 Julian Ramírez-Bello1*
Julian Ramírez-Bello1* José M. Fragoso2
José M. Fragoso2 Isidro Alemán-Ávila1Silvia Jiménez-Morales3
Isidro Alemán-Ávila1Silvia Jiménez-Morales3 Alma D. Campos-Parra4Rosa Elda Barbosa-Cobos5
Alma D. Campos-Parra4Rosa Elda Barbosa-Cobos5 José Moreno6
José Moreno6- 1Unidad de Investigación, Hospital Juárez de México, Mexico City, Mexico
- 2Laboratorio de Biología Molecular, Instituto Nacional de Cardiología, Mexico City, Mexico
- 3Laboratorio de Genómica del Cáncer, Instituto Nacional de Medicina Genómica, Mexico City, Mexico
- 4Laboratorio de Genómica, Instituto Nacional de Cancerología, Mexico City, Mexico
- 5Servicio de Reumatología, Hospital Juárez de México, Mexico City, Mexico
- 6Dirección de Investigación, Hospital Juárez de México, Mexico City, Mexico
Introduction: BLK has been identified as a risk factor to rheumatoid arthritis (RA) primarily in Asian or European-derived populations. However, this finding has not been evaluated in other populations such as Latin-Americans, except for Colombians. On the other hand, BANK1 single nucleotide variants (SNVs) have been scarcely studied in RA patients.
Objective: The aim of this study was to determine whether the BLK rs2736340T/C, rs13277113A/G, and BANK1 rs10516487G/A (R61H) and rs3733197G/A (A383T) polymorphisms are risk factors to RA in a sample of patients from Central Mexico.
Materials and Methods: We studied 957 women; 487 controls and 470 patients with RA by means of a TaqMan® SNP genotyping assay with fluorescent probes for the BLK rs13277113A/G, rs2736340T/C and BANK1 10516487G/A (R61H) and rs3733197G/A (A383T) variants.
Result: The BLK rs2736340T/C and rs13277113A/G variants were associated with risk for RA: C vs T; OR 1.39, p = 0.001, and G vs A; OR 1.37, p = 0.004, respectively. In addition, there was also an association between BANK1 R61H and RA: A vs G; OR 1.49, p = 0.003, but no with BANK1 A383T. We also identified an interaction significant between genotypes of BLK rs2736340T/C-BANK1 rs10516487G/A and RA: OR 1.65, p = 0.0001.
Conclusions: Our data suggest that both BLK and BANK1 confer susceptibility to RA in Mexican patients. The individual association of BANK1 rs1054857G/A with RA had not been previously reported in a particular population (except for pooled patients from several countries), therefore, our study presents the first evidence of association between this BANK1 variant and RA.
Introduction
Genetic association to autoimmune diseases (ADs) includes variants of genes coding for proteins involved in B lymphocyte antigen receptor signaling, of which BLK and BANK1 are components of the B-cell signalosome (Suthers and Sarantopoulos, 2017). The BLK protein is a src family non-receptor tyrosine kinase mainly expressed by B-cells, where besides B-cell receptor signaling, it plays a role in development (Akerblad and Sigvardsson, 1999; Tretter et al., 2003), meanwhile, the BANK1 protein is an adaptor/scaffold primarily expressed in B-lymphocytes, which plays an important role in activation and signaling (Kozyrev et al., 2008; Castillejo-Lopez et al., 2012). It has been shown that BLK, similar to other members of the src family interacts with BANK1 (Castillejo-Lopez et al., 2012). Thus, BLK and BANK1 proteins have an important role in both B-cell signaling and activation.
In 2009 and 2011, two genome wide association studies (GWAS) carried out in European and Asian derived-populations, respectively, identified some single nucleotide variants (SNVs) in the BLK gene to be associated with risk for rheumatoid arthritis (RA) (Gregersen et al., 2009; Freudenberg et al., 2011). However, other GWA or candidate gene studies have failed to replicate these findings (Génin et al., 2013; Jiang et al., 2014; Orozco et al., 2014; Bossini-Castillo et al., 2015; Zhu et al., 2016; Julià et al., 2016; Saxena et al., 2017), except in patients from United Kingdom, Colombia, and China (Deshmukh et al., 2011; Orozco et al., 2011; Viatte et al., 2012; Huang et al., 2017). On the other hand, the BANK1 gene, which was identified as an important risk factor for systemic lupus erythematosus (SLE) (Kozyrev et al., 2008), does not appear to be associated with RA through GWA (Gregersen et al., 2009; Freudenberg et al., 2011; Jiang et al., 2014; Orozco et al., 2014; Bossini-Castillo et al., 2015; Julià et al., 2016; Saxena et al., 2017) or candidate gene studies (Suarez-Gestal et al., 2009; Orozco et al., 2011; Génin et al., 2013; Huang et al., 2017), except for the BANK1 rs3733197G/A (Ala383Thr; non-synonymous polymorphism) SNV, which showed an association with RA in Spanish and Argentine patients (Orozco et al., 2009).
Although different BLK and BANK1 SNVs have been examined in patients with RA, the results are uncertain because some reports show an association with this AD (Gregersen et al., 2009; Orozco et al., 2009; Deshmukh et al., 2011; Freudenberg et al., 2011; Orozco et al., 2011; Viatte et al., 2012; Huang et al., 2017), while others have not replicated this finding (Suarez-Gestal et al., 2009; Génin et al., 2013; Jiang et al., 2014; Orozco et al., 2014; Bossini-Castillo et al., 2015; Julià et al., 2016; Zhu et al., 2016; Saxena et al., 2017). In addition, BLK (except in patients from Colombia) and BANK1 (except in one study which evaluated patients from Argentina and Mexico) variants have been scarcely explored in different Latin-American populations with RA (Orozco et al., 2009; Deshmukh et al., 2011). Therefore, our aim was to examine the possible associations of BLK rs13277113A/G and rs2736340T/C and BANK1 rs10516487C/T (R61H) and rs3733197G/A (A383T) SNVs with RA in Mexican patients, in addition to the interactions between BLK and BANK1 genotypes and this AD.
Material and Methods
Patients and Controls
The present study included 957 individuals from Central Mexico (from Mexico City, States of Mexico, Morelos, Hidalgo, and Puebla), 470 with RA and 487 controls. Because our sample was formed by a proportion of women/men affected with RA of 93.6% and 6.3% (data not shown), respectively, and to avoid any possible bias as a result of the low number of males with possibly distinct features, we decided to exclude men from our study. Previous studies in Mexico have shown a similar proportion of women/men affected with RA (Muñoz-Valle et al., 2017; Zaragoza-García et al., 2019), which is very different from what was reported in Caucasians, where RA is three times more frequent in women than in men (Scott et al., 2010). Therefore, all cases and controls were unrelated women >18 years old. Healthy individuals included in this study were obtained from the blood bank laboratory of Hospital Juárez de México (HJM) and had no family history of ADs or inflammatory diseases, and no personal history of obesity, hypertension, cancer, and allergy. All RA patients fulfilled the 2010 ACR-EULAR criteria (Aletaha et al., 2010) and were recruited from the Department of Rheumatology, HJM. Patients and controls were matched by ancestry and gender. Studies were conducted in compliance with the Declaration of Helsinki. Additionally, all individuals included in our study signed a written informed consent. This protocol was approved by the Ethics and Research Committees of the HJM (registry number 0446/18-I).
