 Tanel Traks1,2*
Tanel Traks1,2* Maris Keermann1,3
Maris Keermann1,3 Maire Karelson1,3
Maire Karelson1,3 Ranno Rätsep4Ene Reimann5,6Helgi Silm1,3
Ranno Rätsep4Ene Reimann5,6Helgi Silm1,3 Eero Vasar2,4
Eero Vasar2,4 Sulev Kõks2,5,6Külli Kingo1,3
Sulev Kõks2,5,6Külli Kingo1,3- 1Department of Dermatology, University of Tartu, Tartu, Estonia
- 2Centre of Excellence for Translational Medicine, University of Tartu, Tartu, Estonia
- 3Clinic of Dermatology, Tartu University Hospital, Tartu, Estonia
- 4Department of Physiology, University of Tartu, Tartu, Estonia
- 5Department of Pathophysiology, University of Tartu, Tartu, Estonia
- 6Department of Reproductive Biology, Estonian University of Life Sciences, Tartu, Estonia
Background: The members of Toll-like receptor (TLR) family are responsible for recognizing various molecular patterns associated with pathogens. Their expression is not confined to immune cells and have been detected in skin cells such as keratinocytes and melanocytes. As part of a generated response to pathogens, TLRs are involved in inducing inflammatory mediators to combat these threats. It is therefore not surprising that TLRs have been implicated in inflammatory skin diseases, including atopic dermatitis and psoriasis. Likewise, as key players in autoimmunity, they have been associated with a number of autoimmune diseases. Based on this, the role of TLRs in vitiligo could be suspected, but is yet to be clearly established.
Methods: In order to conduct a genetic association analysis, 30 SNPs were selected from TLR1-TLR8 and TLR10 regions to be genotyped in Estonian case-control cohort consisting of 139 vitiligo patients and 307 healthy control individuals. The patients were further analyzed in subgroups based on sex, age of onset, occurrence of vitiligo among relatives, extent of depigmented areas, vitiligo progression activity, appearance of Köbner's phenomenon, existence of halo naevi, and incidence of spontaneous repigmentation.
Results: The most notable finding came with SNP rs179020 situated in TLR7 gene, that was associated in entire vitiligo (Padj = 0.0065) and also several subgroup analyses. Other single marker and haplotype analyses pointed to TLR3, TLR4, and TLR10 genes.
Conclusions: This study investigated the genetic regions of nine TLR genes in relation to vitiligo susceptibility. The main results were the associations of TLR7 SNPs with vitiligo, while several other associations were obtained from the remaining TLR gene regions. This suggests that in addition to other inflammatory skin diseases, TLRs affect the development of vitiligo, thus making them interesting targets for future research.
Introduction
Vitiligo is a chronic disease manifested by distinctive lightening of the skin and the cause to these depigmented areas is in the loss of melanocytes. It is considered to be a complex disease, as there is a multitude of genetic and environmental factors involved in this process (Boissy and Spritz, 2009). The precise set of contributors is yet to be determined, but the general consensus approaches it as an autoimmune disease (Le Poole and Luiten, 2008). This notion is supported by the findings that vitligo susceptibilty genes primarily include immune-related genes and by the frequent co-occurrence with other autoimmune diseases (Spritz, 2011). It also implies that a shared genetic background with different self-reactive pathologies can be expected. At the same time, the importance of environmental risk factors remains, in that the concordance in monozygotic twins is only 23% (Alkhateeb et al., 2003).
Toll-like receptors (TLRs) function as critical components of innate immune system by recognizing various molecular motifs associated with pathogens or tissue injury (Takeda et al., 2003). This leads to the induction of inflammatory and antimicrobial innate immune responses as well as antigen-specific adaptive immune mechanisms (Akira and Takeda, 2004). Crucial to this system is the discrimination between self and foreign antigens and regulatory control over downstream immune activation. The breakdown of these processes causes autoimmunity and TLRs are suggested to have a pivotal function in this pathology (Pradhan et al., 2012; Liu et al., 2014). Indeed, they have been associated with various autoimmune diseases and vitiligo among them (Kang et al., 2009; Yu et al., 2009). Specifically, TLR3 that is expressed in melanocytes may sense viral infection and induce apoptosis along with local immune response, that contributes to vitiligo development (Yu et al., 2009). TLR4 in melanocytes could react to endogenous heat-shock proteins and initiate autoimmunity (Yu et al., 2009). Activation of TLR7 may trigger melanocyte apoptosis and thus the appearance of vitiligo symptoms (Kang et al., 2009).
The expression of TLRs has been detected in keratinocytes, melanocytes, and Langerhans cells of the skin (Hari et al., 2010). As could be expected, they have been implicated in a several inflammatory skin diseases such as atopic dermatitis, psoriasis, and acne vulgaris (Miller, 2008; Hari et al., 2010). On a genetic level, only a few studies have been conducted, yielding support for TLR2 involvement in atopic dermatitis (Oh et al., 2009) and TLR9 in psoriasis treatment response (Romaní et al., 2015). Still, a recent report has found TLR2 and TLR4 single-nucleotide polymorphisms (SNPs) to be associated with vitiligo (Karaca et al., 2013).
Considering this background, the aim of this study was to explore possible genetic associations between TLR gene polymorphisms and vitiligo. Thirty SNPs from six genetic loci containing TLR1-TLR8 and TLR10 genes were selected for genotyping and subsequent association analyses.
Materials and Methods
Study Sample
To assemble the study sample, 139 vitiligo patients and 307 healthy control individuals were enrolled at the Department of Dermatology, University of Tartu, Estonia. All subjects were unrelated, of Caucasian origin, and living in Estonia. Vitiligo diagnosis was based on characteristic skin depigmentation at typical locations and whiteness of skin lesions under Wood's lamp. To conduct additional analyses, the patients were separated into subgroups according to several characteristics. Female (n = 94) and male (n = 45) patients were analyzed independently against their respective controls (n = 168; n = 139). Early onset vitiligo (n = 41) was assigned in case the symptoms appeared before the age of 20 and late onset (n = 98) in case 20 or after. Familial vitiligo (n = 36) was determined by occurrence of vitiligo in patients' relatives and the absence of vitiligo among them indicated the sporadic cases (n = 102). One patient fell in neither category, since family data was not available. The extent of affected areas was the basis of next two groups: extent < 10% (n = 71) and extent ≥ 10% (n = 68). The patients were classified to have active vitiligo (n = 96) in case new areas of depigmentation had appeared during the previous 3 months and stable vitiligo (n = 43) if new areas or enlargement of previously existing depigmentation had not occurred during this period. Patients with Köbner's phenomenon, manifested by development of new vitiligo patches at sites of skin injury, comprised the Köbner positive group (n = 23). The occurrence of halo naevi (n = 19) and spontaneous repigmentation (n = 38) were the last distinguishing factors. The control group was recruited at University of Tartu from medical students, health care personnel and patients presenting at the dermatological outpatient clinic with mild expression of either facial teleangiectasis or skin tags.
The Human Research Ethics Committee of the University of Tartu approved the study and informed consent was obtained from all participants.
