 Edward A. Ruiz-Narváez1*†
Edward A. Ruiz-Narváez1*† Lara Sucheston-Campbell2,3†
Lara Sucheston-Campbell2,3† Jeannette T. Bensen4
Jeannette T. Bensen4 Song Yao5
Song Yao5 Stephen Haddad1
Stephen Haddad1 Christopher A. Haiman6Elisa V. Bandera7Esther M. John8
Christopher A. Haiman6Elisa V. Bandera7Esther M. John8 Leslie Bernstein9
Leslie Bernstein9 Jennifer J. Hu10Regina G. Ziegler11Sandra L. Deming12Andrew F. Olshan4Christine B. Ambrosone5Julie R. Palmer1
Jennifer J. Hu10Regina G. Ziegler11Sandra L. Deming12Andrew F. Olshan4Christine B. Ambrosone5Julie R. Palmer1 Kathryn L. Lunetta13
Kathryn L. Lunetta13- 1Slone Epidemiology Center, Boston University, Boston, MA, USA
- 2College of Pharmacy, The Ohio State University, Columbus, OH, USA
- 3College of Veterinary Medicine, The Ohio State University, Columbus, OH, USA
- 4Department of Epidemiology, Gillings School of Global Public Health, University of North Carolina at Chapel Hill, Chapel Hill, NC, USA
- 5Department of Cancer Prevention and Control, Roswell Park Cancer Institute, Buffalo, NY, USA
- 6Department of Preventive Medicine, Keck School of Medicine, University of Southern California/Norris Comprehensive Cancer Center, Los Angeles, CA, USA
- 7Rutgers Cancer Institute of New Jersey, New Brunswick, NJ, USA
- 8Cancer Prevention Institute of California, Fremont, CA, USA
- 9Division of Cancer Etiology, Department of Population Science, Beckman Research Institute, City of Hope, Duarte, CA, USA
- 10Sylvester Comprehensive Cancer Center and Department of Public Health Sciences, University of Miami Miller School of Medicine, Miami, FL, USA
- 11 Epidemiology and Biostatistics Program, Division of Cancer Epidemiology and Genetics, National Cancer Institute, Bethesda, MD, USA
- 12Vanderbilt Epidemiology Center, Vanderbilt University and the Vanderbilt-Ingram Cancer Center, Nashville, TN, USA
- 13Department of Biostatistics, Boston University School of Public Health, Boston, MA, USA
Recent genetic admixture coupled with striking differences in incidence of estrogen receptor (ER) breast cancer subtypes, as well as severity, between women of African and European ancestry, provides an excellent rationale for performing admixture mapping in African American women with breast cancer risk. We performed the largest breast cancer admixture mapping study with in African American women to identify novel genomic regions associated with the disease. We conducted a genome-wide admixture scan using 2,624 autosomal ancestry informative markers (AIMs) in 3,629 breast cancer cases (including 1,968 ER-positive, 1093 ER-negative, and 601 triple-negative) and 4,658 controls from the African American Breast Cancer Epidemiology and Risk (AMBER) Consortium, a collaborative study of four large geographically different epidemiological studies of breast cancer in African American women. We used an independent case-control study to test for SNP association in regions with genome-wide significant admixture signals. We found two novel genome-wide significant regions of excess African ancestry, 4p16.1 and 17q25.1, associated with ER-positive breast cancer. Two regions known to harbor breast cancer variants, 10q26 and 11q13, were also identified with excess of African ancestry. Fine-mapping of the identified genome-wide significant regions suggests the presence of significant genetic associations with ER-positive breast cancer in 4p16.1 and 11q13. In summary, we identified three novel genomic regions associated with breast cancer risk by ER status, suggesting that additional previously unidentified variants may contribute to the racial differences in breast cancer risk in the African American population.
Introduction
A majority of the close to 100 breast cancer risk loci that have been identified to date were found using genome-wide approaches in populations of European ancestry women (Maxwell and Nathanson, 2013; Michailidou et al., 2015; Couch et al., 2016). Follow-up studies in non-European populations are necessary to understand the population-specific risk associated with these loci. Further, genome-wide associations studies in non-European populations are required to identify race-specific genetic variation associated with breast cancer subtypes (Feng et al., 2014). Sample size remains one of the main limiting factors in performing genome-wide association studies (GWAS) in non-Europeans. Admixture mapping, a method for discovery of genomic regions that is based on linkage disequilibrium in populations with recent population admixture, requires far fewer samples and markers than GWAS to detect loci associated with phenotypic variation (Rosenberg et al., 2003; Smith and O’Brien, 2005; Winkler et al., 2010). Early admixture mapping efforts in African American women with breast cancer were likely under-powered, in particular to detect loci associated with breast cancer subtypes (Fejerman et al., 2009). However, this approach, in conjunction with fine mapping, has been successful at identifying genetic associations with other complex traits in recently admixed populations (Zhu et al., 2005; Freedman et al., 2006; Nalls et al., 2008; Manuck et al., 2011; Bensen et al., 2014; Jeff et al., 2014; Parker et al., 2014).
African American women have higher breast cancer mortality rates than US women of European ancestry, particularly among younger women, attributable to diagnosis at a later stage often with larger tumors and lymph node metastases (Chlebowski et al., 2005; Hershman et al., 2005; Grann et al., 2006; Fejerman and Ziv, 2008). In addition, women of African ancestry are at greater risk of having estrogen receptor negative (ER-) and triple negative (ER-, progesterone receptor negative, PR-; and human epidermal growth factor receptor 2 negative, HER2-) breast cancer at all ages compared with European American women (DeSantis et al., 2014). ER- and triple negative cancers are more aggressive, and have fewer treatment options than ER+ disease (Clarke et al., 2012). These differences in incidence, severity and mortality between women of African and European ancestry suggest that there may be different underlying genetic contributions to breast cancer risk in African American and European American women. Recent genetic admixture coupled with these striking racial differences provides excellent rationale for performing admixture mapping in African American women with breast cancer.
To this end, we performed a genome-wide admixture mapping in germline DNA in the African American Breast Cancer Epidemiology and Risk (AMBER) Consortium, a collaborative project from four of the largest studies of breast cancer in African American women (Palmer et al., 2014a). This represents the largest breast cancer admixture mapping study in African American women to date. We discuss our findings in the context of those from the previous admixture mapping study of breast cancer (Fejerman et al., 2009) and of known breast cancer risk loci identified among European Americans recently confirmed in African Americans.
Materials and Methods
Ethics Approval and Consent to Participate
The CBCS was approved by the Institutional Review Board at the University of North Carolina at Chapel Hill School of Medicine. The WCHS was approved by the Institutional Review Boards at the University of Medicine and Dentistry of New Jersey (presently Rutgers University), Mount Sinai School of Medicine, and Roswell Park Cancer Institute. The BWHS was approved by the Institutional Review Board at the Boston University School of Medicine. The MEC was approved by the Institutional Review Boards of the University of Hawaii and University of Southern California. Written informed consent was obtained from each participant.
Study Population
Two case-control studies, the Carolina Breast Cancer Study (CBCS) and the Women’s Circle of Health Study (WCHS), and two prospective cohort studies, the Black Women’s Health Study (BWHS) and the Multi-Ethnic Cohort (MEC) comprise the AMBER Consortium (Palmer et al., 2014a,b; Bandera et al., 2015); the four studies are described below.
