 Błażej Misiak
Błażej Misiak Laura Ricceri
Laura Ricceri Maria M. Sąsiadek1
Maria M. Sąsiadek1- 1Department of Genetics, Wrocław Medical University, Wrocław, Poland
- 2Centre for Behavioural Sciences and Mental Health, Istituto Superiore di Sanità, Rome, Italy
Transposable elements (TEs) are highly repetitive DNA sequences in the human genome that are the relics of previous retrotransposition events. Although the majority of TEs are transcriptionally inactive due to acquired mutations or epigenetic processes, around 8% of TEs exert transcriptional activity. It has been found that TEs contribute to somatic mosaicism that accounts for functional specification of various brain cells. Indeed, autonomous retrotransposition of long interspersed element-1 (LINE-1) sequences has been reported in the neural rat progenitor cells from the hippocampus, the human fetal brain and the human embryonic stem cells. Moreover, expression of TEs has been found to regulate immune-inflammatory responses, conditioning immunity against exogenous infections. Therefore, aberrant epigenetic regulation and expression of TEs emerged as a potential mechanism underlying the development of various mental disorders, including autism spectrum disorders (ASD), schizophrenia, bipolar disorder, major depression, and Alzheimer’s disease (AD). Consequently, some studies revealed that expression of some sequences of human endogenous retroviruses (HERVs) appears only in a certain group of patients with mental disorders (especially those with schizophrenia, bipolar disorder, and ASD) but not in healthy controls. In addition, it has been found that expression of HERVs might be related to subclinical inflammation observed in mental disorders. In this article, we provide an overview of detrimental effects of transposition on the brain development and immune mechanisms with relevance to mental disorders. We show that transposition is not the only mechanism, explaining the way TEs might shape the phenotype of mental disorders. Other mechanisms include the regulation of gene expression and the impact on genomic stability. Next, we review current evidence from studies investigating expression and epigenetic regulation of specific TEs in various mental disorders. Most consistently, these studies indicate altered expression of HERVs and methylation of LINE-1 sequences in patients with ASD, schizophrenia, and mood disorders. However, the contribution of TEs to the etiology of AD is poorly documented. Future studies should further investigate the mechanisms linking epigenetic processes, specific TEs and the phenotype of mental disorders to disentangle causal associations.
Introduction
Mental disorders represent complex phenotypes and are the leading causes of global disease burden (Vigo et al., 2016). The phenotype complexity of mental disorders manifests in symptomatic and biological overlap, impeding a diagnostic process that is based on a clinical consensus without a crucial role of biological markers. Heritability rates of mental disorders are high, exceeding 80% in twin studies of schizophrenia and bipolar disorder (Cardno and Gottesman, 2000; McGuffin et al., 2003; Misiak et al., 2016). However, monogenic determinants with high penetrance rates have not been identified so far, and the concept of major mental disorders as the polygenic phenotypes prevails in the research approaches. Consequently, a paradigm shift toward investigating polygenic signatures, gene × environment (G × E) interactions and epigenetic mechanisms has been widely observed in the recent years.
The term ‘epigenetics’ refers to a number of reversible mechanisms that impacts gene expression without altering DNA sequence, and include DNA methylation and hydroxymethylation at the CpG islands, histone modifications as well as the regulation by microRNA species. It is now increasingly being recognized that the brain development is a complex process during which there is an increased sensitivity to the regulatory effects of epigenetic mechanisms (Nagy and Turecki, 2012). In light of existing evidence, major mental disorders, especially schizophrenia and autism spectrum disorders (ASD), are perceived as the neurodevelopmental disorders, occurring due to the effects of various genetic and environmental factors that affect critical periods of the brain development (Meredith, 2015).
Transposable elements (TEs) are the highly repetitive DNA sequences that constitute more than 50% of the human genome and contain about 52% of all CpG dinucleotides (Cordaux and Batzer, 2009; Su et al., 2012). Therefore, methylation at TEs is believed to serve as a proxy measure of global DNA methylation. Some TEs share similarity to exogenous viral agents and thus they are called endogenous retroviruses (Griffiths, 2001). Only about 7% of TEs are transcriptionally active (Oja et al., 2008). It has been estimated that approximately 0.27% of human genetic diseases are caused by retrotransposition (Callinan and Batzer, 2006).
Less is known about the contribution of TEs to the etiology of mental disorders. However, there is accumulating evidence that retrotransposition plays an important role in shaping somatic mosaicism that accounts for functional specification of brain cells (Baillie et al., 2011; Poduri et al., 2013). For instance, it has been reported that the transposition of long interspersed element (LINE)-1 sequences may play a role in differentiation of neurons during the brain development (Muotri et al., 2010). Moreover, this sequence exerts autonomous retrotransposition activity in the neural rat progenitor cells from the hippocampus, the human fetal brain and the human embryonic stem cells (Muotri et al., 2005; Coufal et al., 2009). Therefore, aberrant epigenetic regulation of TEs has been hypothesized to play an important role in the development of mental disorders. In this article, we provide an overview of transposition processes with relevance to major psychiatric disorders. Next, we review human and animal model studies investigating expression and epigenetic regulation of TEs in various mental disorders. Finally, we provide a summary of evidence with future directions and potential translation of findings to personalized precision medicine.
Brief Overview of TEs in the Human Genome – Classification and Nomenclature
Classification of TEs in the human genome was shown in Figure 1. A detailed description of the structure and function of various TEs can be found elsewhere (Darby and Sabunciyan, 2014). All TEs can be divided into two subgroups – type I TEs (retrotransposons) and type II TEs (DNA transposons). Type I TEs can be divided into two subgroups – long terminal repeat (LTR) elements, represented by the human endogenous retroviruses (HERVs) and non-LTR sequences that include LINEs, short interspersed elements (SINEs) and processed pseudogenes (Dewannieux and Heidmann, 2005). Retrotransposons act via RNA intermediates that are converted to DNA sequences before transposition (reverse transcription) (Munoz-Lopez and Garcia-Perez, 2010). Type II TEs encode enzymes required for insertion and excision, enabling direct transposition processes without the use of RNA intermediates (Pray, 2008b). Some TEs are autonomous and encode all enzymes that are necessary for transposition, while the rest of them require a transcriptional activity of other transposons. Type II TEs have lost a transposition activity (Darby and Sabunciyan, 2014).
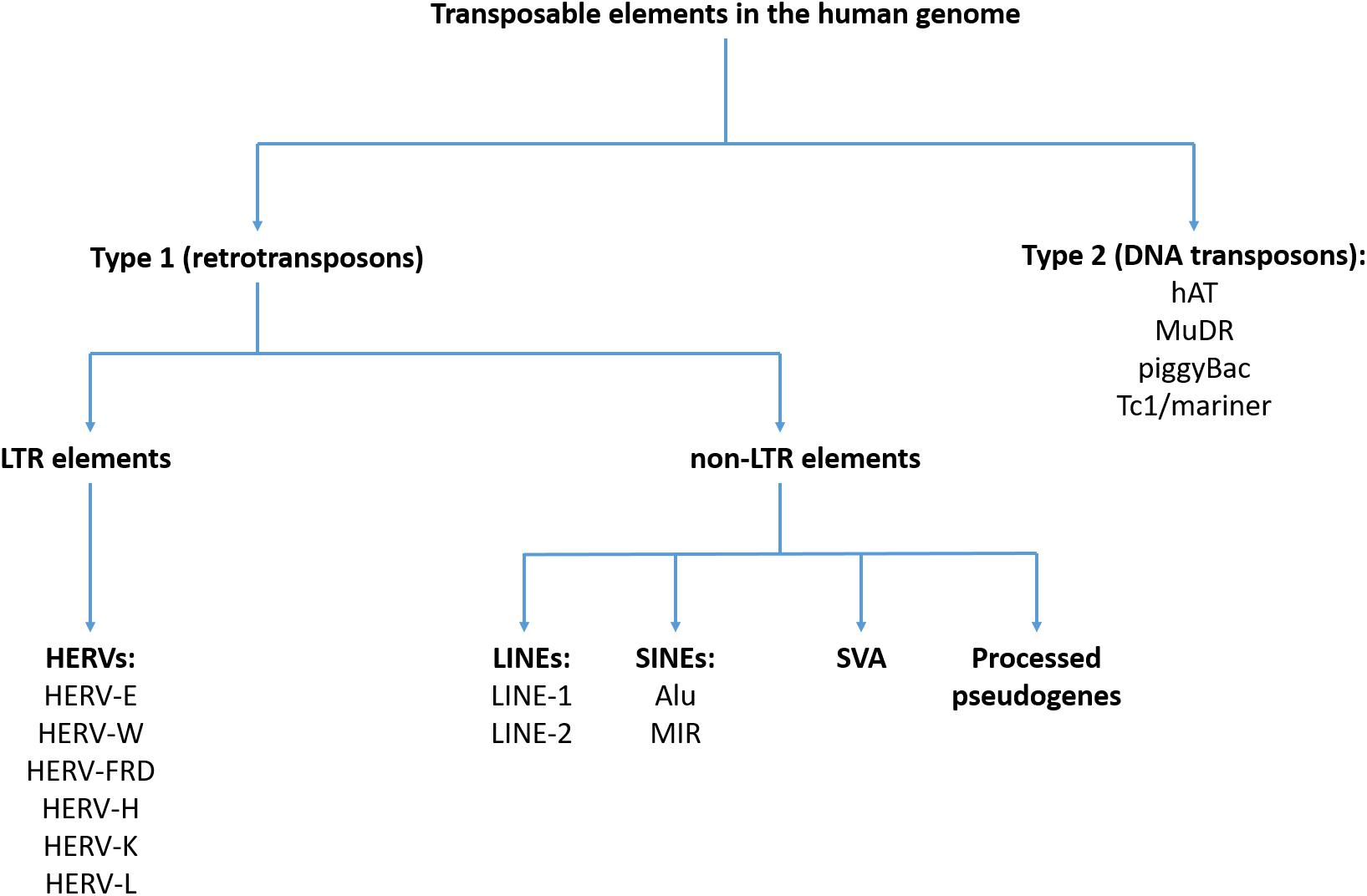
Figure 1. Classification of transposable elements in the human genome.
