 Sufang Peng
Sufang Peng Hang Su
Hang Su Tianzhen Chen
Tianzhen Chen Xiaotong Li
Xiaotong Li Jiang Du
Jiang Du Haifeng Jiang
Haifeng Jiang Min Zhao
Min Zhao- 1Shanghai Mental Health Center, Shanghai Jiao Tong University School of Medicine, Shanghai, China
- 2Shanghai Key Laboratory of Psychotic Disorders, Shanghai, China
- 3CAS Center for Excellence in Brain Science and Intelligence Technology (CEBSIT), Chinese Academy of Sciences, Shanghai, China
Objects: To explore the long-term influence of methamphetamine abuse on metabolomics character, with gas chromatography-mass spectrometry (GS-MS) technology, and the potential regulatory network using the bioinformatics method.
Methods: Forty withdrawal methamphetamine abusers (WMA) were recruited from Shanghai Gaojing Forced Isolation Detoxification Institute. Forty healthy controls (HC) were recruited from society. GS-MS technology was used to detect metabolic products in serum. A bioinformatics method was used to build a regulatory network. Q-PCR was used to detect the candidate gene expressions, and ELISA was used to detect the regulatory enzyme expressions.
Results: Four pathways were significantly changed in the MA compared to the HC: (1) the arginine synthesis pathway, (2) alanine, aspartic acid and glutamate metabolic pathway, (3) cysteine and methionine metabolic pathway, and (4) the ascorbate and aldarate pathway (enrichment analysis p < 0.05, Impactor factor > 0.2). When focusing on the ‘Alanine, aspartate, and glutamate metabolism’ pathway, a regulatory network was established, and the expression of candidate regulatory genes and enzymes was verified. It was found that the expression of DLG2 (Discs large MAGUK scaffold protein 2), PLA2G4 (Phospholipase A2 group IVE), PDE4D (Phosphodiesterase 4D), PDE4B (Phosphodiesterase 4B), and EPHB2 (Ephrin type-B receptor 2) were significantly different between the two groups (p < 0.05), However, after adjusting for age and BMI, only DLG2, PLA2G4, and EPHB2 remained significant (p < 0.05). The expression of enzymes was not significantly different (p > 0.05).
Conclusion: Methamphetamine abuse influences the metabolic process in the long term, and DLG2, PLA2G4, and EPHB2 may regulate the glutamate metabolism pathway.
Introduction
Methamphetamine (METH) is a widely abused addictive psychostimulant, which accounts for a considerable share of global disease burden. METH abuse is increasing rapidly (Yang X. et al., 2018), especially in East and South-East Asia, and Oceania. Long-term abuse of METH causes serious physical and mental damage. Compared to other psychostimulants, METH is better at penetrating the center nervous system (CNS) and has a longer duration of action (Yang X. et al., 2018), making METH more addictive while the damage to the CNS is more serious and persistent. METH addicts often suffer from permanent psychotic symptoms (Mellsop et al., 2019) and cognitive decline, even when deprived from drugs, bringing great difficulties for patients who try to return to society. These symptoms are mostly attributed to METH-induced long-term neurotoxicity and excitotoxicity (Ashok et al., 2017). The neurochemical mechanisms underlying neurotoxicity and excitotoxicity are complex and not well understood. Glutamate, as an important excitatory neurotransmitter is thought to activate various downstream signaling pathways, for example the apoptotic pathway, and eventually produce neurotoxicity (Courtney and Ray, 2014; Yang X. et al., 2018). A previous study found that methamphetamine may result in glutamine system disturbance (Tasic et al., 2017), and glutamate + glutamine in the right inferior frontal cortex is decreased in methamphetamine users (Zheng et al., 2016).
Metabolomics is a potentially powerful tool for understanding the global biological consequences of disease and drug administration. In recent years, with the improvement of metabolomics technology, metabolomics research in psychotic diseases has made great progress (Bortolato et al., 2009; Zhang et al., 2016; Proitsi et al., 2017; Rodríguez Cerdeira et al., 2017). However, in the field of substance addiction, metabolomics research is still in its initial stages. A few studies focus on methamphetamine addiction, using animals as study subjects. Energy metabolic disturbance is believed to be the biochemical base of toxicity and addictive property of MTHE (Zhang et al., 2016). Neurotransmitters in serum or plasma are also disrupted by METH, especially in the glutamate-glutamine-GABA system (Rist et al., 2017). Nevertheless, metabolic character differences between animals and humans still exist. This study uses withdrawal methamphetamine abusers (WMA) as objects and explore the metabolomics characters and possible regulatory mechanisms that may be the basis of the long-term neurotoxicity of methamphetamine.
Materials and Methods
Ethics Statement
Informed consent of the participants was obtained after the nature of the procedures used in the study had been fully explained. The study was carried out in accordance with the last version of the Declaration of Helsinki, and the protocol was approved by the Ethics Committee of Shanghai Mental Health Center (SMHC IRB 2016-11R). All participants were identified using numbers, instead of names.
Research Participants
Forty male WMA were recruited from Shanghai Gaojing Forced Isolation Detoxification Institute in July 2017, and 40 male healthy controls were recruited from general society. The entire sample consisted of a Chinses Han population. The WMA accepted assessment from two senior psychiatric doctors and met the diagnosis criterion for methamphetamine abuse disorder according to the Diagnostic and Statistical Manual of mental disorders, fifth edition (DSM-5). Participants did not have the opportunity to use any illicit substance during the last 2 months. They completed a self-report form that included basic demographic information and accepted an interview of Addiction Severity Index (ASI). Exclusion criteria for both groups were: (1) history of craniocerebral trauma or epileptic seizure; (2) metal implants in the brain or an implanted pacemaker; (3) history of major physical diseases, organic mental disorders, and severe mental disorders. People in the control group did not have any history of illegal substance abuse. Venous blood was collected at 7:30–8:00 am before breakfast.
Gas Chromatography – Mass Spectroscopy (GC-MS)
After standing at room temperature for 30 min, venous blood was centrifuged at 4°C, 3000g, for 15 min. The upper layer of the serum was collected. Sample preparation, GC-MS experimental process, and GC-MS data processing were carried out using previously published methods (Morris et al., 2012; Li et al., 2014). Compound identification was performed by comparing the retention indices and mass spectral data with those previously generated from reference standards of known structures present in the JiaLib metabolite database using the proprietary software XploreMET. The current JiaLib comprises over 1,200 mammalian metabolites over a 15-year accumulation. The reference chemicals present in JiaLib were commercially purchased from Sigma-Aldrich (St. Louis, MO, United States), Santa Cruz (Dallas, TX, United States), Nu-Chek Prep (Elysian, MN, United States), and synthesized in the laboratory. XploreMET adopted a specific algorithm hypergeometric, which can enrich the information of metabolites, weight the contribution of metabolites, and focus on the key metabolic network. The P and impact value was used to measure the importance of metabolic pathway.
