 Devon O’Rourke
Devon O’Rourke Cody E. FitzGerald
Cody E. FitzGerald Charles C. Traverse
Charles C. Traverse Vaughn S. Cooper
Vaughn S. Cooper- 1Molecular, Cellular, and Biomedical Sciences, University of New Hampshire, Durham, NH, USA
- 2Department of Microbiology, University of Texas at Austin, Austin, TX, USA
Experimental evolution paired with modern sequencing can be a powerful approach to identify the mechanisms by which bacteria adapt to discrete environmental conditions found in nature or during infections. We used this approach to identify mechanisms enabling biofilm specialists of the opportunistic respiratory pathogen Burkholderia cenocepacia to regain planktonic fitness. Seven mutants producing wrinkly (W) small-colony variants by mutations in the wrinkly-spreader operon (wsp) cluster, but with varying duration of biofilm adaptation, served as ancestors of this experiment. Following planktonic growth, each W ancestor produced smooth (S) mutants with distinct fitness effects across planktonic, biofilm, and dispersal-phase environments. The causes of the S phenotype traced to mutations in three gene clusters: wsp, Bcen2424_1436, an uncharacterized two-component transcriptional regulator which appears to be critical for wsp signaling, and a cohort of genes involved in polysaccharide synthesis. The genetic pathway from W to S also associated with evolutionary history in the biofilm environment. W mutants isolated from long-term biofilm selection usually produced S types via secondary wsp mutations, whereas S types evolved from less adapted W ancestors by a wider scope of mutations. These different genetic pathways to suppress the W phenotype suggest that prolonged biofilm adaptation limits routes to subsequent planktonic adaptation, despite common initial mechanisms of biofilm adaptation. More generally, experimental evolution can be used as a nuanced screen for gain-of-function mutations in multiple conditions that illustrate tensions that bacteria may face in changing environments or hosts.
Introduction
Two bedrock techniques of genetic analysis are the mutant screen for a particular novel phenotype and the suppressor screen for mutants reverting in phenotype. Commonly these methods have involved transposon or chemical mutagenesis, which may produce severe and even unrealistic mutations that hamper or eliminate the possibility of reversion. Today, combining experimental evolution with modern sequencing can accomplish these same goals with potentially greater realism and sample a broader spectrum of genetic variation (Elena and Lenski, 2003; Kawecki et al., 2012; Barrick and Lenski, 2013). Moreover, the kinds of mutations captured by experimental evolution can produce insights into the function of the altered transcript. For instance, gain-of-function mutations in hypothetical open reading frames can chart a short path to identifying the function of an undescribed gene, and the location of mutations within a known gene can demonstrate the relative importance of different domains for a particular function. Put another way, the evolution of several non-synonymous mutations in an unstudied genome region that are associated with adaptation can illustrate more about the relevant environmental pressures on bacterial physiology than the effects of a loss-of-function (LOF) gene knockout. One added benefit of experimental evolution is that the trajectories of contending mutations can explain the relative strength of selection for that phenotype (Cooper, 2007; Lang et al., 2013; Traverse et al., 2013), although this study focuses on clones representing certain contending mutants.
One environmental transition that likely generates significant selection for many bacteria is the switch from surface attachment to dispersal in a planktonic environment. Life on surfaces, or in biofilms, may dominate the lifestyle of some species in certain environments, but appear less critical in other environments that enable more rapid growth in mass-action conditions. The means by which bacteria adapt to changing conditions that favor either biofilm formation or dispersal underlie a variety of important processes, such as the development of antimicrobial resistance (Jeannot et al., 2008; Guo and Rowe-Magnus, 2010; Sawasdidoln et al., 2010; Lee et al., 2013), the construction of durable microbial communities, and colonization of new environments or hosts (Danhorn and Fuqua, 2007; Lee et al., 2013). Although many mechanisms involved in biofilm formation have been defined for a range of species (Abee et al., 2011; Fazli et al., 2014), research describing the genetic and physiological paths regulating dispersal is less comprehensive, as reviewed by (Kaplan, 2010; McDougald et al., 2012).
Recently, we explored the genetic causes of Wrinkly (W) small colony variants (SCVs) in the biofilm-forming species, Burkholderia cenocepacia, using experimental evolution as a screen for these adaptive phenotypes in a laboratory model of biofilm selection (Poltak and Cooper, 2011; Traverse et al., 2013). Burkholderia SCVs are associated with worse patient prognoses during pulmonary infections of persons with cystic fibrosis (CF) (Haussler et al., 2003; Silva et al., 2011), and correlate with higher biofilm productivity in our laboratory model (Poltak and Cooper, 2011). This model of selection involves a daily cycle of attachment to a polystyrene bead, biofilm formation, dispersal, and reattachment (Poltak and Cooper, 2011), and W colonies are among the first variants to evolve in each replicate population. We quantified relative fitness and biofilm phenotypes of many W mutants and found them to be uniformly highly adaptive, and sequenced their genomes to determine their genetic bases (Cooper et al., 2014). The sequences revealed exceptional parallelism in mutations affecting the operon orthologous to the well-characterized Pseudomonas wsp (wrinkly-spreader) operon: 29 of 35 causative, non-synonymous mutations affected the sensor protein, WspA, or the terminal sensor histidine kinase, WspE (Bantinaki et al., 2007; Cooper et al., 2014). Identical missense mutations occurred in multiple replicates and highlighted residues important for system function. This study implied that the missense mutations led to constitutive activation of signaling in contrast to the LOF mutations that de-repress the system observed in Pseudomonas (McDonald et al., 2009). This study also showed that different W mutants inhabit a complementary niche by attaching early to the plastic surface and facilitating attachment of other cells (Traverse et al., 2013; Cooper et al., 2014). Nevertheless, the proximate, structural cause of the W phenotype and its regulation in Burkholderia have been unresolved.
When testing the stability of the W phenotype outside of biofilms, we noticed that W populations grown in planktonic cultures produced Smooth (S) colonies at a rate of approximately 1/500. These mutants, which were heritable suppressors, motivated this study of the genetic and evolutionary mechanisms that permit biofilm specialists to regain planktonic fitness. We predicted that the duration of time that W mutants had adapted to biofilm growth would influence their paths to S suppression. We used the classic design of evolving populations founded from multiple ancestors differing in their adaptive history (Travisano et al., 1995; Kryazhimskiy et al., 2014), and conducted a suppressor screen using experimental evolution and genome sequencing to determine what kinds of mutations, and in what genes, enable biofilm specialists to subsequently adapt in a planktonic environment. We were specifically interested in whether this phenotypic suppression involved mutations that quantitatively modulated prior biofilm adaptations or rather occurred by qualitative LOF mutations that eliminated these adaptations. Furthermore, we investigated whether the ecological consequences of the W→S switch in multiple environments varied with evolutionary history, and predicted that mutants derived from more adapted ancestors would exhibit greater tradeoffs associated with suppression.
Here we present evidence suggesting that the length of biofilm adaptation limits the number of pathways by which SCV’s may evolve to restore niche breadth and balance biofilm and planktonic growth. More derived W ancestors tended to produce S mutants by secondary wsp mutations that reduced biofilm production but retained high fitness in each of the experimental environments. In contrast to our prediction, these mutants produced a discrete phenotypic switch with limited pleiotropic consequences. On the other hand, S mutants from more naïve ancestors revealed more systematic tradeoffs that largely restored the ancestral phenotype and eliminated any prior fitness gains in the biofilm environment. Furthermore, the mutations producing the S phenotype also altered biofilm composition and the ability to disperse from the biofilm, revealing the likely target of selection. Finally, this work identifies a novel two-component pathway that we predict integrates Wsp-mediated signaling with the transcriptional activation of biofilm-related genes leading to the W phenotype. Using experimental evolution to conduct both a forward and reverse genetic screen therefore provided evidence of how Burkholderia may balance biofilm development as well as dispersal.
Materials and Methods
Ancestral Strains and Experimental Conditions
The Burkholderia cepacia complex (Bcc) is a group of genetically diverse pathogens capable of infecting both plants and humans; notably patients with CF or chronic granulomatous disease (CGD) are at elevated risk of infection from environmental or nosocomial exposure, and suffer increased morbidity and mortality (Mahenthiralingam et al., 2005; Vial et al., 2011). While the strain used for this study, B. cenocepacia HI2424, was obtained from an onion field rather than from a human infection, the strain type represents an epidemic clone (LiPuma et al., 2002). As described previously (Ellis and Cooper, 2010), this ancestor was marked with lacZ to enable competitively neutral, blue-white screening of plated mixtures during direct competitions.
Although this lacZ-marked WT represents the original ancestor to all mutants described herein, our mutants were generated via a pair of independent evolution experiments conducted previously in our laboratory. As illustrated in Figure 1, the mutants produced in our suppression study were isolated from both short-term evolution (STE; Cooper et al., 2014) and long-term evolution (LTE) experiments (Traverse et al., 2013). The general approach, summarized below, was followed for both evolution experiments and has been described previously (Poltak and Cooper, 2011): ancestral clones were revived from frozen stock and grown for 24 h on a roller drum (RD; New Brunswick TC-7 model) at 37°C in 5 mL tryptic soy (T-soy; Fisher) broth using 18 mm × 150 mm test tubes. 50 μL of vortexed broth was subcultured in 5 mL M9 minimal media supplemented with 3% galactose (3% GMM) containing a sterile 7 mm polystyrene bead (American Education Products) and incubated at 37°C on the RD for 24 h. For the LTE experiment, a single Lac+ clone was inoculated into growth media. The bead was transferred daily to a new tube with fresh media and an oppositely marked bead, thus selecting for populations that were capable of daily biofilm formation and dispersal. Subsequent STE experiments were altered such that oppositely marked ancestral populations were mixed to identify beneficial mutants through marker divergence. Specifically, beads and biofilm colonists were transferred to a tube with phosphate buffered saline (PBS), vortexed, and marked (Lac+) and unmarked (Lac-) ancestors were added in equal 500 μL volumes to 4 mL of 3% GMM in tubes with a new bead, and incubated at 37°C for 24 h on the RD. Populations undergo approximately eight generations per day in this model. Populations were sampled by removing the 48-h bead, diluting 1:100,000 in PBS, and plating on half-strength T-soy agar plates containing X-gal. Plates were incubated for 24 h at 37°C, then at room temperature for 48 h. Unique colony morphologies were isolated and streaked to ensure selection of heritable traits. Freezer stocks were generated for each mutant isolate and stored in 8% DMSO at -80°C. Though a variety of phenotypes were generated from these STE and LTE experiments (Poltak and Cooper, 2011), only SCVs termed Wrinkly (W) were analyzed in this study.