The CBCS is a population-based case control study of women ages 20–74, conducted in 44 counties in North Carolina that began in 1993 (Newman et al., 1995). Breast cancer cases were identified using Rapid Case Ascertainment in cooperation with the NC Central Cancer Registry. Controls were identified using Division of Motor Vehicles lists (for women under age 65) and Health Care Financing Administration lists (for women 65 and older). The age range of study participants was 20–74 years. CBCS contributed 1,408 African–American cases and 615 controls to this analysis.
The WCHS includes both European and African American women, 20–75 years of age, newly diagnosed with a first primary, and histologically confirmed breast cancer. Cases were identified from 10 counties in New Jersey through the New Jersey State Cancer Registry and through major metropolitan hospitals in New York City serving a large minority population. Controls, frequency matched to cases by 5-years age categories and self-reported race, were identified using random digit dialing in New York City and New Jersey, as well as from the community (Ambrosone et al., 2009; Bandera et al., 2013). Recruitment in New York City took place between 2003 and 2008. Recruitment at New Jersey started in 2006 and is ongoing. WCHS contributed 821 African American breast cancer cases and 834 African American controls to this analysis.
The BWHS is an ongoing prospective cohort study of 59,000 US women aged 21–69 years from across the United States who self-identify as “black” (Rosenberg et al., 1995). Beginning in 1995, women completed a 14-page postal health questionnaire. Biennial postal questionnaires are mailed to participants to update information. During 2004–2007, we obtained saliva samples as a source of DNA from 26,814 participants using the mouthwash-swish method (Cozier et al., 2004). BWHS contributed 901 cases and 2,249 controls to this analysis.
The MEC study is a prospective cohort that includes over 215,000 individuals from Hawaii and California (primarily Los Angeles) assembled between 1993 and 1996 (Kolonel et al., 2000; Pike et al., 2002). African Americans, Native Hawaiians, Japanese, Latinos and European Americans comprise the predominant population. Blood samples were collected from incident breast cancer cases starting in 1994 and identified by cohort linkage to Surveillance, Epidemiology and End Results (SEER) registries, as well as a random sample of MEC participants to serve as controls for genetic analyses. MEC contributed genetic data from 499 African American cases and 960 African American controls to this analysis.
Genotyping and Quality Control
Genotyping of constitutional DNA of CBCS, WCHS, and BWHS samples was performed by the Center for Inherited Disease Research (CIDR) using the Illumina Human Exome Beadchip v1.1 array with custom content. The standard content of this array includes more than 240,000 coding variants, as well as tag SNPs for GWAS hits, a grid of common variants, and ancestry informative markers (AIMs). A more complete description of the exome chip design and standard content is available from http://genome.sph.umich.edu/wiki/Exome_Chip_Design.
Genotype quality control for this study has been described previously (Laurie et al., 2010). Genotype data were available for 2,624 autosomal AIMs (Supplementary Table 1) included as standard content in the exome array and typed in AMBER participants, including 3,130 cases, 1,674 ER+ cases and 963 ER- cases (515 triple negative) and 3,698 controls. In addition, genetic data from 499 cases (130 ER- including 86 triple negative, 294 ER+, 75 ER unknown) and 960 controls in the MEC study were available from previous genotyping on an earlier version of the exome array that contained the same AIMs. Final sample size for the current admixture analyses was 8,287 participants: 3,629 cases (1,093 ER- including 601 triple negative, 1,968 ER+, and 568 ER status unknown) and 4,658 controls.
We assessed population stratification by calculating principal components of genetic variation using the smartpca program in the EIGENSOFT package (Patterson et al., 2006). Supplementary Figure 1 shows a plot of the top two principal components of the participants stratified by study. No obvious differences in distribution of these principal components were observed across studies.
Admixture Mapping
We used ADMIXMAP software version 3.8.3103 (Hoggart et al., 2004; Montana and Pritchard, 2004; Reich and Patterson, 2005) to estimate African locus-specific ancestry and determine excess sharing across the genome. ADMIXMAP uses a Markov chain Monte Carlo algorithm to generate a score test at each position in the genome that measures local ancestry associations with a phenotype (Hoggart et al., 2004). Admixture mapping is generally carried out using case-only and/or case-control analyses. The case-only approach compares locus-specific ancestry in each chromosomal position with average genome-wide ancestry. The case-control method tests for differences of locus-specific ancestry between cases and controls adjusting for individual admixture and other covariates. Case-only analysis has greater statistical power than case-control assuming that deviation in ancestry between cases and controls is not due to population stratification (Shriner, 2013). We therefore first performed a case-only admixture scan and then a case-control admixture analysis to confirm local ancestry associations from the case-only results. Case-control analyses were adjusted for global individual admixture, study, age (10-year groups), geographic region, and source of constitutional DNA (Oragene-saliva, blood, mouthwash-saliva). Statistical significance was assessed using Z statistics, with a threshold of |Z| > 4.0 considered genome-wide statistically significant. A negative Z-score indicates that higher African (lower European) ancestry at that particular locus is associated with higher breast cancer risk while a positive Z-score indicates that lower African (higher European) ancestry is associated with higher breast cancer risk. We performed analyses of breast cancer overall, and by ER and triple negative status. For each analysis, we ran ADMIXMAP using 2,000 burn-in iterations and 8,000 follow-on iterations. The percentage of African ancestry for each individual was determined using ADMIXMAP and compared across controls, all cases and by ER status as well as by site, using logistic regression adjusting for the covariates described above.
Association Analyses
In order to identify SNPs that may help explain any observed association between local African ancestry and risk of breast cancer regions meeting the genome-wide significant criterion (|Z| > 4) in AMBER, fine-mapping analyses were performed using typed and imputed data from the African American Breast Cancer (AABC) consortium, which included participants from six epidemiological studies (Chen et al., 2011, 2013) after excluding MEC, WCHS, and CBCS subjects which were included in AMBER. Regional association analyses were conducted in 2,114 cases (1,116 ER+, 591 ER-, and 407 unknown ER status) and 1,911 controls typed on the Illumina Human1M-Duo BeadChip [34] and imputed using MACH with HAPMAP2 as a reference. Within each genome-wide significant admixture peak a Bonferroni correction for the number of directly genotyped markers tested was used to determine statistical significance (Palmer et al., 2016).
Results
Table 1 shows the participant characteristics by study site in cases and controls. A total of 3,629 breast cancer cases (1,968 ER+ cases, 1,093 ER- cases including 601 triple negative, and 568 unknown ER status) and 4,658 controls were included in the present analysis. CBCS and WCHS cases had the highest proportion of African ancestry (84.0%) and MEC the lowest (78.0%). Mean African ancestry, adjusted for age, geographic region, DNA source and study site was 81.3% for controls and 81.2% for cases (P = 0.74 for difference of means). A total of 2,624 autosomal AIMs were analyzed (Supplementary Table 1).