Bioinformatics Analysis
Based on the results of untargeted metabolomics, a glutamate metabolism pathway was noticed. Glutamate is a very important excitatory neurotransmitter, and a previous study has confirmed that glutamate + glutamine in the right inferior frontal cortex was decreased in methamphetamine users (Zheng et al., 2016). In serum or plasma, the glutamate-glutamine-GABA system was also disrupted by METH (Rist et al., 2017). So, we chose the ‘alanine, aspartic acid and glutamate metabolic pathway’ to build the regulatory network.
The four differential expressed metabolites were treated as candidate metabolites. We collected 133 risk genes from previous genome-wide association studies (Heikkinen et al., 2018; Irwin et al., 2018). The molecular network was constructed by connecting candidate metabolites, enzymes, intermediate proteins, and risk genes. First, we investigated the relative enzymes linked to these candidate metabolic compounds according to the KEGG database and the Human Metabolome Database (HMDB). To further explore correlation networks linked to the relative enzymes and risk genes, we searched the functional proteins network database (STRING v10.5) setting the requirement that the score must be more than 700 and that no more than one intermediate protein must be present between the relative enzyme and risk gene. Finally, we built a network including four enzymes: ASNS (Asparagine synthetase), AGT (Angiotensinogen), GAD1 (Glutamate decarboxylase 1), and GAD2 (Glutamate decarboxylase 2); 13 genes: DLG2 (Discs large MAGUK scaffold protein 2), NRG1 (Neuregulin 1), PLA2G4E (Phospholipase A2 group IVE), STX8 (Syntaxin 8), NEDD4L (NEDD4 like E3 ubiquitin protein ligase), PRKG1 (Protein kinase cGMP-dependent 1), PDE4D (Phosphodiesterase 4D), PDE4B (Phosphodiesterase 4B), SERPINA5 (Serpin family A member 5), PDE6C (Phosphodiesterase 6C), CD55 (CD55 molecule), EPHB2 (Ephrin type-B receptor 2), and RAPGEF5 (Rap guanine nucleotide exchange factor 5), and several intermediate proteins.
RNA Extraction and Gene Expression Analysis
RNA was extracted from the serum samples with Trizol reagent (Invitrogen, Carlsbad, CA, United States) according to the previous protocol (Gan et al., 2018), and stored at −80°C for subsequent cDNA synthesis. Following RNA quantitation, 1ug of total RNA was used for cDNA synthesis (5 × All-In-One RT Master Mix, ABM, Canada), and the quantitative gene expression analysis was performed using TB Green Fast qPCR Mix (TaKaRa, Shiga, Japan), and Lightcycle® 96 real-time PCR machine (Roche Diagnostics GmbH, Germany). Primers are listed in Table 1. GAPDH was used as a housekeeper gene. The relative gene expression was calculated using the 2−△CT method (Kohno et al., 2016). The CT values of duplicates were averaged for the final analysis.

Table 1. Primers used in Q-PCR.
Elisa
The serum levels of AGT, GAD1, GAD2, and ASNS were assessed using an ELISA kit (catalog number SEC904Hu, Cloud-Clone Corp, Shanghai, China for GAD1, and catalog numbers MBS726474, MBS2501551, MBS090144; Mybiosource, Southern California, San Diego, United States, for AGT, GAD2, and ASNS, respectively).
All assays were performed according to the manufacturer’s directions and were performed in duplicate. The average value of the duplicates was used as the final value.
Statistical Analysis
General data was presented as mean and standard error of the mean (Mean ± SD), and analyzed using SSPSS 22.0. Data was analyzed using the student t test, a was 0.05 and Cohen’s d was used to calculate the effect size of the t test. 2–△CT was used to calculate the relative expression of genes, and the expression ratio of the WMA group relative to the control group was calculated as previously described (Kohno et al., 2016).
Metabolomics data was managed with the XploreMET system (v3.0, Metabo-Profile, Shanghai, China), and R was used to analyze the data. The NIST 2017 database was used for metabolite certification and Metaboanalyst 3.0 was used for the metabolic pathway analysis. Partial least squares discriminant analysis (PSL-DA) was performed, and a t test was used after log transformation. Differential expressed metabolites were screened according to fold change (FC) and p value. FC > 1. 5 and p < 0. 05 showed that the metabolite was significantly differentially expressed. Based on the selected differential metabolites, a pathway analysis was performed using a hypergeometric test and KEGG pathways (Yang W. et al., 2018) and the p values were calculated as follows:
PtwHits is the hit number of differential metabolites in a certain metabolic pathway. TotalAll is the number of metabolites in all pathways of a certain species, such as Homo sapiens. PtwAll is the total number of metabolites in a certain pathway and TotalHits is the hit number of differential metabolites in all pathways. A significant p value means significantly changed metabolic pathway activities that need to be focused on. For multiple tests, a false discovery rate (FDR) was applied to correct the raw p values.
Results
Demography Characteristic
The demographic characteristic are shown in Table 2. Age, BMI, and education level between the two groups were not significantly different. However, WMA individuals have lower marriage rates and higher unemployment (p < 0.05).