SNP Selection and Genotyping
SNPbrowser version 3.5 was used for SNP selection and for SNPlex™ (Applied Biosystems) assay pool design. The SNPs were located in six loci that contain the genes TLR1-TLR8 and TLR10 (Table 1). The SNPs were selected to evenly cover each locus and non-synonymous SNPs were always preferred. Genomic DNA was extracted from 9 ml blood samples and Applied Biosystems SNPlex™ method was used for genotyping (Tobler et al., 2005).
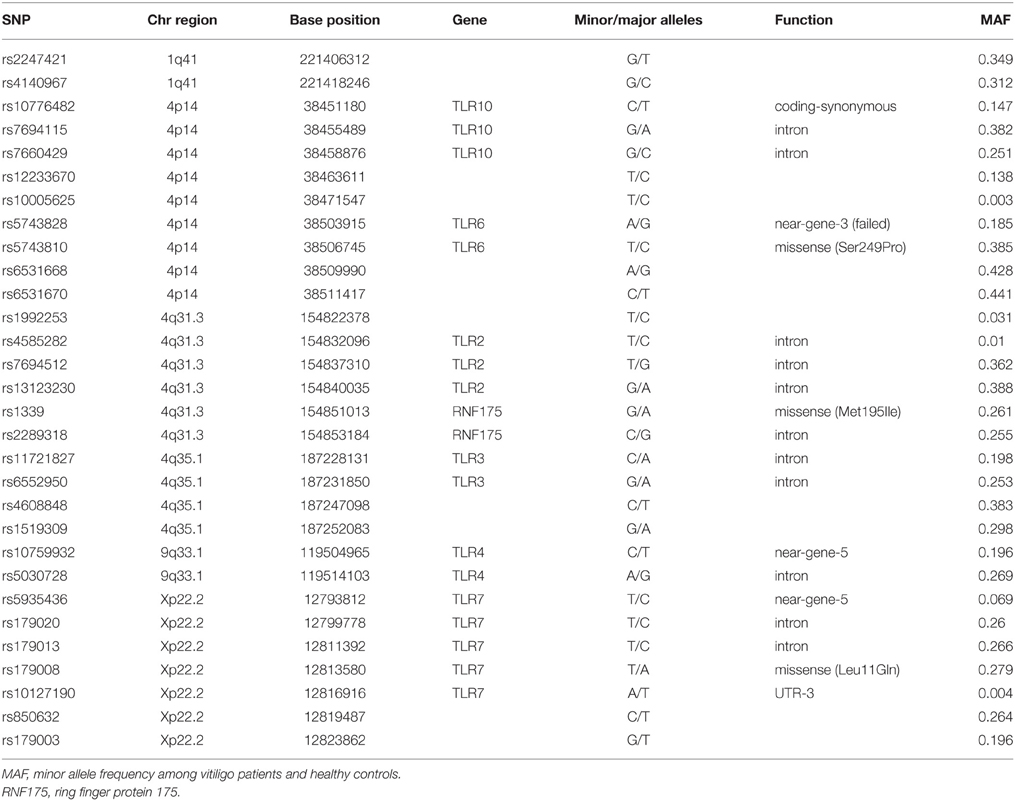
Table 1. Characteristics of studied SNPs.
Data Analysis
The Haploview v4.2 program was used for Hardy-Weinberg equilibrium (HWE) calculations in control group and also for allelic association and haplotype association tests between groups of patients and controls (Barrett et al., 2005). The Solid spine of LD algorithm integrated in Haploview v4.2 was applied to define the haplotype blocks and the resulting blocks were used in the haplotype association test. Differences in allele or haplotype frequencies between cases and controls were assessed by chi square test. The statistical significance threshold was set to 0.05 for all tests. Ten thousand permutations were performed to correct p-values for errors of multiple testing. SNPs of each of the studied regions were analyzed separately.
Results
The genotyping procedure provided data for 30 SNPs and 29 of them qualified for subsequent statistical analysis. A single SNP rs5743828 was excluded due to deviation from Hardy-Weinberg equilibrium. All the remaining markers met the inclusion criteria for minor allele frequency (MAF > 1%) and Hardy-Weinberg equilibrium (Hardy-Weinberg p > 0.01).
Allelic Association Analysis
Single marker associations were present in all studied regions except for chromosome 1q41. Four associations were revealed when analysing the entire vitiligo group and the rest were produced when analysing by different subgroups. The results obtained from entire vitiligo, early onset, late onset, familial, sporadic, extent < 10%, extent ≥ 10%, active, and stable vitiligo analyses are presented in Table 2.
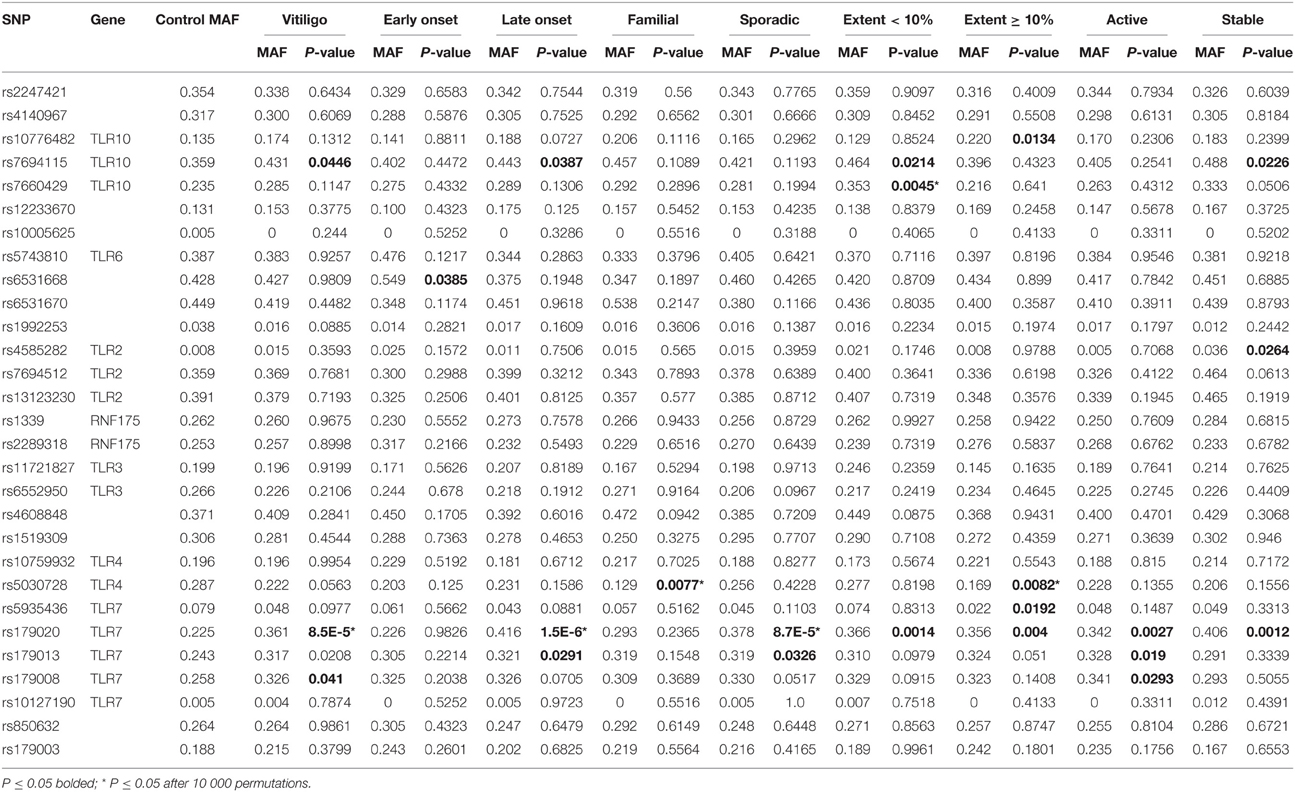
Table 2. Results of allelic association analysis.
