 Nathalie S. Nagalingum1*
Nathalie S. Nagalingum1* Nunzio Knerr2Shawn W. Laffan3
Nunzio Knerr2Shawn W. Laffan3 Carlos E. González-Orozco2,4
Carlos E. González-Orozco2,4 Andrew H. Thornhill2,5Joseph T. Miller2Brent D. Mishler6
Andrew H. Thornhill2,5Joseph T. Miller2Brent D. Mishler6- 1National Herbarium of New South Wales, Royal Botanic Gardens and Domain Trust, Sydney, NSW, Australia
- 2Centre for Australian National Biodiversity Research, Commonwealth Scientific and Industrial Research Organisation Plant Industry, Canberra, ACT, Australia
- 3Centre for Ecosystem Science, School of Biological, Earth and Environmental Sciences, University of New South Wales, Sydney, NSW, Australia
- 4Institute for Applied Ecology and Collaborative Research Network for Murray-Darling Basin Futures, University of Canberra, ACT, Australia
- 5Australian Tropical Herbarium, James Cook University, Cairns, QLD, Australia
- 6University and Jepson Herbaria, and Department of Integrative Biology, University of California, Berkeley, CA, USA
Because ferns have a wide range of habitat preferences and are widely distributed, they are an ideal group for understanding how diversity is distributed. Here we examine fern diversity on a broad-scale using standard and corrected richness measures as well as phylogenetic indices; in addition we determine the environmental predictors of each diversity metric. Using the combined records of Australian herbaria, a dataset of over 60,000 records was obtained for 89 genera to infer richness. A molecular phylogeny of all the genera was constructed and combined with the herbarium records to obtain phylogenetic diversity patterns. A hotspot of both taxic and phylogenetic diversity occurs in the Wet Tropics of northeastern Australia. Although considerable diversity is distributed along the eastern coast, some important regions of diversity are identified only after sample-standardization of richness and through the phylogenetic metric. Of all of the metrics, annual precipitation was identified as the most explanatory variable, in part, in agreement with global and regional fern studies. However, precipitation was combined with a different variable for each different metric. For corrected richness, precipitation was combined with temperature seasonality, while correlation of phylogenetic diversity to precipitation plus radiation indicated support for the species-energy hypothesis. Significantly high and significantly low phylogenetic diversity were found in geographically separate areas. These separate areas correlated with different climatic conditions such as seasonality in precipitation. The phylogenetic metrics identified additional areas of significant diversity, some of which have not been revealed using traditional taxonomic analyses, suggesting that different ecological and evolutionary processes have operated over the continent. Our study demonstrates that it is possible and vital to incorporate evolutionary metrics when inferring biodiversity hotspots from large compilations of data.
Introduction
Biodiversity hotspots occur because organisms have non-random patterns of distribution, and identifying and explaining these hotspots have long been of interest to biologists, ecologists, and biogeographers (Magurran and McGill, 2011). In the past, patterns have been identified based on anecdotal observations, but with the availability of large datasets and informatic tools, these patterns can now be inferred using analytical methods. Oftentimes these analytical approaches have confirmed previous observations, however, they have also revealed new patterns, particularly at continental or global scales (Currie and Paquin, 1987; Crisp et al., 2001; Francis and Currie, 2003; Hawkins et al., 2003; Kier et al., 2005, 2009; Kreft and Jetz, 2007; Kreft et al., 2010).
The most widely used measure of biodiversity is taxon diversity, also known as richness, which is a count of the number of terminal taxa (typically species but sometimes genera or other taxonomic levels) that occur in a particular region. Richness is calculated from occurrence data with geographic locations. However, richness estimates are influenced by sampling size and effort. Therefore, a number of metrics have been developed addressing these issues in order to obtain more accurate estimates (Magurran, 2004; Maurer and McGill, 2011).
Alternatively, there are measures of biodiversity that incorporate phylogeny with geography. These phylogenetic measures can be calculated for diversity and endemism, and are advantageous because use of a phylogeny takes into account evolutionary history (Faith, 1992; Moritz and Faith, 2002; Rosauer et al., 2009). Thus, phylogenetic measures provide an estimate of how much of the evolutionary history is represented in a particular region, and is referred to as phylogenetic diversity. Taxon-based measures use counts of species or genera as units, but in phylogenetic measures the units are the branch lengths connecting the terminal taxa in a region. Using a species as a terminal does not add substantially more branch length than using a genus, and thus, phylogenetic measures are not particularly sensitive to the taxonomic level chosen for analysis or to splitting and lumping of taxa (Rosauer et al., 2009). Identifying regions with high diversity is critical to conservation efforts, and the inclusion of phylogenetic indices can identify areas that standard metrics do not (Forest et al., 2007; Hendry et al., 2010; Winter et al., 2013).
Key to understanding distribution patterns is how they correlate with environmental conditions. Environmental variables can be climatic, such as rainfall, radiation, and temperature, or physical, such as soil type and topography. Numerous variables have been investigated in relation to richness, but rarely for phylogenetic indices. In several cases, the variables most strongly correlated with plant richness are mean annual temperature, water availability, and evapotranspiration, as well as topography (Currie and Paquin, 1987; Francis and Currie, 2003; Hawkins et al., 2003; Kreft and Jetz, 2007; Kreft et al., 2010). Although these variables are strong predictors of richness and may be causal, historical factors could also be responsible for current distribution patterns (Qian and Ricklefs, 2004; Wiens and Donoghue, 2004). Here we explore the relationships between richness, phylogenetic diversity, and environmental variables, and address the potential role of evolutionary history through the inclusion of phylogenetic indices.
Because ferns have a broad range of habitat preferences, spanning tropical rainforests to deserts, they are widely distributed and are therefore an ideal group for understanding how diversity is distributed. Most of our understanding of fern distribution derives from transects documenting richness and its relationship to environmental variables (Dzwonko and Kornaś, 1994; Lwanga et al., 1998; Tuomisto and Poulsen, 2000; Kessler, 2002a; Aldasoro et al., 2004). Fern richness is greatest in areas of high topographic relief and complexity, high evapotranspiration, and with many rain days (Kessler, 2010). Special focus has been given to the relationship between elevational gradients and richness, with richness peaking along mid-regions of slopes (suggesting the presence of the mid-domain effect) (Kessler, 2001, 2002b, 2010; Hemp, 2002; Kromer et al., 2005; Kluge and Kessler, 2006, 2011; Watkins, 2006; Kessler et al., 2011). However, a study compiling data from multiple altitudinal transects across the globe indicates that climate (water availability and temperature) rather than the mid-domain effect has better explanatory power for species richness (Kessler et al., 2011). Furthermore, the mid-domain effect may be more relevant to particularly high elevation regions such as in the Andean tropical regions, but may be less relevant in flat and vast continents such as Australia.
Overall, there are few synthetic studies of fern diversity and their relationship to the environment over large regions; although there are studies in the Iberian Peninsula, New Zealand, and Australia, but all focusing on richness alone (Lehmann et al., 2002; Bickford and Laffan, 2006; Moreno Saiz and Lobo, 2008). At broad scales, these studies found that water availability was correlated with regions of greatest richness, and also variably identified mean annual temperature, radiation, and topography (environmental heterogeneity) as important. At these scales, it is unclear what the corresponding patterns of phylogenetic diversity are, and how they relate to the environment.
Here we examine the patterns of fern diversity using richness and phylogenetic diversity indices. In particular we test for areas of significant randomized phylogenetic diversity against a null model. We also assess the patterns of diversity observed for correlations with environmental variables. We have chosen Australia as the study area because: the continent encompasses a broad range of habitat types, the unique availability of distributional data due to the efforts of Australia's Virtual Herbarium, and a completed floristic treatment of the ferns, which means that the taxonomy has largely been standardized across states (Australian Biological Resources Study, 1998).
Material and Methods
Geographic Data
Records of fern collections held in Australian herbaria are available in Australia's Virtual Herbarium (AVH) (http://avh.ala.org.au/) and were downloaded for this study. The download totaled 84,134 records. Using Google Refine version 2.5 (http://code.google.com/p/google-refine/), the dataset was cleaned to remove non-fern records (e.g., algae, lycophytes, and angiosperms), foreign collections (as well as Norfolk and Macquarie Islands), cultivated material, weeds, and garden escapees. The ferns were restricted to the Classes Marattiopsida and Polypodiopsida (Smith et al., 2006), and geographic ranges were examined to determine if there were potential misidentifications for specimens found outside of their known geographic range (Australian Biological Resources Study, 1998). The taxonomy was reconciled against a classification for extant ferns (Smith et al., 2006), for a total of 386 species in 89 genera and 25 families. Misspellings in the taxonomy were also corrected. Hybrids, doubtfully identified taxa (with either cf., aff., or ?), and specimens assigned only to genus-level were discarded. Records lacking geographic coordinates were excluded from the dataset, and latitude and longitude values of the remaining records were transformed into an Albers equal area coordinate system (European Petroleum Survey Group code EPSG3577). Following all of these spatial verification and cleaning steps, the dataset consisted of 63,230 records with greatest sampling in eastern Australia (Figure 1A).
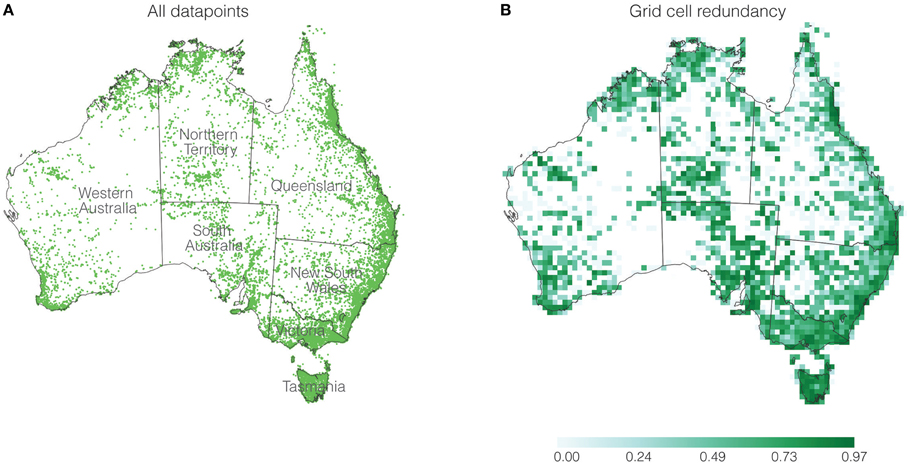
Figure 1. (A) Distribution of geographic records, and (B) redundancy for each grid cell. The scale bar represents redundancy values.
