 Conceição L. V. Santos
Conceição L. V. Santos Bertrand Pourrut
Bertrand Pourrut José M. P. Ferreira de Oliveira3
José M. P. Ferreira de Oliveira3- 1Department of Biology, Faculty of Sciences, University of Porto, Porto, Portugal
- 2Laboratoire Génie Civil et géo-Environnement - Groupe ISA, Lille, France
- 3Laboratory of Biotechnology and Cytometry, Centre for Environmental and Marine Studies, University of Aveiro, Aveiro, Portugal
The systematic study of genotoxicity in plants induced by contaminants and other stress agents has been hindered to date by the lack of reliable and robust biomarkers. The comet assay is a versatile and sensitive method for the evaluation of DNA damages and DNA repair capacity at single-cell level. Due to its simplicity and sensitivity, and the small number of cells required to obtain robust results, the use of plant comet assay has drastically increased in the last decade. For years its use was restricted to a few model species, e.g., Allium cepa, Nicotiana tabacum, Vicia faba, or Arabidopsis thaliana but this number largely increased in the last years. Plant comet assay has been used to study the genotoxic impact of radiation, chemicals including pesticides, phytocompounds, heavy metals, nanoparticles or contaminated complex matrices. Here we will review the most recent data on the use of this technique as a standard approach for studying the genotoxic effects of different stress conditions on plants. Also, we will discuss the integration of information provided by the comet assay with other DNA-damage indicators, and with cellular responses including oxidative stress, cell division or cell death. Finally, we will focus on putative relations between transcripts related with DNA damage pathways, DNA replication and repair, oxidative stress and cell cycle progression that have been identified in plant cells with comet assays demonstrating DNA damage.
Plant Comet Assay: General Considerations
The first reports on the use of comet assay in plants date from the 1990's (e.g., Cerda et al., 1993; Koppen and Verschaeve, 1996; Navarrete et al., 1997; Koppen and Angelis, 1998).
Despite similarities with other eukaryotic systems, namely animal models, the comet assay protocols for plants take into account relevant differences including the presence of a rigid cell wall in plant cells. The localized presences of characteristic meristematic regions (e.g., the concentration of highly dividing cells in the root apex) and the fact that root is usually the organ directly in contact with contaminated soil and water, have also influenced the establishment of plant comet assays in ecotoxicological approaches. Technical details concerning plant comet assays in different organs and species have been thoroughly reviewed by Gichner et al. (2009).
For almost a decade, the comet assay remained restricted to some toxicological studies and to a few model species including Allium cepa, Nicotiana tabacum, Vicia faba, and Arabidopsis thaliana (for review, Gichner et al., 2009; Ventura et al., 2013).
Plant comet assay has been applied to an increasing variety of adverse conditions. Some recent reviews on this subject (Gichner et al., 2009; Ventura et al., 2013) revised most relevant advances in plant comet assay up to 5 years ago. Since then an increasing interest for comet assay in plants was shown (136 articles published between 2010 and March 2015 vs. 89 between 1995 and 2009). Therefore, here we will mostly emphasize most relevant advances within the last 5 years, and highlight current applications of this technique in plant (eco) toxicological studies. We will also discuss advances on genetic studies involving DNA damage and repair.
Basic Principles and Methodologies
Comet assays traditionally use cell suspensions, which are embedded in agarose on a microscope slide, and exposed to lysis by exposure to detergent and high salt solutions (for review Collins et al., 2008; Azqueta et al., 2009). Lysis allows removing membranes and soluble cell components, leaving a supercoiled DNA nucleoid (Azqueta et al., 2011b). When submitted to electrophoretic conditions, DNA fragments will migrate toward the anode, forming a typical “comet tail.” The amount of strand breaks is overall proportional to the amount of DNA in the tail respectively to the DNA remaining in the head (Hovhannisyan, 2010).
However, in plants, the presence of a cell wall causes technical issues for performing the comet assay on plant tissues. To overcome these problems, a simple and efficient mechanical extraction to isolate cell nuclei was developed by Cerda et al. (1993), and then improved by Koppen and Angelis (1998), Navarrete et al. (1997), and Gichner and Plewa (1998). Since then, most of the researchers used directly those protocols or derived versions, such as described in Gichner and Plewa (1998). Recently, Pourrut et al. (2015) identified the key steps of comet assay in plants and proposed an optimized protocol to increase its reliability and its throughput. In the case of plant chopping, particular attention has to be paid to the presence of chloroplasts as they are important sources of free radicals and oxidative damage. For example, the first article on plant comet assay testing chemicals used isolated nuclei of Vicia faba root cells (Koppen and Verschaeve, 1996). In cellular assays, plants exposed to suspected genotoxicants are processed for nuclei isolation and analysis, whereas in acellular assays, nuclei from non-stressed plants are isolated and then incubated with the genotoxicants, before comet assay analysis.
The use of protocol variants allows detecting a wide range of DNA damages (see for review Angelis et al., 1999; Collins et al., 2008). Briefly, an alkaline treatment (referred hereafter as A/A) and electrophoresis at pH 13 or higher allows the detection of most single and double DNA strand breaks (SSBs and DSBs) and also alkali-labile sites. When the unwinding and subsequent electrophoresis are performed using a buffer pH~7–8, the comet assay is called “neutral” (N/N). A crucial difference is that at alkaline conditions, apurinic/apyrimidinic sites are more easily subjected to break (for details refer to Azqueta et al., 2011b). Other pH-variants (e.g., A/N) have meanwhile been introduced as alternative comet assays.
Moreover, the information provided by comets may also be increased by exposing the DNA to enzymes recognizing a specific lesion, e.g., formamidopyrimidine DNA glycosylase, Endonuclease III, thereby originating specific breaks. However, despite their strong interest and their early introduction in plant studies (Menke et al., 2000), these enzymes are still not much used in plants.
Comets may then be visualized by microscopy, by using a suitable DNA-binding dye, e.g., fluorescent dyes or silver staining. Data can be analyzed by visual scoring, ranging from 0 to 4 according to the damage class, or using computer-based image analysis (e.g., the software http://casplab.com/) that allows the quantification of several comet parameters, including the tail DNA %, tail length, tail extension moment or Olive tail movement (Azqueta et al., 2011b). Criteria for the best scoring approaches are however debatable (e.g., Azqueta et al., 2011a), but independently of the approach and scoring, it is consensual that this technique allows collecting data suitable for robust statistical analyses.
Radiation
Plants are prone to DNA damage upon exposure to radiation from natural or anthropogenic sources. For this reason, the analysis of DNA damage in irradiated plants is a topic of growing interest and sensitive methods for detection of DNA damage have been applied (Table 1).
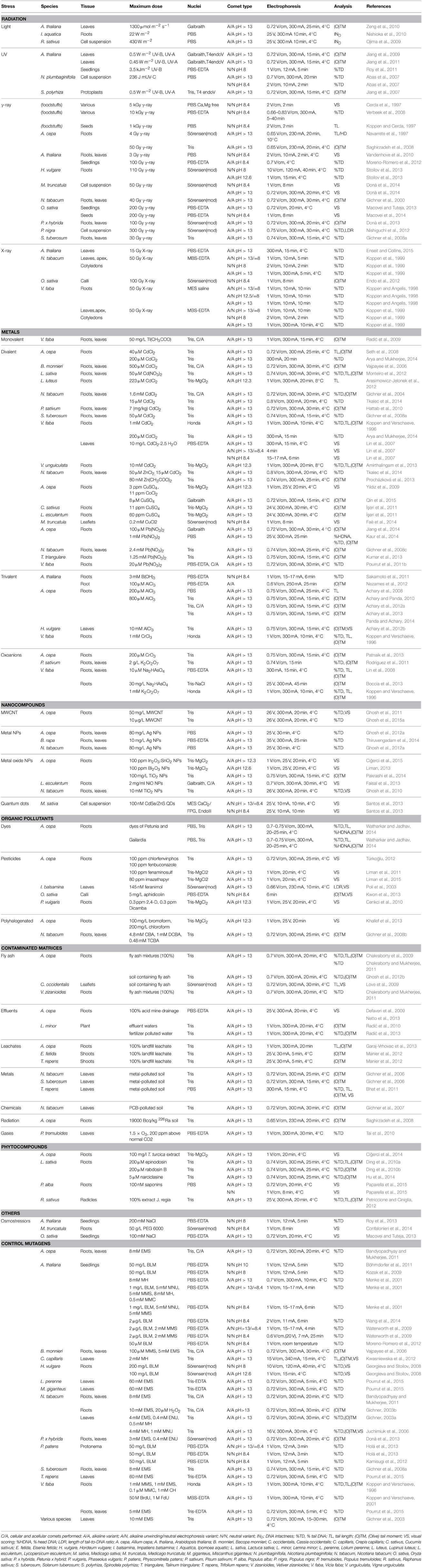
Table 1. Overview of comet assay studies in plant toxicology.
The effects of light excess on plant DNA using comet assay were firstly investigated by Ojima et al. (2009) on Raphanus sativus protoplasts. These authors demonstrated that light excess causes DNA degradations mediated by oxidative stress. In 2010, Nishioka et al. confirmed the role of reactive oxidative species (ROS) in light excess-induced DNA damages in Ipomoea aquatica root protoplasts, and correlated DNA damages observed by comet assay with chlorophyll degradation. However, these two studies did not take into consideration the potential role of UV in light-induced DNA damages. In a study designed to investigate UV-A and UV-B effects, Jiang et al. (2007) performed comet to detect specific DNA lesions as well as pyrimidine dimers formation (using T4 endonuclease V) in irradiated Spirodela polyrhiza protoplasts. These results were confirmed later in Arabidopsis thaliana root tip cells (Jiang et al., 2009, 2011). Jiang et al. (2011) also demonstrated that UV-B-induced DNA damage results in the delay of G1-to-S transition of plant cell cycle. However, by using a neutral comet assay (N/N variant), Roy et al. (2011) showed that UV-B-induced lesions were reversible, particularly in A. thaliana wild-type (Col-0), compared to DNA polymerase λ UV-B sensitive mutants. UV-C was also shown to induce both SSBs and DSBs in Arabidopsis plumbaginifolia protoplasts (Abas et al., 2007). These authors also highlighted the usefulness of the comet assay as an analytical tool for the analysis of repair kinetics in protoplasts. These results were confirmed by Bilichak et al. (2014) on A. thaliana protoplasts.
Besides natural exposure to radiation, plants are also irradiated for industrial purposes. For example, gamma (γ)-rays are used to increase seed vigor and/or enhance plant tolerance to environmental stresses. Navarrete et al. (1997) pioneered the comet research in plants through the optimization of different steps in the comet assay applied to γ-irradiated A. cepa roots. Moreover, Cerda et al. (1997), Koppen and Cerda (1997) and Verbeek et al. (2008) optimized the comet assay to screen DNA damage in γ-irradiated seeds, dried fruits and spices. At the same period, Gichner et al. (2000, 2008a) used the A/A variant to study the effects of the γ-rays in irradiated tobacco and potato plants, respectively.