The SNPs of region 4p14 provided 10 associations. SNP rs7694115 was associated in entire vitiligo group [p = 0.0446, odds ratio (OR) 1.35, 95% confidence interval (CI) 1.01–1.81] and also in stable (p = 0.0226, OR 1.7, CI 1.07–2.69), male (p = 0.006, OR 1.96, CI 1.21–3.17), extent < 10% (p = 0.0214, OR 1.55, CI 1.06–2.24), and late onset (p = 0.0387, OR 1.42, CI 1.02–1.97) subgroups. SNP rs10776482 was associated in extent ≥ 10% (p = 0.0134, OR 1.81, CI 1.13–2.9) and Köbner (p = 0.023, OR 2.28, CI 1.1–4.71) subgroups. SNP rs7660429 was associated in extent < 10% (p = 0.0045, OR 1.77, CI 1.19–2.64), rs12233670 in male (p = 0.0095, OR 2.28, CI 1.21–4.29), and rs6531668 in early onset (p = 0.0385, OR 1.63, CI 1.02–2.59) subgroups. The association of rs7660429 in extent < 10% remained significant after correcting for multiple testing (p = 0.0469).
In 4q31.3 region, a single SNP rs4585282 was associated in stable (p = 0.0264, OR 4.5, CI 1.05–19.17) and Köbner (p = 0.0162, OR 6.07, CI 1.14–32.27) subgroups.
A single SNP rs6552950 of 4q35.1 region was associated in male subgroup (p = 0.0213, OR 0.49, CI 0.27–0.91).
Next, a single SNP rs5030728 of 9q33.1 region was associated in female (p = 0.0384, OR 0.62, CI 0.39–0.98), familial (p = 0.0077, OR 0.37, CI 0.17–0.79), and extent ≥ 10% (p = 0.0082, OR 0.51, CI 0.3–0.84) subgroups. The last two associations remained significant after correcting for multiple testing (p = 0.0165 and p = 0.0227, respectively).
The most notable results occurred in Xp22.2 region and concerned the SNP rs179020. It was associated in entire vitiligo group (p = 8.5E-5, OR 1.95, CI 1.39–2.73) and also active (p = 0.0027, OR 1.8, CI 1.22–2.64), stable (p = 0.0012, OR 2.36, CI 1.39–4.03), female (p = 0.0483, OR 1.52, CI 1–2.31), male (p = 8.8E-5, OR 3.1, CI 1.73–5.55), sporadic (p = 8.7E-5, OR 2.1, CI 1.44–3.06), extent < 10% (p = 0.0014, OR 1.99, CI 1.3–3.06), extent ≥ 10% (p = 0.004, OR 1.91, CI 1.22–2.97), Köbner (p = 0.0265, OR 2.3, CI 1.08–4.9), halo (p = 0.0056, OR 2.69, CI 1.3–5.54), and late onset (p = 1.5E-6, OR 2.46, CI 1.69–3.56) subgroups. The results in entire vitiligo, male, sporadic, and late onset groups remained significant after correcting for multiple testing (p = 0.0065, p = 0.0479, p = 0.0078, and p = 0.0004, respectively). In addition, two other SNPs on this region were associated in entire vitiligo group. These were rs179013 (p = 0.0208, OR 1.45, CI 1.06–1.98) and rs179008 (p = 0.041, OR 1.39, CI 1.01–1.9). The former was also associated in active (p = 0.019, OR 1.52, CI 1.07–2.17), sporadic (p = 0.0326, OR 1.46, CI 1.03–2.07) and late onset (p = 0.0291, OR 1.48, CI 1.04–2.1) vitiligo subgroups and the latter in active vitiligo subgroup (p = 0.0293, OR 1.48, CI 1.04–2.12). Finally, rs5935436 was associated with vitiligo in females (p = 0.0102, OR 0.3, CI 0.11–0.79) and in extent ≥ 10% group (p = 0.0192, OR 0.27, CI 0.08–0.87).
Haplotype Association Analysis
Haplotype blocks were formed by SNPs of all studied regions. Similarly, to single marker analysis, haplotype associations were revealed in all regions except for 1q41. Seven associations were obtained in entire vitiligo group and others occurred in different vitiligo subgroups. The composition of haplotype blocks and results of haplotype association analysis in entire vitiligo group are presented in Table 3.
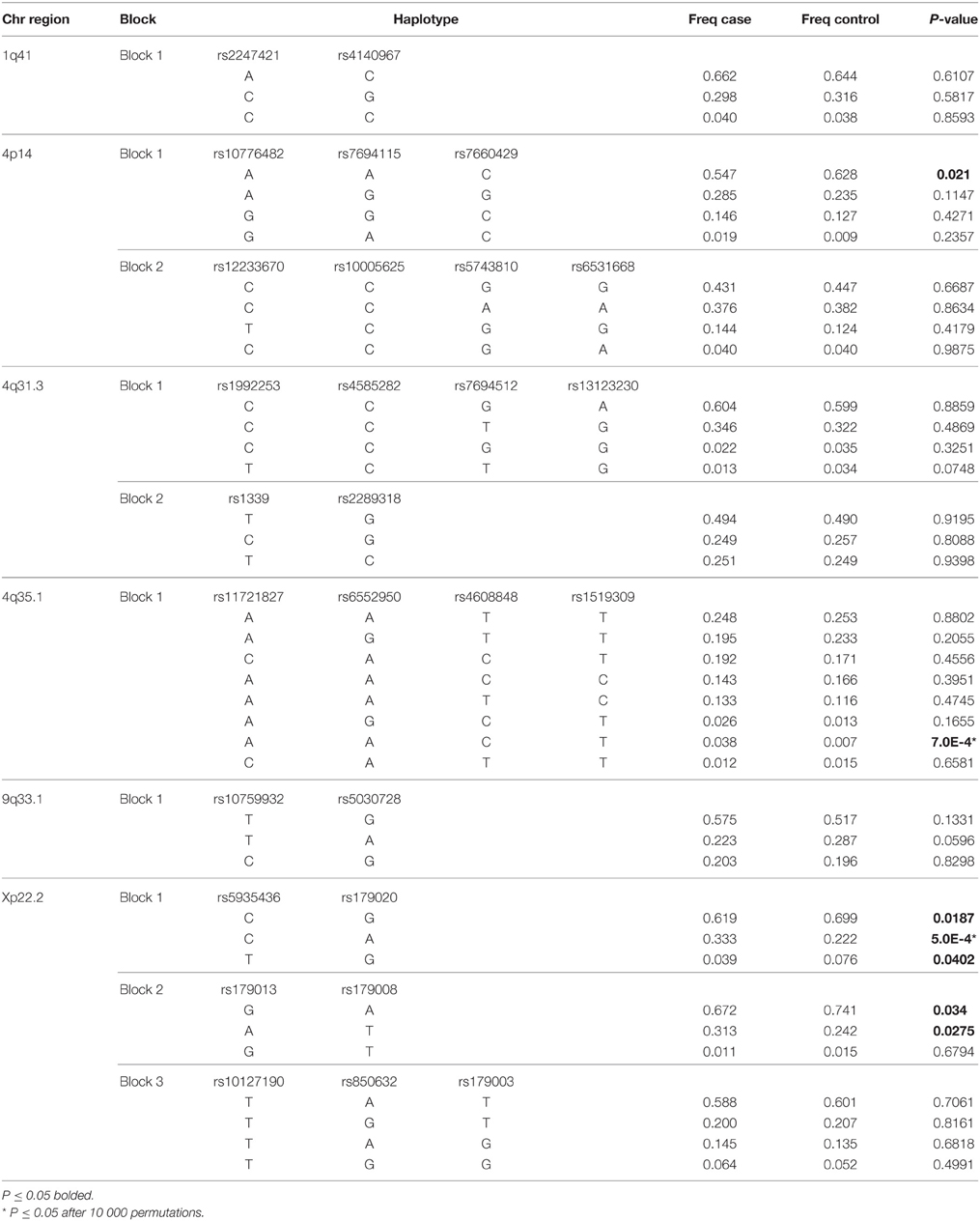
Table 3. Results of haplotype analysis in entire vitiligo group.