Species and genus richness were analyzed using grid cell sizes of 50 × 50 km and 100 × 100 km, which showed similar patterns (Figures S1A vs. S1B, S1C vs. S1D); thus we use the finer-scale, 50 × 50 km, cells for all of the analyses presented in order to minimize aggregation of the environmental data. Inspection of species vs. genus richness (Figures S1A vs. S1C) show that the patterns obtained are congruent. Therefore, the genus level results are presented herein because the phylogenetic diversity analyses were conducted at the genus level as there are currently inadequate molecular data to generate species-level phylogenies. A general relationship between species richness and higher taxon richness is found in many other studies as well (Williams and Gaston, 1994; Williams et al., 1997; Aldasoro et al., 2004; Currie and Francis, 2004).
Using the 50 × 50 km grid cell size resulted in a total of 1986 grid cells across Australia with at least one fern record. When the data were aggregated within the grid cells, the 63,230 records were reduced to 18,050 unique occurrences. As an example, the best-collected genus Cheilanthes comprised 6466 records but after aggregation into the 1986 grid cells there were 1320 unique occurrences. As the calculations only take into account occurrence in a cell, and not abundance values, there are multiple redundant records of the same taxon in each grid cell (Figure 1B). Redundancy, a measure of sampling quality, is scaled from zero to one (Garcillán and Ezcurra, 2003; Laffan et al., 2010). Redundancy values close to zero indicate possible under-sampling, while those close to one indicate well-sampled cells. Redundancy shows some variability across the continent, most closely linked to regions of population density (Figure 1B). There are some regions with values closer to zero, but since ferns are rarely found in these arid regions, the lack of duplicate sampling is likely not influencing the patterns we observe.
Molecular Data and Phylogenetic Analyses
All of the available sequences for each genus were downloaded from Genbank, regardless of species or geographic origin. Initially seven chloroplast markers were assessed for potential use in the phylogenetic analyses, but of these only three markers (atpA, atpB, and rbcL) were selected because they had the lowest amount of missing data. There were 420 sequences for atpA, 1117 sequences for atpB, and 2454 sequences for rbcL; the sequences were aligned using MAFFT version 6.860b (Katoh and Toh, 2008). Using each of these markers separately, a maximum likelihood phylogeny was constructed using GARLI (Genetic Algorithm for Rapid Likelihood Inference) version 0.951 (Zwickl, 2006), and set to terminate automatically using default parameters. In order to select one representative sequence for each genus, each of the three phylogenies was examined to determine that the sequences representing a genus were monophyletic. In cases where a genus was paraphyletic, this was typically due to an outlier sequence; thus, the phylogenetic position of the genus was checked against published phylogenies to verify that the sequence was indeed an outlier, and then it was discarded. Subsequently, two additional criteria were applied to select a representative sequence: first, a sequence was obtained from an Australian species, and second, if there were no Australian species, another species was selected if all three markers were available (or otherwise two markers).
New sequences were generated for seven genera not previously represented in Genbank, with GenBank accession numbers KP164480-KP164497 (Table S1). The following primers were used as amplification and sequencing primers for rbcL: ESRBCL1F, ESRBCL628F, ESRBCL654R, ESRBCL1361R; atpB: ESATPB172F, ESATPE45R; and atpA: ESATPF412F, ESATPF412F, ESTRNR46F, ESTRNR46F (the latter two as reverse primers) (Schuettpelz et al., 2006; Schuettpelz and Pryer, 2007). In addition, sequencing only primers were used for atpB: ATPB1163F, ATPB910R; and atpA: ESATPA856F, ESATPA877R (Schuettpelz et al., 2006; Schuettpelz and Pryer, 2007). PCR amplification was performed at 95°C for 10 min; 30 cycles of 94°C for 30 s, 64°C for 1 min, and 72°C for 45 s; and one cycle at 72°C for 5 min.
One representative sequence per marker for each of the 89 genera was assembled into a matrix for a total of 3893 base pairs. In this matrix, atpA and atpB were each missing sequences for four genera, and rbcL was complete. In this final matrix, the genus Actinostachys was incomplete at two of the three markers, Anogramma, Colysis, Cyclosorus, and Marattia were incomplete at one of three markers, and all other genera were complete for all three markers (Table S1). Molecular phylogenetic analysis of the partitioned, concatenated three markers were conducted using RAxML-HPC 7.3.2 using the CIPRES online portal (Miller et al., 2010). The supermatrix and resultant trees are deposited in TreeBase as study #15499. Overall, there was strong support for most of the nodes in the phylogeny (Figure S2), and the phylogenetic relationships were verified against a tree derived from the most well sampled and complete molecular dataset of ferns to date (Schuettpelz and Pryer, 2007).
To assess the impact of taxonomic changes on the results, we compared the PD values from this study to the PD values of an updated tree. In this updated tree, multiple recent taxonomic changes were incorporated: Doodia and Pteridoblechnum were omitted because they were synonymized into Blechnum (Perrie et al., 2014); the new genus Telmatoblechnum, which is a segregate of Blechnum, was included (Perrie et al., 2014); Oenotrichia and Coveniella were omitted because they had been synonymized into Lastreopsis (Labiak et al., 2014a,b); the new genus Parapolystichum, which is a segregate of Lastreopsis, was added (Labiak et al., 2014a,b); and Revwattsia was removed because it was synonymized into Dryopteris (McKeown et al., 2012). When we reanalyzed the data with the new tree, the range of PD values did not change significantly. In fact, when we subtracted the original PD value from the new PD value (calculated within the same grid cell) the differences ranged from −0.00000050 to 0.00000050, and the mean difference across all 1986 grid cells was −0.00000006. As a comparison, the PD values in this study were 0.01879–0.9308. It is clear, therefore, that recent taxonomic changes have little impact on the results.
Analysis of Diversity
The phylogeny derived from the three markers and the geographic data were imported into Biodiverse version 0.17 (Laffan et al., 2010). Several measures of diversity were calculated: species richness (SR), genus richness (GR), Margalef genus richness (MR), phylogenetic diversity (PD), and randomized phylogenetic diversity (PDrand).
The richness measures (SR and GR) are a direct count of the number of taxa occurring in each grid cell. To correct for uneven sampling effort among grid cells, the Margalef diversity metric is used. This metric standardizes richness across grid cells by dividing richness (R - 1) by the natural log of the number of samples (N) in a grid cell, thus RMargalef = (R − 1)/ln N (Magurran, 2004; Maurer and McGill, 2011).
Phylogenetic diversity (PD) measures the amount of the phylogenetic tree that is represented in a grid cell (Faith, 1992), which represents the sum of branch lengths in that grid cell. Specifically, this measure takes all of the terminal taxa that are found in one grid cell and sums the branches connecting them along a path to and including the root node. We depict PD as a proportion of the total tree length. PD is expected to be correlated with richness since with more terminal taxa there are more branch lengths to be summed (Tucker and Cadotte, 2013)—thus in rich areas the PD is expected to be larger. Indeed, we found that the GR and PD values within each grid cell are highly correlated; the generalized linear model (GLM) yielded an r2 equivalent of 0.895.
To test for statistical significance of the PD results, we used a randomization test (Mishler et al., 2014). First, all of the 18,050 unique occurrences (see “Geographic Data”) are pooled, and records from this pool are randomly assigned (without replacement) to the grid cells based on a constraint that the number of unique occurrences per grid cell are kept constant. This has the effect of keeping the number of terminal taxa per cell constant, for example, if a grid cell had 11 unique occurrences, then 11 unique occurrences from the pool were randomly assigned to that grid cell. In addition, the range size of each terminal taxon was kept constant. Second, PDrand is recalculated for each grid cell using the randomly assigned unique occurrences and the original phylogeny. These two steps yield new PDrand values for each grid cell. This process is repeated 999 times to give a distribution of PDrand values, and the original PDrand value is compared to the 999 PDrand values. If the original PD falls in the upper or lower 2.5% of the 1000 values, the PDrand of that cell is judged statistically significant. This method is similar to a randomly generated null community, known as null model 1 in the software Phylocom (Webb et al., 2008), or the null constrained model (Kembel and Hubbell, 2006). We recognize that this is one of several possible null models (Gotelli, 2000), each of which tests different questions, and is associated with different assumptions, but this model is appropriate for our objective to identify cells with significantly high or low PD values. Furthermore, we conducted an additional analysis where we standardized the PD value by the number of taxa (PD/richness), referred to as relative phylogenetic diversity (PDrel) by Davies et al. (2007). The PDrel randomization results are identical to the PD randomizations, indicating the robustness of the metrics; herein we refer to the PD randomizations only.
Environmental Predictors of Diversity
Eleven environmental variables were assessed against the diversity indices. These variables encompass temperature, precipitation, topography and substrates (Table 1). The 11 variables were selected because they are “independent” and representative of the predominant conditions. The climatic data were from selected BIOCLIM layers described and developed as part of ANUCLIM version 5.1 (Hutchinson et al., 2000). The soil layers were obtained from a database that was generated as part of a national survey (National Land and Water Resources Audit, 2002).