DNA Extraction
Peripheral blood samples (5–8 ml of EDTA-treated) were used to isolate genomic DNA from leukocytes by means of the Invisorb Bood Universal Kit (Stratec Molecular GmbH, Berlin, Germany), according to manufacturer's specifications. DNA samples were quantified, diluted (5 ng/µl), and stored at −20°C until needed.
Genotyping
We used a TaqMan® SNP genotyping assay (Applied Biosystems, Foster City, CA) for the BLK rs13277113A/G and rs2736340T/C and BANK1 rs10516487C/T (R61H) and rs3733197G/A (A383T) genotypes. The Bio-Rad CFX Manager 3.1 software implemented in the CFX96 Touch TM Real-Time PCR of Bio-Rad (Bio-Rad, California, USA) was used to determinate the BLK and BANK1 genotypes in an allelic discrimination plot. The distribution of each BLK and BANK1 genotype was determined by two independent researchers. Additionally, 70% of all samples (including cases and controls) were genotyped twice with a reproducibility of 100%. PCR conditions for each amplicon and sample were as follows: 10 ng DNA per sample, 2.5 μl of TaqMan® Universal Master Mix (2X) (Applied Biosystems, Foster City, CA), 2.435 μl of nuclease-free water, and 0.065 μl of TaqMan probes (Applied Biosystems). The PCR protocol for amplification was as follows: pre-PCR (one cycle); at 50°C for 2 min and at 95°C for 8 min, followed by 45 cycles of denaturing at 95°C for 15 s and annealing an extension at 60°C for 1 min.
Statistical Analysis
We used the Finetti software to evaluate the Hardy-Weinberg equilibrium (HWE) for all BLK and BANK1 SNVs (a p-value < 0.05 in both cases and controls was indicative of deviation from HWE) (https://ihg.gsf.de/cgi-bin/hw/hwa1.pl). We used the Epidat program (http://www.sergas.es/MostrarContidos_N3_T01.aspx?IdPaxina=62715) to examine the genetic association between BLK and BANK1 polymorphisms and RA under the allelic, codominant, dominant, and recessive genetic models. All results were corrected through the Bonferroni correction test (0.05/4 SNVs; p = 0.0125), with a p-value between 0.05 and 0.0125 considered as nominal significance, and p-value ≤0.0125 as significant. Moreover, all p-values obtained of different inheritance genetic models were evaluated by logistic regression and adjusted for age and city of origin. The Haploview program was used to obtain the haplotypes and the linkage disequilibrium (LD) among BLK and BANK1 markers (Barrett et al., 2005). Interactions between genotypes of high and low risk of the four BLK and BANK1 variants for both cases and controls were obtained by the multifactor dimensionality reduction (MDR) program, which allows the evaluation of the quality of gene-gene interactions to be measured by means of two-way contingency tables (Moore et al., 2006). Quanto software (http://hydra.usc.edu/gxe) was used to determine the statistical power of our study. We took into account the minor allele frequency (MAF) of the four BLK and BANK1 SNVs in controls, in addition to a recessive genetic model, the proportion of cases-controls, an odd ratio (OR) of 2.0, the prevalence of RA in Mexicans (Peláez-Ballestas et al., 2011) as well as the sample size.
Results
HWE and Statistical Power in our Study Population
The BLK and BANK1 genotypes were in HWE both in patients with RA and controls, except for BANK1 rs3733197G/A in controls, which had a weak deviation from HWE (p = 0.02). We identified a statistical power >99% in our study taking into account the MAF of the four BLK and BANK1 variants in controls (data not shown).
Allele and Genotype Frequencies of the BLK Polymorphisms and Association Analysis
BLK rs13277113A and rs2736340T major alleles were associated with RA (G vs A; OR 1.37, p = 0.002, and C vs T; OR 1.39, p = 0.001, respectively [Table 1]). We also identified an association of BLK rs13277113A/G with RA under the recessive (OR 1.52, p = 0.0013), and codominant (OR 1.52, p = 0.0055) models but not under the dominant model (Table 1). Because the BLK rs13277113A/G variant is in high LD with rs2736340T/C (r2 ≈ 1), we also observed a similar association between BLK rs2736340T/C and RA, C v T; OR 1.39, p = 0.001, and under the recessive and codominant models, the OR were 1.55, (p = 0.0008) and 2.58 (p = 0.0036), respectively (Table 1).
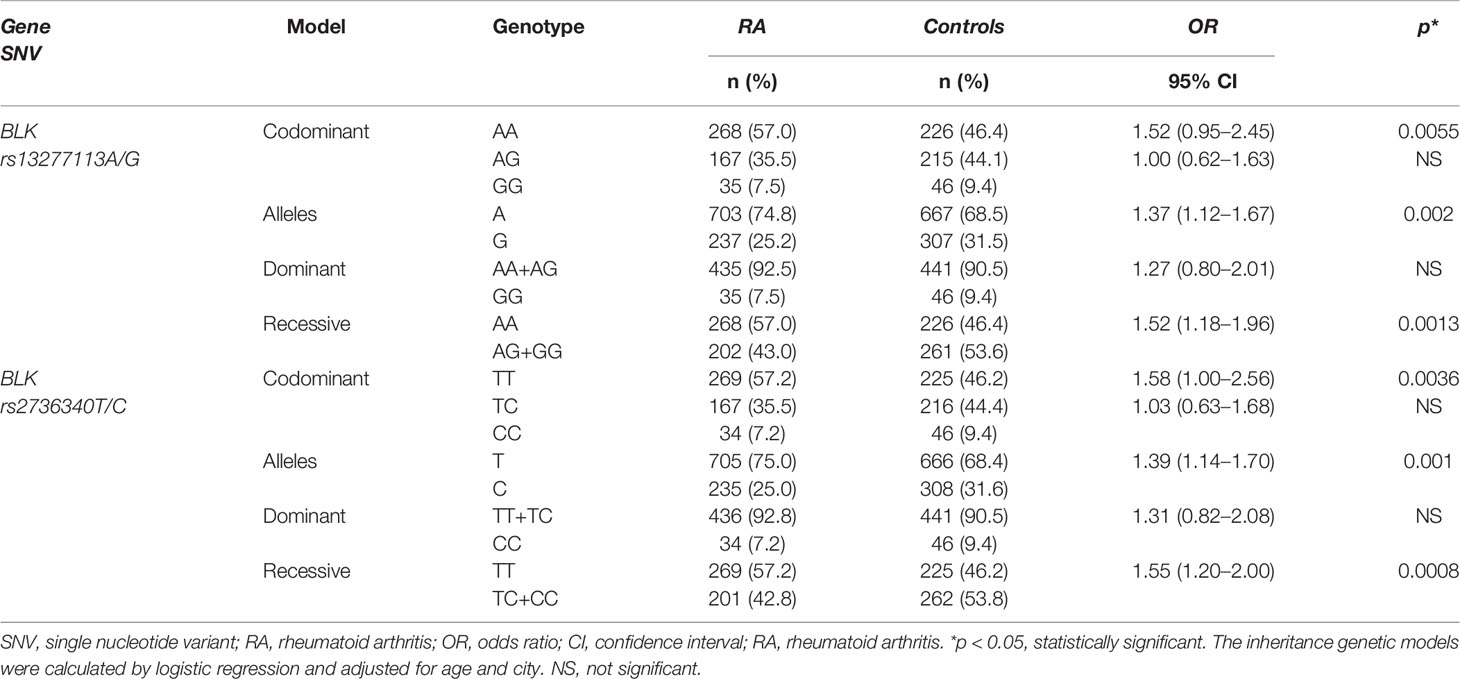
Table 1 Genotypic and allelic frequencies of the BLK rs13277113A/G and rs2736340T/C SNVs and association analysis in RA patients and healthy individuals.