Haplotype AAC of 4p14 region that included alleles of rs10776482, rs7694115, and rs7660429 was associated in entire vitiligo group (p = 0.021, OR 0.71, CI 0.53–0.95) along with stable (p = 0.0228, OR 0.59, CI 0.37–0.93), male (p = 0.0103, OR 0.54, CI 0.33–0.87), extent < 10% (p = 0.019, OR 0.64, CI 0.44–0.93), Köbner (p = 0.0427, OR 0.54, CI 0.3–0.99), and late onset (p = 0.0149, OR 0.67, CI 0.48–0.93) subgroups. Haplotype AGG of the same block was associated in extent < 10% subgroup (p = 0.0031, OR 1.8, CI 1.22–2.67) and this remained significant after correcting for multiple testing (p = 0.0331). The other block of this region was formed by rs12233670, rs10005625, rs5743810, and rs6531668. Exception to this was the male vitiligo analysis, where rs6531670 was also added to this block and haplotype TCGGT provided the only association (p = 0.021, OR 2.15, CI 1.11–4.16). The other exception in block formation was the female vitiligo analysis where the first block included rs10776482, rs7694115, rs7660429, rs12233670, and rs5743810, and the second block included rs6531668 and rs6531670.
Two haplotype blocks were formed by SNPs of 4q31.3 region. The first block was usually composed of rs1992253, rs4585282, rs7694512, and rs13123230 and the second block of rs1339 and rs2289318. This was not the case in stable, extent < 10% and early onset analyses, where the last SNP of block 1 rs13123230 was transferred to block 2. Additionally, in familial vitiligo analysis rs7694512 of block 1 was also transferred to block 2 and in male vitiligo analysis the first SNP of block 1 was omitted, leaving rs4585282, rs7694512, and rs13123230 in that block. The only haplotype associations found in this region came from block 1, where haplotypes CCG and CCT were associated with stable vitiligo (p = 0.034, OR 0.61, CI 0.39–0.97 and p = 0.0217, OR 1.7, CI 1.07–2.68, respectively).
The SNPs of 4q35.1 region formed a single haplotype block. It contained all four SNPs of this region, except for male vitiligo analysis, where the last SNP rs1519309 was not included in the block. The strongest associations were uncovered for haplotype AACT in entire vitiligo group (p = 7.0E-4, OR 5.84, CI 1.85–18.45) and in active (p = 1.0E-4, OR 7.22, CI 2.25–23.23), female (p = 0.0018, OR 10.35, CI 1.7–63.1), sporadic (p = 0.0051, OR 5.31, CI 1.44–19.67), extent = 10% (p = 8.0E-4, OR 7.5, CI 1.87–30.05), spontaneous repigmentation (p = 1.0E-4, OR 10.24, CI 2.38–44.02) and late onset (p = 0.0018, OR 5.95, CI 1.69–20.96) subgroups. Notably, all these seven associations remained significant after correcting for multiple testing (p = 0.0068, p = 0.0016, p = 0.0139, p = 0.0394, p = 0.0114, p = 0.0023, and p = 0.0203, respectively). Haplotype AGCT was associated in familial (p = 0.0092, OR 4.3, CI 1.29–14.35), Köbner (p = 9.0E-4, OR 6.13, CI 1.82–20.57) and early onset (p = 0.0306, OR 3.52, CI 1.04–11.9) subgroups. The result in Köbner group remained significant after correcting for multiple testing (p = 0.0141). Haplotypes AAT and AGT were associated with vitiligo in males (p = 0.0225, OR 1.75, CI 1.08–2.83 and p = 0.0286, OR 0.51, CI 0.27–0.94, respectively).
The two SNPs of 9q33.1 region formed a haplotype block in each of the analyzed groups. Haplotype TA was associated in female (p = 0.0401, OR 0.63, CI 0.4–0.98), familial (p = 0.0098, OR 0.39, CI 0.19–0.82) and extent ≥ 10% (p = 0.0086, OR 0.51, CI 0.3–0.85) subgroups. The last two associations remained significant after correcting for multiple testing (p = 0.0198 and p = 0.0234, respectively).