The HERV sequences have likely existed as exogenous infectious factors; however, they have lost this activity due to acquired mutations (Bannert and Kurth, 2006). These TEs constitute 8% of the human genome and contain genes that are conservative for all retroviruses, including the gag, pro, pol, and env genes (Lander et al., 2001; Vargiu et al., 2016). The gag gene encodes proteins that build up matrix, capsid and nucleocapsid. Pro and pol encode protease, reverse transcriptase and integrase. In turn, the env gene is expressed to surface and transmembrane proteins. The HERV sequences in the human genome represent three classes of retroviruses: class I (e.g., HERV-E, HERV-W, HERV-FRD, and HERV-H), class II (e.g., HERV-K), and class III (e.g., HERV-L). This classification is based on the similarity to exogenous retroviruses. The HERV-K sequences are the youngest and exert the highest transcriptional activity. The HERV sequences can provide promoters, enhancers, repressors, poly-A signals and alternative splicing sites for human transcripts (Vargiu et al., 2016).
The LINEs that represent non-LTR elements, possess an autonomous retrotransposition activity and include LINE-1 and LINE-2 sequences. These sequences make up approximately 21% of the human genome (Lander et al., 2001; Schumann et al., 2010). The LINE-1 sequences contain their own promoters and encode two open reading frame proteins – ORF1 that is an RNA-binding protein and ORF2 with endonuclease and reverse transcriptase activities. They are the most abundant sequences from the LINE family, making up 18% of the human genome (Lander et al., 2001). The majority of LINE-1 sequences are transcriptionally inactive. The LINE-2 sequences in the human genome are highly defective and can encode either one or two ORF proteins (Darby and Sabunciyan, 2014).
The SINEs are active and non-autonomous TEs, represented by the Alu and the Mammalian wide Interspersed Repeat (MIR) elements (11 and 3% of the human genome, respectively). The Alu sequences were named based on sharing a common cleavage site for the AluI restriction enzyme (Houck et al., 1979). The Alu sequences are active but require the reverse transcriptase that is encoded by LINE-1 sequences (Pray, 2008a). In turn, the MIR elements are inactive. It has recently been shown that SINEs may form more complex sequences that are classified as the SVA retrotransposons. The SVA sequences have been formed by coupling the SINEs, a variable number of tandem repeats and the Alu retrotransposons. The SVAs also require the LINE-1 expression for mobilization. These sequences contribute to about 0.1% of the human genome and are the most active group of retrotransposons (Ostertag et al., 2003; Wang et al., 2005).
In turn, pseudogenes are DNA sequences that are related to real genes but they have lost at least some protein-coding abilities. It has been found that mRNA of pseudogenes can be reverse transcribed by the proteins encoded by LINE-1 sequences and transferred into other regions of the genome, creating processed pseudogenes. It has been estimated that the human genome consists of over 7,800 pseudogenes (Zhang et al., 2003). In case of integration close to active promoters, processed pseudogenes can be further transcribed. As listed by Kazazian (2014), they share the following characteristics: (1) their sequences are similar to the transcribed part of the parent gene; (2) they lack all or most introns; (3) they contain a poly-A tail attached to the 3′-most transcribed nucleotide; and (4) they are flanked at their 5′ and 3′ terminals by target site duplications of 5–20 nucleotides.
Finally, little is known about type II TEs (DNA transposons) that have never been active in the human genome. Type II TEs include the hAT, MuDR, piggyBac, and Tc1/mariner sequences (Munoz-Lopez and Garcia-Perez, 2010). These transposons do not act via RNA intermediates and encode enzymes that enable their mobilization. Due to their inactivity their causal role in the etiology of human diseases is less likely (Darby and Sabunciyan, 2014).
Insights Into Potential Mechanisms Underlying the Role of TEs in Mental Disorders
A recent review of human monogenic diseases that occur due to retrotransposition suggests that only the transposition of LINE-1, Alu, and SVA sequences might be deleterious, underlying the development of monogenic diseases (Kaer and Speek, 2013). Retrotransposition might affect various gene regions via altering their sequence or influencing expression activity. For instance, the Alu sequences contain several stop codons that may result in a truncated protein (Mighell et al., 1997). This mechanism has been discovered in patients with hemophilia B caused by transferring the Alu-Ya5 element into a protein coding region of the factor IX gene (Vidaud et al., 1993). In case of transposition into promoter regions, these sequences might impact gene expression. Another scenario originates from sequence homology that can promote homologous recombination, leading to insertions and deletions. Finally, the SVA tandems can mobilize exons, contributing to complex rearrangements.
However, the effects of alterations in DNA sequence triggered by retrotransposition have not been found to underlie the development of common mental disorders. In the majority of studies of patients with mental disorders (reviewed in detail below), altered expression and/or epigenetic regulation of retrotransposons have been reported. There are several epigenetic processes that act as defense mechanism against retrotransposition, including DNA methylation, histone modifications, small RNA-mediated regulation and post-transcriptional silencing by DICER and siRNA (Lapp and Hunter, 2016). Indeed, the majority of TEs in the human genome are hypermethylated (Pray, 2008b). Although DNA methylation acts as a defense mechanism, it cannot be excluded that hypermethylation of newly inserted TEs can lead to further changes in chromatin conformation, triggering changes in the expression of adjacent genes. It is most likely that retrotransposition occurs during early development when epigenetic marks are removed (Darby and Sabunciyan, 2014). There are also some well characterized histone modifications, including trimethylation of lysine 9 and lysine 27 at histone H3 (H3K9me3 and H3K27me3, respectively), which lead to heterochromatin formation and transcriptional silencing of TEs (Day et al., 2010; Baker et al., 2012).
It should be noted that only a small subset of TEs has been reported to be involved in retrotransposition. For instance, only 30–60 LINE-1 sequences in diploid cells are capable of retrotransposition (Sassaman et al., 1997). In addition, the majority of LINE-1 sequences are methylated to a certain degree. It has been found that LINE-1 methylation might impact gene expression via specific mechanisms [for review see (Kitkumthorn and Mutirangura, 2011)]. Firstly, LINE-1 sequences may produce unique RNA transcripts that act beyond the LINE-1 location. Alternatively, the reverse LINE-1 promoter can transcribe unique DNA sequences beyond the 5′ end of LINE-1. The second scenario is that intragenic LINE-1 RNAs decrease the expression of host gene via the nuclear RNA-induced silencing complexes. Indeed, it has been found that the Argonaute-2 (AGO2) protein targets intronic LINE-1 pre-mRNA complexes leading to down-regulation of gene expression in cancer cells (Aporntewan et al., 2011).
Global DNA hypomethylation that progresses with aging has been associated with genomic instability (Jung and Pfeifer, 2015). Hypomethylated genome regions are prone to accumulate various types of DNA lesions that include oxidative damage, depurination, depyrimidation and pathologic endogenous double-strand breaks (Mutirangura, 2019). The latter ones are now believed to act as intermediate products that drive genomic instability (Mutirangura, 2019). Accumulating evidence indicates that methylation of TEs might protect against genomic instability processes. For instance, it has been demonstrated that Alu siRNA increases Alu methylation levels, lowers endogenous DNA damage and increases DNA resistance to DNA damaging agents (Patchsung et al., 2018). Similarly, LINE-1 hypomethylation may contribute to genomic instability via interactions with the ATM gene expression (Kitkumthorn and Mutirangura, 2011). However, the contribution of a reduction in the Alu methylation to genomic instability might be greater than that of LINE-1 or HERV sequences (Jintaridth and Mutirangura, 2010).
It remains largely unknown how changes in the expression of TEs might contribute to the development of mental disorders. It has been hypothesized that the presence of TEs in the human genome provides immunity against several infectious agents. Indeed, the mechanisms that contributed to HERV insertions are analogous to those used for replication by exogenous retroviruses (Grandi and Tramontano, 2018). Therefore, changes in the expression of TEs, e.g., via epigenetic processes, might impact immune responses and make the host more liable to exogenous infections. There is evidence that HERV-derived peptides may interact with innate immunity via various mechanisms. For instance, HERV proteins are able to interact with pattern recognition receptors (PRRs) that play a pivotal role in antiviral responses (Hurst and Magiorkinis, 2015). Emerging evidence indicates that exogenous viruses, including herpesviruses and influenza virus, might modulate the expression of HERV sequences. This mechanism might play a protective role and has been reviewed in detail by Grandi and Tramontano (2018). In brief, HERV transcripts might interact with homologous RNA from exogenous retroviruses, leading to the formation of molecules that are recognized by PRRs, acting as innate immunity sensors. The similarity of HERV proteins to those exogenous retroviruses allow them to compete with cellular receptors. This similarity might also trigger complementation events that impair formation of viral particles after cellular infection. On the other site, HERV proteins may suppress innate immunity. It has been reported that HERV-K proteins inhibit the activation of T cells (Morozov et al., 2013) as well as reduce the levels of interleukin-6 and Toll-like Receptor 7 (Laska et al., 2017).
Transposable Elements and Their Epigenetic Regulation in Mental Disorders
As mentioned above, expression of TEs might play an important role in shaping immune responses against exogenous infections. Aberrant immune-inflammatory responses have been reported in several mental disorders. Also, a number of exogenous infections have been found to impact a risk of mental disorders. Below, we review studies investigating TEs and their epigenetic regulation in specific mental disorders, starting from the rationale of these studies that is based on the contribution of immune-inflammatory processes. A summary of human studies was provided in Table 1.