Later, Böhmdorfer et al. (2011) used this technique to study DSB formation in Arabidopsis homologous recombination deficient mutants subjected to γ-rays. On the other hand, Vandenhove et al. (2010) applied low γ-radiation dose rates for long periods to Arabidopsis plants. Despite the growth limitations and induction of oxidative stress response, the low applied radiation dose applied did not induce DNA damages measurable by the comet assay. Moreover, Macovei et al. (2014) demonstrated the occurrence of DSBs in rice (Oryza sativa L.) seedlings after exposure to γ-rays concomitant with a difference in expression profiles of three miRNAs, and an increase of reactive oxygen species (ROS) levels. Combining the use of the comet assay, and the expression of genes encoding DNA repair-related proteins, Nishiguchi et al. (2012) investigated the mechanisms of γ-radiation-induced DNA degradation and repair in Lombardy poplar (Populus nigra var. italica). Donà et al. (2014) studied further the mechanisms associated with plant sensitivity to γ-irradiation. By comparison of A/N and N/N variants of the comet assay in Medicago truncatula, these authors argued that active repair of DSBs occurred in treated cells. However, SSB repair did not occur and SSBs continued to accumulate as a consequence of increasing ROS levels. It is necessary to point that the distinction by comet assay of DSBs and SSBs is not trivial, since the neutral assay with prolonged protease digestion at high temperature will more likely only detect DSBs. The research team demonstrated in Petunia x hybrida treated with low and high-dose γ-irradiation that the level of DNA strand breaks was higher in the high-dose group. However, after 2 h the two groups showed identical amounts of strand breaks, suggesting a faster initial DNA repair in the high-dose group.
Alkaline and neutral DNA comet assays were also used to estimate both the levels of DNA damages and the repair potential in the barley lines T-1586 and D-2946 after exposure to γ-rays and Li ions (Stoilov et al., 2013). The authors found that the mutant line D-2946 was more sensitive to γ-radiation, supporting that susceptibility to this radiation is genotype dependent. Overall, these data support that the genotype, radiation dose and time of radiation exposure are crucial factors that determine the effects of radiation on DNA integrity.
In comparison to γ-rays, comet assay has been little used to evaluate DNA damages induced by X-rays. Using alkaline comet assay, Koppen and Angelis (1998) demonstrated that X-rays induce a linear increase of DNA content in the comet tail of irradiated V. faba plants. Endo et al. (2012) reported that X-ray exposure in calli of Oryza sativa resulted in a dose-dependent increase of DSBs, as shown by neutral comet assay. Recently, Enseit and Collins (2015) studied the effect of low dose radiations on DNA repair mechanisms using alkaline comet assay. They identified two phases of DNA repair after acute exposures of 5 and 15 Gy (“rapid” and “slow” phases). With lower exposures (2 Gy and lower), they also highlighted that “rapid” repair was so fast that it was difficult to detect.
Concerning radioactive contaminations, Saghirzadeh et al. (2008) successfully demonstrated that very high levels of natural radioactivity (e.g., by accumulation of 226Ra) presented by soils were significantly genotoxic to A. cepa roots, with DNA damages measured by comet assay and compared to the effects of increasing γ-ray doses.
Metals
Most of the contaminated sites worldwide are contaminated with heavy metals. In Europe, heavy metals contaminated almost 50% of the investigated sites (Panagos et al., 2013). Exposure to metals may induce a variety of direct and indirect phytotoxic effects (e.g., Silva et al., 2010). In general metals induce more severe symptoms in roots than in leaves, since roots are in direct contact with the soil and generally with the toxic contaminant.
The first comet assays evaluating metal genotoxicity in plants were pioneered by Koppen and Verschaeve (1996) which studied chromium (Cr) and cadmium (Cd) genotoxicity in V. faba. These authors showed a dose-dependent increase in DNA damage. More recently, Cd-induced DNA degradations were also observed in Trifolium repens (Bhat et al., 2011), Lactuca sativa (Monteiro et al., 2012), Lupinus luteus (Arasimowicz-Jelonek et al., 2012), Vigna unguiculata (Amirthalingam et al., 2013), N. tabacum (Tkalec et al., 2014), V. faba and A. cepa (Arya and Mukherjee, 2014). However, dose-dependent responses were not clearly observed in these studies. This could be explained by the fact that these authors lead hydroponic studies and used very high and environmental-unrealistic concentrations of cadmium. Monteiro et al. (2012) suggested that these high concentrations could induce Cd-DNA adducts that lead to DNA-DNA/DNA-protein cross-links, and/or formation of longer DNA fragments, and/or impairment of DNA repair mechanisms, which could explain these results. Interestingly, the only study using soil spiked with environmental-realistic concentrations of cadmium (Hattab et al., 2010), demonstrated a dose-dependent increase in DNA damages in P. sativum. Tkalec et al., (2014) and Amirthalingam et al. (2013) also used the comet assay to understand Cd-induced genotoxicity mechanisms. They suggested the implication of oxidative stress while Arasimowicz-Jelonek et al. (2012) showed that scavenging the endogenous nitric oxide (NO) pool during Cd stress, despite reducing the programmed cell death, did not affect the degree of DNA damages evidenced by comet assay. Recently, comet assay was used to investigate the difference of sensitivity to Cd exposure of A. cepa and V. faba (Arya and Mukherjee, 2014). The results indicated that exposure to Cd induced slight dose-dependent increase in chromosomal aberrations, DNA fragmentation and micronucleus frequency in both A. cepa and V. faba. However, V. faba appeared more sensitive than A. cepa toward Cd-induced genotoxicity, which was correlated to the increased level of oxidative stress in root tissues.
Along with Cd, aluminum (Al) genotoxicity has been the most studied during the last years. Achary et al. (2008, 2012a) and Achary and Panda (2010) demonstrated dose-dependent DNA damage induced by Al exposure on A. cepa roots. These results were confirmed later on Hordeum vulgare (Achary et al., 2012b) and Andropogon virginicus (Ezaki et al., 2013). These studies also highlighted the implication of oxidative stress in Al genotoxicity. Comet assay was also used to investigate the mechanisms of Al genotoxicity, underscoring the role of cell wall-bound NADH-PX in the Al oxidative burst-mediated (Achary et al., 2012a), and the role of signal transduction mediated by Ca2+ (Achary et al., 2013) and MAP Kinases (Panda and Achary, 2014) in Al-induced cell death and DNA damage. Interestingly, these authors also described the occurrence of adaptation responses that involved oxidative stress, and that root cells conditioned with low doses of Al (<10 μM Al3+) developed adaptive responses and protection mechanisms against genotoxic effects of the mutagenic agents methylmercuric chloride (MMCl) and ethyl methanesulfonate (EMS) (Achary et al., 2013). Moreover, the role of DNA damage in Al-dependent root growth inhibition was also investigated in A. thaliana mutants (Rounds and Larsen, 2008; Nezames et al., 2012).
The phytotoxicity of lead (Pb) including genotoxic aspects was reviewed by Pourrut et al. (2011a). Using comet assay, Gichner et al. (2008c) were the first to demonstrate dose-dependent Pb-induced DNA damage in N. tabacum in hydroponic and soil experiments. These results were confirmed on Talinum triangulare roots and correlated with Pb-induced oxidative stress (Kumar et al., 2013). However, both studies used very high and environmentally-unrealistic concentrations of Pb. More interestingly, dose-dependent Pb-induced DNA damage were also observed with lower and environmentally-realistic concentrations of Pb (<20 μM Pb) in V. faba plants (Pourrut et al., 2011b). Moreover, these authors also confirmed the role of oxidative stress in this damage process, since co-incubation with antioxidant vitamin E or the NADPH-oxidase inhibitor dephenylene iodonium inhibited DNA damage and micronuclei formation in exposed roots (Pourrut et al., 2011b). Recently, two studies performed on A. cepa confirmed the role of oxidative stress in lead-induced genotoxicity and that DNA damages are also tightly linked to the cell cycle (Jiang et al., 2014; Kaur et al., 2014).
Similarly, the micronutrient copper (Cu) was shown to induce significant DNA damages in A. cepa roots (Yıldız et al., 2009; Qin et al., 2015). Very high concentrations of copper chloride also increased DNA fragmentations in P. sativum roots but not in leaves (Hattab et al., 2010). Similarly to the above-cited metals, Cu-induced DNA damages were associated with cytotoxic damages involving oxidative stress in Lycopersicon esculentum and Cucumis sativus roots (İşeri et al., 2011) and other chromosome aberrations in A. cepa roots (Yıldız et al., 2009). Recently, Faè et al. (2014) used the neutral comet assay to demonstrate the overexpression efficiency of the DNA repair gene MtTdp2a for enhancing plant tolerance to Cu exposure in Medicago truncatula mutants.
By using the comet assay, Lin et al. (2008) proved that arsenate (10 μM) induced DNA damages in V. faba leaves and roots, in a dose-dependent manner and that these effects were associated with oxidative stress. Sturchio et al. (2011) confirmed As genotoxicity in V. faba roots grown on sandy and clay-loamy soil spiked with arsenate. In the same species, Boccia et al. (2013) combined the comet assay with infrared (FTIR), and near infrared (FTNIR) spectroscopy, to show that arsenate (20 and 30 mg/L) induced DNA damages which were associated with structural changes of different functional groups, suggesting the possible replacement of phosphate by arsenate in DNA.
The plant comet assay also contributed to clarify the effects of several other metals in plant DNA damages (Table 1). For example, Radić et al. (2009) demonstrated that the rare metal thallium (Tl), released to the environment as a by-product of Fe and Zn refining processes, induces DNA damages together with oxidative damages in V. faba seedlings. The comet assay was also helpful in demonstrating that boron (B) toxicity mechanism in plants involves DSBs and possibly replication blocks, with plant condensin II playing a critical role in DNA damages repair (Sakamoto et al., 2011). Rodriguez et al. (2011) and Rodriguez (2011) used a battery of genotoxic and cytotoxic biomarkers to assess Cr (VI) toxicity in pea, and were able to correlate Cr (VI)-induced DNA damages (demonstrated by comet assay) with cell cycle arrest at the G2/M checkpoint and with clastogenicity assessed by flow cytometry (Rodriguez, 2011, PhD thesis). Moreover, Patnaik et al. (2013) showed by alkaline comet assay that induction of DNA damage by Cr (VI) was dose-dependent in A. cepa. However, in plants exposed to 1-day treatment followed by 4-day recovery, no effects were found by comet assay. On the same plant species, cobalt (Co) was shown to induce significant DNA damages (Yıldız et al., 2009).
Besides some more established physiological analyses, the comet assay has also been conducted to determine the differential toxic effects affecting different plant organs. Procházková et al. (2013) showed that in N. tabacum zinc (Zn) induces higher DNA damages in roots compared to leaves. This differential effect was possibly attributable to the higher accumulation of Zn (II) in roots, compared to shoots. Tkalec et al. (2014) also observed these effects in N. tabacum. However, these authors also shown that, when Zn was added in the culture medium in combination with Cd, this metal conversely exhibited a protective effects against Cd-induced DNA damages.
It is worth noting that the interest of using the comet assay as a reliable biomarker on ecotoxicological assays is increasing, and Bandyopadhyay and Mukherjee (2011) applied both acellular and cellular comet tests to compare A. cepa and N. tabacum as toxicity models in rapid monitoring Cd-induced genotoxicity. Monteiro et al. (2012) used a battery of tests including the comet assay, to determine differences associated with organ dependence in Cd toxicity. The authors used Lactuca sativa and integrated cytostaticity/genotoxicity and oxidative stress data, where parameters measured by the comet assay (e.g., tail moment) were demonstrated to be relevant genotoxicity biomarkers. Despite still restricted to a few number, some studies have already used plant comet in field ecotoxicology assays of soils contaminated with metals (see Section “Contaminated Matrices” below).
Nanocompounds
Plant comet assays are also increasingly used to assess the phytotoxicity of small-scale materials (Table 1), e.g., nanomaterials and in particular nanoparticles (NPs). Nanomaterials possess unique properties suitable for a wide range of industrial applications. For this reason and due to their intense uses and subsequent release to the environment, they are currently classified as emerging contaminants. One example of emerging nanomaterials are carbon nanotubes, that depending on the physical properties can pose cytotoxicity to mammalian and plant cells (Ghosh et al., 2011). Ghosh et al. (2011, 2015a) demonstrated a correlation between DNA strand breaks and the concentration of multi-walled carbon nanotubes in A. cepa, supporting the genotoxic potential of this type of nanomaterials.