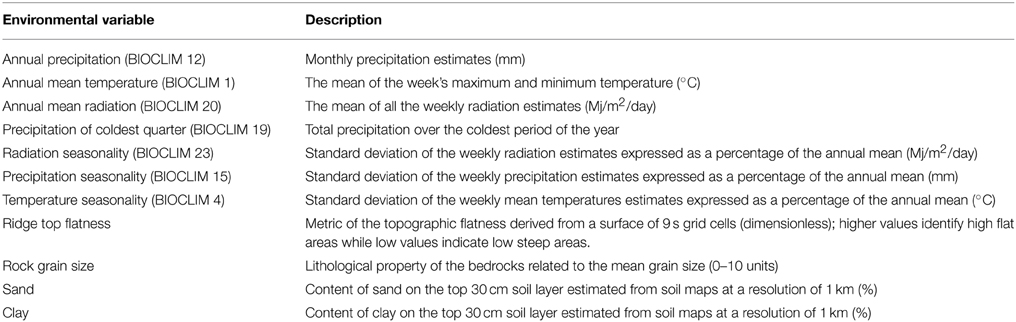
Table 1. Explanation of environmental variables used in this study, see Methodsd.
The predictors were tested against all of the indices as single variables (e.g., mean annual temperature). The top five single variables (based on the Akaike Information Criterion, AIC, see below) were examined further as interaction variables (e.g., mean annual temperature × topography), and as additive variables (e.g., mean annual temperature + topography). Some of the grid cells did not have environmental data and were excluded from the analyses, leaving 1913 grid cells in this part of the study. Most of these points were on the coast.
Generalized Linear Models
Because the indices do not have a normal distribution, standard linear models (LM) were not appropriate for analyzing their relationship to the 11 environmental variables. Based on examination of the data, a Poisson distribution best described the GR data, and a gamma distribution was most appropriate for the PD data. Consequently, generalized linear models (GLMs) were used because they allow for data that are not normally distributed, and a GLM was calculated for each variable and diversity metric with their appropriate distributions. A different approach was required for PDrand results since they are either significant or not, that is p ≤ 0.05. The statistically significant values were transformed into 1 and all other values were set to 0. These transformed data were then analyzed using GLMs with binomial distributions; the binomial is the only distribution that accepts categorical data, whereas all other distributions require continuous data. The higher and lower than expected PDrand were analyzed separately (high PDrand = 1 or low PDrand = 1). In total there were 40 high PDrand values and 84 low PDrand values. For each GLM, the following were recorded: AIC, percent of deviance explained (equivalent to r2), the statistical significance of the z-score of Wald's test, and whether the slope of the line was positive or negative. AIC values within 3 units of the best model were considered equally informative.
Detecting Spatial Autocorrelation Using Moran's I
To test whether there are biases due to spatial autocorrelation, Moran's I was calculated on the residuals of each standard linear model (LM). Spatial autocorrelation can yield misleading results because points that are close to each other will tend to share the same taxa, and are not independent (Dormann et al., 2007). Testing for spatial autocorrelation required a series of steps.
Firstly, a matrix was created that establishes which points are neighbors (referred to as a neighbors list), thus identifying the adjacent grid cells that could be affected by spatial autocorrelation. A distance criterion based on grid cell size was used to define the nearest neighbors. Since the grid cells were 50 × 50 km, all of the eight adjoining grids are in the radius of 75 km and so this latter value was set as the distance value for defining the nearest neighbors. Of the 1913 grid cells, 18 cells did not have neighbors within 75 km, and were subsequently excluded from the analyses (Figure S3). For the PDrand data, neighbors were defined at either 150 or 300 km because the radius of 75 km resulted in too few neighbors to draw any meaningful conclusions. There were 40 PDrand high values, and when using the smaller radius size, 15 of these points were deleted because there were no neighbors. In contrast, using the larger radius size required deletion of only four neighbors (Figures S4A,B). For the PDrand low analyses there were 84 values, and at the 150 km radius nine points were removed, while four were removed at the 300 km level (Figures S4C,D).
Secondly, weights were assigned to the neighbor relationships (via a spatial weights matrix). All of the relationships were given equal values of 1. Finally, the Moran's I global tests were run using the spatial weights matrix and a LM for each of the environmental variables and each of the indices. Moran's I values around zero indicate that there is no spatial autocorrelation in the residuals of the LMs and so neighbors have random values that are not linked. Moran's I values toward 1 indicate a positive correlation where adjoining grid cells are likely to share the same value (high-high; low-low); values toward -1 indicate a negative correlation suggesting that neighbors are more likely to have opposite values (low-high). The p-values of the Moran's I results were also recorded.
Accounting for Spatial Autocorrelation Using Spatial Autoregressive Models
To account for spatial autocorrelation, spatial autoregressive (SAR) models were used (Dormann et al., 2007; Kissling and Carl, 2007). There are three types of SARs, each of which account for autocorrelation via the addition of an extra term, a covariance matrix, to a standard linear model (LM). (1) SARlag is the lagged response model, which accounts for spatial autocorrelation by adding a term for spatial autocorrelation in the response variable (in this study, the richness indices), (2) SARmix is the lagged mixed model accounting for autocorrelations in both the indices and the environmental variables (the response and predictor variables), and (3) SARerr is a spatial error model that accounts for autocorrelation via an error term (i.e., in neither the indices nor the variables) (Dormann et al., 2007). For our data, the three SAR models (SARerr, SARlag, and SARmix) identified the same variables as the best models, however, SARmix occasionally yielded some differences from the other two models (not shown). Given the high error rate in SARmix identified in earlier studies, this result is not surprising and is likely to be a type I error (Dormann et al., 2007; Kissling and Carl, 2007). Tests of all three models indicate that SARerr has the least bias and error among all the SAR models (Kissling and Carl, 2007). Therefore, we present the results of SARerr here.
In the same steps as the calculations of the Moran's I, a neighbor list was constructed, followed by a spatial weights matrix; both were identical to that used for calculating the Moran's I. The SARs were then calculated using a LM for each of the environmental variables and each of the indices, together with the spatial weights matrix and the extra autocorrelation term. AICs were calculated for each model to determine which of the environmental variables best fitted each of the indices. The significance value for each SAR model was also recorded. SARs were calculated for all of the indices except for PDrand, which are statistical significance values and are therefore not appropriate for analysis using a LM.
The GLMs, Moran's I, and the SARs were calculated using the spdep package version 0.5–56 (Bivand, 2014) in R version 2.15.3 (R Core Team, 2014).
Results
Richness
The majority of fern richness is found along the eastern coast in Australia (Figure 2A). The greatest richness occurs in the Wet Tropics in Queensland with 80 genera (red-orange, Figure 2A), while the Border Ranges (between Queensland and New South Wales) is the second richest area with approximately 45 genera (yellow, Figure 2A). There is a continuous tract of low richness (approximately 30–40 genera, light blue Figure 2A) extending from Tasmania along the east coast of the continent. However, the vast majority of Australia has poor richness (less than 10 genera, dark blue, Figure 2A). Correcting for sampling using Margalef richness (MR) (Figure 2B) intensifies the patterns observed in uncorrected richness (Figure 2A). Namely, the east coast tract of low richness increases to medium richness with patches of medium-high richness. These patches correspond to the Border Ranges and Sydney Sandstone regions.
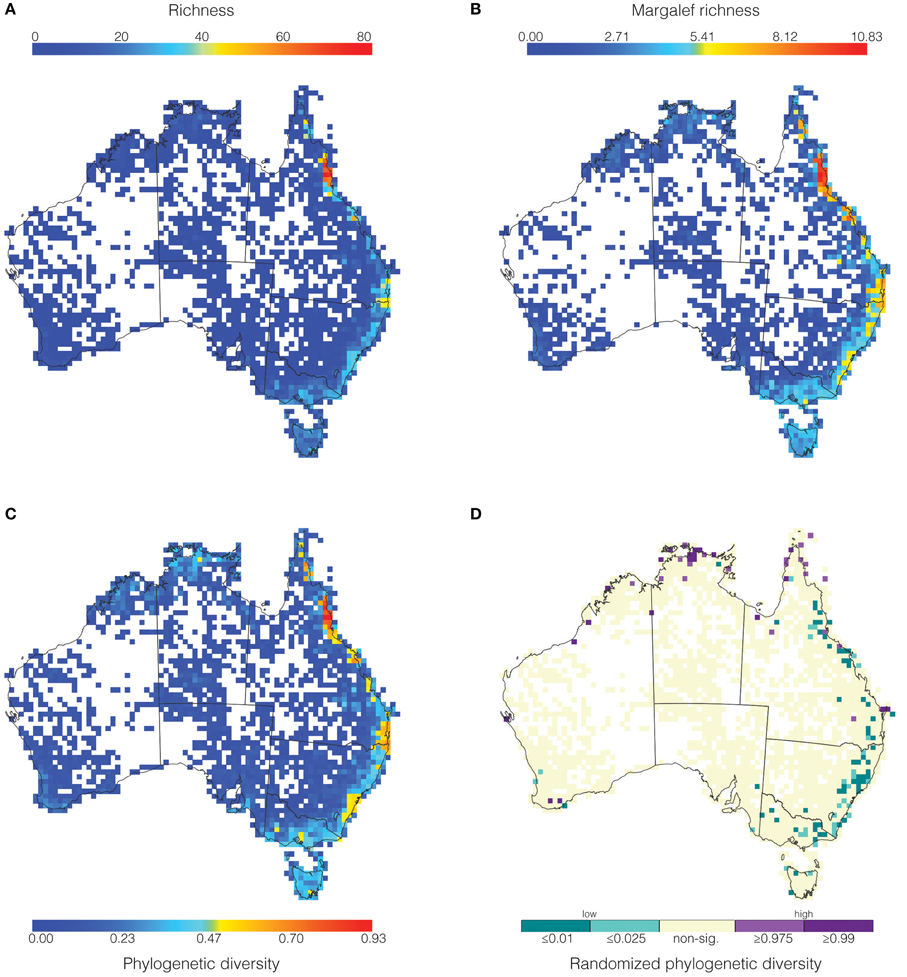
Figure 2. Maps showing grid cells values for (A) richness, (B) Margalef richness, (C) phylogenetic diversity (as a proportion of total phylogenetic diversity), and (D) significant phylogenetic diversity identified via randomization.
