frequencies ranged from 2.2 to 48.6%. Nominal associations were found for CNR1, SLC30A8, GCK, and PCSK1 SNPs with systolic blood pressure, insulin and glucose, and for CNR1, SLC30A8, KCNJ11, and PCSK1 SNPs with adiponectin and leptin (p < 0.05). P-values greater than 0.0014 were considered significant. Association of SNPs rs10485170 of CNR1 and rs5215 of KCNJ11 with adiponectin and leptin, respectively, reached near significance (p = 0.002). Significant association (p = 0.001) was observed between plasma leptin and rs5219 of KCNJ11.
Introduction
Epidemiological studies have shown that obesity is associated with an increased risk of mortality (Flegal et al., 2007) including via increased risk of T2D and cardiovascular disease (Ogden et al., 2007). Obesity and T2D have strong genetic components (Rankinen et al., 2006; Sandholt et al., 2010). Recent genome-wide association studies (GWAS) have identified single nucleotide polymorphisms (SNPs) in or near a large number of genes that are related to obesity and T2D phenotypes (Tung and Yeo, 2011). The mechanism by which variation in these genes influences body weight and biomarkers of glucose metabolism is not yet known.
The prevalence of T2D in Mexico in 2000 was 10.9%. By the year 2012 this prevalence almost doubled to 19%. The prevalence of being overweight or obese (BMI ≥ 25 kg/m2) is higher in Mexican women (73.0%) than men (69.4%) similar to obesity alone (BMI ≥ 30 kg/m2) (Gutierrez et al., 2012). These national statistics are reflected in the regions within the country, including the state of Yucatán. Strong predictors of diabetes included a family history of the disease, an increased body mass index, elevated liver-enzyme levels, smoking status, and reduced measures of insulin secretion and action.
As our study subjects are from the Yucatán peninsula, we are interested in the results from a significant study on the state of health in the population of Mérida, Yucatán performed in 1999, which revealed some noteworthy statistics when compared to the prevalence for metabolic and cardiovascular risk factors around Mexico documented in 2000. For adults between 20 and 75 years of age, 45% of men and 73% of women were overweight (BMI ≥ 27.8 for men and ≥ 27.3 for women), and 19.5% of men and 41.1% of women were obese (BMI ≥ 30). T2D was seen in 6.1% of men and 7.4% of women (Arroyo et al., 1999). This may reflect a particular trend for a relatively higher index of these risk factors in the general population of the Yucatán compared to the rest of México as it is seen in more recent epidemiologic studies (Arroyo et al., 2007; Gutierrez et al., 2012). Therefore, intrinsic genetic factors should not be ignored as the Yucatán population has a very unique Maya-Hispanic racial admixture that has experienced very little out-migration.
The genetic factors involved in the development of obesity and T2D include SNPs in genes that encode proteins influencing body composition as well as fat and glucose metabolism (Hirschhorn and Daly, 2005). These includes those influencing: (a) fatty acid oxidation and glucose uptake by adiponectin through increased AMPK and PPAR-α ligand activities (ADIPOR1, ADIPOR2) (Yamauchi et al., 2003; Crimmins and Martin, 2007), (b) proinsulin and proglucagon processing and cleavage (PCSK1) (Choquet et al., 2013), (c) activation of the CB1 receptor to increase de novo lipogenesis in the liver (CNR1) (Osei-Hyiaman et al., 2005), (d) stimulation of the Wnt signaling pathway with activation of B-catenin and target genes in the nucleus, repressing proglucagon synthesis in enteroendocrine cells (TCF7L2) (Jin and Liu, 2008), (e) regulation of glucose metabolism in liver and pancreas (GCK) (Wang et al., 2013), (f) a KATP channel that plays a major role in insulin secretion (KCNJ11) (Olson and Terzic, 2010), and (g) regulation of insulin secretion in humans through a pancreatic β-cell-specific zinc transporter (SLC30A8) (Chimienti et al., 2006). These genes were carefully selected due to their possible role in the etiology of obesity and T2D, in addition to their association with insulin-mediated glucose profile and an abnormal expression of adipose tissue endocrine and inflammatory factors. Such parameters have been well-documented as obesity-related metabolic complication leading to cardiovascular pathology and endothelial dysfunction.
