 Peter F. Cowman
Peter F. Cowman- Department of Ecology and Evolutionary Biology, Yale University, New Haven, CT, USA
Biodiversity patterns across the marine tropics have intrigued evolutionary biologists and ecologists alike. Tropical coral reefs host 1/3 of all marine species of fish on 0.1% of the ocean’s surface. Yet our understanding of how mechanistic processes have underpinned the generation of this diversity is limited. However, it has become clear that the biogeographic history of the marine tropics has played an important role in shaping the diversity of tropical reef fishes we see today. In the last decade, molecular phylogenies and age estimation techniques have provided a temporal framework in which the ancestral biogeographic origins of reef fish lineages have been inferred, but few have included fully sampled phylogenies or made inferences at a global scale. We are currently at a point where new sequencing technologies are accelerating the reconstruction and the resolution of the Fish Tree of Life. How will a complete phylogeny of fishes benefit the study of biodiversity in the tropics? Here, I review the literature concerning the evolutionary history of reef-associated fishes from a biogeographic perspective. I summarize the major biogeographic and climatic events over the last 65 million years that have regionalized the tropical marine belt and what effect they have had on the molecular record of fishes and global biodiversity patterns. By examining recent phylogenetic trees of major reef associated groups, I identify gaps to be filled in order to obtain a clearer picture of the origins of coral reef fish assemblages. Finally, I discuss questions that remain to be answered and new approaches to uncover the mechanistic processes that underpin the evolution of biodiversity on coral reefs.
Introduction
A latitudinal gradient in species diversity is a common feature of many taxonomic groups, both terrestrial and marine (Willig et al., 2003; Hillebrand, 2004). However, a longitudinal gradient in species diversity is also apparent across the marine tropics. Fishes exemplify this diversity gradient (Hughes et al., 2002; Tittensor et al., 2010) driven largely by patterns of species richness associated with tropical coral reef habitats. Species richness of reef associated fishes forms an enigmatic “bullseye” pattern centered on the Indo-Australian Archipelago (IAA; Figure 1A). This region has also been called several other names (reviewed by Hoeksema, 2007), but its position at the center of this species richness gradient has given it status as the largest marine biodiversity hotspot, covering two thirds of the global equatorial tropics (Bellwood et al., 2012). Unlike terrestrial biodiversity hotspots (Myers, 1988; Myers et al., 2000), centers of endemism are not concordant with the center of highest species diversity, whether endemic species are defined by regional checklists (Figure 1B), or the extent of their geographic range (Hughes et al., 2002; but see Mora et al., 2003). Traditional hotspot analysis of the marine environment has identified endemic centers under high levels of threat (Roberts et al., 2008), however these 10 defined areas of endemism exclude some areas that have the high diversity of overlapping, wide ranging species. In addition to the distinctive biodiversity gradient, the tropics have been divided into a number of realms, regions, provinces and eco-regions based on shared environmental characteristics (Spalding et al., 2007), composition of endemic taxa (Briggs and Bowen, 2012), or measures of species dissimilarity (Kulbicki et al., 2013). Although these differing regional schemes are based on present day patterns, it appears that the division of regional assemblages across the tropics is linked to its biogeographic history and the formation of several historical barriers to dispersal (Cowman and Bellwood, 2013a,b). While environmental clines in sea surface temperature are linked to latitudinal variation in diversity (Tittensor et al., 2010), the extensive tectonic, eustatic, climatic, oceanographic and geomorphological (TECOG; Bellwood et al., 2012) processes have played an important role in the origin and maintenance of the tropical biodiversity gradient spanning both deep and shallow times scales (Renema et al., 2008; Pellissier et al., 2014).
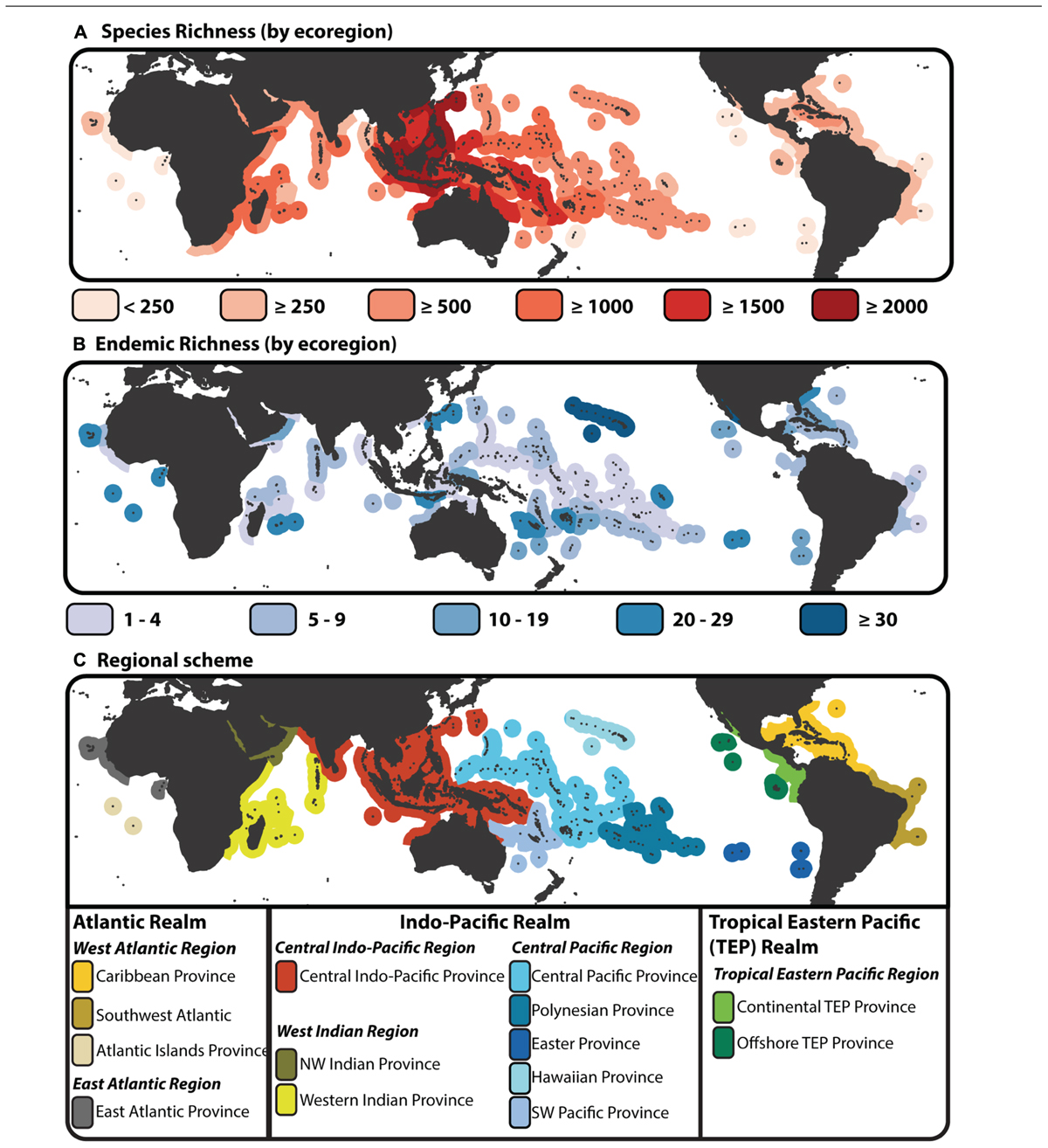
FIGURE 1. Species richness, endemism and provinciality of tropical reef fishes. (A) Map of species biodiversity by tropical ecoregion (Spalding et al., 2007) with color gradient denoting areas of high species richness (dark red) to areas of low species richness (light red). (B) Map of endemic species by ecoregion. Under this scheme a species is endemic if it is only found in a single ecoregion, i.e., a regional assessment of endemism rather that designated by percent of area comparison (Hughes et al., 2002). Species richness and endemic estimates are based on species counts from the “checklist” × “all species” dataset of Kulbicki et al. (2013). (C) Biogeographic delineation of tropical Realms, Regions, and Provinces based on species dissimilarity analysis of Kulbicki et al. (2013). This biogeographic scheme is base on checklists as base units (see Kulbicki et al., 2013), however here the scheme is imposed onto the tropical ecoregions of Spalding et al. (2007).
The mechanistic processes that underpinned this history and how they have generated such biodiversity has inspired much debate over the last 40 years (Potts, 1985; Briggs, 1999; Bellwood and Meyer, 2009a; Cowman and Bellwood, 2013a) with numerous hypotheses being proposed (Bellwood et al., 2012), but little consensus. The answers to key questions regarding where species have originated and the processes that have promoted speciation and extinction in tropical clades remain unclear. The popularity of phylogenetics, fossil-calibrated age estimation techniques, and the availability of geographic information have allowed biologists to examine the history of the taxa that form the marine biodiversity hotspot. In the case of coral reef fishes and the IAA biodiversity hotspot, what have we learned in the last decade? While new genomic sequencing methods are becoming available (Faircloth et al., 2013) and larger datasets are increasing the resolution of deeper nodes in the Fish Tree of Life (Near et al., 2012), what gaps remain in the evolutionary record of reef associated fishes? How complete is our understanding of the evolution of biodiversity on tropical reefs and which questions will benefit from more sampling, more data, and new analytical approaches?
In this review, I examine the phylogenetic and biogeographic completeness of key families found in reef habitats globally. By exploring the current state of the biogeographic history of tropical reef fishes I highlight where further analysis and discussion is needed, and what new questions require answers.
Evolution of Fishes on Coral Reefs – Filling in the Gaps
Bellwood and Wainwright (2002) discussed the biogeographic history of fishes on coral reefs. They stated that from the integration of systematics, biogeography, ecology, and paleontology a new understanding of the nature of reef fishes would arise. Twelve years on, the integration of methods and multiple datasets has cast a wide net across the fields of reef fish ecology, evolution and biogeography to give vast insight into the important phases of evolution of coral reef fishes over the past 400 million years (Bellwood et al., in press). A major part of this insight has come from the combination of molecular phylogenetics and the fossil record to form a temporal framework in which to ask questions regarding the origin and tempo of reef fish diversification. Although only a handful of new fossils have been described with reef affinities in the last decade (Carnevale, 2006; Micklich et al., 2009; Bannikov, 2010) the fossil record of early reef fish forms continues to provide a wealth of information on the early origins of reef association in teleost fishes. New analytical techniques have revealed the origins and diversification of anatomical features (Friedman, 2010), important morphological transitions (Goatley et al., 2010), and the emergence of essential functional roles on coral reefs (Bellwood et al., 2014a,b). However, it is the utility of fossils as calibrations points on molecular phylogenies that have allowed the evolution of reef associated lineages to be studied on an absolute timescale. In particular, while the origins of several reef fish groups can be found in the fossil deposits of the Monte Bolca Lagerstätten (50 mya; Bellwood, 1996) the crown ages and the diversification of major lineages that lack a fossil record have only been examined with the aid of calibrated chronograms.
There has been debate over what characterizes a ‘coral reef’ fish (Bellwood, 1998; Robertson, 1998), but a general list of ‘reef’ fish families (Table 1) identifies those groups that are characteristic of a modern reef assemblage (both coral and rocky reefs), regardless of geographic location (Bellwood and Wainwright, 2002). Indeed, species counts of these families on coral reefs around the world are found in relatively similar proportions (Bellwood and Hughes, 2001). Although these nine fish families found on coral reefs are often used as model groups to address questions regarding diversification on coral reefs, there are at least 35 families of acanthomorph fishes that can be considered ‘reef associated’ (Price et al., 2014). Some of these families are monotypic (e.g., Zanclidae) while other can be entirely reef dwelling but not globally distributed (e.g., Siganidae). Interestingly, the most diverse fish family found on reefs, the Gobiidae, containing over 2000 species, has several lineages confined to coral reefs (Herler et al., 2011), yet remains off the list of traditional reef fish families. Its exclusion from this list may be related to their consistent undersampling in geographic surveys (Ackerman and Bellwood, 2000), made ever more difficult by their cryptic nature and many undescribed species. Nonetheless, this non-traditional reef fish family may provide a good model to study speciation and biodiversity on coral reefs (Rüber et al., 2003; Taylor and Hellberg, 2005). While this review focuses on those nine families classically recognized as reef fish families, other lineages found on (and off) reefs might provide further insight into the evolution of tropical biodiversity. The utility of these families and lineages should be determined by several factors, most importantly, the level at which they have been sampled for phylogenetic reconstruction. As the nine reef fish families are prominent on coral reef around the globe they have been examined with phylogenetic methods and increasing levels of genetic data over the past two decades.
