 Sandra Rosas-Madrigal1
Sandra Rosas-Madrigal1 María Teresa Villarreal-Molina2
María Teresa Villarreal-Molina2 José Flores-Rivera3
José Flores-Rivera3 Verónica Rivas-Alonso3
Verónica Rivas-Alonso3 Luis Rodrigo Macias-Kauffer4
Luis Rodrigo Macias-Kauffer4 Graciela Ordoñez5María del Carmen Chima-Galán6
Graciela Ordoñez5María del Carmen Chima-Galán6 Víctor Acuña-Alonzo7Gastón Macín-Pérez7Rodrigo Barquera8
Víctor Acuña-Alonzo7Gastón Macín-Pérez7Rodrigo Barquera8 Julio Granados9Ricardo Valle-Rios10
Julio Granados9Ricardo Valle-Rios10 Teresa Corona3Alessandra Carnevale1
Teresa Corona3Alessandra Carnevale1 Sandra Romero-Hidalgo11*
Sandra Romero-Hidalgo11*- 1Laboratorio de Enfermedades Mendelianas, Instituto Nacional de Medicina Genómica, Mexico City, Mexico
- 2Laboratorio de Genómica de Enfermedades Cardiovasculares, Instituto Nacional de Medicina Genómica, Mexico City, Mexico
- 3Laboratorio Clínico de Enfermedades Neurodegenerativas, Instituto Nacional de Neurología y Neurocirugía “Manuel Velasco Suarez” (INNN), Mexico City, Mexico
- 4Unidad de Genómica de Poblaciones Aplicada a La Salud, Facultad de Química, UNAM/INMEGEN, Mexico City, Mexico
- 5Laboratorio de Neuroinmunología, INNN, Mexico City, Mexico
- 6División de Medicina Genómica, Centro Médico Nacional “20 de Noviembre”, ISSSTE, Mexico City, Mexico
- 7Escuela Nacional de Antropología e Historia, Mexico City, Mexico
- 8Department of Archaeogenetics, Max Planck Institute for the Science of Human History, Jena, Germany
- 9Departamento de Trasplantes, Instituto Nacional de Ciencias Medicas y Nutrición “Salvador Zubirán”, Mexico City, Mexico
- 10División de Investigación, Facultad de Medicina, Unidad de Investigación en Inmunología y Proteómica, Hospital Infantil de México Federico Gómez, Universidad Nacional Autónoma de México, Mexico City, Mexico
- 11Departamento de Genómica Computacional, Instituto Nacional de Medicina Genómica (INMEGEN), Mexico City, Mexico
Neuromyelitis Optica Spectrum Disorder (NMOSD) is a demyelinating autoimmune disease of the central nervous system, more prevalent in individuals of non-European ancestry. Few studies have analyzed genetic risk factors in NMOSD, and HLA class II gene variation has been associated NMOSD risk in various populations including Mexicans. Thymopoietin (TMPO) has not been tested as a candidate gene for NMOSD or other autoimmune disease, however, experimental evidence suggests this gene may be involved in negative selection of autoreactive T cells and autoimmunity. We thus investigated whether the missense TMPO variant rs17028450 (Arg630Cys, frequent in Latin America) is associated with NMOSD, and whether this variant shows an interaction with HLA-class II rs9272219, previously associated with NMOSD risk. A total of 119 Mexican NMOSD patients, 1208 controls and 357 Native Mexican individuals were included. The HLA rs9272219 “T” risk allele frequency ranged from 21 to 68%, while the rs17028450 “T” minor allele frequency was as high as 18% in Native Mexican groups. Both rs9272219 and rs17028450 were significantly associated with NMOSD risk under additive models (OR = 2.48; p = 8 × 10–10 and OR = 1.59; p = 0.0075, respectively), and a significant interaction between both variants was identified with logistic regression models (p = 0.048). Individuals bearing both risk alleles had an estimated 3.9-fold increased risk of NMOSD. To our knowledge, this is the first study reporting an association of TMPO gene variation with an autoimmune disorder and the interaction of specific susceptibility gene variants, that may contribute to the genetic architecture of NMOSD in admixed Latin American populations.
Introduction
As other autoimmune diseases, Neuromyelitis Optica Spectrum Disorder (NMOSD) is a multifactorial disorder that results from complex interactions between genetic and environmental factors. Its worldwide prevalence has been estimated to range between ∼0.5 and 4/100,000 and may be up to 10/100,000 depending on geographical location and ethnicity. It has been consistently suggested that NMOSD is more frequent in non-European populations. East Asians and populations with African ancestry have reported a higher prevalence of this disease (Hor et al., 2020).
In Latin America, admixed populations with lower proportions of European ancestry have higher relative frequencies of NMOSD (Alvarenga et al., 2017). We recently proposed that Native American ancestry contributes to NMOSD susceptibility, as Mexican NMOSD patients showed significantly higher proportions of Native American ancestry than controls. In addition, we performed a genome-wide association study (GWAS) identifying a NMOSD risk genetic variant within HLA class II region (rs9272219) that is in linkage disequilibrium with HLA DRB1∗16:02, previously associated with NMOSD in Southern Han Chinese, Japanese and Brazilian populations (Wang et al., 2011; Yoshimura et al., 2013; Kay et al., 2019; Romero-Hidalgo et al., 2020). Other HLA class II risk alleles have been associated with NMOSD in different ethnic groups. A limited number of candidate gene association studies have also reported associations of NMOSD with individual polymorphisms in a few genes involved in immune function, including PD-1, IL-17, IL-7R, CD226, and CD58 (Asgari et al., 2012; Liu et al., 2012; Wang et al., 2012; Kim et al., 2014; Zhuang et al., 2015).