Chronic-degenerative and metabolic diseases related to nutrition, lifestyle changes, and genetic predisposition are increasing at alarming rates. Diseases such as arterial hypertension, T2D, dislipidemias, and obesity, all traditionally viewed as risk factors for coronary heart disease and cerebrovascular disease, now constitute a rapidly advancing, world-wide epidemic (Roger et al., 2011). Many studies indicate that México is in the midst of an epidemiological transition that began in the 1950s. It is experiencing a dramatic decrease in mortality from infectious diseases and a sustained increase in mortality from chronic-degenerative diseases (Rivera et al., 2002). National statistics show that the primary cause of general mortality in Mexico is heart disease (Sanchez-Castillo et al., 2005). These nationally representative surveys are reflected in the regions within the country. Since 1990, the primary cause of death in the adult population in Yucatan has been heart disease (Arroyo et al., 2007). Studies of cardiovascular risk factor's prevalence conducted in the Yucatan, also indicate that the prevalence of such conditions are significantly present in this population when compared to those reported in such Mexican national surveys (Arroyo et al., 1999).
In the present study we hypothesize that a population of unrelated individuals from the Yucatan, sharing a common cultural environment and perhaps peculiar intrinsic genetic factors, may help detect associations between obesity and type 2-related genes and risk factor phenotypes. Thus, a community-based cohort from the Yucatan Peninsula was utilized to examine the associations between the genetic polymorphisms in obesity and T2D-related genes and risk factors of metabolic origin.
Materials and Methods
Study Population
We recruited individuals born and living in the metropolitan area of Merida, Yucatan, a region of southeastern Mexico. These unrelated individuals served as probands used to recruit extended families for the Genetics of Metabolic Disease in Mexico (GEMM) family study (Bastarrachea et al., 2007, 2012). Families were recruited based on family size rather than on any disease status.
Phenotyping
All phenotypes studied constitute risk factors for T2D and obesity. All participants gave written informed consent to participate in this study. The study was approved by the ethical committee of the School of Medicine, Autonomous University of the Yucatan.
The clinical examination included basic anthropometric measures. Blood samples were collected after an overnight fast of at least 12 h. Stature was measured to the nearest centimeter, and weight was measured to the nearest 0.1 kg, with the subject in light clothing and without shoes. BMI (kg/m2) was calculated from this data. Waist circumference was measured to the nearest centimeter with a steel tape measure placed midway between the highest point of the iliac crest and the lowest point of the costal margin. The systolic (first phase) and diastolic (fifth phase) blood pressures were measured to the nearest millimeter Hg with a manually operated sphygmomanometer on the right arm of the seated participant. Three readings were recorded for each individual, and the subject's blood pressure was defined as the average of the second and third readings. Serum samples were obtained from whole blood after clotting.
Upon arrival at Texas Biomedical Research Institute (TBRI), in San Antonio, TX, samples were inventoried. Biochemical phenotypes measured included glucose, insulin, interleukin 6 (IL-6), interleukin 1 beta (IL1β), tumor necrosis factor alpha (TNFα), leptin, and adiponectin. Most of the biochemical phenotypes were analyzed on a Luminex 100 IS platform, consisting of an advanced optometric flux designed to analyze up to 17 different cytokines and chemokines in less than 2 h in ~25 μ L of plasma. An automated clinical chemistry analyzer was used as well as an Immulite 1000, which run ELISA and RIA analysis using ~10 μ L of serum.
SNP Selection
A total of 39 SNPs from 10 genes were selected based on their associations in previously reported GWA studies across various populations (Hirschhorn and Daly, 2005). These genes are adiponectin (ADIPOQ) (Takahashi et al., 2000), adiponectin receptors 1 and 2 (ADIPOR1 and ADIPOR2) (Yamauchi et al., 2003; Crimmins and Martin, 2007), proprotein convertase subtilisin/kexin type 1(PCSK1) (Choquet et al., 2013), cannabinoid receptor 1 (CNR1) (Osei-Hyiaman et al., 2005), fat mass and obesity associated (FTO) (Frayling et al., 2007), glucokinase (GCK) (Matsutani et al., 1992; Wang et al., 2013), potassium inwardly-rectifying channel, subfamily J, member 11 (KNCJ11) (Olson and Terzic, 2010), solute carrier family 30 (zinc transporter), member 8 (SLC30A8) (Chimienti et al., 2006) and transcription factor like7-like 2 (TCF7L2) (Jin and Liu, 2008).