Allele and Genotype Frequencies of the BANK1 Polymorphisms and Association Analysis
Our data shows that the BANK1 rs10516487G/A variant is a risk factor to RA. Of note, we identified BANK1 rs10516487A as the major allele associated with susceptibility to this AD: A vs G; OR 1.49, p = 0.003 (Table 2). We also observed an association under the recessive (OR 1.47, p = 0.01) and codominant (OR 2.16, p = 0.012) models and a nominal significance under the dominant model (OR 2.78, p = 0.036) (Table 2). No association was identified between BANK1 rs3733197G/A and RA under any genetic model (Table 2).
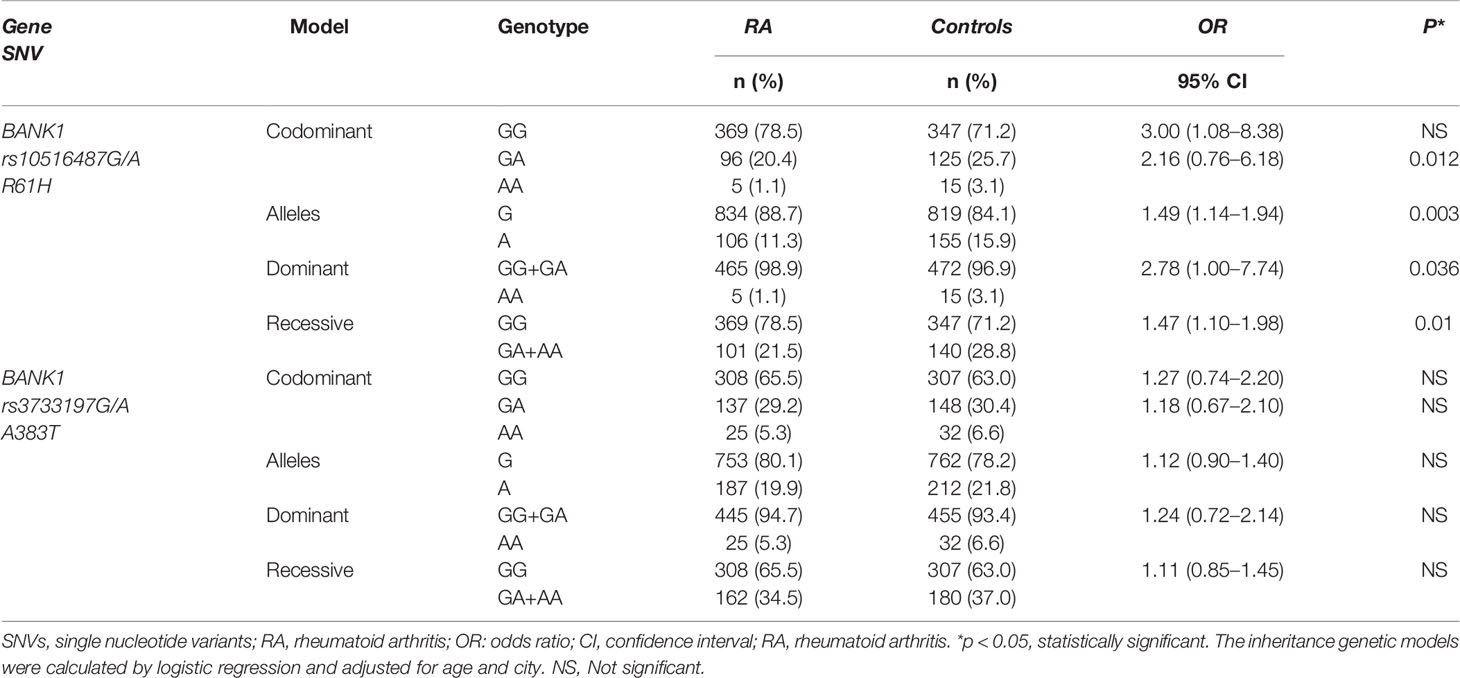
Table 2 Genotypic and allelic frequencies of the BANK1 rs10516487G/A and rs3733197G/A SNVs and association analysis in RA patients and healthy individuals.
Haplotypes and LD Analysis
We identified a pair of haplotypes between the two BLK SNVs in RA patients because of a strong LD among them (data not shown), the TA haplotype, which carries the major alleles of the BLK rs2736340T/C and rs13277113A/G SNVs showed an association with susceptibility to RA; OR 1.36, p = 0.0023, pc 0.0024 after 100,000 permutations (data not shown). We also identified an association between the AA haplotype carrying the BANK1 rs10516487G/A and rs3733197G/A minor alleles and protection against RA; OR 0.68, p = 0.01 (pc = 0.038) (Table 3), indicating that the BANK1 rs10516487G major allele together with the rs3733197A minor allele; GA haplotype, is not associated with RA. Regarding the BANK1 variants, there was no LD between the two BANK1 SNVs (data not shown).
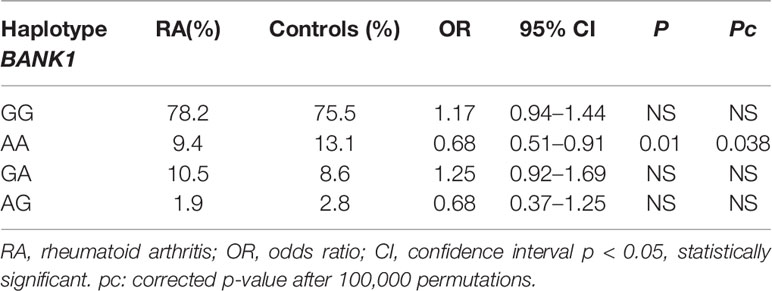
Table 3 Haplotype frequencies and association analysis between BANK1 SNVs in RA patients and controls. The order of the BANK1 SNVs is: rs10516487G/A and rs3733197G/A.