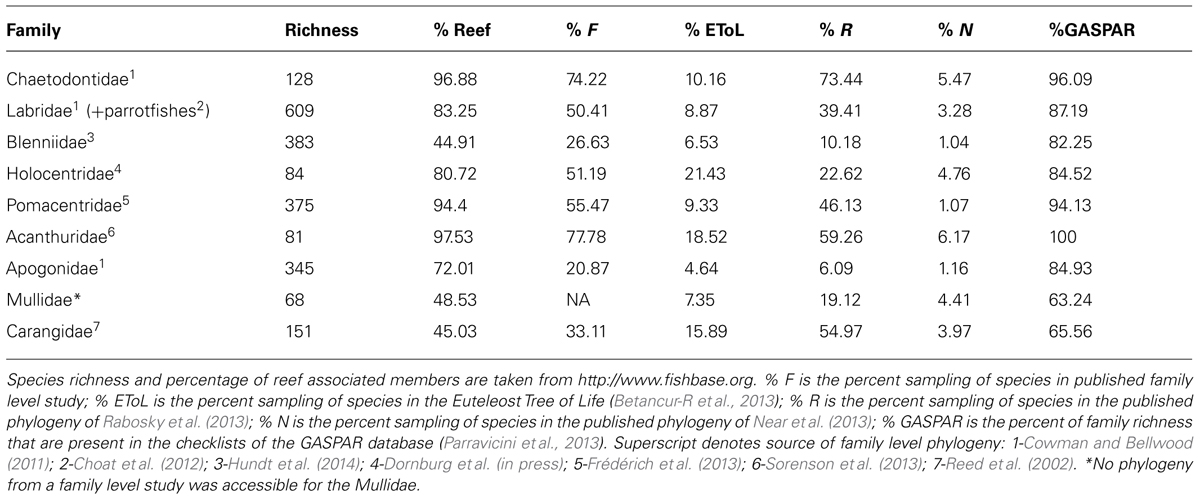
TABLE 1. Diversity, phylogenetic and geographic sampling of the nine characteristic reef fish families.
Early molecular phylogenetic studies within reef fish groups contained a small number of taxa in select genera (Lacson and Nelson, 1993; McMillan et al., 1999; Bernardi et al., 2001; Reed et al., 2002). Later, the combination of generic level phylogenies with relaxed clock methods (Sanderson, 2003) allowed the estimation of ages of divergence within several reef-associated lineages (Bellwood et al., 2004; Bernardi et al., 2004; Klanten et al., 2004; Barber and Bellwood, 2005; Read et al., 2006). With improved sequencing efforts at the family level (Westneat and Alfaro, 2005; Cooper et al., 2009; Thacker and Roje, 2009), molecular datasets have given insight into the crown origins of reef fish groups and the tempo at which they have diversified (Westneat and Alfaro, 2005; Alfaro et al., 2007; Fessler and Westneat, 2007; Cowman et al., 2009; Frédérich et al., 2013). Even though the characteristic nine families have been the focus of many phylogenetic studies (albeit some more than others), as of yet, not one of these families is represented by a fully sampled, species level phylogeny (Table 1). While the majority of major lineages and genera are sampled within these phylogenetic studies, the level at which species within these lineages are sampled varies dramatically (Figure 2). Those families that have more completely sampled phylogenies have achieved it through the combination of multiple sequence datasets and the use of supermatrix phylogenetic methods. The combination of datasets for the butterflyfish family Chaetodontidae (Fessler and Westneat, 2007; Bellwood et al., 2010) has resulted in a phylogeny that is over 70% complete (Table 1; Cowman and Bellwood, 2011). Similarly, the family Acanthuridae is nearly complete (76%) through the combination of previously published and new sequence data (Sorenson et al., 2013). Other families have been the focus of several phylogenetic studies, incrementally increasing taxon sampling as more data or specimens become available, e.g., the wrasses, family Labridae (now inclusive of odacids and parrotfishes; Westneat and Alfaro, 2005; Alfaro et al., 2009; Cowman et al., 2009; Kazancioglu et al., 2009; Cowman and Bellwood, 2011); and the damselfishes, family Pomacentridae (Cooper et al., 2009; Cowman and Bellwood, 2011; Frédérich et al., 2013). Within the Labridae and Pomacentridae, shallower lineages have also been examined with increased sampling to explore a variety of evolutionary and ecological questions (Smith et al., 2008; Choat et al., 2012; Hodge et al., 2012; Litsios et al., 2012). Other families, such as the Blenniidae and the Apogonidae have been plagued by taxonomic issues that are only beginning to be addressed with more taxa and multi-locus datasets (Thacker and Roje, 2009; Hundt et al., 2014). The incomplete phylogenetic sampling for reef fishes is exacerbated by the rate of new species descriptions and identification of cryptic species (Zapata and Robertson, 2006; Mora et al., 2008; Bowen et al., 2013). Recently, Allen (2014) reviewed the systematics of Indo-Pacific coral reef fishes over the past three decades to reveal that over 1,400 new species have been described with an average of 51.3 new species description per year since 2010.
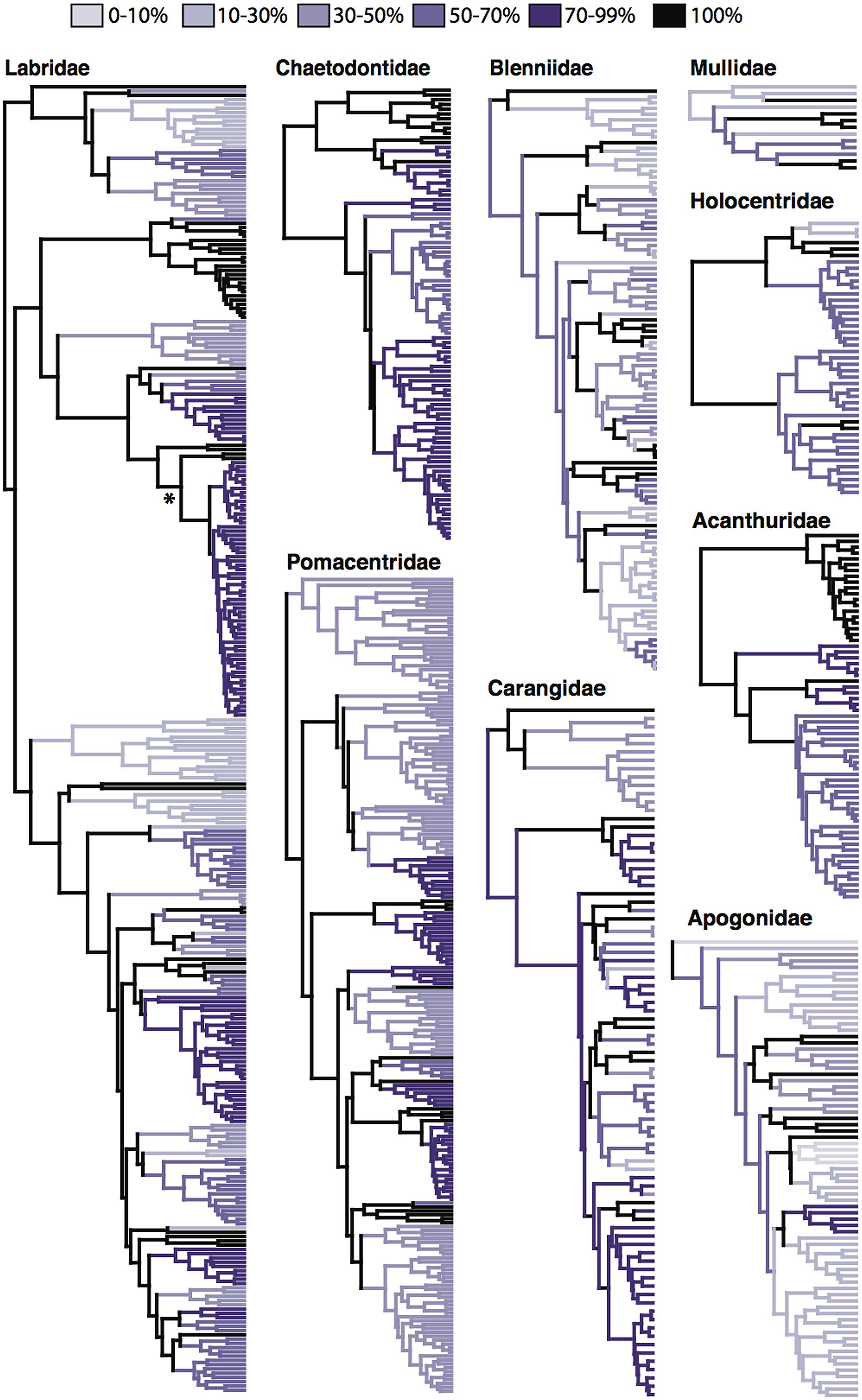
FIGURE 2. Phylogenetic sampling of characteristic reef fish families. Published chronologies of the nine characteristic reef fish families found globally on coral reefs (Bellwood and Wainwright, 2002). Sources of these trees can be found in Table 1. Level of taxon sampling per lineage is denoted by color with black branches completely sample. Percent sampling was calculated by a per genus basis with species counts taken from Fishbase (http://www.fishbase.org). In the cases of the families Labridae, Chaetodontidae, Pomacentridae and Apogonidae lineage richness estimates were taken from Cowman and Bellwood (2011). Asterisk indicates node where the parrotfish phylogeny of Choat et al. (2012) was grafted to the Labridae tree of Cowman and Bellwood (2011).
The incomplete sampling observed in these reef fish families appears to have been a general symptom seen across all fishes when compared to other vertebrate branches of the Tree of Life (Thomson and Shaffer, 2010a). However, there have been three recent efforts in reconstructing the Fish Tree of Life (Betancur-R et al., 2013; Near et al., 2013; Rabosky et al., 2013) with new sequencing methods (Faircloth et al., 2013) providing an exciting avenue for future phylogenomic research in fishes. These ‘top down’ approaches to reconstructing the Fish Tree of Life have greatly improved the resolution of deep nodes and divergences in the major fish groups, including those with reef affinities. These datasets have included varying degrees of taxon sampling of reef associated lineages (Table 1), depending on the core aim of the study. The chronogram of Near et al. (2013) concentrated on sampling all families of acanthomorphs with as complete a molecular matrix as possible. While it does not have high species level sampling of reef fish lineages, it has allowed the exploration of rates of transition of fish lineages (at the family level) on and off of reefs over the past 100 million years (Price et al., 2014). The chronogram of Rabosky et al. (2013) closely matches the sampling effort of family level studies of the nine characteristic groups, achieved by mining the published sequence data available on GenBank. In the cases of the families Carangidae and Mullidae, this concatenated super-matrix approach included more species than any other published phylogeny for each family (Table 1). These large-scale phylogenetic studies employing supermatrix methods have also allowed the identification of the closest sister families to prominent reef fish families. However, disagreement among these large phylogenies still exists for some families. For example, the closest sister group to the Labridae changes from being the family Centrogenyidae (Betancur-R et al., 2013), to the family Ammodytidae (Rabosky et al., 2013), to the family Gerridae (Near et al., 2013). This highlights the utility of supermatrix approaches, but caution is still needed in their implementation (Thomson and Shaffer, 2010b). In the case of fishes, more work remains to resolve some of these early diverging lineages at the top of the percomorph “bush” (Nelson, 1989), where the origin of several reef associated lineages are found. While these top–down approaches continue to reveal the early evolution of reef fishes, ‘bottom–up’ studies concentrating on the origins of extant lineages have provided a framework to examine the diversification of reef fishes over the past 65 million years.
Diversification of Fishes on Tropical Reefs
Phylogenetic sampling and the resolution of reef fish lineages remains a key issue for future research. However, for those groups that have been the focus of age estimation studies, some general, concordant patterns have emerged. The stem lineages of many reef lineages extend back into the Cretaceous (Near et al., 2013) while the crown origins are strongly associated with the aftermath of the K-Pg boundary mass extinction event (∼65 ma; Bellwood et al., in press). A recent study of family level transitions into reef habitat and associated morphological divergence has outlined two waves of colonization before and after the K-Pg boundary (Price et al., 2014). Initial colonization of lineages before the K-Pg boundary (90–72 mya) was accompanied with morphological divergence of clades, while the subsequent wave of reef colonization (65–56 mya) appears to saturate with increasing convergence in morphospace (Price et al., 2014). Patterns of reef invasions within families are likely to be more dynamic (Price et al., 2014) with trophic evolution showing increasing association between fishes and the reef benthos (Cowman et al., 2009; Bellwood et al., 2010). From the appearance of more generalist trophic modes in the Eocene/Oligocene, new and novel trophic modes began to appear on reefs in the Miocene with the trophic system in place by 7 mya (Cowman et al., 2009; Bellwood et al., 2010). Some reef fish lineages have diversified ecologically by expanding into novel areas of morphospace (Friedman, 2010; Price et al., 2011) while others have exhibited convergent radiations across similar trophic strategies (Frédérich et al., 2013). An association with coral reef habitat appears to both promote clade diversity, with higher reef occupancy linked to faster rates of diversification (Alfaro et al., 2007; Cowman and Bellwood, 2011), and increased rates of morphological diversification within lineages (Price et al., 2011, 2013). Lineage diversity and morphological divergence do not appear to be related in these groups (Cowman et al., 2009; Price et al., 2011), however key innovations have been linked to increased diversity in some clades (Kazancioglu et al., 2009; Litsios et al., 2012; Wainwright et al., 2012). In addition, an over arching link between rate of body size evolution and rate of diversification appears to be a general trend across the fish tree of life (Rabosky et al., 2013), but its affect on the evolution of reef clades has not been examined.