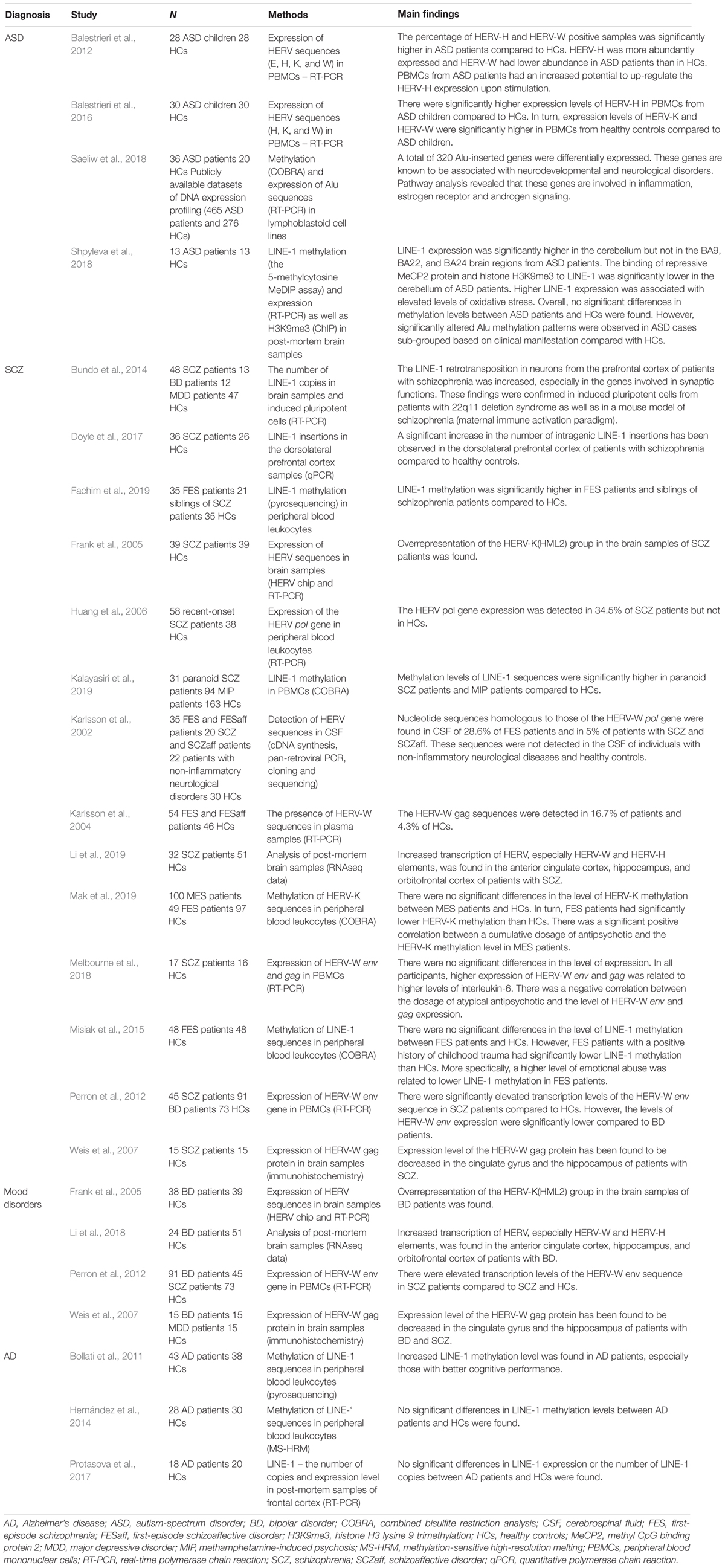
Table 1. Overview of human studies investigating the role of TEs in mental disorders.
Autism Spectrum Disorders (ASD)
Overexpression of HERV-H has been observed in peripheral blood mononuclear cells (PBMC) of children with ASD (Balestrieri et al., 2012, 2016). Similar findings have also been observed in two different ASD mouse models – inbred BTBR T+tf/J mice and CD-1 outbred mice prenatally exposed to valproic acid. In both of these mouse models, the expression of several endogenous retrovirus (ERV) families (ETnI, ETnII-α, ETnII-β, ETnII-γ, MusD, and IAP) was significantly higher in comparison with corresponding controls (Cipriani et al., 2018). Interestingly, the studies in mouse models provide additional information on the potential use of ERV sequences as biomarkers: (i) a higher expression of ERV was observed both in the peripheral blood mononuclear cells and the brain, suggesting that altered profile of peripheral ERV sequences may reflect similar alterations at the brain level; (ii) ERV overexpression in ASD mouse models is detectable from prenatal stage till the adulthood and (iii) ERV overexpression in ASD mouse models is also accompanied by increased expression of pro-inflammatory cytokines and Toll-like receptors. Furthermore, a subsequent study in one of the models (mice prenatally exposed to valproic acid) provided evidence that higher levels of ERVs are also detectable in the offspring (second and third generations) of those mice exposed prenatally to valproic acid (Tartaglione et al., 2018).
Also LINE-1 retrotransposons have been associated with ASD (Shpyleva et al., 2018; Suarez et al., 2018). The levels of LINE-1 ORF1 and ORF2 transcripts have been investigated in four brain regions of patients with idiopathic autism (the frontal cortex, anterior cingulate, auditory cortex, and cerebellum). Elevated LINE-1 expression together with lower binding affinity of repressive MeCP2 protein and histone H3K9me3 to LINE-1 sequences was observed only in the cerebellum, suggesting a lessening of epigenetic repression and consequently an increase in chromatin accessibility. Interestingly, the increase in LINE-1 expression was also inversely correlated with glutathione redox status, consistent with reports indicating that LINE-1 expression is increased under pro-oxidant conditions (Shpyleva et al., 2018). The overexpression of LINE-1 within a single brain region is suggestive of a mosaicism-like impact of retrotransposons and definitively needs further investigation. In partial agreement with the findings of increased LINE-1 expression in ASD, data concerning LINE-1 methylation status in lymphoblastoid peripheral cells have provided evidence of reduced methylation in a subgroup of patients with severe language impairment (Tangsuwansri et al., 2018).
It has also been shown that the Alu sequence, the most abundant of all TEs in the human genome, deserves further research in ASD (Saeliw et al., 2018). Indeed, this study investigated the Alu methylation and expression in lymphoblastoid peripheral cells from ASD patients. Although the global methylation of Alu subfamilies was not significantly different between ASD and control group, when ASD samples were divided according to phenotypic subgroups, methylation patterns of the AluS subfamily were different from those in relative controls in two of the ASD subgroups, and within one of the subgroup (mild phenotype), the Alu expression was correlated with methylation status. Despite the limited sample size (particularly of subgroups), these data suggest that classification of ASD patients in phenotypic subgroups may represent a useful tool in investigating associations of TEs with the highly heterogeneous ASD diagnostic construct.
Schizophrenia-Spectrum Disorders
It has been clearly demonstrated that winter-spring seasonality of birth as well as prenatal and postnatal infections increase a risk of developing schizophrenia (McGrath and Welham, 1999; Davies et al., 2003; Khandaker et al., 2013). Moreover, the largest genome-wide association study revealed that variation within the HLA genes is strongly associated with schizophrenia susceptibility (Ripke et al., 2014). Finally, schizophrenia patients present with several indices of subclinical inflammation in terms of pro-inflammatory cytokine profiles (Miller et al., 2011; Frydecka et al., 2018), alterations of lymphocyte counts (Miller et al., 2013; Karpiǹski et al., 2016, 2018) and elevated levels of C-reactive protein (CRP) (Fernandes et al., 2016). On the basis of a meta-analysis, Arias et al. (2012) found a higher prevalence of infections with several agents, including Human Herpesvirus (HHV) 2, Borna Disease Virus, Chlamydia pneumoniae, Chlamydia psittaci, and Toxoplasma gondii in patients with schizophrenia compared to healthy controls.
Accumulating evidence indicates altered expression of HERV sequences in patients with schizophrenia. Karlsson et al. (2002) found nucleotide sequences homologous to those of the HERV-W pol gene in the cerebrospinal fluid (CSF) of 28.6% of first-episode schizophrenia patients and in 5% of patients with chronic schizophrenia. These sequences were not detected in the CSF of individuals with non-inflammatory neurological diseases and healthy controls. Increased levels of HERV-W-related gag and pol transcripts and a higher prevalence of the gag and pol antigenemia in peripheral blood from patients with schizophrenia compared to healthy controls have been reported by several studies (Karlsson et al., 2004; Huang et al., 2006; Perron et al., 2008; Yao et al., 2008). The study by Perron et al. (2008) also revealed significantly higher rates of positive HERV-W env antigenemia in patients with schizophrenia than in healthy controls. The HERV-W gag and env antigenemia has been also associated with subclinical inflammation in terms of elevated levels of CRP and pro-inflammatory cytokines (Perron et al., 2008; Melbourne et al., 2018). Interestingly, Huang et al. (2011) found that overexpression of the HERV-W env in the human U251 glioma cells up-regulated a number of schizophrenia-associated genes, including those that encode brain-derived neurotrophic factor, neurotrophic tyrosine kinase receptor type 2 and the dopamine D3 receptor as well as increased the phosphorylation of cyclic adenosine monophosphate response element-binding protein. In this study, mRNA of the HERV-W env gene was detected in plasma from 42 out of 118 recent-onset schizophrenia patients but not in healthy controls. There is also evidence that the expression of HERV-W env induces calcium influx and down-regulates the DISC1 gene expression in the human neuroblastoma cells (Chen et al., 2018). Interestingly, expression level of the HERV-W gag protein has been found to be decreased in the cingulate gyrus and the hippocampus of patients with schizophrenia (Weis et al., 2007). However, a recent analysis of RNA-seq data in the human post mortem brain samples revealed increased transcription of HERV, especially HERV-W and HERV-H elements, in the anterior cingulate cortex, hippocampus and orbitofrontal cortex of patients with schizophrenia and bipolar disorder (Li et al., 2019). Interestingly, the HERV sequences within the ERVWE1 gene (7q21.2) exhibited the highest levels of transcription across all brain regions examined in this analysis. The env gene in this locus encodes syncytin-1, expressed at high levels in the human placenta (Blond et al., 2000; Mi et al., 2000). However, altered expression of this gene has been reported in the areas of active demyelination in patients with multiple sclerosis (Mameli et al., 2007). At this point, it should be noted that myelin alterations are widely observed in patients with schizophrenia (Mighdoll et al., 2015). Although initial results regarding expression of the HERV-W sequences in schizophrenia patients are promising, caution should be taken on the way these results are being interpreted. Indeed, the majority of studies in this field analyzed the overall expression of HERV-W sequences without investigating specific HERV-W loci. Moreover, no conclusive association between the HERV-W expression and other human pathologies has been documented so far [for review see (Grandi and Tramontano, 2017)].