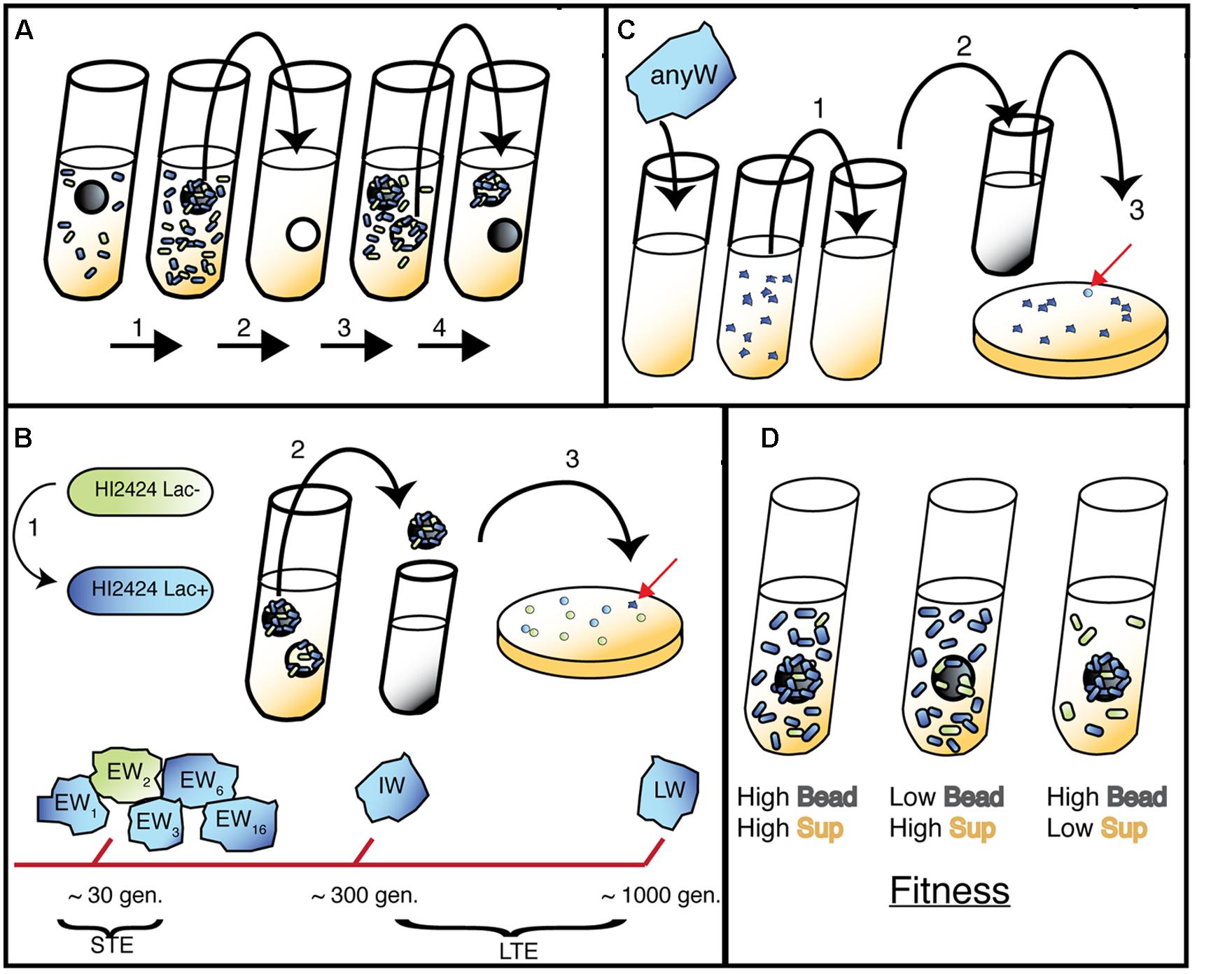
FIGURE 1. Bead model of biofilm evolution. (A) Bead-transfer method used in both evolution experiments. (1) A fresh bead is added to an inoculated tube containing marked (long-term evolution, LTE) or both marked and unmarked (short-term evolution, STE) WT populations and grown for 24 h. (2) Bead is colonized by biofilm cells and transferred to a fresh tube containing a sterile bead and growth media. (3) Bacteria disperse from original bead and attach to new bead. (4) New bead is selected and process repeats; note that 48-h bead is archived to sample representative population. (B) (1) Lac+ strain used in LTE and STE experiments, Lac- used in STE only. (2) Following daily transfer of colonized bead, remaining 48 h bead was removed and vortexed in PBS. (3) Populations were plated and screened for different morphologies related to adaptive traits (red arrow indicates a novel W mutant). Early Wrinkly (EW) mutants were detected at ∼30 generations, while Intermediate Wrinkly (IW) and Late Wrinkly (LW) were detected at ∼300 and 1000 generations. (C) Isolation of Smooth mutants. 1. W colonies were serially passaged in planktonic environment for three days. (2) Samples were diluted and plated. (3) Smooth colonies were detected and isolated. (D) Predicted fitness effects of adaptive mutants (blue cells) in sup and bead environment, with three outcomes presented: from left to right, early colonization and rapid dispersal; rapid dispersal but delayed colonization; early colonization and slow dispersal.
Experimental Evolution and Mutant Collection
Seven different W mutants were evolved to produce S variants, as follows. W cultures were streaked directly from freezer stock; a second streak was completed from a single isolate to ensure a homogenous clone was selected. This isolate was used to inoculate 5 mL of Tsoy broth, and incubated for 24 h at 37°C in the RD. 50 μL of the 24 h Tsoy broth was used to subculture a 5 mL tube containing 3% GMM without any bead, and incubated for 24 h. Transfers were repeated for 3 days in a planktonic environment, during which samples of broth were plated on strength Tsoy agar and examined for potential reversion of phenotype. S mutants were isolated from an agar plate, stocked at -80°C in 8% DMSO, and subjected to a bead-to-bead transfer in 3% GMM media for two days to ensure that the colony phenotype was heritable. Mutants listed with the same Wrinkly ancestor were generated from independent replicates; however, multiple mutants generated from a single culture were discarded from analysis due to an inability to verify their independent origin prior to sequencing.
Relationships Between Colony Morphology and Mucoidy
All members of the Bcc have demonstrated capacity to produce a smooth, high EPS-producing phenotype termed “mucoid,” which is the result of enhanced expression of EPS (Zlosnik et al., 2008). The Mucoid phenotype is a result of enhanced expression of EPS. Following procedures described previously (Zlosnik et al., 2008), freezer stock of each Smooth was streaked on agar plates with minimal media consisting of 0.5 gL-1 yeast extract and 4 gL-1 mannitol to differentiate Mucoid (M) from Non-mucoid (N). Streaks were performed in duplicate, and observed at 24 and 48 h to confirm the production of EPS. Known positive and negative controls were obtained from a previous study that confirmed the presence and absence of EPS (Silva et al., 2011). Only isolates that produced copious amounts of EPS that diffused well outside of the streaked area were characterized as M.
Genome Sequencing
Genomes of all isolates were sequenced as described for both the STE (Cooper et al., 2014) and LTE (Traverse et al., 2013) experiments. Briefly, mutant isolates were grown on a bead, vortexed in PBS, and genomic DNA was extracted using the DNeasy Blood and Tissue Kit (Qiagen) protocol for Gram-negative bacteria. Libraries were prepared following standard protocols (Nugen). LTE genomes were sequenced by the Joint Genome Institute with an Illumina GAII using paired-end 36 bp reads (Traverse et al., 2013), while STE and all S mutant genomes were sequenced on the Illumina HiSeq2500 at the Hubbard Center for Genome Studies at the University of New Hampshire. Reads were mapped to the previously sequenced B. cenocepacia HI2424 reference genome (LiPuma et al., 2002) revealing > 100x coverage, for each clone. Mutations were identified using the breseq pipeline with default settings, focusing only on high-confidence mutations, although marginal calls or missing coverage were evaluated by visual inspection of aligned reads when present at high frequency or when the gene was mutated in more than one clone (Deatherage and Barrick, 2014).
Fitness Assays
Methods for fitness assays were reported previously for both STE (Cooper et al., 2014) and LTE (Traverse et al., 2013) experiments, but now include a new environmental condition highlighted below. As previously, three- or fourfold replicate competitions were conducted between oppositely marked ancestor and mutants. All samples were revived from freezer stock in 5 mL Tsoy and incubated at 37°C for 24 h on a RD. 50 μL of Tsoy culture was added to 1 mL of PBS, vortexed, and 50 μL of the dilute sample was conditioned in 5 mL 3% GMM either with a bead or without (termed the planktonic environment) for 24 h on a RD at 37°C. For the bead environment, the single bead conditioned in GMM was removed with sterile forceps, transferred to 1 mL PBS and vortexed; the planktonic environment was completed by transferring 50 μL into the PBS dilution tube and vortexing. Competitions between evolved isolates and ancestral WT were conducted by adding 250 μL of mutant culture and 750 μL of WT to 4 mL GMM. Samples were vortexed, 100 μL of broth was removed and diluted 1:10,000 in PBS, and plated on Tsoy agar with X-gal. Cultures were incubated at 37°C in a RD for 24 h. Following incubation, planktonic tubes were vortexed and 50 μL of culture was added to 1 mL PBS and diluted; dilution samples were plated at both 1:10,000 and 1:100,000 on Tsoy agar with X-gal. Bead tubes followed a similar dilution protocol, with the bead and biofilm community being transferred to the PBS tube instead. A new environment, termed ‘sup’ for supernatant, was introduced in this experiment and represented the vortexed remains once the bead was removed from the T = 24 h tube. Similar to the planktonic culture, 50 μL of the broth remaining in the bead tube was transferred to a PBS tube, diluted, and plated. All plates were incubated for 24 h at 37°C and then 48 h at room temperature before colonies were enumerated. Colony forming units (CFUs) at T = 0 and T = 24 h were used to determine relative fitness as the selection rate constant, which is the difference (mutant – ancestor) in the natural logarithm of realized growth over 24 h (Lenski et al., 1991).
Biofilm and Exopolysaccharide (EPS) Assays
Biofilm production was measured via crystal violet absorbance (O’Toole, 2011). Briefly, freezer stock was revived in 5 mL of Tsoy, and grown overnight for 24 h at 37°C in a RD. Tsoy broth was added to 3% GMM to create a 5 mL solution with an optical density (O.D.) of 0.1, with 200 μL of diluted sample added to wells of a 96-well plate (Corning Costar). Four replicates were conducted for each treatment. Following 24 h of incubation at 37°C, biofilm production was measured by crystal violet staining, with 550 nm absorbance values recorded for all samples.
Relative EPS production was quantified following a hybrid of methods described previously (Ueda and Wood, 2009). This growth media is different than that used to measure biofilm production and quantify fitness, and was selected for its ability to differentiate EPS production among mutants. Isolate colonies were revived from freezer stock in 5 mL of Tsoy and grown overnight for 24 h at 37°C in a RD. Tsoy broth was added to respective 96-well plate containing 200 μL of mannitol media (MM) with 40 μg/mL Congo red dye such that all flasks had initial O.D. of 0.1. Inoculated wells were incubated for 24 h at 37°C, after which the contents of wells were pelleted in a centrifuge at 4,000 rpm for 10 min at 4°C. One hundred microliters of supernatant were transferred to a new 96-well plate, and contents were again centrifuged. Finally, 50 μL of this supernatant was added to a new 96-well plate, and absorbance was measured at 490 nm. Relative EPS production was reported as the difference in absorbance between blank (Congo Red and mannitol) and sample.