SNP Genotyping
DNA was isolated from frozen buffy coats using organic solvents. Single nucleotide polymorphisms (SNPs) were typed using the multiplex VeraCode technology from Illumina according to the manufacturer's protocol (Illumina, San Diego, CA). Details of the technology are given in Voruganti et al. (2010). Briefly, the technology is based on allele-specific primer extension. Raw data, consisting of intensities of fluorescence, were imported into the analysis software Bead Studio (Illumina). Cluster calls were checked for accuracy and genotypes were exported as text files for further use in association analysis. Replica samples were included as controls for genotyping and allele calling consistencies.
Statistical Analyses
Descriptive statistics and sex-specific differences (Students t-test) were calculated using statistical package for social sciences (SPSS, version 10.0; SPSS Inc., Chicago, USA). Genotype frequencies for each SNP were calculated using a maximum likelihood estimation method and were tested for departures from Hardy-Weinberg equilibrium. Estimates of linkage disequilibrium (LD) between SNPs were determined by calculating pair-wise D' and r2 statistics. As a first step in investigating the association between the SNPs in candidate genes and variation in diabetes-related phenotypes, we employed a measured genotype analysis (Boerwinkle et al., 1986), as implemented in the software package sequential oligogenic linkage analysis routines (SOLAR) (Almasy and Blangero, 1998). This approach extends the classical variance components-based biometrical model to account for both the sporadic effects of unrelated individuals and the main effects of SNP genotypes. For each SNP, we compared this saturated model with a null model in which the main effect of the SNP is constrained to zero. The test statistic, twice the difference in loge (likelihood) between the saturated model and the SNP-specific null, is distributed as a chi-square with one degree of freedom. Based on LD between SNPs, the effective number of SNPs was 32, which was used to calculate the empirical p-value (Ma et al., 2012). After accounting for SNPs in high LD, the p-value of 0.0014 was considered significant.
The T2D and obesity phenotypes were the dependent variables and SNPs were modeled using an additive genetic model (coded 0, 1, and 2 copies of the minor allele). SNP by environment interaction term was added as an additional covariate to the model which was originally adjusted for age, sex, age * sex age2, age2 sex (interaction of age with sex, age squared, and interaction of age squared with sex) and the SNP. (Age squared was included to model possible non-linear effects such as accelerated changes in later life, etc. The interactions model possible differential effects of age in men and women, etc.) All environments (internal and external) were modeled as dichotomous variables.
Results
The study was conducted in 259 (149 women and 110 men) unrelated individuals from the GEMM Family Study. Sex-specific distribution showed significant differences in adiposity-related measures such as body weight, waist circumference, total body fat, leptin, and adiponectin. Systolic blood pressure was also significantly different between men and women, with men having higher pressure. Men also had higher body weight and waist circumference. However, women had significantly higher values of body fat, leptin, and adiponectin. Sex-specific descriptives are shown in Table 1.
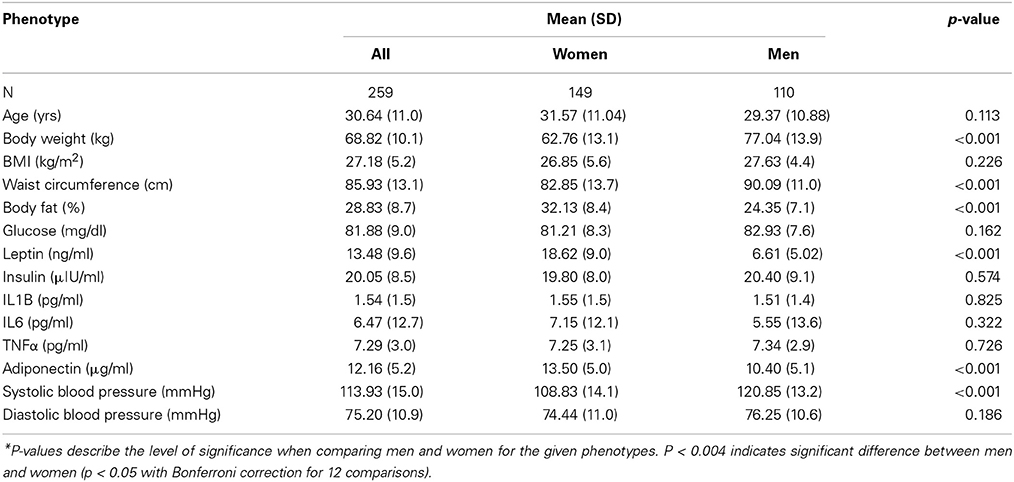
Table 1. Descriptive statistics of phenotypes used in the study.