The increasing amount of NPs in groundwater and soil has raised environmental concerns regarding their putative toxicity and fate through food chains. A large group of NP contaminants include toxic or reactive metals NPs. One of the most relevant pioneer studies of NPs genotoxicity in plants was done with TiO2 NPs in A. cepa (Ghosh et al., 2010). In this study the comet assay was used to assess DNA damages and this endpoint was combined with oxidative stress endpoints (e.g., malondialdehyde level). Moreover, in A. cepa roots, TiO2 NPs induced DNA damages confirmed by comet assay and correlated with the occurrence of chromosomal aberrations (Pakrashi et al., 2014).
Silver nanoparticles (AgNPs) were shown to induce DNA damages in A. cepa and N. tabacum with more pronounced effects in roots than in shoots (Ghosh et al., 2012a).
Recently, using higher NPs concentrations, Thiruvengadam et al. (2015) also demonstrated a dose-dependent increase in DNA damages in Brassica rapa ssp. rapa, and this result was confirmed by DNA laddering and TUNEL assays.
Bismuth (III) oxide NPs increased the nuclear DNA damages in A. cepa plants. These data supported the concomitant observation of chromosomal aberrations and mitotic aberrations in the same tissues (Liman, 2013).
The alkaline comet assay showed an increase of DNA damages in tomato seedlings exposed to NiO-NPs up to 2 mg/ml (Faisal et al., 2013). In this study the authors also used the plant comet assay test to assess the percentage of necrotic and apoptotic cells, however, these conclusions must be regarded carefully as the validity of the comet assay in identifying apoptotic cells remains a matter of discussion (Collins et al., 2008).
Indium (III) oxide and tin (IV) oxide is a mixture widely used in industrial coating. A significant increase in DNA damages was recently observed of A. cepa root meristematic cells exposed to doses up to 100 ppm of indium tin oxide suspension (Ciğerci et al., 2015).
Besides metal oxide NPs, quantum dots form another type of nanomaterials increasingly prevalent in the environment. Quantum dots are nanomaterials used in electronics which possess semiconducting properties, composed for example of arsenic (As), selenium (Se) and tellurium (Te) in various proportions. Despite their increasing prevalence in the environment, the toxicity of quantum dots in plants is largely unknown. In a pioneer study, Santos et al. (2013) used a battery of tests and gene expression related with DNA repair, and demonstrated that 10 nM 3-mercaptopropanoic coated-CdSe/ZnS quantum dots were cytotoxic and genotoxic to Medicago sativa cells. In this and other pioneer studies, the comet assay can play a pivotal role as a tool to assess environmental impacts of suspected emerging nanocontaminants.
Organic Pollutants
Several researchers have used the comet assay to monitor DNA damages induced in plants by numerous organic pollutants (Table 1). The most common organic chemical contaminants include reactive compounds, e.g., alkylating agents, azo dyes, cyclic aromatic hydrocarbons and chemicals incorporated in pesticides and herbicides.
The comet assay was recently used to better understand the role of homologous recombination and genome stability during DNA replication. Comet assay was used to study, in alfalfa, broad bean, lentil, miscanthus, onion, potato, tobacco, sugar beet and wheat, how different agents including ethyl methanesulfonate (EMS) and/or H2O2 induce DNA damages (Gichner et al., 2008a; Bandyopadhyay and Mukherjee, 2011; Pourrut et al., 2015). Due to their dose-dependent genotoxic effects, EMS and H2O2 became largely used as positive controls in plant comet assays, providing further robustness to the assay (Gichner et al., 2008a; Bandyopadhyay and Mukherjee, 2011; Pourrut et al., 2015). Similarly, the dose-dependent induction of DNA damages by compounds such as N-methyl-N-nitroso-urea (MNU), methyl methanesulfonate (MMS) and mitomycin C (MMC) (e.g., Menke et al., 2001; Juchimiuk et al., 2006) supported the wide use of these compounds as positive controls.
Azo dyes are important xenobiotic compounds, largely used in textile industry. Their putative genotoxicity was recently demonstrated in Petunia grandiflora and Gaillardia grandiflora by comet assay, in a pioneer study of plant–plant association for phytoremediation involving the treatment of textile dyes (Watharkar and Jadhav, 2014). Recently, it was demonstrated that bromoform (which may occur during disinfection processes of water) and chloroform (>25 μg/mL) increased chromosome aberrations and DNA damages, this last one assessed by comet assay in A. cepa roots (Khallef et al., 2013). Also chlorobenzoic acids (CBAs) may be found in soils contaminated with polychlorinated biphenyls (PCBs), and have mutagenic and carcinogenic effects in animals. Gichner et al. (2008b) demonstrated that the levels of CBAs inducing leaf withering or death also induced DNA migration in the comet assay.
In the last decade, several pesticides were demonstrated to induce DNA damages in plant cells (e.g., Poli et al., 2003). Endosulfan is an organochlorine pesticide widely used, and its genotoxicity was demonstrated in white clover (Trifolium repens) roots after exposure to doses up to 10 mg/L (Liu et al., 2009). The use of comet assay on A. cepa roots also demonstrated the genotoxic effects of the organophosphate insecticide/acaricide chlorfenvinphos and the triazole fungicide fenbuconazole (Türkoğlu, 2012). The experiment included tests/parameters such as the mitotic index, mitotic phase, chromosomal abnormalities, 2C DNA content (pg) and the plant comet assay on root meristem cells of A. cepa. Results indicated a robust negative correlation between both pesticides-induced DNA damage and 2C DNA amount. On the same plant model, Liman et al. (2013) studied the genotoxicity of the aromatic diazo fungicide and micro-biocide fenaminosulf. Comet assay clearly indicated a dose-dependent genotoxicity of Fenaminosulf in the root meristematic cells of A. cepa, which was confirmed by Mitotic index analysis. Herbicide genotoxicity was also evaluated by comet assay. Cenkci et al. (2010) demonstrated dose-dependent DNA-damages in common bean (Phaseolus vulgaris) roots used treated by two herbicides 2,4-D (2,4-dicholorophenoxyacetic acid) and Dicamba (3,6-dichloro-2-methoxybenzoic acid). These results were confirmed in the same study by RAPD analysis. Recently, Liman et al. (2015) also observed a dose-dependent DNA degradation induced by the imidazolinone herbicide Imazethapyr in A. cepa roots.
Antibiotics were also shown to induce DNA damages in plant cells. For example, the cytostatic effects of the antibiotic bleomycin (a DNA damaging glycopeptide) were demonstrated in plants, e.g., barley (Georgieva and Stoilov, 2008; Stoilov et al., 2013). Bleomycin also induced DNA oxidative damages and single and double strand breaks in the wild moss Physcomitrella lines and in the lig4 mutant (Holá et al., 2013). Similarly, MMC induced a dose-dependent increase in DNA damages in Arabidopsis plants (Menke et al., 2001).
Contaminated Matrices
Despite the promising data concerning the robustness and suitability of the comet assay for screening metal-induced DNA damages in plant cells, its use to assess the genotoxicity of poly-contaminated matrices, including samples of contaminated soils, of leakages or fly ashes, remains scarce (Table 1). In a pioneer study, Gichner et al. (2006) used the alkaline comet assay to demonstrate DNA damages in both N. tabacum and Solanum tuberosum plants exposed to soils contaminated with a mixture of Cd, Cu, Pb, and Zn. Also, soil samples polluted with polychlorinated biphenyls were shown to induce DNA damages in tobacco plants (Gichner et al., 2007). These authors concluded that comet assays may be used for monitoring the DNA-damaging effects of environmental pollutants.
In a microcosm study, and using T. repens as plant model, Manier et al. (2012) found a dose-dependent increase in DNA damages in plants exposed to soil contaminated with landfill leachate. Garaj-Vrhovac et al. (2013) used the comet assay to validate two new methods of leachate treatment, which induced less DNA damages in A. cepa roots than the untreated landfill leachate. Comet assay was also used to evaluate the efficiency of new treatment technology to decrease acid mine drainage genotoxicity. Defaveri et al. (2009) and Netto et al. (2013) used A. cepa roots, and different biomarkers including DNA damages and other cytotoxic and physiological biomarkers, while Radić et al. (2010) used the aquatic species Lemna minor. In a previous study, these authors demonstrated in Lemna minor that the tail moment assessed by the plant comet assay and parameters related to oxidation were suitable as biomarkers for environmental monitoring of the toxicity of industrial effluents in Croatia (Radić et al., 2010). Importantly, the same group (Radić et al., 2013) found comparable responses in fish and Lemna minor regarding DNA damage and oxidative stress, after exposure to polluted surface water contaminated by a fertilizer factory effluent rich in fluorides, metals, and polycyclic aromatic hydrocarbons. The authors highlighted that their results imply that conventional chemical analysis should be extended to genotoxicity/toxicity biological assays to better predict potential health hazard.
Fly ashes are generated during combustion, and include fine particles, with different sizes, rising to the atmosphere. Their complex constitution raised questions on their genotoxicity to animals and plants. Love et al. (2009) demonstrated, based upon comet assay results, that higher levels of DNA damages were found in leaves of Cassia occidentalis exposed to fly ash, compared to non-exposed controls. The authors suggested that these DNA damages might be associated with foliar concentrations of As and Ni absorbed from the fly ash. Ghosh et al. (2012b) studied the genotoxicity in A. cepa of soil samples contaminated with metal-rich fly ashes from a thermal power plant in India and concluded that the observed DNA damages could be correlated to the presence of toxic metals. Also, Chakraborty et al. (2009) studied the genotoxic effects of fly ash comparing the comet assay and the Allium test in this model species. The authors supported the combination of these two techniques in monitoring assays. The same group used the comet to validate the relevance of Vetiveria zizanioides as a good candidate for remediation of fly ash dumpsites (Chakraborty and Mukherjee, 2011). They demonstrated this plant could grow in the presence of fly ash without any genotoxic effects in comparison to A. cepa which exhibited a very high DNA degradation (>80%). Later, this research group used comet assay on A. cepa to monitor the remediation efficiency of V. zizanioides on fly ash amended soils (Ghosh et al., 2015b). They showed that this plant was able to strongly mitigate the genotoxic potential of these soils. These results were also confirmed by a reduction in micronuclei formation, binucleate cells and chromosomal aberrations.
The effects of air contaminants on plant DNA-damages have also been studied in the last years. For example, Populus tremuloides clones exposed to air enriched with O3 alone, or CO2 + O3 showed increased DNA damages levels above background as measured by the comet assay, but these effects were genotype dependent (Tai et al., 2010).
Phytocompounds
A wide number of phytocompounds (including alkaloids, phenolic compounds, glycosides, flavonoids, anthocyanins, etc) may have cytotoxic and genotoxic effects or have protective roles against stressing conditions in a wide number of species, including humans. The way phytocompounds influence oxidative stress balances, and regulate programmed cell death pathways and cell cycle chekpoints, support their wide therapeutic use (e.g., Ascenso et al., 2013; Ferreira de Oliveira et al., 2014). Recently, the interest of using comet assay to monitor genotoxic effects of some phytocompounds on other plant species has emerged (Table 1). For example, Petriccione and Ciniglia (2012) demonstrated the occurrence of a dose-dependent accumulation of DNA damages in Raphanus sativus (radish) radicles treated with Juglans regia husk water extracts. It should be noted that the authors stressed the need of performing accurate and appropriate statistical evaluations of comet results, an emerging topic of discussion. Ciğerci et al. (2014) also used alkaline comet assay to demonstrate the genotoxicity of Thermopsis turcica extracts on A. cepa roots. They showed dose-dependent DNA damages which were confirmed by RAPD profile analysis.