Statistical Analyses
Analysis of variance (ANOVA) and post hoc Tukey tests were performed with JMP software (JMP®, Version 11.0. SAS Institute Inc., Cary, NC, USA, 1989–2007). A P-value of <0.05 was considered statistically significant. Post hoc comparisons between experimental factors were delineated by different letters if statistically different, and share the same letter if indistinguishable.
Results
Isolation of S Mutants from Genetically Distinct W Ancestors
Mutants with a W phenotype, which exhibited nearly invariant high fitness in a model biofilm environment (Cooper et al., 2014), were significantly worse in a planktonic environment lacking a plastic bead for attachment (Figure S1). We predicted this tradeoff generated strong selection for mutants that restore planktonic fitness. Seven W mutants with different genotypes and evolutionary histories were each used to found several replicate populations that were passaged under planktonic conditions for 3 days and screened for colony variants. Of these W ancestors, five had previously evolved during biofilm selection for 32–64 generations and acquired single wsp mutations (Cooper et al., 2014), one evolved for 315 generations and acquired at least one secondary mutation in rpoC, and one W ancestor evolved for 1050 generations and acquired at least eight mutations (Traverse et al., 2013). Twenty mutants with S morphology from these different ancestors were collected and labeled ‘E’ for arising from an Early W ancestor, ‘I’ for arising from the Intermediate W ancestor, or ‘L’ for the Late ancestor. Once these phenotypes were confirmed to be heritable, the genomes of these mutants were sequenced to determine the genetic basis of suppression (Table 1) and to identify any additional mutations acquired during the 3 days of passaging in growth media (Table S1). Isolates were labeled according to their duration of evolution (Early-E, Intermediate-I, Late-L), colony morphology (W or S), production of EPS [Mucoid, M, or Non-mucoid (N)], and ancestor (numeral).
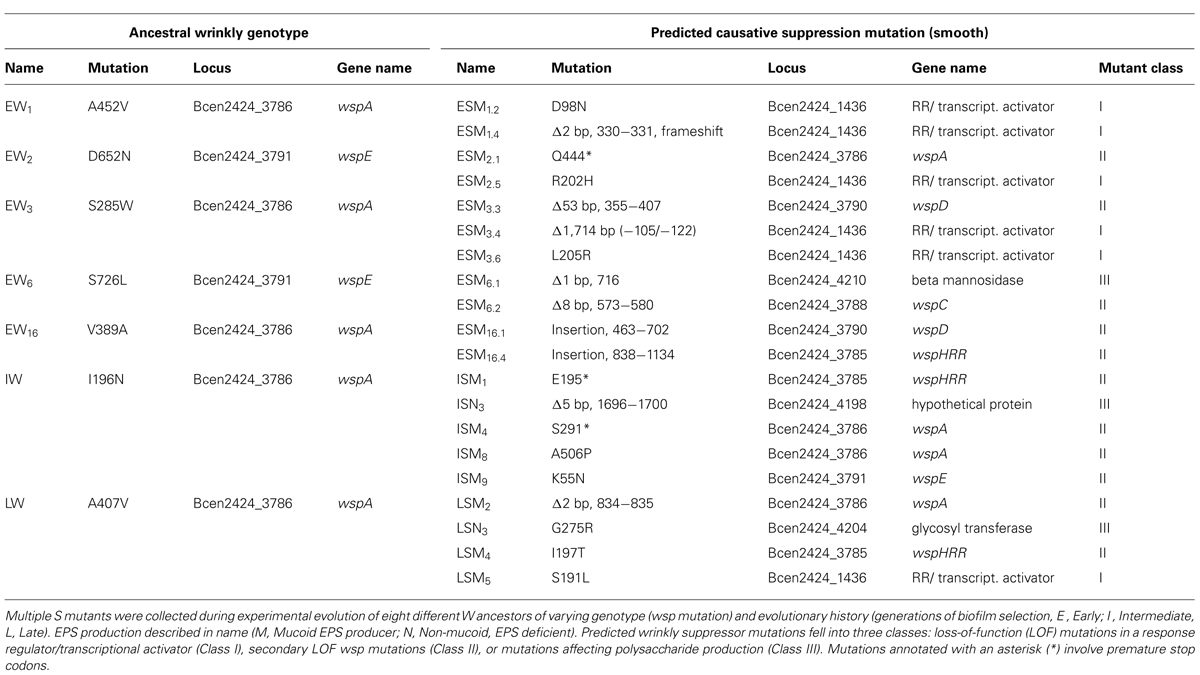
TABLE 1. Mutations predicted to produce Smooth (S) colonies from various Wrinkly (W) ancestors.
Mutations Suppressing the W Phenotype form Three Classes
All seven W ancestors used in this study possess a mutation in either the wspA or wspE genes (Table 1; Traverse et al., 2013; Cooper et al., 2014). Of the 20 suppression mutants of these W ancestors, 11 isolates acquired a secondary wsp mutation that likely converted the colony phenotype from wrinkly to smooth, but only three of these new mutations occurred in the same wsp gene as the initial wsp mutation. The remaining nine S mutants involved non-wsp mutations. Given the strong parallelism among independent mutants, we confidently attribute missense mutations and a single deletion in an undescribed two-component transcriptional regulator, Bcen2424_1436 (termed “1436” moving forward), to be the genetic cause of six suppression events. Notably, all five SNP’s within 1436 clustered within putative REC and OmpR domains (Figure S2). Though wsp and 1436 together accounted for 17 of the 20 suppression mutations, three additional mutations in gene clusters associated with polysaccharide production illustrated additional paths by which the W phenotype can revert to the ancestral smooth colony type. These three mutations did not occur in the same operon, however, nor do their annotations suggest that they contribute exclusively with EPS production (Winsor et al., 2008). We collectively refer to these three genes as PPA for “Polysaccharide Production-Associated,” given their putative functions.
Importantly, the types of W suppression mutations (wsp, 1436, or PPA) were not equally distributed among isolates of different selective histories. Among the I and L mutants that evolved from a more derived ancestor (i.e., following a greater period of biofilm selection), six of nine acquired a secondary suppression mutation in wsp and only one mutant featured a mutation in 1436. Suppressor mutations identified in the ES isolates that evolved from W ancestors with more limited histories of biofilm selection occurred as many times in 1436 as in wsp, demonstrating alternative evolutionary paths to S. It should be noted that the frequency with which any of these S suppression phenotypes occurred was rare – often several plates with over 200 colonies of W phenotypes would have just a single smooth-colony mutant among them. Though the rate of mutation to the S phenotype was not quantified, E, I, and L ancestors each produced S mutants with low frequency (data not shown).
Although all S mutants acquired only a single mutation in either wsp, 1436, or PPA, additional missense or non sense mutations were identified in six of 21 isolates (Table S1). Three isolates had single SNPs, two isolates acquired a pair of unique SNPs each, and one isolate contained a SNP and a 2-base-pair missense substitution. The mutated genes were diverse, including glycosyltransferases, restriction enzymes, transcriptional regulators, and flagellar assembly proteins. These additional mutations may be beneficial and could contribute to the fitness of S mutants. More likely, however, is that these additional mutations may be effectively neutral and the presence of these mutations can be attributed to genetic hitchhiking with the three classes of suppression mutations, especially given that most S mutants contained only a single causative mutation.
Fitness Effects of Suppressor Mutants Differ by Genotype and among Environments
Mutations in the same gene or operon may nonetheless produce different phenotypes or growth affinities, especially if secondary mutations are not neutral, so we quantified fitness in three environments by direct competitions between S mutants and the original HI2424 ancestor. These environments were the selective biofilm environment with a plastic bead in a test tube, a planktonic environment lacking a bead, and (new to this study) a third condition termed ‘sup’ to indicate the supernatant phase of the bead-containing environment. The sup condition provides a unique opportunity to quantify the tendency of each mutant to disperse from an established biofilm or grow in the immediate suspension. Because the bead condition exclusively measures attached cells, and the sup condition the cells in the planktonic phase suspended above the biofilm, comparing fitness in the sup and bead conditions enables predictions about the life history strategy of an isolate. For example, mutants with high fitness in both the biofilm and sup environments suggest a strategy of early attachment and early dispersal. In contrast, a mutant with high fitness in the biofilm but low fitness in the sup condition suggests later dispersal timing, or alternatively, reduced ability to disperse. Lastly, mutants with low fitness in the biofilm but high fitness in the sup condition could be late or low biofilm formers and/or early dispersers. These assays of fitness across environments coupled with defined genotypes provided the opportunity to explore genetic mechanisms involved with distinct biofilm life history strategies (Figure 1).
Fitness effects of S mutants varied significantly with the duration of adaptation of the W parental genotypes in each of the three environments tested (Figure 2; Tables S2 and S3). The fitness of ES mutants was similar to WT in all environments, suggesting that the suppression mechanism largely
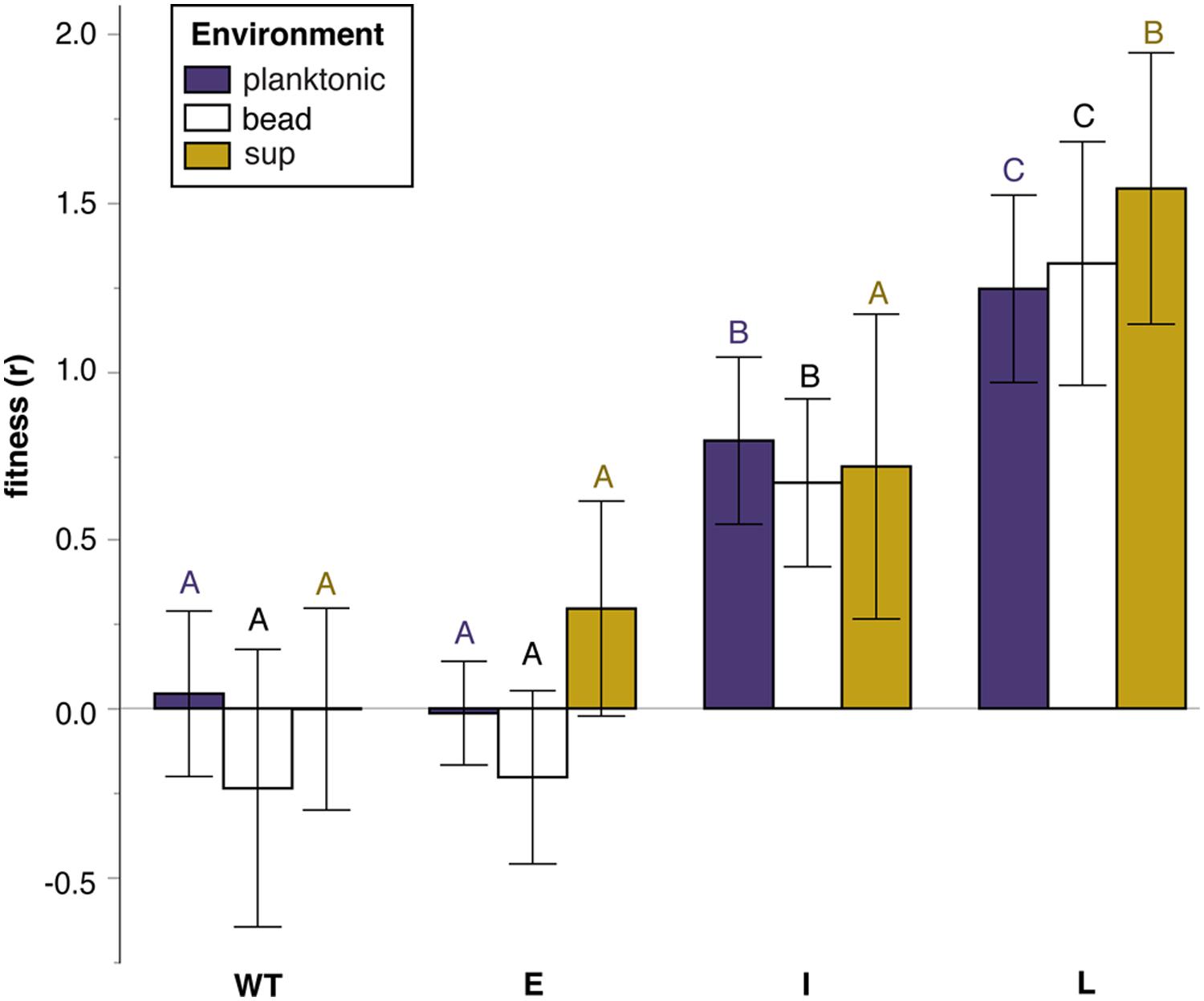
FIGURE 2. Relative fitness (selection rate, r ) of S mutants grouped by the duration of selection of theirW ancestor in a biofilm environment: Early (E) ~30 generations, Intermediate (I) ~300 generations, Late (L) ~1000 generations. Error bars are 95% CI. Tukey post hoc tests were used for pairwise comparisons. Unique letters within environments reflect significant differences.