By the end of the Eocene, major lineages leading to present day genera and tribes within reef fish families were in place for many reef fish families (Cowman and Bellwood, 2011). After what may have been a cryptic extinction event near the Eocene/Oligocene boundary, coinciding with the origin of the butterflyfishes (∼33 mya), a rebound in cladogenesis within reef associated lineages during the Oligocene/Miocene underpinned much of the extant diversity seen on todays reefs (Cowman and Bellwood, 2011). Several lineages within the Labridae, Pomacentridae, Apogonidae and Chaetodontidae display significantly more diversity than expected, with the most reef-associated lineages appearing more resistant to higher extinction rates than their non-reef counterparts (Cowman and Bellwood, 2011). Elevated cladogenesis has previously been identified in several marine fish lineages (Rüber and Zardoya, 2005), with reef association or habitat shifts suggested to be the underlying mechanism. Later, the relationship between reef association and elevated rates of diversification was demonstrated in Tetraodontiformes (Alfaro et al., 2007), marine gastropods (Williams and Duda, 2008), and more recently in sharks (Sorenson et al., 2014). Whether reef habitats promote this diversity through elevated speciation, or relaxed extinction remains to be seen. As extinction rates are notoriously difficult to estimate from molecular phylogenies in the absence of a paleontological record (Quental and Marshall, 2009, 2010; Rabosky, 2009b), the vital evidence in the form of Miocene fossils for many reef fish lineages, at least, remains out of reach. The expansion of coral reef habitat in the Miocene may have promoted cladogenesis, and provided a refuge from extinction, two processes that may vary on both temporal and geographic scales (Cowman and Bellwood, 2013a).
The majority of extant coral reef fishes examined by Cowman and Bellwood (2011, 2013a) are of Miocene age (23–5 mya; Figure 3) with some possibly being older than the IAA hotspot (Renema et al., 2008). While some geographic variation in the reconstructed ages of lineages exists (Figure 3), the older ages of extant species challenged the early suggestion that sea level fluctuations during the Pleistocene was a major factor in the origin of modern coral reef assemblages (Potts, 1985). For reef fishes, the majority of cladogenetic events occurred in the Miocene but speciation still continues in several groups from the Pleistocene onward (Rocha and Bowen, 2008). Pleistocene speciation in these groups may be linked to patterns of barrier vicariance (Bowen et al., 2013; Cowman and Bellwood, 2013b), peripheral budding (Hodge et al., 2012), and more recent fluctuations in coral reef stability (Pellissier et al., 2014). Pleistocene processes may very well have played an active role in the evolution of butterflyfishes (McMillan and Palumbi, 1995), which display younger global ages of extant lineages (∼2.6 mya) than labrids (∼6.7 mya) and pomacentrids (∼6.7 mya), particularly in the Indian Ocean and IAA hotspot (Figure 3). It is likely that when the gaps in taxonomic sampling of these reef fish families are filled, and cryptic species are identified, the inclusion of unsampled lineages closer to the present may enhance the role played by speciation in the Pleistocene and the importance of peripheral locations in promoting biodiversity. Processes at work in the Miocene appear to be the main source of origination of modern reef fish biodiversity patterns, while processes maintaining this pattern are prominent from the Pliocene/Pleistocene. However, to gain a clearer picture of the magnitude of these processes across the marine tropics, studies with a biogeographic focus have been important.
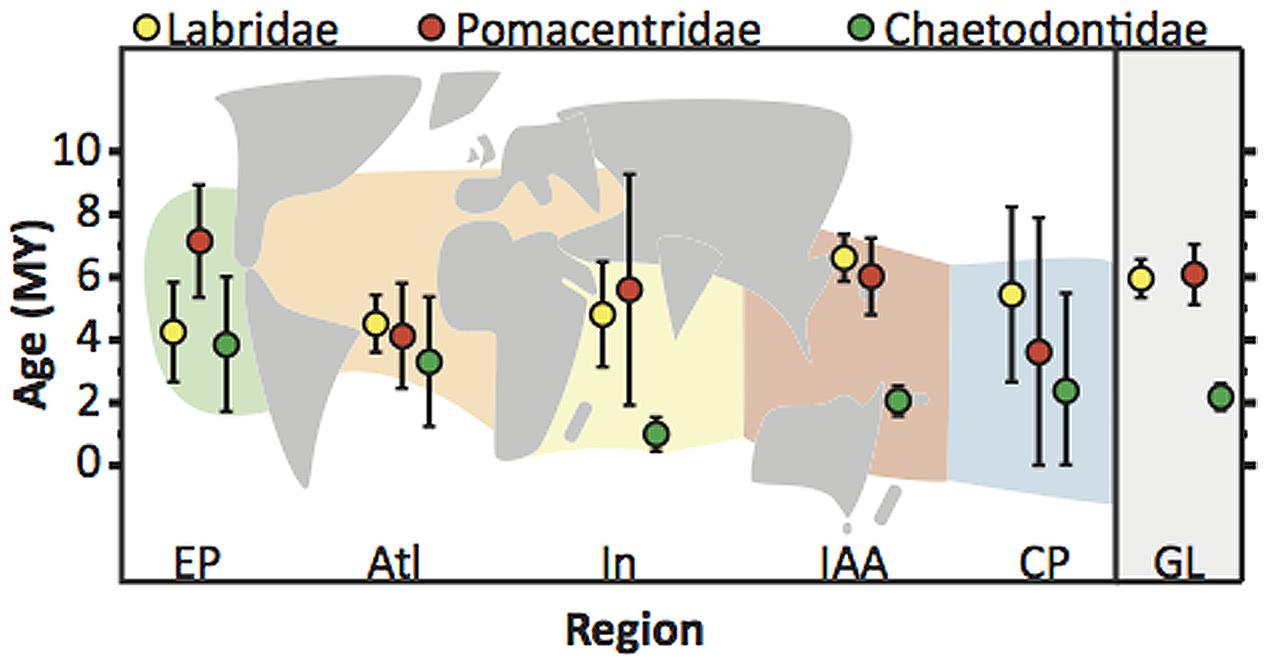
FIGURE 3. Biogeographic ages of species of the families Labridae, Pomacentridae, and Chaetodontidae. Plot shows mean (circle) and 95% CI (whiskers) of the distribution of ages of origination of extant lineages in each biogeographic region, and globally for the Labridae, Pomacentridae, Chaetodontidae (data from Cowman and Bellwood, 2013a). Underlying schematic map shows regional scheme used by Cowman and Bellwood (2013a) for ancestral biogeographic reconstruction. This scheme differs from the regional scheme of Kulbicki et al. (2013) shown in Figure 1C. Age distributions for each region represent the ages of extant lineages that originated in that particular region accounting for ancestral biogeographic reconstruction (see Cowman and Bellwood, 2013a). EP, East Pacific; Atl, Atlantic; In, Indian Ocean; IAA, IAA Hotspot; CP, Central Pacific Islands; GL, Global.
Biogeography and Biodiversity
As with the phylogenetic history of reef fishes, the biogeographic history is reliant on sampling, specifically, knowledge of the current extend of reef fish species ranges. In this regard, we are fortunate to have had many skilled ichthyologists throughout the decades collecting geographic information on reef fish distributions (Allen, 2014). Several initiatives have been actively cataloging the diversity found on and off coral reefs, e.g., Atlas of Living Australia1; IUCN red list2; the Global Biodiversity Information Facility3; Map of Life4; and Ocean Biogeographic Information System5. A recent effort to construct a global database for tropical reef fishes has resulted in over 6300 records for reef fishes across 169 locations (GASPAR database; Kulbicki et al., 2013; Parravicini et al., 2013). This database has been used to explore global predictors of reef fish species richness (Parravicini et al., 2013); global biogeography of reef fishes (Kulbicki et al., 2013); human mediated losses of phylogenetic and functional diversity (D’agata et al., 2014); and the role of stable reef habitat in preserving reef fish diversity (Pellissier et al., 2014).
Of the valid nominal species in the nine reef fish families, these geographic checklists (Kulbicki et al., 2013; Parravicini et al., 2013) include the vast majority of them, ranging from 63% of carangid species, to 100% of acanthurid species (Table 1). These data are likely to include the majority, if not all reef associated members of these families. In combination with a fully sampled phylogeny of reef fishes, these geographic data would allow us to tease apart some of the questions that have been partially answered so far regarded the origins of tropical biodiversity. Unfortunately, fully sampled phylogenies for important groups are still out of reach. The incomplete and clade biased phylogenetic sampling also translates into a bias in geographic sampling (Figure 4). Those charismatic families such as the Labridae and Chaetodontidae that have been the focus of several papers from a variety of research groups have more even phylogenetic sampling across biogeographic ecoregions, with over 50% of taxa present in each region represented in a published phylogeny (Figures 4A,B). A sampling bias can be observed among ocean basins, and among families, where some families (Apogonidae, Blenniidae; Figures 4E,F) have higher phylogenetic sampling in Indo-Pacific locations, whereas others (Pomacentridae, Mullidae, Carangidae; Figures 4C,H,I) show higher phylogenetic sampling in the Atlantic locations. Overall, the families Apogonidae, Blenniidae, Mullidae and Carangidae show concerning levels of lower phylogenetic resolution across tropical reef habitats (Figures 4E,F,H,I) with many regions showing below 10% phylogenetic sampling of ecoregion assemblages. Nonetheless, the geographic data available, regardless of its sampling in phylogenetic trees, have been fruitful in delineating biogeographic regions across the marine tropics.
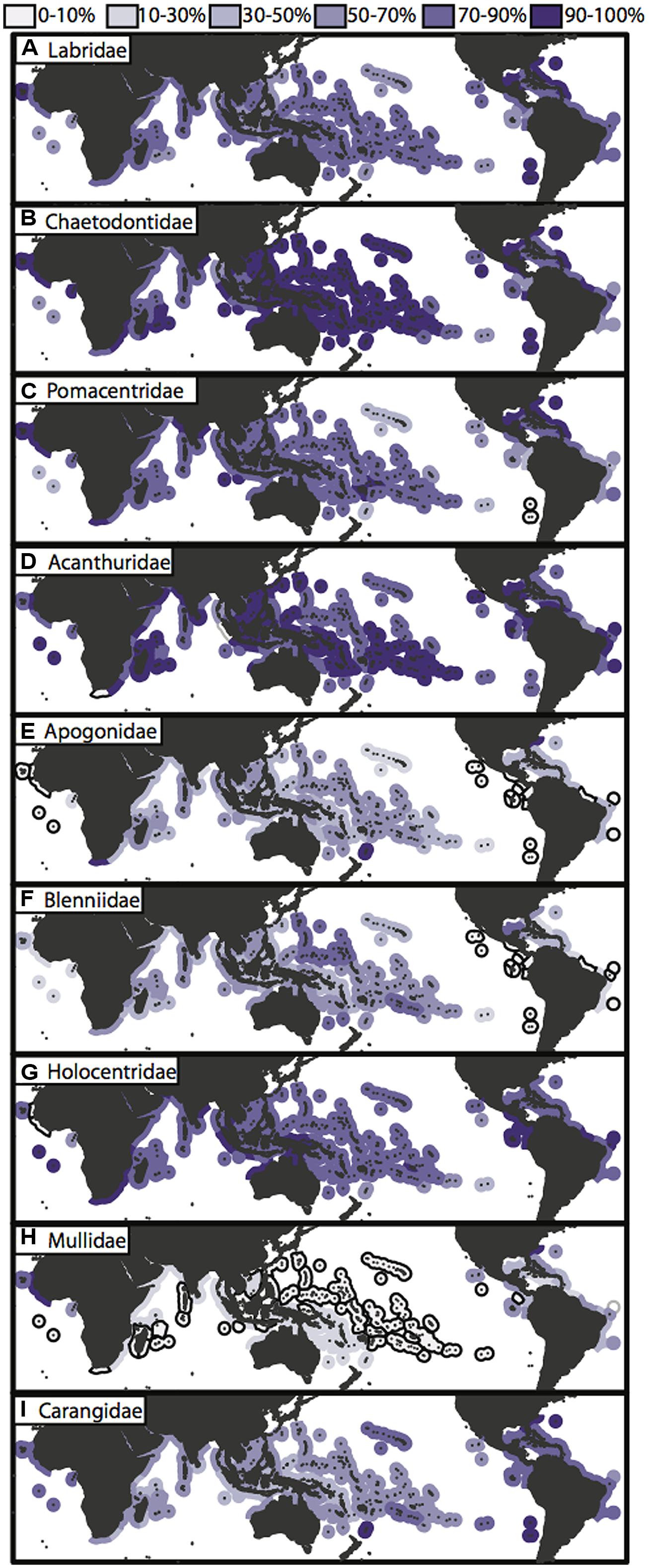
FIGURE 4. Phylogenetic sampling of nine reef fish families across the marine tropics. (A–I) Global maps of tropical ecoregions displaying phylogenetic sampling of species assemblages for each of the nine characteristic reef fish families. Species richness for each family within ecoregions is base on species checklist of species counts from the “checklist” × “all species” dataset of Kulbicki et al. (2013) and phylogenetic sampling is based on taxon sampling of each published family phylogeny (Figure 2; Table 1). Ecoregions that have <10% of the family species pool represented are outlined in black.