Phylogenetic Diversity
Phylogenetic diversity (PD) (Figure 2C) shows similar patterns to richness (Figure 2A), but most especially to MR (Figure 2B). In agreement with the two richness metrics, PD also shows the Wet Tropics as the greatest hotspot with 90% (0.90) of the phylogeny represented. Notable similarities between MR and PD are the medium-high values in the Border Ranges and the Sydney Sandstone regions, with about 50–60% of the tree present for PD (Figure 2C). Also, the Northern Territory and Tasmania have corresponding regions of low MR and low PD (Figures 2B,C light blue). As for the richness metrics, the overwhelming pattern is that most of the continent has poor PD (dark blue, Figure 2C), and that the east coast hosts most of the fern PD (light blue-yellow-orange-red, Figure 2C).
Statistically Significant Phylogenetic Diversity
The phylogenetic diversity randomizations showed that cells with significantly high PDrand were mostly separate from those with low PDrand (Figure 2D). The high PDrand regions are concentrated toward the north of Australia, in the most northern parts of the Northern Territory and Queensland (purples, Figure 2D), whereas the low PDrand regions are aggregated along the east coast (teals, Figure 2D). We note that PDrand and PD standardized by richness (not shown) yielded identical results, indicating that PDrand is not sensitive to sampling bias.
The significantly high PDrand cells are characterized by the overrepresentation of taxa with long branches representing disparate clades of the phylogeny. Principally these are the combination of a long-branched early diverging lineage (Schizaeales) and a long-branched derived family, Pteridaceae. The genus Lindsaea is typically found in these cells, and in some cells the early diverging lineages Salviniales and Gleichneiales are present too. Alternatively, significantly high PDrand cells can have one taxon representing a particularly long branch, either Acrostichum or Ceratopteris. Conversely, the significantly low PDrand cells can have one taxon with an especially short branch, for example Abrodictyum in the early diverging Hymenophyllaceae. Examination of the taxonomic composition of the cells that have significantly low PDrand reveals three additional ways in which low PDrand can arise. First, there are few taxa in that cell, and they are all restricted to one family, such as Pteridaceae. Second, there is a moderate number of taxa but they are restricted to one clade, the tree fern + polypod clade, with the early diverging lineages not represented. Thirdly, again there is moderate to high numbers of taxa, but here the early diverging lineages are represented, while the eupolypods I (composed of Nephrolepidaceae, Lomariopsidaceae and Polypodiaceae) are missing.
Predictors
When the indices were correlated with the environmental variables, the GLMs for richness and PD indicated that annual precipitation together with mean annual radiation, as additive or interactive variables, is the best predictor (Table 2). In the case of Margalef richness, the GLM with the lowest AIC was annual precipitation together with seasonality in temperature. However, Moran's I values from 0.48 to 0.59 show that these results are biased by spatial autocorrelation (Table 2). When spatial autocorrelation is taken into account using SARs, the best predictor for richness changes to annual precipitation by topography (ridge top). For both Margalef richness and PD, the models recovered using GLM and SAR are identical. All of the best performing models are explained by a positive interaction, and all included a term for annual precipitation, indicating the importance of water in determining the diversity of ferns (Table 2). Regardless of the metric used, the two-factor models outperformed all of the single-factor models.
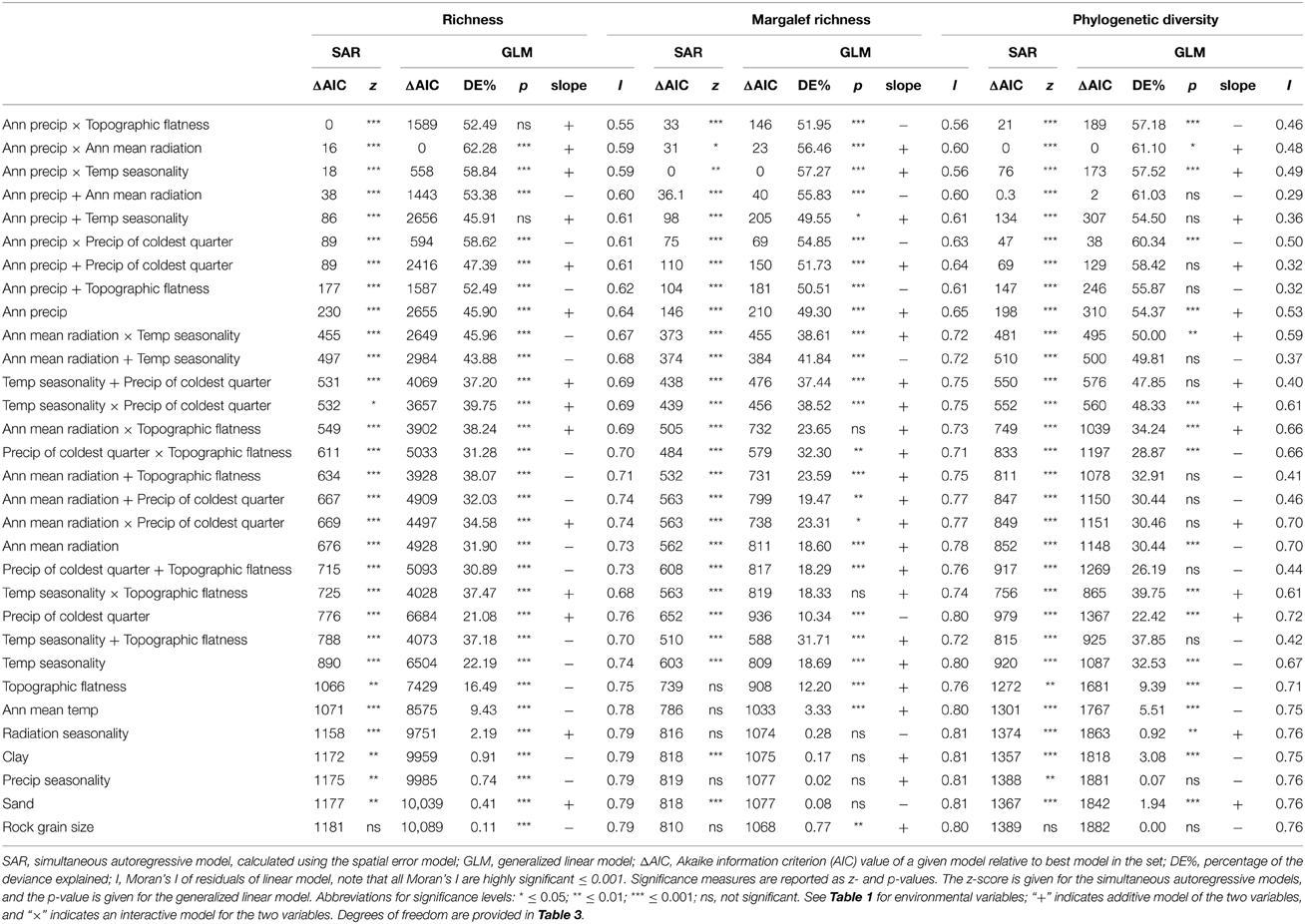
Table 2. Statistical relationships between environmental variables and the richness, Margalef richness and phylogenetic diversity metrics.
The randomizations of PD are explained by a different set of environmental factors compared to the other indices (Table 3). In addition, high PDrand and low PDrand are each explained by different environmental factors. The high PDrand areas correlate with mean annual radiation added/by temperature seasonality (0 compared to 2 ΔAIC respectively, Table 3), and a close model was temperature seasonality plus precipitation in the coldest quarter (3 ΔAIC, Table 3). Overall, the highest performing models for high PDrand all had temperature seasonality in common. The low PDrand areas also correlate with temperature seasonality, but by annual precipitation. For 150 or 300 km values at which neighbors were defined, Moran's I values were all close to zero indicating that spatial autocorrelation was not present in these datasets (Table 3).
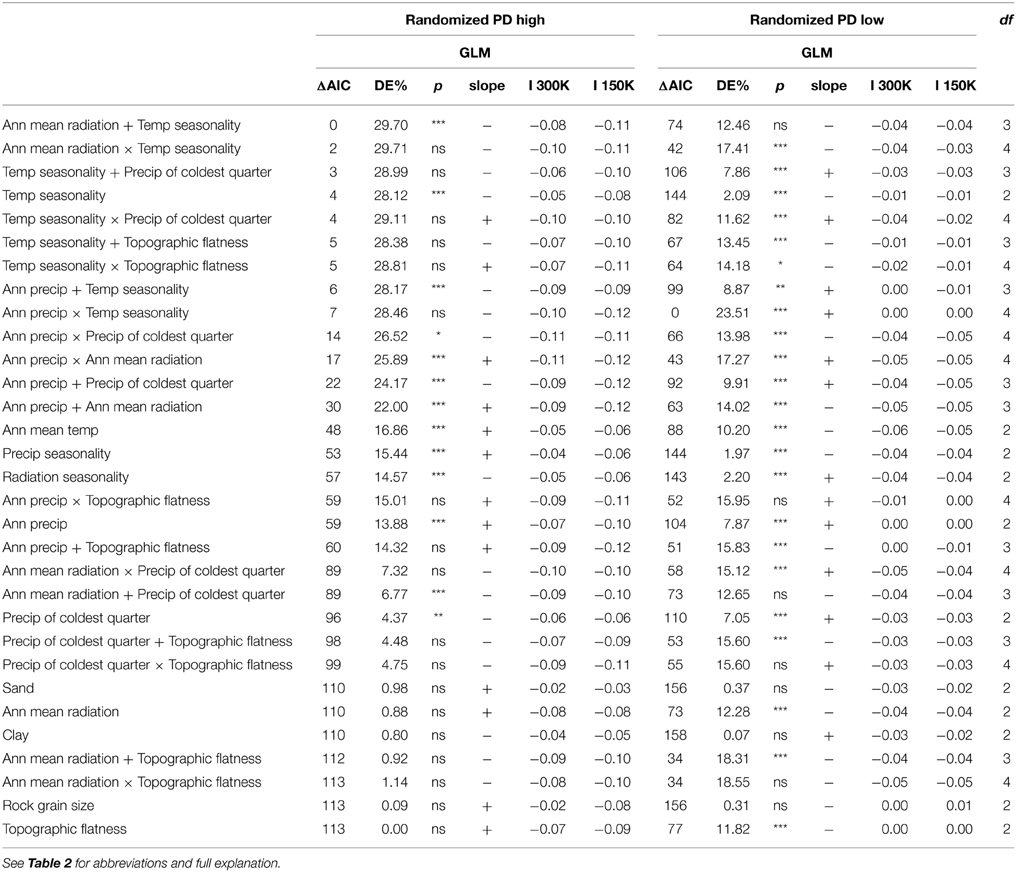
Table 3. Statistical relationships between environmental variables and PDrand, randomized phylogenetic diversity, with higher (high PDrand) or lower (low PDrand) than expected PD for statistically significant cells only.