Thymopoietin (TMPO) has not been previously explored as a candidate gene for NMSOD or other autoimmune disease. TMPO is located on chromosome 12q22, is highly expressed in immune cells (BioGP, 2020) and plays an important role in T cell differentiation (Basch and Goldstein, 1974; Silva et al., 2009). It encodes thymopoietin or LAP2 known to be involved in nuclear architecture and chromatin organization by interacting with structural nuclear proteins such as lamins (Lunin et al., 2020). LAP2 binds to lamin B (Foisner and Gerace, 1993) and regulates the correct distribution of lamin B in the nucleus through its carboxy-terminal domain (Brady et al., 2018). On the other hand, lamin B plays a role in maintaining cortical and medullar compartments of the thymus by inducing gene expression in Thymic Epithelial Cells (TECs) (Yue et al., 2019). Importantly, altered lamin B function may induce the generation of ectopic transcripts encoding Peripheral-Tissue Antigens (PTAs) by influencing the Autoimmune Regulator (AIRE), leading to autoimmunity (Abramson et al., 2010). Thus, genetic variants affecting interaction of LAP2 and lamin B may indirectly affect the normal function of AIRE in the thymus.
Rs17028450 is a missense polymorphism (Arg690Cys) in the TMPO gene, and the 690 Cys protein was found to decrease the interaction LAP2 and lamin A proteins in vitro (Taylor et al., 2005). Interestingly, the derived allele is most frequent in Latin American populations (14%), and less common in Asians, Africans and Europeans (<2%). Thus, we aimed to evaluate whether the TMPO rs17028450 missense variant is associated with NMOSD, and whether this variant interacts with HLA class II variation affecting NMOSD susceptibility in the admixed Mexican population.
Materials and Methods
Study Population
Microarray data of 119 previously described admixed Mexican NMOSD patients (79% female) and 1,208 controls (61% female) (Romero-Hidalgo et al., 2020) were used for the analyses. In addition, TMPO rs17028450 genotypes were determined using Taqman probes in 85 systemic lupus erythematosus (SLE) patients recruited from the Instituto Nacional de Ciencias Médicas y Nutrición “Salvador Zubirán,” and 104 multiple sclerosis (MS) patients recruited from the DNA bank of the Genomic Medicine Division at the Centro Médico Nacional 20 de Noviembre del Instituto de Seguridad y Servicios Sociales de los Trabajadores del Estado (ISSSTE). SLE patients were diagnosed according to the 1982 SLE classification criteria (Watanabe et al., 1985) and MS was diagnosed according to McDonald criteria (Sadaka et al., 2012). In addition, TMPO genotypes from 3 different Native Mexican groups were analyzed: 53 previously described Nahuas and 19 Totonacs (Romero-Hidalgo et al., 2017) using Taqman probes, and 82 Mayans with type 2 diabetes recruited from rural health centers in the State of Yucatan with available exome data. Finally, allele frequencies of HLA rs9272219 were evaluated in 138 Nahuas, 24 Totonacs, 45 Zapotecs and 68 Mayas using available microarray data (Romero-Hidalgo et al., 2017). Protocols for each cohort were approved by their respective Institutional Ethics and Research Committees (No. 66/14, No. DIC/491/14).
Genotyping
Genomic DNA was extracted from blood samples with the QIAamp DNA Blood Midi/Maxi kit (Quiagen) according to the supplier’s instructions. DNA integrity, purity and concentrations were determined by NanoDrop® spectrophotometry (NanoDrop One/Onec Thermo scientific). The TMPO rs17028450 single nucleotide polymorphism (SNP) was genotyped by Taqman® assays (Life Technologies Company) using a viiaTM seven real-time PCR instrument. Genotype were assigned automatically by measuring allele specific fluorescence using TaqMan® GenotyperTM Software (Applied Biosystems). PCR mix included 15 ng of genomic DNA, 0.45 μM of oligonucleotide (VIC/FAM AGAAGTATGCAAAGTAATTAAAAAG[C/T]GTGGAAATAA ACACTAGTAAAATTA), 2.5 μl of TaqMan master mix (Applied Biosystems, Foster city, CA, United States) and ddH2O for a final volume of 5 μl per reaction. The amplification protocol was as follows: one cycle of 10 min at 95°C (denaturing), followed by 40 cycles of 15 s at 95°C (denaturing), and 1 min at 60°C (annealing) and 1 min at 72°C (extension). TMPO rs17028450 genotypes from a subset of 19 NMOSD samples were validated by Sanger sequencing according to the standard protocol used at the High Technology Core of National Institute of Genomic Medicine (HTC_INMEGEN). Primer sequences were: Forward 5′-tcagcagttggacttagcactc and Reverse 5′-tgtcctaggtataaaggaggatgc. No discordant genotypes were found.
Statistical Analyses
Logistic regression models were used to test associations of NMOSD with single SNPs under additive and dominant inheritance models, and to test gene-gene interactions adding a multiplicative interaction term in the corresponding logistic regression model. Finally, a genetic risk score (GRS) was tested by summing the “T” risk alleles of HLA rs9272219 and TMPO rs17028450 variants (0, 1, 2, 3, or 4 risk alleles). A weighted GRS was also calculated as the sum of the effect estimates from a logistic regression analysis with an additive inheritance model, multiplied by the number of risk alleles (Igo et al., 2019). All logistic regression models were adjusted for sex and two principal components as covariates. The principal component was obtained from Romero-Hidalgo et al. (2020). Because age was not available for all NMOSD cases, it was not included as covariate in the analyses. An independent t-test was used to compare mean GRS values between cases and controls. Statistical analyses were performed using R software environment (R-Software, 2021). The Geography of Genetic Variants (GGV) were used to map the geographic distribution of the genetic variants (Marcus and November, 2017).