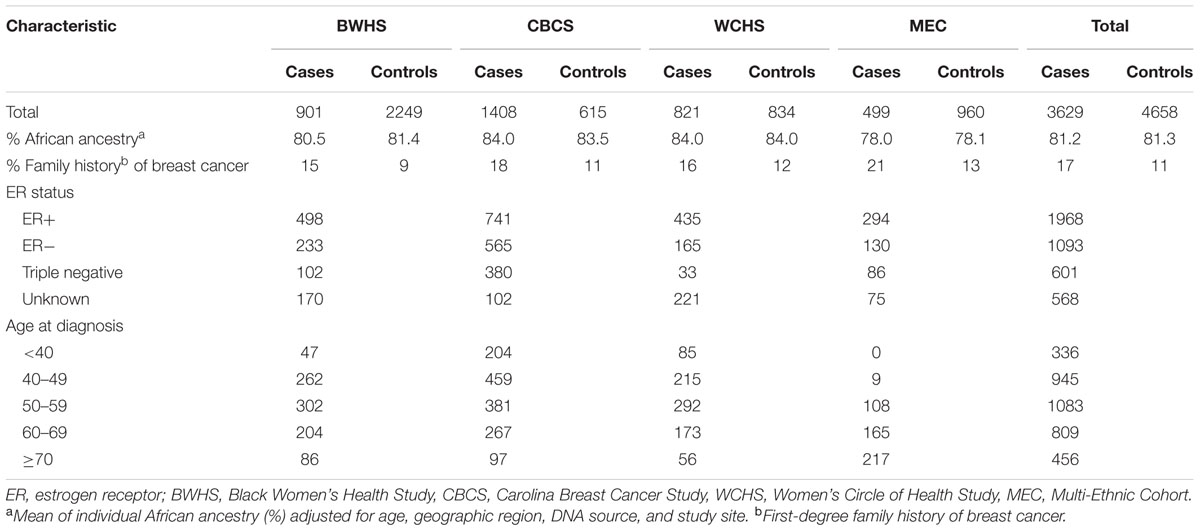
TABLE 1. Characteristics of participants by study in the AMBER consortium.
Admixture Mapping
Table 2 shows the four genome-wide significant regions in case-only analysis (|Z| ≥ 4, P < 6 × 10-5) that also had confirmatory results in case-control analysis (|Z|≥ 2.0, P < 0.05). We identified two novel genome-wide significant regions, showing evidence of excess African ancestry associated with ER+ breast cancer on 4p16.3 and 17q25.1. The 10q26 and 11q13.2 regions both contain SNPs previously identified in published GWAS of breast cancer in European ancestry women (Hunter et al., 2007; Turnbull et al., 2010), and fine-mapping efforts have identified independent signals in African American women (Chen et al., 2011).
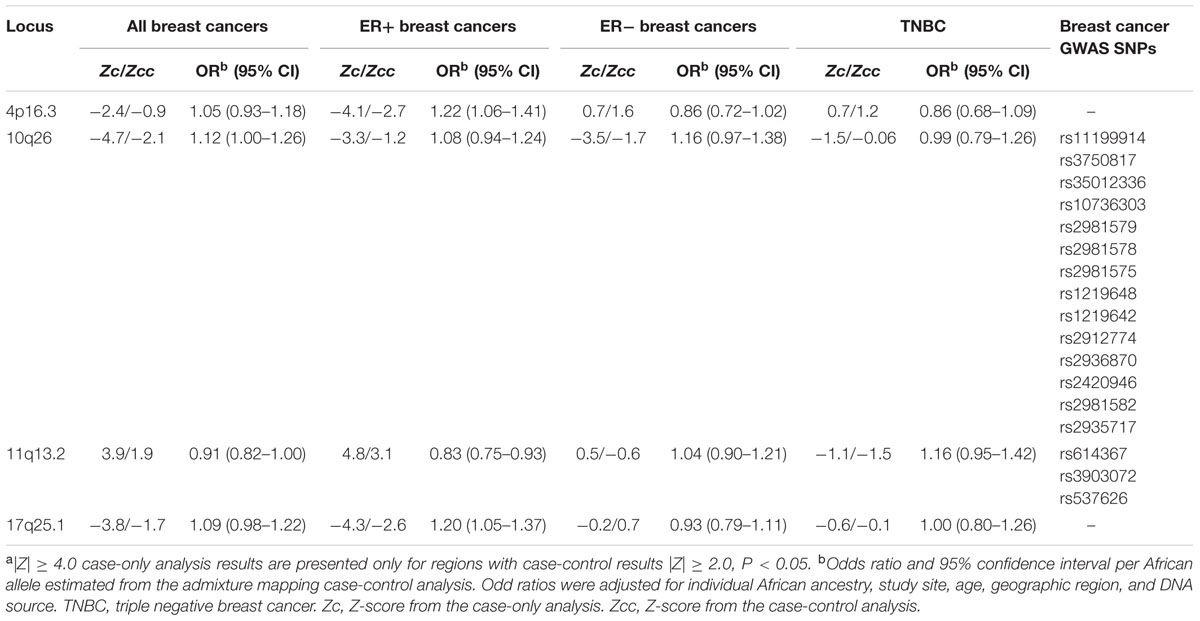
TABLE 2. Genome wide significant regions of excess African ancestry (negative Z-score) and reduced (positive Z-score) associated with breast cancer at |Z| > 4 in case-only analyses and corresponding case-control resultsa.
Supplementary Table 2 contains admixture results for all 2,624 AIMs and breast cancer subtypes. Supplementary Table 3 shows admixture results with suggestive evidence of association, defined as 4 > |Z|≥ 3.5 in case-only and |Z|≥ 2.0 in case-control analyses. Of these 13 suggestive regions, seven contain at least one genome-wide significant marker previously associated with breast cancer (Hindorff et al., 2015).
Association Analyses
We used an independent study (African American Breast Cancer Consortium, AABC) to test for association of SNPs in the admixture associated regions identified for ER+ and overall breast cancer. We defined significant association at P < 5.8 × 10-6 after Bonferroni correction for multiple testing based on adjustment for the 8,653 typed markers included. AABC does not have information about triple negative breast cancer, thus we did not follow up the three regions showing suggestive association with this subtype. We report on the most significant associations in each genomic region identified in the admixture mapping (Table 3; Supplementary Figure 2).
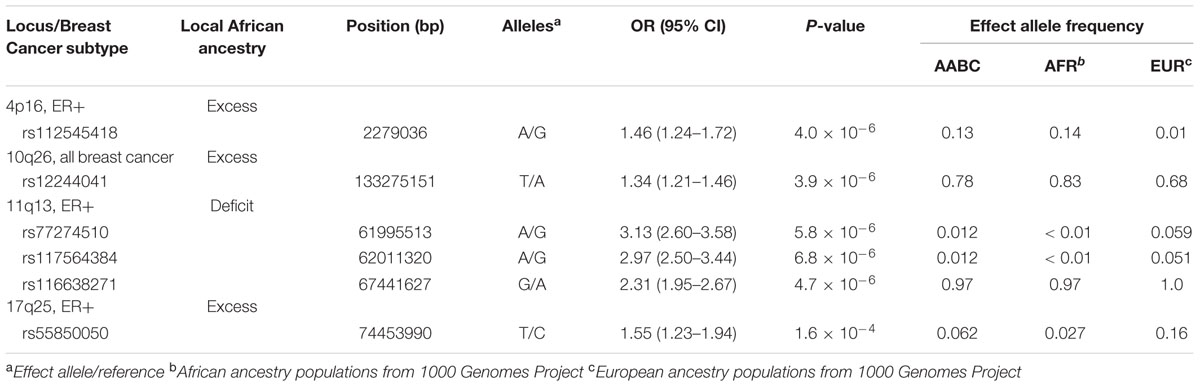
TABLE 3. Most significant single independent SNP associations in the African American Breast Cancer (AABC) consortium within genomic regions identified in admixture mapping of AMBER.