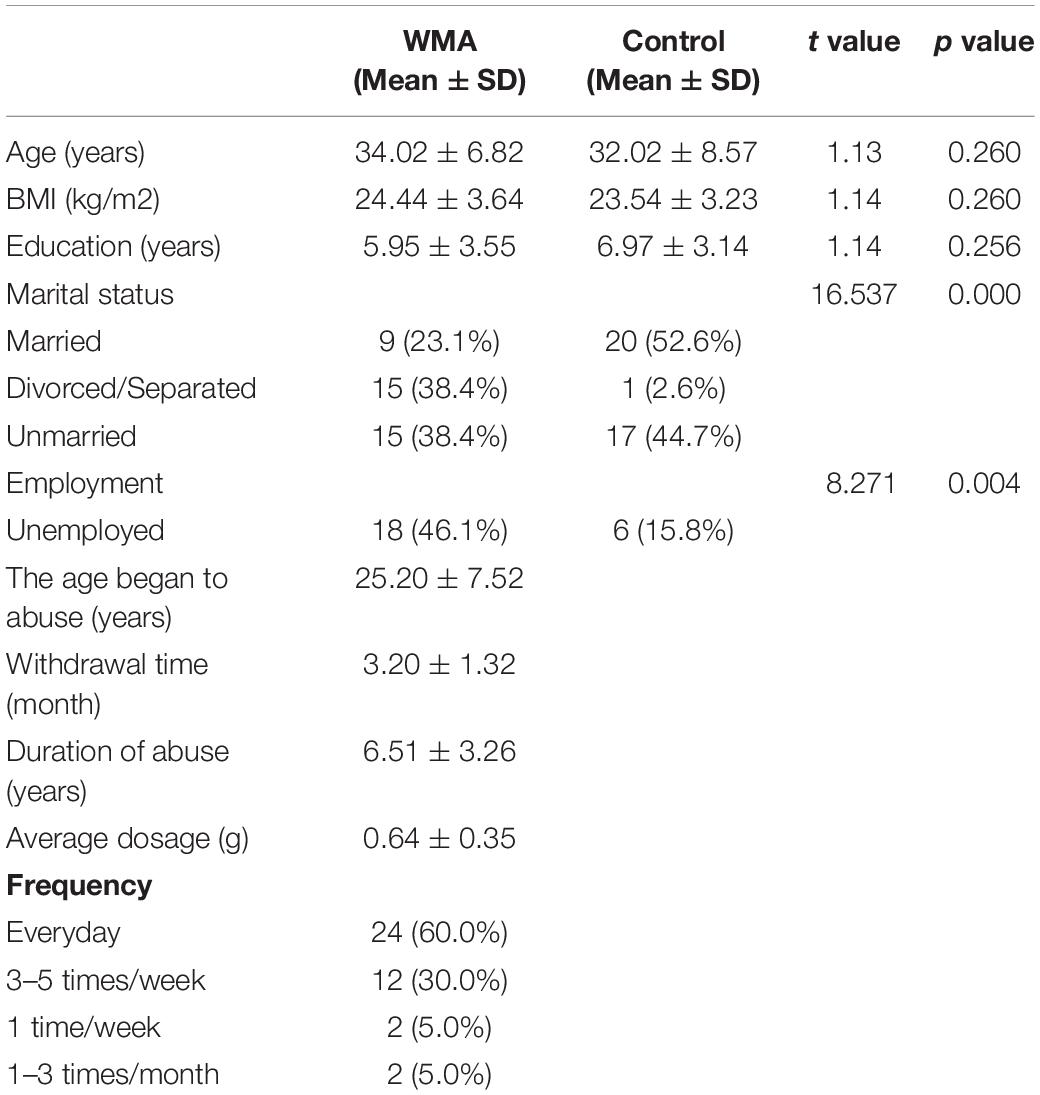
Table 2. Demographic characteristics of subjects.
Metabolomics Profile and Pathways
Thirty-five differentially expressed metabolites were screened by t test (p < 0.05), in which 21 metabolites could be found by searching metabolic pathways in the KEGG information database (as shown in Table 3). Since age and BMI may influence the metabolic function, we adjusted age and BMI. After that, there were 19 metabolites differentially expressed in two groups, including two kinds of alcohols, seven kinds of amino acids, one kind of carbohydrate, one kind of fatty acid, three kinds of nucleotides, and five kinds of organic acids.
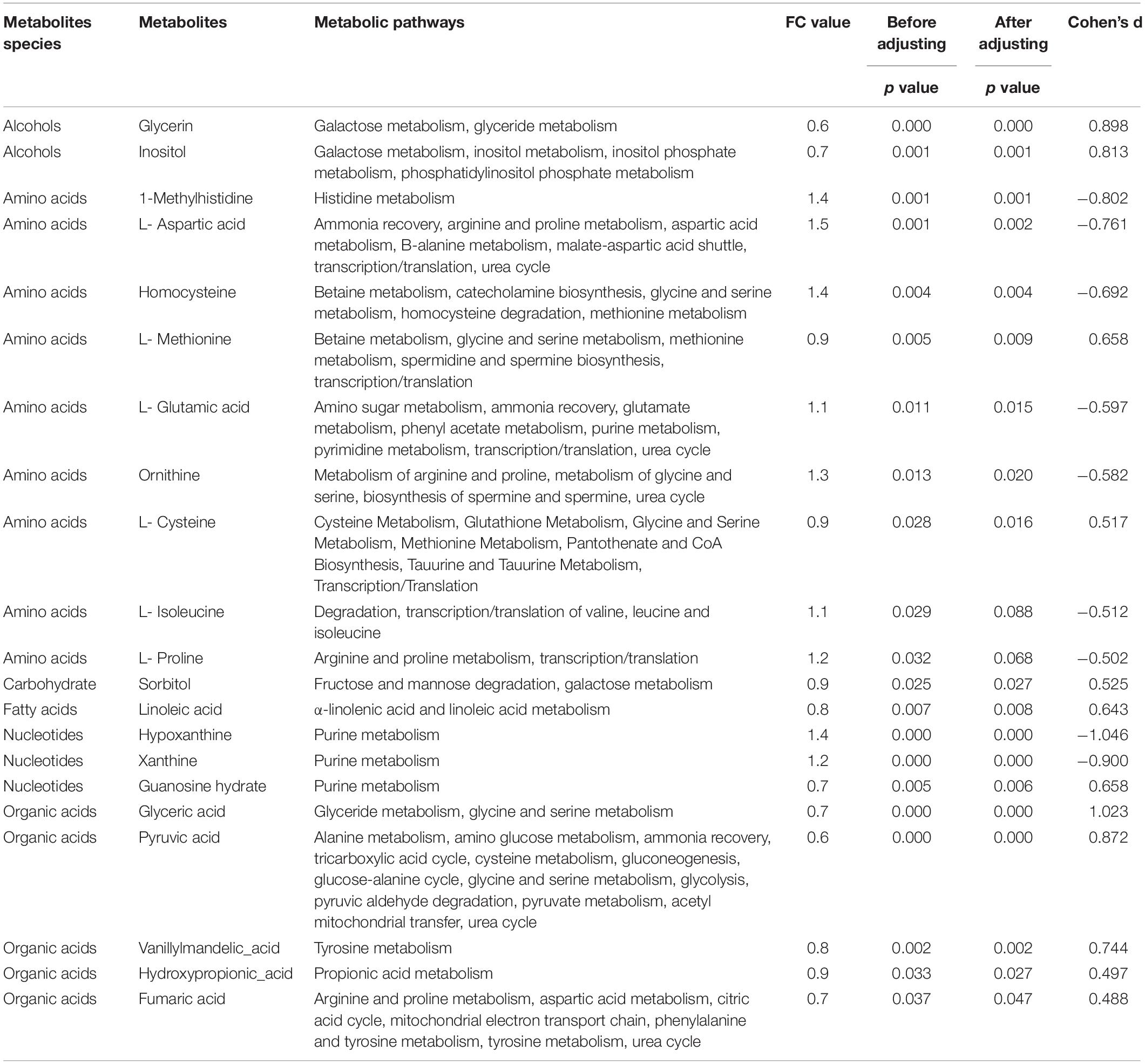
Table 3. Differentially expressed metabolites in pathways.