Of the total 39 SNPs genotyped, 7 were located within or near TCF7L2, 5 from FTO, PCSK1, and ADIPOQ, 4 from SLC30A8 and ADIPOR2, 3 from CNR1 and GCK, 2 from KCNJ11, and one from ADIPOR1. One SNP, ADIPOR2m63422, was dropped from our study due to genotyping error. Minor allele frequency of the remaining 39 SNPs ranged from 2.2 to 48.6% (Table 2). Nominal associations were found for CNR1, SLC30A8, GCK, and PCSK1 SNPs with systolic blood pressure, insulin and glucose, and for CNR1, SLC30A8, KCNJ11, and PCSK1 SNPs with adiponectin and leptin (p < 0.05). Association of SNPs rs10485170 of CNR1 and rs5215 of KCNJ11 with adiponectin and leptin, respectively, reached a p-value of 0.002. After adjusting for multiple testing, the association between plasma leptin levels and rs5219 of KNCJ11 remained significant (Table 3). Minor alleles of rs10485170 and rs5215 were associated with lower levels of adiponectin and leptin, respectively. In contrast, minor allele of rs5219 was associated with higher levels of leptin (Table 4). The SNP by environment interaction was significant for SNP by sex interaction for rs6469675 of SLC30A8 and systolic blood pressure (p = 0.02) and SNP by BMI interaction for rs2268576 of GCK and percent body fat (p = 0.03).
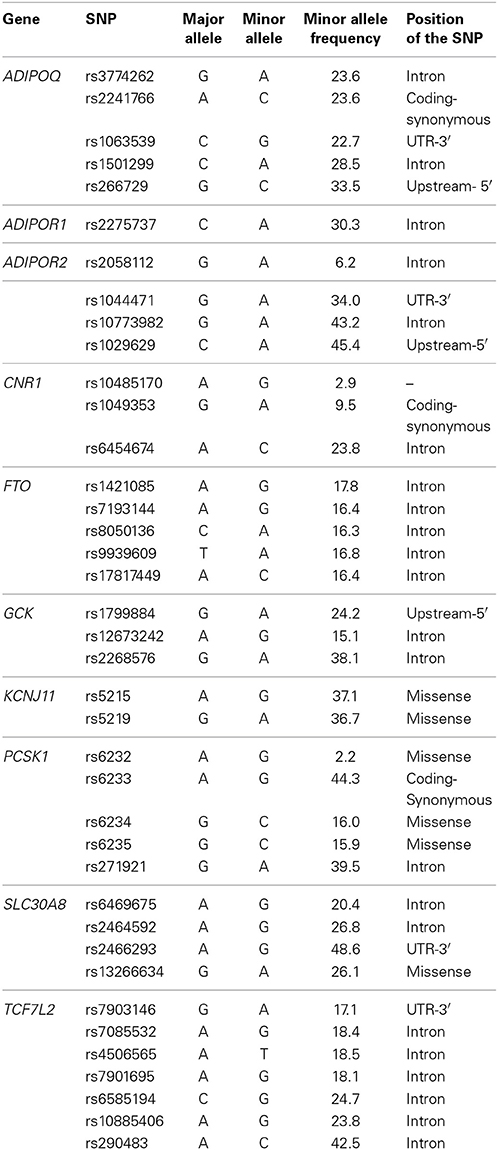
Table 2. Genotype frequencies.
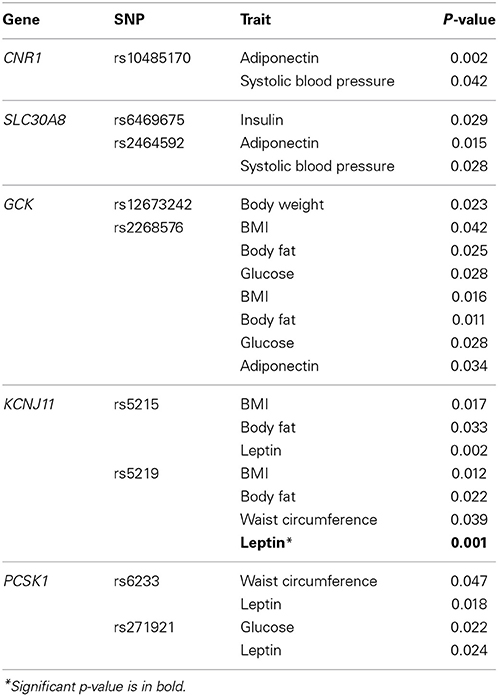
Table 3. Measured genotype analysis results.