Biogeographic science has an important role in the guidance of biodiversity conservation (Whittaker et al., 2005). Dividing the tropics into discrete regions has proven to be a difficult process (Mouillot et al., 2013), but it is a necessary step toward critically evaluating and implementing conservation priorities (Whiting et al., 2000; Olson et al., 2001). With the predictions of a grim future ahead for coral reef systems under a changing climate (Hughes et al., 2003), the biogeographic delineation of the marine tropics and how regional assemblages have formed through time is paramount to our understanding of biodiversity maintenance. In the past decade, several studies have provided a schematic break down of regions across the tropical belt based on differing criteria (Spalding et al., 2007; Briggs and Bowen, 2012; Kulbicki et al., 2013). Most recently, Kulbicki et al. (2013) used a hierarchical approach to delineating tropical reef regions based on species dissimilarity. Using species checklists across 169 locations (Parravicini et al., 2013), their results identify three realms (Atlantic, Central Indo-Pacific, Tropical East Pacific; Figure 1C), each with varying degrees of structure within those delineated regions and provinces (Figure 1C; Kulbicki et al., 2013). The Central Indo-Pacific region, within the Indo-Pacific realm, was characterized by lower within region dissimilarity, while neighboring regions (Western Indian and Central Pacific) could be broken down further into provinces (although some internal structure is seen when analyses were based on ecoregions as base units; see Kulbicki et al., 2013). While the IAA (or Coral Triangle) may be delineated as the area containing the highest proportion of reef fish species (Briggs and Bowen, 2012), in terms of species composition there is no strong evidence delineating it as a separate entity in the Central Indo-Pacific (Kulbicki et al., 2013). The IAA biodiversity hotspot may not be a defined region based on species dissimilarity. But, an area the extent of the IAA at the center of the highest number of overlapping species ranges must have played a significant role on an evolutionary scale in the generation of current day biodiversity patterns. On a shallow timescale, the complex role of the IAA and Coral Triangle region has been illustrated through numerous population level and phylogeographic studies (reviews by Carpenter et al., 2011). While the extant ranges of reef associated fishes can statistically delineate regions of dissimilar assemblages, the lines of division are highly dependent on the method used (Leprieur et al., 2012; Mouillot et al., 2013), and there remains no consensus on which method or regional scheme is best. It is likely that the appropriate biogeography scheme will depend on the question being addressed. From a macroevolutionary perspective, whether any present day scheme for biogeographic delineation has a meaning for past diversification and biogeographic evolution has yet to be addressed.
The evolution of reef fish biodiversity patterns is likely to be concordant with the evolution of coral reef habitats. Higher diversification rates of reef associated fish lineages have been demonstrated (Cowman and Bellwood, 2011) and transitions onto coral reefs appear important for accelerated morphological evolution (Price et al., 2014). However, a direct (or indirect) link between the diversification of corals and the diversification of their associated fish lineages has yet to be recognized (Duchene et al., 2013). From a biogeographic perspective, the spatial and temporal distribution of coral taxa and the platforms they construct may provide insight into the evolution of reef fishes that inhabit them. Extent of coral reef area (Bellwood and Hughes, 2001) and its stability through time (Pellissier et al., 2014) have been highlighted as significant predictors of extant reef fish biodiversity. The fossil record of reef building corals highlight differences among ocean basins (Budd, 2000; Wallace and Rosen, 2006). The Atlantic and Caribbean fossil reef biota display high turnover of coral species and extinction of reef habitat (Budd, 2000), while the Indo-Pacific fossil biota displays a history of eastward movement linked to tectonic activity (Wilson and Rosen, 1998; Renema et al., 2008) with modern coral taxa in the Central Indo-Pacific consisting of Tethyan relicts and recent speciation events (Wallace and Rosen, 2006). Such data could be used to model the spatial and temporal dynamics of coral reef habitat allowing us to test more explicit biogeographic scenarios and hypothesis related to tropical biodiversity (e.g., Ree and Sanmartín, 2009).
The IAA Biodiversity Hotspot – A Center of Confusion
Although compositionally the IAA hotspot may not currently present a geographic entity within the Central Indo-Pacific Realm (Kulbicki et al., 2013), the area has historically been recognized as a center of biodiversity in the Indo-Pacific (Ekman, 1953). In an effort to understand the processes that have been important in producing the diversity pattern across the Indo-Pacific and the associated center of high diversity, three hypotheses became popular in the early 1980s, originally formulated to explain the biodiversity of reef building corals (summerized by Potts, 1985). These ‘center of’ hypotheses have been co-opted in the context of reef fish biodiversity. They have been expanded and modified to explain the extensive and overlapping widespread ranges seen in several reef fish groups (Hughes et al., 2002; Connolly et al., 2003). The details of each of these, and other hypotheses have been reviewed by Bellwood et al. (2012). Both phylogenetic and population level studies of reef fish taxa have highlighted evidence describing the IAA (or the Coral Triangle) as a center of origin (Briggs, 2003; Timm and Kochzius, 2008), a center of overlap (Hubert et al., 2012; Gaither and Rocha, 2013), or a center of accumulation/survival (Barber and Bellwood, 2005; Kool et al., 2011).
Each hypothesis has made predictions about the location of origin of species, their age, and their trajectory of range expansion or change (see Bellwood et al., 2012). Primarily, species with restricted endemic ranges have been important in the assessment of these hypotheses, but even the study of endemic taxa has been fraught with debate (Bellwood and Meyer, 2009a,b; Briggs, 2009). Even how an endemic range is defined can lead to conflicting patterns of endemism (Hughes et al., 2002; Mora et al., 2003). Bellwood and Meyer (2009a,b) highlighted the diffuse ages of endemic taxa, whether they are found inside or outside the IAA. Endemic taxa can be young (neo-endemics) or old (palaeo-endemics) and as such their use to delineate areas of species geographic origin should be cautious. Indeed the ages of endemic coral reef fishes in several families do not differ significantly from those of more widespread species (Hodge et al., 2014). Instead of using endemic species as a tool in pinpointing locations of species origin it is becoming clear that understanding how processes of isolation and extinction have lead to current patterns of endemism along side widespread species is an important step in the study of reef fish biodiversity.
Recent studies have begun to highlight that the processes that promote, maintain and diffuse biodiversity in the marine tropics are more dynamic in nature with multiple drivers acting both in concert, and decoupled across temporal and geographic scales (Bowen et al., 2013; Cowman and Bellwood, 2013a). The question has changed from which hypothesis is most accurate, to when and where the processes they invoke have been most prevalent and how they have interacted to produce the biodiversity we see today. To this end, it may be time to mute the discussion about ‘centers of’ in the field of reef fish biodiversity in favor of directly examining and modeling rates of speciation, extinction and dispersal in a temporal and geographic framework. Such methods have been advantageous in investigating terrestrial diversity patterns on global scales (Jetz et al., 2012; Rolland et al., 2014). If these different processes have played an active role in the development of tropical biodiversity but on different temporal and spatial scales, then several biogeographic areas may have historically acted as sources or sinks (or both) for biodiversity at different periods in time. For example, the Atlantic realm, like the IAA hotspot, can be considered a center for species origination (Cowman and Bellwood, 2013a), but its history of isolation from the Indo-Pacific (Floeter et al., 2001; Joyeux et al., 2001) and extinction (O’Dea et al., 2007) has contributed to its lower diversity when compared to the Indo-Pacific. Both the Indian Ocean and the Central Pacific regions have higher standing diversity of reef fishes than the Atlantic. However, most of their diversity has been derived through expansions of lineages from the IAA. But peripheral locations in both these regions have also been sites of species origination (Hodge et al., 2013). In addition, within the Indo-Pacific realm, it remains unclear if the IAA hotspot actually has experienced higher rates of speciation than adjacent regions (Bellwood and Meyer, 2009b; Litsios et al., 2014). While speciation has certainly occurred within the IAA hotspot, peripheral locations are also important sources of new species (Bowen et al., 2013; Hodge et al., 2014). None of these hypotheses can be disregarded, but nor can any one of them solely explain the IAA biodiversity pattern (Rosen, 1984; Palumbi, 1997; Halas and Winterbottom, 2009; Hoeksema, 2009). Halas and Winterbottom (2009), comparing reconstructed area relationships of cladograms of fishes, corals and molluscs, found little congruence among these taxa and little evidence for any of the core models examined, despite these groups displaying very similar patterns of diversity across the tropics (Roberts et al., 2008). Several studies have asked what present day geographic or environmental factors explain the variation in the IAA diversity pattern (Mora et al., 2003; Tittensor et al., 2010; Parravicini et al., 2013), but it appears that examining historical factors may have more explanatory power when examining the origin and maintenance of biodiversity in the marine tropics (Renema et al., 2008; Pellissier et al., 2014). While the history of tropical biodiversity may remain clouded until complete phylogenies are available, concordant patterns in currently published data for tropical reef fishes has allowed key events in the history of the tropics to be recognized.
Though the IAA hotspot is enigmatic, it has not been a unique pattern through time. It represents the modern manifestation of a pattern that has existed for at least the past 50 million years (Renema et al., 2008). The center of biodiversity has ‘hopped’ from a Tethyan location (Paleocene), to an Arabian/IAA hotspot (Eocene/Oligocene), to its current location in the IAA (Miocene; Renema et al., 2008). This biogeographic re-centering of biodiversity was associated with a sequence of TECOG events (Bellwood et al., 2012), dynamic processes controlling the origin and survival of species (Cowman and Bellwood, 2011, 2013a) resulting in the establishment of a trophic system characteristic of modern coral reefs. These processes resulted in the contraction and expansion of carbonate platforms, the evolution of the coral species that built them, and their associated fish lineages.
The earliest fossil records of lineages leading to modern coral reef fishes and the coral genus Acropora are found in close proximity in the Late Paleocene/Early Eocene deposits of Europe and the Western Indian Ocean (Bellwood, 1996; Wallace and Rosen, 2006). These deposits can be realistically extrapolated to be associated with the ancestral hotspot centered in the Western Tethys seaway (Renema et al., 2008). No fossil Acropora are currently know from the Eocene of the Indo-West Pacific. While this could be an observational artifact, this gap in the coral record corresponds to a geographic gap with fewer shallow water habitat for coral growth in the Indo-Pacific at that time (Wilson and Rosen, 1998). It is not until the Late Oligocene/Early Miocene (∼26 mya) where we see the first fossil evidence of coral species of the genus Acropora occurring in the IAA (Wallace and Rosen, 2006). From this time, the tectonic collision of Australian and South East Asian plate fragments favored localized isolation and origination of new coral taxa and the expansion of carbonate platforms in the IAA (Wilson and Rosen, 1998). It is during this time we also see the demise of carbonate platforms in Europe and the Mediterranean deposits (Wallace and Rosen, 2006) and the collapse of the ancestral Tethyan and Arabian biodiversity hotspots (Renema et al., 2008). This collapse of the ancestral hotspots is associated with an eastward shift in fossil deposits of reef associated organisms (Renema et al., 2008) and the expansion of carbonate platforms in the IAA (Wallace and Rosen, 2006). A period of high extinction may be visible in the molecular record of some reef fish groups coinciding with a decrease in fossil numbers of all marine taxa (Cowman and Bellwood, 2011). More fossil data for focal fish groups is required to confirm this pattern, however, a total evidence approach including fossil taxa as dated tips in an ancestral biogeographic framework for the family Holocentridae holds promising insight (Dornburg et al., in press).
The Miocene epoch represents an important phase in the evolution of the IAA biodiversity hotspot (Bellwood et al., in press), with the expansion of both coral reef platforms (Wallace and Rosen, 2006) and associated fish lineages (Cowman and Bellwood, 2013a). As a result of tectonic activity we see the final closure of the Tethys seaway, known as the Terminal Tethyan Event (TTE, 18–12 mya; Steininger and Rögl, 1979) and the development of the Isthmus of Panama (IOP; Coates and Obando, 1996) isolating the Atlantic and Caribbean from the Indo-Pacific. The development and closure of these ‘hard’ land barriers would have been associated with climatic upheaval (Hallam, 1994; Montes et al., 2012) and extinction in reef locations (McCoy and Heck, 1976; Budd, 2000; O’Dea et al., 2007). This has led to a diffuse pattern of vicariance in the molecular record of some reef fish families (Cowman and Bellwood, 2013b). The TTE and the IOP barriers in conjunction with the expanse of ocean known as the East Pacific Barrier (EPB; Bellwood and Wainwright, 2002) have left a lasting mark on modern tropical reef fish assemblages (Kulbicki et al., 2013). However, some recent dispersal from the Indian Ocean into the Atlantic has been detected (Rocha et al., 2005a; Bowen et al., 2006), with several lineages maintaining gene flow across the EPB (Lessios and Robertson, 2006).