According to the results of the Metabolic Pathway Enrichment Analysis (MPEA), four pathways were found to be significantly changed; (1) arginine synthesis pathway, (2) alanine, aspartic acid and glutamate metabolic pathway, (3) cysteine and methionine metabolic pathways, and (4) ascorbate and aldarate pathway (enrichment analysis p < 0.05,Impactor factor > 0.2) (Figure 1).
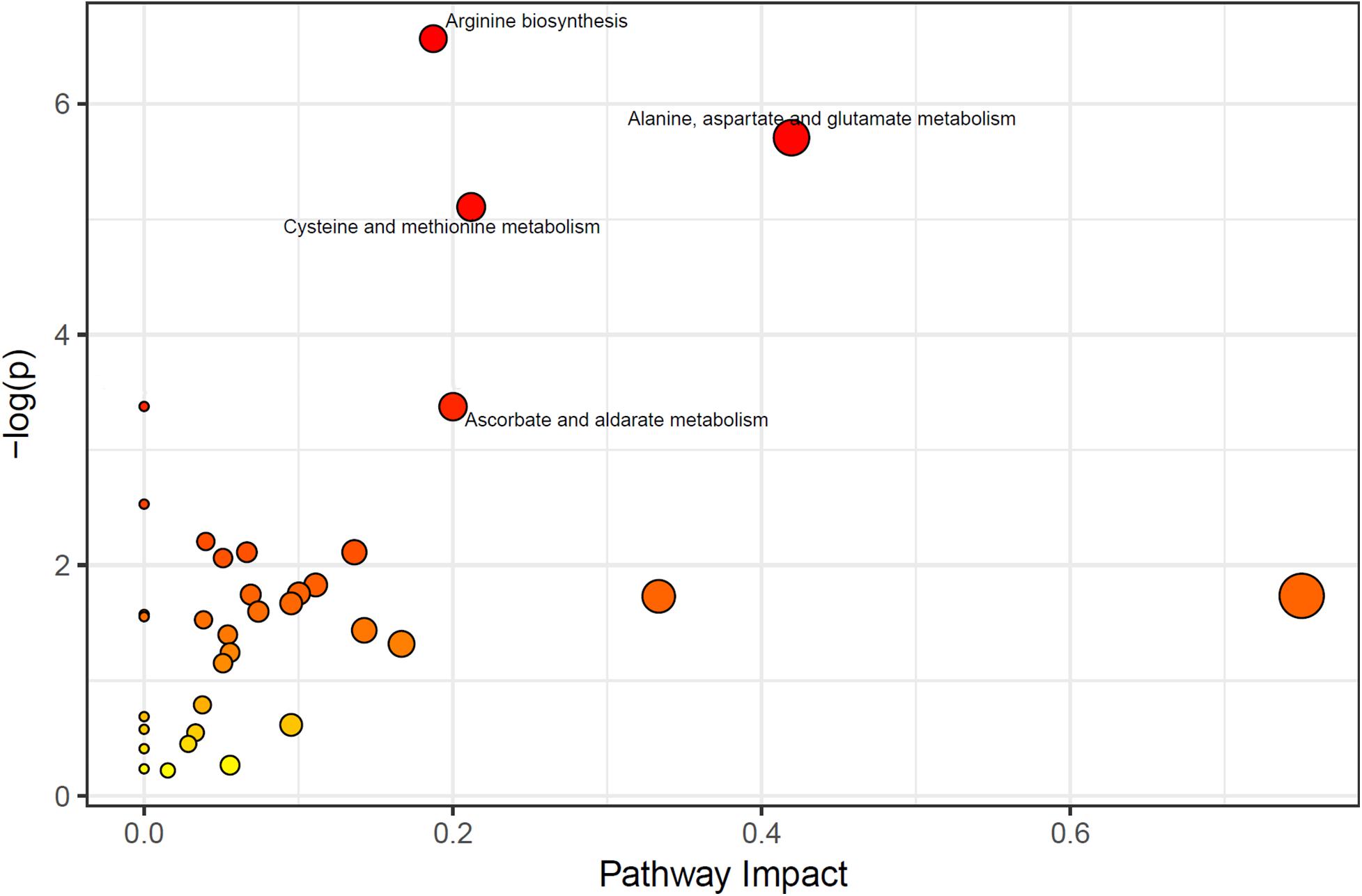
Figure 1. Significant pathways (the pathways which p < 0.05, and Impactor factor > 0.02 were marked out).
Glutamine Metabolites Regulatory Network
Previous research found that methamphetamine may result in glutamine system disturbance (Tasic et al., 2017), either in the brain and serum or in plasma (Zheng et al., 2016; Rist et al., 2017). This study also found a change in the glutamate pathway. We therefore focused on the ‘Alanine, aspartate, and glutamate metabolism’ pathway, and based on the bioinformatics method, the regulatory network was built. As shown in Figure 2, we found three metabolites (pyruvate, L-aspartate, and L-glutamine), which can be regulated by four enzymes, and which can be related to 13 candidate genes. Each candidate gene influences the regulatory enzyme through one intermediate protein (Figure 2).