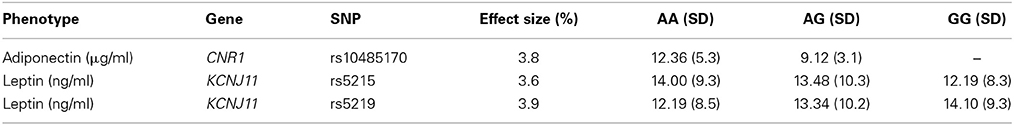
Table 4. Genotype-specific phenotype means for significant or near significant associations.
Discussion
The main finding of this study is the association of the KCNJ11 SNP rs5219 with circulating levels of leptin. KCNJ11 encodes a KATP channel kir6.2 that plays an important role in insulin secretory function in pancreatic β-cells (Olson and Terzic, 2010). Interestingly, leptin is known to inhibit insulin secretion by activating these channels in pancreatic β-cells (Spanswick et al., 2000). Leptin and insulin also modulate the KATP channel function in hypothalamic neurons involved in food intake and body weight (Spanswick et al., 1997; Miki et al., 2001). GWAS have identified several gene polymorphisms to be associated with obesity and T2D risk factors. Our SNPs were selected based on those studies. In the current study, we not only replicated the previously identified association, but also found association of these SNPs with phenotypes not reported in prior GWAS. Besides rs5219 of KCNJ11, rs5215 also showed strong association with leptin levels, reflecting the role of leptin in KATP channel modulation of insulin secretion (Kieffer et al., 1997; Olson and Terzic, 2010).
It is worth mentioning that in our data the rs10485170 SNP from gene CNR1 was not significantly associated with circulating levels of adiponectin (p < 0.002). Previous studies on this gene, which codes for an endocannabinoid receptor 1 that is involved in appetite and energy homeostasis (Despres et al., 2006), have nevertheless shown association with obesity-related phenotypes (Pi-Sunyer et al., 2006; Russo et al., 2007). In rodent models, cannabinoid receptor1 (CB1) inhibits the production and release of adiponectin (Matias et al., 2006). In a study in Romania, SNP rs1049353 was associated with lower adiponectin levels (Dinu et al., 2011). In addition, two human studies did not find evidence of association between CNR1 expression and plasma adiponectin levels (Lofgren et al., 2007; Pagano et al., 2007). Further research is needed in order to elucidate the link between the polymorphisms of the CNR1 gene and adiponectin levels.
Findings from this cohort study suggest that a particular unrelated population born and living in the Yucatan, a region of the southeast in Mexico, sharing the same environment, could help target genes involved in complex traits. The fact that we were able to obtain positive results for associations previously reported in the literature suggests that the study of this Yucatan population can be very useful in research on genetics of common, complex, highly prevalent diseases.
Author Contributions
Victor M. Hernandez-Escalante, Hugo A. Laviada-Molina, and Fernanda Molina-Segui collected the samples and prepared the database; Shelley A. Cole, Karin Haack, and John W. Kent Jr. processed samples in the laboratory; Edna J. Nava-Gonzalez, Marguerite J. Mezzles, V. Saroja Voruganti, and Juan C. Lopez-Alvarenga performed genetic and epidemiological analyses; Raul A. Bastarrachea, Victor M. Hernandez-Escalante, Edna J. Nava-Gonzalez, and Marguerite J. Mezzles wrote the manuscript; Anthony G. Comuzzie and Esther C. Gallegos-Cabriales proofread the manuscript and incorporated changes and corrections for the final draft.
Conflict of Interest Statement
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Acknowledgments
We thank the Consejo Nacional de Ciencia y Tecnologia (CONACYT) of the Mexican Federal Government (Basic Sciences Fund 2006-01 Agreement 59431) and the Health Foundation of Mexico Yucatan Chapter for the grant to finance this study.