Discussion
Comparing Diversity Metrics and Hotspots
Using the largest fern dataset assembled to date, we find that the Wet Tropics is the most diverse region identified using three different diversity metrics (red cells in Figures 2A–C). When richness is compared to PD there are regions that have greater phylogenetic diversity than richness. However, when uneven sampling effort is accounted for, PD and richness (measured as Margalef richness) are generally in agreement. Additional regions with significant diversity are the Border Ranges and Sydney Sandstone, both in coastal eastern Australia (light blue in Figure 2A vs. orange-yellow in Figure 2C), and Kakadu–Alligator Rivers in the Northern Territory, the Kimberly region in northwest Western Australia, southwest Western Australia, and Tasmania (dark blue in Figure 2A vs. light blue-yellow in Figure 2C).
The disparity between uncorrected richness and phylogenetic diversity indicates that richness is not necessarily predictive of phylogenetic diversity. Studies elsewhere have shown a disparity between richness and phylogenetic diversity, sometimes when sampling has been accounted for (Davies et al., 2007; Forest et al., 2007; Huang et al., 2011). Such disparity emphasizes that caution is needed when using richness alone as a metric of diversity. Thus, we focus on Margalef richness results because the sampling bias has been corrected. From a conservation viewpoint, diversity must be assessed using sample standardization for richness as well as using phylogenetic diversity. Regions with high phylogenetic diversity can harbor unrecognized diversity value (Moritz and Faith, 2002; Rosauer and Mooers, 2013), and may be culturally and medicinally useful too (Forest et al., 2007).
In an earlier study, fern richness across Australia was documented at the species scale and using a dataset approximately half the size of the present study (Bickford and Laffan, 2006). Differences in the datasets are most evident in the increased sampling in inland regions. Regardless of the dataset size (Figure S1A our study, vs. Figure 2A, Bickford and Laffan, 2006), or the taxonomic level (Figure 1A our study vs. Figure S1A our study), there remains greatest richness along the east coast of Australia with a substantial hotspot in the Wet Tropics. This Wet Tropics hotspot was earlier detected for a dataset of vascular plants, comprising principally of angiosperms (Crisp et al., 2001). However, other significant angiosperm hotspots, including the significant southwest Western Australia hotspot, are not shared with ferns (Crisp et al., 2001; González-Orozco et al., 2011, 2014; Schmidt-Lebuhn et al., 2012; Kooyman et al., 2013); this is not surprising given the more arid conditions in these latter hotspots, as well as the dependence on water for ferns during the reproductive phase of their life cycle, and preference for moist conditions. The inconsistency among hotspots of various floristic groups indicates that hotspots need to be inferred on a case-by-case basis. These may be the result of the dissimilar ecological preferences as well as different diversification histories. Interestingly, two angiosperm hotspots, the Border Ranges and Sydney Sandstone, are only observed as fern hotspots when using Margelef richness and phylogenetic diversity. On the other hand, liverworts and mosses show greatest diversity along the east coast (Stevenson et al., 2012), largely matching the pattern seen in the ferns (Nagalingum et al., 2014). These corresponding patterns likely reflect the more critical requirement for water of all of these seed-free plant groups, and its greater availability in these regions.
Explaining the Distribution of Diversity
Differences in diversity distribution have been attributed to a variety of mechanisms. In ferns, the overwhelming pattern is the near-absence of diversity in the central arid interior of the continent, and a hotspot in the Wet Tropics. The Wet Tropics likely represents the ancestral niche for ferns, whereas survival in arid biomes require a suite of adaptations that have arisen in only two of the 89 genera examined here. This pattern fits the ancestral niche hypothesis that predicts that greater diversity will be present in an ancestral niche because more taxa have accumulated here as the group has remained in that niche; at the same time, the group is unable to disperse to other niches without the evolution of suitable adaptations (Crisp et al., 2009; Wiens et al., 2010). In addition, a range of historical factors has been used to explain the previously observed greater diversity in the tropics. These range from elevated speciation rates and decreased extinction rates (compared to extra-tropical regions; the cradle-museum hypothesis), a greater geographical extent of tropical rainforests in the past (the area effect), and an older age of the tropics (the age effect) (Qian and Ricklefs, 2004; Wiens and Donoghue, 2004). Although we were not able to conduct a detailed examination of traits and historical factors with the present dataset, it is likely that both have shaped the distribution patterns we observed for the ferns.
Biodiversity hotspots and underlying differences in diversity distribution are commonly linked to current climatic and environmental conditions—although this has led to discussion between the role of current versus historical factors (Currie and Paquin, 1987; Latham and Ricklefs, 1993; Qian and Ricklefs, 2000; Francis and Currie, 2003; Ricklefs, 2004). Regardless of the scale, from global modeling studies (Kreft and Jetz, 2007; Kreft et al., 2010) to regional meta-analyses (Lehmann et al., 2002; Bickford and Laffan, 2006), plot-based elevational transects (Kessler, 2000, 2001; Bhattarai et al., 2004; Grytnes and Beaman, 2006; Kluge et al., 2006; Kessler et al., 2011) and regional plot-surveys (Aldasoro et al., 2004), water is the leading determinant for fern richness. Indeed, all of the best performing models in our analyses show that the two richness metrics as well as phylogenetic diversity are explained by annual precipitation combined with a different variable (Table 2). At a global scale, water availability combined with elevation and temperature are most important for richness of pteridophytes (ferns and lycophytes), and also for plants in general (Kreft and Jetz, 2007; Kreft et al., 2010).
A global analysis of pteridophytes indicates that greatest richness occurs in wet tropical regions with increasing elevation, referred to as topographic complexity (Kessler, 2010; Kreft et al., 2010). The role of elevation is confirmed in our analyses with topography combined with annual precipitation best explaining fern richness (but we note that topography is not included the best-performing model in corrected richness and phylogenetic diversity; Table 2). Other regional analyses have also demonstrated the importance of elevation (Dzwonko and Kornaś, 1994; Ferrer-Castan and Vetaas, 2005). It is thought that topography contributes to greater richness because increasing elevations are associated with variability in substrates (including microhabitats), climate, and environment, all yielding a greater diversity of niches for more taxa (Aldasoro et al., 2004; Moran, 2008; Moreno Saiz and Lobo, 2008; Kessler, 2010; Kessler et al., 2011). However, topography alone does not necessarily predict hotspots. In New Zealand, richness is greatest in the North Island (Lehmann et al., 2002), which is considerably less mountainous than the South Island. Instead, the North Island hotspot is associated with the warmest climate zone. At particularly high elevations, the elevational-richness relationship is not observed likely because frost limits fern diversity (Kessler, 2000, 2001; Lehmann et al., 2002; Bhattarai et al., 2004; Grytnes and Beaman, 2006; Kluge et al., 2006; Kessler et al., 2011).
Regions with greater Margalef richness are explained by increases in both precipitation and seasonal temperature extremes. This is in contrast to uncorrected richness, which correlates to increases in precipitation and topography. The difference suggests that sampling biases affect the richness results. These biases may be due to greater sampling effort in topographically complex areas, and the removal of the poorly sampled cells (via the Margalef calculation). The relationship between seasonal temperatures in fern diversity is surprising compared to other findings for ferns (Kessler, 2010) and to trees (Currie and Paquin, 1987). However, seasonality has been identified as a mechanism that enables plants requiring different niches to co-occur (Pausas et al., 2003).
The positive relationship between radiation and phylogenetic diversity suggests that available energy is a limiting factor, conforming to the species-energy hypothesis. Several studies have concluded that plant productivity controls richness (Currie, 1991 and references therein), and for ferns, the link between productivity and species richness has been mechanistically attributed to competition and niche availability (Kessler et al., 2014). However, the mechanisms explaining fern phylogenetic diversity and radiation are unresolved. It has been observed that there is a predominance of epiphytes in tropical rainforests (Gentry and Dodson, 1987; Gentry, 1992). Thus, with low light availability in tropical rainforests, perhaps phylogenetic breadth is limited to epiphytes (which occur in only a few fern clades, Schuettpelz and Pryer, 2009); whereas with greater radiation there is more light available to support a broader range from epiphytic to terrestrial forms. Overall, the relationship between plant phylogenetic diversity and environment is poorly understood with the exception of few analyses (Williams et al., 2010). It remains to be determined if radiation is important for other plant groups, and if radiation is causally linked to phylogenetic diversity.