In terms of coral reef ecology, the Miocene holds the origins of many novel feeding modes (Cowman et al., 2009; Bellwood et al., 2010), and an escalation in herbivory and detritivory that have become essential services performed by fishes on healthy coral reefs (Hughes et al., 2011). In the Labridae, coral reef associated lineages show significantly higher rates of trophic ecomorphological evolution with over a third of that diversity seen within trophic modes only found on coral reefs (Price et al., 2011). A switch to consuming low quality food items has been linked to higher rates of diversification in several coral reef lineages with origins in the Oligo-Miocene (Lobato et al., 2014). This reflects fossil evidence showing the transition of reef fish forms to exploiting the epilithic algal matrix, an underutilized resource on coral reef flats (Bellwood et al., 2014a).
By the end of the Miocene, the center of fish diversity has taken shape in the IAA (Cowman and Bellwood, 2013a) and important trophic components are in place on coral reefs (Cowman et al., 2009; Price et al., 2011; Bellwood et al., 2014a). Speciation continues in several lineages from the Pliocene to Recent time periods. Expansion of lineage ranges from the IAA to adjacent regions is common (Cowman and Bellwood, 2013a) with vicariance and speciation in peripheral locations (Hodge et al., 2014). In the Atlantic realm, Pliocene speciation has been described in several reef associated genera (Floeter et al., 2008), with evidence of ecological speciation (Rocha et al., 2005b). While ecological speciation is likely to be ongoing in the Indo-Pacific, recent studies reflect a complex history of sympatric, allopatric and parapatric speciation (Rocha and Bowen, 2008; Choat et al., 2012; Hodge et al., 2012, 2013) with rapid dispersal potential (Quenouille et al., 2011) blurring the geographic history of speciation.
Macroevolution and Macroecology on Tropical Reefs
Albeit incomplete, dated phylogenies combined with biogeographic distributions can detect the initial origins of ancestral reef fish lineages, their extinction and survival with shifting centers of biodiversity, and proliferation within expanding habitat. From these patterns it is possible to identify temporal and spatial variation in rates of speciation, extinction and dispersal and how this variation has resulted in the current biodiversity gradient. Measuring the net rate of diversification and how it varies through time has become an important metric in the integrated study of macroevolution and macroecology (Rabosky, 2009a). Methods to model variation in diversification rates in light of ecological processes has seen dramatic advancement in the last decade (reviewed by Morlon, 2014). In particular, recent interest and debate has grown around whether diversity in clades or assemblages can increase unbounded or if it can be limited by ecological or other factors (Rabosky, 2009a; Morlon et al., 2010). Only a handful of studies have explicitly examined rate variation in reef fish lineages with the comparison of constant and rate variable models of diversification (Rüber and Zardoya, 2005; Alfaro et al., 2007, 2009; Cowman and Bellwood, 2013a; Litsios et al., 2014; Lobato et al., 2014), but none have considered the effects of ecological or other factors in limiting biodiversity among tropical regions. If limiting factors do govern the capacity for biodiversity in clades and communities, variation in tropical reef fish biodiversity may have little to do with rates of speciation or extinction and more to do with the capacity of regions to support biodiversity. Rather, clades or communities have experienced different phases in their rate of diversification where they initial radiate and then slowdown as a limit is approached (Rabosky, 2009a). Variation in where and when clades have radiated would led to the observed patterns in tropical biodiversity. If clades have varied in the timing of their radiating phase among geographic regions this might manifest itself as differences in the ages of lineage origination among regions. This may be the case for some families where data is available (Figure 3), however these data are still from incompletely sampled phylogenies.
This radiation and subsequent slowdown is also termed “density-dependent” diversification and can resemble a “niche-filling” process. Such a process was recently uncovered in the trophic diversification of several tropical reef fish families (Lobato et al., 2014) where a switch to low quality food items by several lineages resulted in significant diversification. This highlights the potential for ecological opportunity on reefs to shape lineage diversification. However, it is unclear if this potential has manifested as an actual limit on diversification as many reef fish lineages do not display a slowdown in diversification rate toward the present (Cowman and Bellwood, 2011). While there is evidence of speciation rates decaying over time it appears that limits of diversity in several groups have yet to be realized (Morlon et al., 2010).
We have yet to definitively identify within a complete phylogenetic framework how rates of net diversification on tropical reefs have been altered by ecological or biogeographical processes. If such processes have underpinned the radiation of fishes on coral reefs it may change our understanding of the origins of biodiversity and what factors are important in maintaining diversity in the present.
Tropical Biodiversity and Rates of Molecular Evolution
Across several taxonomic groups there is consistent evidence of a link between the rate of molecular evolution and the observed biodiversity of clades (Fontanillas et al., 2007; Lanfear et al., 2010b; Duchene and Bromham, 2013). This pattern is not universal (Goldie et al., 2011) and has yet to be critically evaluated across the Fish Tree of Life. However, a recent study of genomic variation in African cichlids highlights several molecular mechanisms that may be linked to the enigmatic and rapid diversification of the group (Brawand et al., 2014). In the context of biodiversity patterns there is a tangle web among ecological traits, diversification and molecular rate (Dowle et al., 2013). There are a large number of characteristics, ecological and environmental, that can potentially shape the rate at which genes evolve with numerous hypotheses put forward (Bromham, 2011).
When exploring the link between molecular rate and diversity there are three main explanations that have been discussed (Barraclough and Savolainen, 2001). First, there is something about the process of speciation itself that increases the rate of molecular evolution (Venditti and Pagel, 2010). If the rate of speciation is associated with the rate at which populations divide or become isolated, then a reduction in the effective population size could increase the rate of substitution of nearly neutral mutations (Bromham, 2011). Second, the direction of causation could be the opposite where changes at the genomic level drive rates of speciation, and as such directly influence macroevolutionary patterns (Bromham, 2011). Higher mutation rates would result in faster accumulation of incompatibilities among hybrids and hasten the reproductive isolation among populations. Lastly, the association between diversity and molecular rate could be indirect, where a third factor promotes an increase in the rate of molecular evolution and the diversification rate. Methods are available for testing these scenarios (reviewed by Lanfear et al., 2010b) and results tend to show evidence for the rate of mutation influencing diversification (Lancaster, 2010; Lanfear et al., 2010a; Duchene and Bromham, 2013). These hypotheses have yet to be examined in fishes and may provide insight into the underlying mechanics of speciation on coral reefs.
If speciation drives the rate of molecular evolution through population subdivision, higher diversity and molecular rates in reef associated fish lineages could be driven by the fragmentation of habitat and peripheral isolation, both process that have been reported in evolutionary studies of tropical reef fishes (Hodge et al., 2012; Pellissier et al., 2014). If correct, endemic range species should show faster rates of molecular evolution when compared with a widespread sister lineage. Endemic range species and isolated populations within widespread species have displayed increased genetic structure and haplotype diversity than their widespread counterparts (Hobbs et al., 2013). Whether this is a reflection of a fast molecular rate remains to be seen.
If mutation rate drives speciation rate, would this mean that coral reefs provide the molecular fuel for speciation? It has already been demonstrated that coral reefs promote both the diversification of lineages (Alfaro et al., 2007; Cowman and Bellwood, 2011; Sorenson et al., 2014) and morphological diversity (Price et al., 2011, 2013) so it is not unrealistic that they would also speed molecular evolution. But not all lineages found on coral reefs are morphologically diverse, nor are they all biodiverse. If a similar pattern is found in the molecular rates of coral reef fish clades, where only some lineages identify with faster rates, their proliferation may be due to intrinsically higher mutation rates. This higher rate would allow populations that are briefly or partially separated by any number of mechanisms to become reproductively isolated faster. The IAA hotspot may be exceptionally diverse because its complex series of archipelagos and shallow basins provide more opportunity for population separation than elsewhere. A higher mutation rate could also provide more genomic variation for selection to act upon (Bromham, 2011) for adaptation and separation along ecological axis (Schluter and Conte, 2009). Ecological speciation has been documenting on coral reefs (Rocha et al., 2005b), and a link between adaptations to new niches and high diversity (Lobato et al., 2014). For this scenario, there would not be anything particularly special about coral reef association other than it enabling those lineages with higher mutation rates to promote lineage diversification. The IAA being at the center of the diversity gradient would be a consequence of more reef habitat, which has previously been shown as a significant predictor of variation in reef fish diversity (Bellwood and Hughes, 2001). A situation where coral reefs have acted as a medium for the direct influence of molecular rate on diversification is very different from a third scenario where an indirect factor associated with coral reef habitats promotes faster molecular rates, and independently higher diversification. In comparing reef and non-reef habitats, or tropical versus temperate latitudes, there are a number of indirect factors that could promote both molecular rate and diversification (Dowle et al., 2013). However, across the tropical belt it may be difficult to deduce what particular factors mediate the link on coral reefs in the IAA and not on reef in other regions. As with models of diversification, it is likely that when these hypotheses are examined in depth, the processes at play will be more dynamic and possibly include more than one explanation.
Conclusion
In reviewing the current state of phylogenies and historical biogeography of tropical reef fishes I have summarized a series of historical events that have underpinned the origins and proliferation of reef fish biodiversity in the tropics. This review also highlights several groups that require increased sampling and further analysis. While some focal groups are almost completely sampled, an additional push is needed to obtain complete species level sampling. Although the traditional nine coral reef fish families have been important models in the exploration of marine speciation and evolution on coral reefs, there are other fish families and lineages that may provided as much, if not more insight into the origins of tropical biodiversity. It is in this respect that a robust and well-resolved Fish Tree of Life will be beneficial to both the examination and comparison of evolutionary rates among discrete tropical clades found on and off reefs, and the investigation of overarching patterns of tropical diversification. I suggest that future research concerning the macroevolutionary patterns of fishes found on coral reefs examine the historical variation in rates of speciation, extinction, and dispersal among biogeographic regions and across multiple lineages. Further discussion is needed to evaluate how hypotheses concerning the origin and maintenance of biodiversity are modeled to account for the interaction between macroecology, macroevolution and molecular processes.
There are several questions that offer exciting pathways for future research:
- How has temporal and spatial variation in rates of speciation, extinction and dispersal lead to present day patterns of tropic fish biodiversity?
- Do present day biogeographic delineations reflect the evolutionary history of the tropical belt, and which scheme is best?
- Are there limits to the biodiversity of tropical regions and if so, how are these limits linked to the diversity of clades and regional assemblages?
- Do tropical clades experience increase rates of molecular change on coral reefs and how does this link to patterns of biodiversity across the tropical belt?
With increasing access to genomic methods, there is a unique opportunity to reconstruct the evolutionary history of all fishes to the level of resolution that is available in other vertebrate clades. Within this framework we can move beyond categorizing patterns and predictors of extant biodiversity, and statistically examine the evolutionary history under hypotheses driven models of diversification.
Conflict of Interest Statement
The author declares that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Acknowledgments
I would like to thank J. Tanner, D. Bellwood, J. Hodge and members of the Macroevolution and Macroecology group at the Australian National University for helpful discussion, and G. Bernardi and L. Rüber for valuable comments on earlier drafts. I would like to thank F. Santini for access to the Acanthuridae phylogeny; B. Frédérich for access to the Pomacentridae phylogeny; P. Hundt for access to the Blenniidae phylogeny; A. Dornburg for access of the Holocentridae phylogeny; and S. Klanten for access to the parrotfish phylogeny. I thank M Kulbicki and V Parravicini for access to data on species distributions and endemism obtained from the GASPAR program. The GASPAR program is part of the CESAB initiative financed by the Foundation pour la Recherche en Biodiversité (FRB). This review was funded from the Gaylord Donnelley Postdoctoral Environment Fellowship administered by the Yale Institute for Biospheric Studies (YIBS).