Results
Figure 1 shows the worldwide derived allele frequencies of HLA class II rs9272219 and TMPO rs17028450 based on populations included in 1,000 genomes project. Clearly, TMPO rs17028450 is an ancestry-specific polymorphism almost private to the Americas. The frequency of the derived “T” allele is lower than 2% in Asian, African and European populations, and 10.9 and 14.1% in Mexican American and Peruvian populations, respectively. In contrast, the HLA rs9272219 “T” allele is frequent in all populations, ranging from 13.4% in South and East Asian populations to 47% in Sierra Leone and Mexican Americans. In Native Mexican populations, the HLA “T” risk allele ranged from 20.6% in Mayans to 67.8% in the Zapotec population, whereas, the TMPO derived allele showed a more homogenous allele frequency among Native Mexican populations (11–18%).
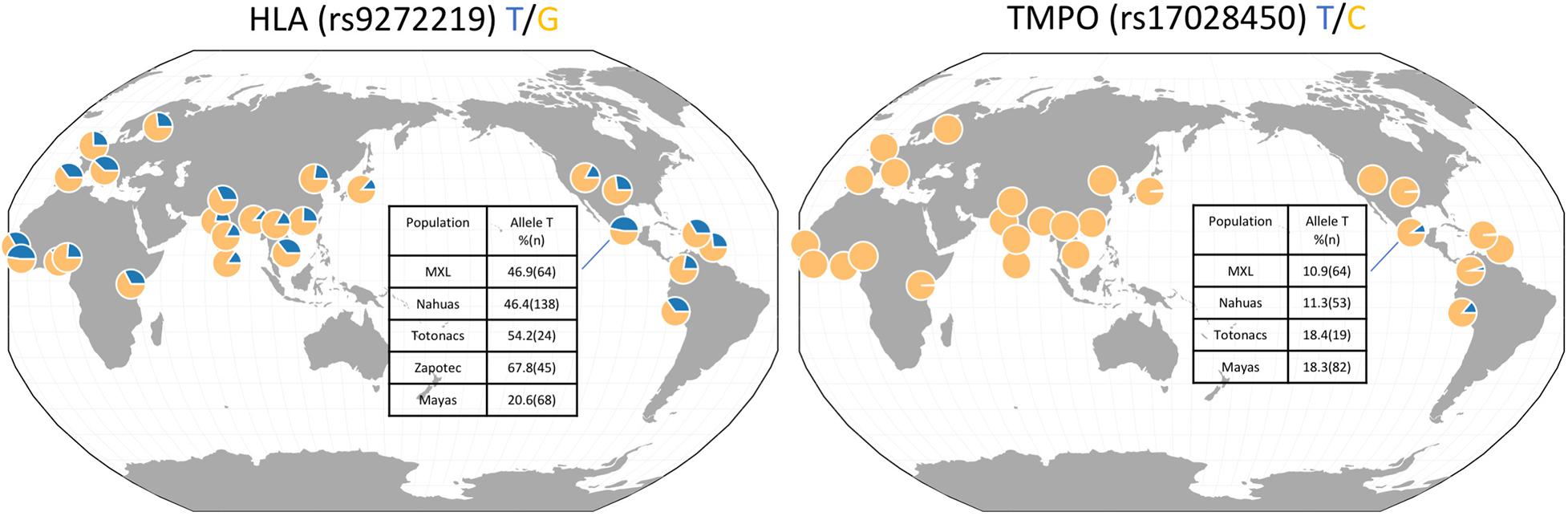
Figure 1. HLA class II rs9272219 and TMPO rs17028450 allele frequencies in worldwide populations and in Native Mexican groups.
For the case-control association study, a total of 119 NMOSD patients (79% female) and 1,208 apparently healthy controls (61% female) were included in the analysis. As shown in Table 1, HLA rs9272219 and TMPO rs17028450 risk allele frequencies were higher in NMOSD patients (67.2 and 23.5%, respectively) as compared to controls (41 and 13.5%, respectively). Additive and dominant inheritance models were evaluated for both genetic variants and all showed significant associations in the admixed Mexican population. The HLA “T” allele showed significant association with NMOSD risk (OR = 2.48; p = 8 × 10–10 and OR = 2.95; p = 0.0001, additive and dominant models, respectively). The TMPO “T” allele was also significantly associated with NMOSD risk (OR = 1.59; p = 0.0075 and OR = 1.88; p = 0.002, additive and dominant models, respectively).

Table 1. Association of HLA rs9272219 and TMPO rs17028450 with NMOSD in the Mexican Population.
Because TMPO rs17028450 was found to be associated with NMOSD, we explored the allele frequencies of this variant in patients with other autoimmune diseases. The “T” allele frequency was 12.2% in a group of 104 MS patients, and 15.8% in a group of 85 SLE patients, however allele frequencies were not significantly different as compared to controls (13.5%).
In order to assess a possible HLA-TMPO gene interaction we compared HLA risk variant frequencies in NMOSD cases and controls, stratified by the absence or presence of the TMPO risk allele (“CC” vs. “CT/TT” genotypes) (Figure 2A). Interestingly, the distribution of cases and controls across the three HLA genotypes (GG, GT, TT) differed according to the presence or absence of the TMPO risk allele. We observed a case-control ratio of 2.7 (0.48/0.18) in individuals with HLA “TT” genotype without considering TMPO genotypes. However, when stratified by the absence or presence of the TMPO risk allele, case control ratios in HLA “TT” genotypes were 1.8 (0.25/0.14) in the absence, and 4.6 (0.23/0.05) in the presence of the TMPO “T” risk allele. Similarly, the case-control ratio in individuals bearing the TMPO “T” risk allele is 1.8 (0.44/0.25). However, when stratified by the absence or presence of the HLA “T” risk allele case-control ratios were 0.4 (0.3/0.8) and 2.41 (0.41/0.17), respectively. According to the logistic regression model, the interaction was significant when both SNPs are considered dominant (p = 0.048) and only show a tendency for the HLA (additive)-TMPO (dominant) scenario (p = 0.148; Table 2).