Depending on the analyzed group, two or three haplotype blocks were formed by SNPs of Xp22.2 region. Three blocks were present in entire vitiligo group, and active, male, sporadic, extent < 10% and late onset subgroups. Block 1 included rs5935436 and rs179020, block 2 included rs179013 and rs179008, and block 3 included the remaining rs10127190, rs850632 and rs179003. The strongest associations involved block 1 haplotype CA in entire vitiligo (p = 5.0E-4, OR 1.75, CI 1.28–2.4), active (p = 0.0109, OR 1.59, CI 1.11–2.29), male (p = 0.0026, OR 2.23, CI 1.31–3.79), sporadic (p = 8.0E-4, OR 1.81, CI 1.28–2.57), extent < 10% (p = 0.0094, OR 1.7, CI 1.13–2.54), and late onset (p = 2.6E-5, OR 2.1, CI 1.48–2.98) vitiligo groups. From these, the associations in entire vitiligo, sporadic, and late onset vitiligo groups remained significant after correcting for multiple testing (p = 0.0244, p = 0.04, p = 0.0035, respectively). Haplotype CG of the same block was associated in entire vitiligo (p = 0.0187, OR 0.7, CI 0.52–0.94), male (p = 0.0018, OR 0.45, CI 0.28–0.75), sporadic (p = 0.025, OR 0.68, CI 0.49–0.95), extent < 10% (p = 0.0246, OR 0.65, CI 0.44–0.95) and late onset (p = 0.0028, OR 0.6, CI 0.43–0.84) vitiligo groups. The third haplotype of this block TG was associated with entire vitligo (p = 0.0402, OR 0.5, CI 0.25–0.98) as well as sporadic (p = 0.035, OR 0.42, CI 0.18–0.97) and late onset vitiligo (p = 0.0246, OR 0.38, CI 0.16–0.91). Block 2 haplotypes gave more modest associations when haplotype GA was associated in entire vitiligo (p = 0.034, OR 0.72, CI 0.53–0.98) and active vitiligo (p = 0.0214, OR 0.66, CI 0.47–0.94) groups and haplotype AT in entire vitiligo (p = 0.0275, OR 1.42, CI 1.04–1.95), active (p = 0.0272, OR 1.49, CI 1.05–2.12), sporadic (p = 0.0449, OR 1.43, CI 1.01–2.02) and late onset (p = 0.0408, OR 1.45, CI 1.02–2.06) vitiligo groups. In seven subgroups, namely stable, familial, extent ≥ 10%, Köbner, halo naevi, spontaneous repigmentation and early onset vitiligo the first two haplotype blocks were combined into a single haplotype block containing four SNPs. Haplotype CAAT of this extended block was associated in stable (p = 5.0E-4, OR 6.15, CI 1.95–19.46), familial (p = 0.0212, OR 4.54, CI 1.13–18.31), and early onset (p = 0.0029, OR 5.01, CI 1.56–16.15) subgroups. Haplotypes CGGA and CAGA were both associated in extent ≥ 10% (p = 0.034, OR 0.66, CI 0.45–0.97 and p = 0.0388, OR 1.55, CI 1.02–2.35, respectively) and halo naevi (p = 0.0191, OR 0.42, CI 0.2–0.89 and p = 0.012, OR 2.34, CI 1.18–4.62, respectively) subgroups and haplotype TGGA only in extent ≥ 10% subgroup (p = 0.0374, OR 0.3, CI 0.09–0.99). One additional version of this first haplotype block was present in female vitiligo analysis. In this instance, SNP rs10127190 was also added to the extended haplotype block and thus it contained five SNPs. Haplotypes TGGAT (p = 0.0085, OR 0.26, CI 0.09–0.76) and CAATT (p = 0.0302, OR 3.26, CI 1.06–10.01) were both associated with vitiligo in females.
Discussion
The genotyping procedure in Estonian case-control sample provided data for 29 SNPs. Four allelic and seven haplotype associations were revealed when analysing the entire vitiligo group. In addition, 31 allelic and 48 haplotype associations were present in different subgroups. In terms of genetic location, results with various strength were obtained from all studied regions, except for 1q41 that contains TLR5 gene.
The most notable SNP in 4p14 region was rs7694115. It was associated in entire vitiligo analysis as well as four subgroup analyses. The given marker is situated in the intron of TLR10 gene. Two other SNPs of TLR10 (rs10776482 and rs7660429) and one SNP located in 5′ direction from TLR10 (rs12233670) were associated in vitiligo subgroups. Of note, rs7660429 and haplotype AGG (rs10776482A, rs7694115G, and rs7660429G) produced a strong association specifically in extent < 10% group, that survived the adjustment for multiple testing. TLR10 protein is expressed by B cells, plasmacytoid dendritic cells and regulatory T cells, but the specific function remains to be clarified since the identity of its ligand has not been established (Hasan et al., 2005; Bell et al., 2007). Previous genetic studies have associated TLR10 with inflammatory diseases such as asthma (Lazarus et al., 2004), Crohn's disease (Abad et al., 2011; Morgan et al., 2012) and Ménière's disease, that is suspected to have autoimmune properties (Requena et al., 2013). Interestingly, associated SNPs of all these studies fall in the range of here described haplotype block 1 consisting of rs10776482, rs7694115, and rs7660429. It is therefore plausible, that the presently associated SNPs are in linkage disequilibrium with variants that cause changes in TLR10 function and eventually affect disease outcome. There was still another SNP rs6531668 in 4p14 region that was associated in a single subgroup. It is positioned in TLR6 gene, that has been associated with asthma (Tantisira et al., 2004) and coronary artery disease (Hamann et al., 2013).
One SNP from 4q31.3 region displayed modest associations in vitiligo subgroups. That was rs4585282 located in intron 2 of TLR2 gene. The respective protein TLR2 recognizes a wide array of antigens and has been suggested to play an instrumental role in the development of self-reactive diseases (Borrello et al., 2011). In the skin, TLR2 is expressed in antigen-presenting Langerhans cells, as well as keratinocytes and melanocytes (Yu et al., 2009; Hari et al., 2010). Polymorphisms in this gene have been associated with autoimmune diseases, namely type 1 diabetes, allergic asthma (Bjornvold et al., 2009), severe ulcerative colitis (Pierik et al., 2006) and also atopic dermatitis, that is proposed to hold autoimmune qualities (Ahmad-Nejad et al., 2004; Cipriani et al., 2014). More intriguingly, the Arg753Gln (rs5743708) polymorphism was associated with vitiligo in Turkish patients (Karaca et al., 2013). It is located 14 kb from the currently associated rs4585282 and is a missense mutation causing arginine to glutamine substitution in TLR2 peptide. The haplotype block 1 that contains rs4585282 did not reach over the position of Arg753Gln and the possible relationship between these two polymorphisms together with effects on TLR2 activity remain to be determined.
The four SNPs of 4q35.1 were in the region of TLR3 gene. SNP rs6552950 from TLR3 intron produced the only allelic association with vitiligo among males. However, several strong haplotype associations appeared in entire vitiligo and subgroup analyses, that remained statistically significant after correcting for multiple testing. The TLR3 receptor recognizes double-stranded (ds)RNA during viral infection and initiates the production of type I interferons in response. It has been shown that TLR3 is also expressed in melanocytes and stimulation with dsRNA leads to apoptosis (Yu et al., 2011). To date, polymorphisms of TLR3 have been associated with several chronic inflammatory diseases e.g., rheumatoid arthritis (Laska et al., 2014a), systemic lupus erythematosus (SLE) (Laska et al., 2014b) and osteoarthritis (Yang et al., 2013), the first two of which are autoimmune diseases. Interestingly, the associated polymorphisms in these reports (rs3775296, rs5743312, rs3775291, and rs3775290) fall in the range of a haplotype block formed by the SNPs genotyped in this study. Therefore, it is possible that these associations signal the existence of certain causal polymorphisms within the block, such as the non-synonymous rs3775291, that affect vitiligo pathology.