Footnotes
- ^http://www.ala.org.au
- ^http://www.iucnredlist.org
- ^http://www.gbif.org
- ^http://www.mol.org
- ^http://www.iobis.org
References
Ackerman, J., and Bellwood, D. (2000). Reef fish assemblages: a re-evaluation using enclosed rotenone stations. Mar. Ecol. Ser. 206, 227–237. doi: 10.3354/meps206227
Alfaro, M. E., Brock, C. D., Banbury, B. L., and Wainwright, P. C. (2009). Does evolutionary innovation in pharyngeal jaws lead to rapid lineage diversification in labrid fishes? BMC Evol. Biol. 14:1–14. doi: 10.1186/1471-2148-9-255
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Alfaro, M. E., Santini, F., and Brock, C. D. (2007). Do reefs drive diversification in marine teleosts? Evidence from the pufferfish and their allies (order Tetraodontiformes). Evolution 61, 2104–2126. doi: 10.1111/j.1558-5646.2007.00182.x
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Allen, G. R. (2014). Review of Indo-Pacific coral reef fish systematics: 1980 to 2014. Ichthyol. Res. 1–7. doi: 10.1007/s10228-014-0411-1
Bannikov, A. F. (2010). Bellwoodilabrus landinii n. gen., n. sp., a new genus and species of labrid fish (Teleostei, Perciformes) from the Eocene of Monte Bolca. Geodiversitas 32, 201–220. doi: 10.5252/g2010n2a2
Barber, P. H., and Bellwood, D. R. (2005). Biodiversity hotspots: evolutionary origins of biodiversity in wrasses (Halichoeres: Labridae) in the Indo-Pacific and new world tropics. Mol. Phylogenet. Evol. 35, 235–253. doi: 10.1016/j.ympev.2004.10.004
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Barraclough, T. G., and Savolainen, V. (2001). Evolutionary rates and species diversity in flowering plants. Evolution 55, 677–683. doi: 10.1554/0014-3820 (2001)055[0677:ERASDI]2.0.CO;2
Bellwood, D. R. (1996). The Eocene fishes of Monte Bolca: the earliest coral reef fish assemblage. Coral Reefs 15, 11–19. doi: 10.1007/BF01626074
Bellwood, D. R. (1998). What are reef fishes? – Comment on the report by D. R. Robertson: do coral-reef fish faunas have a distinctive taxonomic structure? Coral Reefs 17, 187–189. doi: 10.1007/s003380050114
Bellwood, D. R., Goatley, C. H. R., Brandl, S. J., and Bellwood, O. (2014a). Fifty million years of herbivory on coral reefs: fossils, fish and functional innovations. Proc. Biol. Sci. 281, 20133046. doi: 10.1098/rspb.2013.3046
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Bellwood, D. R., Hoey, A. S., Bellwood, O., and Goatley, C. H. R. (2014b). Evolution of long-toothed fishes and the changing nature of fish-benthos interactions on coral reefs. Nat. Commun. 5, 3144. doi: 10.1038/ncomms4144
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Bellwood, D. R., Goatley, C. H. R., Cowman, P. F., and Bellwood, O. (in press). “The evolution of fishes on coral reefs: fossils, phylogenies and functions,” in Ecology of Fishes on Coral Reefs, ed. C. Mora (Cambridge: Cambridge University Press).
Bellwood, D. R., and Hughes, T. P. (2001). Regional-scale assembly rules and biodiversity of coral reefs. Science 292, 1532–1535. doi: 10.1126/science.1058635
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Bellwood, D. R., Klanten, S., Cowman, P. F., Pratchett, M. S., Konow, N., and van Herwerden, L. (2010). Evolutionary history of the butterflyfishes (f: Chaetodontidae) and the rise of coral feeding fishes. J. Evol. Biol. 23, 335–349. doi: 10.1111/j.1420-9101.2009.01904.x
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Bellwood, D. R., and Meyer, C. P. (2009a). Endemism and evolution in the Coral Triangle: a call for clarity. J. Biogeogr. 36, 2010–2012. doi: 10.1111/j.1365-2699.2009.02167.x
Bellwood, D. R., and Meyer, C. P. (2009b). Searching for heat in a marine biodiversity hotspot. J. Biogeogr. 36, 569–576. doi: 10.1111/j.1365-2699.2008.02029.x
Bellwood, D. R., Renema, W., and Rosen, B. R. (2012). “Biodiversity hotspots, evolution and coral reef biogeography,” in Biotic Evolution and Environmental Change in Southeast Asia, eds D. Gower, K. Johnson, J. Richardson, B. Rosen, L. Rüber, and S. Williams (Cambridge: Cambridge University Press), 2–32.
Bellwood, D. R., van Herwerden, L., and Konow, N. (2004). Evolution and biogeography of marine angelfishes (Pisces: Pomacanthidae). Mol. Phylogenet. Evol. 33, 140–155. doi: 10.1016/j.ympev.2004.04.015
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Bellwood, D. R., and Wainwright, P. C. (2002). “The history and biogeography of fishes on coral reefs,” in Coral Reef Fishes: Dynamics and Diversity in a Complex Ecosystem, ed. P. Sale (London: Academic Press), 5–32.
Bernardi, G., Bucciarelli, G., Costagliola, D., Robertson, D. R., and Heiser, J. B. (2004). Evolution of coral reef fish Thalassoma spp. (Labridae). 1. Molecular phylogeny and biogeography. Mar. Biol. 144, 369–375. doi: 10.1007/s00227-003-1199-0
Bernardi, G., Holbrook, S. J., and Schmitt, R. J. (2001). Gene flow at three spatial scales in a coral reef fish, the three-spot dascyllus, Dascyllus trimaculatus. Mar. Biol. 138, 457–465. doi: 10.1007/s002270000484
Betancur-R, R., Broughton, R. E., Wiley, E. O., Carpenter, K., López, J. A., Li, C.,et al. (2013). The tree of life and a new classification of bony fishes. PLoS Curr. 5:1–41 doi: 10.1371/currents.tol.53ba26640df0ccaee75bb165c8c26288
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Bowen, B. W., Muss, A., Rocha, L. A., and Grant, W. S. (2006). Shallow mtDNA coalescence in Atlantic pygmy angelfishes (genus Centropyge) indicates a recent invasion from the Indian Ocean. J. Hered. 97, 1–12. doi: 10.1093/jhered/esj006
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Bowen, B. W., Rocha, L. A., Toonen, R. J., and Karl, S. A. (2013). The origins of tropical marine biodiversity. Trends Ecol. Evol. 28, 359–366. doi: 10.1016/j.tree.2013.01.018
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Brawand, D., Wagner, C. E., Li, Y. I., Malinsky, M., Keller, I., Fan, S.,et al. (2014). The genomic substrate for adaptive radiation in African cichlid fish. Nature 513, 375–381. doi: 10.1038/nature13726
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Briggs, J. C. (1999). Coincident biogeographic patterns: Indo-West Pacific Ocean. Evolution 53, 326–335. doi: 10.2307/2640770
Briggs, J. C. (2009). Diversity, endemism and evolution in the Coral Triangle. J. Biogeogr. 36, 2008–2010. doi: 10.1111/j.1365-2699.2009.02146.x
Briggs, J. C. (2003). Marine centres of origin as evolutionary engines. J. Biogeogr. 30, 1–18. doi: 10.1046/j.1365-2699.2003.00810.x
Briggs, J. C., and Bowen, B. W. (2012). A realignment of marine biogeographic provinces with particular reference to fish distributions. J. Biogeogr. 39, 12–30. doi: 10.1111/j.1365-2699.2011.02613.x
Bromham, L. (2011). The genome as a life-history character: why rate of molecular evolution varies between mammal species. Philos. Trans. R. Soc. Lond. B Biol. Sci. 366, 2503–2513. doi: 10.1098/rstb.2011.0014
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Budd, A. F. (2000). Diversity and extinction in the Cenozoic history of Caribbean reefs. Coral Reefs 19, 25–35. doi: 10.1007/s003380050222
Carnevale, G. (2006). Morphology and biology of the Miocene butterflyfish Chaetodon ficheuri (Teleostei: Chaetodontidae). Zool. J. Linn. Soc. 146, 251–267. doi: 10.1111/j.1096-3642.2006.00203.x
Carpenter, K. E., Barber, P. H., Crandall, E. D., Ablan-lagman, M. C. A., Mahardika, G. N., Manjaji-Matsumoto, B. M.,et al. (2011). Comparative phylogeography of the coral triangle and implications for marine management. J. Mar. Biol. 2011, 1–14. doi: 10.1155/2011/396982
Choat, J. H., Klanten, O. Y. A. S., Van Herwerden, L., Robertson, D. R., Clements, K. D., and van Herwerden, L. (2012). Patterns and processes in the evolutionary history of parrotfishes (Family Labridae). Biol. J. Linn. Soc. 107, 529–557. doi: 10.1111/j.1095-8312.2012.01959.x
Coates, A. G., and Obando, J. A. (1996) “The geologic evolution of the Central American Isthmus,” in Evolution and Environment in Tropical America, eds J. B. C. Jackson, A. F. Budd, and A. G. Coates (Chicago, IL: University of Chicago Press), 21–56.
Connolly, S. R., Bellwood, D. R., and Hughes, T. P. (2003). Indo-Pacific biodiversity of coral reefs: deviations from a mid-domain model. Ecology 84, 2178–2190. doi: 10.1890/02-0254
Cooper, W. J., Smith, L. L., and Westneat, M. W. (2009). Exploring the radiation of a diverse reef fish family: phylogenetics of the damselfishes (Pomacentridae), with new classifications based on molecular analyses of all genera. Mol. Phylogenet. Evol. 52, 1–16. doi: 10.1016/j.ympev.2008.12.010
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Cowman, P. F., and Bellwood, D. R. (2011). Coral reefs as drivers of cladogenesis: expanding coral reefs, cryptic extinction events, and the development of biodiversity hotspots. J. Evol. Biol. 24, 2543–2562. doi: 10.1111/j.1420-9101.2011.02391.x
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Cowman, P. F., and Bellwood, D. R. (2013a). The historical biogeography of coral reef fishes: global patterns of origination and dispersal. J. Biogeogr. 40, 209–224. doi: 10.1111/jbi.12003
Cowman, P. F., and Bellwood, D. R. (2013b). Vicariance across major marine biogeographic barriers: temporal concordance and the relative intensity of hard versus soft barriers. Proc. Biol. Sci. 280, 20131541. doi: 10.1098/rspb.2013.1541
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Cowman, P. F., Bellwood, D. R., and van Herwerden, L. (2009). Dating the evolutionary origins of wrasse lineages (Labridae) and the rise of trophic novelty on coral reefs. Mol. Phylogenet. Evol. 52, 621–31. doi: 10.1016/j.ympev.2009.05.015
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
D’agata, S., Mouillot, D., Kulbicki, M., Andréfouët, S., Bellwood, D. R., Cinner, J. E.,et al. (2014). Human-mediated loss of phylogenetic and functional diversity in coral reef fishes. Curr. Biol. 24, 555–560. doi: 10.1016/j.cub.2014.01.049
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Dornburg, A., Moore, J., Beaulieu, M., Eytan, R., and Near, T. J. (in press). The impact of shifts in marine biodiversity hotspots on patterns of range evolution: evidence from the Holocentridae (squirrelfishes and soldierfishes). Evolution doi: 10.1016/j.cub.2014.01.049
Dowle, E. J.,Morgan-Richards, M., and Trewick, S. A. (2013). Molecular evolution and the latitudinal biodiversity gradient. Heredity (Edinb.) 110, 501–510. doi: 10.1038/hdy.2013.4
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Duchene, D., and Bromham, L. (2013). Rates of molecular evolution and diversification in plants: chloroplast substitution rates correlate with species-richness in the Proteaceae. BMC Evol. Biol. 13:65. doi: 10.1186/1471-2148-13-65
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Duchene, D., Klanten, S. O., Munday, P. L., Herler, J., and Van Herwerden, L. H. (2013). Phylogenetic evidence for recent diversification of obligate coral-dwelling gobies compared with their host corals. Mol. Phylogenet. Evol. 69, 123–132. doi: 10.1016/j.ympev.2013.04.033
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Ekman, S. (1953). Zoogeography of the Sea. London: Sigwick and Jackson. doi: 10.1186/1471-2148-13-65
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Faircloth, B. C., Sorenson, L., Santini, F., and Alfaro, M. E. (2013). A phylogenomic perspective on the radiation of ray-finned fishes based upon targeted sequencing of ultraconserved elements (UCEs). PLoS ONE 8:e65923. doi: 10.1371/journal.pone.0065923
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Fessler, J. L., and Westneat, M. W. (2007). Molecular phylogenetics of the butterflyfishes (Chaetodontidae): taxonomy and biogeography of a global coral reef fish family. Mol. Phylogenet. Evol. 45, 50–68. doi: 10.1016/j.ympev.2007.05.018
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Floeter, S. R., Guimaraes, R. Z. P., Rocha, L. A., Ferreira, C. E. L., Rangel, C. A., and Gasparini, J. L. (2001). Geographic variation in reef-fish assemblages along the Brazilian coast. Glob. Ecol. Biogeogr. 10, 423–431. doi: 10.1046/j.1466-822X.2001.00245.x
Floeter, S. R., Rocha, L. A., Robertson, D. R., Joyeux, J. C., Smith-Vaniz, W. F., Wirtz, P.,et al. (2008). Atlantic reef fish biogeography and evolution. J. Biogeogr. 35, 22–47. doi: 10.1111/j.1365-2699.2007.01790.x
Fontanillas, E., Welch, J. J., Thomas, J. A., and Bromham, L. (2007). The influence of body size and net diversification rate on molecular evolution during the radiation of animal phyla. BMC Evol. Biol. 7:95. doi: 10.1186/1471-2148-7-95
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Frédérich, B., Sorenson, L., Santini, F., Slater, G. J., and Alfaro, M. E. (2013). Iterative ecological radiation and convergence during the evolutionary history of damselfishes (Pomacentridae). Am. Nat. 181, 94–113. doi: 10.1086/668599
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Friedman, M. (2010). Explosive morphological diversification of spiny-finned teleost fishes in the aftermath of the end-Cretaceous extinction. Proc. Biol. Sci. 277, 1675–1683. doi: 10.1098/rspb.2009.2177
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Gaither, M. R., and Rocha, L. A. (2013). Origins of species richness in the Indo-Malay-Philippine biodiversity hotspot: evidence for the centre of overlap hypothesis. J. Biogeogr. 40, 1638–1648. doi: 10.1111/jbi.12126
Goatley, C. H. R., Bellwood, D. R., and Bellwood, O. (2010). Fishes on coral reefs: changing roles over the past 240 million years. Paleobiology 36, 415–427. doi: 10.1666/09035.1
Goldie, X., Lanfear, R., and Bromham, L. (2011). Diversification and the rate of molecular evolution: no evidence of a link in mammals. BMC Evol. Biol. 11:286. doi: 10.1186/1471-2148-11-286
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Halas, D., and Winterbottom, R. (2009). A phylogenetic test of multiple proposals for the origins of the East Indies coral reef biota. J. Biogeogr. 36, 1847–1860. doi: 10.1111/j.1365-2699.2009.02103.x
Hallam, A. (1994). An Outline of Phanerozoic Biogeography. Oxford: Oxford University Press. doi: 10.1111/j.1365-2699.2009.02103.x
Herler, J., Munday, P., and Hernaman, V. (2011). “Gobies on coral reefs,” in The Biology of Gobies, eds R. A. Patzner,J. L. Van Tassell,M. Kovačić, and B. G. Kapoor (New Hampshire: Science Publishers),493–529.