4p16 and ER-Positive (ER+) Breast Cancer
One SNP in the 4p16 region exceeded multiple test correction threshold, rs112545418 in ZFYVE28; the A allele showed a 46% risk increase compared to the G allele (P = 4.0 × 10-6). Consistent with our admixture mapping results (i.e., excess of local African ancestry at 4p16 associated with higher risk of ER+ breast cancer), the risk A-allele is more frequent in 1000 Genomes African ancestry samples (AFR), 14%, and AABC, 13%, than in European ancestry (EUR) samples, 1% (Table 3).
10q26 and Overall Breast Cancer
The most significant association with overall breast cancer, rs12244041, at P = 2.9 × 10-6, is located over 10 Mb from the well-known breast cancer associated FGFR2 gene and is unlinked to SNPs in this region (r2 = 0.01). The risk T-allele at this SNP is associated with 87% higher risk of breast cancer, and it is more frequent in AABC (79%) and AFR populations (83%) compared to EUR populations (68%) (Table 3). The frequency differences by race are consistent with our admixture mapping results that excess of local African ancestry at 10q26 is associated with higher risk of all breast cancer.
11q13 and ER+ Breast Cancer
Three variants, rs116638271 in ALDH3B2, rs77274510 in RP11-703H8.9, and rs117564384 in SCGB1D2 in the chromosome 11q13 region were significantly associated with ER+ breast cancer after correction for multiple testing (Table 3). The latter two are only 15,807 bp apart and are in modest LD (r2 = 0.66) in 1000 Genomes AFR populations, while rs116638271, resides ∼5.4 Mb from rs117564384 and is not correlated (r2 = 0) with either SNP. In agreement with the admixture mapping results (i.e., deficit of local African ancestry at 11q13 associated with higher risk of ER+ breast cancer) the risk alleles of the three SNPs were less frequent in 1000 Genomes AFR vs. EUR populations (Table 3). For rs77274510 and rs117564384, the risk alleles had <1% frequency in 1000 Genomes AFR subjects compared to 5.9 and 5.1%, respectively, in EUR samples. For rs116638271 the risk allele had 97% frequency in AFR vs. 100% in EUR. In other words, the protective rs116638271 allele against ER+ breast cancer seems to be specific of AFR populations with a frequency of about 3% but 0% in EUR populations.
The genome wide significant ER+ associated region on chromosome 11 was also suggestive of association in the admixture analyses of overall breast cancer (Z = 3.9 in case-only analysis, and Z = 1.9 with P = 0.058 in case-control analysis). This region contains three overall breast cancer associated variants within an 8.6 Kb region, rs75313515 (P = 2.4 × 10-6), rs75771004 (P = 3.2 × 10-6) and rs116667969 (P = 3.5 × 10-6) that are significant after correction for multiple testing. The first two SNPs are in strong LD while rs116667969 is very weakly correlated with both SNPs (r2 ∼0.40). The minor alleles range from 2.5 to 4% in our analyses; however, none of the variant alleles at these three SNPs are present in the EUR 1000G populations. These three SNPs are within 10 Kb of rs116638271 (r2 = 0.40), one of the three SNPs under this peak shown to be associated with ER+ breast, which suggests that this region contains multiple independently associated variants.
17q25 and ER+ Breast Cancer
No single variant was significantly associated with ER+ breast cancer after correction for multiple testing. The risk T-allele, at rs55850050, the most significant SNP in the chromosome 17 region, conferred a 55% increased susceptibility to ER+ breast cancer (P = 1.6 × 10-4). The direction of the risk was contrary to what was expected based on the admixture results (i.e., excess of local African ancestry at 17q25 associated with higher risk of ER+ breast cancer), as the risk T-allele was less frequent in 1000 Genomes AFR (2.7%) vs. EUR populations (16.1%) (Table 3). Notably it resides adjacent to BRCA1 (17q21.1) and 13 Mb from the 17q23.1 region which contains breast cancer susceptibility SNPs (Ahmed et al., 2009; Kelemen et al., 2009; Lin et al., 2012).
Regions of Suggestive Genome-Wide Significance
Overall Breast Cancer
The 9q34 region showed excess local African ancestry associated with risk of overall breast cancer (Supplementary Table 3). It does not harbor known susceptibility breast cancer variants and no SNPs were associated with overall breast cancer at nominal P < 0.05. This region contains multiple Crohn’s disease, ulcerative colitis and Type 2 diabetes loci (Supplementary Table 4).
ER+ Breast Cancer
We found an excess of local African ancestry in the chromosome region 8q24 associated with ER-positive breast cancer (Supplementary Table 3). The 8q24 locus contains breast cancer susceptibility loci as well as loci for other cancers including ovarian, prostate, colorectal, glioma, lymphomas and leukemias, bladder and kidney (Supplementary Table 4). No single SNP reached significance after correction for multiple testing. Two SNPs, rs147582253 (P = 1.9 × 10-5) and rs10112657 (P = 5.8 × 10-5) showed suggestive associations with ER+ breast cancer. The direction of the associations were consistent with the admixture mapping results, as the risk alleles of both SNPs, rs147582253-G (1.7% in African vs. 0% in European populations) and rs10112657-T (3.3% in African vs. 0% in European populations), were more frequent in African ancestry vs. European ancestry populations.
ER-Negative (ER-) Breast Cancer
We found three regions, 2q24, 5p13, and 18q23, showing suggestive evidence of association with ER- breast cancer (Supplementary Table 3); both 2q24 and 5p13 contain or are adjacent to breast cancer susceptibility variants. The 2q24 region showed a deficit of local African ancestry associated with ER- breast cancer. The admixture signal in 2q24 is less than 4 Mb from two recently identified SNPs, rs2016394, and rs1550623, associated with breast cancer in European ancestry women in the Collaborative Oncological Gene-Environment Study (COGS) (Michailidou et al., 2013). However, these two variants are not specific to ER- breast cancer (Michailidou et al., 2013); thus, they do not explain the admixture signal in 2q24. Our fine-mapping efforts found two independent signals associated with ER- breast cancer. Both protective alleles, rs78550178-G (99% frequency in African vs. 84% in European populations) and rs62172609-G (76% frequency in African vs. 31% in European populations), were more frequent in African ancestry vs. European ancestry populations as expected according to the direction of the admixture signal.
Region 5p15.33, showing excess African ancestry associations with ER- breast cancer, contains the TERT gene, which harbors a well-known ER- specific breast cancer associated variant, rs10069690 (Haiman et al., 2011; Garcia-Closas et al., 2013; Michailidou et al., 2013; Purrington et al., 2014). The T allele of rs10069690 was our most significant association with ER- breast cancer (P = 2.2 × 10-5). The T allele is far more frequent in African populations (66%) versus Europeans (28%). We found some evidence of one additional independent signal in TERT, represented by the risk increasing C allele of rs37004 (P = 6.8 × 10-5, 99% frequency in AFR vs. 78% in EUR) at ∼1 Mb from rs10069690. This variant has been shown to be associated with lung cancer risk in the Genetic Association and Mechanisms in Oncology (GAME-ON) consortium (Karami et al., 2015).