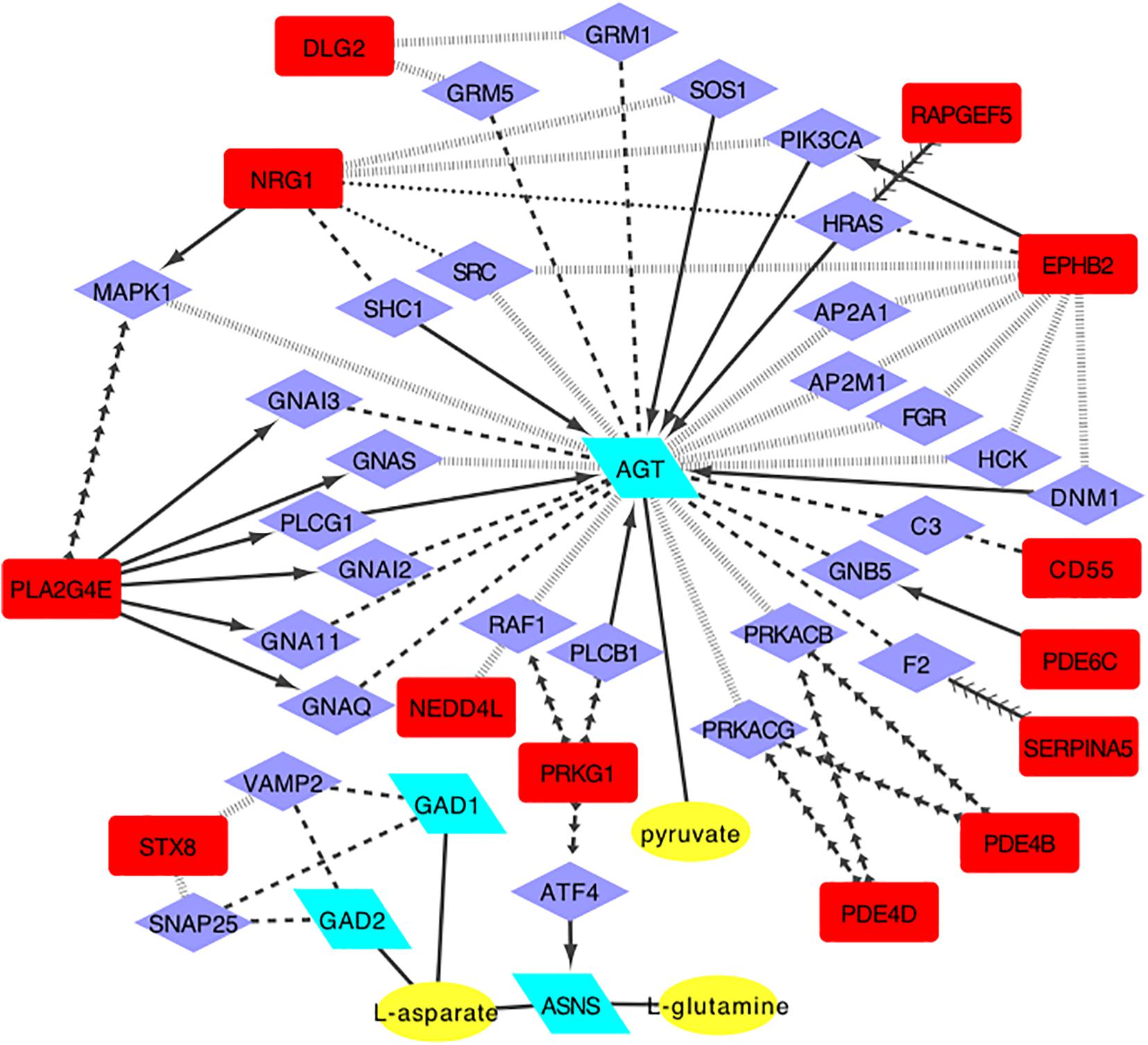
Figure 2. The regulatory network model for alanine, aspartic acid and glutamate metabolism pathway(enzyme, posttranslational modification, activate, inhibition, catalysis, reaction, and combine. Yellow oval, metabolites of glutamate pathway; Green parallelogram, regulatory enzymes; Purple diamond, intermediate proteins; Red square, candidate genes).
Candidate Gene Expression
According to the regulatory network, we used Q-PCR to verify the 13 candidate genes expressed. 2–△ CT was used to calculate the relative expression of genes (Kohno et al., 2016) (Table 4). DLG2, PLA2G4, PDE4D, PDE4B, and EPHB2 were significantly differentially expressed between the two groups (p < 0.05). However, after adjusting for age and BMI, only DLG2, PLA2G4, and EPHB2 remained significant (p < 0.05). STX8, RAPGEF5, and PDE6C were not detected because of their low expression.
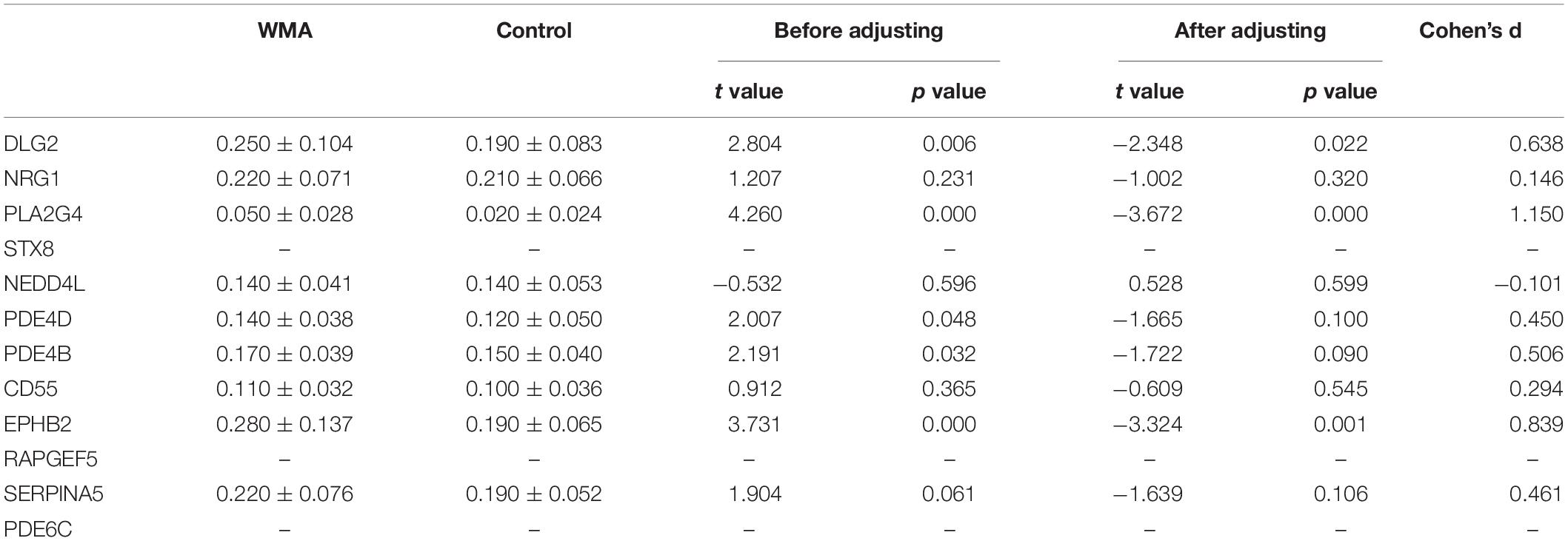
Table 4. Expression of candidate genes.
For the DLG2, PLA2G4, PDE4D, PDE4B, and EPHB2 genes, the expression in the WMA group was 1.473, 4.226, 1.334, 1.153, and 1.595 times higher than that in control group, respectively (Figure 3).