restored WT fitness from the high biofilm-specific benefit of the W phenotype (Cooper et al., 2014). However, IS and LS mutants displayed significant fitness increases (Figure 2). Specifically, LS mutants were significantly adaptive in all three environments, whereas IS mutants were adaptive in both bead and planktonic environments. These results revealed that the relative fitness of S suppressor mutants varied both as a function of duration of biofilm adaptation and the environment in which they compete. Although mutants derived from ancestors that had previously evolved in the biofilm model for longer periods tended to be better adapted in all environments, the two non-mucoid isolates (ISN3 and LSN3) were outliers in the sup environment (Figure 3). These findings suggested that mutational mechanism as well as selective history could explain fitness variance.
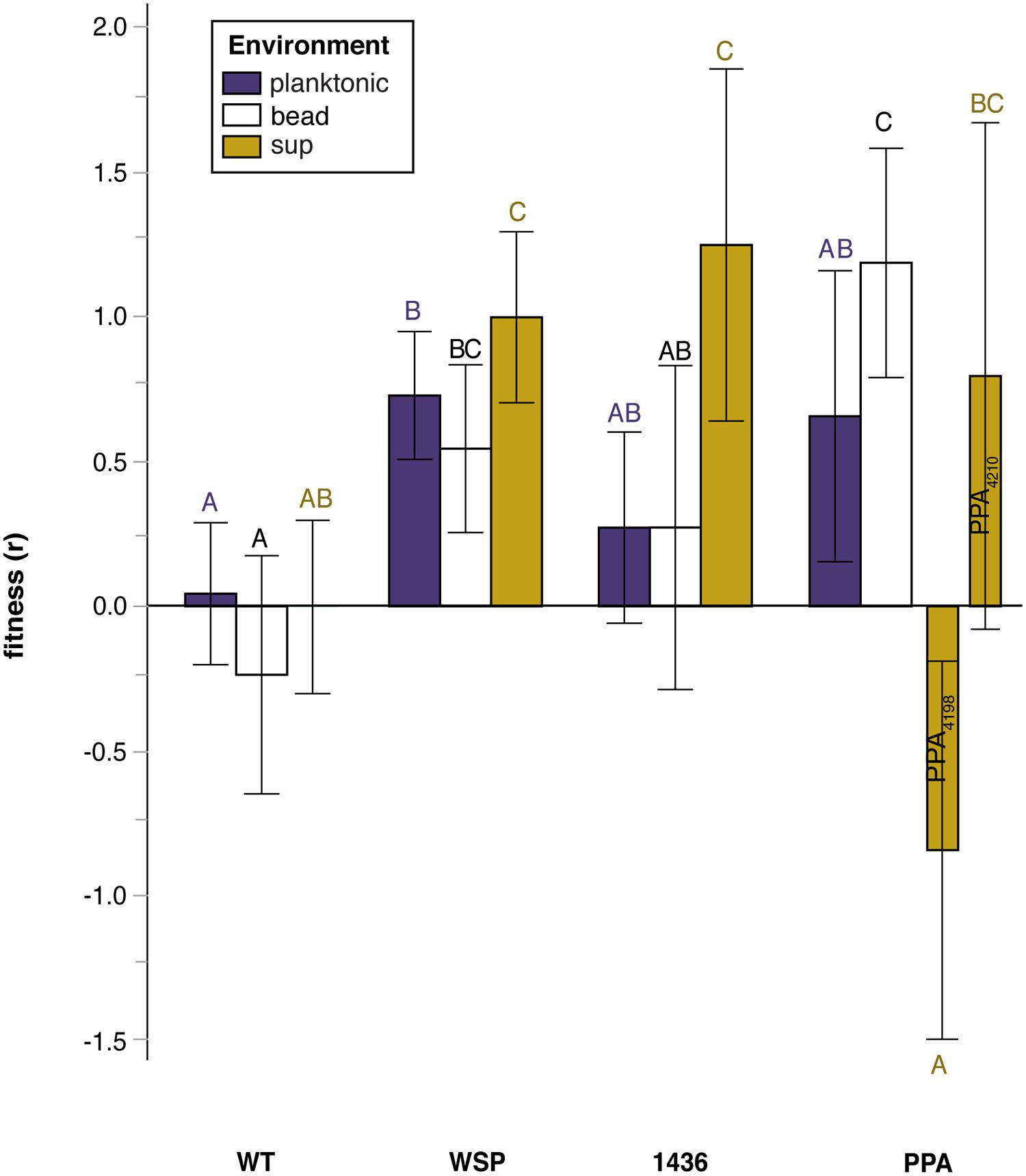
FIGURE 3. Relative fitness (selection rate, r) in three environments of mutants grouped by their suppression mutation class: wild-type (WT), two-component transcriptional regulator (1436), wrinkly-spreader operon (wsp), and putative polysaccharide production associated (PPA) genes. Note that mutation class PPA has been split for the ‘sup’ environment to distinguish between populations by duration of prior biofilm selection: ISN3 (intermediate) compared to LSN3 (late). Tukey post hoc tests were used for pairwise comparisons, with unique letters reflecting significant differences. Error bars are 95% CI of the mean.
We thus evaluated whether mutational class rather than the duration of biofilm selection explained fitness differences among suppressors (Figure 3; Table S3). Relative fitness varied significantly among mutational classes, with each group more fit than WT in at least one environment; however, only suppressors with a secondary mutation in wsp were significantly more fit across all environments, including the planktonic phase. The sup environment revealed the greatest variation in fitness among mutation classes: wsp and 1436 mutants were more fit than WT, but PPA mutants that were non-mucoid were less fit than WT.
Biofilm Phenotypes Associated with Suppression
As predicted by the tradeoff model between biofilm and planktonic growth, mean biofilm production varied significantly between W and S mutants (Figure 4; Tables S4 and S5). Prior studies of mutants from long-term selection in our model demonstrated that biofilm production tended to increase with further generations of selection (Poltak and Cooper, 2011; Cooper et al., 2014). Here, biofilm production of S suppressor mutants also varied with prior selective history (Figure 4A): LS mutants produced more biofilm than WT and other mutants, but IS and ES mutants produced similar biofilm as WT (Table S4). More evidently, biofilm output differed among mutational classes: both wsp and 1436 suppression mutants produced much less biofilm, whereas PPA mutations maintained biofilm productivity equal to wrinkly ancestors (Figure 4B; Table S5). This suggests that PPA mutations are not critical for biofilm production, whereas the two-component transcriptional regulator 1436, like wsp, is directly involved in the activation of biofilm production.
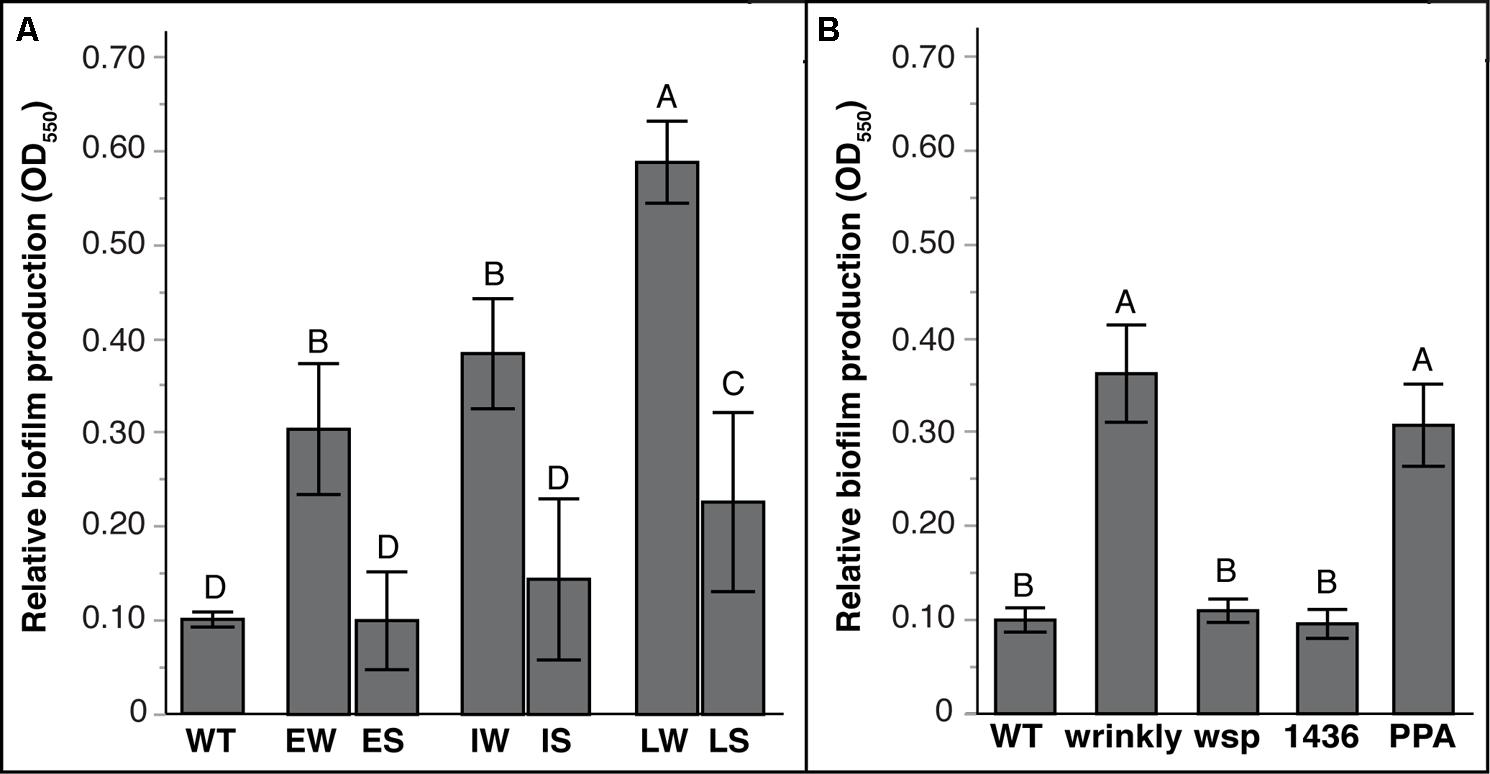
FIGURE 4. Biofilm production varies with duration of biofilm adaptation and by mutation class. Mean biofilm production with 95% CI shown. Tukey post hoc tests were used for pairwise comparisons, with unique letters reflecting significant differences. (A) Mean biofilm production by selective history of ancestors: Early Wrinkly (EW), Early Smooth (ES), Intermediate Wrinkly (IW), Intermediate Smooth (IS), Late Wrinkly (LW), Late Smooth (LS). (B) Biofilm production by mutation class: wild-type (WT), two-component transcriptional regulator (1436), wsp operon, and putative PPA genes.