The 18q23 region is novel and showed an excess of local African ancestry associated with risk of ER- breast cancer, however the most significantly associated single SNP, rs58839649 at P = 1.5 × 10-4, did not pass multiple test correction; the minor G-allele was associated with a 60% increased risk of ER- breast cancer with a frequency in AABC of 10, 13% in AFR populations, and absent in EUR populations. This region contains prostate susceptibility SNPs (Eeles et al., 2013) and has been identified by linkage analyses as a region co-segregating with familial Glioma (Liu et al., 2012) (Supplementary Table 4).
Triple-Negative Breast Cancer
We found evidence of association of excess African ancestry with triple negative disease at three locations: 2q37.1, 3p24.2, and 18p11 (Supplementary Table 3). While higher African ancestry in the 2q37 and 3p24 loci seems to increase risk of ER- breast cancer as well, higher African ancestry in the 18p11 locus appears to be associated only with risk of triple negative breast cancer. The 3p24.2 region contains well-established breast cancer associated SNPs (Matsumoto et al., 1997, 2000; Yang et al., 2001; Ahmed et al., 2009; Lin et al., 2012). However, there are no reports of variants at 2q37.1 or 18p11 associated with breast cancer or breast cancer subtypes. Given these regions of excess African ancestry are associated with disease; it may be possible that these locations harbor breast cancer susceptibility alleles only present in African ancestry populations.
Discussion
In this large genome-wide admixture scan in African American women, we found evidence of associations of local African ancestry with breast cancer in several genomic regions. Tests of genetic associations in the identified regions in an independent sample suggest that continental ancestry specific genetic variation within these regions may be associated with breast cancer subtypes.
4p16 and ER-Positive Breast Cancer
This region contains no previously reported genome wide significant breast cancer loci, but loci for bladder cancer, as well as other traits and diseases include triglycerides and cholesterol levels, bone mineral density, type 2 diabetes and Parkinson’s disease (Supplementary Table 4). The most strongly associated SNP in 4p16, rs112545418, in ZFYVE28, was in moderate linkage disequilibrium (r2 = 0.48) with a nearby variant rs17132398, associated with ER-positive breast cancer at P = 1.8 × 10-4. Analysis of rs17132398 using RegulomeDB database (Boyle et al., 2012) showed that this SNP is located inside an enhancer region across multiple cell lines, and is likely to affect binding of transcription factors and expression of ZFYVE28, which is a negative regulator of epidermal growth factor receptor (EGFR) signaling (Mosesson et al., 2009). The low frequency of the rs112545418 risk A-allele (1%) and the corresponding risk T-allele of rs17132398 (0.4%) in European ancestry populations may explain why it has not been identified in previous GWAS of breast cancer.
10q26 and Overall Breast Cancer
The 10q26 showed excess of local African ancestry associated with risk of overall breast cancer and was our strongest admixture association with overall breast cancer. This region contains well established SNPs in FGFR2 associated with increased risk of breast cancer (Chen et al., 2011). A recent fine-mapping of FGFR2 (Meyer et al., 2013) identified three independent signals in Europeans and East Asians: the first signal represented by rs35054928, the second signal by rs45631563, and the third one by rs2981578 (Meyer et al., 2013). The rs2981578 was previously identified as the SNP with the strongest association with overall and ER+ breast cancer in FGFR2 in African Americans (Chen et al., 2011), and it was also shown recently by us using AMBER consortium data (Ruiz-Narvaez et al., 2016). Interestingly, the excess of African ancestry in breast cancer cases extends over both FGFR2 and a second region spanning from 131 to 134 Mb on chromosome 10. This second region contains our significant variant, rs12244041 (Table 3; Supplementary Figure 2), as well as a non-synonymous variant, rs148966337, previously shown to be associated with both overall and ER- breast cancer (Haiman et al., 2013). This is a less common variant (1.9%) that was only present in the African American women from the MEC study, but completely absent in women of other continental ancestries (Haiman et al., 2013).
11q13 and ER-Positive Breast Cancer
Although the chromosome 11q13 region has been previously shown to contain breast cancer SNPs (Michailidou et al., 2013; Ahsan et al., 2014), all three of the SNPs we found within the admixture-identified region were independent of the published GWAS-identified SNPs. One of the three SNPs in the chromosome 11 region, rs117564384, resides in SCGB1D2. SCGB1D2 expression is high in and almost exclusive to mammary tissue (Culleton et al., 2007), significantly differs between histologically normal breast tissue and breast cancer tissue (Zubor et al., 2015), and it also has been used as a marker to detect disseminated tumor cells (DTC) in breast cancer (Brown et al., 2006; Lacroix, 2006).
In summary, this is the largest admixture analysis of African American breast cancer, both in number of subjects and markers, to date. One limitation of this report is the lack of characterization of tumors by grade and stage that may show differential association with locus-specific African ancestry. Nevertheless, the current study includes more than twice the number cases and a higher density of AIMs throughout the genome compared to the previous admixture mapping of breast cancer in AA women (Fejerman et al., 2009). In addition, the use of controls in the present study allowed us to confirm significant results from the case-only scan. Fine-mapping of the identified regions showed that multiple genomic variants may be within admixture associated regions. Our results suggest that several loci with small to moderate effects rather than a few with strong effects are responsible for the differences of incidence of ER subtypes among US white and black women.
Author Contributions
ER-N and LS-C performed the statistical analyses and drafted the manuscript. KL supervised the statistical analyses. JB, SY, SH, CH, EB, AO, CA, JP, and KL participated in data analysis and interpretation and provided critical feedback. CH, EJ, LB, JH, RZ, and SD provided data for fine-mapping. CA and JP conceived of the study and participated in the design and coordination of the study. All authors read and approved the final manuscript.
Funding
The research conducted by the AMBER Consortium is funded by the National Institutes of Health and Foundation grants: P01 CA151135 (CBA, JRP, AFO); R01 CA058420 (LR); UM1 CA164974 (JRP, LR); R01 CA098663 (JRP); R01 CA100598 (CBA); R01 CA185623 (EVB, CCH, KD); UM1 CA164973 (LLM, LKM, CAH); R01 CA54281 (LNK); R01 CA063464 (BH); P50 CA58223 (MAT, AFO); U01 CA179715 (MAT, AFO); Department of Defense Breast Cancer Research Program, Era of Hope Scholar Award Program W81XWH-08-1-0383 (CAH); the Susan G. Komen for the Cure Foundation (MAT, AFO); the Breast Cancer Research Foundation (CBA); and the University Cancer Research Fund of North Carolina (MAT, AFO).
Conflict of Interest Statement
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
The reviewer JP and handling Editor declared their shared affiliation, and the handling Editor states that the process nevertheless met the standards of a fair and objective review.
Acknowledgments
We thank participants and staff of the contributing studies. We wish also to acknowledge the late Robert Millikan, DVM, MPH, Ph.D., who was instrumental in the creation of this consortium. Pathology data were obtained from numerous state cancer registries (Arizona, California, Colorado, Connecticut, Delaware, District of Columbia, Florida, Georgia, Hawaii, Illinois, Indiana, Kentucky, Louisiana, Maryland, Massachusetts, Michigan, New Jersey, New York, North Carolina, Oklahoma, Pennsylvania, South Carolina, Tennessee, Texas, and Virginia). The results reported do not necessarily represent their views or the views of the NIH.