Genetic Interactions Between BLK and BANK1 Genotypes and RA
The distribution of interactions between the BLK rs2736340T/C-rs13277113A/G and BANK1 rs10516487G/A-rs3733197G/A genotypes in RA patients and controls are shown in Figure 1. We identified three genetic interactions, of which the best model was represented by BLK rs2736340T/C and BANK1 rs10516487G/A (testing accuracy 0.5443, and cross-validation consistency 10/10), this interaction also showed an association with susceptibility to RA (OR 1.65, p = 0.0001) (Table 4). Of note, both BLK rs36340T/C (this variant showed almost a complete LD, r2≈1, with BLK rs13277113A/G) and BANK1 rs10516487G/A variants were previously identified to be associated with RA (Tables 1 and 2). Because of strong LD between BLK rs2736340T/C and rs13277113A/G SNVs, we identified in the dendrogram analysis a redundancy between these markers (data not shown). In addition, we also identified the BANK1 rs3733197G/A variant with an independent effect (data not shown). The second best interaction model was formed by the BLK rs2736340T/C-rs13277113A/G plus BANK1 rs10516487G/A-rs3733197G/A genotypes (testing accuracy; 0.5381, and cross-validation consistency; 10/10), this interaction was also associated with RA (OR 1.79, p < 0.0001). Finally, we did not observe a synergistic interaction between the four BLK an BANK1 variants (data not shown).
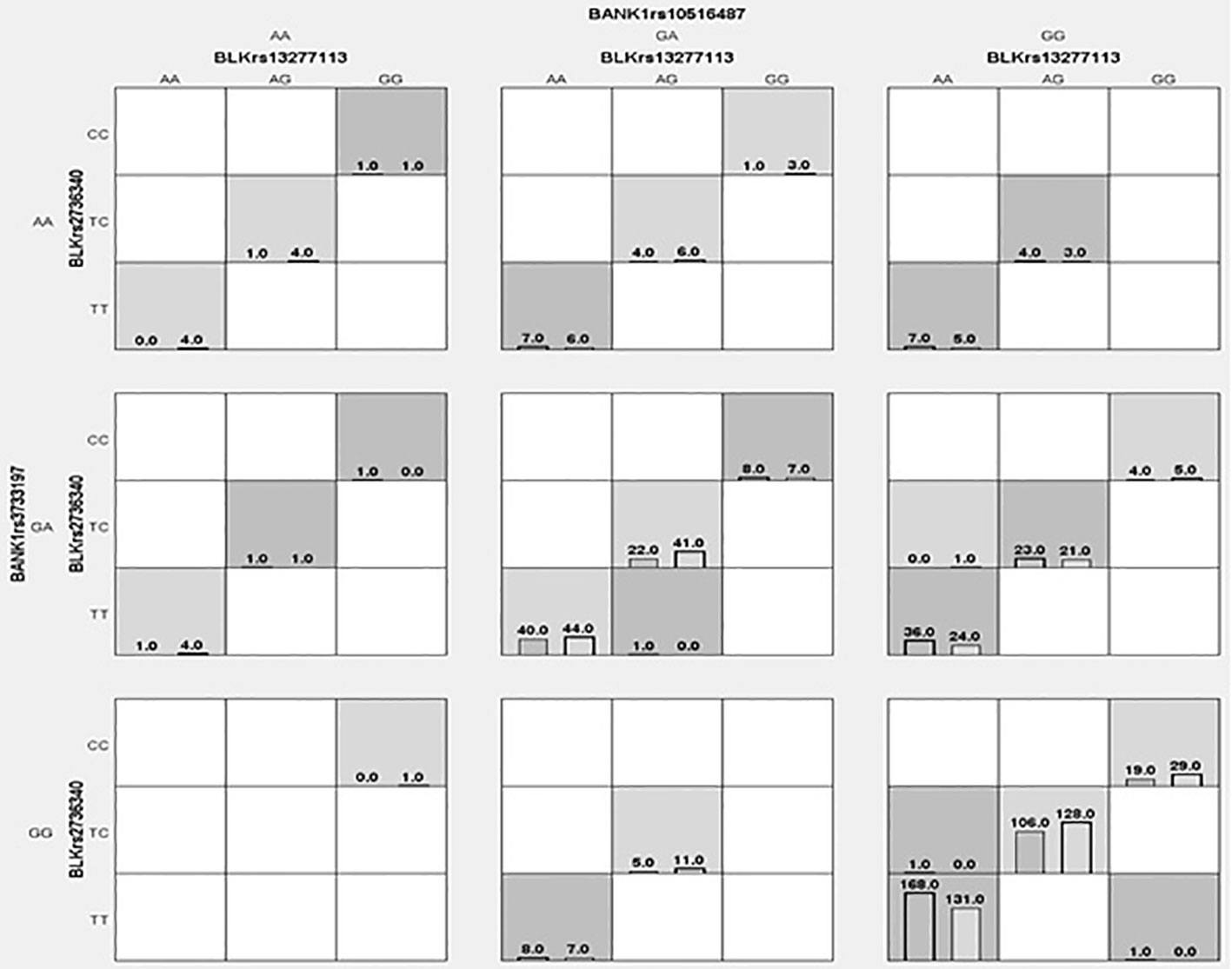
Figure 1 Distribution of genotypes in cases and controls for BLK rs13277113A/G-rs2736340T/C and BANK1 rs10516487G/A-rs3733197G/A. Each cell shows counts of cases (left) and controls (right). Dark-shaded cells represent “high-risk” genotypes, meanwhile, lighter-shaded cells represent “low-risk” genotypes. White-shades cells represent empty, that is, there are no cases-controls with genotypes.
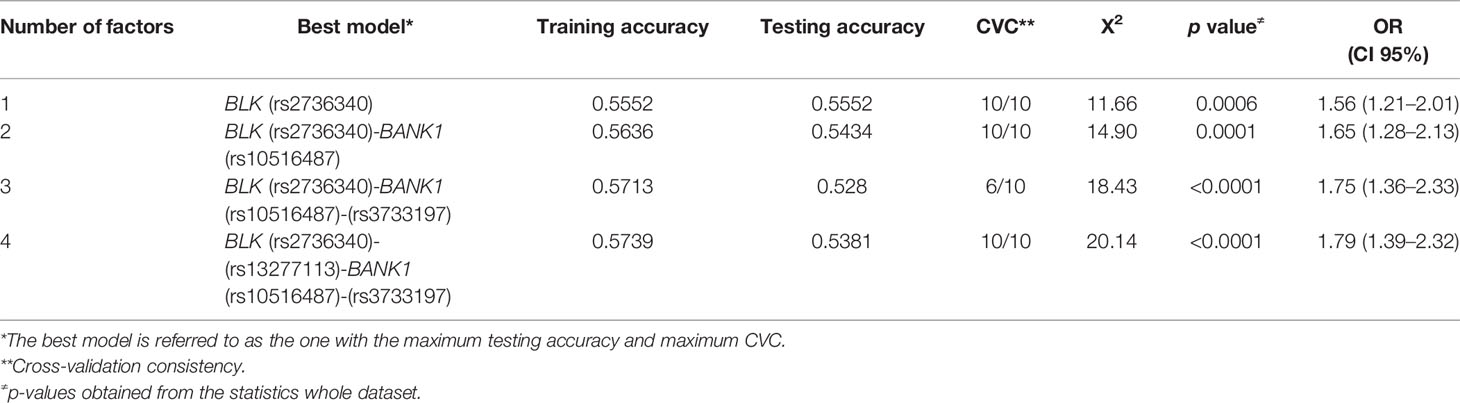
Table 4 Gene-gene interaction models between BLK and BANK1 SNVs in cases with RA and controls.