Hillebrand, H. (2004). On the generality of the latitudinal diversity gradient. Am. Nat. 163, 192–211. doi: 10.1086/381004
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Hobbs, J.-P., van Herwerden, L., Jerry, D., Jones, G., and Munday, P. (2013). High genetic diversity in geographically remote populations of endemic and widespread coral reef angelfishes (genus: Centropyge). Diversity 5, 39–50. doi: 10.3390/d5010039
Hodge, J. R., Read, C. I., Bellwood, D. R., and van Herwerden, L. (2013). Evolution of sympatric species: a case study of the coral reef fish genus Pomacanthus (Pomacanthidae). J. Biogeogr. 62, 653–663. doi: 10.1111/jbi.12124
Hodge, J. R., Read, C. I., van Herwerden, L., and Bellwood, D. R. (2012). The role of peripheral endemism in species diversification: evidence from the coral reef fish genus Anampses (Family: Labridae). Mol. Phylogenet. Evol. 62, 653–663. doi: 10.1016/j.ympev.2011.11.007
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Hodge, J. R., van Herwerden, L., and Bellwood, D. R. (2014). Temporal evolution of coral reef fishes: global patterns and disparity in isolated locations. J. Biogeogr. 41, 2115–2127. doi: 10.1111/jbi.12356
Hoeksema, B. (2007). “Delineation of the Indo-Malayan centre of maximum marine biodiversity: the coral triangle,” in Biogeography, Time, and Place: Distributions, Barriers, and Islands, ed. W. Renema (Dordrecht: Springer),117–178. doi: 10.1111/jbi.12356
Hoeksema, B. (2009). West-east variation in the Indonesian reef coral fauna: lines of division or zones of transition? in Proceedings of World Ocean Conference, Manado, 1–10.
Hubert, N.,Meyer, C. P., Bruggemann, H. J., Guérin, F., Komeno, R. J. L., Espiau, B.,et al. (2012). Cryptic diversity in Indo-Pacific coral-reef fishes revealed by DNA-barcoding provides new support to the centre-of-overlap hypothesis. PLoS ONE 7:e28987. doi: 10.1371/journal.pone.0028987
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Hughes, T. P., Baird, A. H., Bellwood, D. R., Card, M., Connolly, S. R., Folke, C.,et al. (2003). Climate change,human impacts,and the resilience of coral reefs. Science 301, 929–933. doi: 10.1126/science.1085046
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Hughes, T. P., Bellwood, D. R., Baird, A. H., Brodie, J., Bruno, J. F., and Pandolfi, J. M.et al.(2011). Shifting base-lines,declining coral cover,and the erosion of reef resilience: comment on Sweatman et al (2011). Coral Reefs 30, 653–660. doi: 10.1007/s00338-011-0787-6
Hughes, T. P., Bellwood, D. R., and Connolly, S. R. (2002). Biodiversity hotspots, centres of endemicity, and the conservation of coral reefs. Ecol. Lett. 5, 775–784. doi: 10.1046/j.1461-0248.2002.00383.x
Hundt, P. J., Iglésias, S. P., Hoey, A. S., and Simons, A. M. (2014). A multilocus molecular phylogeny of combtooth blennies (Percomorpha: Blennioidei: Blenniidae): multiple invasions of intertidal habitats. Mol. Phylogenet. Evol. 70, 47–56. doi: 10.1016/j.ympev.2013.09.001
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Jetz, W., Thomas, G. H., Joy, J. B., Hartmann, K., and Mooers, A. O. (2012). The global diversity of birds in space and time. Nature 491, 444–448. doi: 10.1038/nature11631
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Joyeux, J.-C., Floeter, S. R., Ferreira, C. E. L., and Gasparini, J. L. (2001). Biogeography of tropical reef fishes: the South Atlantic puzzle. J. Biogeogr. 28, 831–841. doi: 10.1046/j.1365-2699.2001.00602.x
Kazancioglu, E., Near, T. J., Hanel, R., and Wainwright, P. C. (2009). Influence of sexual selection and feeding functional morphology on diversification rate of parrotfishes (Scaridae). Proc. Biol. Sci. 276, 3439–3446. doi: 10.1098/rspb.2009.0876
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Klanten, S. O., van Herwerden, L., Choat, J. H., and Blair, D. (2004). Patterns of lineage diversification in the genus Naso (Acanthuridae). Mol. Phylogenet. Evol. 32, 221–235. doi: 10.1016/j.ympev.2003.11.008
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Kool, J. T., Paris, C. B., Barber, P. H., and Cowen, R. K. (2011). Connectivity and the development of population genetic structure in Indo-West Pacific coral reef communities. Glob. Ecol. Biogeogr. 20, 695–706. doi: 10.1111/j.1466-8238.2010.00637.x
Kulbicki, M., Parravicini, V., Bellwood, D. R., Arias-Gonzàlez, E., Chabanet, P., Floeter, S. R.,et al. (2013). Global biogeography of reef fishes: a hierarchical quantitative delineation of regions. PLoS ONE 8:e81847. doi: 10.1371/journal.pone.0081847
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Lacson, J., and Nelson, S. (1993). Genetic distances among fishes of the genus Siganus (Siganidae) from the Western Pacific Ocean. Mar. Biol. 116, 187–192.
Lancaster, L. T. (2010). Molecular evolutionary rates predict both extinction and speciation in temperate angiosperm lineages. BMC Evol. Biol. 10:162. doi: 10.1186/1471-2148-10-162
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Lanfear, R., Welch, J. J., and Bromham, L. (2010a). Watching the clock: studying variation in rates of molecular evolution between species. Trends Ecol. Evol. 25, 495–503. doi: 10.1016/j.tree.2010.06.007
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Lanfear, R., Ho, S. Y. W., Love, D., and Bromham, L. (2010b). Mutation rate is linked to diversification in birds. Proc. Natl. Acad. Sci. U.S.A. 107, 20423–20428. doi: 10.1073/pnas.1007888107
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Leprieur, F., Albouy, C., De Bortoli, J., Cowman, P. F., Bellwood, D. R., and Mouillot, D. (2012). Quantifying phylogenetic beta diversity: distinguishing between “True” turnover of lineages and phylogenetic diversity gradients. PLoS ONE 7:e42760. doi: 10.1371/journal.pone.0042760
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Lessios, H. A., and Robertson, D. R. (2006). Crossing the impassable: genetic connections in 20 reef fishes across the eastern Pacific barrier. Proc. Biol. Sci. 273, 2201–2208. doi: 10.1098/rspb.2006.3543
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Litsios, G., Pearman, P. B., Lanterbecq, D., Tolou, N., and Salamin, N. (2014). The radiation of the clownfishes has two geographical replicates. J. Biogeogr. 41, 2140–2149. doi: 10.1111/jbi.12370
Litsios, G., Sims, C. A., Wüest, R. O., Pearman, P. B., Zimmermann, N. E., and Salamin, N. (2012). Mutualism with sea anemones triggered the adaptive radiation of clownfishes. BMC Evol. Biol. 12:212. doi: 10.1186/1471-2148-12-212
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Lobato, F. L., Barneche, D. R., Siqueira, A. C., Liedke, A. M. R., Lindner, A., Pie, M. R.,et al. (2014). Diet and diversification in the evolution of coral reef fishes. PLoS ONE 9:e102094. doi: 10.1371/journal.pone.0102094
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
McCoy, E. D., and Heck, K. L. (1976). Biogeography of corals, seagrasses, and mangroves: an alternative to the center of origin concept. Syst. Zool. 25, 201. doi: 10.2307/2412488
McMillan, W. O., and Palumbi, S. R. (1995). Pacific butterflyfishes concordant evolutionary patterns among Indo-West. Proc. R. Soc. London 260, 229–236. doi: 10.2307/2412488
McMillan, W. O., Weigt, L. A., and Palumbi, S. R. (1999). Color pattern evolution, assortative mating, and genetic differentiation in brightly colored butterflyfishes. Evolution 53, 247–260. doi: 10.1098/rspb.1995.0085
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Micklich, N. R., Tyler, J. C., Johnson, G. D., Świdnicka, E., and Bannikov, A. F. (2009). First fossil records of the tholichthys larval stage of butterfly fishes (Perciformes, Chaetodontidae), from the Oligocene of Europe. Paläontol. Z. 83, 479–497. doi: 10.1007/s12542-009-0031-7
Montes, C., Cardona, A., McFadden, R., Moron, S. E., Silva, C. A., Restrepo-Moreno, S.,et al. (2012). Evidence for middle Eocene and younger land emergence in central Panama: implications for Isthmus closure. Geol. Soc. Am. Bull. 124, 780–799. doi: 10.1130/B30528.1
Mora, C., Chittaro, P. M., Sale, P. F., Kritzer, J. P., and Ludsin, S. A. (2003). Patterns and processes in reef fish diversity. Nature 421, 933–936. doi: 10.1038/nature01421.1
Mora, C., Tittensor, D. P., and Myers, R. A. (2008). The completeness of taxonomic inventories for describing the global diversity and distribution of marine fishes. Proc. Biol. Sci. 275, 149–55. doi: 10.1098/rspb.2007.1315
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Morlon, H. (2014). Phylogenetic approaches for studying diversification. Ecol. Lett. 17, 508–25. doi: 10.1111/ele.12251
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Morlon, H., Potts, M. D., and Plotkin, J. B. (2010). Inferring the dynamics of diversification: a coalescent approach. PLoS Biol. 8:e1000493. doi: 10.1371/journal.pbio.1000493
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Mouillot, D., De Bortoli, J., Leprieur, F., Parravicini, V., Kulbicki, M., and Bellwood, D. R. (2013). The challenge of delineating biogeographical regions: nestedness matters for Indo-Pacific coral reef fishes. J. Biogeogr. 40, 2228–2237. doi: 10.1111/jbi.12194
Myers, N. (1988). Threatened biotas: “hot spots” in tropical forests. Environmentalist 8, 187–208. doi: 10.1111/jbi.12194
Myers, N., Mittermeier, R. A., Mittermeier, C. G., da Fonseca, G. A., and Kent, J. (2000). Biodiversity hotspots for conservation priorities. Nature 403, 853–858. doi: 10.1038/35002501
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Near, T. J., Dornburg, A., Eytan, R. I., Keck, B. P., Smith, W. L., Kuhn, K. L.,et al. (2013). Phylogeny and tempo of diversification in the superradiation of spiny-rayed fishes. Proc. Natl. Acad. Sci. U.S.A. 110, 12738–1243. doi: 10.1073/pnas.1304661110
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Near, T. J., Eytan, R. I., Dornburg, A., Kuhn, K. L., Moore, J. A, Davis, M. P.,et al. (2012). Resolution of ray-finned fish phylogeny and timing of diversification. Proc. Natl. Acad. Sci. U.S.A. 109, 13698–13703. doi: 10.1073/pnas.1206625109
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Nelson, G. (1989). “Phylogeny of major fish groups,” in The Hierarchy of Life, eds B. Fernholm, K. Bremer, and H. Jörnvall (Amsterdam: Excerpta Medica), 325–336.