A single SNP rs5030728 from 9q33.1 region was associated in familial, extent ≥ 10% and female subgroups. Likewise, haplotype TA consisting of rs10759932T and rs5030728A was associated in the same groups. Notably, the results from familial and extent ≥ 10% groups remained significant when corrections for multiple testing were applied. The former SNP is located near the 5′ end of TLR4 gene and the latter is an intronic SNP of TLR4. This receptor is known for recognizing lipopolysaccharide from gram-negative bacteria, which induces the production of proinflammatory cytokines and Type I interferons (Lu et al., 2008). As a contributor to TLR2/TLR4 pathway, it has been attributed a pivotal function in the pathogenesis of autoimmune diseases (Liu et al., 2014). Similarly, to TLR2 and TLR3, it has been detected in Langerhans cells, keratinocytes and melanocytes (Hari et al., 2010). Polymorphisms of TLR4 have been associated with Crohn's disease, ulcerative colitis (Shen et al., 2010b), asthma (Zhang et al., 2011) and atherosclerosis (Kiechl et al., 2002). Remarkably, the Asp299Gly (rs4986790) polymorphism was also associated with vitiligo among Turkish patients (Karaca et al., 2013). It is positioned 1 kb from rs5030728 and while not situated within here described haplotype block, it may still be in linkage disequilibrium with these SNPs. Hence, it is possible that the current associations with non-coding SNPs reflect the effect of missense Asp299Gly on TLR2 activity. Conversely, they may have an independent impact and rs5030728 has been suggested to play a role in metabolic syndrome (Cuda et al., 2011) and inflammatory bowel disease treatment response (Bank et al., 2014). The other contributor to TA haplotype described above, rs10759932, has been associated with prostate cancer (Cheng et al., 2007), chorionic plate inflammation (Simhan et al., 2008) and atopy (Penders et al., 2010).
The final studied region was in Xp22.2 that contained TLR7 and TLR8 genes. The strongest associations were obtained with SNP rs179020 located in intron 2 of TLR7. Notably, this SNP remained significant after correcting for multiple testing in entire vitiligo and three subgroup analyses: male, sporadic, and late onset vitiligo. Furthermore, allele rs5935436C together with rs179020A formed a haplotype CA, that withstood the correction in all these groups except vitiligo males. Three other SNPs from TLR7 region—rs5935436, rs179013, and rs179008—produced weaker allelic associations in entire vitiligo and subgroup analyses. TLR7 is an intracellular receptor located in endosomes, that binds single-stranded RNA and induces the production of inflammatoty cytokines (Diebold et al., 2004). In addition to immune cells, its expression has been demonstrated in melanocytes (Kang et al., 2009). It is intriguing to note, that topical administration of TLR7 agonist imiquimod has been shown to be associated with vitiligo-like hypopigmentation (Brown et al., 2005). Considering that TLR7 activation may lead to apoptosis (Meyer et al., 2003), this effect could be due to TLR7 induced loss of melanocytes. Concurrently, the role of TLR7 in other autoimmune diseases has been widely reported, especially in the case of SLE (Deane et al., 2007; O'Neill et al., 2009). The proposed mechanism involves activation of autoreactive B cells through TLR7 signaling, while TLR9 acts as a restrictor of this process and the role of TLR8 is yet to be clarified (Soni et al., 2014). On a genetic level, TLR7 SNPs have been associated with Graves' disease (Xiao et al., 2015) and SLE (Shen et al., 2010a; Enevold et al., 2014). Among the four SNPs associated in this study, rs179008 has recently been associated with asthma (Møller-Larsen et al., 2008), multiple sclerosis (Enevold et al., 2010), and SLE (dos Santos et al., 2012). It is a non-synonymous mutation, where A to T substitution causes glycine to leucine change in TLR7 peptide (Gln11Leu). Currently, T was the associated allele, whereas previously both A and T were shown to confer disease susceptibility (Møller-Larsen et al., 2008; dos Santos et al., 2012). In silico analyses by Møller-Larsen and colleagues suggested, that changing Gln11 to Leu shortens the N-region and extends the H-region of TLR7 signal peptide, thus potentially affecting post-translational processing (Møller-Larsen et al., 2008). Taken together, these genetic associations of four TLR7 SNPs could indicate alterations in TLR7 functioning, but the precise mechanisms remain to be determined in future studies.
In conclusion, the present study revealed a number of genetic associations between TLR gene SNPs and vitiligo susceptibility. The main findings emerged with TLR7 SNP 179020 and TLR3 haplotype AACT. SNPs of TLR10 and TLR4 were associated in specific subgroups, indicating their possible role in certain subphenotypes. However, the interpretation of these results should account for the small sample used, while the functional effects of studied SNPs remain to be elucidated in future experiments.
Conflict of Interest Statement
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Acknowledgments
This study was financially supported by Estonian Research Council grant PUT177, Estonian Ministry of Education and Research targeted financing (SF0180043s07), and European Regional Development Fund. We would like to thank the patients and volunteers for participation.
References
Abad, C., González-Escribano, M. F., Diaz-Gallo, L. M., Lucena-Soto, J. M., Márquez, J. L., Leo, E., et al. (2011). Association of Toll-like receptor 10 and susceptibility to Crohn's disease independent of NOD2. Genes Immun. 12, 635–642. doi: 10.1038/gene.2011.41
Ahmad-Nejad, P., Mrabet-Dahbi, S., Breuer, K., Klotz, M., Werfel, T., Herz, U., et al. (2004). The toll-like receptor 2 R753Q polymorphism defines a subgroup of patients with atopic dermatitis having severe phenotype. J. Allergy Clin. Immunol. 113, 565–567. doi: 10.1016/j.jaci.2003.12.583
Akira, S., and Takeda, K. (2004). Toll-like receptor signalling. Nat. Rev. Immunol. 4, 499–511. doi: 10.1038/nri1391
Alkhateeb, A., Fain, P. R., Thody, A., Bennett, D. C., and Spritz, R. A. (2003). Epidemiology of vitiligo and associated autoimmune diseases in Caucasian probands and their families. Pigment Cell Res. 16, 208–214. doi: 10.1034/j.1600-0749.2003.00032.x
Bank, S., Andersen, P. S., Burisch, J., Pedersen, N., Roug, S., Galsgaard, J., et al. (2014). Associations between functional polymorphisms in the NFkappaB signaling pathway and response to anti-TNF treatment in Danish patients with inflammatory bowel disease. Pharmacogenomics J. 14, 526–534. doi: 10.1038/tpj.2014.19
Barrett, J. C., Fry, B., Maller, J., and Daly, M. J. (2005). Haploview: analysis and visualization of LD and haplotype maps. Bioinformatics 21, 263–265. doi: 10.1093/bioinformatics/bth457
Bell, M. P., Svingen, P. A., Rahman, M. K., Xiong, Y., and Faubion, W. A. Jr. (2007). FOXP3 regulates TLR10 expression in human T regulatory cells. J. Immunol. 179, 1893–1900. doi: 10.4049/jimmunol.179.3.1893
Bjørnvold, M., Munthe-Kaas, M. C., Egeland, T., Joner, G., Dahl-Jørgensen, K., Njølstad, P. R., et al. (2009). A TLR2 polymorphism is associated with type 1 diabetes and allergic asthma. Genes Immun. 10, 181–187. doi: 10.1038/gene.2008.100
Boissy, R. E., and Spritz, R. A. (2009). Frontiers and controversies in the pathobiology of vitiligo: separating the wheat from the chaff. Exp. Dermatol. 18, 583–585. doi: 10.1111/j.1600-0625.2008.00826.x
Borrello, S., Nicolo, C., Delogu, G., Pandolfi, F., and Ria, F. (2011). TLR2: a crossroads between infections and autoimmunity? Int. J. Immunopathol. Pharmacol. 24, 549–556.