Discussion
The study of gene polymorphism associations to diseases in different ethnic groups is an important means to close down on the likelihood that particular genes and specific variants to the disease. Here we studied two SNVs of the BLK gene, and two of BANK1 (both of them part of the B cell signalosome) for their association to RA in Mexican patients. Association of BLK SNVs to RA was initially documented in North American and Korean patients (Gregersen et al., 2009; Freudenberg et al., 2011), and later in British, Colombian, and Han Chinese patients (Deshmukh et al., 2011; Orozco et al., 2011; Viatte et al., 2012; Huang et al., 2017). Nevertheless, other GWA or candidate gene studies carried out in some Arab countries and Europeans from the Netherlands and Spain, and even another Chinese group failed to replicate this finding (Suarez-Gestal et al., 2009; Génin et al., 2013; Jiang et al., 2014; Bossini-Castillo et al., 2015; Julià et al., 2016; Zhu et al., 2016; Saxena et al., 2017). As far as we know, the only study conducted in Latin-Americans thus far, found an association of both BLK rs13277113A/G and rs2736340T/C SNVs and risk for RA in Colombians (Deshmukh et al., 2011). Our current findings confirm such association in Mexican RA patients. On the other hand, in European-derived populations, BLK association to RA has only been identified in patients from the UK (Orozco et al., 2011; Viatte et al., 2012). It is important to note that in Chinese population, one candidate gene study found an association between BLK rs13277113A/G and RA (Huang et al., 2017), which was not previously identified in a GWAS (Jiang et al., 2014). GWAS allow the identification of many genes associated with any disease, including RA or other ADs (Gregersen et al., 2009; Freudenberg et al., 2011; Kozyrev et al., 2008; Rodríguez-Elías et al., 2016). However, in these studies, several hundreds or thousands of SNVs are removed, particularly because of their high error rates, such as those with an excess missing genotype (Anderson et al., 2010). Possible explanations for this are: a) the probes for the genotyping of BLK rs13277113A/G and rs2736340T/C (or SNVs in high LD for these variants) SNVs might have been lost in that process, b) the controls had a deviation from the HWE, c) these variants had no associations at genome-wide significance level (p < 5×10−8), among others. All these caveats must be considered to explain the controversial results found among the aforementioned studies. To solve this, it would be useful to carry out replications with candidate gene studies in the different populations to confirm or rule out these findings. It is important to note that in controls from North America, Spain (or European-derived populations), Colombia, and Africa, BLK rs2736340T or the BLK rs13277113A are the minor alleles (Hom et al., 2008; Gregersen et al., 2009; Orozco et al., 2009; Deshmukh et al., 2011; Sánchez et al., 2011; Génin et al., 2013; Guthridge et al., 2014; Martínez-Bueno et al., 2018) whereas in our population these were the major alleles as reported in the 1000 Genomes Project for Mexicans (Mexican-Americans from Los Angeles, CA) (https://www.ncbi.nlm.nih.gov/variation/tools/1000genomes/) as well as in other Asian populations like Japanese and Chinese (Génin et al., 2013; Dang et al., 2016; Huang et al., 2017) (Table 5). We conclude, hence, that in our population the BLK rs2736340T and/or rs13277113A major alleles are associated to risk for RA.
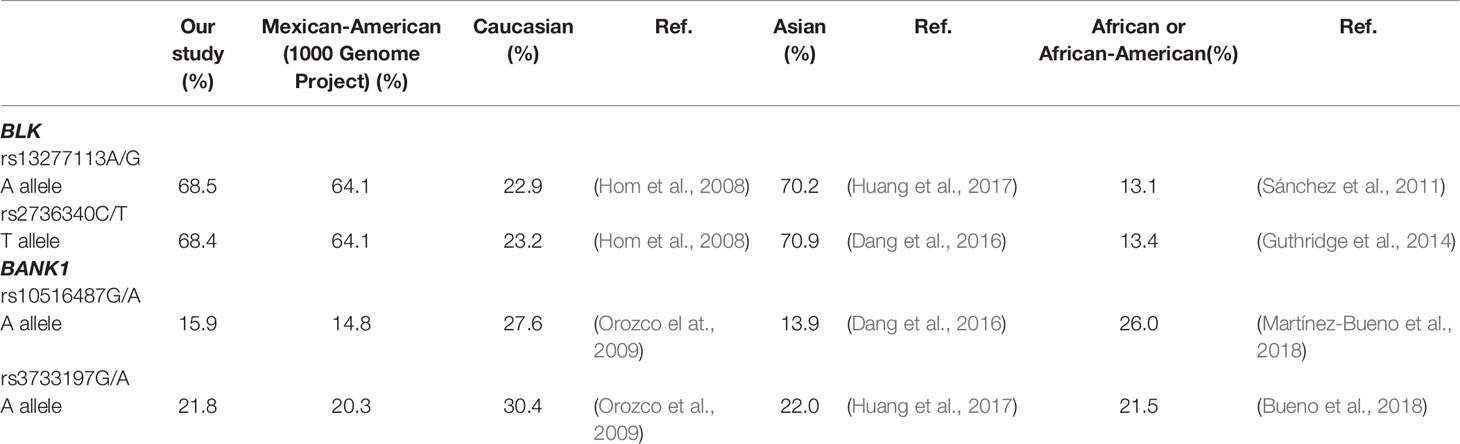
Table 5 Frequency of the BLK rs13277113G, rs2736340T and BANK1 rs10516487A-rs3733197A alleles associated with RA susceptibility in our study, in Mexican-Americans who lives in the Ángeles as well as in European, Asian, and African (or African-American)-derived populations. All data presented here are from control individuals.
Functionally, the rs13277113A allele has been associated with lower levels of BLK mRNA in transformed B-cell lines (Hom et al., 2008). However, it is possible that this BLK SNV is in LD with another BLK variant truly associated with risk for RA. In this manner, BLK rs13277113A appears to belong to a haplotype associated with RA, such as the BLK rs922483 variant (which is in high LD; r2 > 0.7 with both BLK rs13277113 and rs2736340T/C), which affects the BLK mRNA and protein expression early in B cell ontogeny (Simpfendorfer et al., 2012). Moreover, another study found that the rs922483 and rs1382568 variants located within the proximal BLK promoter and in the upstream alternative BLK promoter, respectively (both BLK variants are in high LD with rs13277113; r2 > 0.74 in Europeans), affect the promoter activity in B progenitor cell lines (Guthridge et al., 2014).