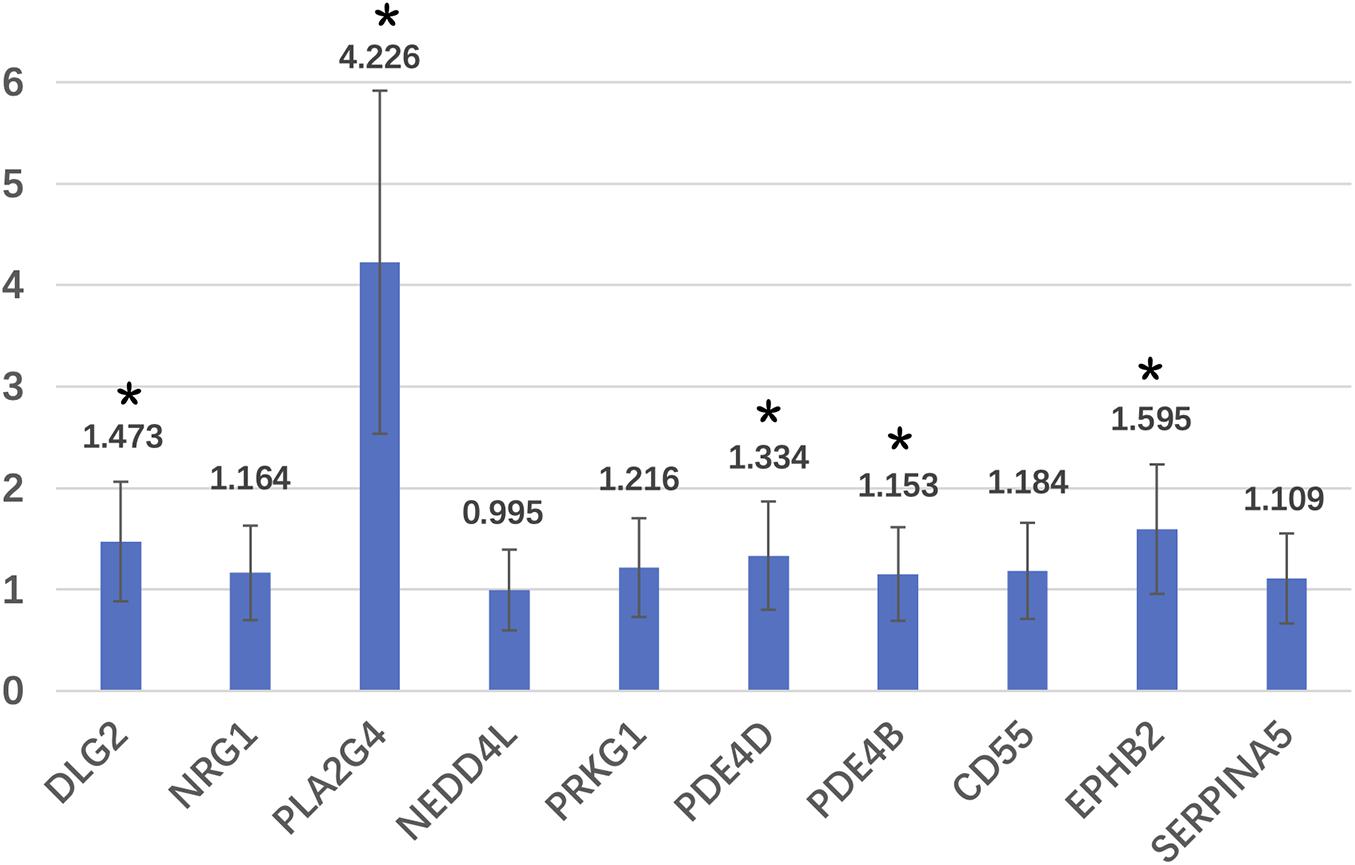
Figure 3. Relative expression fold of candidate genes. *Means that these genes are significantly different expressed in two groups.
Regulatory Enzyme Expression
ELISA was used to detect the regulatory enzyme expression. As Table 5 shows, no significant difference was found between the two groups (p > 0.05).
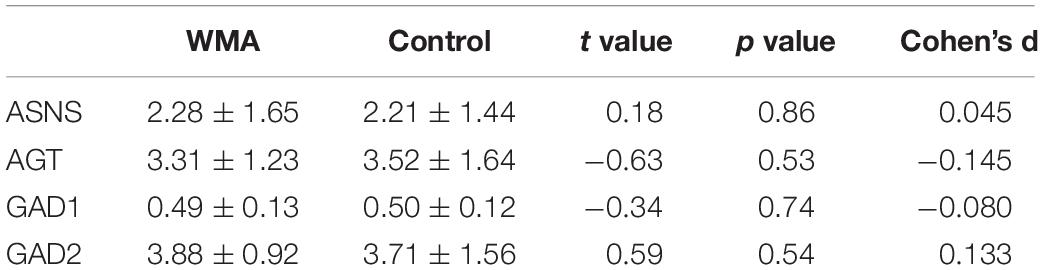
Table 5. Expression of candidate enzymes.
Discussion
This study found that peripheral metabolites of WMA patients were mainly changed in amino acid metabolism. The most obvious pathways were; (1) the arginine synthesis pathway, (2) the alanine, aspartic acid and glutamate metabolic pathways, (3) the cysteine and methionine metabolic pathways, and (4) the purine metabolic pathway.
Previous studies on methamphetamine addicted animals found that abnormal energy metabolism was the biochemical basis of toxicity and addiction (Cservenka and Ray, 2017; Moallem et al., 2018). An increase in energy metabolism was found in the serum of amphetamine-administered rats. After 5 consecutive days of administration, the difference between the administration and control group decreased (Moallem et al., 2018). After high dose, high frequency, or long time administration, the energy reserve may be depleted (Moallem et al., 2018). The patients in this study were basically deprived from METH for about 2 months. This may explain why abnormal energy metabolism was not observed—the energy reserves may have been consumed.
In addition, amphetamines cause serum/plasma neurotransmitter disturbances mainly in the glutamate-glutamine-GABA system (Moallem et al., 2018). Glutamate-Glutamine disturbance was observed both in withdrawal animals and in the WMA patients (Shima et al., 2011; Chen et al., 2015; Su et al., 2018), and this disturbance may be related to a continuous search for drugs and the high relapse risk (Zheng et al., 2014). Glutamate, aspartic acid, and methionine are elevated in rat neuroblastoma cells 48 h after methamphetamine treatment (Bu et al., 2013). These studies showed that the effect of methamphetamine on the glutamate system could last for a long time. In this study, the “alanine, aspartic acid, and glutamate pathway” was found to be abnormal, and the expression of related metabolites was significantly changed, which is consistent with previous studies.
We chose the glutamate metabolic pathway and used bioinformatics methods to establish a regulatory network model, which predicts that GAD1, GAD2, ASNS, and AGT may directly regulate the glutamate pathway metabolites- pyruvate, L-aspartate, L-glutamine. The regulatory enzyme is regulated by DLG2, NRG1, PLA2G4E, STX8, NEDD4L, PRKG1, PDE4D, PDE4B, SERPINA5, PDE6C, CD55, EPHB2, and RAPGEF5 genes only through one intermediate protein. We verified the expression of the candidate genes and enzymes, and finally found five regulatory genes that were differentially expressed in the two groups, namely DLG2, PLA2G4, PDE4D, PDE4B, and EPHB2 genes. After adjusting for age and BMI, only DLG2, PLA2G4, and EPHB2 remained significant.