Brown, T., Zirvi, M., Cotsarelis, G., and Gelfand, J. M. (2005). Vitiligo-like hypopigmentation associated with imiquimod treatment of genital warts. J. Am. Acad. Dermatol. 52, 715–716. doi: 10.1016/j.jaad.2004.10.861
Cheng, I., Plummer, S. J., Casey, G., and Witte, J. S. (2007). Toll-like receptor 4 genetic variation and advanced prostate cancer risk. Cancer Epidemiol. Biomarkers Prev. 16, 352–355. doi: 10.1158/1055-9965.EPI-06-0429
Cipriani, F., Ricci, G., Leoni, M. C., Capra, L., Baviera, G., Longo, G., et al. (2014). Autoimmunity in atopic dermatitis: biomarker or simply epiphenomenon? J. Dermatol. 41, 569–576. doi: 10.1111/1346-8138.12464
Cuda, C., Badawi, A., Karmali, M., and El-Sohemy, A. (2011). Polymorphisms in Toll-like receptor 4 are associated with factors of the metabolic syndrome and modify the association between dietary saturated fat and fasting high-density lipoprotein cholesterol. Metabolism 60, 1131–1135. doi: 10.1016/j.metabol.2010.12.006
Deane, J. A., Pisitkun, P., Barrett, R. S., Feigenbaum, L., Town, T., Ward, J. M., et al. (2007). Control of toll-like receptor 7 expression is essential to restrict autoimmunity and dendritic cell proliferation. Immunity 27, 801–810. doi: 10.1016/j.immuni.2007.09.009
Diebold, S. S., Kaisho, T., Hemmi, H., Akira, S., and Reis e Sousa, C. (2004). Innate antiviral responses by means of TLR7-mediated recognition of single-stranded RNA. Science 303, 1529–1531. doi: 10.1126/science.1093616
dos Santos, B. P., Valverde, J. V., Rohr, P., Monticielo, O. A., Brenol, J. C., Xavier, R. M., et al. (2012). TLR7/8/9 polymorphisms and their associations in systemic lupus erythematosus patients from southern Brazil. Lupus 21, 302–309. doi: 10.1177/0961203311425522
Enevold, C., Nielsen, C. H., Jacobsen, R. S., Hermansen, M. L., Molbo, D., Avlund, K., et al. (2014). Single nucleotide polymorphisms in genes encoding toll-like receptors 7, 8 and 9 in Danish patients with systemic lupus erythematosus. Mol. Biol. Rep. 41, 5755–5763. doi: 10.1007/s11033-014-3447-4
Enevold, C., Oturai, A. B., Sørensen, P. S., Ryder, L. P., Koch-Henriksen, N., Bendtzen, K., et al. (2010). Polymorphisms of innate pattern recognition receptors, response to interferon-beta and development of neutralizing antibodies in multiple sclerosis patients. Mult. Scler. 16, 942–949. doi: 10.1177/1352458510373264
Hamann, L., Koch, A., Sur, S., Hoefer, N., Glaeser, C., Schulz, S., et al. (2013). Association of a common TLR-6 polymorphism with coronary artery disease - implications for healthy ageing? Immun. Ageing 10:43. doi: 10.1186/1742-4933-10-43
Hari, A., Flach, T. L., Shi, Y., and Mydlarski, P. R. (2010). Toll-like receptors: role in dermatological disease. Mediators Inflamm. 2010:437246. doi: 10.1155/2010/437246
Hasan, U., Chaffois, C., Gaillard, C., Saulnier, V., Merck, E., Tancredi, S., et al. (2005). Human TLR10 is a functional receptor, expressed by B cells and plasmacytoid dendritic cells, which activates gene transcription through MyD88. J. Immunol. 174, 2942–2950. doi: 10.4049/jimmunol.174.5.2942
Kang, H. Y., Park, T. J., and Jin, S. H. (2009). Imiquimod, a Toll-like receptor 7 agonist, inhibits melanogenesis and proliferation of human melanocytes. J. Invest. Dermatol. 129, 243–246. doi: 10.1038/jid.2008.184
Karaca, N., Ozturk, G., Gerceker, B. T., Turkmen, M., and Berdeli, A. (2013). TLR2 and TLR4 gene polymorphisms in Turkish vitiligo patients. J. Eur. Acad. Dermatol. Venereol. 27:e85–e90. doi: 10.1111/j.1468-3083.2012.04514.x
Kiechl, S., Lorenz, E., Reindl, M., Wiedermann, C. J., Oberhollenzer, F., Bonora, E., et al. (2002). Toll-like receptor 4 polymorphisms and atherogenesis. N. Engl. J. Med. 347, 185–192. doi: 10.1056/NEJMoa012673
Laska, M. J., Hansen, B., Troldborg, A., Lorenzen, T., Stengaard-Pedersen, K., Junker, P., et al. (2014a). A non-synonymous single-nucleotide polymorphism in the gene encoding Toll-like Receptor 3 (TLR3) is associated with sero-negative Rheumatoid Arthritis (RA) in a Danish population. BMC Res. Notes 7:716. doi: 10.1186/1756-0500-7-716
Laska, M. J., Troldborg, A., Hansen, B., Stengaard-Pedersen, K., Junker, P., Nexø, B. A., et al. (2014b). Polymorphisms within Toll-like receptors are associated with systemic lupus erythematosus in a cohort of Danish females. Rheumatology (Oxford). 53, 48–55. doi: 10.1093/rheumatology/ket316
Lazarus, R., Raby, B. A., Lange, C., Silverman, E. K., Kwiatkowski, D. J., Vercelli, D., et al. (2004). TOLL-like receptor 10 genetic variation is associated with asthma in two independent samples. Am. J. Respir. Crit. Care Med. 170, 594–600. doi: 10.1164/rccm.200404-491OC
Le Poole, I. C., and Luiten, R. M. (2008). Autoimmune etiology of generalized vitiligo. Curr. Dir. Autoimmun. 10, 227–243. doi: 10.1159/000131485
Liu, Y., Yin, H., Zhao, M., and Lu, Q. (2014). TLR2 and TLR4 in autoimmune diseases: a comprehensive review. Clin. Rev. Allergy Immunol. 47, 136–147. doi: 10.1007/s12016-013-8402-y
Lu, Y. C., Yeh, W. C., and Ohashi, P. S. (2008). LPS/TLR4 signal transduction pathway. Cytokine 42, 145–151. doi: 10.1016/j.cyto.2008.01.006
Meyer, T., Nindl, I., Schmook, T., Ulrich, C., Sterry, W., and Stockfleth, E. (2003). Induction of apoptosis by Toll-like receptor-7 agonist in tissue cultures. Br. J. Dermatol. 149(Suppl. 66), 9–14. doi: 10.1046/j.0366-077X.2003.05632.x
Miller, L. S. (2008). Toll-like receptors in skin. Adv. Dermatol. 24, 71–87. doi: 10.1016/j.yadr.2008.09.004
Møller-Larsen, S., Nyegaard, M., Haagerup, A., Vestbo, J., Kruse, T. A., Borglum, A. D., et al. (2008). Association analysis identifies TLR7 and TLR8 as novel risk genes in asthma and related disorders. Thorax 63, 1064–1069. doi: 10.1136/thx.2007.094128