On the other hand, the two BANK1 rs10516487C/T (R61H) and rs3733197G/A (A383T) variants have been scarcely studied in patients with RA (Orozco et al., 2009; Suarez-Gestal et al., 2009; Orozco et al., 2011; Génin et al., 2013; Huang et al., 2017), having found only one association with BANK1 rs3733197G/A in RA patients from Spain and Argentina (Orozco et al., 2009). In that same study, a sample of Mexican RA patients was examined for BANK1 rs10516487C/T, rs3733197G/A, and rs17266594T/C showing no association. There was, however, an association of RA with BANK1 rs10516487G/A and rs3733197G/A when the cases and controls of all the countries were pooled. Our current results identified an individual association between BANK1 rs10516487G/A and RA in Mexican patients, which is at a variance with what was published by Orozco et al. (2009). A possible explanation for this discrepancy could be the different sample sizes between both studies. Our study included 470 RA patients and 487 controls, whereas they studied 278 patients and 272 controls. In addition, our cases and controls are from Central Mexico, whereas Orozco et al. did not reveal the place of origin of their cases and controls (Orozco et al., 2009). The Mexican Republic is a large country with a great population admixture of several ethnic origins, which are not evenly distributed, e.g. Mexico City, located in central Mexico, is formed by approximately 50% Amerindian, 45% Caucasian, and 5% African ancestry (Martinez-Marignac et al., 2007). On the other hand, regions located in Western or Southeastern Mexico have a markedly different ethnic composition (Martínez-Cortés et al., 2012; Rincón et al., 2016). As far as we can tell, this is the first study showing an individual association between BANK1 rs1051648T/C and RA. Although some authors have not found association between different BANK1 or BLK alleles and risk for RA, in Spanish and Chinese patients an association of an interaction of certain BANK1-BLK genotype with RA has been reported (Génin et al., 2013; Huang et al., 2017). Moreover, is has been documented a physical and genetic interaction between BLK and BANK1 (Castillejo-Lopez et al., 2012), which would support our finding of a BLK-BANK1 genetic interaction associated to RA (Castillejo-Lopez et al., 2012; Génin et al., 2013; Huang et al., 2017). According to our results, the best model of interaction was BLK rs2736340T/C-BANK1 rs10516487G/A.
Among the four studied SNPs, it has been reported that rs13277113A/G and rs2736340T/C could have a potential eQTL effect on FAM167A and BLK genes in monocytes and lymphoblastic cell lines suggesting that these variants are involved in the physiopathology of RA and SLE (Deng et al., 2013a; Deng et al., 2013b). In addition, because both BLK variants studied here map near the promoter, they were examined in silico by means of expression quantitative trait locus (eQTL) studies (Schmiedel et al., 2018), which point out to a possible overrepresentation of T helper cells, predominantly Th17 and almost any other T helpers, and for the BLK-FAM167A locus for naïve B cells (for rs13277113A/G). Although for rs2736340T/C the eQTL associations are similar, there is a slight difference for the BLK only on that T cell overrepresentation is Th17 followed by T follicular helper cells. Thus, we can conclude that these associations clearly point to a role of an altered adaptive immunity conferring risk to develop RA. For the BANK1 variants we did not do an eQTL analysis, because both of them affect the ORF and not the regulatory regions.
A study about the functional consequence of the non-synonymous BANK1 rs10516487G/A variant (located in exon 2 with a change of arginine to histidine at amino acid position 61 [R61H] of BANK1) reported that the product of the BANK1 rs10516487A allele lacks a binding site for the SRp40 splicing enhancer protein, thereby affecting the splicing efficiency of BANK1 isoforms (Kozyrev et al., 2012). In addition, the BANK1 rs10516487GG genotype produces more BANK1 mRNA and protein than the rs10516487AA genotype. Moreover, the full-length isoform transcript BANK1-R61 generates a larger protein complex and self-association and multimerization than BANK1-H61 (Kozyrev et al., 2012). Another study showed the functional effect of the SLE-associated TGG risk haplotype which is formed by BANK1 rs17266594T/C, rs10516487G/A, and rs3733197G/A. The first variant is located in a putative branch point of intron 1, the second and third variants (R61H and A383T) are located in exon 2 and 7, respectively. Thus, the TGG risk haplotype is formed by T allele of the rs17266594T/C variant and by Arg61 and Ala383. That study showed that this risk haplotype leads to a decrease of B cell receptor signaling in Ramos B cells and in peripheral B cells. Finally, it was also described in BANK1 risk carriers that this haplotype appears to result on an increased number of memory B cells (Dam et al., 2016).
Of note, the lack of clinical and laboratory information (bone erosions, rheumatoid nodules, autoantibodies such as anti-CCPs, rheumatoid factor) as well as the absence of mRNA or protein expression studies to correlate with the four BLK and BANK1 variants studied, are some limitations of our study. Thus, at present, it is not possible to determine if these variants are associated with particular traits of RA patients.
In summary, our data indicates that both BLK as BANK1 SNVs studied herein are associated with risk to RA in Mexican patients. On the other hand, the individual association of BANK1 rs1054857G/A with RA found in the present study had not been identified in a particular population, although one study reported association between this variant and RA only in pooled patients from several countries. The main limitations in our study is the lack of clinical traits and serological markers in or RA patients, as well as the absence of ancestry informative-markers.
Data Availability Statement
The data has been uploaded to EVA repository Project: PRJEB36187 Analyses: ERZ1284135 www.ebi.ac.uk/eva/?eva-study=PRJEB36187.
Ethics Statement
This study was conducted in compliance with the Declaration of Helsinki. All individuals included in our study signed a written informed consent. This protocol was approved by the Ethics and Research Committees of the HJM (registry number 0446/18-I). The patients/participants provided their written informed consent to participate in this study.
Author Contributions
JR-B contributed to the study design, acquired the funding, data analyzed, and wrote the article. IA-Á performed the methodology and contributed with the experiments. JF and AC-P performed the data analysis. JM contributed to the data analysis and wrote the article. SJ-M contributed to the data analysis. RB-C contributed in the explanation of the project to the patients and in the collection of samples.
Conflict of Interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Acknowledgments
The authors are grateful to the patients and controls for donating their samples and to Ricardo Godínez-Aguilar for his help with the eQTL analysis.
References
Akerblad, P., Sigvardsson, M. (1999). Early B cell factor is an activator of the B lymphoid kinase promoter in early B cell development. J. Immunol. 163, 5453–5461.