Another clinically important phenotype that appeared to change with the shift from W to S colony types was their mucoidy (Silva et al., 2011; Zlosnik et al., 2011). When grown on agar plates containing mannitol, Mucoid (M) clones will produce significant quantities of exopolysaccharide (EPS) whereas Non-mucoids (N) will appear opaque and produce little or no visible liquid on the plate. Within 48 h, WT HI2424 and each W ancestor were capable of producing robust EPS. While 18 of 20 suppression mutants also formed copious quantities of EPS, two suppression mutants, ISN3 and LSN3, were non-mucoid. These two isolates each acquired unique suppression mutations in genes associated with EPS production (Fazli et al., 2013). Interestingly, a frameshift mutation in the other affected PPA gene, Bcen2424_4210, was sufficient to suppress the W phenotype without the loss of EPS production. This alternative compensatory mutational path was identified in mutants derived from EW, IW, and LW ancestors. These results illustrate that EPS production may be retained independently of the W phenotype, but mutations in at least two genes were sufficient to reduce both EPS production and suppress the W phenotype.
The extent of EPS production produced by these mutants also correlated with the mutational mode of suppression (Figure 5; Tables S6 and S7). PPA mutants were distinct as a group and among mutants within this group: mutants ISN3 and LSN3 both acquired mutations in a newly described EPS cluster (Fazli et al., 2013) and produced EPS at significantly lower levels than 1436 and wsp mutants (Figure 5B). PPA mutant ESM6.2 acquired a frameshift mutation in a gene putatively encoding a beta-mannosidase that is outside that putative EPS cluster, and produced the greatest amount of EPS among all mutants. To our surprise, and for uncertain reasons, W mutants produced less EPS than S mutants with 1436 or wsp mutations (Figure 5).
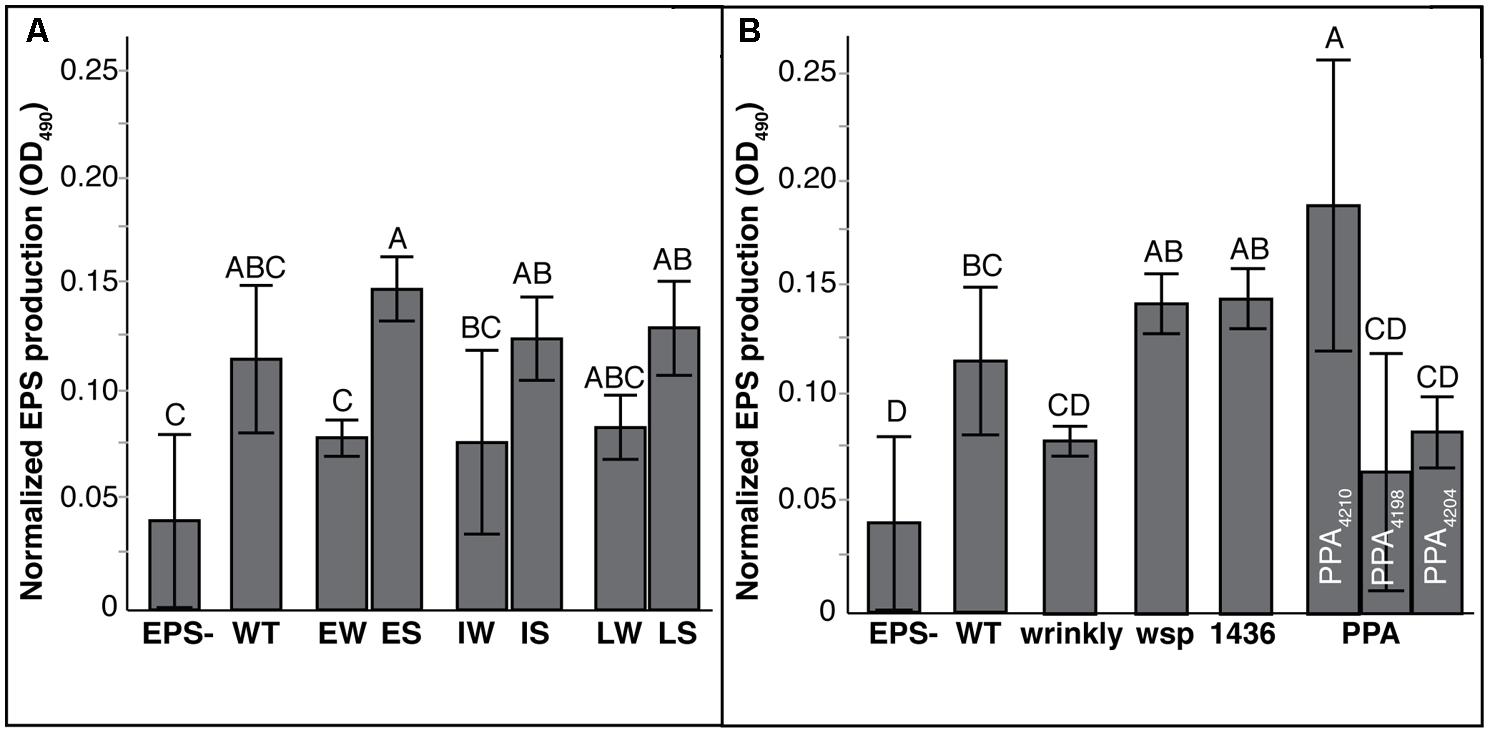
FIGURE 5. Exopolysaccharide (EPS) production differs by mutation class but not prior history of biofilm adaptation. EPS production is expressed as the mean difference between the blank and supernatant of culture, with 95% CI. Tukey post hoc tests were used for pairwise comparisons, with unique labels reflecting significant differences. (A) EPS production by generation of biofilms: Early Wrinkly (EW), Early Smooth (ES), Intermediate Wrinkly (IW, Intermediate Smooth (IS), Late Wrinkly (LW), Late Smooth (LS). (B) EPS production by mutation class: wild-type (WT), wrinkly, two-component transcriptional regulator (1436), wsp, and putative PPA genes. PPA mutants are plotted separated to show significant differences between mutations affecting putative EPS loci (4198 and 4204) and non-EPS loci (4210).
Discussion
It is now widely accepted that the biofilm lifestyle is the dominant mode of growth and survival for most bacteria (O’Toole et al., 2000; Karatan and Watnick, 2009). For decades, however, research has been primarily directed toward study of microbes in well-mixed planktonic cultures. Therefore, the understanding of bacterial physiology in naturalistic conditions may be tenuous at best. Fortunately, thee last 15 years have seen many genetic screens yield substantial insight into processes governing biofilm community assembly (Kuramitsu et al., 2007; Ryder et al., 2007; Irie and Parsek, 2008; McDougald et al., 2012; Fazli et al., 2014) and integrity (Spiers and Rainey, 2005; Borlee et al., 2010). Such models have demonstrated complexity in biofilm regulation, suggesting that these opportunistic pathogens exhibit multiple overlapping signaling networks that influence the fine-tuning of the composition and expression of biofilm components (Petrova and Sauer, 2009; Fazli et al., 2014).
It seems probable, though, that planktonic and biofilm growth as they are typically modeled in the laboratory represent extreme conditions. Rather, bacterial populations may fluctuate between motile and sessile states at a range of frequencies. Understanding of how bacteria genetically adapt to growth in a biofilm, but then successfully disperse to grow planktonically or to re-initiate biofilm formation, is much more limited, largely because of a lack of methods to realistically probe the phenotypes under selection in these variable environments. Most experiments have involved genetic screens for losses of biofilm production or planktonic growth. These studies tend to find mutations that block transitions between extreme phenotypes, but these findings lack nuance and tend not to find gain-of-function mutations. However, when transposon mutagenesis and the less biased method of experimental evolution were combined to probe P. fluorescens biofilm production, mutations in relatively few genes explained much of the variation in biofilm assembly (McDonald et al., 2009; Callahan et al., 2014). These studies suggest that perhaps only a handful of gene products are under positive selection for the production of certain biofilms. Yet little research has investigated how bacteria might subsequently adapt to a more motile lifestyle, as would be required to disperse and colonize a new environment. Moreover, suppression studies of mutants produced by transposons cannot realistically capture these lifestyle reversals because the initial step inherently prevents back mutations. Therefore, the experimental evolution approach presented here provides greater potential for investigating the genes and pathways that regulate biofilm growth as well as dispersal to resume planktonic growth.
Adaptive Genetic Mechanisms That Restore Dispersal Capacity for Biofilm Specialists
We previously characterized the genetic and ecological origins of the Wrinkly (W) phenotype in B. cenocepacia from many independently evolved mutants with varying histories of adaptation to biofilm growth on plastic beads (Traverse et al., 2013; Cooper et al., 2014). The cause of the W phenotype was nearly always a single missense mutation in either the wspA or wspE gene. Despite this evidence, the downstream targets of the wsp cluster and the causes of the wrinkly phenotype in B. cenocepacia have been unsolved, which motivated this study for mutations that would suppress this phenotype. Here we show that three classes of mutations can suppress the W phenotype and restore planktonic fitness and the ancestral colony phenotype. The first group, secondary mutations in the wsp operon, was most expected, although the locations of these suppressor mutations suggest that residues predicted to be involved in WspA dimerization (ISM8), WspE phosphorelay (ISM9), or WspHRR kinase activity (LSM4) are essential for Wsp system function (Table 1). Next, we discovered mutations in a novel transcriptional regulator, Bcen2424_1436. Importantly, all five SNP mutations occurred within predicted functional activation and response domains, demonstrating that losses of function in residues predicted to mediate phosphorylation or DNA binding produce similar phenotypes (Table 1; Figure S2). In addition, mutations in three other genes implicated in polysaccharide synthesis produced the S phenotype from W ancestors, indicating that the W phenotype likely depends more on downstream effects of Wsp signaling rather than the Wsp proteins directly. Importantly, nearly all of these suppressor mutations, which include nine indels and three premature stop codons, likely produce qualitative losses of function (Table 1). Evidently, the strong experimental selection to restore planktonic fitness favored LOF mutations that would be expected to limit subsequent evolution of W-type biofilm specialists.