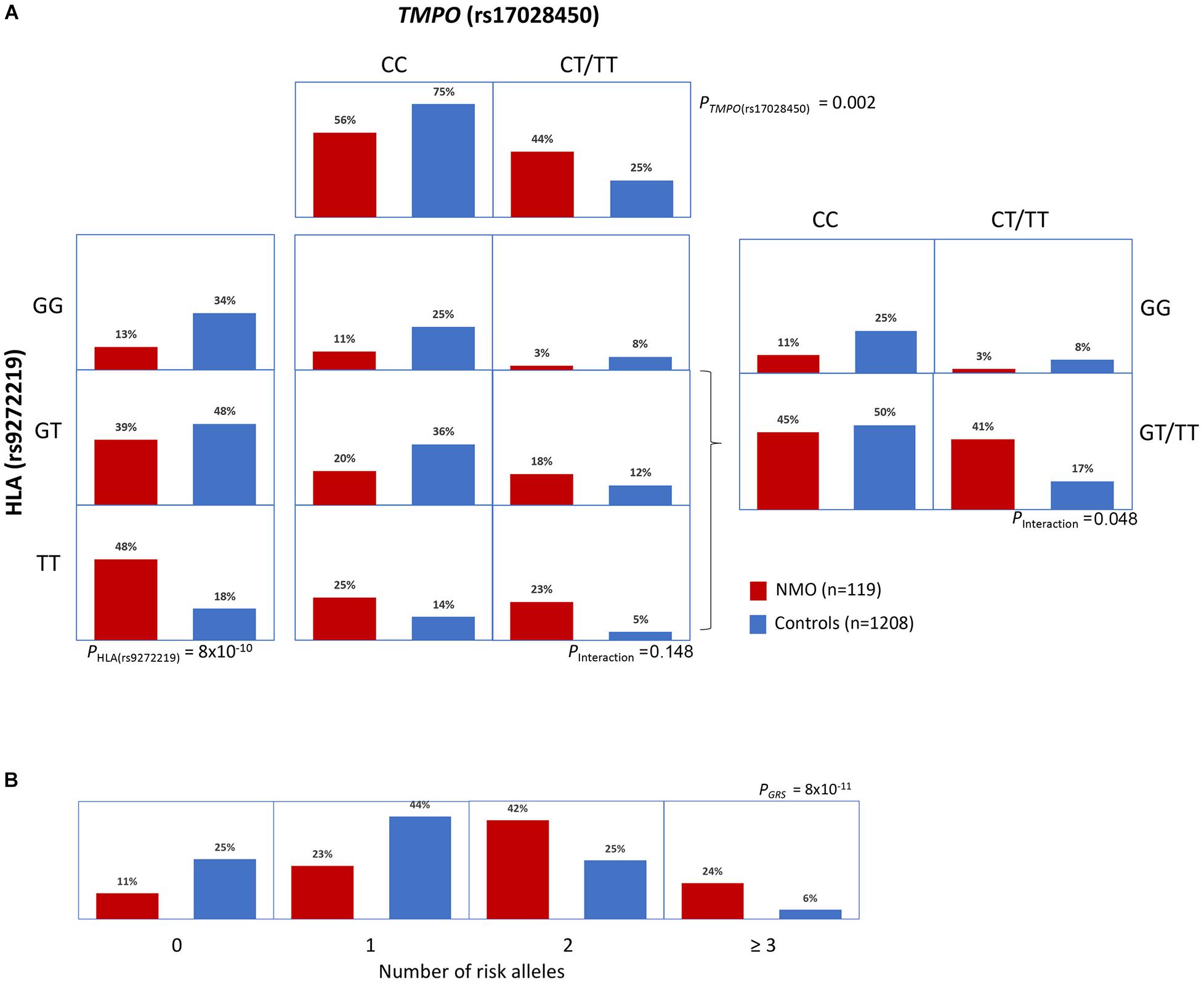
Figure 2. Interaction and genetic risk score analyses. (A) Distribution of NMOSD cases and controls across genotypes for HLA rs9272219 and TMPO rs17028450 genetic variants. The TMPO variant was analyzed assuming a dominant inheritance model mainly because of the reduced number of TMPO “TT” homozygous individuals. (B) Distribution of cumulative risk alleles (HLA rs9272219 and TMPO rs17028450) in cases and controls.