Supplementary Material
The Supplementary Material for this article can be found online at: http://journal.frontiersin.org/article/10.3389/fgene.2016.00170
TABLE 1 | Number of ancestral informative markers (AIMs) per chromosome.
TABLE 2 | Admixture mapping results for all 2624 ancestral informative markers and breast cancer subtypes in the AMBER consortium.
TABLE 3 | Genomic regions with suggestive excess (negative Z-Score) and reduced (positive Z-score) African ancestry associated with breast cancer at 4 >|Z| ≤ 3.5 in case-only analyses and corresponding case-control results.
TABLE 4 | Phenotypic traits with genome-wide significant associations at the identified breast cancer loci.
References
Ahmed, S., Thomas, G., Ghoussaini, M., Healey, C. S., Humphreys, M. K., Platte, R., et al. (2009). Newly discovered breast cancer susceptibility loci on 3p24 and 17q23.2. Nat. Genet. 41, 585–590. doi: 10.1038/ng.354
Ahsan, H., Halpern, J., Kibriya, M. G., Pierce, B. L., Tong, L., Gamazon, E., et al. (2014). A genome-wide association study of early-onset breast cancer identifies PFKM as a novel breast cancer gene and supports a common genetic spectrum for breast cancer at any age. Cancer Epidemiol. Biomarkers Prev. 23, 658–669. doi: 10.1158/1055-9965.EPI-13-0340
Ambrosone, C. B., Ciupak, G. L., Bandera, E. V., Jandorf, L., Bovbjerg, D. H., Zirpoli, G., et al. (2009). Conducting molecular epidemiological research in the age of HIPAA: a multi-institutional case-control study of breast cancer in african-american and European-American women. J. Oncol. 2009, 871250. doi: 10.1155/2009/871250
Bandera, E. V., Chandran, U., Hong, C. C., Troester, M. A., Bethea, T. N., Adams-Campbell, L. L., et al. (2015). Obesity, body fat distribution, and risk of breast cancer subtypes in African American women participating in the AMBER consortium. Breast Cancer Res. Treat. 150, 655–666. doi: 10.1007/s10549-015-3353-z
Bandera, E. V., Chandran, U., Zirpoli, G., McCann, S. E., Ciupak, G., and Ambrosone, C. B. (2013). Rethinking sources of representative controls for the conduct of case-control studies in minority populations. BMC Med. Res. Methodol. 13:71. doi: 10.1186/1471-2288-13-71
Bensen, J. T., Xu, Z., McKeigue, P. M., Smith, G. J., Fontham, E. T., Mohler, J. L., et al. (2014). Admixture mapping of prostate cancer in African Americans participating in the North Carolina-Louisiana Prostate Cancer Project (PCaP). Prostate 74, 1–9. doi: 10.1002/pros.22722
Boyle, A. P., Hong, E. L., Hariharan, M., Cheng, Y., Schaub, M. A., Kasowski, M., et al. (2012). Annotation of functional variation in personal genomes using RegulomeDB. Genome Res. 22, 1790–1797. doi: 10.1101/gr.137323.112
Brown, N. M., Stenzel, T. T., Friedman, P. N., Henslee, J., Huper, G., and Marks, J. R. (2006). Evaluation of expression based markers for the detection of breast cancer cells. Breast Cancer Res. Treat. 97, 41–47. doi: 10.1007/s10549-005-9085-8
Chen, F., Chen, G. K., Millikan, R. C., John, E. M., Ambrosone, C. B., Bernstein, L., et al. (2011). Fine-mapping of breast cancer susceptibility loci characterizes genetic risk in African Americans. Hum. Mol. Genet. 20, 4491–4503. doi: 10.1093/hmg/ddr367
Chen, F., Chen, G. K., Stram, D. O., Millikan, R. C., Ambrosone, C. B., John, E. M., et al. (2013). A genome-wide association study of breast cancer in women of African ancestry. Hum. Genet. 132, 39–48. doi: 10.1007/s00439-012-1214-y
Chlebowski, R. T., Chen, Z., Anderson, G. L., Rohan, T., Aragaki, A., Lane, D., et al. (2005). Ethnicity and breast cancer: factors influencing differences in incidence and outcome. J. Natl. Cancer Inst. 97, 439–448. doi: 10.1093/jnci/dji346
Clarke, C. A., Keegan, T. H., Yang, J., Press, D. J., Kurian, A. W., Patel, A. H., et al. (2012). Age-specific incidence of breast cancer subtypes: understanding the black-white crossover. J. Natl. Cancer Inst. 104, 1094–1101. doi: 10.1093/jnci/djs264
Couch, F. J., Kuchenbaecker, K. B., Michailidou, K., Mendoza-Fandino, G. A., Nord, S., Lilyquist, J., et al. (2016). Identification of four novel susceptibility loci for oestrogen receptor negative breast cancer. Nat. Commun. 7:11375. doi: 10.1038/ncomms11375
Cozier, Y. C., Palmer, J. R., and Rosenberg, L. (2004). Comparison of methods for collection of DNA samples by mail in the Black Women’s Health Study. Ann. Epidemiol. 14, 117–122. doi: 10.1016/S1047-2797(03)00132-7
Culleton, J., O’Brien, N., Ryan, B. M., Hill, A. D., McDermott, E., O’Higgins, N., et al. (2007). Lipophilin B: a gene preferentially expressed in breast tissue and upregulated in breast cancer. Int. J. Cancer 120, 1087–1092. doi: 10.1002/ijc.22471
DeSantis, C., Ma, J., Bryan, L., and Jemal, A. (2014). Breast cancer statistics, 2013. CA Cancer J. Clin. 64, 52–62. doi: 10.3322/caac.21203
Eeles, R. A., Olama, A. A., Benlloch, S., Saunders, E. J., Leongamornlert, D. A., Tymrakiewicz, M., et al. (2013). Identification of 23 new prostate cancer susceptibility loci using the iCOGS custom genotyping array. Nat. Genet. 45, e381–e382. doi: 10.1038/ng.2560
Fejerman, L., Haiman, C. A., Reich, D., Tandon, A., Deo, R. C., John, E. M., et al. (2009). An admixture scan in 1,484 African American women with breast cancer. Cancer Epidemiol. Biomarkers Prev. 18, 3110–3117. doi: 10.1158/1055-9965.EPI-09-0464
Fejerman, L., and Ziv, E. (2008). Population differences in breast cancer severity. Pharmacogenomics 9, 323–333. doi: 10.2217/14622416.9.3.323
Feng, Y., Stram, D. O., Rhie, S. K., Millikan, R. C., Ambrosone, C. B., John, E. M., et al. (2014). A comprehensive examination of breast cancer risk loci in African American women. Hum. Mol. Genet. 23, 5518–5526. doi: 10.1093/hmg/ddu252
Freedman, M. L., Haiman, C. A., Patterson, N., McDonald, G. J., Tandon, A., Waliszewska, A., et al. (2006). Admixture mapping identifies 8q24 as a prostate cancer risk locus in African-American men. Proc. Natl. Acad. Sci. U.S.A. 103, 14068–14073. doi: 10.1073/pnas.0605832103