The striking molecular parallelism among 1436 mutants illustrates the power of experimental evolution to identify functional roles for predicted proteins of unknown function. Among the six S 1436 mutants, one substitution occurred five amino acids from a KPV motif known to be a conserved part of the phosphate receiver domain, while four others each contained mutations at conserved residues associated with DNA binding (Marchler-Bauer et al., 2013; Figure S2). No other mutations occurred in these genotypes, which offers a clear prediction that the S phenotype results from either blocking the stimulation of 1436 by phosphorylation, or by eliminating its function as a DNA-binding transcriptional activator. This 1436 gene is found in many Burkholderia species and is always accompanied by a putative membrane-associated two-component response regulator with a histidine kinase domain, named Bcen2424_1438 in strain HI2424. This study therefore enables the additional prediction that a previously undefined two-component system, in which Bcen2424_1438 receives a periplasmic signal that it transduces to 1436 to activate transcription, plays a central role in modulating biofilm attachment and dispersal in Burkholderia.
The possible interactions between this 1438/1436 two-component system and the wsp system are less clear, but this study provides considerable circumstantial evidence. Again, recalling that all W ancestors began with a single wsp mutation, 17 of 20 S suppressors were caused by secondary mutations in either wsp or 1436. Importantly, S mutants with either wsp or 1436 mutations produced similar phenotypes and fitness profiles across all environments (Figure 3), which indicates that that these gene products likely cooperate in a single path regulating biofilm development in our experimental model. We therefore hypothesize that 1436, which has a N-terminal CheY-like response regulator and a C-terminal winged helix-turn-helix DNA binding effector domain, is directly responsible for activating transcription of biofilm associated processes, and is activated via phosphorylation of the wsp pathway by wspHRR (Figure 6). We previously predicted, based on very strong similarity to the Pseudomonas wsp system (Cooper et al., 2014), that a phosphorylation cascade is initiated upon surface growth being detected via WspA, leading ultimately to wspHRR activation. Here we predict that the phosphate transfer from wspHRR to 1436 completes the cascade and allows 1436 to bind and activate the transcription of genes that initiate biofilm production. However, the fact that 1436 is part of an operon containing a periplasmic protein, Bcen2424_1438, with its own predicted Chase2 and PAS sensory domains, suggests that alternative phosphorelay paths may activate overlapping biofilm-related genes. This 1438/1436 two-component system is broadly similar to EnvZ/OmpR and CreB/C in multiple well-studied bacterial genomes (Winsor et al., 2008), but is not the reciprocal best match of either of these sensor kinases that mediate central metabolism and play roles in both resistance and biofilm formation (Prigent-Combaret et al., 2001; Zamorano et al., 2014). Thus, the Wsp system may serve as the master regulator of surface detection in B. cenocepacia HI2424 as it does in Pseudomonas (O’Connor et al., 2012), but 1436 may integrate both Wsp signaling and additional sensed inputs. Clearly, further work detailing the transcripts activated by 1436 as well as the kinases that activate it will clarify the complex regulatory networks that govern Burkholderia biofilm development.
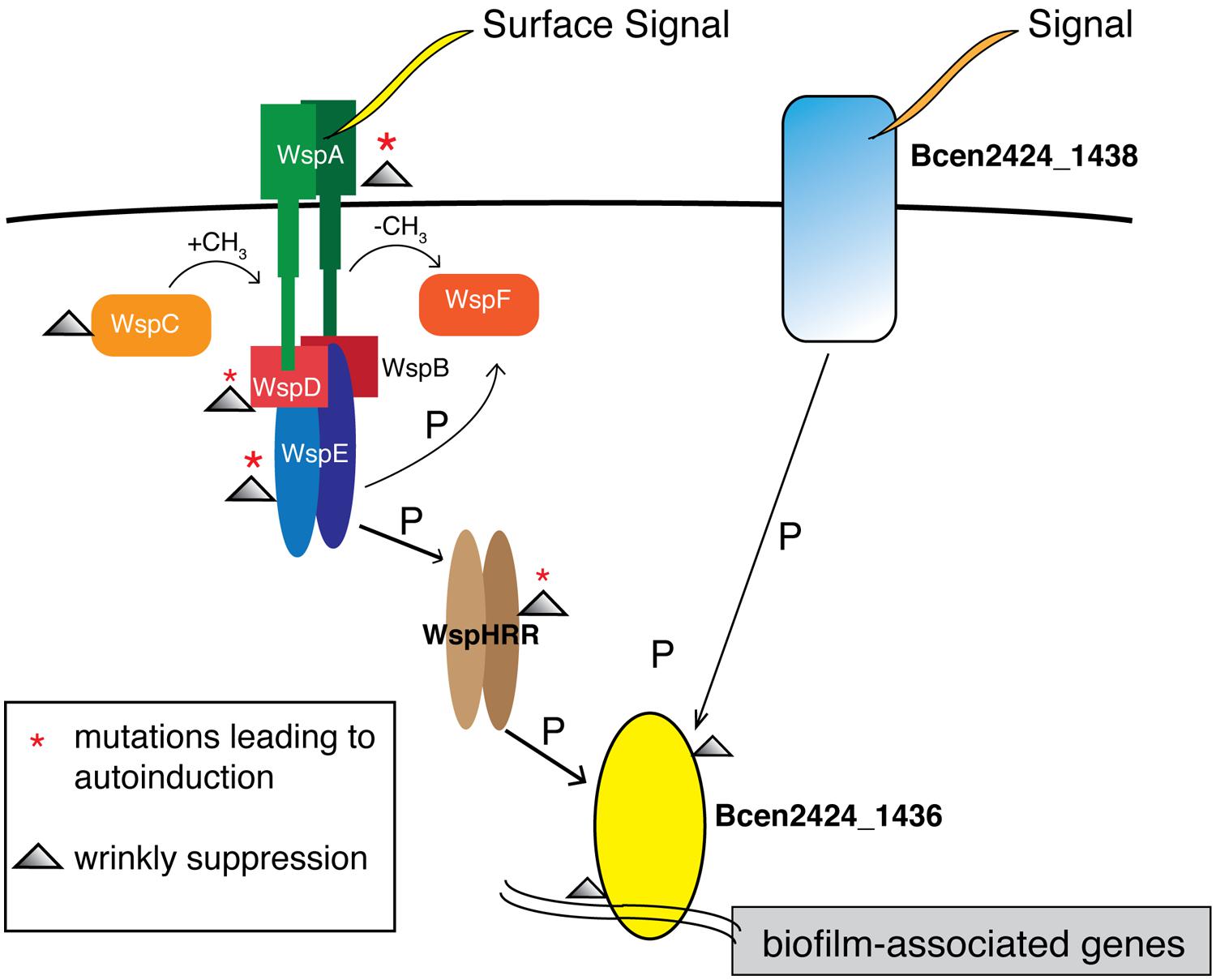
FIGURE 6. Model of biofilm regulation in Burkholderia cenocepacia building upon the Wsp model in Pseudomonas. Surface signaling at WspA leads to a phosphorylation cascade, activating WspHRR. W mutants in this study possessed mutations in wspA and wspE, leading to autoinduction of this process; S mutants possessed secondary mutations that also clustered in wsp or in Bcen2424_1436, as highlighted by symbols in the legend. We predict that the DNA-binding response-regulator Bcen2424_1436 is phosphorylated by WspHRR, which stimulates biofilm production by transcriptional activation. As part of a predicted two-component system, the Bcen2424_1436 protein also likely receive signals from a transmembrane sensor histidine kinase, Bcen2424_1438, which has conserved domains matching a periplasmic domain, a PAS fold, and a HATPase domain.
This study also illustrates parallels and differences between the Wsp system in Burkholderia and in Pseudomonas, in which the proximate cause of the W morphology is cellulose overproduction (Spiers and Rainey, 2005), and the ultimate source involves mutations that de-repress the diguanylate cyclase (DGC) WspR, leading to elevated levels of cyclic-di-GMP (Hickman et al., 2005; McDonald et al., 2009). In addition to Wsp, numerous other systems have been found in a wide range of bacteria to regulate biofilm formation via c-di-GMP levels (McDougald et al., 2012; Srivastava and Waters, 2012; Fazli et al., 2014). In contrast, the Burkholderia Wsp system does not contain a cognate DGC, so if altered cyclic di-GMP production is also the ultimate selective mechanism, the regulatory connection is indirect. This sensory system may therefore have been coopted in Burkholderia to activate transcription more directly by phosphorelay, with the DGC module being replaced by a histidine kinase that we called wspHRR (Cooper et al., 2014; Figure 6). Given our prediction that ancestral W mutants had constitutively active Wsp systems, a logical means of suppression would involve secondary wsp mutations, which we found 11 times. Six other mutations affected 1436, which demonstrate that its function is essential for Wsp signaling. However, the three mutations affecting genes related to the proximate cause of the smooth morphology, polysaccharide synthesis, tell a similar story to that of Pseudomonas. In both of these biofilm-forming bacteria, there appear to be relatively few regulatory ways to produce a hyper-biofilm producing specialist, but more ways involving both regulation and structural components to undo this specialization.
Effects of History on Adaptation to a New Environment: Paths and Tradeoffs
Another goal of this study was to evaluate the contingent effects of evolutionary history on adaptation, and to test the prediction that greater biofilm specialization would constrain adaptation to planktonic conditions. The results of this study generally support the notion that an organism’s life history is important but not sufficient to predict its evolutionary trajectory. We found that the suppression of an evolved W phenotype and its associated low fitness in planktonic conditions can arise by mutations in three major gene classes and furthermore, that the history of biofilm adaptation may limit the potential number of targets available to revert these phenotypes. While increased specialization appears to limit adaptive paths in a different environment, mutants that manage such a step appear nonetheless to benefit from their accumulated mutational history. Indeed, only S mutants evolved from more derived W ancestors were fitter than WT across all environments, in effect broadening their niche in fluctuating biofilm and planktonic environments and showing little evidence of tradeoffs. Overall, mutants ofW clones isolated following LTE in our model were fittest in nearly all comparisons (Figure 2), suggesting that their past selection in fluctuating biofilm environment favored genotypes that were broadly beneficial.