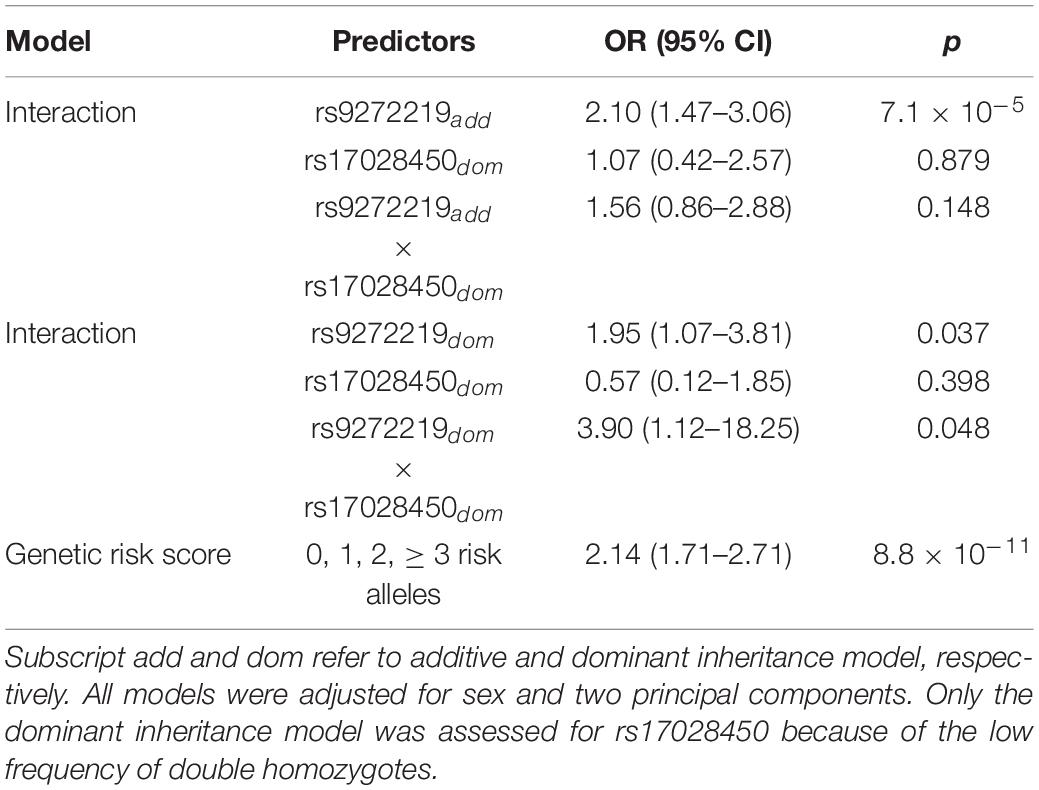
Table 2. HLA rs9272219 and TMPO rs17028450 in NMOSD, interactions and genetic risk score analyses.
We used a GRS to analyze the cumulative contribution of HLA and TMPO variants (Figure 2B). The mean GRS was significantly higher in cases (1.82 ± 0.97) than in controls (1.12 ± 0.86; p = 4.6 × 10–12) and showed a strong and significant association with NMOSD risk, for both the unweighted (OR = 2.14; p = 8.8 × 10–11) and weighted models (OR = 2.78; p = 2.5 × 10–11).
Discussion
As other autoimmune diseases, NMOSD is a multifactorial disorder that results from complex interactions between genetic and environmental factors. First considered a variation of MS, it was identified as a distinct autoimmune entity after the discovery of positive antiaquaporin-4 antibodies in 2004. Although significant progress has been made in the clinical and epidemiological characterization of NMOSD, genetic and environmental risk factors involved in this disorder are still unclear, in part because of its low worldwide prevalence (Lennon et al., 2005; Paul et al., 2020).
Only four genome-wide association studies have been conducted in NMOSD, including a small number of cases per study (∼200). In spite of their low statistical power, three independent GWAS identified SNPs within the MHC region associated with NMOSD in individuals of European ancestry (Estrada et al., 2018), in Admixed Mexicans (Romero-Hidalgo et al., 2020) and in the Japanese population (Matsushita et al., 2020); while a GWAS in Koreans did not identify any variants associated with NMOSD susceptibility with genome-wide significance (Kim et al., 2010). The GWAS in the Mexican and Japanese populations reported similar results, as both identified a genome-wide significant association with rs1964995, although the lead SNP in the Mexican population was rs9272219 (Matsushita et al., 2020; Romero-Hidalgo et al., 2020). Moreover, both studies reported HLA DRB1∗16:02 and DRB1∗08:02 as NMOSD susceptibility alleles. It is noteworthy that rs9272219 risk allele frequencies in 3 of the Native Mexican populations (Nahuas, Totonacs, and Zapotecs) are among the highest reported worldwide. Interestingly, rs9272219 and rs1964995 have been associated with other autoimmune diseases such as rheumatoid arthritis and dermatomyositis in different populations (Eleftherohorinou et al., 2011; Hao et al., 2014).
Few non-HLA candidate gene studies have been conducted in NMOSD (Asgari et al., 2012; Liu et al., 2012; Wang et al., 2012; Kim et al., 2014; Zhuang et al., 2015), and as far as we know, TMPO has not been previously identified as a susceptibility gene in GWAS, nor has it has been studied as a candidate gene for any autoimmune disorder. Although experimental evidence on the role of TMPO and LAP2 in autoimmunity is limited, it has been demonstrated that LAP2 binds to lamin B1 in vitro (Foisner and Gerace, 1993). Furthermore, the carboxi-terminal domain of LAP2 is required to maintain the correct nuclear distribution of lamins A, B1 and B2 in cancer cells (Brady et al., 2018) suggesting that TMPO mutations may destabilize lamin distribution in certain cell types. This might be relevant since lamin B1 overexpression reduces the expression of the autoimmune regulator (AIRE), which in turn could inhibit the expression of a battery of peripheral-tissue self-antigens (PTAs), an essential for immunological tolerance by promoting the development of autoreactive T cells in thymic stromal cells (Abramson et al., 2010).
Notably, rs17028450 was first identified in a patient with dilated cardiomyopathy (DCM), and the LAP2 630Cys form of the protein was found to decrease the interaction of LAP2 with the lamin A terminus in vitro (Taylor et al., 2005). However, it is unknown whether this amino acid substitution affects LAP2 interaction with lamin B1, has other functional consequences in other specific cell types, or is involved in susceptibility to autoimmunity. The rs17028450 minor “T” allele is frequent in Latin Americans but rare in other continental populations (<2%), with frequencies as high as 18% in Native Mexican groups including Nahuas, Totonacs, and Mayans. To our knowledge, this is the first report associating TMPO gene variation with an autoimmune disease (NMOSD) using a candidate gene approach, under both additive (OR = 1.59, p = 0.0075) and dominant (OR = 1.88, p = 0.002) models.