The alkaloid narciclasine (extracted from N. tazetta) was recently shown to inhibit plant growth of Oryza sativa, A. thaliana, Brassica rapa or Lactuca sativa (Hu et al., 2014). The comet assay, complemented with the terminal deoxynucleotidyl transferase dUTP nick end labeling (TUNEL) assay, showed a narciclasine dose-effect response in lettuce seedlings, and this triggered DNA damages may involve increased oxidative stress (Hu et al., 2014). Contrarily, anthocyanins protected DNA integrity (detected by comet assay) in Arabidopsis plants during prolonged exposure to high-light (1300 mmol/m2/s) (Zeng et al., 2010).
Epinodosin, and rabdosin B, diterpenoids isolated from Isodon japonica, exhibited a biphasic dose-dependent effect on Lactuca sativa root growth. The inhibitory effects of both compounds found at higher doses was paralleled with an increase of DNA damages and an inhibition of root cell mitotic activity or retardation of the cell cycle, respectively (Ding et al., 2010a,b). Other terpenes (saponins) extracted from Medicago sativa were shown to induce SSBs and DSBs in Populus alba cell cultures (Paparella et al., 2015). Very interestingly, these authors demonstrated that for all 11 tested saponins, neutral comet assay resulted in similar DSBs patterns, indicating a general response to saponin-induced genotoxic stress, not related to the specific structure of these molecules. Differently, the evaluation of DNA damages performed with alkaline comet assay provided distinct profiles depending on the tested saponin.
Comet assay was also used to evaluate the effect of the phytohormone salicylic acid. Interestingly, Yan et al. (2013) demonstrated that salicylic acid can generate DNA damages in the absence of a genotoxic agent in A. thaliana, supporting that activation of DNA damage responses is an intrinsic component of the plant defense responses.
Comet Assay and Putative Genetic Associations
The comet assay has contributed to elucidate the DNA repair mechanisms involved in the response to external stress factors. A variety of methodologies can be used to investigate DNA repair mechanisms in plants (Azqueta et al., 2009), the most common being the study of plants exposed to DNA mutagens and comparison of plant strains deficient in specific DNA repair pathways. Ionizing radiation and a variety of genotoxins specifically induce DSBs and are frequently analyzed together with the action of radiomimetic compounds, such as bleomycin (e.g., Menke et al., 2001; Waterworth et al., 2009; Böhmdorfer et al., 2011; Wang et al., 2014), zeocin (Nishiguchi et al., 2012), or MMS (e.g., Menke et al., 2001; Vajpayee et al., 2006; Waterworth et al., 2009). Other mutagens frequently used to study DNA repair and strand breaks include agents that induce point mutations, e.g., N-ethyl-N-nitrosourea (ENU), MNU, or EMS (e.g., Menke et al., 2001; Donà et al., 2013), and the DNA crosslinking agent MMC (e.g., Koppen and Verschaeve, 1996; Menke et al., 2001).
In the past, plant strains deficient in DNA repair pathways have been analyzed by comet assay for their DNA repair capability under specific genotoxic stress. The first observation of biphasic DSB repair in plants with extremly rapid first phase was by Kozak et al. (2009). This approach, led to the identification of A. thaliana AtRad18 (SMC6B) and AtRad21.1 (SYN2) as important effectors in early repair of DSBs, after treatment with bleomycin (Kozak et al., 2009). Also important, through the use of comet functional assays, Moreno-Romero et al. (2012) showed that Arabidopsis mutant plants quickly repaired the DNA damage produced by bleomycin and γ-rays, and that they showed preferential use of non-conservative mechanisms. Moreover, in Arabidopsis knock-down strains of DNA ligase I, Waterworth et al. (2009) found by neutral comet assay that the LIG1 knock-down strains were less efficient in the repair of DSBs compared to wild-type, suggesting that the AtLIG1 gene is involved also in DSB repair pathway.
Several transcripts related with DNA damage pathways, DNA replication, and repair, oxidative stress and cell cycle progression have been identified in plant cells associated with alterations in comet assay profiles. Some of the most relevant studies in wildtype plants are summarized in Table 2. For example, Endo et al. (2006) demonstrated that Arabidopsis fas mutants showed increased levels of DNA DSBs. The authors proposed that the induction of DNA DSBs and enhanced transcription of genes involved in Homologous Recombination (HR) might occur during S phase and stimulate HR in fas mutants. Also, levels of formed DSBs were compared in rice wild type plants vs. an aphidicolin-sensitive phenotype. Without aphidicolin treatment, both WT and osrecql4-2 mutants produced very low levels of DSBs, but these increased in the mutants after treatment (Kwon et al., 2013).
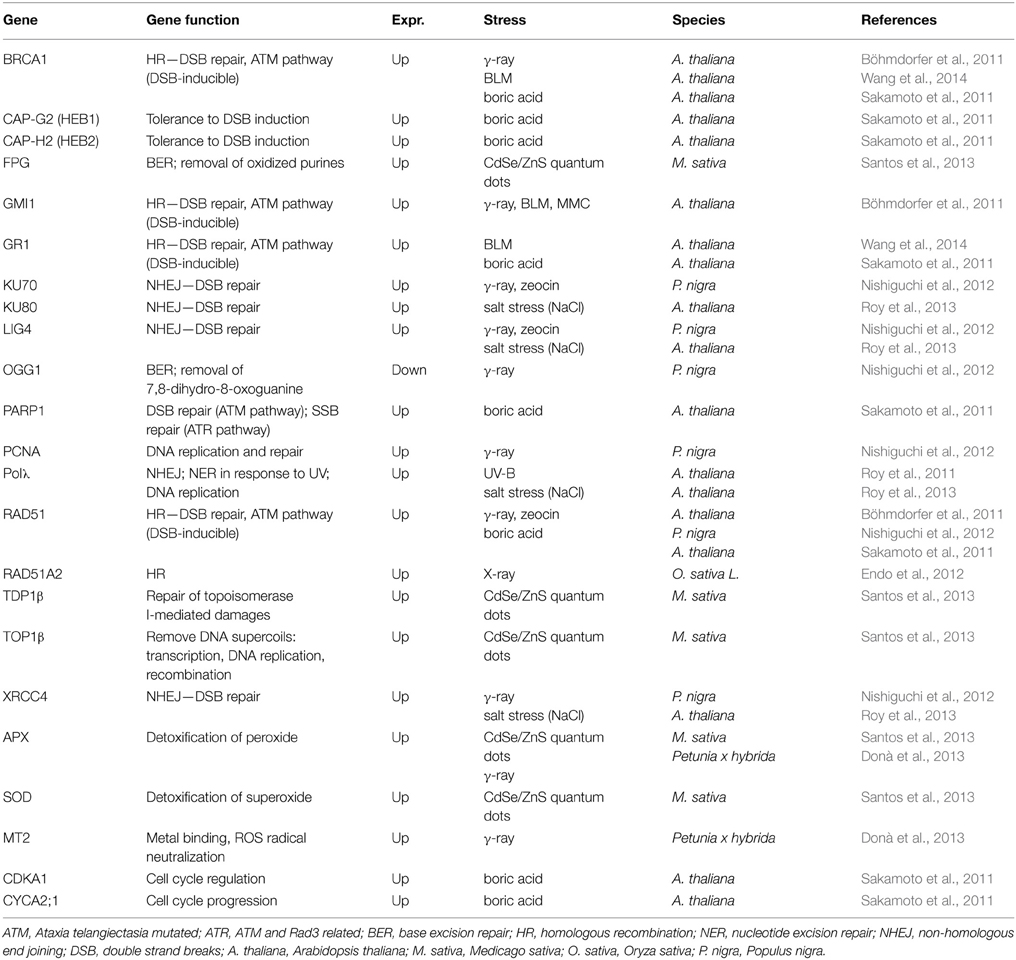
Table 2. Genes differentially expressed in comet assay positive plants.
Böhmdorfer et al. (2011) studied the involvement of γ-irradiation and MMC induced one protein (GMI1), a structural-maintenance-of-chromosomes-hinge domain-containing protein in mechanisms of somatic homologous recombination in Arabidopsis mutant lines. Comet assay demonstrated that the gmi1 mutants had a reduced rate of DNA DSB repair during the early recovery phase after exposure to bleomycin. Also Yao et al. (2013) used the comet assay to show an increase of DNA damage levels in Arabidopsis sdg2 mutants, containing a mutation at SET DOMAIN GROUP 2, necessary for global genome wide deposition of histone H3 lysine 4 trimethylation in chromatin. With these results, authors contributed to elucidate the regulation of SDG2-mediated H3K4me3 on chromatin structure and genome integrity in plants.
Sakamoto et al. (2011) studied Arabidopsis mutants (heb1-1 and heb2-1) hypersensitive to excess of boron (B). Excess of B induced DNA damages and affected the expression of HEB1 and HEB2, which encode respectively the CAP-G2 and CAP-H2 subunits of the condensin II protein complex, important in maintenance of chromosome structure. These results suggested that DSBs are a cause of B toxicity and that condensin II reduces the incidence of DSBs (Sakamoto et al., 2011).
Santos et al. (2013) demonstrated in Medicago sativa that exposure to increasing concentrations of MPA-CdSe/ZnS quantum dots, led to an increase of DNA damages, and up-regulated the transcription of the DNA repair enzymes formamidopyrimidine DNA glycosylase, tyrosyl-DNA phosphodiesterase I and DNA topoisomerase I.
Roy et al. (2011, 2013) reported that Arabidopsis atpolλ mutant lines exposed to UV-B radiation or to high salinity and MMC treatment s showed higher accumulation of DSBs than wild-type plants and a delayed repair of DSBs. This fact suggested the requirement of Pol λ in DSB repair in plants. Gamma irradiated Populus nigra suspension-cultured cells showed increased levels of DNA damage and increase of the transcripts PnRAD51, PnLIG4, PnKU70, PnXRCC4, and PnPCNA while PnOGG1 mRNA was repressed (Nishiguchi et al., 2012). On the other hand, Donà et al. (2013) tested genotoxic effects of γ-irradiation and found significant fluctuations on the levels of DSB and different capacities of DNA repair, together with dose-rate-dependent changes in the expression of the genes PhMT2 (encoding for a type 2 metallothionein) and PhAPX (encoding for a cytosolic isoform of ascorbate peroxidase).
Probing FISH techniques have been successfully applied to comet assay preparations to detect specific DNA lesions, nuclear organizer regions (NORs) and telomeric regions in V. faba (Menke et al., 2000) or 5S/25S rDNA in Crepis capillaris (Kwasniewska et al., 2012).
Salt, drought and osmotic stress are ever more emerging as abiotic defies intimately related with soil overuse and climate changes (e.g., Santos et al., 2002; Brito et al., 2003). Salt stress induction of DNA damages has been explored in e.g., Arabidopsis mutants by Roy et al. (2013) who supported the role of Polλ in DNA damages repair. Salt stress and/or radiation induction of DNA damages was studied in rice by Macovei and collaborators who also evaluated the expression of OsXPB2, OsXPD, OsTFIIS, and OsTFIIS-like genes (Macovei and Tuteja, 2013; Macovei et al., 2014). Recently, Balestrazzi et al. (2014) demonstrated in Medicago truncatula plants that a prolonged exposure to osmotic stress can cause unwanted DNA damages, while negatively affected the expression profiles of genes involved in DNA repair, namely MtTdp1 (tyrosyl-DNA phosphodiesterase), top1 (DNA topoisomerase I), MtTFIIS (transcription elongation factor II-S) and MtTFIIS-like. So, despite comet assay has not been consistently applied to these environmental stresses in plants, the available data of their interference with DNA integrity, opens a perspective of their use in the near future. Also, Confalonieri et al. (2014) demonstrated that in Medicago truncatula the MtTdp2α-gene overexpression prevented the accumulation of DSBs in absence or presence of osmotic stress, and that the MtMRE11, MtRAD50 and MtNBS1 genes that are involved in DSB sensing/repair, being up-regulated in the MtTdp2α-overexpressing plants grown under physiological conditions, were no further up-regulated under osmotic stress (Confalonieri et al., 2014).