Less is known about other families of HERVs in patients with schizophrenia. Frank et al. (2005) found overrepresentation of the HERV-K(HML2) group in brain samples of patients with schizophrenia and bipolar disorder. Our group also tested peripheral blood methylation levels of HERV-K sequences in first-episode and multi-episode schizophrenia patients (Mak et al., 2019). We found significantly lower levels of HERV-K methylation in first-episode schizophrenia patients compared to healthy controls. These alterations were not observed in multi-episode schizophrenia patients. Moreover, we did not find an association between HERV-K methylation levels and the deficit schizophrenia subtype that refers to a subgroup of patients with enduring and persistent negative symptoms. However, we found a significant positive correlation between the dosage of antipsychotics and HERV-K methylation levels in multi-episode schizophrenia patients. These findings imply that the HERV-K methylation might normalize in the course of schizophrenia. It is also likely that antipsychotic drugs might impact methylation and expression of HERV-K sequences. In contrast to our findings, Diem et al. (2012) found no significant effects of valproic acid, haloperidol, risperidone and clozapine on the HERV-K expression levels in the human brain cell lines. However, valproic acid was found to strongly up-regulate expression of HERV-W and ERV9 elements.
Some studies also investigated methylation status and expression levels of non-LTR sequences in patients with schizophrenia. Bundo et al. (2014) demonstrated increased LINE-1 retrotransposition in neurons from the prefrontal cortex of patients with schizophrenia, especially in the genes involved in synaptic functions. These findings were confirmed in induced pluripotent cells from patients with 22q11 deletion syndrome as well as in a mouse model of schizophrenia (maternal immune activation paradigm). In agreement with these results, a significant increase in the number of intragenic LINE-1 insertions has been observed in the dorsolateral prefrontal cortex of patients with schizophrenia compared to healthy controls (Doyle et al., 2017). Over-representation of these insertions appeared within the gene ontologies called “cell projection” and “postsynaptic membrane,” suggesting their role in the brain development. In some studies, LINE-1 methylation was tested in peripheral blood leukocytes of patients with schizophrenia, providing mixed findings (Misiak et al., 2015; Li et al., 2018; Fachim et al., 2019; Kalayasiri et al., 2019). The study by our group revealed lower LINE-1 methylation only in patients with first-episode schizophrenia and a positive history of childhood trauma. Among various childhood adversities, emotional trauma was most strongly associated with the LINE-1 methylation status. These results are in agreement with a previous study, showing that the LINE-1 methylation might be involved in resilience and susceptibility to develop post-traumatic stress disorder (Rusiecki et al., 2012). Moreover, increased expression of LINE-1 in response to stress has been reported in various cell lines (Li and Schmid, 2001; Capomaccio et al., 2010). Lower LINE-1 methylation levels in patients with schizophrenia and bipolar disorder were also reported by Li et al. (2018). Other studies revealed hypermethylation of LINE-1 sequences in patients with first-episode psychosis, paranoid schizophrenia and methamphetamine-induced paranoia (Fachim et al., 2019; Kalayasiri et al., 2019).
Mood Disorders
A recent systematic review indicates that prenatal infections might impact the risk of bipolar disorder (Marangoni et al., 2016). However, this observation is based on a lower number of studies compared to studies addressing the impact of prenatal infections on schizophrenia risk. There is evidence that influenza infection during pregnancy is associated with a fourfold increase in the risk of bipolar disorder in the offspring (Parboosing et al., 2013). Another study demonstrated that prenatal flu exposure increases the risk of bipolar disorder with psychotic features (Canetta et al., 2014). However, no association was found between prenatal infections with HHV-1, HHV-2, Cytomegalovirus or Toxoplasma gondii and bipolar disorder risk (Mortensen et al., 2011). Maternal infections in the second trimester might also contribute to the development of depressive symptoms in the adolescent offspring (Murphy et al., 2017). However, the impact of specific infectious agents has not been tested so far.
Although all major mental disorders are characterized by co-existing subclinical inflammation, some differences, regarding specific pro-inflammatory markers can be indicated (Goldsmith et al., 2016; Misiak et al., 2019). Therefore, it might be hypothesized that the mechanisms leading to subclinical inflammation in bipolar disorder, major depression and schizophrenia-spectrum disorders are different. However, studies investigating expression of TEs do not support this hypothesis. For instance, over-expression of HERV-K sequences has been reported in brain samples of patients with bipolar disorder and schizophrenia (Frank et al., 2005). Similarly, decreased expression of the HERV-W gag protein has been reported in the cingulate gyrus and hippocampus of patients with schizophrenia, bipolar disorder, and major depression (Weis et al., 2007). Finally, hypomethylation of LINE-1 elements in peripheral blood has been observed in patients with bipolar disorder and schizophrenia (Li et al., 2018). Some differences have been detected with respect to the expression of HERV-W sequences. Indeed, Perron et al. (2012) found elevated transcription levels of the HERV-W env sequence in the peripheral blood of patients with bipolar disorder and schizophrenia compared to healthy controls. Expression levels of the HERV-W env sequence were also significantly higher in patients with bipolar disorder than in those with schizophrenia.
Alzheimer’s Disease
There is a general consensus that aging processes are associated with progressive loss of global DNA methylation and site-specific DNA hypermethylation (Jung and Pfeifer, 2015). Similarly, TEs are subjected to profound epigenetic modifications during aging that appear in the context of organismal and cellular senescence (Cardelli, 2018). For instance, age-related loss of Alu and HERV-K methylation has been well-documented (Bollati et al., 2009; Jintaridth and Mutirangura, 2010; Gentilini et al., 2013). Moreover, it has been found that the expression of HERV-H, HERV-K and HERV-W changes during the lifespan with distinct patterns (Balestrieri et al., 2015). Importantly, the study by Gentilini et al. (2013) demonstrated that age-related loss of Alu methylation was less apparent in the offspring of centenarians, suggesting the effects of genetic factors associated with longevity. In turn, studies investigating changes of LINE-1 methylation have provided mixed findings (Bollati et al., 2009; Talens et al., 2012; Cho et al., 2015). Finally, there is evidence that chromatin of Alu, SVA, and LINE-1 becomes relatively more open in senescent cells (De Cecco et al., 2013).
Age-related changes in epigenetic modifications of TEs have provided basis for investigating alterations of these processes in Alzheimer’s disease. In a single study of early-onset Alzheimer’s disease family, it has been reported that large genomic rearrangements might affect the presenilin-1 gene via the mechanisms involving recombination stimulated by the Alu sequence (Hiltunen et al., 2000). However, subsequent studies have not provided compelling evidence regarding the contribution of TEs to the etiology of Alzheimer’s disease. There is only one study, showing increased LINE-1 methylation in peripheral blood leukocytes of patients with Alzheimer’s disease, especially those with better cognitive performance, compared to healthy controls (Bollati et al., 2011). However, the authors did not find significant between-group differences in the levels of Alu and SAT-α methylation. Other studies did not confirm these findings regarding LINE-1 methylation (Hernández et al., 2014; Protasova et al., 2017). Alterations of other TEs in patients with Alzheimer’s disease have not been tested so far.
Summary of Evidence and Future Directions
Although specific retrotransposition events that may account for mental disorders in the manner observed in case of Mendelian diseases have not been identified so far, accumulating evidence indicates the involvement of altered expression and epigenetic regulation of TEs in the pathophysiology of schizophrenia, mood disorders and ASD. Most consistently, previous studies indicate altered expression of HERVs and methylation of LINE-1 sequences. However, specific findings are similar in patients with various mental disorders and thus their use as biomarkers is largely limited. Moreover, the direction of causality is yet to be determined. For instance, it cannot be excluded that altered expression of HERV appears as a consequence of other epigenetic dysregulations that are widely observed in mental disorders. Additionally, severe mental disorders, including schizophrenia and mood disorders, are associated with high prevalence rates of somatic comorbidities, including autoimmune diseases, type 2 diabetes and cardiovascular diseases that have also been associated with altered epigenetic regulation of TEs (Cash et al., 2011; De Hert et al., 2011; Misiak et al., 2013; Nestler et al., 2016; Zhao et al., 2018). Interestingly, there are studies showing that the expression of various HERV sequences appears in a certain subgroup of patients with schizophrenia but not in healthy controls. These findings are consistent with previous studies, showing that immune alterations can be observed only in a subgroup of patients characterized by poor response to treatment and support the concept of psychosis subtypes (Frydecka et al., 2015; Mondelli et al., 2015; Fillman et al., 2016). Other clinical correlates of subclinical inflammation in schizophrenia include, i.e., more severe cognitive deficits (Misiak et al., 2017b), persistent negative symptoms (Goldsmith et al., 2018) and certain neurostructural abnormalities (Najjar and Pearlman, 2015). However, so far studies investigating expression and epigenetic regulation of TEs in schizophrenia have been based on relatively small samples without comprehensive clinical assessment. Similarly, studies investigating the expression of TEs in patients with bipolar disorder did not control for mood status and a severity of psychopathological symptoms.
Another important point is that causal inferences between TEs and mental disorders cannot be established. Firstly, it remains unknown what are the critical periods when alterations in epigenetic regulation and expression of TEs appear. Therefore, future studies should examine epigenetic processes that regulate expression of TEs in patients at early stages of mental disorders or individuals from clinical high risk groups. This is particularly important since several lifestyle characteristics that are highly prevalent among patients with mental disorders, e.g., cigarette smoking and poor dietary habits, might impact TEs per se (Miglino et al., 2014; Miousse et al., 2015). Secondly, the role of HERVs in shaping innate immunity also remains problematic with respect to understanding causal associations. On one side, expression of HERVs might condition resistance to exogenous infections; on the other, exogenous retroviruses have been found to impact the expression of HERVs. Therefore, it remains unknown whether altered expression profiles of HERVs in mental disorders represent cause or consequence of exogenous infections. Future studies should necessarily examine the biological nature and the extent of associations between immune alterations in mental disorders and expression of various TEs.