Interaction effects are difficult to assess because the number of multilocus genotype combinations increases exponentially as additional SNPs are considered, and larger sample sizes are needed to estimate the corresponding effects. However, it is important to seek interactions since the effect of a genetic variant can be masked by the effect of other variants (VanderWeele, 2010). Because of this difficulty, and particularly for diseases of low prevalence, interactions have been sought using the candidate gene approach. Using this approach, we observed an interaction between HLA rs9272219 and TMPO rs17028450 with an estimated odds ratio of 3.9 for individuals bearing both risk alleles. To our knowledge, this is the first study reporting gene interactions affecting NMOSD risk. Epistatic interactions between HLA and non-HLA genetic variants have been previously reported for other autoimmune diseases including SLE, MS, rheumatoid arthritis, ankylosing spondylitis and psoriasis (Matzaraki et al., 2017; Diaz-Gallo et al., 2018; Hanson et al., 2020). In addition, a significant interaction between r9273363 (HLA DQB1) and SNPs rs9272219 (HLA DQA1) was reported in RA and type 1 diabetes, and this HLA region was identified as epigenetically active in B cells (Woo et al., 2017). Since Lamin B1 and LAP2 are expressed in the thymus, the LAP2 690Cys variant may indirectly affect AIRE function in medullary TECs, perhaps altering the presentation of AQP4 or other antigens in the thymus, allowing autoreactive T cells that recognize these antigens to escape negative selection. If this occurs in epigenetically active B cells expressing the HLA DQA1 rs9272219 “T” allele, this may favor the production of anti-AQP4 or other autoantibodies.
Some limitations of the study must be pointed out. Firstly, the association between TMPO rs17028450 with NMOSD requires confirmation in other populations, particularly in Latin Americans because of the higher frequency of this variant and the relatively higher prevalence of NMOSD. Secondly, although rs17028450 was not associated with MS or SLE (p > 0.26), it should be tested in larger cohorts of MS and SLE cases, and in other autoimmune diseases. Finally, further association and functional studies are required to confirm the interaction observed between HLA rs9272219 and TMPO rs17028450 and to elucidate how these variants may confer NMOSD susceptibility in combination. Although the sample size is small, our data support a model whereby the contribution of co-inherited risk alleles changes the individual’s predisposition to NMOSD, as probably occurs in many immune cell-mediated diseases.
Data Availability Statement
The raw data for the NMO patients analyzed in the current study are available from the corresponding author on reasonable request. Access to the control dataset used in this manuscript was obtained through a formal request to the Consortium for the Analysis of the Diversity and Evolution of Latin America steering committee.
Ethics Statement
The studies involving human participants were reviewed and approved by protocols for each cohort were approved by their respective Institutional Ethics Committees: Instituto Nacional de Medicina Gen mica, Instituto Nacional de Neurología y Neurocirugía “Manuel Velasco Suarez”, Instituto Nacional de Ciencias Medicas y Nutrición “Salvador Zubirán” and Centro Médico Nacional 20 de Noviembre del Instituto de Seguridad y Servicios Sociales de los Trabajadores del Estado. The patients/participants provided their written informed consent to participate in this study.
Author Contributions
SR-M performed the genotyping. SR-M, AC, and SR-H conceived and designed study. SR-H and LM-K performed data analysis. TC, JF-R, and VR-A were responsible for recruiting NMOSD patients. GO and MC-G were responsible for providing multiple sclerosis patients samples. JG was responsible for providing systemic lupus erythematosus patients samples. VA-A, GM-P, and RB were responsible for providing Native Mexicans samples. RV-R contributed to analysis and interpretation of results. SR-M, MV-M, and SR-H wrote the manuscript. All authors contributed to the article and approved the submitted version.
Funding
This work was supported by Consejo Nacional de Ciencia y Tecnología (CONACyT) (grant no. SALUD-2013-01-201206 to SR-H).
Conflict of Interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Acknowledgments
We thank members of CANDELA Consortium Steering Committee (Maria-Cátira Bortolini, Carla Gallo, Gabriel Bedoya, Francisco Rothhammer, Rolando González-Jose, Samuel Canizales-Quinteros, and Andrés Ruiz-Linares) for sharing genomic data.