Interestingly, we found different environmental factors each explaining richness, corrected richness, and phylogenetic diversity. As noted above annual precipitation is common to all of the best models—for richness it combines with topography, for corrected richness it interacts with temperature seasonality, while for phylogenetic diversity it combines with mean annual radiation (Table 2). In general, assessment of fern diversity with environmental factors is conducted for single variables, which unsurprisingly identifies precipitation/humidity as the most important factor (Aldasoro et al., 2004; Bickford and Laffan, 2006; Kessler, 2010). Using two variables, we find that models with single variables were always outperformed by those with two variables (Table 2). We also employed spatial autoregression, which accounted for non-independence of the data by incorporating a spatial term into the models (Dormann et al., 2007; Kissling and Carl, 2007). The models for Margalef richness and phylogenetic diversity were identical using a general linear model and the spatial autoregression model; however, the models differed for richness (Table 2). This result indicates that multivariate models as well as spatial terms need to be considered when inferring the relationship with diversity and environment.
Many fern species distributions are shaped by rock and soil types (Kessler, 2010 and references therein), however, our analyses indicate that fern diversity hotspots are not related to rock and soil substrate (Table 2). In fact, rock grain size, sand, and clay all performed extremely poorly in our models. On the other hand, soil fertility (measured as C/N ratio) best explained fern richness for plots across Uganda (Lwanga et al., 1998). It is possible that fertility may have a strong relationship to richness at the continental-scale, but soil nutrient profiles have been difficult to obtain for larger areas, and future studies are needed to further examine the relationship (Kessler, 2010).
Differences in Phylogenetic Representation Over the Landscape
As discussed above, richness is not necessarily predictive of phylogenetic diversity, despite their general correlation. In this study, we used a randomized phylogenetic diversity test to identify quantitatively places where the expected correlation does not hold. This method discerns whether it has a significantly higher or lower representation of the phylogeny compared to a random sampling of the same number of taxa. In the case of significantly high PDrand (randomized phylogenetic diversity) there is more of the phylogeny represented in that grid cell than expected; this is otherwise known as “phylogenetic overdispersion.” Possible explanations include ecological competition that excludes close relatives, or biogeographic processes creating refugia (Webb et al., 2002; Hennequin et al., 2014). Alternatively, when there is significantly low PDrand there is less of the phylogeny represented than expected, and this is referred to as “phylogenetic clustering.” Possible explanations include ecological filtering where close relatives have the same habitat requirements, or evolutionarily recent radiations (Webb et al., 2002; Hennequin et al., 2014).
It is also important to note that PD is not necessarily predictive of significant PDrand. For example, some regions, such as the Wet Tropics, are high in PD yet significantly low in PDrand (perhaps due to ecological factors as discussed below). Conversely, some areas, such as the Kakadu–Alligator Rivers region in the Northern Territory, have only low to moderate levels of PD, but significantly high PDrand (perhaps due to ecological and biogeographical factors as discussed below). The ability to detect such regions with unusually low or high values of PD shows the value of the PDrand test used here.
We found that there are two strong geographic patterns in the distribution of high and low PDrand cells. The high PDrand cells are clustered in the north of the continent (Figure 2D, purples), whereas the low PDrand cells are concentrated along the east coast (Figure 2D, teals). Given the strong geographic pattern, it is not surprising that there are differences in the environmental conditions between these two regions (Table 3). High PDrand regions are negatively associated with seasonality in temperature, which means that high PDrand is linked to temperature stability. Equable, stable temperatures may be more favorable to a wider range of lineages across the phylogeny because they do not need to evolve adaptations to extremes in temperature.
Significantly low PDrand is positively associated with annual precipitation and temperature seasonality. The latter is indicative of extremes in temperature, however, not all fern clades are able to tolerate cold conditions. Indeed in low PDrand grid cells families that prefer warmer temperatures are absent, such as the Polypodiaceae (with the exception of Grammatis, which occur in less than one-quarter of low PDrand cells); while families that are cold tolerant, such as the Blechnaceae and Dennstaedtiaceae are present. Alternatively, the absence of Polypodiaceae may be due to the absence of suitable host trees for this largely epiphytic group. It is also possible that such extremes in conditions limit dispersal due to the absence of suitable traits (Zanne et al., 2014), which for ferns include easily dispersed spores and underground rhizomes, and also promote extinction, although extinction may not necessarily be related to temperature.
Concluding Remarks
Using the largest fern dataset to date, we have identified several areas as hotspots for richness, phylogenetic diversity, and significantly high or low randomized phylogenetic diversity across Australia. Notably, the use of several metrics identifies different or additional areas of importance. The PDrand measure identifies novel areas of diversity significance compared to the two other metrics we used, and these regions have not been revealed in any other analyses. We suggest that these areas are of particular evolutionary and conservation importance and a detailed analysis of them is needed in future studies. Notably environmental predictors explain the distribution of various hotspots, and in turn, the different metrics are predicted by different environmental variables. However, with the onset of changing climate, the conditions that support greatest diversity today will change in the future, e.g., increased and decreased rainfall across Australia (Bureau of Meteorology and CSIRO, 2014), and will likely impact species distributions and thus, diversity patterns.
We note that these broad scale continental-level patterns were obtained using digitized herbarium records. With increasing digitization efforts across the globe we will be able to conduct even larger scale analyses (without having to rely on modeled distributions, Lehmann et al., 2002; Kreft and Jetz, 2007) as has been performed for global marine distribution records (Tittensor et al., 2010). Furthermore, our study is a proof-of-concept that it is possible and vital to incorporate evolutionary metrics when inferring biodiversity hotspots from large compilations of data.
Author Contributions
NSN and BDM designed the study. NSN, NK, and AHT assembled the data. NSN, NK, SWL, and CEG-O analyzed the data. NSN prepared the manuscript, and all authors edited the manuscript.
Conflict of Interest Statement
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Acknowledgments
We thank Carolyn Connelly from the Royal Botanic Garden and Domain Trust for assistance with laboratory techniques, Nada Sankowsky for providing fern specimens, and CSIRO (Australia) for a Distinguished Visiting Scientist Award to BDM. Brian Enquist, Sabine Hennequin, Paul Manos, Michelle McMahon, Michael Sanderson and Maurizio Rossetto are thanked for helpful discussions. The reviewers are thanked for their comments on the manuscript.
Supplementary Material
The Supplementary Material for this article can be found online at: http://www.frontiersin.org/journal/10.3389/fgene.2015.00132/abstract
References
Aldasoro, J. J., Cabezas, F., and Aedo, C. (2004). Diversity and distribution of ferns in sub-Saharan Africa, Madagascar and some islands of the South Atlantic. J. Biogeogr. 31, 1579–1604. doi: 10.1111/j.1365-2699.2004.01106.x
Australian Biological Resources Study. (1998). “Ferns, gymnosperms and allied groups,” in Flora of Australia, Vol. 48, ed P. M. McCarthy (Melbourne, VIC: CSIRO), 766.
Bhattarai, K. R., Vetaas, O. R., and Grytnes, J. A. (2004). Fern species richness along a central Himalayan elevational gradient, Nepal. J. Biogeogr. 31, 389–400. doi: 10.1046/j.0305-0270.2003.01013.x
Bickford, S. A., and Laffan, S. W. (2006). Multi-extent analysis of the relationship between pteridophyte species richness and climate. Global Ecol. Biogeogr. 15, 588–601. doi: 10.1111/j.1466-8238.2006.00250.x
Bivand, R. (2014). spdep: Spatial Dependence: Weighting Schemes, Statistics and Models. Available online at: http://CRAN.R-project.org/package=spdep
Crisp, M. D., Arroyo, M. T. K., Cook, L. G., Gandolfo, M. A., Jordan, G. J., Mcglone, M. S., et al. (2009). Phylogenetic biome conservatism on a global scale. Nature 458, 754–756. doi: 10.1038/nature07764
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Crisp, M. D., Laffan, S., Linder, H. P., and Monro, A. (2001). Endemism in the Australian flora. J. Biogeogr. 28, 183–198. doi: 10.1046/j.1365-2699.2001.00524.x
Currie, D. J. (1991). Energy and large-scale patterns of animal-and plant-species richness. Am. Nat. 27–49. doi: 10.1086/285144
Currie, D. J., and Francis, A. P. (2004). Regional versus climatic effect on taxon richness in angiosperms: reply to Qian and Ricklefs. Am. Nat. 163, 780–785. doi: 10.1086/383596
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Currie, D. J., and Paquin, V. (1987). Large-scale biogeographical patterns of species richness of trees. Nature 329, 326–327. doi: 10.1038/329326a0