Finally, more global concordance patterns of different TEs expression in mental disorders are yet to be examined: this could provide further insight into specificity of methylation patterns across different TEs and provide additional information of their use as potential biomarkers. At this point, it is important to note that similar DNA methylation patterns have been described in brain samples and peripheral blood leukocytes of patients with schizophrenia (Van Den Oord et al., 2016).
Another direction for the field is to disentangle the effects of stressful life events on epigenetic regulation of TEs expression. Early-life stress is a known risk factor for mood and psychotic disorders as well as correlates with a number of biological dysregulations in adults (Misiak et al., 2017a; Bielawski et al., 2019; Jaworska-Andryszewska and Rybakowski, 2019). Acute stress has been found to increase the levels of H3K9me3 as well as decrease the levels of H3K9me1 and H3K27me3 in the dentate gyrus and the CA1 layer of the hippocampus in rats (Milne et al., 2009). In turn, chronic restraint stress for 21 days mildly increased the levels of H3Kme4 and reduced the levels of H3K9me3 in the dentate gyrus. Treatment with fluoxetine reversed changes in the levels of H3K9me3 during chronic restraint stress. More specifically, the same group found that acute stress had increased H3K9me3 enrichment at SINEs (Baker et al., 2012). In turn, our group found lower methylation of LINE-1 sequences in peripheral blood leukocytes of patients with first-episode schizophrenia reporting a positive history of childhood trauma (Misiak et al., 2015). In light of these findings, future studies should further examine the effects of stress on the expression of TEs in patients from various clinical groups and preclinical studies could contribute to this aim.
Author Contributions
BM and MS conceived the concept of this article. BM wrote the Sections “Introduction”, “Insights Into Potential Mechanisms Underlying the Role of TEs in Mental Disorders”, “Schizophrenia-Spectrum Disorders” and “Mood Disorders”. LR wrote the Section “Autism Spectrum Disorders (ASD)”. MS prepared the Sections “Brief Overview of TEs in the Human Genome – Classification and Nomenclature” and “Alzheimer’s Disease”. BM, LR, and MS prepared the Section “Summary of Evidence and Future Directions”. All authors contributed to the manuscript revision, read, and approved the submitted version.
Funding
This work was supported by the statutory project funded by the Wrocław Medical University, Wrocław, Poland (Task Number: ST-A.290.17.032).
Conflict of Interest Statement
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
References
Aporntewan, C., Phokaew, C., Piriyapongsa, J., Ngamphiw, C., Ittiwut, C., Tongsima, S., et al. (2011). Hypomethylation of intragenic LINE-1 represses transcription in cancer cells through AGO2. PLoS One 6:e17934. doi: 10.1371/journal.pone.0017934
Arias, I., Sorlozano, A., Villegas, E., Luna, J., de, D., McKenney, K., et al. (2012). Infectious agents associated with schizophrenia: a meta-analysis. Schizophr. Res. 136, 128–136. doi: 10.1016/j.schres.2011.10.026
Baillie, J. K., Barnett, M. W., Upton, K. R., Gerhardt, D. J., Richmond, T. A., De Sapio, F., et al. (2011). Somatic retrotransposition alters the genetic landscape of the human brain. Nature 479, 534–537. doi: 10.1038/nature10531
Baker, M. E. R., Pfaff, D. W., McEwen, B. S., Seligsohn, M., Hunter, R. G., Datson, N. A., et al. (2012). Acute stress and hippocampal histone H3 lysine 9 trimethylation, a retrotransposon silencing response. Proc. Natl. Acad. Sci. U.S.A. 109, 17657–17662. doi: 10.1073/pnas.1215810109
Balestrieri, E., Arpino, C., Matteucci, C., Sorrentino, R., Pica, F., Alessandrelli, R., et al. (2012). HERVs expression in autism spectrum disorders. PLoS One 7:e48831. doi: 10.1371/journal.pone.0048831
Balestrieri, E., Cipriani, C., Matteucci, C., Capodicasa, N., Pilika, A., Korea, I., et al. (2016). Transcriptional activity of human endogenous retrovirus in Albanian children with autism spectrum disorders. New Microbiol. 39, 228–231.
Balestrieri, E., Pica, F., Matteucci, C., Zenobi, R., Sorrentino, R., Argaw-Denboba, A., et al. (2015). Transcriptional activity of human endogenous retroviruses in human peripheral blood mononuclear cells. Biomed Res. Int. 2015:164529. doi: 10.1155/2015/164529
Bannert, N., and Kurth, R. (2006). The evolutionary dynamics of human endogenous retroviral families. Annu. Rev. Genomics Hum. Genet. 7, 149–173. doi: 10.1146/annurev.genom.7.080505.115700
Bielawski, T., Misiak, B., Moustafa, A., and Frydecka, D. (2019). Epigenetic mechanisms, trauma, and psychopathology: targeting chromatin remodeling complexes. Rev. Neurosci. doi: 10.1515/revneuro-2018-0055 [Epub ahead of print].
Blond, J. L., Lavillette, D., Cheynet, V., Bouton, O., Oriol, G., Chapel-Fernandes, S., et al. (2000). An envelope glycoprotein of the human endogenous retrovirus HERV-W is expressed in the human placenta and fuses cells expressing the type D mammalian retrovirus receptor. J. Virol. 74, 3321–3329. doi: 10.1128/jvi.74.7.3321-3329.2000
Bollati, V., Galimberti, D., Pergoli, L., Dalla Valle, E., Barretta, F., Cortini, F., et al. (2011). DNA methylation in repetitive elements and Alzheimer disease. Brain Behav. Immun. 25, 1078–1083. doi: 10.1016/j.bbi.2011.01.017
Bollati, V., Schwartz, J., Wright, R., Litonjua, A., Tarantini, L., Suh, H., et al. (2009). Decline in genomic DNA methylation through aging in a cohort of elderly subjects. Mech. Ageing Dev. 130, 234–239. doi: 10.1016/j.mad.2008.12.003
Bundo, M., Toyoshima, M., Okada, Y., Akamatsu, W., Ueda, J., Nemoto-Miyauchi, T., et al. (2014). Increased L1 retrotransposition in the neuronal genome in schizophrenia. Neuron 81, 306–313. doi: 10.1016/j.neuron.2013.10.053
Callinan, P. A., and Batzer, M. A. (2006). Retrotransposable elements and human disease. Genome Dis. 1, 104–115. doi: 10.1159/000092503
Canetta, S. E., Bao, Y., Co, M. D. T., Ennis, F. A., Cruz, J., Terajima, M., et al. (2014). Serological documentation of maternal influenza exposure and bipolar disorder in adult offspring. Am. J. Psychiatry 171, 557–563. doi: 10.1176/appi.ajp.2013.13070943
Capomaccio, S., Verini-Supplizi, A., Galla, G., Vitulo, N., Barcaccia, G., Felicetti, M., et al. (2010). Transcription of LINE-derived sequences in exercise-induced stress in horses. Anim. Genet. 41(Suppl. 2), 23–27. doi: 10.1111/j.1365-2052.2010.02094.x
Cardelli, M. (2018). The epigenetic alterations of endogenous retroelements in aging. Mech. Ageing Dev. 174, 30–46. doi: 10.1016/j.mad.2018.02.002
Cardno, A. G., and Gottesman, I. I. (2000). Twin studies of schizophrenia: from bow-and-arrow concordances to star wars Mx and functional genomics. Am. J. Med. Genet. Semin. Med. Genet. 97, 12–17. doi: 10.1002/(sici)1096-8628(200021)97:1<12::aid-ajmg3>3.3.co;2-l
Cash, H. L., Mcgarvey, S. T., Houseman, E. A., Marsit, C. J., Hawley, N. L., Lambert-Messerlian, G. M., et al. (2011). Cardiovascular disease risk factors and DNA methylation at the LINE-1 repeat region in peripheral blood from Samoan Islanders. Epigenetics 6, 1257–1264. doi: 10.4161/epi.6.10.17728
Chen, Y., Yan, Q., Zhou, P., Li, S., and Zhu, F. (2018). HERV-W env regulates calcium influx via activating TRPC3 channel together with depressing DISC1 in human neuroblastoma cells. J. Neurovirol. 25, 101–113. doi: 10.1007/s13365-018-0692-7
Cho, Y. H., Woo, H. D., Jang, Y., Porter, V., Christensen, S., Hamilton, R. F., et al. (2015). The association of LINE-1 hypomethylation with age and centromere positive micronuclei in human lymphocytes. PLoS One 10:e0133909. doi: 10.1371/journal.pone.0133909
Cipriani, C., Ricceri, L., Matteucci, C., De Felice, A., Tartaglione, A. M., Argaw-Denboba, A., et al. (2018). High expression of Endogenous Retroviruses from intrauterine life to adulthood in two mouse models of Autism Spectrum Disorders. Sci. Rep. 8:629. doi: 10.1038/s41598-017-19035-w
Cordaux, R., and Batzer, M. A. (2009). The impact of retrotransposons on human genome evolution. Nat. Rev. Genet. 10, 691–703. doi: 10.1038/nrg2640
Coufal, N. G., Garcia-perez, J. L., Peng, G. E., Yeo, G. W., Mu, Y., Lovci, M. T., et al. (2009). L1 retrotransposition in human neural progenitor cells Nicole. Nature 460, 1127–1131. doi: 10.1038/nature08248.L1