This evidence that past adaptive gains in the long-term biofilm model were robust to mutations affecting the W phenotype led us to test for fitness epistasis between wsp-associated mutations and other mutations found in the I and L genotypes (Khan et al., 2011; Flynn et al., 2013). Although the experimental design was not originally designed to evaluate this possibility and is therefore not ideal, we found no evidence of interactions for fitness among genotypes either having or lacking the set of mutations acquired during LTE (Table S8). This finding suggests the remarkable possibility that biofilm-adapted genotypes may gain or lose the SCV phenotype of W with little or no effect on other adaptations. If SCV phenotypes may generally evolve without influencing other adaptations, this could explain their frequent recovery from diverse biofilm-associated infections (Haussler et al., 2003; Starkey et al., 2009; Lee et al., 2011; Silva et al., 2011; Mayer-Hamblett et al., 2014).
Despite consistent benefits of prior adaptations in our laboratory system, S genotypes nevertheless varied in phenotypes and fitness depending on their mutational path adapting to a planktonic lifestyle. For instance, mutants of 1436 were most fit in the supernatant environment, reflecting early dispersal. We predict that these 1436 mutants fail to activate transcription of various biofilm-related genes and effectively fail to respond to the Wsp signal of surface attachment, and thus disperse too early to be sampled on the bead. In contrast, fitness effects of wsp double-mutants are consistently elevated relative to WT across all three environments and follow a similar logic: whereas the initial wsp mutation leads to constitutive attachment and high biofilm fitness, we predict the second suppressor mutation blunts this surface-attachment signal and restores the ability to disperse and achieve high fitness in the sup environment. This conclusion is complicated by the fact that more wsp double-mutants arose in more derived genotypes that were originally more fit. Despite these distinctions, the similar fitness profiles among 1436 and wsp mutants (Figure 3), as well as their similar reductions in biofilm and EPS production (Figures 4 and 5), support our model that they are integrated in a pathway initiating biofilm production (Figure 6). Thus, interrupting the signal of being attached to a surface may be a major step facilitating the transition from a biofilm-adapted to planktonic-adapted mode.
In general, the S suppression mutants produced consistently lower levels of biofilm and EPS regardless of prior adaptive history (Figures 4 and 5). These biofilm traits contrast with fitness in attaching to the bead, in which more evolved mutants may nevertheless benefit from other adaptive mutations in spite of reduced biofilm and EPS production. However, PPA mutants produced distinct fitness and phenotype effects that suggest the proximate cause of wrinkly colonies. Two PPA mutants enhanced fitness in attaching to the bead but reduced fitness in the supernatant of the biofilm. These mutants affected homologs of a newly described EPS cluster, Bcam1330-1341 (Fazli et al., 2013), reduced EPS production relative to WT, and became non-mucoid. The nature of these mutations is illuminating: one involved a deletion in a hypothetical protein that may be responsible for cleaving the final product for secretion into the matrix, while the other involved a missense mutation in a putative glycosyl transferase (Fazli et al., 2013). The third PPA mutation, a frameshift in beta-mannosidase, actually increased EPS production and this mutant was more fit in the supernatant environment; we predict this mutation disrupts mannose degradation and enhances polysaccharide synthesis. The fact that these mutations converted the wrinkly colony phenotype into smooth types suggests strongly that that the affected loci are downstream targets of the wsp/1436 regulon and major constituents of the biofilm. They may also play roles in balancing biofilm life history, as EPS synthesis may be required for dispersal in addition to biofilm assembly. Given the purported hygroscopic properties of these polysaccharides, an absence or even partial reduction in EPS production may substantially increase the viscosity of the matrix and prevent dispersal (McDougald et al., 2012). Further, because the PPA mutants retain the ancestral, constitutively activated Wsp system, the inability of the N types to form EPS could be combining with greater synthesis of other polymers, such as cellulose, that may prevent dispersal.
To conclude, this study began as a survey of the mutations that explained the relatively frequent evolution of smooth mutant colonies from wrinkly, SCV ancestors during growth in planktonic culture. We wondered whether the causes of these smooth suppressor mutants differed with the ancestral genotype and the duration of prior biofilm selection on these wrinkly ancestors; furthermore, we predicted that greater biofilm selection would generate greater tradeoffs across environments for these suppressors owing to stronger antagonistic pleiotropy. We paired short-term experimental evolution with high-throughput sequencing to survey the mechanisms of planktonic adaptation and suppression of the W phenotype, and conducted a range of assays of fitness in multiple environments and of biofilm-associated phenotypes. The evolved mutations identified mechanisms by which multiple previously uncharacterized genes contribute to the balance of biofilm formation and planktonic growth. Similarly, the fitness assays contribute to a narrative that shows how these various genotypes and phenotypes may evolve in response to a common selective force. Evolutionary history was generally predictive of fitness and biofilm production, but not for EPS synthesis. The specific mutational paths taken appear to be more predictive still, such that despite any advantageous mutational heritage derived from prolonged biofilm adaptation, a single base pair substitution in a few targets appears sufficient to dramatically alter phenotype, polysaccharide production, and fitness across a range of environments.
These findings may have broader implications for the evolutionary genetics of microbial populations. Perhaps most striking, the suppression of more adapted W mutants disproportionately involved secondary wsp mutants, which implies the evolution of constraint on alternative genetic pathways. Nevertheless, this constraint did not produce greater tradeoffs; rather, it appears that selection in the new planktonic environment limited the sampling of beneficial mutations to those producing few if any costs across environments. Perhaps with continued adaptation in one environment, the distribution of effects of beneficial mutants found in a new environment becomes increasingly skewed toward direct suppressors and revertants that undo prior specializing mutations. Consequently, genotypes relatively naïve in the first environment may be compensated by a range of mutations in the second, but not so in the case of a more derived genotype. Therefore, with increasing history of selection in a single environment, options for the evolutionary return trip may become few indeed.
Conflict of Interest Statement
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Supplementary Material
The Supplementary Material for this article can be found online at: http://www.frontiersin.org/journal/10.3389/fgene.2015.00018/abstract
References
Abee, T., Kovács,Á. T., Kuipers, O. P., and Van Der Veen, S. (2011). Biofilm formation and dispersal in Gram-positive bacteria. Curr. Opin. Biotechnol. 22, 172–179. doi: 10.1016/j.copbio.2010.10.016
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Bantinaki, E., Kassen, R., Knight, C. G., Robinson, Z., Spiers, A. J., and Rainey, P. B. (2007). Adaptive divergence in experimental populations of Pseudomonas fluorescens. III. Mutational origins of wrinkly spreader diversity. Genetics 176, 441–453. doi: 10.1534/genetics.106.069906
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Barrick, J. E., and Lenski, R. E. (2013). Genome dynamics during experimental evolution. Nat. Rev. Genet. 14, 827–839. doi: 10.1038/nrg3564
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Borlee, B. R., Goldman, A. D., Murakami, K., Samudrala, R., Wozniak, D. J., and Parsek, M. R. (2010). Pseudomonas aeruginosa uses a cyclic-di-GMP-regulated adhesin to reinforce the biofilm extracellular matrix. Mol. Microbiol. 75, 827–842. doi: 10.1111/j.1365-2958.2009.06991.x
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Callahan, B. J., Fukami, T., and Fisher, D. S. (2014). Rapid evolution of adaptive niche construction in experimental microbial populations. Evolution 68, 3307–3316. doi: 10.1111/evo.12512
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Cooper, T. F. (2007). Recombination speeds adaptation by reducing competition between beneficial mutations in populations of Escherichia coli. PLoS Biol. 5:e225. doi: 10.1371/journal.pbio.0050225
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Cooper, V. S., Staples, R. K., Traverse, C. C., and Ellis, C. N. (2014). Parallel evolution of small colony variants in Burkholderia cenocepacia biofilms. Genomics 104, 447–452. doi: 10.1016/j.ygeno.2014.09.007
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Danhorn, T., and Fuqua, C. (2007). Biofilm formation by plant-associated bacteria. Annu. Rev. Microbiol. 61, 401–422. doi: 10.1146/annurev.micro.61.080706.093316
Deatherage, D. E., and Barrick, J. E. (2014). Identification of mutations in laboratory-evolved microbes from next-generation sequencing data using breseq. Methods Mol. Biol. 1151, 165–188. doi: 10.1007/978-1-4939-0554-6_12
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Elena, S. F., and Lenski, R. E. (2003). Evolution experiments with microorganisms: the dynamics and genetic bases of adaptation. Nat. Rev. Genet. 4, 457–469. doi: 10.1038/nrg1088
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Ellis, C. N., and Cooper, V. S. (2010). Experimental adaptation of Burkholderia cenocepacia to onion medium reduces host range. Appl. Environ. Microbiol. 76, 2387–2396. doi: 10.1128/AEM.01930-09
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Fazli, M., Almblad, H., Rybtke, M. L., Givskov, M., Eberl, L., and Tolker-Nielsen, T. (2014). Regulation of biofilm formation in Pseudomonas and Burkholderia species. Environ. Microbiol. 16, 1961–1981. doi: 10.1111/1462-2920.12448
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Fazli, M., Mccarthy, Y., Givskov, M., Ryan, R. P., and Tolker-Nielsen, T. (2013). The exopolysaccharide gene cluster Bcam1330-Bcam1341 is involved in Burkholderia cenocepacia biofilm formation, and its expression is regulated by c-di-GMP and Bcam1349. Microbiologyopen 2, 105–122. doi: 10.1002/mbo3.61
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Flynn, K. M., Cooper, T. F., Moore, F. B. G., and Cooper, V. S. (2013). The environment affects epistatic interactions to alter the topology of an empirical fitness landscape. PLoS Genet. 9:e1003426. doi: 10.1371/journal.pgen.1003426
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Guo, Y., and Rowe-Magnus, D. A. (2010). Identification of a c-di-GMP-regulated polysaccharide locus governing stress resistance and biofilm and rugose colony formation in Vibrio vulnificus. Infect. Immun. 78, 1390–1402. doi: 10.1128/IAI.01188-09
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Haussler, S., Lehmann, C., Breselge, C., Rohde, M., Classen, M., Tummler, B.,et al. (2003). Fatal outcome of lung transplantation in cystic fibrosis patients due to small-colony variants of the Burkholderia cepacia complex. Eur. J. Clin. Microbiol. Infect. Dis. 22, 249–253.