References
Abramson, J., Giraud, M., Benoist, C., and Mathis, D. (2010). Aire’s partners in the molecular control of immunological tolerance. Cell 140, 123–135. doi: 10.1016/j.cell.2009.12.030
Alvarenga, M. P., Schimidt, S., and Alvarenga, R. P. (2017). Epidemiology of neuromyelitis optica in Latin America. Mult. Scler. J. Exp. Transl. Clin. 3:2055217317730098. doi: 10.1177/2055217317730098
Asgari, N., Nielsen, C., Stenager, E., Kyvik, K. O., and Lillevang, S. T. (2012). HLA, PTPN22 and PD-1 associations as markers of autoimmunity in neuromyelitis optica. Mult. Scler. 18, 23–30. doi: 10.1177/1352458511417480
Basch, R. S., and Goldstein, G. (1974). Induction of T-cell differentiation in vitro by thymin, a purified polypeptide hormone of the thymus. Proc. Natl. Acad. Sci. U. S. A. 71, 1474–1478. doi: 10.1073/pnas.71.4.1474
BioGP (2020). BioGPS [Online]. Available online at: http://biogps.org/#goto=genereport&id=7112 (accessed December 10, 2020)
Brady, G. F., Kwan, R., Bragazzi Cunha, J., Elenbaas, J. S., and Omary, M. B. (2018). Lamins and lamin-associated proteins in gastrointestinal health and disease. Gastroenterology 154, 1602–1619.e1. doi: 10.1053/j.gastro.2018.03.026
Diaz-Gallo, L. M., Ramskold, D., Shchetynsky, K., Folkersen, L., Chemin, K., Brynedal, B., et al. (2018). Systematic approach demonstrates enrichment of multiple interactions between non-HLA risk variants and HLA-DRB1 risk alleles in rheumatoid arthritis. Ann. Rheum. Dis. 77, 1454–1462. doi: 10.1136/annrheumdis-2018-213412
Eleftherohorinou, H., Hoggart, C. J., Wright, V. J., Levin, M., and Coin, L. J. (2011). Pathway-driven gene stability selection of two rheumatoid arthritis GWAS identifies and validates new susceptibility genes in receptor mediated signalling pathways. Hum. Mol. Genet. 20, 3494–3506. doi: 10.1093/hmg/ddr248
Estrada, K., Whelan, C. W., Zhao, F., Bronson, P., Handsaker, R. E., Sun, C., et al. (2018). A whole-genome sequence study identifies genetic risk factors for neuromyelitis optica. Nat. Commun. 9:1929. doi: 10.1038/s41467-018-04332-3
Foisner, R., and Gerace, L. (1993). Integral membrane proteins of the nuclear envelope interact with lamins and chromosomes, and binding is modulated by mitotic phosphorylation. Cell 73, 1267–1279. doi: 10.1016/0092-8674(93)90355-t
Hanson, A. L., International Genetics of Ankylosing Spondylitis Consortium, Vukcevic, D., Leslie, S., Harris, J., Le Cao, K. A., et al. (2020). Epistatic interactions between killer immunoglobulin-like receptors and human leukocyte antigen ligands are associated with ankylosing spondylitis. PLoS Genet. 16:e1008906. doi: 10.1371/journal.pgen.1008906
Hao, G. F., Li, Y. S., Liu, J. L., and Wo, M. Y. (2014). Association of HLA-DQA1 (rs9272219) with susceptibility to rheumatoid arthritis in a Han Chinese population. Int. J. Clin. Exp. Pathol. 7, 8155–8158.
Hor, J. Y., Asgari, N., Nakashima, I., Broadley, S. A., Leite, M. I., Kissani, N., et al. (2020). Epidemiology of neuromyelitis optica spectrum disorder and its prevalence and incidence worldwide. Front. Neurol. 11:501. doi: 10.3389/fneur.2020.00501
Igo, R. P. Jr., Kinzy, T. G., and Cooke Bailey, J. N. (2019). Genetic risk scores. Curr. Protoc. Hum. Genet. 104:e95. doi: 10.1002/cphg.95
Kay, C. S. K., Scola, R. H., Arndt, R. C., Lorenzoni, P. J., and Werneck, L. C. (2019). HLA-alleles class I and II associated with genetic susceptibility to neuromyelitis optica in Brazilian patients. Arq. Neuropsiquiatr. 77, 239–247. doi: 10.1590/0004-282X20190031
Kim, H. J., Park, H. Y., Kim, E., Lee, K. S., Kim, K. K., Choi, B. O., et al. (2010). Common CYP7A1 promoter polymorphism associated with risk of neuromyelitis optica. Neurobiol. Dis. 37, 349–355. doi: 10.1016/j.nbd.2009.10.013
Kim, J. Y., Bae, J. S., Kim, H. J., and Shin, H. D. (2014). CD58 polymorphisms associated with the risk of neuromyelitis optica in a Korean population. BMC Neurol. 14:57. doi: 10.1186/1471-2377-14-57
Lennon, V. A., Kryzer, T. J., Pittock, S. J., Verkman, A. S., and Hinson, S. R. (2005). IgG marker of optic-spinal multiple sclerosis binds to the aquaporin-4 water channel. J. Exp. Med. 202, 473–477. doi: 10.1084/jem.20050304
Liu, C., Wang, G., Liu, H., Li, Y., Li, J., Dai, Y., et al. (2012). CD226 Gly307Ser association with neuromyelitis optica in Southern Han Chinese. Can. J. Neurol. Sci. 39, 488–490. doi: 10.1017/s0317167100014001
Lunin, S., Khrenov, M., Glushkova, O., Parfenyuk, S., Novoselova, T., and Novoselova, E. (2020). Precursors of thymic peptides as stress sensors. Expert. Opin. Biol. Ther. 20, 1461–1475. doi: 10.1080/14712598.2020.1800636
Marcus, J. H., and November, J. (2017). Visualizing the geography of genetic variants. Bioinformatics 33, 594–595. doi: 10.1093/bioinformatics/btw643.