DLG2 plays a very important role in synaptic development and stability of plasticity (McClay et al., 2013; Su et al., 2020). The function of the DLG2 gene is associated with complex cognitive and learning tasks (Parsegian and See, 2014), and is also associated with developmental and intellectual disorders (Christopher et al., 2007) such as Parkinson’s disease (Darna et al., 2015). The DLG2 gene polymorphism was relevant for methamphetamine addiction (Irwin et al., 2018). This study also found that the expression of the DLG2 gene was up-regulated in WMA subjects. There was a long-term cognitive impairment in methamphetamine addicted populations (Crocker et al., 2014), as well as dysfunction of the glutamate system (Maker et al., 2018). The DLG2 gene may play an important role in the development, plasticity, and stability of glutamatergic synapses and may further affect the cognitive function of WMA patients.
The protein coded by the PLA2G4E gene is involved in the regulation of membrane-mediated transport and affects phospholipase activity (Zhu et al., 2016). PLA2G4E is strongly expressed in neurons and regulates endogenous cannabinoids, by mobilizing intracellular calcium (Zhu et al., 2016). The endogenous cannabinoid system also regulates the function of the glutamate system. Cannabinoid 1 receptors affect glutamatergic and GABAergic regulation of anxiety and fear responses (Zheng et al., 2011; Nithianantharajah et al., 2013). The PLA2G4E gene expression changed the most in this study, reaching 4.226 times that of the control group. That may influence cannabinoid, glutamate, and GABA system functions.
The PDE4D and PDE4B genes belong to the phosphodiesterase (PDE family) superfamily, and play an important role in the degradation of cAMP (Reggiani et al., 2017). Since cAMP is an important second messenger, PDE4D/4B may play a role in signal transduction by regulating cAMP concentration. PDE4D/4B is widely expressed in the human brain. Previous studies have shown that PDE4D/4B is associated with neuropsychiatric disorders such as depression, schizophrenia (Wu et al., 2018), methamphetamine addiction (Irwin et al., 2018), alcohol-induced neuroinflammation (Ikeda et al., 2013), and stroke (Zhong et al., 2016). Since signal transduction of neurotransmitters in the brain, such as dopamine, glutamate, and GABA, etc., requires the participation of second messenger cAMP, some researchers believe that the phosphodiesterase family (PDE) may be used as an important drug target for schizophrenia and affective disorder (Wu et al., 2018). This study found that the expression of PDE4D/4B in the peripheral blood of WMA patients was significantly higher than that of the control group. However, after adjusting for age and BMI, this difference was no longer significant. The expression ratios of the WMA group compared to the control group were also close to 1, showing that these two genes may not be important regulatory genes in the glutamine system of WMA.
The EPHB2 gene encodes Ephrin type-B receptor 2, which is a tyrosine kinase receptor involved in triggering and maturation. In mature brains, EPHB2 is highly abundant in large dendritic axes and in the frontal cortex and dendritic spines in the hippocampus, and is also involved in the regulation of glutamate receptor distribution and excitatory synapse formation (Su et al., 2017). EPHB2 also regulates synaptic plasticity and leads to anxiety behavior (Capestrano et al., 2014) and reversal of cognitive deficits (Ruehle et al., 2012). EPHB2 reverses cognitive dysfunction in the Alzheimer’s disease model, possibly related with glutamate receptor-NMDA’s function (N-methyl-D-aspartic acid receptor), and EPHB2 is a key regulator of synaptic localization of NMDA receptors (Hu et al., 2017). Knockout EPHB2 induces depression-like behavior and memory impairment in mice, which is mediated by NMDA receptor subtypes (Zhen et al., 2018). This study found a significant increase of the EPHB2 gene in WMA patients, which elicits that an abnormal expression of the EPHB2 gene may be a potential mechanism of glutamate system dysfunction and emotional or cognitive symptoms of methamphetamine addicts.
The expression of candidate regulatory enzymes was not differentially expressed between two groups. However, the activity of enzymes was not detected, which may influence the metabolites. Another complex regulatory mechanism could not be excluded.
Conclusion
MA use has a long-term and extensive impact on the human body, which can be reflected by the peripheral metabolites. The glutamate system metabolic disturbance may be regulated by the genes that are associated with multiple pathophysiological processes such as synaptic plasticity, signal transduction, cAMP degradation, phosphorylation, and synaptic localization, which may be related to clinical symptoms of withdrawal MA, such as long-term cognitive impairment, anxiety and fear responses, and so on.
There were some limitations in this study. First, only one time point was selected for the exploration of serum metabolomics in the addiction state of methamphetamine. If long-term follow-up can be performed during the acute phase and withdrawal period, the changes in metabolomics characteristics over time will be observed and the results will be more systematic. Second, the exploration of the mechanism is limited to biochemical and gene regulation mechanisms, and if it can be combined with the analysis of central neurotransmitter changes, it will be more in-depth. Third, the enzyme activity was not detected. Fourth, the sample size was limited, and all the enrolled participants were from the Shanghai area only. The conclusion can therefore only be extended to Shanghai Han nationality. A multi-center collaboration may therefore facilitate further discoveries.
Data Availability Statement
The raw data supporting the conclusions of this article will be made available by the authors.
Ethics Statement
The studies involving human participants were reviewed and approved by Ethics Committee of Shanghai Mental Health Center. The patients/participants provided their written informed consent to participate in this study.
Author Contributions
SP: conceptualization, data curation, formal analysis, methodology, writing – original draft, review, and editing. HS: investigation, project administration, and supervision. TC: investigation and project administration. XL: investigation and project administration. JD: conceptualization, methodology, project administration, resources, and supervision. HJ: conceptualization, methodology, project administration, resources, and supervision. MZ: conceptualization, funding acquisition, methodology, resources, supervision, and writing – review and editing. All authors contributed to the article and approved the submitted version.
Funding
This work was supported by the National Natural Science Foundation (81771436), Shanghai Key Laboratory of Psychotic Disorders (13DZ2260500), Shanghai Municipal Science and Technology Major Project (2018SHZDZX05), Shanghai Clinical Research Center for Mental Health (19MC1911100), Shanghai Shenkang Hospital Development Center (SHDC2020CR3045B), and Shanghai Intelligent Engineering Technology Research Center for Addiction and rehabilitation (19DZ2255200).
Conflict of Interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Acknowledgments
We appreciate the support from Dr. Xingxing Jian, Shanghai Bioinformation Technology Research Center for the bioinformation analysis and Dr. Qingzhong Wang, Chinese Academy of Sciences for gene expression.