Conclusions
In this review we have highlighted most relevant studies that used comet assay in plants to study the impact of stress conditions on plant DNA damages. This work was mostly focused on the most recent major advances in the last five, regarding conventional and emerging contaminants and complex matrices. The recent advances in the use of the plant comet assay to both a larger number of plant species, and a larger number of conditions, support the use of this technique as a robust and sensitive technique to assess DNA damages induced by stress conditions. Data also support that this simple and robust technique may be a powerful tool to complement conventional and -omics tools in situ environmental pollution monitoring. Moreover, new fields of research using plant comet assay are open, not only in environmental studies, but also in plant physiology, as this technique may help elucidating pathways involved in plant development, cell cycle/programmed cell death, or even plant disease resistance. Also, it remains an important field of research deciphering genetic mechanisms underlying processes related with DNA damage/repair, in which comet assay will have undoubtedly a crucial role.
Conflict of Interest Statement
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Acknowledgments
Thanks are due to Fundação para Ciência e Tecnologia for JO fellowship SFRH/BPD/74868/2010; for COMPETE/FEDER, BioRem, FCT/PTDC/AAC-AMB/112804/2009.
References
Abas, Y., Touil, N., Kirsch-Volders, M., Angenon, G., Jacobs, M., and Famelaer, I. D. (2007). Evaluation of UV damage at DNA level in Nicotiana plumbaginifolia protoplasts using single cell gel electrophoresis. Plant Cell Tiss. Organ. Cult. 91, 145–154. doi: 10.1007/s11240-007-9257-9
Achary, V. M., Jena, S., Panda, K. K., and Panda, B. (2008). Aluminium induced oxidative stress and DNA damages in root cells of Allium cepa L. Ecotoxicol. Environ. Saf. 70, 300–310. doi: 10.1016/j.ecoenv.2007.10.022
Achary, V. M., and Panda, B. B. (2010). Aluminium-induced DNA damages and adaptive response to genotoxic stress in plant cells are mediated through reactive oxygen intermediates Mutagenesis 25, 201–209. doi: 10.1093/mutage/gep063
Achary, V. M., Parinandi, N. L., and Panda, B. B. (2012a). Aluminum induces oxidative burst, cell wall NADH peroxidase activity, and DNA damages in root cells of Allium cepa L. Environ. Mol. Mutagen. 53, 550–560. doi: 10.1002/em.21719
Achary, V. M., Parinandi, N. L., and Panda, B. B. (2012b). Oxidative biomarkers in leaf tissue of barley seedlings in response to aluminum stress. Ecotoxicol. Environ. Saf. 75, 16–26. doi: 10.1016/j.ecoenv.2011.08.015
Achary, V. M., Parinandi, N. L., and Panda, B. B. (2013). Calcium channel blockers protect against aluminium-induced DNA damages and block adaptive response to genotoxic stress in plant cells. Mutat. Res. Genet. Toxicol. Environ. Mutagen. 751, 130–138. doi: 10.1016/j.mrgentox.2012.12.008
Amirthalingam, T., Velusamy, G., and Pandian, R. (2013). Cadmium-induced changes in mitotic index and genotoxicity on Vigna unguiculata (Linn.) Walp. J Environ. Chem. Ecotoxicol. 5, 57–62. doi: 10.5897/JECE11.008
Angelis, K. J., Dusinská, M., and Collins, A. R. (1999). Single cell gel electrophoresis: detection of DNA damage at different levels of sensitivity. Electrophoresis. 20, 2133–2138.
Arasimowicz-Jelonek, M., Floryszak-Wieczorek, J., Deckert, J., Ruciñska-Sobkowiak, R., Gzyl, J., Pawlak-Sprada, S., et al. (2012). Nitric oxide implication in cadmium-induced programmed cell death in roots and signaling response of yellow lupine plants. Plant Physiol. Biochem. 58, 124–134. doi: 10.1016/j.plaphy.2012.06.018
Arya, S. K., and Mukherjee, A. (2014). Sensitivity of Allium cepa and Vicia faba towards cadmium toxicity. J. Soil Sci. Plant Nutr. 14, 447–458.
Ascenso, A., Pinho, S., Euletério, C., Praça, F., Oliveira, H., Bentley, M., et al. (2013). Lycopene from tomatoes: vesicular nanocarrier formulations for dermal delivery. J. Agr. Food Chem. 61, 7284–7293. doi: 10.1021/jf401368w
Azqueta, A., Meier, S., Priestley, C., Guzkow, K., Brunborg, G., Sallette, J., et al. (2011a). The influence of scoring method on variability in results obtained with comet assay. Mutagenesis 26, 393–399. doi: 10.1093/mutage/geq105
Azqueta, A., Shaposhnikov, S., and Collins, A. (2009). “Detection of oxidized DNA using DNA repair enzymes,” in The Comet Assay in Toxicology, 1st Edn. eds A. Dhawan and D. Anderson, (London: RSC Publishing).
Azqueta, A., Shaposhnikov, S., and Collins, A. (2011b). “DNA repair measured by comet assay,” in DNA repair, ed I. Kruman (Rijeka: Intech).
Balestrazzi, A., Confalonieri, M., Macovei, A., and Carbonera, D. (2014). Seed imbibition in Medicago truncatula Gaertn.: expression profiles of DNA repair genes in relation to PEG-mediated stress. J. Plant Physiol. 168, 706–713. doi: 10.1016/j.jplph.2010.10.008
Bandyopadhyay, A., and Mukherjee, A. (2011). Sensitivity of Allium and Nicotiana in cellular and acellular comet assays to assess differential genotoxicity of direct and indirect acting mutagens. Ecotoxicol. Environ. Saf. 74, 860–865. doi: 10.1016/j.ecoenv.2010.12.002
Bhat, T., Ansari, M. Y. K., Choudhary, S., Aslam, R., and Alka, A. (2011). Synergistic cytotoxic stress and DNA damages in clover (Trifolium repens) exposed to heavy metal soil from automobile refining shops in Kashmir-Himalaya. ISRN Toxicol. 2011:109092. doi: 10.5402/2011/109092
Bilichak, A., Yao, Y., Titov, V., Golubov, A., and Kovalchuk, I. (2014). Genome stability in the uvh6 mutant of Arabidopsis thaliana. Plant Cell Rep. 33, 979–991. doi: 10.1007/s00299-014-1580-0
Boccia, P., Meconi, C., Mecozzi, M., and Sturchio, E. (2013). Molecular modifications induced by inorganic arsenic in Vicia faba investigated by FTIR, FTNIR spectroscopy and genotoxicity testing. Toxicol. Environ. Health A. 76, 281–290. doi: 10.1080/15287394.2013.757211
Böhmdorfer, G., Schleiffer, A., Brunmeir, R., Ferscha, S., Nizhynska, V., Kozák, J., et al. (2011). GMI1, a structural-maintenance-of-chromosomes-hinge domain-containing protein, is involved in somatic homologous recombination in Arabidopsis. Plant J. 67, 420–433. doi: 10.1111/j.1365-313X.2011.04604.x
Brito, G., Costa, A., Fonseca, H. M. A. C., and Santos, C. (2003). Response of Olea europaea ssp. maderensis in vitro shoots exposed to osmotic stress. Sci. Hortic. 97, 411–417. doi: 10.1016/S0304-4238(02)00216-9
Cenkci, S., Yıldız, M., Ciğerci, I., Bozdağ, A., Terzi, H., and Terzi, E. (2010). Evaluation of 2,4-D and Dicamba genotoxicity in bean seedlings using comet and RAPD assays. Ecotoxicol. Environ. Saf. 73, 1558–1954. doi: 10.1016/j.ecoenv.2010.07.033
Cerda, H., Delincée, H., Haine, H., and Rupp, H. (1997). The DNA ‘comet assay’ as a rapid screening technique to control irradiated food. Mutat. Res. 375, 167–181. doi: 10.1016/S0027-5107(97)00012-2
Cerda, H. v., Hofsten, B., and Johanson, K. (1993). “Identification of irradiated food by microelectrophoresis of DNA from single cells,” in Proceedings of the workshop on Recent Advances on Detection of Irradiated Food, eds M. Leonardi, J. J. Raffi, and J. J. Belliardo (Brussels: Commission of the European Communities), 401–405.
Chakraborty, R., and Mukherjee, A. (2011). Technical note: vetiver can grow on coal fly ash without DNA damages. Int. J. Phytoremediation 13, 206–214. doi: 10.1080/15226510903535171
Chakraborty, R., Mukherjee, A. K., and Mukherjee, A. (2009). Evaluation of genotoxicity of coal fly ash in Allium cepa root cells by combining comet assay with the Allium test. Environ. Monit. Assess. 153, 351–357. doi: 10.1007/s10661-008-0361-z
Ciğerci, I., Cenkci, S., Kargıoglu, M., and Konuk, M. (2014). Genotoxicity of Thermopsis turcica on Allium cepa L. roots revealed by alkaline comet and random amplified polymorphic DNA assays. Cytotechnology. doi: 10.1007/s10616-014-9835-8. [Epub ahead of print].