Matsushita, T., Masaki, K., Isobe, N., Sato, S., Yamamoto, K., Nakamura, Y., et al. (2020). Genetic factors for susceptibility to and manifestations of neuromyelitis optica. Ann. Clin. Transl. Neurol. 7, 2082–2093. doi: 10.1002/acn3.51147
Matzaraki, V., Kumar, V., Wijmenga, C., and Zhernakova, A. (2017). The MHC locus and genetic susceptibility to autoimmune and infectious diseases. Genome Biol. 18:76. doi: 10.1186/s13059-017-1207-1
Paul, S., Mondal, G. P., Bhattacharyya, R., Ghosh, K. C., and Bhat, I. A. (2020). Neuromyelitis optica spectrum disorders. J. Neurol. Sci. 420:117225. doi: 10.1016/j.jns.2020.117225
Romero-Hidalgo, S., Flores-Rivera, J., Rivas-Alonso, V., Barquera, R., Villarreal-Molina, M. T., Antuna-Puente, B., et al. (2020). Native American ancestry significantly contributes to neuromyelitis optica susceptibility in the admixed Mexican population. Sci. Rep. 10:13706. doi: 10.1038/s41598-020-69224-3
Romero-Hidalgo, S., Ochoa-Leyva, A., Garciarrubio, A., Acuna-Alonzo, V., Antunez-Arguelles, E., Balcazar-Quintero, M., et al. (2017). Demographic history and biologically relevant genetic variation of Native Mexicans inferred from whole-genome sequencing. Nat. Commun. 8:1005. doi: 10.1038/s41467-017-01194-z
R-Software (2021). R Software Environment. Available at: https://www.r-project.org/
Sadaka, Y., Verhey, L. H., Shroff, M. M., Branson, H. M., Arnold, D. L., Narayanan, S., et al. (2012). 2010 McDonald criteria for diagnosing pediatric multiple sclerosis. Ann. Neurol. 72, 211–223. doi: 10.1002/ana.23575
Silva, A. B., Aw, D., and Palmer, D. B. (2009). Neuropeptides and thymic hormones in the Xenopus thymus. Front. Biosci. (Landmark Ed.) 14, 1990–2003. doi: 10.2741/3358
Taylor, M. R., Slavov, D., Gajewski, A., Vlcek, S., Ku, L., Fain, P. R., et al. (2005). Thymopoietin (lamina-associated polypeptide 2) gene mutation associated with dilated cardiomyopathy. Hum. Mutat. 26, 566–574. doi: 10.1002/humu.20250
VanderWeele, T. J. (2010). Epistatic interactions. Stat. Appl. Genet. Mol. Biol. 9:1. doi: 10.2202/1544-6115.1517
Wang, H., Dai, Y., Qiu, W., Zhong, X., Wu, A., Wang, Y., et al. (2011). HLA-DPB1 0501 is associated with susceptibility to anti-aquaporin-4 antibodies positive neuromyelitis optica in southern Han Chinese. J. Neuroimmunol. 233, 181–184. doi: 10.1016/j.jneuroim.2010.11.004
Wang, H., Zhong, X., Wang, K., Qiu, W., Li, J., Dai, Y., et al. (2012). Interleukin 17 gene polymorphism is associated with anti-aquaporin 4 antibody-positive neuromyelitis optica in the Southern Han Chinese–a case control study. J. Neurol. Sci. 314, 26–28. doi: 10.1016/j.jns.2011.11.005
Watanabe, I., Sagawa, A., Yasuda, I., Tanimura, K., Mukai, M., Shimizu, M., et al. (1985). [The 1982 revised criteria of American Rheumatism Association for the classification of systemic lupus erythematosus: a review of 126 patients]. Nihon Naika Gakkai Zasshi 74, 1529–1534.
Woo, H. J., Yu, C., and Reifman, J. (2017). Collective genetic interaction effects and the role of antigen-presenting cells in autoimmune diseases. PLoS One 12:e0169918. doi: 10.1371/journal.pone.0169918
Yoshimura, S., Isobe, N., Matsushita, T., Yonekawa, T., Masaki, K., Sato, S., et al. (2013). Distinct genetic and infectious profiles in Japanese neuromyelitis optica patients according to anti-aquaporin 4 antibody status. J. Neurol. Neurosurg. Psychiatry 84, 29–34. doi: 10.1136/jnnp-2012-302925
Yue, S., Zheng, X., and Zheng, Y. (2019). Cell-type-specific role of lamin-B1 in thymus development and its inflammation-driven reduction in thymus aging. Aging Cell 18:e12952. doi: 10.1111/acel.12952
Zhuang, J. C., Wu, L., Qian, M. Z., Cai, P. P., Liu, Q. B., Zhao, G. X., et al. (2015). Variants of Interleukin-7/Interleukin-7 receptor alpha are associated with both neuromyelitis optica and multiple sclerosis among Chinese han population in Southeastern China. Chin. Med. J. (Engl.) 128, 3062–3068. doi: 10.4103/0366-6999.169093
Keywords: NMOSD, TMPO, HLA, genetic interaction, rs17028450, rs9272219
Citation: Rosas-Madrigal S, Villarreal-Molina MT, Flores-Rivera J, Rivas-Alonso V, Macias-Kauffer LR, Ordoñez G, Chima-Galán MdC, Acuña-Alonzo V, Macín-Pérez G, Barquera R, Granados J, Valle-Rios R, Corona T, Carnevale A and Romero-Hidalgo S (2021) Interaction of HLA Class II rs9272219 and TMPO rs17028450 (Arg690Cys) Variants Affects Neuromyelitis Optica Spectrum Disorder Susceptibility in an Admixed Mexican Population. Front. Genet. 12:647343. doi: 10.3389/fgene.2021.647343
Received: 29 December 2020; Accepted: 23 June 2021;
Published: 15 July 2021.
Edited by:
Michael Dean, National Cancer Institute at Frederick, United StatesReviewed by:
Alessandro Gialluisi, Istituto Neurologico Mediterraneo Neuromed, Scientific Institute for Research, Hospitalization and Healthcare (IRCCS), ItalyCarolina Bekker Mendez, La Raza National Medical Center, Mexico
Julian Ramírez-Bello, Hospital Juárez de México, Mexico
Copyright © 2021 Rosas-Madrigal, Villarreal-Molina, Flores-Rivera, Rivas-Alonso, Macias-Kauffer, Ordoñez, Chima-Galán, Acuña-Alonzo, Macín-Pérez, Barquera, Granados, Valle-Rios, Corona, Carnevale and Romero-Hidalgo. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Sandra Romero-Hidalgo, c3JvbWVyb0Bpbm1lZ2VuLmdvYi5teA==