Garcia-Closas, M., Couch, F. J., Lindstrom, S., Michailidou, K., Schmidt, M. K., Brook, M. N., et al. (2013). Genome-wide association studies identify four ER negative-specific breast cancer risk loci. Nat. Genet. 45, e391–e392. doi: 10.1038/ng.2561
Grann, V., Troxel, A. B., Zojwalla, N., Hershman, D., Glied, S. A., and Jacobson, J. S. (2006). Regional and racial disparities in breast cancer-specific mortality. Soc. Sci Med. 62, 337–347. doi: 10.1016/j.socscimed.2005.06.038
Haiman, C. A., Chen, G. K., Vachon, C. M., Canzian, F., Dunning, A., Millikan, R. C., et al. (2011). A common variant at the TERT-CLPTM1L locus is associated with estrogen receptor-negative breast cancer. Nat. Genet. 43, 1210–1214. doi: 10.1038/ng.985
Haiman, C. A., Han, Y., Feng, Y., Xia, L., Hsu, C., Sheng, X., et al. (2013). Genome-wide testing of putative functional exonic variants in relationship with breast and prostate cancer risk in a multiethnic population. PLoS Genet. 9:e1003419. doi: 10.1371/journal.pgen.1003419
Hershman, D., McBride, R., Jacobson, J. S., Lamerato, L., Roberts, K., Grann, V. R., et al. (2005). Racial disparities in treatment and survival among women with early-stage breast cancer. J. Clin. Oncol. 23, 6639–6646. doi: 10.1200/JCO.2005.12.633
Hindorff, L. A., MacArthur, J., Morales, J., Junkins, H. A., Hall, P. N., Klemm, A. K., et al. (2015). A Catalog of Published Genome-Wide Association Studies. Available at: www.genome.gov/gwastudies [accessed April 30, 2015]
Hoggart, C. J., Shriver, M. D., Kittles, R. A., Clayton, D. G., and McKeigue, P. M. (2004). Design and analysis of admixture mapping studies. Am. J. Hum. Genet. 74, 965–978. doi: 10.1086/420855
Hunter, D. J., Kraft, P., Jacobs, K. B., Cox, D. G., Yeager, M., Hankinson, S. E., et al. (2007). A genome-wide association study identifies alleles in FGFR2 associated with risk of sporadic postmenopausal breast cancer. Nat. Genet. 39, 870–874. doi: 10.1038/ng2075
Jeff, J. M., Armstrong, L. L., Ritchie, M. D., Denny, J. C., Kho, A. N., Basford, M. A., et al. (2014). Admixture mapping and subsequent fine-mapping suggests a biologically relevant and novel association on chromosome 11 for type 2 diabetes in African Americans. PLoS ONE 9:e86931. doi: 10.1371/journal.pone.0086931
Karami, S., Han, Y., Schumacher, F. R., Kote-Jarai, Z., Lindstrom, S., Witte, J. S., et al. (2015). Abstract 4609: Risk loci in telomere structure and maintenance genes across five cancer types: GAME-ON Consortium. Cancer Res. 75, 4609–4609. doi: 10.1158/1538-7445.am2015-4609
Kelemen, L. E., Wang, X., Fredericksen, Z. S., Pankratz, V. S., Pharoah, P. D., Ahmed, S., et al. (2009). Genetic variation in the chromosome 17q23 amplicon and breast cancer risk. Cancer Epidemiol. Biomarkers Prev. 18, 1864–1868. doi: 10.1158/1055-9965.EPI-08-0486
Kolonel, L. N., Henderson, B. E., Hankin, J. H., Nomura, A. M., Wilkens, L. R., Pike, M. C., et al. (2000). A multiethnic cohort in Hawaii and Los Angeles: baseline characteristics. Am. J. Epidemiol. 151, 346–357. doi: 10.1093/oxfordjournals.aje.a010213
Lacroix, M. (2006). Significance, detection and markers of disseminated breast cancer cells. Endocr. Relat. Cancer 13, 1033–1067. doi: 10.1677/ERC-06-0001
Laurie, C. C., Doheny, K. F., Mirel, D. B., Pugh, E. W., Bierut, L. J., Bhangale, T., et al. (2010). Quality control and quality assurance in genotypic data for genome-wide association studies. Genet. Epidemiol. 34, 591–602. doi: 10.1002/gepi.20516
Lin, C. Y., Ho, C. M., Bau, D. T., Yang, S. F., Liu, S. H., Lin, P. H., et al. (2012). Evaluation of breast cancer susceptibility loci on 2q35, 3p24, 17q23 and FGFR2 genes in Taiwanese women with breast cancer. Anticancer. Res. 32, 475–482.
Liu, Y., Melin, B. S., Rajaraman, P., Wang, Z., Linet, M., Shete, S., et al. (2012). Insight in glioma susceptibility through an analysis of 6p22.3, 12p13.33-12.1, 17q22-23.2 and 18q23 SNP genotypes in familial and non-familial glioma. Hum. Genet. 131, 1507–1517. doi: 10.1007/s00439-012-1187-x
Manuck, T. A., Lai, Y., Meis, P. J., Sibai, B., Spong, C. Y., Rouse, D. J., et al. (2011). Admixture mapping to identify spontaneous preterm birth susceptibility loci in African Americans. Obstet. Gynecol. 117, 1078–1084. doi: 10.1097/AOG.0b013e318214e67f
Matsumoto, S., Kasumi, F., Sakamoto, G., Onda, M., Nakamura, Y., and Emi, M. (1997). Detailed deletion mapping of chromosome arm 3p in breast cancers: a 2-cM region on 3p14.3-21.1 and a 5-cM region on 3p24.3-25.1 commonly deleted in tumors. Genes Chromosomes Cancer 20, 268–274. doi: 10.1002/(SICI)1098-2264(199711)20:3<268::AID-GCC7>3.3.CO;2-T
Matsumoto, S., Minobe, K., Utada, Y., Furukawa, K., Onda, M., Sakamoto, G., et al. (2000). Loss of heterozygosity at 3p24-p25 as a prognostic factor in breast cancer. Cancer Lett. 152, 63–69. doi: 10.1016/S0304-3835(99)00431-0
Maxwell, K. N., and Nathanson, K. L. (2013). Common breast cancer risk variants in the post-COGS era: a comprehensive review. Breast Cancer Res. 15, 212. doi: 10.1186/bcr3591
Meyer, K. B., O’Reilly, M., Michailidou, K., Carlebur, S., Edwards, S. L., French, J. D., et al. (2013). Fine-scale mapping of the FGFR2 breast cancer risk locus: putative functional variants differentially bind FOXA1 and E2F1. Am. J. Hum. Genet. 93, 1046–1060. doi: 10.1016/j.ajhg.2013.10.026
Michailidou, K., Beesley, J., Lindstrom, S., Canisius, S., Dennis, J., Lush, M. J., et al. (2015). Genome-wide association analysis of more than 120,000 individuals identifies 15 new susceptibility loci for breast cancer. Nat. Genet. 47, 373–380. doi: 10.1038/ng.3242
Michailidou, K., Hall, P., Gonzalez-Neira, A., Ghoussaini, M., Dennis, J., Milne, R. L., et al. (2013). Large-scale genotyping identifies 41 new loci associated with breast cancer risk. Nat. Genet. 45, e351–e352. doi: 10.1038/ng.2563
Montana, G., and Pritchard, J. K. (2004). Statistical tests for admixture mapping with case-control and cases-only data. Am. J. Hum. Genet. 75, 771–789. doi: 10.1086/425281