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Davies, R. G., Orme, C. D. L., Webster, A. J., Jones, K. E., Blackburn, T. M., and Gaston, K. J. (2007). Environmental predictors of global parrot (Aves: Psittaciformes) species richness and phylogenetic diversity. Global Ecol. Biogeogr. 16, 220–233. doi: 10.1111/j.1466-8238.2007.00282.x
Dormann, C., McPherson, J. M., Araújo, M. B., Bivand, R., Bolliger, J., Carl, G., et al. (2007). Methods to account for spatial autocorrelation in the analysis of species distributional data: a review. Ecography 30, 609–628. doi: 10.1111/j.2007.0906-7590.05171.x
Dzwonko, Z., and Kornaś, J. (1994). Patterns of species richness and distribution of pteridophytes in Rwanda (Central Africa): a numerical approach. J. Biogeogr. 21, 491–501. doi: 10.2307/2845653
Faith, D. P. (1992). Conservation evaluation and phylogenetic diversity. Biol. Conserv. 61, 1–10. doi: 10.1016/0006-3207(92)91201-3
Ferrer-Castan, D., and Vetaas, O. R. (2005). Pteridophyte richness, climate and topography in the Iberian Peninsula: comparing spatial and nonspatial models of richness patterns. Global Ecol. Biogeogr. 14, 155–165. doi: 10.1111/j.1466-822X.2004.00140.x
Forest, F., Grenyer, R., Rouget, M., Davies, T. J., Cowling, R. M., Faith, D. P., et al. (2007). Preserving the evolutionary potential of floras in biodiversity hotspots. Nature 445, 757–760. doi: 10.1038/nature05587
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Francis, A. P., and Currie, D. J. (2003). A globally consistent richness-climate relationship for angiosperms. Am. Nat. 161, 523–536. doi: 10.1086/368223
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Garcillán, P. P., and Ezcurra, E. (2003). Distribution and species richness of woody dryland legumes in Baja California, Mexico. J. Veg. Sci. 14, 475–486. doi: 10.1111/j.1654-1103.2003.tb02174.x
Gentry, A. H. (1992). Tropical forest biodiversity: distributional patterns and their conservational significance. Oikos 19–28. doi: 10.2307/3545512
Gentry, A. H., and Dodson, C. H. (1987). Diversity and biogeography of neotropical vascular epiphytes. Ann. Mo. Bot. Gard. 74, 205–233. doi: 10.2307/2399395
González-Orozco, C. E., Laffan, S. W., and Miller, J. T. (2011). Spatial distribution of species richness and endemism of the genus Acacia in Australia. Aust. J. Bot. 59, 601–608. doi: 10.1071/BT11112
González-Orozco, C. E., Thornhill, A. H., Knerr, N. J., Laffan, S. W., and Miller, J. T. (2014). Biogeographical regions and phytogeography of the eucalypts. Divers. Distrib. 20, 46–58. doi: 10.1111/ddi.12129
Gotelli, N. J. (2000). Null model analysis of species co-occurrence patterns. Ecology 81, 2606–2621. doi: 10.1890/0012-9658(2000)081[2606:NMAOSC]2.0.CO;2
Grytnes, J.-A., and Beaman, J. H. (2006). Elevational species richness patterns for vascular plants on Mount Kinabalu, Borneo. J. Biogeogr. 33, 1838–1849. doi: 10.1111/j.1365-2699.2006.01554.x
Hawkins, B. A., Field, R., Cornell, H. V., Currie, D. J., Guégan, J. F., Kaufman, D. M., et al. (2003). Energy, water, and broad-scale geographic patterns of species richness. Ecology 84, 3105–3117. doi: 10.1890/03-8006
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Hemp, A. (2002). Ecology of the pteridophytes on the southern slopes of Mt. Kilimanjaro–I. Altitudinal distribution. Plant Ecol. 159, 211–239. doi: 10.1023/A:1015569125417
Hendry, A. P., Lohmann, L. G., Conti, E., Cracraft, J., Crandall, K. A., Faith, D. P., et al. (2010). Evolutionary biology in biodiversity science, conservation, and policy: a call to action. Evolution 64, 1517–1528. doi: 10.1111/j.1558-5646.2010.00947.x
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Hennequin, S., Kessler, M., Lindsay, S., and Schneider, H. (2014). Evolutionary patterns in the assembly of fern diversity on the oceanic Mascarene Islands. J. Biogeogr. 41, 1651–1663. doi: 10.1111/jbi.12339
Huang, J., Chen, B., Liu, C., Lai, J., Zhang, J., and Ma, K. (2011). Identifying hotspots of endemic woody seed plant diversity in China. Divers. Distrib. 18, 673–688. doi: 10.1111/j.1472-4642.2011.00845.x
Hutchinson, M. F., Houlder, D. J., Nix, H. A., and McMahon, J. P. (2000). Hutchinson: ANUCLIM Users Guide Version 5.1. Canberra: Centre for Resource and Environmental Studies, Australian National University.
Katoh, K., and Toh, H. (2008). Recent developments in the MAFFT multiple sequence alignment program. Brief. Bioinform. 9, 286–298. doi: 10.1093/bib/bbn013
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Kembel, S. W., and Hubbell, S. P. (2006). The phylogenetic structure of a neotropical forest tree community. Ecology 87, S86–S99. doi: 10.1890/0012-9658(2006)87[86:TPSOAN]2.0.CO;2
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Kessler, M. (2010). “Biogeography of ferns,” in Fern Ecology, eds K. Mehltreter, L. R. Walker, and J. M. Sharpe (Cambridge: Cambridge University Press), 23–60.
Kessler, M. (2000). Elevational gradients in species richness and endemism of selected plant groups in the central Bolivian Andes. Plant Ecol. 149, 181–193. doi: 10.1023/A:1026500710274
Kessler, M. (2001). Pteridophyte species richness in Andean forests in Bolivia. Biodivers. Conserv. 10, 1473–1495. doi: 10.1023/A:1011811224595
Kessler, M. (2002a). Range size and its ecological correlates among the pteridophytes of Carrasco National Park, Bolivia. Global Ecol. Biogeogr. 11, 89–102. doi: 10.1046/j.1466-822X.2002.00176.x
Kessler, M. (2002b). The elevational gradient of Andean plant endemism: varying influences of taxon-specific traits and topography at different taxonomic levels. J. Biogeogr. 149, 1159–1165. doi: 10.1046/j.1365-2699.2002.00773.x
Kessler, M., Kluge, J., Hemp, A., and Ohlemüller, R. (2011). A global comparative analysis of elevational species richness patterns of ferns. Global Ecol. Biogeogr. 20, 868–880. doi: 10.1111/j.1466-8238.2011.00653.x
Kessler, M., Salazar, L., Homeier, J., and Kluge, J. (2014). Species richness-productivity relationships of tropical terrestrial ferns at regional and local scales. J. Ecol. 102, 1623–1633. doi: 10.1111/1365-2745.12299
Kier, G., Kreft, H., Lee, T. M., Jetz, W., Ibisch, P. L., Nowicki, C., et al. (2009). A global assessment of endemism and species richness across island and mainland regions. Proc. Natl. Acad. Sci. U.S.A. 106, 9322–9327. doi: 10.1073/pnas.0810306106
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Kier, G., Mutke, J., Dinerstein, E., Ricketts, T. H., Küper, W., Kreft, H., et al. (2005). Global patterns of plant diversity and floristic knowledge. J. Biogeogr. 32, 1107–1116. doi: 10.1111/j.1365-2699.2005.01272.x
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Kissling, W. D., and Carl, G. (2007). Spatial autocorrelation and the selection of simultaneous autoregressive models. Global Ecol. Biogeogr. 17, 59–71. doi: 10.1111/j.1466-8238.2007.00334.x
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Kluge, J., and Kessler, M. (2006). Fern endemism and its correlates: contribution from an elevational transect in Costa Rica. Divers. Distrib. 12, 535–545. doi: 10.1111/j.1366-9516.2006.00231.x
Kluge, J., and Kessler, M. (2011). Influence of niche characteristics and forest type on fern species richness, abundance and plant size along an elevational gradient in Costa Rica. Plant Ecol. 212, 1109–1121. doi: 10.1007/s11258-010-9891-x
Kluge, J., Kessler, M., and Dunn, R. R. (2006). What drives elevational patterns of diversity? A test of geometric constraints, climate and species pool effects for pteridophytes on an elevational gradient in Costa Rica. Global Ecol. Biogeogr. 15, 358–371. doi: 10.1111/j.1466-822X.2006.00223.x
Kooyman, R. M., Rossetto, M., Sauquet, H., and Laffan, S. W. (2013). Landscape patterns in rainforest phylogenetic signal: isolated islands of refugia or structured continental distributions? PLoS ONE 8:e80685. doi: 10.1371/journal.pone.0080685
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Kreft, H., and Jetz, W. (2007). Global patterns and determinants of vascular plant diversity. Proc. Natl. Acad. Sci. U.S.A. 104, 5925–5930. doi: 10.1073/pnas.0608361104
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Kreft, H., Jetz, W., Mutke, J., and Barthlott, W. (2010). Contrasting environmental and regional effects on global pteridophyte and seed plant diversity. Ecography 33, 408–419. doi: 10.1111/j.1600-0587.2010.06434.x
Kromer, T., Kessler, M., Gradstein, S. R., and Acebey, A. (2005). Diversity patterns of vascular epiphytes along an elevational gradient in the Andes. J. Biogeogr. 32, 1799–1809. doi: 10.1111/j.1365-2699.2005.01318.x
Labiak, P. H., Sundue, M. A., Rouhan, G., and Moran, R. C. (2014a). New combinations in Lastreopsis and Parapolystichum (Dryopteridaceae). Brittonia 67, 79–86. doi: 10.1007/s12228-014-9351-3
Labiak, P. H., Sundue, M. A., Rouhan, G., Hanks, J. G., Mickel, J. T., and Moran, R. C. (2014b). Phylogeny and historical biogeography of the lastreopsid ferns (Dryopteridaceae). Am. J. Bot. 101, 1207–1228. doi: 10.3732/ajb.1400071
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Laffan, S. W., Lubarsky, E., and Rosauer, D. F. (2010). Biodiverse, a tool for the spatial analysis of biological and related diversity. Ecography 33, 643–647. doi: 10.1111/j.1600-0587.2010.06237.x
Latham, R. E., and Ricklefs, R. E. (1993). Global patterns of tree species richness in moist forests: energy-diversity theory does not account for variation in species richness. Oikos 67, 325–333. doi: 10.2307/3545479
Lehmann, A., Leathwick, J., and Overton, J. M. (2002). Assessing New Zealand fern diversity from spatial predictions of species assemblages. Biodivers. Conserv. 11, 2217–2238. doi: 10.1023/A:1021398729516
Lwanga, J. S., Balmford, A., and Badaza, R. (1998). Assessing fern diversity: relative species richness and its environmental correlates in Uganda. Biodivers. Conserv. 7, 1387–1398. doi: 10.1023/A:1008865518378
Magurran, A. E., and McGill, B. J. (2011). “Challenges and opportunities in the measurement and assessment of biological diversity,” in Biological Diversity: Frontiers in Measurement and Assessment, eds A. E. Magurran and B. J. McGill (Oxford: Oxford University Press), 1–7.