O’Dea, A., Jackson, J. B. C., Fortunato, H., Smith, J. T., D’Croz, L., Johnson, K. G.,et al. (2007). Environmental change preceded Caribbean extinction by 2 million years. Proc. Natl. Acad. Sci. U.S.A. 104, 5501–5506. doi: 10.1073/pnas.0610947104
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Olson, D. M., Dinerstein, E., Wikramanayake, E. D., Burgess, N. D., Powell, G. V. N., Underwood, E. C.,et al. (2001). Terrestrial ecoregions of the world: a new map of life on earth. Bioscience 51, 933–938. doi: 10.1073/pnas.0610947104
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Palumbi, S. R. (1997). Molecular biogeography of the Pacific. Coral Reefs 16, S47–S52. doi: 10.1007/s003380050241
Parravicini, V., Kulbicki, M., Bellwood, D. R., Friedlander, A. M., Arias-Gonzalez, J. E., Chabanet, P.,et al. (2013). Global patterns and predictors of tropical reef fish species richness. Ecography (Cop.) 36, 1254–1262. doi: 10.1111/j.1600-0587.2013.00291.x
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Pellissier, L., Leprieur, F., Parravicini, V., Cowman, P. F., Kulbicki, M., Litsios, G.,et al. (2014). Quaternary coral reef refugia preserved fish diversity. Science 344, 1016–1019. doi: 10.1126/science.1249853
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Potts, D. C. (1985). “Sea level fluctuations and speciation in Scleractina,” in Proceedings of the Fifth International Coral Reef Congress, Tahiti.
Price, S. A., Holzman, R., Near, T. J., and Wainwright, P. C. (2011). Coral reefs promote the evolution of morphological diversity and ecological novelty in labrid fishes. Ecol. Lett. 14, 462–469. doi: 10.1111/j.1461-0248.2011.01607.x
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Price, S. A., Schmitz, L., Oufiero, C. E., Eytan, R. I., Dornburg, A., Smith, W. L.,et al. (2014). Two waves of colonization straddling the K-Pg boundary formed the modern reef fish fauna. Proc. Biol. Sci. 281, 20140321. doi: 10.1098/rspb.2014.0321
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Price, S. A., Tavera, J. J., Near, T. J., and Wainwright, P. C. (2013). Elevated rates of morphological and functional diversification in reef-dwelling haemulid fishes. Evolution 67, 417–428. doi: 10.1111/j.1558-5646.2012.01773.x
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Quenouille, B., Hubert, N., Bermingham, E., and Planes, S. (2011). Speciation in tropical seas: allopatry followed by range change. Mol. Phylogenet. Evol. 58, 546–552. doi: 10.1016/j.ympev.2010.12.009
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Quental, T. B., and Marshall, C. R. (2009). Extinction during evolutionary radiations: reconciling the fossil record with molecular phylogenies. Evolution 63, 3158–3167. doi: 10.1111/j.1558-5646.2009.00794.x
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Quental, T. B., and Marshall, C. R. (2010). Diversity dynamics: molecular phylogenies need the fossil record. Trends Ecol. Evol. 25, 434–441. doi: 10.1016/j.tree.2010.05.002
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Rabosky, D. L. (2009a). Ecological limits and diversification rate: alternative paradigms to explain the variation in species richness among clades and regions. Ecol. Lett. 12, 735–743. doi: 10.1111/j.1461-0248.2009.01333.x
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Rabosky, D. L. (2009b). Extinction rates should not be estimated from molecular phylogenies. Evolution 4, 1816–1824. doi: 10.1111/j.1558-5646.2009.00926.x
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Rabosky, D. L., Santini, F., Eastman, J., Smith, S. A., Sidlauskas, B., Chang, J.,et al. (2013). Rates of speciation and morphological evolution are correlated across the largest vertebrate radiation. Nat. Commun. 4, 1–8. doi: 10.1038/ncomms2958
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Read, C. I., Bellwood, D. R., and van Herwerden, L. (2006). Ancient origins of Indo-Pacific coral reef fish biodiversity: a case study of the leopard wrasses (Labridae: Macropharyngodon). Mol. Phylogenet. Evol. 38, 808–819. doi: 10.1016/j.ympev.2005.08.001
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Ree, R. H., and Sanmartín, I. (2009). Prospects and challenges for parametric models in historical biogeographical inference. J. Biogeogr. 36, 1211–1220. doi: 10.1111/j.1365-2699.2008.02068.x
Reed, D. L., Carpenter, K. E., and deGravelle, M. J. (2002). Molecular systematics of the Jacks (Perciformes: Carangidae) based on mitochondrial cytochrome b sequences using parsimony, likelihood, and Bayesian approaches. Mol. Phylogenet. Evol. 23, 513–524.
Renema, W., Bellwood, D. R., Braga, J. C., Bromfield, K., Hall, R., Johnson, K. G.,et al. (2008). Hopping hotspots: global shifts in marine biodiversity. Science 321, 654–657. doi: 10.1126/science.1155674
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Roberts, C. M., McClean, C. J., Veron, J. E. N., Hawkins, J. P., Allen, G. R., McAllister, D. E.,et al. (2008). Marine biodiversity hotspots and conservation priorities for tropical reefs. Science 295, 1280–1284. doi: 10.1126/science.1067728
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Robertson, D. R. (1998). Do coral-reef fish faunas have a distinctive taxonomic structure? Coral Reefs 17, 179–186. doi: 10.1007/s003380050113
Rocha, L. A., Robertson, D. R., Rocha, C. R., Van Tassell, J. L., Craig, M. T., Bowen, B. W.,et al. (2005a). Recent invasion of the tropical Atlantic by an Indo-Pacific coral reef fish. Mol. Ecol. 14, 3921–3928. doi: 10.1111/j.1365-294X.2005.02698.x
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Rocha, L. A., Robertson, D. R., Roman, J., and Bowen, B. W. (2005b). Ecological speciation in tropical reef fishes. Proc. Biol. Sci. 272, 573–579. doi: 10.1098/2004.3005
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Rocha, L. A., and Bowen, B. W. (2008). Speciation in coral-reef fishes. J. Fish. Biol. 72, 1101–1121. doi: 10.1111/j.1095-8649.2007.01770.x
Rolland, J., Condamine, F. L., Jiguet, F., and Morlon, H. (2014). Faster speciation and reduced extinction in the tropics contribute to the mammalian latitudinal diversity gradient. PLoS Biol. 12:e1001775. doi: 10.1371/journal.pbio.1001775
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Rosen, B. R. (1984). “Reef coral biogeography and climate through the late Cainozoic: just islands in the sun or a critical pattern of islands?” in Fossil and Climate, ed. P. Brenchley (Chichester: John Wiley and Sons), 201–262.
Rüber, L., Tassell, J., and Zardoya, R. (2003). Rapid speciation and ecological divergence in the American seven-spined gobies (Gobiidae, Gobiosomatini) inferred from a molecular phylogeny. Evolution 57, 1584–1598. doi: 10.1111/j.0014-3820.2003.tb00366.x
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Rüber, L., and Zardoya, R. (2005). Rapid cladogenesis in marine fishes revisited. Evolution 59, 1119–1127. doi: 10.1111/j.0014-3820.2005.tb01048.x
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Sanderson, M. (2003). r8s: inferring absolute rates of molecular evolution and divergence times in the absence of a molecular clock. Bioinformatics 19, 301–302.
Schluter, D., and Conte, G. L. (2009). Genetics and ecological speciation. Proc. Natl. Acad. Sci. U.S.A. 9955–9962. doi: 10.1073/pnas.0901264106
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Smith, L. L., Fessler, J. L., Alfaro, M. E., Streelman, J. T., and Westneat, M. W. (2008). Phylogenetic relationships and the evolution of regulatory gene sequences in the parrotfishes. Mol. Phylogenet. Evol. 49, 136–152. doi: 10.1016/j.ympev.2008.06.008
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Sorenson, L., Santini, F., and Alfaro, M. E. (2014). The effect of habitat on modern shark diversification. J. Evol. Biol. 27, 1536–1548. doi: 10.1111/jeb.12405
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Sorenson, L., Santini, F., Carnevale, G., and Alfaro, M. E. (2013). A multi-locus timetree of surgeonfishes (Acanthuridae, Percomorpha), with revised family taxonomy. Mol. Phylogenet. Evol. 68, 150–160. doi: 10.1016/j.ympev.2013.03.014
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Spalding, M. D., Fox, H. E., Allen, G. R., Davidson, N., Ferdaña, Z. A., Finlayson, M.,et al. (2007). Marine ecoregions of the world: a bioregionalization of coastal and shelf areas. Bioscience 57, 573. doi: 10.1641/B570707
Steininger, F. F., and Rögl, F. (1979). The paratethys history. A contribution towards the Neogene geodynamics of the alpine orogene. Ann. Geol. Pays Hell. 3, 1153–1165. doi: 10.1641/B570707
Taylor, M. S., and Hellberg, M. E. (2005). Marine radiations at small geographic scales: speciation in neotropical reef gobies (Elacatinus). Evolution 59, 374–385. doi: 10.1554/04-590
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Thacker, C. E., and Roje, D. M. (2009). Phylogeny of cardinalfishes (Teleostei: Gobiiformes: Apogonidae) and the evolution of visceral bioluminescence. Mol. Phylogenet. Evol. 52, 735–745. doi: 10.1016/j.ympev.2009.05.017
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Thomson, R. C., and Shaffer, H. B. (2010a). Rapid progress on the vertebrate tree of life. BMC Biol. 8:19. doi: 10.1186/1741-7007-8-19
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Thomson, R. C., and Shaffer, H. B. (2010b). Sparse supermatrices for phylogenetic inference: taxonomy, alignment, rogue taxa, and the phylogeny of living turtles. Syst. Biol. 59, 42–58. doi: 10.1093/sysbio/syp075
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Timm, J., and Kochzius, M. (2008). Geological history and oceanography of the Indo-Malay Archipelago shape the genetic population structure in the false clown anemonefish (Amphiprion ocellaris). Mol. Ecol. 17, 3999–4014. doi: 10.1111/j.1365-294X.2008.03881.x
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Tittensor, D. P., Mora, C., Jetz, W., Lotze, H. K., Ricard, D., Berghe, E. V.et al. (2010). Global patterns and predictors of marine biodiversity across taxa. Nature 466, 1098–1101. doi: 10.1038/nature09329
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Venditti, C., and Pagel, M. (2010). Speciation as an active force in promoting genetic evolution. Trends Ecol. Evol. 25, 14–20. doi: 10.1016/j.tree.2009.06.010
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Wainwright, P. C., Smith, W. L., Price, S. A., Tang, K. L., Sparks, J. S., Ferry, L. A.,et al. (2012). The evolution of pharyngognathy: a phylogenetic and functional appraisal of the pharyngeal jaw key innovation in labroid fishes and beyond. Syst. Biol. 61, 1001–1027. doi: 10.1093/sysbio/sys060
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Wallace, C. C., and Rosen, B. R. (2006). Diverse staghorn corals (Acropora) in high-latitude Eocene assemblages: implications for the evolution of modern diversity patterns of reef corals. Proc. Biol. Sci. 273, 975–982. doi:10.1098/rspb.2005.3307
Westneat, M. W., and Alfaro, M. E. (2005). Phylogenetic relationships and evolutionary history of the reef fish family Labridae. Mol. Phylogenet. Evol. 36, 370–390. doi: 10.1016/j.ympev.2005.02.001
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Whiting, A. S., Lawler, S. H., Horwitz, P., and Crandall, K. A. (2000). Biogeographic regionalization of Australia: assigning conservation priorities based on endemic freshwater crayfish phylogenetics. Anim. Conserv. 3, 155–163. doi: 10.1016/j.ympev.2005.02.001
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Whittaker, R. J., Araujo, M. B., Paul, J., Ladle, R. J., Watson, J. E. M., and Willis, K. J. (2005). Conservation biogeography: assessment and prospect. Divers. Distrib. 11, 3–23. doi: 10.1111/j.1366-9516.2005.00143.x
Williams, S. T., and Duda, T. F. (2008). Did tectonic activity stimulate oligomiocene speciation in the Indo-West Pacific. Evolution 62, 1618–1634. doi: 10.1111/j.1558-5646.2008.00399.x
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Willig, M. R., Kaufman, D. M., and Stevens, R. D. (2003). Latitudinal gradients of biodiversity: pattern, process, scale, and synthesis. Annu. Rev. Ecol. Evol. 34, 273–309. doi: 10.1146/annurev.ecolsys.34.012103.44032
Wilson, M. E. J. and Rosen, B. R. (1998). “Implications of paucity of corals in the Paleogene of SE Asia: plate tectonics or centre of origin?,” in Biogeography and Geological Evolution of SE Asia, eds R. Hall and J. D. Holloway (Leiden: Backhuys Publishers), 165–195.
Keywords: coral reef fishes, ancestral biogeography, marine tropics, phylogeny, diversification
Citation: Cowman PF (2014) Historical factors that have shaped the evolution of tropical reef fishes: a review of phylogenies, biogeography, and remaining questions. Front. Genet. 5:394. doi: 10.3389/fgene.2014.00394
Received: 01 August 2014; Accepted: 27 October 2014;
Published online: 13 November 2014.
Edited by:
James Edward Richardson, Royal Botanic Garden Edinburgh, UKReviewed by:
Giacomo Bernardi, University of California, Santa Cruz, USALukas Rüber, Naturhistorisches Museum der Burgergemeinde Bern, Switzerland
Copyright © 2014 Cowman. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) or licensor are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Peter F. Cowman, Department of Ecology and Evolutionary Biology, Yale University, 21 Sachem Street, New Haven, CT 06511, USA e-mail:cGV0ZXIuY293bWFuQHlhbGUuZWR1