References
Almasy, L., and Blangero, J. (1998). Multipoint quantitative-trait linkage analysis in general pedigrees. Am. J. Hum. Genet. 62, 1198–1211. doi: 10.1086/301844
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Arroyo, P., Fernández, V., Loría, A., Pardio, J., Laviada, H., Vargas-Ancoma, L., et al. (2007). Obesity, body morphology, and blood pressure in urban and rural population groups of Yucatan. Salud Publica Mex. 49, 274–285. doi: 10.1590/S0036-36342007000400007
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Arroyo, P., Pardio, J., Fernandez, V., Vargas-Ancona, L., Canul, G., and Loria, A. (1999). Obesity and cultural environment in the Yucatán region. Nutr. Rev. 57, S78–S83.
Bastarrachea, R. A., Gallegos-Cabriales, E. C., Nava-González, E. J., Haack, K., Voruganti, V. S., Charlesworth, J., et al. (2012). Integrating genomic analysis with the genetic basis of gene expression: preliminary evidence of the identification of causal genes for cardiovascular and metabolic traits related to nutrition in Mexicans. Adv. Nutr. 3, 596S–604S. doi: 10.3945/an.112.001925
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Bastarrachea, R. A., Kent, J. W. Jr., Rozada, G., Cole, S. A., Alvarenga, J. L., Aradillas, C., et al. (2007). Heritability and genetic correlations of metabolic disease-related phenotypes in Mexico: preliminary report from the GEMM family study. Hum. Biol. 78, 121–130. doi: 10.1353/hub.2007.0021
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Boerwinkle, E., Chakraborty, R., and Sing, C. F. (1986). The use of measured genotype information in the analysis of quantitative phenotypes in man. I. Models and analytical methods. Ann. Hum. Genet. 50, 181–194.
Chimienti, F., Favier, A., and Seve, M. (2006). ZnT-8, a pancreatic beta-cell-specific zinc transporter. Biometals 18, 313–317. doi: 10.1007/s10534-005-3687-9
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Choquet, H., Kasberger, J., Hamidovic, A., and Jorgenson, E. (2013). Contribution of common PCSK1 genetic variants to obesity in 8,359 subjects from multi-ethnic American population. PLoS ONE 8:e57857. doi: 10.1371/journal.pone.0057857
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Crimmins, N. A., and Martin, L. J. (2007). Polymorphisms in adiponectin receptor genes ADIPOR1 and ADIPOR2 and insulin resistance. Obes. Rev. 8, 419–423. doi: 10.1111/j.1467-789X.2007.00348.x
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Despres, J. P., Lemieux, I., and Almeras, N. (2006). Contribution of CB1 blockade to the management of high-risk abdominal obesity. Int. J. Obes. 30, S44–S52. doi: 10.1038/sj.ijo.0803278
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Dinu, I. R., Popa, S. G., Moţa, M., Mota, E., Stanciulescu, C., Ioana, M., et al. (2011). The association of the rs1049353 polymorphism of the CNR1 gene with hypoadiponectinemia. Rom. J. Morphol. Embryol. 52, 791–795.
Flegal, K. M., Graubard, B. I., Williamson, D. F., and Gail, M. G. (2007). Cause-specific excess deaths associated with underweight, overweight, and obesity. JAMA 298, 2028–2037. doi: 10.1001/jama.298.17.2028
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Frayling, T. M., Timpson, N. J., Weedon, M. N., Zeggini, E., Freathy, R. M., Lindgren, C., et al. (2007). A common variant in the FTO gene is associated with body mass index and predisposes to childhood and adult obesity. Science 316, 889–894. doi: 10.1126/science.1141634
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Gutierrez, J. P., Rivera-Dommarco, J., Shamah-Levy, T., Villalpando-Hernandez, S., Franco, A., Cuevas-Nasu, L., et al. (2012). Encuesta Nacional de Salud y Nutricion 2012. Resultados Nacionales. Cuernavaca, Mexico: Instituto Nacional de Salud Pública (MX).
Hirschhorn, J. N., and Daly, M. J. (2005). Genome-wide association studies for common diseases and complex traits. Nat. Rev. Genet. 6, 95–108. doi: 10.1038/nrg1521
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Jin, T., and Liu, L. (2008). The Wnt signaling pathway effector TCF7L2 and type 2 diabetes mellitus. Mol. Endocrinol. 22, 2383–2392. doi: 10.1210/me.2008-0135
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Kieffer, T. J., Heller, R. S., Leech, C. A., Holz, G. G., and Habener, J. F. (1997). Leptin suppression of insulin secretion by the activation of ATP-sensitive K+ channels in pancreatic beta-cells. Diabetes 46, 1087–1093.