Aletaha, D., Neogi, T., Silman, A. J., Funovits, J., Felson, D. T., Bingham, C. O., 3rd, et al. (2010). 2010 Rheumatoid arthritis classification criteria: an American College of Rheumatology/European League Against Rheumatism collaborative initiative. Arthritis. Rheumatol. 62, 2569–2581. doi: 10.1002/art.27584
Anderson, C. A., Pettersson, F. H., Clarke, G. M., Cardon, L. R., Morris, A. P., Zondervan, K. T. (2010). Data quality control in genetic case-control association studies. Nat. Protoc. 5, 1564–1573. doi: 10.1038/nprot.2010.116
Barrett, J. C., Fry, B., Maller, J., Daly, M. J. (2005). Haploview: analysis and visualization of LD and haplotype maps. Bioinformatics. 21, 263–265. doi: 10.1093/bioinformatics/bth457
Bossini-Castillo, L., de Kovel, C., Kallberg, H., van ‘t Slot, R., Italiaander, A., Coenen, M., et al. (2015). A genome-wide association study of rheumatoid arthritis without antibodies against citrullinated peptides. Ann. Rheumatol. Dis. 74, e15. doi: 10.1136/annrheumdis-2013-204591
Castillejo-Lopez, C., Delgado-Vega, A. M., Wojcik, J., Kozyrev, S. V., Thavathiru, E., Wu, Y. Y., et al. (2012). Genetic and physical interaction of the B-cell systemic lupus erythematosus-associated genes BANK1 and BLK. Ann. Rheumatol. Dis. 71, 136–142. doi: 10.1136/annrheumdis-2011-200085
Dam, E. M., Habib, T., Chen, J., Funk, A., Glukhova, V., Davis-Pickett, M., et al. (2016). The BANK1 SLE-risk variants are associated with alterations in peripheral B cell signaling and development in humans. Clin. Immunol. 173, 171–180. doi: 10.1016/j.clim.2016.10.018
Dang, J., Li, J., Xin, Q., Shan, S., Bian, X., Yuan, Q., et al. (2016). Gene-gene interaction of ATG5, ATG7, BLK and BANK1 in systemic lupus erythematosus. Int. J. Rheumatol. Dis. 19, 1284–1293. doi: 10.1111/1756-185X.12768
Deng, F. Y., Lei, S. F., Zhu, H., Zhang, Y. H., Zhang, Z. L. (2013a). Integrative analyses for functional mechanisms underlying associations for rheumatoid arthritis. J. Rheumatol. 40, 1063–1068. doi: 10.3899/jrheum.121119
Deng, F. Y., Lei, S. F., Zhang, Y. H., Zhang, Z. L., Guo, Y. F. (2013b). Functional relevance for associations between genetic variants and systemic lupus erythematosus. PLoS. One 8, e53037. doi: 10.1371/journal.pone.0053037
Deshmukh, H. A., Maiti, A. K., Kim-Howard, X. R., Rojas-Villarraga, A., Guthridge, J. M., Anaya, J. M., et al. (2011). Evaluation of 19 autoimmune disease-associated loci with rheumatoid arthritis in a Colombian population: evidence for replication and gene-gene interaction. J. Rheumatol. 38, 1866–1870. doi: 10.3899/jrheum.110199
Freudenberg, J., Lee, H. S., Han, B. G., Shin, H. D., Kang, Y. M., Sung, Y. K., et al. (2011). Genome-wide association study of rheumatoid arthritis in Koreans: population-specific loci as well as overlap with European susceptibility loci. Arthritis. Rheumatol. 63, 884–893. doi: 10.1002/art.30235
Génin, E., Coustet, B., Allanore, Y., Ito, I., Teruel, M., Constantin, A., et al. (2013). Epistatic interaction between BANK1 and BLK in rheumatoid arthritis: results from a large trans-ethnic meta-analysis. PLoS. One 8, e61044. doi: 10.1371/journal.pone.0061044
Gregersen, P. K., Amos, C. I., Lee, A. T., Lu, Y., Remmers, E. F., Kastner, D. L., et al. (2009). REL, encoding a member of the NF-kappaB family of transcription factors, is a newly defined risk locus for rheumatoid arthritis. Nat. Genet. 41, 820–823. doi: 10.1038/ng.395
Guthridge, J. M., Lu, R., Sun, H., Sun, C., Wiley, G. B., Dominguez, N., et al. (2014). Two functional lupus-associated BLK promoter variants control cell-type- and developmental-stage-specific transcription. Am. J. Hum. Genet. 94, 586–598. doi: 10.1016/j.ajhg.2014.03.008
Hom, G., Graham, R. R., Modrek, B., Taylor, K. E., Ortmann, W., Garnier, S., et al. (2008). Association of systemic lupus erythematosus with C8orf13-BLK and ITGAM–ITGAX. N. Engl. J. Med. 358, 900–909. doi: 10.1056/NEJMoa0707865
Huang, H., Huang, S. C., Hua, D. J., Sun, Q. Q., Cen, H., Xin, X. F. (2017). Interaction analysis between BLK rs13277113 polymorphism and BANK1 rs3733197 polymorphism, MMEL1/TNFRSF14 rs3890745 polymorphism in determining susceptibility to rheumatoid arthritis. Autoimmunity. 50, 403–408. doi: 10.1080/08916934.2017.1377191
Jiang, L., Yin, J., Ye, L., Yang, J., Hemani, G., Liu, A. J., et al. (2014). Novel risk loci for rheumatoid arthritis in Han Chinese and congruence with risk variants in Europeans. Arthritis. Rheumatol. 66, 1121–1132. doi: 10.1002/art.38353
Julià, A., González, I., Fernández-Nebro, A., Blanco, F., Rodriguez, L., González, A., et al. (2016). A genome-wide association study identifies SLC8A3 as a susceptibility locus for ACPA-positive rheumatoid arthritis. Rheumatol. (Oxford) 55, 1106–1111. doi: 10.1093/rheumatology/kew035
Kozyrev, S. V., Abelson, A. K., Wojcik, J., Zaghlool, A., Linga, Reddy, M. V., Sanchez, E., et al. (2008). Functional variants in the B-cell gene BANK1 are associated with systemic lupus erythematosus. Nat. Genet. 40, 211–216. doi: 10.1038/ng0408-484
Kozyrev, S. V., Bernal-Quirós, M., Alarcón-Riquelme, M. E., Castillejo-López, C. (2012). The dual effect of the lupus-associated polymorphism rs10516487 on BANK1 gene expression and protein localization. Genes. Immun. 13, 129–138. doi: 10.1038/gene.2011.62
Martínez-Bueno, M., Oparina, N., Dozmorov, M. G., Marion, M. ,. C., Comeau, M. E., Gilkeson, G., et al. (2018). Trans-ethnic mapping of BANK1 identifies two independent SLE-risk linkage groups enriched for Co-transcriptional Splicing Marks. Int. J. Mol. Sci. 19, E2331. doi: 10.3390/ijms19082331
Martínez-Cortés, G., Salazar-Flores, J., Fernández-Rodríguez, L. G., Rubi-Castellanos, R., Rodríguez-Loya, C., Velarde-Félix, J. S., et al. (2012). Admixture and population structure in Mexican-Mestizos based on paternal lineages. J. Hum. Genet. 57, 568–574. doi: 10.1038/jhg.2012.67