Hickman, J. W., Tifrea, D. F., and Harwood, C. S. (2005). A chemosensory system that regulates biofilm formation through modulation of cyclic diguanylate levels. Proc. Natl. Acad. Sci. U.S.A. 102, 14422–14427. doi: 10.1073/pnas.0507170102
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Irie, Y., and Parsek, M. R. (2008). Quorum sensing and microbial biofilms. Curr. Top. Microbiol. Immunol. 322, 67–84. doi: 10.1007/978-3-540-75418-3_4
Jeannot, K., Elsen, S., Kohler, T., Attree, I., Van Delden, C., and Plesiat, P. (2008). Resistance and virulence of Pseudomonas aeruginosa clinical strains overproducing the MexCD-OprJ efflux pump. Antimicrob. Agents Chemother. 52, 2455–2462. doi: 10.1128/AAC.01107-07
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Kaplan, J. B. (2010). Biofilm dispersal: mechanisms, clinical implications, and potential therapeutic uses. J. Dent. Res. 89, 205–218. doi: 10.1177/0022034509359403
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Karatan, E., and Watnick, P. (2009). Signals, regulatory networks, and materials that build and break bacterial biofilms. Microbiol. Mol. Biol. Rev. 73, 310–347. doi: 10.1128/MMBR.00041-08
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Kawecki, T. J., Lenski, R. E., Ebert, D., Hollis, B., Olivieri, I., and Whitlock, M. C. (2012). Experimental evolution. Trends Ecol. Evol. (Amst.) 27, 547–560. doi: 10.1016/j.tree.2012.06.001
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Khan, A. I., Dinh, D. M., Schneider, D., Lenski, R. E., and Cooper, T. F. (2011). Negative epistasis between beneficial mutations in an evolving bacterial population. Science 332, 1193–1196. doi: 10.1126/science.1203801
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Kryazhimskiy, S., Rice, D. P., Jerison, E. R., and Desai, M. M. (2014). Microbial evolution. Global epistasis makes adaptation predictable despite sequence-level stochasticity. Science 344, 1519–1522. doi: 10.1126/science.1250939
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Kuramitsu, H. K., He, X., Lux, R., Anderson, M. H., and Shi, W. (2007). Interspecies interactions within oral microbial communities. Microbiol. Mol. Biol. Rev. 71, 653–670. doi: 10.1128/MMBR.00024-07
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Lang, G. I., Rice, D. P., Hickman, M. J., Sodergren, E., Weinstock, G. M., Botstein, D.,et al. (2013). Pervasive genetic hitchhiking and clonal interference in forty evolving yeast populations. Nature 500, 571–574. doi: 10.1038/nature12344
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Lee, B., Schjerling, C. K., Kirkby, N., Hoffmann, N., Borup, R., Molin, S.,et al. (2011). Mucoid Pseudomonas aeruginosa isolates maintain the biofilm formation capacity and the gene expression profiles during the chronic lung infection of CF patients. APMIS 119, 263–274. doi: 10.1111/j.1600-0463.2011.02726.x
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Lee, K. W. K., Periasamy, S., Mukherjee, M., Xie, C., Kjelleberg, S., and Rice, S. A. (2013). Biofilm development and enhanced stress resistance of a model, mixed-species community biofilm. ISME J. 8, 894–907. doi: 10.1038/ismej.2013.194
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Lenski, R. E., Rose, M. R., Simpson, S. C., and Tadler, S. C. (1991). Long-term experimental evolution in Escherichia coli. I. Adaptation and divergence during 2,000 generations. Am. Nat. 138, 1315–1341. doi: 10.1086/285289
LiPuma, J. J., Spilker, T., Coenye, T., and Gonzalez, C. F. (2002). An epidemic Burkholderia cepacia complex strain identified in soil. Lancet 359, 2002–2003. doi: 10.1016/S0140-6736(02)08836-0
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Mahenthiralingam, E., Urban, T. A., and Goldberg, J. B. (2005). The multifarious, multireplicon Burkholderia cepacia complex. Nat. Rev. Microbiol. 3, 144 –156. doi: 10.1038/nrmicro1085
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Marchler-Bauer, A., Zheng, C., Chitsaz, F., Derbyshire, M. K., Geer, L. Y., Geer, R. C.,et al. (2013). CDD: conserved domains and protein three-dimensional structure. Nucleic Acids Res. 41, D348–D352. doi: 10.1093/nar/gks1243
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Mayer-Hamblett, N., Ramsey, B. W., Kulasekara, H. D., Wolter, D. J., Houston, L. S., Pope, C. E.,et al. (2014). Pseudomonas aeruginosa phenotypes associated with eradication failure in children with cystic fibrosis. Clin. Infect. Dis. 59, 624–631. doi: 10.1093/cid/ciu385
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
McDonald, M. J., Gehrig, S. M., Meintjes, P. L., Zhang, X.-X., and Rainey, P. B. (2009). Adaptive divergence in experimental populations of Pseudomonas fluorescens. IV. Genetic constraints guide evolutionary trajectories in a parallel adaptive radiation. Genetics 183, 1041–1053. doi: 10.1534/genetics.109.107110
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
McDougald, D., Rice, S. A., Barraud, N., Steinberg, P. D., and Kjelleberg, S. (2012). Should we stay or should we go: mechanisms and ecological consequences for biofilm dispersal. Nat. Rev. Microbiol. 10, 39–50.
O’Connor, J. R., Kuwada, N. J., Huangyutitham, V., Wiggins, P. A., and Harwood, C. S. (2012). Surface sensing and lateral subcellular localization of WspA, the receptor in a chemosensory-like system leading to c-di-GMP production. Mol. Microbiol. 86, 720–729. doi: 10.1111/mmi.12013
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
O’Toole, G. A. (2011). Microtiter dish biofilm formation assay. J. Vis. Exp. 47, 2437. doi: 10.3791/2437
O’Toole, G., Kaplan, H. B., and Kolter, R. (2000). Biofilm formation as microbial development. Annu. Rev. Microbiol. 54, 49–79. doi: 10.1146/annurev.micro.54.1.49
Petrova, O. E., and Sauer, K. (2009). A novel signaling network essential for regulating Pseudomonas aeruginosa biofilm development. PLoS Pathog. 5:e1000668. doi: 10.1371/journal.ppat.1000668
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Poltak, S. R., and Cooper, V. S. (2011). Ecological succession in long-term experimentally evolved biofilms produces synergistic communities. ISME J. 5, 369–378. doi: 10.1038/ismej.2010.136
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Prigent-Combaret, C., Brombacher, E., Vidal, O., Ambert, A., Lejeune, P., Landini, P.,et al. (2001). Complex regulatory network controls initial adhesion and biofilm formation in Escherichia coli via regulation of the csgD gene. J. Bacteriol. 183, 7213–7223. doi: 10.1128/JB.183.24.7213-7223.2001
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Ryder, C., Byrd, M., and Wozniak, D. J. (2007). Role of polysaccharides in Pseudomonas aeruginosa biofilm development. Curr. Opin. Microbiol. 10, 644–648. doi: 10.1016/j.mib.2007.09.010
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Sawasdidoln, C., Taweechaisupapong, S., Sermswan, R. W., Tattawasart, U., Tungpradabkul, S., and Wongratanacheewin, S. (2010). Growing Burkholderia pseudomallei in biofilm stimulating conditions significantly induces antimicrobial resistance. PLoS ONE 5:e9196. doi: 10.1371/journal.pone.0009196
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Silva, I. N., Ferreira, A. S., Becker, J. D., Zlosnik, J. E., Speert, D. P., He, J.,et al. (2011). Mucoid morphotype variation of Burkholderia multivorans during chronic cystic fibrosis lung infection is correlated with changes in metabolism, motility, biofilm formation and virulence. Microbiology 157, 3124–3137. doi: 10.1099/mic.0.050989-0
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Spiers, A. J., and Rainey, P. B. (2005). The Pseudomonas fluorescens SBW25 wrinkly spreader biofilm requires attachment factor, cellulose fibre and LPS interactions to maintain strength and integrity. Microbiology 151, 2829–2839. doi: 10.1099/mic.0.27984-0
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Srivastava, D., and Waters, C. M. (2012). A tangled web: regulatory connections between quorum sensing and cyclic Di-GMP. J. Bacteriol. 194, 4485–4493. doi: 10.1128/JB.00379-12
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Starkey, M., Hickman, J. H., Ma, L., Zhang, N., De Long, S., Hinz, A.,et al. (2009). Pseudomonas aeruginosa rugose small colony variants have adaptations likely to promote persistence in the cystic fibrosis lung. J. Bacteriol. 191, 3492–503. doi: 10.1128/JB.00119-09
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Traverse, C. C., Mayo-Smith, L. M., Poltak, S. R., and Cooper, V. S. (2013). Tangled bank of experimentally evolved Burkholderia biofilms reflects selection during chronic infections. Proc. Natl. Acad. Sci. U.S.A. 110, E250–E259. doi: 10.1073/pnas.1207025110
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Travisano, M., Mongold, J. A., Bennett, A. F., and Lenski, R. E. (1995). Experimental tests of the roles of adaptation, chance, and history in evolution. Science 267, 87–90. doi: 10.1126/science.7809610
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Ueda, A., and Wood, T. K. (2009). Connecting quorum sensing, c-di-GMP, pel polysaccharide, and biofilm formation in Pseudomonas aeruginosa through tyrosine phosphatase TpbA (PA3885). PLoS Pathog. 5:e1000483. doi: 10.1371/journal.ppat.1000483
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Vial, L., Chapalain, A., Groleau, M. C., and Deziel, E. (2011). The various lifestyles of the Burkholderia cepacia complex species: a tribute to adaptation. Environ. Microbiol. 13, 1–12. doi: 10.1111/j.1462-2920.2010.02343.x
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Winsor, G., Khaira, B., Van Rossum, T., Lo, R., Whiteside, M., and Brinkman, F. (2008). The Burkholderia genome database: facilitating flexible queries and comparative analyses. Bioinformatics 24, 2803 –2804. doi: 10.1093/bioinformatics/btn524
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Zamorano, L., Moyà, B., Juan, C., Mulet, X., Blázquez, J., and Oliver, A. (2014). The Pseudomonas aeruginosa CreBC two-component system plays a major role in the response to β-Lactams, fitness, biofilm growth, and global regulation. Antimicrob. Agents Chemother. 58, 5084–5095. doi: 10.1128/AAC.02556-14
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Zlosnik, J. E., Costa, P. S., Brant, R., Mori, P. Y., Hird, T. J., Fraenkel, M. C.,et al. (2011). Mucoid and nonmucoid Burkholderia cepacia complex bacteria in cystic fibrosis infections. Am. J. Respir. Crit. Care Med. 183, 67–72. doi: 10.1164/rccm.201002-0203OC
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Zlosnik, J. E., Hird, T. J., Fraenkel, M. C., Moreira, L. M., Henry, D. A., and Speert, D. P. (2008). Differential mucoid exopolysaccharide production by members of the Burkholderia cepacia complex. J. Clin. Microbiol. 46, 1470–1473. doi: 10.1128/JCM.02273-07
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Keywords: biofilm, Burkholderia cepacia, dispersal, adaptation, evolution, exopolymeric substances, contingency
Citation: O’Rourke D, FitzGerald CE, Traverse CC and Cooper VS (2015) There and back again: consequences of biofilm specialization under selection for dispersal. Front. Genet. 6:18. doi: 10.3389/fgene.2015.00018
Received: 10 November 2014; Paper pending published: 03 December 2014; Accepted: 14 January 2015;
Published online: 11 February 2015.
Edited by:
Olivier Antoine Tenaillon, Institut National de la Santé et de la Recherche Médicale, FranceReviewed by:
Antonio Carvajal-Rodriguez, Universidad de Vigo, SpainIsabel Gordo, Instituto Gulbenkian de Ciência, Portugal
Copyright © 2015 O’Rourke, FitzGerald, Traverse and Cooper. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) or licensor are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Vaughn S. Cooper, Molecular, Cellular, and Biomedical Sciences, University of New Hampshire, 46 College Road, 212 Rudman Hall, Durham, NH, USA e-mail:dmF1Z2huLmNvb3BlckB1bmguZWR1