Morgan, A. R., Lam, W. J., Han, D. Y., Fraser, A. G., and Ferguson, L. R. (2012). Genetic variation within TLR10 is associated with Crohn's disease in a New Zealand population. Hum. Immunol. 73, 416–420. doi: 10.1016/j.humimm.2012.01.015
Oh, D. Y., Schumann, R. R., Hamann, L., Neumann, K., Worm, M., and Heine, G. (2009). Association of the toll-like receptor 2 A-16934T promoter polymorphism with severe atopic dermatitis. Allergy 64, 1608–1615. doi: 10.1111/j.1398-9995.2009.02066.x
O'Neill, L. A., Bryant, C. E., and Doyle, S. L. (2009). Therapeutic targeting of Toll-like receptors for infectious and inflammatory diseases and cancer. Pharmacol. Rev. 61, 177–197. doi: 10.1124/pr.109.001073
Penders, J., Thijs, C., Mommers, M., Stobberingh, E. E., Dompeling, E., Reijmerink, N. E., et al. (2010). Host-microbial interactions in childhood atopy: toll-like receptor 4 (TLR4), CD14, and fecal Escherichia coli. J. Allergy Clin. Immunol. 125, 231-6 e1–5. doi: 10.1016/j.jaci.2009.10.011
Pierik, M., Joossens, S., Van Steen, K., Van Schuerbeek, N., Vlietinck, R., Rutgeerts, P., et al. (2006). Toll-like receptor-1, -2, and -6 polymorphisms influence disease extension in inflammatory bowel diseases. Inflamm. Bowel Dis. 12, 1–8. doi: 10.1097/01.MIB.0000195389.11645.ab
Pradhan, V. D., Das, S., Surve, P., and Ghosh, K. (2012). Toll-like receptors in autoimmunity with special reference to systemic lupus erythematosus. Indian J. Hum. Genet. 18, 155–160. doi: 10.4103/0971-6866.100750
Requena, T., Gazquez, I., Moreno, A., Batuecas, A., Aran, I., Soto-Varela, A., et al. (2013). Allelic variants in TLR10 gene may influence bilateral affectation and clinical course of Meniere's disease. Immunogenetics 65, 345–355. doi: 10.1007/s00251-013-0683-z
Romaní, J., Juliá, M., Lozano, F., Muñoz-Santos, C., Guilabert, A., Carrascosa, J. M., et al. (2015). Toll-like receptor 9 promoter polymorphism as a predictive factor of narrow-band UVB phototherapy response in patients with psoriasis. Photodermatol. Photoimmunol. Photomed. 31, 98–103. doi: 10.1111/phpp.12160
Shen, N., Fu, Q., Deng, Y., Qian, X., Zhao, J., Kaufman, K. M., et al. (2010a). Sex-specific association of X-linked Toll-like receptor 7 (TLR7) with male systemic lupus erythematosus. Proc. Natl. Acad. Sci. U.S.A. 107, 15838–15843. doi: 10.1073/pnas.1001337107
Shen, X., Shi, R., Zhang, H., Li, K., Zhao, Y., and Zhang, R. (2010b). The Toll-like receptor 4 D299G and T399I polymorphisms are associated with Crohn's disease and ulcerative colitis: a meta-analysis. Digestion 81, 69–77. doi: 10.1159/000260417
Simhan, H. N., MacPherson, T., Caritis, S. N., and Krohn, M. A. (2008). Maternal and fetal Toll-like receptor 4 genotype and chorionic plate inflammatory lesions. Am. J. Obstet. Gynecol. 199, 400 e1–e4. doi: 10.1016/j.ajog.2008.07.043
Soni, C., Wong, E. B., Domeier, P. P., Khan, T. N., Satoh, T., Akira, S., et al. (2014). B cell-intrinsic TLR7 signaling is essential for the development of spontaneous germinal centers. J. Immunol. 193, 4400–4414. doi: 10.4049/jimmunol.1401720
Spritz, R. A. (2011). Recent progress in the genetics of generalized vitiligo. J. Genet. Genomics. 38, 271–278. doi: 10.1016/j.jgg.2011.05.005
Takeda, K., Kaisho, T., and Akira, S. (2003). Toll-like receptors. Annu. Rev. Immunol. 21, 335–376. doi: 10.1146/annurev.immunol.21.120601.141126
Tantisira, K., Klimecki, W. T., Lazarus, R., Palmer, L. J., Raby, B. A., Kwiatkowski, D. J., et al. (2004). Toll-like receptor 6 gene (TLR6): single-nucleotide polymorphism frequencies and preliminary association with the diagnosis of asthma. Genes Immun. 5, 343–346. doi: 10.1038/sj.gene.6364096
Tobler, A. R., Short, S., Andersen, M. R., Paner, T. M., Briggs, J. C., Lambert, S. M., et al. (2005). The SNPlex genotyping system: a flexible and scalable platform for SNP genotyping. J. Biomol. Tech. 16, 398–406.
Xiao, W., Liu, Z., Lin, J., Li, J., Wu, K., Ma, Y., et al. (2015). Association of Toll-like receptor 7 and 8 gene polymorphisms with Graves' disease in Chinese Cantonese population. Tissue Antigens 85, 29–34. doi: 10.1111/tan.12479
Yang, H. Y., Lee, H. S., Lee, C. H., Fang, W. H., Chen, H. C., Salter, D. M., et al. (2013). Association of a functional polymorphism in the promoter region of TLR-3 with osteoarthritis: a two-stage case-control study. J. Orthop. Res. 31, 680–685. doi: 10.1002/jor.22291
Yu, N., Zhang, S., Sun, T., Kang, K., Guan, M., and Xiang, L. (2011). Double-stranded RNA induces melanocyte death via activation of Toll-like receptor 3. Exp. Dermatol. 20, 134–139. doi: 10.1111/j.1600-0625.2010.01208.x
Yu, N., Zhang, S., Zuo, F., Kang, K., Guan, M., and Xiang, L. (2009). Cultured human melanocytes express functional toll-like receptors 2-4, 7 and 9. J. Dermatol. Sci. 56, 113–120. doi: 10.1016/j.jdermsci.2009.08.003
Keywords: vitiligo, autoimmune, Toll-like receptor, SNP, genetic association study
Citation: Traks T, Keermann M, Karelson M, Rätsep R, Reimann E, Silm H, Vasar E, Kõks S and Kingo K (2015) Polymorphisms in Toll-like receptor genes are associated with vitiligo. Front. Genet. 6:278. doi: 10.3389/fgene.2015.00278
Received: 14 May 2015; Accepted: 19 August 2015;
Published: 09 September 2015.
Edited by:
Mitesh Dwivedi, Uka Tarsadia University, IndiaReviewed by:
Yan Wu, No. 1 Hospital of China Medical University, ChinaRanya Moustafa Hassan, Suez Canal University, Egypt
Copyright © 2015 Traks, Keermann, Karelson, Rätsep, Reimann, Silm, Vasar, Kõks and Kingo. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) or licensor are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Tanel Traks, Department of Dermatology, University of Tartu, Raja 31, 50417 Tartu, Estonia,dGFuZWwudHJha3NAdXQuZWU=