Ciğerci, I., Liman, R., Ozgül, E., and Konuk, M. (2015). Genotoxicity of indium tin oxide by Allium and Comet tests. Cytotechnology 67, 157–163. doi: 10.1007/s10616-013-9673-0
Collins, A., Azqueta, A., Brunborg, G., Gaivão, I., Giovannelli, L., Kruszewski, M., et al. (2008). The comet assay: topical issues. Mutagenesis 23, 143–151. doi: 10.1093/mutage/gem051
Confalonieri, M., Faè, M., Balestrazzi, A., Donà, M., Macovei, A., Valassi, A., et al. (2014). Enhanced osmotic stress tolerance in Medicago truncatula plants overexpressing the DNA repair gene MtTdp2α (tyrosyl-DNA phosphodiesterase 2). Plant Cell Tiss. Organ. Cult. 116, 187–203. doi: 10.1007/s11240-013-0395-y
Defaveri, T. M., da Silveira, F. Z., Bortolotto, T., Geremias, R., Zocche, J. J., and Pich, C. T. (2009). “Evaluation of acid mine drainage treatment using Artemia sp. and Allium cepa as bioindicators of toxicity and genotoxicity,” in Joint Conference of the 26th Annual Meetings of the American Society of Mining and Reclamation and 11th Billings Land Reclamation Symposium (Billings). doi: 10.13140/2.1.4068.9929
Ding, L., Jing, H., Qin, B., Qi, L., Li, J., Wang, T., et al. (2010b). Regulation of cell division and growth in roots of Lactuca sativa L. seedlings by the ent-kaurane diterpenoid rabdosin B. J. Chem. Ecol. 36, 553–563. doi: 10.1007/s10886-010-9783-5
Ding, L., Jing, H., Wang, T., Li, J., and Liu, G. (2010a). Regulation of root growth in Lactuca sativa L. seedlings by the ent-kaurane diterpenoid Epinodosin. J. Plant Growth Regul. 29, 419–427. doi: 10.1007/s00344-010-9154-z
Donà, M., Ventura, L., Balestrazzi, A., Buttafava, A., Carbonera, D., Confalonieri, M., et al. (2014). Dose-dependent reactive species accumulation and preferential double-strand breaks repair are featured in the γ-ray response in Medicago truncatula cells. Plant Mol. Biol. Rep. 32, 129–141. doi: 10.1007/s11105-013-0635-7
Donà, M., Ventura, L., Macovei, A., Confalonieri, M., Savio, M., Giovannini, A., et al. (2013). Gamma irradiation with different dose rates induces different DNA damages responses in Petunia x hybrid cells. J. Plant Physiol. 170, 780–787. doi: 10.1016/j.jplph.2013.01.010
Endo, M., Ishikawa, Y., Osakabe, K., Nakayama, S., Kaya, H., Araki, T., et al. (2006). Increased frequency of homologous recombination and T-DNA integration in Arabidopsis CAF-1 mutants. EMBO J. 25, 5579–5590. doi: 10.1038/sj.emboj.7601434
Endo, M., Nakayama, S., Umeda-Hara, C., Ohtsuki, N., Saika, H., Umeda, M., et al. (2012). CDKB2 is involved in mitosis and DNA damages response in rice. Plant J. 69, 967–977. doi: 10.1111/j.1365-313X.2011.04847.x
Enseit, J., and Collins, A. R. (2015). DNA repair after X-irradiation: lessons from plants. Mutagenesis 30, 45–50. doi: 10.1093/mutage/geu054
Ezaki, B., Jayaram, K., Higashi, A, and Takahashi, K. (2013). A combination of five mechanisms confers a high tolerance for aluminum to a wild species of Poaceae, Andropogon virginicus L. Environ. Exp. Bot. 93, 35–44. doi: 10.1016/j.envexpbot.2013.05.002
Faè, M., Balestrazzi, A., Confalonieri, M., Donà, M., Macovei, A., Valassi, A., et al. (2014). Copper-mediated genotoxic stress is attenuated by the overexpression of the DNA repair gene MtTdp2a (tyrosyl-DNA phosphodiesterase 2) in Medicago truncatula plants. Plant Cell Rep. 33, 1071–1080 doi: 10.1007/s00299-014-1595-6
Faisal, M., Saquib, Q., Alatar, A. A., Al-Khedhairy, A. A., Hegazy, A. K., and Musarrat, J. (2013). Phytotoxic hazards of NiO-nanoparticles in tomato: a study on mechanism of cell death. J. Hazard. Mater. 250–251, 318–332. doi: 10.1016/j.jhazmat.2013.01.063
Ferreira de Oliveira, J. M. P., Costa, E., Pedrosa, T., Pinto, P., Remedios, C., Oliveira, H., et al. (2014). Sulforaphane-induced cell death is associated with increased oxidative. PLoS ONE 9:e92980. doi: 10.1371/journal.pone.0092980
Garaj-Vrhovac, V., Orešèanin, V., Gajski, G., Geriæ, M., Ruk, D., Kollar, R., et al. (2013). Toxicological characterization of the landfill leachate prior/after chemical and electrochemical treatment: a study on human and plant cells. Chemosphere 93, 939–945. doi: 10.1016/j.chemosphere.2013.05.059
Georgieva, M., and Stoilov, L. (2008). Assessment of DNA strand breaks induced by bleomycin in barley by the comet assay. Environ. Mol. Mutagen. 49, 381–387. doi: 10.1002/em.20396
Ghosh, M., Bhadra, S., Adegoke, A., Bandyopadhyay, M., and Mukherjee, A. (2015a). MWCNT uptake in Allium cepa root cells induces cytotoxic and genotoxic responses and results in DNA hyper-methylation. Mutat. Res. Genet. Toxicol. Environ. Mutagen. 774, 49–58. doi: 10.1016/j.mrfmmm.2015.03.004
Ghosh, M., Bandyopadhyay, M., and Mukherjee, A. (2010). Genotoxicity of titanium dioxide (TiO2) nanoparticles at two trophic levels: plant and human lymphocytes. Chemosphere 81, 1253–1262. doi: 10.1016/j.chemosphere.2010.09.022
Ghosh, M., Chakraborty, A., Bandyopadhyay, M., and Mulherjee, A. (2011). Multi-walled carbon nanotubes (MWCNT): induction of DNA damages in plantand mammalian cells. J. Hazard. Mater. 197, 327–336. doi: 10.1016/j.jhazmat.2011.09.090
Ghosh, M., Sinha, J. M., Chakraborty, A., Malick, S. K., Bandyopadhyay, M., and Mulherjee, A. (2012a). In vitro and in vivo genotoxicity of silver nanoparticles. Mutat. Res. Genet. Toxicol. Environ. Mutagen. 749, 60–69. doi: 10.1016/j.mrgentox.2012.08.007
Ghosh, S., Chatterjee, T., Saha, T., and Mukherjee, A. (2012b). Genotoxicity assessment of soil contamination: a case study from Farakka coal-fired power plant in eastern India. Nucleus 55, 45–50. doi: 10.1007/s13237-012-0052-6
Gichner, T. (2003a). Differential genotoxicity of ethyl methanesulphonate,N-ethyl-N-nitrosourea and maleic hydrazide in tobacco seedlings based on data of the Comet assay and two recombination assays. Mutat. Res. 538, 171–179. doi: 10.1016/S1383-5718(03)00117-7
Gichner, T. (2003b). DNA damages induced by indirect and direct acting mutagens in catalase-deficient transgenic tobacco. Cellular and acellular Comet assays. Mutat. Res. 535, 187–193. doi: 10.1016/S1383-5718(02)00320-0
Gichner, T., Lovecká, P., Kochánková, L., Macková, M., and Demnerová, K. (2007). Monitoring toxicity, DNA damages, and somatic mutations in tobacco plants growing in soil heavily polluted with polychlorinated biphenyls. Mutat. Res. 629, 1–6. doi: 10.1016/j.mrgentox.2006.11.013
Gichner, T., Lovecká, P., and Vrchotova, B. (2008b). Genomic damage induced in tobacco plants by chlorobenzoic acids—Metabolic products of polychlorinated biphenyls. Mutat. Res. Genet. Toxicol. Environ. Mutagen. 657, 140–145. doi: 10.1016/j.mrgentox.2008.08.021
Gichner, T., Patková, K., and Kim, J. (2003). DNA damages measured by the Comet assay in eigh agronomic plants. Biol. Plantarum 47, 185–188. doi: 10.1023/B:BIOP.0000022249.86426.2a
Gichner, T., Patková, Z., Száková, J., and Demnerová, K. (2004). Cadmium induces DNA damages in tobacco roots, but no DNA damages, somatic mutations or homologous recombination in tobacco leaves. Mutat. Res. 559, 49–57. doi: 10.1016/j.mrgentox.2003.12.008
Gichner, T., Patková, Z., Száková, J., and Demnerová, K. (2006). Toxicity and DNA damages in tobacco and potato plants growing on soil polluted with heavy metals. Ecotoxicol. Environ. Saf. 65, 420–426. doi: 10.1016/j.ecoenv.2005.08.006
Gichner, T., Patková, Z., Szaková, J., Znidar, I., and Mukherjee, A. (2008a). DNA damages in potato plants induced by cadmium, ethyl methanesulphonate and γ-rays. Environ. Exp. Bot. 62, 113–119. doi: 10.1016/j.envexpbot.2007.07.013
Gichner, T., and Plewa, M. J. (1998). Induction of somatic DNA damage as measured by single cell gel electrophoresis and point mutation in leaves of tobacco plants. Mutat. Res. 401, 143–152. doi: 10.1016/S0027-5107(98)00003-7
Gichner, T., Ptacek, O., Stavreva, D. A., Wagner, E. D., and Plewa, M. J. (2000). A comparison of DNA repair using the comet assay in tobacco seedlings after exposure to alkylating agents or ionizing radiation. Mutat Res. 470, 1–9. doi: 10.1016/S1383-5718(00)00081-4
Gichner, T., Znidar, I., and Szakova, J. (2008c). Evaluation of DNA damage and mutagenicity induced by lead in tobacco plants. Mutat. Res. Genet. Toxicol. Environ. Mutagen. 652, 186–190. doi: 10.1016/j.mrgentox.2008.02.009
Gichner, T., Znidar, I., Wagner, E., and Plewa, M. (2009). “The use of higher plants in the Comet Assay,” in Issues in Toxicology n° 5 The Comet Assay in Toxicology, eds A. Dhawan and D. Anderson (London: Royal Society of Chemistry), 98–119.
Ghosh, M., Paul, J., Jana, A., De, A., and Mukherjee, A. (2015b). Use of the grass, Vetiveria zizanioides (L.) Nash for detoxification and phytoremediation of soils contaminated with fly ash from thermal power plants. Ecol. Eng. 74, 258–265. doi: 10.1016/j.ecoleng.2014.10.011
Hattab, S., Chouba, L., Kheder, M., and Boussetta, H. (2010). Cadmium− and copper−induced DNA damages in Pisum sativum roots and leaves as determined by the Comet assay. Plant Biosyst. 143, S6–S11. doi: 10.1080/11263500903187035
Holá, M., Kozák, J., Vágnerová, R., and Angelis, K. J. (2013). Genotoxin induced mutagenesis in the model plant Physcomitrella patens. Biomed. Res. Int. 2013:535049. doi: 10.1155/2013/535049
Hovhannisyan, G. (2010). Fluorescence in situ hybridization in combination with the comet assay and micronucleus test in genetic toxicology. Mol. Cytogenet. 3:17. doi: 10.1186/1755-8166-3-17
Hu, Y., Li, J., Yang, L., Nan, W., Cao, X., and Bi, Y. (2014). Inhibition of root growth by narciclasine is caused by DNA damages-induced cell cycle arrest in lettuce seedlings. Protoplasma 51, 1113–1124. doi: 10.1007/s00709-014-0619-y
Jiang, L., Wang, Y., Björn, L. O., and Li, S. (2009). Arabidopsis RADICAL-INDUCED CELL DEATH1 is involved in UV-B signaling. Photochem. Photobiol. Sci. 8, 838–846. doi: 10.1039/b901187k
Jiang, L., Wang, Y., Björn, L. O., and Li, S. (2011). UV-B-induced DNA damage mediates expression changes of cell cycle regulatory genes in Arabidopsis root tips. Planta 233, 831–841. doi: 10.1007/s00425-010-1340-5
Jiang, L., Wang, Y., and Li, S. (2007). Application of the comet assay to measure DNA damage induced by UV radiation in the hydrophyte, Spirodela polyrhiza. Physiol. Plantarum. 129, 652–657. doi: 10.1111/j.1399-3054.2006.00820.x
Jiang, Z., Qin, R., Zhang, H., Zou, J., Shi, Q., Wang, J., et al. (2014). Determination of Pb genotoxic effects in Allium cepa root cells by fluorescent probe, microtubular immunofluorescence and comet assay. Plant Soil 383, 357–372. doi: 10.1007/s11104-014-2183-9
Juchimiuk, J., Gnys, A., and Maluszynska, J. (2006). DNA damages induced by mutagens in plant and human cell nuclei in acellular comet assay. Folia Histochem. Cytobiol. 44, 127–131.