Lofgren, P., Sjolin, E., Wahlen, K., and Hoffstedt, J. (2007). Human adipose tissue cannabinoid receptor 1 gene expression is not related to fat cell function of adiponectin levels. J. Clin. Endocrinol. Metab. 92, 1555–1559. doi: 10.1210/jc.2006-2240
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Ma, L., Andrew, G. C., and Alon, K. (2012). Gene-based testing of interactions in association studies of quantitative traits. PLoS Genet. 9:e1003321. doi: 10.1371/journal.pgen.1003321
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Matias, I., Gonthier, M. P., Orlando, P., Martiadis, V., De Petrocellis, L., Cervino, C., et al. (2006). Regulation, function and dysregulation of endocannainoids in models of adipose and beta-pancreatic cells and in obesity and hyperglycemia. J. Clin. Endocrinol. Metab. 91, 3171–3180. doi: 10.1210/jc.2005-2679
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Matsutani, A., Janssen, R., Donis-Keller, H., and Permutt, M. A. (1992). A polymorphic(CA)n repeat element maps the human glucokinase gene (GCK) to chromosome 7p. Genomics 12, 319–325.
Miki, T., Liss, B., Minami, K., Shiuchi, T., Saraya, A., Kashima, Y., et al. (2001). ATP-sensitive K+ channels in the hypothalamus are essential for the maintenance of glucose homeostasis. Nat. Neurosci. 4, 507–512. doi: 10.1038/87455
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Ogden, C. L., Yanovski, S. Z., Carroll, M. D., and Flegal, K. M. (2007). The epidemiology of obesity. Gastroenterology 132, 2087–2102. doi: 10.1053/j.gastro.2007.03.052
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Olson, T. M., and Terzic, A. (2010). Human KATP channelopathies: diseases of metabolic homeostasis. Pfugers Arch. 460, 295–306. doi: 10.1007/s00424-009-0771-y
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Osei-Hyiaman, D., Depetrillo, M., Pacher, P., Liu, J., Radaeva, S., Batkai, S., et al. (2005). Endocannabinoid activation at hepatic CB1 receptors stimulates fatty acid synthesis and contributes to diet-induced obesity. J. Clin. Invest. 115, 1298–1305. doi: 10.1172/JCI200523057
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Pagano, C., Pilon, C., Calcagno, A., Urbanet, R., Rossato, M., Milan, G., et al. (2007). The endogenous cannabinoid system stimulates glucose uptake in human fat cells via phosphophatidylinositol 3-kinase and calcium-dependent mechanisms. J. Clin. Endocrinol. Metab. 92, 4810–4819. doi: 10.1210/jc.2007-0768
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Pi-Sunyer, F. X., Aronne, L. J., Heshmati, H. M., Devin, J., Rosenstock, J., and RIO-North American Study Group (2006). Effect of rimonobant, a cannabinoid-1 receptor blocker, on weight and cardiometabolic risk factors in overweight or obese patients. JAMA 295, 761–775. doi: 10.1001/jama.295.7.761
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Rankinen, T., Zuberi, A., Chagnon, Y. C., Weisnagel, S. J., Argyropoulos, G., Walts, B., et al. (2006). The human obesity gene map: the 2005 update. Obesity 14, 529–644. doi: 10.1038/oby.2006.71
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Rivera, J. A., Barquera, S., Campirano, F., Campos, I., Safdie, M., and Tovar, V. (2002). Epidemiological and nutritional transition in Mexico: rapid increase of non-communicable chronic diseases and obesity. Public Health Nutr. 5, 113–122. doi: 10.1079/PHN2001282
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Roger, V. L., Go, A. S., Lloyd-Jones, D. M., Adams, R. J., Berry, J. D., Brown, T. M., et al. (2011). Heart disease and stroke statistics–2011 update: a report from the American Heart Association. Circulation 123, e18–e209. doi: 10.1161/CIR.0b013e3182009701
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Russo, P., Strazzullo, P., Cappuccio, F. P., Tregouet, D. A., Lauria, F., Loguercio, M., et al. (2007). Genetic variations at the endocannabinoid type 1 receptor gene (CNR1) are associated with obesity phenotypes in men. J. Clin. Endocrinol. Metab. 92, 2382–2386. doi: 10.1210/jc.2006-2523
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Sanchez-Castillo, C. P., Velasquez-Monroy, O., Lara-Esqueda, A., Berber, A., Sepulveda, J., Tapia-Conver, R., et al. (2005). Diabetes and hypertension increases in a society with abdominal obesity: results of the Mexican national health survey 2000. Public Health Nutr. 8, 53–60. doi: 10.1079/PHN2004659t
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Sandholt, C. H., Sparso, T., Grarup, N., Albrechtsen, A., Almind, K., Hansen, L., et al. (2010). Combined analyses of 20 common obesity susceptibility variants. Diabetes 59, 1667–1673. doi: 10.2337/db09-1042
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Spanswick, D., Smith, M. A., Groppi, V. E., Logan, S. D., and Ashford, M. L. (1997). Leptin inhibits hypothalamic neurons by activation of ATP-sensitive potassium channels. Nature 390, 521–525.