Darby, M. M., and Sabunciyan, S. (2014). Repetitive elements and epigenetic marks in behavior and psychiatric disease. Adv. Genet. 86, 185–252. doi: 10.1016/B978-0-12-800222-3.00009-7
Davies, G., Welham, J., Chant, D., Torrey, E. F., and McGrath, J. (2003). A systematic review and meta-analysis of Northern Hemisphere season of birth studies in schizophrenia. Schizophr. Bull. 29, 587–593. doi: 10.1093/oxfordjournals.schbul.a007030
Day, D. S., Luquette, L. J., Park, P. J., and Kharchenko, P. V. (2010). Estimating enrichment of repetitive elements from high-throughput sequence data. Genome Biol. 11:R69. doi: 10.1186/gb-2010-11-6-r69
De Cecco, M., Criscione, S. W., Peckham, E. J., Hillenmeyer, S., Hamm, E. A., Manivannan, J., et al. (2013). Genomes of replicatively senescent cells undergo global epigenetic changes leading to gene silencing and activation of transposable elements. Aging Cell 12, 247–256. doi: 10.1111/acel.12047
De Hert, M., Correll, C. U., Bobes, J., Cetkovich-Bakmas, M., Cohen, D. A. N., Asai, I., et al. (2011). Physical illness in patients with severe mental disorders. I. Prevalence, impact of medications and disparities in health care. World Psychiatry 10, 52–77. doi: 10.1002/j.2051-5545.2011.tb00014.x
Dewannieux, M., and Heidmann, T. (2005). LINEs, SINEs and processed pseudogenes: parasitic strategies for genome modeling. Cytogenet. Genome Res. 110, 35–48. doi: 10.1159/000084936
Diem, O., Schäffner, M., Seifarth, W., and Leib-Mösch, C. (2012). Influence of antipsychotic drugs on human endogenous retrovirus (HERV) transcription in brain cells. PLoS One 7:e30054. doi: 10.1371/journal.pone.0030054
Doyle, G. A., Crist, R. C., Karatas, E. T., Hammond, M. J., Ewing, A. D., Ferraro, T. N., et al. (2017). Analysis of LINE-1 elements in DNA from postmortem brains of individuals with schizophrenia. Neuropsychopharmacology 42, 2602–2611. doi: 10.1038/npp.2017.115
Fachim, H., Loureiro, C., Corsi-Zuelli, F., Shuhama, R., Louzada-Junior, P., Menezes, P., et al. (2019). GRIN2B promoter methylation deficits in early-onset schizophrenia and its association with cognitive function. Epigenomics 11, 401–410. doi: 10.2217/epi-2018-0127
Fernandes, B. S., Steiner, J., Bernstein, H. G., Dodd, S., Pasco, J. A., Dean, O. M., et al. (2016). C-reactive protein is increased in schizophrenia but is not altered by antipsychotics: meta-analysis and implications. Mol. Psychiatry 21, 554–564. doi: 10.1038/mp.2015.87
Fillman, S. G., Weickert, T. W., Lenroot, R. K., Catts, S. V., Bruggemann, J. M., Catts, V. S., et al. (2016). Elevated peripheral cytokines characterize a subgroup of people with schizophrenia displaying poor verbal fluency and reduced Broca’s area volume. Mol. Psychiatry 21, 1090–1098. doi: 10.1038/mp.2015.90
Frank, O., Giehl, M., Hehlmann, R., Zheng, C., Leib-Mosch, C., and Seifarth, W. (2005). Human endogenous retrovirus expression profiles in samples from brains of patients with schizophrenia and bipolar disorders. J. Virol. 79, 10890–10901. doi: 10.1128/jvi.79.17.10890-10901.2005
Frydecka, D., Krzystek-Korpacka, M., Lubeiro, A., Stramecki, F., Staǹczykiewicz, B., Aleksander Beszłej, J., et al. (2018). Profiling inflammatory signatures of schizophrenia: a cross-sectional and meta-analysis study. Brain Behav. Immun. 71, 28–36. doi: 10.1016/j.bbi.2018.05.002
Frydecka, D., Misiak, B., Pawlak-Adamska, E., Karabon, L., Tomkiewicz, A., Sedlaczek, P., et al. (2015). Interleukin-6: the missing element of the neurocognitive deterioration in schizophrenia? The focus on genetic underpinnings, cognitive impairment and clinical manifestation. Eur. Arch. Psychiatry Clin. Neurosci. 265, 449–459. doi: 10.1007/s00406-014-0533-5
Gentilini, D., Mari, D., Castaldi, D., Remondini, D., Ogliari, G., Ostan, R., et al. (2013). Role of epigenetics in human aging and longevity: genome-wide DNA methylation profile in centenarians and centenarians’ offspring. Age 35, 1961–1973. doi: 10.1007/s11357-012-9463-1
Goldsmith, D. R., Haroon, E., Miller, A. H., Strauss, G. P., Buckley, P. F., and Miller, B. J. (2018). TNF-α and IL-6 are associated with the deficit syndrome and negative symptoms in patients with chronic schizophrenia. Schizophr. Res. 199, 281–284. doi: 10.1016/j.schres.2018.02.048
Goldsmith, D. R., Rapaport, M. H., and Miller, B. J. (2016). A meta-analysis of blood cytokine network alterations in psychiatric patients: comparisons between schizophrenia, bipolar disorder and depression. Mol. Psychiatry 199, 281–284.
Grandi, N., and Tramontano, E. (2017). Type W human endogenous retrovirus (HERV-W) integrations and their mobilization by L1 machinery: contribution to the human transcriptome and impact on the host physiopathology. Viruses 9:E162. doi: 10.3390/v9070162
Grandi, N., and Tramontano, E. (2018). Human endogenous retroviruses are ancient acquired elements still shaping innate immune responses. Front. Immunol. 9:2039. doi: 10.3389/fimmu.2018.02039
Griffiths, D. J. (2001). Endogenous retroviruses in the human genome sequence. Genome Biol. 2, reviews1017.1–reviews1017.5.
Hernández, H. G., Mahecha, M. F., Mejía, A., Arboleda, H., and Forero, D. A. (2014). Global long interspersed nuclear element 1 DNA methylation in a Colombian sample of patients with late-onset Alzheimer’s disease. Am. J. Alzheimers Dis. Other Demen. 29, 50–53. doi: 10.1177/1533317513505132
Hiltunen, M., Helisalmi, S., Mannermaa, A., Alafuzoff, I., Koivisto, A. M., Lehtovirta, M., et al. (2000). Identification of a novel 4.6-kb genomic deletion in presenilin-1 gene which results in exclusion of exon 9 in a Finnish early onset Alzheimer’s disease family: an Alu core sequence-stimulated recombination? Eur. J. Hum. Genet. 8, 259–266. doi: 10.1038/sj.ejhg.5200423
Houck, C. M., Rinehart, F. P., and Schmid, C. W. (1979). A ubiquitous family of repeated DNA sequences in the human genome. J. Mol. Biol. 132, 289–306. doi: 10.1016/0022-2836(79)90261-4
Huang, W., Li, S., Hu, Y., Yu, H., Luo, F., Zhang, Q., et al. (2011). Implication of the env gene of the human endogenous retrovirus W family in the expression of BDNF and DRD3 and development of recent-onset schizophrenia. Schizophr. Bull. 37, 988–1000. doi: 10.1093/schbul/sbp166
Huang, W. J., Liu, Z. C., Wei, W., Wang, G. H., Wu, J. G., and Zhu, F. (2006). Human endogenous retroviral pol RNA and protein detected and identified in the blood of individuals with schizophrenia. Schizophr. Res. 83, 193–199. doi: 10.1016/j.schres.2006.01.007
Hurst, T. P., and Magiorkinis, G. (2015). Activation of the innate immune response by endogenous retroviruses. J. Gen. Virol. 96, 1207–1218. doi: 10.1099/jgv.0.000017
Jaworska-Andryszewska, P., and Rybakowski, J. K. (2019). Childhood trauma in mood disorders: neurobiological mechanisms and implications for treatment. Pharmacol. Rep. 71, 112–120. doi: 10.1016/j.pharep.2018.10.004
Jintaridth, P., and Mutirangura, A. (2010). Distinctive patterns of age-dependent hypomethylation in interspersed repetitive sequences. Physiol. Genomics 41, 194–200. doi: 10.1152/physiolgenomics.00146.2009
Jung, M., and Pfeifer, G. P. (2015). Aging and DNA methylation. BMC Biol. 13:7. doi: 10.1186/s12915-015-0118-4
Kaer, K., and Speek, M. (2013). Retroelements in human disease. Gene 518, 231–241. doi: 10.1016/j.gene.2013.01.008
Kalayasiri, R., Kraijak, K., Mutirangura, A., and Maes, M. (2019). Paranoid schizophrenia and methamphetamine-induced paranoia are both characterized by a similar LINE-1 partial methylation profile, which is more pronounced in paranoid schizophrenia. Schizophr. Res. 208, 221–227. doi: 10.1016/j.schres.2019.02.015
Karlsson, H., McArthur, J., Schroder, J., Torrey, E. F., Bachmann, S., and Yolken, R. H. (2002). Retroviral RNA identified in the cerebrospinal fluids and brains of individuals with schizophrenia. Proc. Natl. Acad. Sci. U.S.A. 98, 4634–4639. doi: 10.1073/pnas.061021998
Karlsson, H., Schröder, J., Bachmann, S., Bottmer, C., and Yolken, R. H. (2004). HERV-W-related RNA detected in plasma from individuals with recent-onset schizophrenia or schizoaffective disorder. Mol. Psychiatry 9, 12–13. doi: 10.1038/sj.mp.4001439
Karpiǹski, P., Frydecka, D., Sasiadek, M. M., and Misiak, B. (2016). Reduced number of peripheral natural killer cells in schizophrenia but not in bipolar disorder. Brain Behav. Immun. 54, 194–200. doi: 10.1016/j.bbi.2016.02.005
Karpiǹski, P., Samochowiec, J., Frydecka, D., Sąsiadek, M. M., and Misiak, B. (2018). Further evidence for depletion of peripheral blood natural killer cells in patients with schizophrenia: a computational deconvolution study. Schizophr. Res. 54, 194–200. doi: 10.1016/j.schres.2018.04.026