Maurer, B. A., and McGill, B. J. (2011). “Measurement of species diversity,” in Biological Diversity: Frontiers in Measurement and Assessment, eds A. E. Magurran and B. J. McGill (Oxford: Oxford University Press), 55–65.
McKeown, M., Sundue, M. A., and Barrington, D. (2012). Phylogenetic analyses place the Australian monotypic Revwattsia in Dryopteris (Dryopteridaceae). PhytoKeys 14, 43–56. doi: 10.3897/phytokeys.14.3446
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Miller, M. A., Pfeiffer, W., and Schwartz, T. (2010). “Creating the CIPRES Science Gateway for inference of large phylogenetic trees,” in Proceedings of the Gateway Computing Environments Workshop (GCE), (New Orleans, LA: IEEE), 1–8.
Mishler, B. D., Knerr, N., González-Orozco, C. E., Thornhill, A. H., Laffan, S. W., and Miller, J. T. (2014). Phylogenetic measures of biodiversity and neo-and paleo-endemism in Australian Acacia. Nat. Commun. 5:4473. doi: 10.1038/ncomms5473
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Moran, R. C. (2008). “Diversity, biogeography, and floristics,” in Biology and Evolution of Ferns and Lycophytes, eds T. A. Ranker and C. H. Haufler (Cambridge: Cambridge University Press), 367–394.
Moreno Saiz, J. C., and Lobo, J. M. (2008). Iberian–Balearic fern regions and their explanatory variables. Plant Ecol. 198, 149–167. doi: 10.1007/s11258-007-9392-8
Moritz, C., and Faith, D. P. (2002). Comparative phylogeography and the identification of genetically divergent areas for conservation. Mol. Ecol. 7, 419–429. doi: 10.1046/j.1365-294x.1998.00317.x
Nagalingum, N. S., Knerr, N., Mishler, B. D., and Cargill, D. C. (2014). Overlapping fern and bryophyte hotspots: assessing ferns as a predictor of bryophyte diversity. Telopea 17, 383–392. doi: 10.7751/telopea20148280
National Land and Water Resources Audit. (2002). Australian Terrestrial Biodiversity Assessment. Canberra: Commonwealth of Australia.
Pausas, J. G., Carreras, J., Ferré, A., and Font, X. (2003). Coarse-scale plant species richness in relation to environmental heterogeneity. J. Veg. Sci. 14, 661–668. doi: 10.1111/j.1654-1103.2003.tb02198.x
Perrie, L. R., Wilson, R. K., Shepherd, L. D., Ohlsen, D. J., Batty, E. L., Brownsey, P. J., et al. (2014). Molecular phylogenetics and generic taxonomy of Blechnaceae ferns. Taxon 63, 745–758. doi: 10.12705/634.13
Qian, H., and Ricklefs, R. E. (2000). Large-scale processes and the Asian bias in species diversity of temperate plants. Nature 407, 180–182. doi: 10.1038/35025052
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Qian, H., and Ricklefs, R. E. (2004). Taxon richness and climate in angiosperms: is there a globally consistent relationship that precludes region effects? Am. Nat. 163, 773–779. doi: 10.1086/383097
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
R Core Team. (2014). R: A Language and Environment for Statistical Computing. Available online at: http://www.R-project.org/
Ricklefs, R. E. (2004). A comprehensive framework for global patterns in biodiversity. Ecol. Lett. 7, 1–15. doi: 10.1046/j.1461-0248.2003.00554.x
Rosauer, D. F., and Mooers, A. O. (2013). Nurturing the use of evolutionary diversity in nature conservation. Trends Ecol. Evol. 28, 322–323. doi: 10.1016/j.tree.2013.01.014
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Rosauer, D. F., Laffan, S. W., Crisp, M. D., Donnellan, S. C., and Cook, L. G. (2009). Phylogenetic endemism: a new approach for identifying geographical concentrations of evolutionary history. Mol. Ecol. 18, 4061–4072. doi: 10.1111/j.1365-294X.2009.04311.x
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Schmidt-Lebuhn, A. N., Knerr, N. J., and González-Orozco, C. E. (2012). Distorted perception of the spatial distribution of plant diversity through uneven collecting efforts: the example of Asteraceae in Australia. J. Biogeogr. 39, 2072–2080. doi: 10.1111/j.1365-2699.2012.02756.x
Schuettpelz, E., and Pryer, K. M. (2009). Evidence for a Cenozoic radiation of ferns in an angiosperm-dominated canopy. Proc. Natl. Acad. Sci. U.S.A. 106, 11200–11205. doi: 10.1073/pnas.0811136106
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Schuettpelz, E., and Pryer, K. M. (2007). Fern phylogeny inferred from 400 leptosporangiate species and three plastid genes. Taxon 56, 1037–1050. doi: 10.2307/25065903
Schuettpelz, E., Korall, P., and Pryer, K. M. (2006). Plastid atpA data provide improved support for deep relationships among ferns. Taxon 55, 897–906. doi: 10.2307/25065684
Smith, A. R., Pryer, K. M., Schuettpelz, E., Korall, P., Schneider, H., and Wolf, P. G. (2006). A classification for extant ferns. Taxon 55, 705–731. doi: 10.2307/25065646
Stevenson, L. A., González-Orozco, C. E., Knerr, N., Cargill, D. C., and Miller, J. T. (2012). Species richness and endemism of Australian bryophytes. J. Bryol. 34, 101–107. doi: 10.1179/1743282012Y.0000000004
Tittensor, D. P., Mora, C., Jetz, W., Lotze, H. K., Ricard, D., Berghe, E. V., et al. (2010). Global patterns and predictors of marine biodiversity across taxa. Nature 466, 1098–1101. doi: 10.1038/nature09329
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Tucker, C. M., and Cadotte, M. W. (2013). Unifying measures of biodiversity: understanding when richness and phylogenetic diversity should be congruent. Divers. Distrib. 19, 845–854. doi: 10.1111/ddi.12087
Tuomisto, H., and Poulsen, A. D. (2000). Pteridophyte diversity and species composition in four Amazonian rain forests. J. Veg. Sci. 11, 383–396. doi: 10.2307/3236631
Watkins, J. (2006). Species richness and distribution of ferns along an elevational gradient in Costa Rica. Am. J. Bot. 93, 73–83. doi: 10.3732/ajb.93.1.73
Webb, C. O., Ackerly, D. D., and Kembel, S. W. (2008). Phylocom: software for the analysis of phylogenetic community structure and trait evolution. Bioinformatics 24, 2098–2100. doi: 10.1093/bioinformatics/btn358
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Webb, C. O., Ackerly, D. D., Mcpeek, M. A., and Donoghue, M. J. (2002). Phylogenies and community ecology. Annu. Rev. Ecol. Syst. 33, 475–505. doi: 10.1146/annurev.ecolsys.33.010802.150448
Wiens, J. J., Ackerly, D. D., Allen, A. P., Anacker, B. L., Buckley, L. B., Cornell, H. V., et al. (2010). Niche conservatism as an emerging principle in ecology and conservation biology. Ecol. Lett. 13, 1310–1324. doi: 10.1111/j.1461-0248.2010.01515.x
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Wiens, J. J., and Donoghue, M. J. (2004). Historical biogeography, ecology and species richness. Trends Ecol. Evol. 19, 639–644. doi: 10.1016/j.tree.2004.09.011
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Williams, K. J., Ferrier, S., Rosauer, D. F., Yeates, D., Manion, G., Harwood, T., et al. (2010). Harnessing Continent-Wide Biodiversity Datasets for Prioritising National Conservation Investment. A report prepared for the Department of Sustainability, Environment, Water, Population and Communities, Canberra.
Williams, P. H., and Gaston, K. J. (1994). Measuring more of biodiversity: can higher-taxon richness predict wholesale species richness? Biol. Conserv. 67, 211–217. doi: 10.1016/0006-3207(94)90612-2
Williams, P. H., Gaston, K. J., and Humphries, C. J. (1997). Mapping biodiversity value worldwide: combining higher-taxon richness from different groups. Proc. R. Soc. B Biol. Sci. 264, 141–148. doi: 10.1098/rspb.1997.0021
Winter, M., Devictor, V., and Schweiger, O. (2013). Phylogenetic diversity and nature conservation: where are we? Trends Ecol. Evol. 28, 199–204. doi: 10.1016/j.tree.2012.10.015
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Zanne, A. E., Tank, D. C., Cornwell, W. K., Eastman, J. M., Smith, S. A., FitzJohn, R. G., et al. (2014). Three keys to the radiation of angiosperms into freezing environments. Nature 506, 89–92. doi: 10.1038/nature12872
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Keywords: Polypodiopsida, Filicopsida, ferns, Australia, conservation, evolution, community, spatial phylogenetics
Citation: Nagalingum NS, Knerr N, Laffan SW, González-Orozco CE, Thornhill AH, Miller JT and Mishler BD (2015) Continental scale patterns and predictors of fern richness and phylogenetic diversity. Front. Genet. 6:132. doi: 10.3389/fgene.2015.00132
Received: 30 June 2014; Accepted: 19 March 2015;
Published: 14 April 2015.
Edited by:
Toby Pennington, Royal Botanic Garden Edinburgh, UKReviewed by:
Mary Gibby, Royal Botanic Garden Edinburgh, UKMichael Kessler, University of Zurich, Switzerland
Copyright © 2015 Nagalingum, Knerr, Laffan, González-Orozco, Thornhill, Miller and Mishler. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) or licensor are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Nathalie S. Nagalingum, Royal Botanic Gardens and Domain Trust, Mrs Macquaries Road, Sydney, NSW 2000, AustraliabmF0aGFsaWUubmFnYWxpbmd1bUByYmdzeWQubnN3Lmdvdi5hdQ==