Spanswick, D., Smith, M. A., Mirshamsi, S., Routh, V. H., and Ashford, M. L. (2000). Insulin activates ATP-sensitive K+ channels in hypothalamic neurons of lean, but not obese rats. Nat. Neurosci. 3, 757–758. doi: 10.1038/77660
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Takahashi, M., Arita, Y., Yamagata, K., Matsukawa, Y., Okutomi, K., Horie, M., et al. (2000). Genomic structure and mutations in adipose-specific gene, adiponectin. Int. J. Obes. 24, 861–868. doi: 10.1038/sj.ijo.0801244
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Tung, Y. C., and Yeo, G. S. (2011). From GWAS to biology: lessons from FTO. Ann. N. Y. Acad. Sci. 1220, 162–171. doi: 10.1111/j.1749-6632.2010.05903.x
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Voruganti, V. S., Cole, S. A., Ebbesson, S. O., Goring, H. H., Haack, K., Laston, S., et al. (2010). Genetic variation in APOJ, LPL and TNFRSF10B affects plasma fatty acid distribution in Alaskan Eskimos. Am. J. Clin. Nutr. 9, 1574–1583. doi: 10.3945/ajcn.2009.28927
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Wang, R., Gao, H., Xu, W., Li, H., Mao, Y., Wang, Y., et al. (2013). Differential expression of genes and changes in glucose metabolism in the liver of liver-specific glucokinase gene knockout mice. Gene 516, 248–254. doi: 10.1016/j.gene.2012.12.036
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Yamauchi, T., Kamon, J., Ito, Y., Tsuchida, A., Yokomizo, T., Kita, S., et al. (2003). Cloning of adiponectin receptors that mediate antidiabetic metabolic effects. Nature 423, 762–769. doi: 10.1038/nature01705
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Keywords: Single Nucleotide Polymorphisms (SNP), association analysis, minor allele frequency, type 2 diabetes, obesity
Citation: Hernandez-Escalante VM, Nava-Gonzalez EJ, Voruganti VS, Kent JW, Haack K, Laviada-Molina HA, Molina-Segui F, Gallegos-Cabriales EC, Lopez-Alvarenga JC, Cole SA, Mezzles MJ, Comuzzie AG and Bastarrachea RA (2014) Replication of obesity and diabetes-related SNP associations in individuals from Yucatán, México. Front. Genet. 5:380. doi: 10.3389/fgene.2014.00380
Received: 31 July 2014; Paper pending published: 23 August 2014;
Accepted: 15 October 2014; Published online: 18 November 2014.
Edited by:
Ramu Elango, Princess Al-Jawhara Centre of Excellence in Research of Hereditary Disorders (PACER-HD), Saudi ArabiaCopyright © 2014 Hernandez-Escalante, Nava-Gonzalez, Voruganti, Kent, Haack, Laviada-Molina, Molina-Segui, Gallegos-Cabriales, Lopez-Alvarenga, Cole, Mezzles, Comuzzie and Bastarrachea. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) or licensor are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Raul A. Bastarrachea, Department of Genetics, Texas Biomedical Research Institute, 7620 NW Loop 410, San Antonio, TX 78227, USA e-mail:cmF1bGJzQHR4YmlvbWVkZ2VuZXRpY3Mub3Jn
†These authors have contributed equally to this work.