Kazazian, H. H. (2014). Processed pseudogene insertions in somatic cells. Mob. DNA 5:20. doi: 10.1186/1759-8753-5-20
Khandaker, G. M., Zimbron, J., Lewis, G., and Jones, P. B. (2013). Prenatal maternal infection, neurodevelopment and adult schizophrenia: a systematic review of population-based studies. Psychol. Med. 43, 239–257. doi: 10.1017/S0033291712000736
Kitkumthorn, N., and Mutirangura, A. (2011). Long interspersed nuclear element-1 hypomethylation in cancer: biology and clinical applications. Clin. Epigenetics 2, 315–330. doi: 10.1007/s13148-011-0032-8
Lander, E. S., Linton, L. M., Birren, B., Nusbaum, C., Zody, M. C., Baldwin, J., et al. (2001). Initial sequencing and analysis of the human genome. Nature 409, 860–921. doi: 10.1038/35057062
Lapp, H. E., and Hunter, R. G. (2016). The dynamic genome: transposons and environmental adaptation in the nervous system. Epigenomics 8, 237–249. doi: 10.2217/epi.15.107
Laska, M. J., Troldborg, A., Hauge, E. M., Bahrami, S., and Stengaard-Pedersen, K. (2017). Human endogenous retroviral genetic element with immunosuppressive activity in both human autoimmune diseases and experimental arthritis. Arthritis Rheumatol. 69, 398–409. doi: 10.1002/art.39867
Li, F., Sabunciyan, S., Yolken, R. H., Lee, D., Kim, S., and Karlsson, H. (2019). Transcription of human endogenous retroviruses in human brain by RNA-seq analysis. PLoS One 14:e0207353. doi: 10.1371/journal.pone.0207353
Li, S., Yang, Q., Hou, Y., Jiang, T., Zong, L., Wang, Z., et al. (2018). Hypomethylation of LINE-1 elements in schizophrenia and bipolar disorder. J. Psychiatr. Res. 107, 68–72. doi: 10.1016/j.jpsychires.2018.10.009
Li, T. H., and Schmid, C. W. (2001). Differential stress induction of individual Alu loci: implications for transcription and retrotransposition. Gene 276, 135–141. doi: 10.1016/s0378-1119(01)00637-0
Mak, M., Samochowiec, J., Frydecka, D., Pełka-Wysiecka, J., Szmida, E., Karpiǹski, P., et al. (2019). First-episode schizophrenia is associated with a reduction of HERV-K methylation in peripheral blood. Psychiatry Res. 271, 459–463. doi: 10.1016/j.psychres.2018.12.012
Mameli, G., Astone, V., Arru, G., Marconi, S., Lovato, L., Serra, C., et al. (2007). Brains and peripheral blood mononuclear cells of multiple sclerosis (MS) patients hyperexpress MS-associated retrovirus/HERV-W endogenous retrovirus, but not human herpesvirus 6. J. Gen. Virol. 88, 264–274. doi: 10.1099/vir.0.81890-0
Marangoni, C., Hernandez, M., and Faedda, G. L. (2016). The role of environmental exposures as risk factors for bipolar disorder: a systematic review of longitudinal studies. J. Affect. Disord. 193, 165–174. doi: 10.1016/j.jad.2015.12.055
McGrath, J. J., and Welham, J. L. (1999). Season of birth and schizophrenia: a systematic review and meta-analysis of data from the Southern Hemisphere. Schizophr. Res. 35, 237–242. doi: 10.1016/s0920-9964(98)00139-x
McGuffin, P., Rijsdijk, F., Andrew, M., Sham, P., Katz, R., and Cardno, A. (2003). The heritability of bipolar affective disorder and the genetic relationship to unipolar depression. Arch. Gen. Psychiatry 60, 497–502.
Melbourne, J. K., Chase, K. A., Feiner, B., Rosen, C., and Sharma, R. P. (2018). Long non-coding and endogenous retroviral RNA levels are associated with proinflammatory cytokine mRNA expression in peripheral blood cells: implications for schizophrenia. Psychiatry Res. 262, 465–468. doi: 10.1016/j.psychres.2017.09.025
Meredith, R. M. (2015). Sensitive and critical periods during neurotypical and aberrant neurodevelopment: a framework for neurodevelopmental disorders. Neurosci. Biobehav. Rev. 50, 180–188. doi: 10.1016/j.neubiorev.2014.12.001
Mi, S., Lee, X., Li, X., Veldman, G. M., Finnerty, H., Racie, L., et al. (2000). Syncytin is a captive retroviral envelope protein involved in human placental morphogenesis. Nature 403, 785–789. doi: 10.1038/35001608
Mighdoll, M. I., Tao, R., Kleinman, J. E., and Hyde, T. M. (2015). Myelin, myelin-related disorders, and psychosis. Schizophr. Res. 161, 85–93. doi: 10.1016/j.schres.2014.09.040
Mighell, A. J., Markham, A. F., and Robinson, P. A. (1997). Alu sequences. FEBS Lett. 417, 1–5. doi: 10.1016/s0014-5793(97)01259-3
Miglino, N., Tamm, M., and Borger, P. (2014). Transposable element LINE1 is activated after exposure to cigarette smoke in primary human lung fibroblasts. Eur. Respir. J. 44:S2023.
Miller, B. J., Buckley, P., Seabolt, W., Mellor, A., and Kirkpatrick, B. (2011). Meta-analysis of cytokine alterations in schizophrenia: clinical status and antipsychotic effects. Biol. Psychiatry 70, 663–671. doi: 10.1016/j.biopsych.2011.04.013
Miller, B. J., Gassama, B., Sebastian, D., Buckley, P., and Mellor, A. (2013). Meta-analysis of lymphocytes in schizophrenia: clinical status and antipsychotic effects. Biol. Psychiatry 73, 993–999. doi: 10.1016/j.biopsych.2012.09.007
Milne, T. A., McEwen, B. S., Hunter, R. G., Pfaff, D. W., and McCarthy, K. J. (2009). Regulation of hippocampal H3 histone methylation by acute and chronic stress. Proc. Natl. Acad. Sci. U.S.A. 106, 20912–20917. doi: 10.1073/pnas.0911143106
Miousse, I. R., Chalbot, M. C. G., Lumen, A., Ferguson, A., Kavouras, I. G., and Koturbash, I. (2015). Response of transposable elements to environmental stressors. Mutat. Res. Rev. Mutat. Res. 765, 19–39. doi: 10.1016/j.mrrev.2015.05.003
Misiak, B., Frydecka, D., Piotrowski, P., and Kiejna, A. (2013). The multidimensional nature of metabolic syndrome in schizophrenia: lessons from studies of one-carbon metabolism and DNA methylation. Epigenomics 5, 317–329. doi: 10.2217/epi.13.22
Misiak, B., Frydecka, D., and Rybakowski, J. K. (2016). Editorial: endophenotypes for schizophrenia and mood disorders: implications from genetic, biochemical, cognitive, behavioral, and neuroimaging studies. Front. Psychiatry 7:83. doi: 10.3389/fpsyt.2016.00083
Misiak, B., Frydecka, D., Staǹczykiewicz, B., and Samochowiec, J. (2019). Editorial: peripheral markers of immune response in major psychiatric disorders: where are we now and where do we want to be? Front. Psychiatry 10:5. doi: 10.3389/fpsyt.2019.00005
Misiak, B., Krefft, M., Bielawski, T., Moustafa, A. A., Sąsiadek, M. M., and Frydecka, D. (2017a). Toward a unified theory of childhood trauma and psychosis: a comprehensive review of epidemiological, clinical, neuropsychological and biological findings. Neurosci. Biobehav. Rev. 75, 393–406. doi: 10.1016/j.neubiorev.2017.02.015
Misiak, B., Staǹczykiewicz, B., Kotowicz, K., Rybakowski, J. K., Samochowiec, J., and Frydecka, D. (2017b). Cytokines and C-reactive protein alterations with respect to cognitive impairment in schizophrenia and bipolar disorder: a systematic review. Schizophr. Res. 192, 16–29. doi: 10.1016/j.schres.2017.04.015
Misiak, B., Szmida, E., Karpiǹski, P., Loska, O., Sąsiadek, M. M., and Frydecka, D. (2015). Lower LINE-1 methylation in first-episode schizophrenia patients with the history of childhood trauma. Epigenomics 7, 1275–1285. doi: 10.2217/epi.15.68
Mondelli, V., Ciufolini, S., Murri, M. B., Bonaccorso, S., Di Forti, M., Giordano, A., et al. (2015). Cortisol and inflammatory biomarkers predict poor treatment response in first episode psychosis. Schizophr. Bull. 41, 1162–1170. doi: 10.1093/schbul/sbv028
Morozov, V. A., Dao Thi, V. L., and Denner, J. (2013). The transmembrane protein of the human endogenous retrovirus - K (HERV-K) modulates cytokine release and gene expression. PLoS One 8:e70399. doi: 10.1371/journal.pone.0070399
Mortensen, P. B., Pedersen, C. B., Mcgrath, J. J., Hougaard, D. M., Nørgaard-Petersen, B., Mors, O., et al. (2011). Neonatal antibodies to infectious agents and risk of bipolar disorder: a population-based case-control study. Bipolar Disord. 13, 624–629. doi: 10.1111/j.1399-5618.2011.00962.x
Munoz-Lopez, M., and Garcia-Perez, J. (2010). DNA transposons: nature and applications in genomics. Curr. Genomics 11, 115–128. doi: 10.2174/138920210790886871
Muotri, A. R., Chu, V. T., Marchetto, M. C. N., Deng, W., Moran, J. V., and Gage, F. H. (2005). Somatic mosaicism in neuronal precursor cells mediated by L1 retrotransposition. Nature 435, 903–910. doi: 10.1038/nature03663
Muotri, A. R., Marchetto, M. C. N., Coufal, N. G., Oefner, R., Yeo, G., Nakashima, K., et al. (2010). L1 retrotransposition in neurons is modulated by MeCP2. Nature 468, 443–446. doi: 10.1038/nature09544