Mosesson, Y., Chetrit, D., Schley, L., Berghoff, J., Ziv, T., Carvalho, S., et al. (2009). Monoubiquitinylation regulates endosomal localization of Lst2, a negative regulator of EGF receptor signaling. Dev. Cell 16, 687–698. doi: 10.1016/j.devcel.2009.03.015
Nalls, M. A., Wilson, J. G., Patterson, N. J., Tandon, A., Zmuda, J. M., Huntsman, S., et al. (2008). Admixture mapping of white cell count: genetic locus responsible for lower white blood cell count in the health ABC and Jackson Heart studies. Am. J. Hum. Genet. 82, 81–87. doi: 10.1016/j.ajhg.2007.09.003
Newman, B., Moorman, P. G., Millikan, R., Qaqish, B. F., Geradts, J., Aldrich, T. E., et al. (1995). The Carolina Breast Cancer Study: integrating population-based epidemiology and molecular biology. Breast Cancer Res. Treat. 35, 51–60. doi: 10.1007/BF00694745
Palmer, J. R., Ambrosone, C. B., and Olshan, A. F. (2014a). A collaborative study of the etiology of breast cancer subtypes in African American women: the AMBER consortium. Cancer Causes Control 25, 309–319. doi: 10.1007/s10552-013-0332-8
Palmer, J. R., Viscidi, E., Troester, M. A., Hong, C. C., Schedin, P., Bethea, T. N., et al. (2014b). Parity, lactation, and breast cancer subtypes in African American women: results from the AMBER Consortium. J. Natl. Cancer Inst. 106, dju237. doi: 10.1093/jnci/dju237
Palmer, N. D., Divers, J., Lu, L., Register, T. C., Carr, J. J., Hicks, P. J., et al. (2016). Admixture mapping of serum vitamin D and parathyroid hormone concentrations in the African American-Diabetes Heart Study. Bone 87, 71–77. doi: 10.1016/j.bone.2016.01.028
Parker, M. M., Foreman, M. G., Abel, H. J., Mathias, R. A., Hetmanski, J. B., Crapo, J. D., et al. (2014). Admixture mapping identifies a quantitative trait locus associated with FEV1 /FVC in the COPDGene study. Genet. Epidemiol. 38, 652–659. doi: 10.1002/gepi.21847
Patterson, N., Price, A. L., and Reich, D. (2006). Population structure and eigenanalysis. PLoS Genet. 2:e190. doi: 10.1371/journal.pgen.0020190
Pike, M. C., Kolonel, L. N., Henderson, B. E., Wilkens, L. R., Hankin, J. H., Feigelson, H. S., et al. (2002). Breast cancer in a multiethnic cohort in Hawaii and Los Angeles: risk factor-adjusted incidence in Japanese equals and in Hawaiians exceeds that in whites. Cancer Epidemiol. Biomarkers. Prev. 11, 795–800.
Purrington, K. S., Slager, S., Eccles, D., Yannoukakos, D., Fasching, P. A., Miron, P., et al. (2014). Genome-wide association study identifies 25 known breast cancer susceptibility loci as risk factors for triple-negative breast cancer. Carcinogenesis 35, 1012–1019. doi: 10.1093/carcin/bgt404
Reich, D., and Patterson, N. (2005). Will admixture mapping work to find disease genes? Philos. Trans. R Soc. Lond. B Biol. Sci. 360, 1605–1607. doi: 10.1098/rstb.2005.1691
Rosenberg, L., Adams-Campbell, L., and Palmer, J. R. (1995). The Black Women’s Health Study: a follow-up study for causes and preventions of illness. J. Am. Med. Womens. Assoc. 50, 56–58.
Rosenberg, N. A., Li, L. M., Ward, R., and Pritchard, J. K. (2003). Informativeness of genetic markers for inference of ancestry. Am. J. Hum. Genet. 73, 1402–1422. doi: 10.1086/380416
Ruiz-Narvaez, E. A., Haddad, S. A., Lunetta, K. L., Yao, S., Bensen, J. T., Sucheston-Campbell, L. E., et al. (2016). Gene-based analysis of the fibroblast growth factor receptor signaling pathway in relation to breast cancer in African American women: the AMBER consortium. Breast Cancer Res. Treat. 155, 355–363. doi: 10.1007/s10549-015-3672-0
Shriner, D. (2013). Overview of admixture mapping. Curr. Protoc. Hum. Genet. Chapter 1:Unit 1. 23. doi: 10.1002/0471142905.hg0123s76
Smith, M. W., and O’Brien, S. J. (2005). Mapping by admixture linkage disequilibrium: advances, limitations and guidelines. Nat. Rev. Genet. 6, 623–632. doi: 10.1038/nrg1657
Turnbull, C., Ahmed, S., Morrison, J., Pernet, D., Renwick, A., Maranian, M., et al. (2010). Genome-wide association study identifies five new breast cancer susceptibility loci. Nat. Genet. 42, 504–507. doi: 10.1038/ng.586
Winkler, C. A., Nelson, G. W., and Smith, M. W. (2010). Admixture mapping comes of age. Annu. Rev. Genomics Hum. Genet. 11, 65–89. doi: 10.1146/annurev-genom-082509-141523
Yang, Q., Yoshimura, G., Nakamura, M., Nakamura, Y., Shan, L., Suzuma, T., et al. (2001). Allelic loss of chromosome 3p24 correlates with tumor progression rather than with retinoic acid receptor beta2 expression in breast carcinoma. Breast Cancer Res. Treat. 70, 39–45. doi: 10.1023/A:1012574305832
Zhu, X., Luke, A., Cooper, R. S., Quertermous, T., Hanis, C., Mosley, T., et al. (2005). Admixture mapping for hypertension loci with genome-scan markers. Nat. Genet. 37, 177–181. doi: 10.1038/ng1510
Keywords: breast cancer, African American women, admixture mapping, genetics, fine-mapping
Citation: Ruiz-Narváez EA, Sucheston-Campbell L, Bensen JT, Yao S, Haddad S, Haiman CA, Bandera EV, John EM, Bernstein L, Hu JJ, Ziegler RG, Deming SL, Olshan AF, Ambrosone CB, Palmer JR and Lunetta KL (2016) Admixture Mapping of African–American Women in the AMBER Consortium Identifies New Loci for Breast Cancer and Estrogen-Receptor Subtypes. Front. Genet. 7:170. doi: 10.3389/fgene.2016.00170
Received: 28 July 2016; Accepted: 07 September 2016;
Published: 21 September 2016.
Edited by:
Logan Walker, University of Otago, New ZealandReviewed by:
Parvin Mehdipour, Tehran University of Medical Sciences, IranJohn Frederick Pearson, University of Otago, New Zealand
Copyright © 2016 Ruiz-Narváez, Sucheston-Campbell, Bensen, Yao, Haddad, Haiman, Bandera, John, Bernstein, Hu, Ziegler, Deming, Olshan, Ambrosone, Palmer and Lunetta. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) or licensor are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Edward A. Ruiz-Narváez, ZXJ1aXpuYXJAYnUuZWR1
†These authors have contributed equally to this work.