Kamisugi, Y., Schaefer, D. G., Kozak, J., Charlot, F., Vrielynck, N., Holá, M., et al. (2012). MRE11 and RAD50, but not NBS1, are essential for gene targeting in the moss Physcomitrella patens. Nucleic Acids Res. 40, 3496–3510. doi: 10.1093/nar/gkr1272
Kaur, G., Singh, H. P., Batish, D. R., and Kohli, R. K. (2014). Pb-inhibited mitotic activity in onion roots involved DNA damage and disruption of oxidative metabolism. Ecotoxicology 23, 1292–1304. doi: 10.1007/s10646-014-1272-0
Khallef, M., Liman, R., Konuk, M., Ciğerci, I. H., Benouareth, D., Tabet, M., et al. (2013). Genotoxicity of drinking water disinfection by-products (bromoform and chloroform) by using both Allium anaphase-telophase and comet tests. Cytotechnology 67, 207–213. doi: 10.1007/s10616-013-9675-y
Koppen, G., and Angelis, K. J. (1998). Repair of X-ray induced DNA damages measured by the Comet assay in roots of Vicia faba. Environ. Mol. Mutagen. 32, 281–285.
Koppen, G., and Cerda, H. (1997). Identification of low-dose irradiated seeds using the neutral comet assay. Lebenson-Wiss Technol. 30, 452–457.
Koppen, G., Toncelli, L. M., Triest, L., and Verschaeve, L. (1999). The comet assay: a tool to study alteration of DNA integrity in developing plant leaves. Mech. Ageing Dev. 110, 13–24. doi: 10.1016/S0047-6374(99)00038-X
Koppen, G., and Verschaeve, L. (1996). The alkaline comet test on plant cells: a new genotoxicity test for DNA strand breaks in Vicia faba root cells. Mutat. Res. 360, 193–200. doi: 10.1016/S0165-1161(96)90017-5
Koppen, G., and Verschaeve, L. (2001). The alkaline single-cell gel electrophoresis/comet assay: a way to study DNA repair in radicle cells of germinating Vicia faba. Folia Biol. 47, 50–54.
Kozak, J., West, C. E., White, C., da Costa-Nunes, J. A., and Angelis, K. J. (2009). Rapid repair of DNA double strand breaks in Arabidopsis thaliana is dependent on proteins involved in chromosome structure maintenance. DNA Repair 8, 413–419. doi: 10.1016/j.dnarep.2008.11.012
Kumar, A., Prasad, M. N., Achary, M. V., and Panda, B. B. (2013). Elucidation of lead-induced oxidative stress in Talinum triangulare roots by analysis of antioxidant responses and DNA damage at cellular level. Environ. Sci. Pollut. Res. Int. 20, 4551–4561. doi: 10.1007/s11356-012-1354-6
Kwasniewska, J., Grabowska, M., Kwasniewski, M., and Kolano, B. (2012). Comet-FISH with rDNA probes for the analysis of mutagen-induced DNA damages in plant cells. Environ. Mol. Mutagen. 53, 369–375. doi: 10.1002/em.21699
Kwon, Y., Abe, K., Endo, M., Osakabe, K., Namie Ohtsuki, N., Ayako Nishizawa-Yoko, A., et al. (2013). DNA replication arrest leads to enhanced homologous recombination and cell death in meristems of rice OsRecQl4 mutants. BMC Plant Biol. 13:62. doi: 10.1186/1471-2229-13-62
Liman, R. (2013). Genotoxic effects of Bismuth (III) oxide nanoparticles by Allium and Comet assay. Chemosphere 93, 269–273. doi: 10.1016/j.chemosphere.2013.04.076
Liman, R., Ciğerci, I. H., and Öztürk, N.S. (2015). Determination of genotoxic effects of Imazethapyr herbicide in Allium cepa root cells by mitotic activity, chromosome aberration, and comet assay. Pest. Biochem. Physiol. 118, 38–42. doi: 10.1016/j.pestbp.2014.11.007
Liman, R., Ciğerci, I. H., Akyıl, D., Eren, Y., and Konuk, M. (2011). Determination of genotoxicity of fenaminosulf by allium and comet tests. Pest. Biochem. Physiol. 99, 61–64. doi: 10.1016/j.pestbp.2010.10.006
Lin, A., Zhang, X., Chen, C., and Cao, Q. (2007). Oxidative stress and DNA damages induced by cadmium accumulation. J. Environ. Sci. 19, 596–602. doi: 10.1016/S1001-0742(07)60099-0
Lin, A., Zhang, X., Zhu, G., and Zhao, F. (2008). Arsenate-induced toxicity: effects on antioxidative enzymes and DNA damages in Vicia faba. Environ. Toxicol. Chem. 27, 413–419. doi: 10.1897/07-266R.1
Liu, W., Zhu, L., Wang, J., Wang, J. H., Xie, H., and Song, Y. (2009). Assessment of the genotoxicity of Endosulfan in Earthworm and White Clover Plants Using the Comet Assay. Arch. Environ. Contam. Toxicol. 56, 742–746. doi: 10.1007/s00244-009-9309-8
Love, A., Tandon, R., Banerjee, B., and Babu, C. (2009). Comparative study on elemental composition and DNA damages in leaves of a weedy plant species, Cassia occidentalis, growing wild on weathered fly ash and soil. Ecotoxicology 18, 791–801. doi: 10.1007/s10646-009-0322-5
Macovei, A., Garg, B., Raikwar, S., Balestrazzi, A., Carbonera, D., Buttafava, A., et al. (2014). Synergistic exposure of rice seeds to different doses of γ-ray and salinity stress resulted in increased antioxidant enzyme activities and gene-specific modulation of TC-NER pathway. Biomed. Res. Int. 2014:676934. doi: 10.1155/2014/676934
Macovei, A., and Tuteja, N. (2013). Different expression of miRNAs targeting helicases in rice in response to low and high dose rate γ-ray treatments. Plant Signal. Behav. 8:e25128. doi: 10.4161/psb.25128
Manier, N., Brulle, F., LeCurieux, F., Vandenbulcke, F., and Deram, A. (2012). Biomarker measurements in Trifolium repens and Eisenia fetida to assess the toxicity of soil contaminated with landfill leachate: a microcosm study. Ecotoxicol. Environ. Saf. 80, 339–348 doi: 10.1016/j.ecoenv.2012.04.002
Menke, M., Angelis, K. J., and Schubert, I. (2000). Detection of specific DNA lesions by a combination of Comet assay and FISH in plants. Environ. Mol. Mutagen. 35, 132–138. doi: 10.1002/(SICI)1098-2280(2000)35:2<132::AID-EM8>3.0.CO;2-G
Menke, M., Chen, I., Angelis, K., and Schubert, I. (2001). DNA damages and repair in Arabidopsis thaliana as measured by the comet assay after treatment with different classes of genotoxins. Mutat. Res. 493, 87–93. doi: 10.1016/S1383-5718(01)00165-6
Monteiro, C., Santos, C., Pinho, S., Oliveira, H., Pedrosa, T., and Dias, C. (2012). Cadmium-induced cyto- and genotoxicity are organ-dependent in lettuce. Chem. Res. Toxicol. 25, 1423–1434. doi: 10.1021/tx300039t
Moreno-Romero, J., Armengot, L., Mar Marquès-Bueno, M., Britt, A., and Carmen Martínez, M. (2012). CK2-defective Arabidopsis plants exhibit enhanced double-strand break repair rates and reduced survival after exposure to ionizing radiation. Plant J. 71, 627–638. doi: 10.1111/j.1365-313X.2012.05019.x
Navarrete, M. H., Carrera, P., Miguel, M., and Torre, C. (1997). A fast Comet assay variant for solid tissue cells. The assessment of DNA damages in higher plants. Mutat. Res. 389, 271–277. doi: 10.1016/S1383-5718(96)00157-X
Netto, E., Madeira, R., Silveira, F., Fiori, M., Angioleto, E., Pich, C., et al. (2013). Evaluation of the toxic and genotoxic potential of acid mine drainage using physicochemical parameters and bioassays. Environ. Toxicol. Pharmacol. 35, 511–516. doi: 10.1016/j.etap.2013.02.007
Nezames, C. D., Sjogren, C. A., Barajas, J. F., and Larsen, P. B. (2012). The Arabidopsis cell cycle checkpoint regulators TANMEI/ALT2 and ATR mediate the active process of aluminium-dependent root growth inhibition. Plant Cell 243, 608–621. doi: 10.1105/tpc.112.095596
Nishiguchi, M., Nanjo, T., and Yoshida, K. (2012). The effects of gamma irradiation on growth and expression of genes encoding DNA repair-related proteins in Lombardy poplar (Populus nigra var. italica). J. Environ. Radioact. 109, 19–28. doi: 10.1016/j.jenvrad.2011.12.024
Nishioka, M., Ojima, Y., Tsushima, Y., Matsumoto, M., and Taya, M. (2010). Evaluation of photo-induced cellular damage using photoautotrophic cultures of puk-bung hairy roots as a sensing tool. J. Biosci. Bioeng. 109, 392–394. doi: 10.1016/j.jbiosc.2009.09.046
Ojima, Y., Nishioka, M., Matsumoto, M., and Taya, M. (2009). Quantification of DNA damage by the comet assay in radish sprouts exposed to excess light irradiation. Biochem. Eng. J. 46, 69–72. doi: 10.1016/j.bej.2009.04.013
İşeri, O., Korpe, D., Yurtcu, E., Sahin, F., and Haberal, M. (2011). Copper-induced oxidative damages, antioxidant response and genotoxicity in Lycopersicum esculentum Mill and Cucumis sativus L. Plant Cell Rep. 40, 1713–1721. doi: 10.1007/s00299-011-1079-x
Pakrashi, S., Jain, N., Dalai, S., Jayakumar, J., Chandrasekaran, P., Raichur, A., et al. (2014). In vivo genotoxicity assessment of titanium dioxide nanoparticles by Allium cepa root tip assay at high exposure concentrations. PLoS ONE 9:e98828. doi: 10.1371/journal.pone.0087789
Panagos, P., Liedekerke, M., Yigini, Y., and Montanarella, L. (2013). Contaminated sites in Europe: review of the current situation based on data collected through a European network. J. Environ. Public Health 2013:158764. doi: 10.1155/2013/158764
Panda, B. B., and Achary, V. M. (2014). Mitogen-activated protein kinase signal transduction and DNA repair network are involved in aluminium-induced DNA damage and adaptive response in root cells of Allium cepa L. Front. Plant Sci. 5:256. doi: 10.3389/fpls.2014.00256
Paparella, S., Tava, A., Avato, P., Biazzi, E., Macovei, A., Biggiogera, M., et al. (2015). Cell wall integrity, genotoxic injury and PCD dynamics in alfalfa saponin-treated white poplar cells highlight a complex link between molecule structure and activity. Phytochemistry 111, 114–123. doi: 10.1016/j.phytochem.2015.01.008
Patnaik, A. R., Achary, V. M. M., and Panda, B. B. (2013). Chromium (VI)-induced hormesis and genotoxicity are mediated through oxidative stress in root cells of Allium cepa L. Plant Growth Regul. 71, 157–170. doi: 10.1007/s10725-013-9816-5
Petriccione, M., and Ciniglia, C. (2012). Comet assay to assess the genotoxicity of Persian walnut (Juglans regia L.) husks with statistical evaluation. Bull. Environ. Contam. Toxicol. 89, 166–71. doi: 10.1007/s00128-012-0637-4
Poli, P., de Mello, M. A., Buschini, A., Castro, V., Restivo, F., Rossi, C., et al. (2003). Evaluation of the genotoxicity induced by the fungicide fenarimol in mammalian and plant cells by use of the single-cell gel electrophoresis assay. Mutat. Res. 540, 57–66. doi: 10.1016/S1383-5718(03)00165-7