Murphy, S. K., Fineberg, A. M., Maxwell, S. D., Alloy, L. B., Zimmermann, L., Krigbaum, N. Y., et al. (2017). Maternal infection and stress during pregnancy and depressive symptoms in adolescent offspring. Psychiatry Res. 257, 102–110. doi: 10.1016/j.psychres.2017.07.025
Mutirangura, A. (ed.). (2019). “A hypothesis to explain how the DNA of elderly people is prone to damage: genome-wide hypomethylation drives genomic instability in the elderly by reducing youth-associated gnome-stabilizing DNA gaps,” in Epigenetics [Working Title], (London: Intechopen). doi: 10.5772/intechopen.83372
Nagy, C., and Turecki, G. (2012). Sensitive periods in epigenetics: bringing us closer to complex behavioral phenotypes. Epigenomics 4, 445–457. doi: 10.2217/epi.12.37
Najjar, S., and Pearlman, D. M. (2015). Neuroinflammation and white matter pathology in schizophrenia: systematic review. Schizophr. Res. 161, 102–112. doi: 10.1016/j.schres.2014.04.041
Nestler, E. J., Peña, C. J., Kundakovic, M., Mitchell, A., and Akbarian, S. (2016). Epigenetic basis of mental illness. Neuroscientist 22, 447–463. doi: 10.1177/1073858415608147
Oja, M., Peltonen, J., Blomberg, J., and Kaski, S. (2008). Methods for estimating human endogenous retrovirus activities from EST databases. BMC Bioinformatics 8(Suppl. 2):S11. doi: 10.1186/1471-2105-8-S2-S11
Ostertag, E. M., Goodier, J. L., Zhang, Y., and Kazazian, H. H. (2003). SVA elements are nonautonomous retrotransposons that cause disease in humans. Am. J. Hum. Genet. 73, 1444–1451. doi: 10.1086/380207
Parboosing, R., Bao, Y., Shen, L., Schaefer, C. A., and Brown, A. S. (2013). Gestational influenza and bipolar disorder in adult offspring. JAMA Psychiatry 70, 677–685. doi: 10.1001/jamapsychiatry.2013.896
Patchsung, M., Settayanon, S., Pongpanich, M., Mutirangura, D., Jintarith, P., and Mutirangura, A. (2018). Alu siRNA to increase Alu element methylation and prevent DNA damage. Epigenomics 10, 175–185. doi: 10.2217/epi-2017-0096
Perron, H., Hamdani, N., Faucard, R., Lajnef, M., Jamain, S., Daban-Huard, C., et al. (2012). Molecular characteristics of Human Endogenous Retrovirus type-W in schizophrenia and bipolar disorder. Transl. Psychiatry 2:e201. doi: 10.1038/tp.2012.125
Perron, H., Mekaoui, L., Bernard, C., Veas, F., Stefas, I., and Leboyer, M. (2008). Endogenous retrovirus type W GAG and envelope protein antigenemia in serum of schizophrenic patients. Biol. Psychiatry 64, 1019–1023. doi: 10.1016/j.biopsych.2008.06.028
Poduri, A., Evrony, G. D., Cai, X., and Walsh, C. A. (2013). Somatic mutation, genomic variation, and neurological disease. Science 341:1237758. doi: 10.1126/science.1237758
Protasova, M. S., Gusev, F. E., Grigorenko, A. P., Kuznetsova, I. L., Rogaev, E. I., and Andreeva, T. V. (2017). Quantitative analysis of L1-retrotransposons in Alzheimer’s disease and aging. Biochemistry 82, 962–971. doi: 10.1134/S0006297917080120
Ripke, S., Neale, B. M., Corvin, A., Walters, J. T. R., Farh, K. H., Holmans, P. A., et al. (2014). Biological insights from 108 schizophrenia-associated genetic loci. Nature 511, 421–427. doi: 10.1038/nature13595
Rusiecki, J. A., Chen, L., Srikantan, V., Zhang, L., Yan, L., Polin, M. L., et al. (2012). DNA methylation in repetitive elements and post-traumatic stress disorder: a case-control study of US military service members. Epigenomics 4, 29–40. doi: 10.2217/epi.11.116
Saeliw, T., Tangsuwansri, C., Thongkorn, S., Chonchaiya, W., Suphapeetiporn, K., Mutirangura, A., et al. (2018). Integrated genome-wide Alu methylation and transcriptome profiling analyses reveal novel epigenetic regulatory networks associated with autism spectrum disorder. Mol. Autism 9:27. doi: 10.1186/s13229-018-0213-9
Sassaman, D. M., Dombroski, B. A., Moran, J. V., Kimberland, M. L., Naas, T. P., DeBerardinis, R. J., et al. (1997). Many human L1 elements are capable of retrotransposition. Nat. Genet. 16, 37–43. doi: 10.1038/ng0597-37
Schumann, G. G., Gogvadze, E. V., Osanai-Futahashi, M., Kuroki, A., Münk, C., Fujiwara, H., et al. (2010). Unique functions of repetitive transcriptomes. Int. Rev. Cell Mol. Biol. 285, 115–188. doi: 10.1016/B978-0-12-381047-2.00003-7
Shpyleva, S., Melnyk, S., Pavliv, O., Pogribny, I., and Jill James, S. (2018). Overexpression of LINE-1 retrotransposons in autism brain. Mol. Neurobiol. 55, 1740–1749. doi: 10.1007/s12035-017-0421-x
Su, J., Shao, X., Liu, H., Liu, S., Wu, Q., and Zhang, Y. (2012). Genome-wide dynamic changes of DNA methylation of repetitive elements in human embryonic stem cells and fetal fibroblasts. Genomics 99, 10–17. doi: 10.1016/j.ygeno.2011.10.004
Suarez, N. A., Macia, A., and Muotri, A. R. (2018). LINE-1 retrotransposons in healthy and diseased human brain. Dev. Neurobiol. 78, 434–455. doi: 10.1002/dneu.22567
Talens, R. P., Christensen, K., Putter, H., Willemsen, G., Christiansen, L., Kremer, D., et al. (2012). Epigenetic variation during the adult lifespan: cross-sectional and longitudinal data on monozygotic twin pairs. Aging Cell 11, 694–703. doi: 10.1111/j.1474-9726.2012.00835.x
Tangsuwansri, C., Saeliw, T., Thongkorn, S., Chonchaiya, W., Suphapeetiporn, K., Mutirangura, A., et al. (2018). Investigation of epigenetic regulatory networks associated with autism spectrum disorder (ASD) by integrated global LINE-1 methylation and gene expression profiling analyses. PLoS One 13:e0201071. doi: 10.1371/journal.pone.0201071
Tartaglione, A. M., Cipriani, C., Chiarotti, F., Perrone, B., Balestrieri, E., Matteucci, C., et al. (2018). Early behavioral alterations and increased expression of endogenous retroviruses are inherited across generations in mice prenatally exposed to valproic acid. Mol. Neurobiol. 56, 3736–3750. doi: 10.1007/s12035-018-1328-x
Van Den Oord, E. J. C. G., Clark, S. L., Xie, L. Y., Shabalin, A. A., Dozmorov, M. G., Kumar, G., et al. (2016). A whole methylome CpG-SNP association study of psychosis in blood and brain tissue. Schizophr. Bull. 42, 1018–1026. doi: 10.1093/schbul/sbv182
Vargiu, L., Rodriguez-Tomé, P., Sperber, G. O., Cadeddu, M., Grandi, N., Blikstad, V., et al. (2016). Classification and characterization of human endogenous retroviruses mosaic forms are common. Retrovirology 13:7. doi: 10.1186/s12977-015-0232-y
Vidaud, D., Vidaud, M., Bahnak, B. R., Siguret, V., Gispert Sanchez, S., Laurian, Y., et al. (1993). Haemophilia B due to a de novo insertion of a human-specific Alu subfamily member within the coding region of the factor IX gene. Eur. J. Hum. Genet. 1, 30–36.
Vigo, D., Thornicroft, G., and Atun, R. (2016). Estimating the true global burden of mental illness. Lancet Psychiatry 3, 171–178. doi: 10.1016/S2215-0366(15)00505-2
Wang, H., Xing, J., Grover, D., Hedges Kyudong, Han, D. J., Walker, J. A., et al. (2005). SVA elements: a hominid-specific retroposon family. J. Mol. Biol. 354, 994–1007. doi: 10.1016/j.jmb.2005.09.085
Weis, S., Llenos, I. C., Sabunciyan, S., Dulay, J. R., Isler, L., Yolken, R., et al. (2007). Reduced expression of human endogenous retrovirus (HERV)-W GAG protein in the cingulate gyrus and hippocampus in schizophrenia, bipolar disorder, and depression. J. Neural Transm. 114, 645–655. doi: 10.1007/s00702-006-0599-y
Yao, Y., Schröder, J., Nellåker, C., Bottmer, C., Bachmann, S., Yolken, R. H., et al. (2008). Elevated levels of human endogenous retrovirus-W transcripts in blood cells from patients with first episode schizophrenia. Genes Brain Behav. 7, 103–112.
Zhang, Z., Harrison, P. M., Liu, Y., and Gerstein, M. (2003). Millions of years of evolution preserved: a comprehensive catalog of the processed pseudogenes in the human genome. Genome Res. 13, 2541–2558. doi: 10.1101/gr.1429003
Keywords: retrotransposon, DNA methylation, LINE-1, Alu, SINE, SVA
Citation: Misiak B, Ricceri L and Sąsiadek MM (2019) Transposable Elements and Their Epigenetic Regulation in Mental Disorders: Current Evidence in the Field. Front. Genet. 10:580. doi: 10.3389/fgene.2019.00580
Received: 10 April 2019; Accepted: 04 June 2019;
Published: 25 June 2019.
Edited by:
Yun Liu, Fudan University, ChinaReviewed by:
Nicole Grandi, University of Cagliari, ItalyApiwat Mutirangura, Chulalongkorn University, Thailand
Copyright © 2019 Misiak, Ricceri and Sąsiadek. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Błażej Misiak, YmxhemVqX21pc2lha0BpbnRlcmlhLnBs; bWJsYXplakBpbnRlcmlhLmV1