Martinez-Marignac, V. L., Valladares, A., Cameron, E., Chan, A., Perera, A., Globus-Goldberg, R., et al. (2007). Admixture in Mexico City: implications for admixture mapping of type 2 diabetes genetic risk factors. Hum. Genet. 120, 807–819. doi: 10.1007/s00439-006-0273-3
Moore, J. H., Gilbert, J. C., Tsai, C. T., Chiang, F. T., Holden, T., Barney, N., et al. (2006). A flexible computational framework for detecting, characterizing, and interpreting statistical patterns of epistasis in genetic studies of human disease susceptibility. J. Theor. Biol. 241, 252–261. doi: 10.1016/j.jtbi.2005.11.036
Muñoz-Valle, J. F., Padilla-Gutiérrez, J. R., Hernández-Bello, J., Ruiz-Noa, Y., Valle, Y., Palafox-Sánchez, C. A., et al. (2017). PTPN22 -1123G > C polymorphism and anti-cyclic citrullinated protein antibodies in rheumatoid arthritis. Med. Clin. (Barc). 149, 95–100. doi: 10.1016/j.medcli.2017.01.025
Orozco, G., Abelson, A. K., González-Gay, M. A., Balsa, A., Pascual-Salcedo, D., García, A., et al. (2009). Study of functional variants of the BANK1 gene in rheumatoid arthritis. Arthritis. Rheumatol. 60, 372–379. doi: 10.1002/art.24244
Orozco, G., Eyre, S., Hinks, A., Bowes, J., Morgan, A. W., Wilson, A. G., et al. (2011). Study of the common genetic background for rheumatoid arthritis and systemic lupus erythematosus. Ann. Rheumatol. Dis. 70, 463–468. doi: 10.1136/ard.2010.137174
Orozco, G., Viatte, S., Bowes, J., Martin, P., Wilson, A. G., Morgan, A. W., et al. (2014). Novel rheumatoid arthritis susceptibility locus at 22q12 identified in an extended UK genome-wide association study. Arthritis. Rheumatol. 66, 24–30. doi: 10.1002/art.38196
Peláez-Ballestas, I., Sanin, L. H., Moreno-Montoya, J., Alvarez-Nemegyei, J., Burgos-Vargas, R., Garza-Elizondo, M., et al. (2011). Epidemiology of the rheumatic diseases in Mexico. A study of 5 regions based on the COPCORD methodology. J. Rheumatol. Suppl. 86, 3–8. doi: 10.3899/jrheum.100951
Rincón, J. F., Cano, D. L., Morales, S. J., Jiménez, M. L., Cobos, R. E., Bello, J. R. (2016). The functional PTPN22 C1858T polymorphism confers risk for rheumatoid arthritis in patients from Central Mexico. Clin. Rheumatol. 35, 1457–1462. doi: 10.1007/s10067-016-3223-z
Rodríguez-Elías, A. K., Maldonado-Murillo, K., López-Mendoza, L. F., Ramírez-Bello, J. (2016). Genetics and genomics in rheumatoid arthritis (RA): An update. Gac. Med. Mex. 152, 218–227.
Sánchez, E., Comeau, M. E., Freedman, B. I., Kelly, J. A., Kaufman, K. M., Langefeld, C. D., et al. (2011). Identification of novel genetic susceptibility loci in African American lupus patients in a candidate gene association study. Arthritis. Rheumatol. 63, 3493–3501. doi: 10.1002/art.30563
Saxena, R., Plenge, R. M., Bjonnes, A. C., Dashti, H. S., Okada, Y., Gad, E., et al. (2017). A Multinational Arab genome-wide association study identifies new genetic associations for rheumatoid arthritis. Arthritis. Rheumatol. 69, 976–985. doi: 10.1002/art.40051
Schmiedel, B. J., Singh, D., Madrigal, A., Valdovino-Gonzalez, A. G., White, B. M., Zapardiel-Gonzalo, J., et al. (2018). Impact of genetic polymorphisms on human immune cell gene expression. Cell. 175, 1701–1715.e16. doi: 10.1016/j.cell.2018.10.022
Scott, D. L., Wolfe, F., Huizinga, T. W. (2010). Rheumatoid arthritis. Lancet. 376, 1094–1108. doi: 10.1016/S0140-6736(10)60826-4
Simpfendorfer, K. R., Olsson, L. M., Manjarrez-Orduño, N., Khalili, H., Simeone, A. M., Katz, M. S., et al. (2012). The autoimmunity-associated BLK haplotype exhibits cis-regulatory effects on mRNA and protein expression that are prominently observed in B cells early in development. Hum. Mol. Genet. 21, 3918–3925. doi: 10.1093/hmg/dds220
Suarez-Gestal, M., Calaza, M., Dieguez-Gonzalez, R., Perez-Pampin, E., Pablos, J. L., Navarro, F., et al. (2009). Rheumatoid arthritis does not share most of the newly identified systemic lupus erythematosus genetic factors. Arthritis. Rheumatol. 60, 2558–2564. doi: 10.1002/art.24748
Suthers, A. N., Sarantopoulos, S. (2017). TLR7/TLR9- and B Cell receptor-signaling crosstalk: promotion of potentially dangerous B cells. Front. Immunol. 8, 775. doi: 10.3389/fimmu.2017.00775
Tretter, T., Ross, A. E., Dordai, D. I., Desiderio, S. (2003). Mimicry of pre-B cell receptor signaling by activation of the tyrosine kinase Blk. J. Exp. Med. 198, 1863–1873. doi: 10.1084/jem.20030729
Viatte, S., Plant, D., Bowes, J., Lunt, M., Eyre, S., Barton, A., et al. (2012). Genetic markers of rheumatoid arthritis susceptibility in anti-citrullinated peptide antibody negative patients. Ann. Rheumatol. Dis. 71, 1984–1990. doi: 10.1136/annrheumdis-2011-201225
Zaragoza-García, O., Navarro-Zarza, J. E., Maldonado-Anicacio, J. Y., Castro-Alarcón, N., Rojas, I. P., Guzmán-Guzmán, I. P. (2019). Hypertriglyceridaemic waist is associated with hyperuricaemia and metabolic syndrome in rheumatoid arthritis patients. Diabetes Metab. Syndr. 13 (1), 722–729. doi: 10.1016/j.dsx.2018.11.053
Keywords: rheumatoid arthritis, single nucleotide variants, gene interaction, susceptibility, association
Citation: Ramírez-Bello J, Fragoso JM, Alemán-Ávila I, Jiménez-Morales S, Campos-Parra AD, Barbosa-Cobos RE and Moreno J (2020) Association of BLK and BANK1 Polymorphisms and Interactions With Rheumatoid Arthritis in a Latin-American Population. Front. Genet. 11:58. doi: 10.3389/fgene.2020.00058
Received: 21 May 2019; Accepted: 17 January 2020;
Published: 20 February 2020.
Edited by:
Taru Tukiainen, Institute for Molecular Medicine Finland (FIMM), FinlandReviewed by:
Guillaume Lettre, Université de Montréal, CanadaArmand Valsesia, Nestle Institute of Health Sciences (NIHS), Switzerland
Copyright © 2020 Ramírez-Bello, Fragoso, Alemán-Ávila, Jiménez-Morales, Campos-Parra, Barbosa-Cobos and Moreno. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Julian Ramírez-Bello, ZHIuanVsaWFuLnJhbWlyZXouaGptQGdtYWlsLmNvbQ==">dr.julian.ramirez.hjm@gmail.com