Pourrut, B., Jean, S., Silvestre, J., and Pinelli, E. (2011b). Lead-induced DNA damages in Vicia faba root cells: potential involvement of oxidative stress. Mutat. Res. Genet. Toxicol. Environ. Mutagen. 726, 123–128. doi: 10.1016/j.mrgentox.2011.09.001
Pourrut, B., Pinelli, E., Celiz Mendiola, V., Silvestre, J., and Douay, F. (2015). Recommendations for increasing alkaline comet assay reliability in plants. Mutagenesis 30, 37–43. doi: 10.1093/mutage/geu075
Pourrut, B., Shahid, M., Dumat, C., Winterton, P., and Pinelli, E. (2011a). Lead uptake, toxicity, and detoxification in plants. Rev. Environ. Contam. Toxicol. 213, 113–136. doi: 10.1007/978-1-4419-9860-6_4
Procházková, D., Wilhelmová, N., Pavlíková, D., Száková, J., and Gichner, T. (2013). Zinc induces DNA damages in tobacco roots. Biol. Plantarum 57, 783–787. doi: 10.1007/s10535-013-0345-x
Qin, R., Wang, C., Chen, D., Björn, L. O., and Li, S. (2015). Copper-induced root growth inhibition of Allium cepa var. agrogarum L. involves disturbances in cell division and DNA damage. Environ. Toxicol. 34, 1045–1105. doi: 10.1002/etc.2884
Radić, S., Cvjetko, P., Glavas, K., Roje, V., Pevalek-Kozlina, B., and Pavlica, M. (2009). Oxidative stress and DNA damages in broad bean (Vicia faba l.) seedlings induced by thallium. Environ. Toxicol. Chem. 28, 189–196. doi: 10.1897/08-188.1
Radić, S., Gregoroviæ, G., Stipanièev, D., Cvjetko, P., Šruta, M., Vujèiæ, V., et al. (2013). Assessment of surface water in the vicinity of fertilizer factory using fish and plants. Ecotoxicol. Environ. Saf. 96, 32–40. doi: 10.1016/j.ecoenv.2013.06.023
Radić, S., Stipanicev, D., Cvjetko, P., Mikeliæ, I. L., Marijanoviæ, M., Rajciæ, M. M., et al. (2010). Ecotoxicological assessment of industrial effluent using duckweed (Lemna minor L.) as a test organism. Ecotoxicol 19, 216–222. doi: 10.1007/s10646-009-0408-0
Rodriguez, E. (2011). Genotoxicity and Cytotoxicity of Cr(VI) and Pb2+ in Pisum sativum, PhD Thesis, University Aveiro (English vers.), Available online at: https://ria.ua.pt/bitstream/10773/7402/1/Tese%20Definitiva%20EJR_imprimir.pdf
Rodriguez, E., Azevedo, R., Fernandes, P., and Santos, C. (2011). Cr(VI) induces DNA damages, cell cycle arrest and polyploidization: flow cytometric and comet assay study in Pisum sativum. Chem. Res. Toxicol. 24, 1040–1047. doi: 10.1021/tx2001465
Rounds, M. A., and Larsen, P. B. (2008). Aluminium-dependent root-growth inhibition in Arabidopsis results from AtATR-regulated cell-cycle arrest. Curr. Biol. 18, 1495–1500. doi: 10.1016/j.cub.2008.08.050
Roy, S., Choudhury, S., Sengupta, N., and Das, K. (2011). AtPolλ, a homolog of mammalian DNA polymerase λ in Arabidopsis thaliana, is involved in the repair of UV-B induced DNA damages through the dark repair pathway. Plant Cell Physiol. 52, 448–467. doi: 10.1093/pcp/pcr002
Roy, S., Choudhury, S., Sengupta, N., and Das, K. (2013). Involvement of AtPolλ in the repair of high salt- and DNA cross-linking agent-induced double strand breaks in Arabidopsis. Plant Physiol. 162, 1195–1210. doi: 10.1104/pp.113.219022
Saghirzadeh, M., Gharaati, M. R., Mohammadi, S. H., and Ghiassi-Nejad, M. (2008). Evaluation of DNA damages in the root cells of Allium cepa seeds growing in soil of high background radiation areas of Ramsar—Iran. J. Environ. Radioact. 99, 1698–1702. doi: 10.1016/j.jenvrad.2008.03.013
Sakamoto, T., Inui, Y., Uraguchi, S., Yoshizumi, T., Matsunaga, S., Mastui, M., et al. (2011). Condensin II alleviates DNA damages and is essential for tolerance of boron overload stress in Arabidopsis. Plant Cell 23, 3533–3546. doi: 10.1105/tpc.111.086314
Santos, A. R., Miguel, A. S., Macovei, A., Maycock, C., Balestrazzi, A., Oliva, A., et al. (2013). CdSe/ZnS quantum dots trigger DNA repair and antioxidant enzyme systems in Medicago sativa cells in suspension culture. BMC Biotechnol. 13, 111–118. doi: 10.1186/1472-6750-13-111
Santos, C. V., Falcão, I. P., Pinto, G. C., Oliveira, H., and Loureiro, J. (2002). Nutrient responses and glutamate and proline metabolism in sunflower plants and calli under Na2SO4 stress. J. Plant Nutr. Soil Sci. 165, 366–372. doi: 10.1002/1522-2624(200206)165:3<366::AID-JPLN366>3.0.CO;2-V
Seth, C., Misra, V., Chauhan, L., and Singh, R. (2008). Genotoxicity of cadmium on root meristem cells of Allium cepa: cytogenetic and Comet assay approach. Ecotoxicol. Environ. Saf. 71, 711–716. doi: 10.1016/j.ecoenv.2008.02.003
Silva, S., Pinto-Carnide, O., Martins-Lopes, P., Matos, M., Guedes-Pinto, H., and Santos, C. (2010). Differential aluminium changes on nutrient accumulation and root differentiation in an Al sensitive vs. tolerant wheat. Environ. Exp. Bot. 68, 91–98. doi: 10.1016/j.envexpbot.2009.10.005
Stoilov, L., Georgieva, M., Manova, V., Liu, L., and Gecheff, K. (2013). Karyotype reconstruction modulates the sensitivity of barley genome to radiation-induced DNA and chromosomal damages. Mutagenesis 28, 153–160. doi: 10.1093/mutage/ges065
Sturchio, E., Boccia, P., Meconi, C., Zanellato, M., Marconi, S., Beni, C., et al. (2011). Effects of arsenic on soil-plant systems. Chem. Eco. (Suppl. 1). 27, 67–78. doi: 10.1080/02757540.2010.536762
Tai, H. H., Percy, K. E., and Karnosky, D. F. (2010). DNA damages in Populus tremuloides clones exposed to elevated O3.Environ. Pollut. 158, 969–976. doi: 10.1016/j.envpol.2009.10.005
Thiruvengadam, M., Gurunathan, S., and Chung, I. M. (2015). Physiological, metabolic, and transcriptional effects of biologically-synthesized silver nanoparticles in turnip (Brassica rapa ssp. rapa L.). Protoplasma. doi: 10.1007/s00709-014-0738-5. [Epub ahead of print].
Tkalec, M., Štefaniæ, P., Cvjetko, P., Šikić, S., Pavlica, M., and Balen, B. (2014). The effects of cadmium-zinc interactions on biochemical responses in tobacco seedlings and adult plants. PLoS ONE 9:e87582. doi: 10.1371/journal.pone.0087582
Türkoğlu, S. (2012). Determination of genotoxic effects of chlorfenvinphos and fenbuconazole in Allium cepa root cells by mitotic activity, chromosome aberration, DNA content, and comet assay. Pestic. Biochem. Physiol. 103, 224–230. doi: 10.1016/j.pestbp.2012.06.001
Vajpayee, P., Dhawan, A., and Shanker, R. (2006). Evaluation of the alkaline Comet assay conducted with the wetlands plant Bacopa monnieri L. as a model for ecogenotoxicity assessment. Environ. Mol. Mutagen. 47, 483–489. doi: 10.1002/em.20217
Vandenhove, H., Vanhoudt, N., Cuypers, A., van Hees, M., Wannijn, J., and Horemans, N. (2010). Life-cycle chronic gamma exposure of Arabidopsis thaliana induces growth effects but no discernable effects on oxidative stress pathways. Plant Physiol. Biochem. 48, 778–786. doi: 10.1016/j.plaphy.2010.06.006
Ventura, L., Giovannini, A., Savio, M., Donà, M., Macovei, A., Buttafava, A., et al. (2013). Single Cell Gel Electrophoresis (Comet) assay with plants: research on DNA repair and ecogenotoxicity testing. Chemosphere 92, 1–9. doi: 10.1016/j.chemosphere.2013.03.006
Verbeek, F., Koppen, G., Schaeken, B., and Verschaeve, L. (2008). Automated detection of irradiated food with the comet assay. Radiat. Prot. Dosimetry. 128, 421–426. doi: 10.1093/rpd/ncm433
Wang, Y., Xiao, R., Haifeng Wang, H., Zhihao Cheng, Z., Li, W., Zhu, G., et al. (2014). The Arabidopsis RAD51 paralogs RAD51B, RAD51D and XRCC2 play partially redundant roles in somatic DNA repair and gene regulation. New Phytol. 201, 292–304. doi: 10.1111/nph.12498
Waterworth, W. M., Kozak, J., Provost, C. M., Bray, C. M., Angelis, K. J., and West, C. E. (2009). DNA ligase 1 deficient plants display severe growth defects and delayed repair of both DNA single and double strand breaks. BMC Plant Biol. 9:79 doi: 10.1186/1471-2229-9-79
Watharkar, A., and Jadhav, J. (2014). Detoxification and decolorization of a simulated textile dye mixture by phytoremediation using Petunia grandiflora and, Gailardia grandiflora: a plant–plant consortial strategy. Ecotoxicol. Environ. Saf. 103, 1–8. doi: 10.1016/j.ecoenv.2014.01.033
Yan, S., Wang, W., Marqués, J., Mohan, R., Saleh, A., Durrant, W., et al. (2013). Salicylic acid activates DNA damages responses to potentiate plant immunity. Mol. Cell 52, 602–610. doi: 10.1016/j.molcel.2013.09.019
Yao, X., Feng, H., Yu, Y., Dong, A., and Shen, W. (2013). SDG2-mediated H3K4 methylation is required for proper Arabidopsis root growth and development. PLoS ONE 8:e56537. doi: 10.1371/journal.pone.0056537
Yıldız, M., Ciğerci, I. H., Konuk, M., Fidan, A. F., and Terzi, H. (2009). Determination of genotoxic effects of copper sulphate and cobalt chloride in Allium cepa root cells by chromosome aberration and comet assays. Chemosphere. 75, 934–938. doi: 10.1016/j.chemosphere.2009.01.023
Keywords: plant comet assay, genotoxicity, metal, phytocompounds, radiation, pollutants, nanoparticles, DNA damages biomarkers
Citation: Santos CLV, Pourrut B and Ferreira de Oliveira JMP (2015) The use of comet assay in plant toxicology: recent advances. Front. Genet. 6:216. doi: 10.3389/fgene.2015.00216
Received: 17 September 2014; Accepted: 02 June 2015;
Published: 30 June 2015.
Edited by:
Sabine Langie, Flemish Institute for Technological Research, BelgiumReviewed by:
Karel J. Angelis, Institute of Experimental Botany Academy of Sciences of the Czech Republic, Czech RepublicGudrun Koppen, Flemish Institute for Technological Research, Belgium
Copyright © 2015 Santos, Pourrut and Ferreira de Oliveira. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) or licensor are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Conceição L. V. Santos, Department of Biology, Faculty of Sciences, University of Porto, Rua do Campo Alegre 1021/1055, 4169-007 Porto, Portugal,Y3NhbnRvc0BmYy51cC5wdA==