References
Ashok, A. H., Mizuno, Y., Volkow, N. D., and Howes, O. D. (2017). Association of stimulant use with dopaminergic alterations in users of cocaine, amphetamine, or methamphetamine a systematic review and meta-analysis. JAMA Psychiatry 74, 511–519. doi: 10.1001/jamapsychiatry.2017.0135
Bortolato, M., Frau, R., Piras, A. P., Luesu, W., Bini, V., Diaz, G., et al. (2009). Methamphetamine induces long-term alterations in reactivity to environmental stimuli: correlation with dopaminergic and serotonergic toxicity. Neurotox. Res. 15, 232–245. doi: 10.1007/s12640-009-9024-2
Bu, Q., Lv, L., Yan, G., Deng, P., Wang, Y., Zhou, J., et al. (2013). NMR-based metabonomic in hippocampus, nucleus accumbens and prefrontal cortex of methamphetamine-sensitized rats. Neurotoxicology 36, 17–23. doi: 10.1016/j.neuro.2013.02.007
Capestrano, M., Mariggio, S., Perinetti, G., Egorova, A. V., Iacobacci, S., Santoro, M., et al. (2014). Cytosolic phospholipase A2e drives recycling through the clathrin-independent endocytic route. J. Cell Sci. 127, 977–993. doi: 10.1242/jcs.136598
Chen, Y. C., Chen, C. K., and Wang, L. J. (2015). Predictors of relapse and dropout during a 12-Week relapse prevention program for methamphetamine users. J. Psychoactive Drugs 47, 317–324. doi: 10.1080/02791072.2015.1071447
Christopher, L. N., Michael, M., Joseph, B. W., Kerstin, A. F., and Wolf, M. E. (2007). Behavioral sensitization to amphetamine is not accompanied by changes in glutamate receptor surface expression in the rat nucleus accumbens Christopher. J. Neurochem. 86, 573–579.
Courtney, K. E., and Ray, L. A. (2014). Methamphetamine: an update on epidemiology, pharmacology, clinical phenomenology, and treatment literature. Drug Alcohol Depend. 1, 11–21. doi: 10.1016/j.drugalcdep.2014.08.003
Crocker, C. E., Bernier, D. C., Hanstock, C. C., Lakusta, B., Purdon, S. E., Seres, P., et al. (2014). Prefrontal glutamate levels differentiate early phase schizophrenia and methamphetamine addiction: a 1 H MRS study at 3 tesla. Schizophr. Res. 157, 231–237.
Cservenka, A., and Ray, L. A. (2017). Self-reported attentional and motor impulsivity are related to age at first methamphetamine use. Addict. Behav. 65, 7–12. doi: 10.1016/j.addbeh.2016.09.008
Darna, M., Beckmann, J. S., Gipson, C. D., Bardo, M. T., and Dwoskin, L. P. (2015). Effect of environmental enrichment on dopamine and serotonin transporters and glutamate neurotransmission in medial prefrontal and orbitofrontal cortex. Brain Res. 1599, 115–125. doi: 10.1016/j.brainres.2014.12.034
Gan, H., Zhao, Y., Jiang, H., Zhu, Y., Chen, T., Tan, H., et al. (2018). A research of methamphetamine induced psychosis in 1,430 individuals with methamphetamine use disorder: clinical features and possible risk factors. Front. Psychiatry 9:551. doi: 10.3389/fpsyt.2018.00551
Heikkinen, N., Karkkainen, O., Laukkanen, E., Kekkonen, V., Kaarre, O., Kivimaki, P., et al. (2018). Changes in the serum metabolite profile correlate with decreased brain gray matter volume in moderate-to-heavy drinking young adults. Alcohol 75, 89–97. doi: 10.1016/j.alcohol.2018.05.010
Hu, R., Wei, P., Jin, L., Zheng, T., Chen, W.-Y., Liu, X.-Y., et al. (2017). Overexpression of EphB2 in hippocampus rescues impaired NMDA receptors trafficking and cognitive dysfunction in Alzheimer model. Cell Death Dis. 8:e2717. doi: 10.1038/cddis.2017.140
Ikeda, M., Okahisa, Y., Aleksic, B., Won, M., Kondo, N., Naruse, N., et al. (2013). Evidence for shared genetic risk between methamphetamine-induced psychosis and schizophrenia. Neuropsychopharmacology 38, 1864–1870. doi: 10.1038/npp.2013.94
Irwin, C., van Reenen, M., Mason, S., Mienie, L. J., Wevers, R. A., Westerhuis, J. A., et al. (2018). The 1H-NMR-based metabolite profile of acute alcohol consumption: a metabolomics intervention study. PLoS One 13:e0196850. doi: 10.1371/journal.pone.0196850
Kohno, M., Okita, K., Morales, A. M., Robertson, C. L., Dean, A. C., Ghahremani, D. G., et al. (2016). Midbrain functional connectivity and ventral striatal dopamine D2-type receptors: link to impulsivity in methamphetamine users. Mol. Psychiatry 21, 1554–1560. doi: 10.1038/mp.2015.223
Li, H., Bu, Q., Chen, B., Shao, X., Hu, Z., Deng, P., et al. (2014). Mechanisms of metabonomic for a gateway drug: nicotine priming enhances behavioral response to cocaine with modification in energy metabolism and neurotransmitter level. PLoS One 9:e87040. doi: 10.1371/journal.pone.0087040
Maker, G. L., Green, T., Mullaney, I., and Trengove, R. D. (2018). Untargeted metabolomic analysis of rat neuroblastoma cells as a model system to study the biochemical effects of the acute administration of methamphetamine. Metabolites 8:38. doi: 10.3390/metabo8020038
McClay, J. L., Adkins, D. E., Vunck, S. A., Batman, A. M., Vann, R. E., Clark, S. L., et al. (2013). Large-scale neurochemical metabolomics analysis identifies multiple compounds associated with methamphetamine exposure. Metabolomics 9, 392–402. doi: 10.1007/s11306-012-0456-y
Mellsop, G., Tapsell, R., and Holmes, P. (2019). Mental health service users’ progression from illicit drug use to schizophrenia in New Zealand. Gen. Psychiatr. 32:e100088. doi: 10.1136/gpsych-2019-100088
Moallem, N. R., Courtney, K. E., and Ray, L. A. (2018). The relationship between impulsivity and methamphetamine use severity in a community sample. Drug Alcohol Depend. 187, 1–7. doi: 10.1016/j.drugalcdep.2018.01.034
Morris, C., O’Grada, C., Ryan, M., Roche, H. M., Gibney, M. J., Gibney, E. R., et al. (2012). The relationship between BMI and metabolomic profiles: a focus on amino acids. Proc. Nutr. Soc. 71, 634–638. doi: 10.1017/s0029665112000699
Nithianantharajah, J., Komiyama, N. H., McKechanie, A., Johnstone, M., Blackwood, D. H., St Clair, D., et al. (2013). Synaptic scaffold evolution generated components of vertebrate cognitive complexity. Nat. Neurosci. 16, 16–24. doi: 10.1038/nn.3276
Parsegian, A., and See, R. E. (2014). Dysregulation of dopamine and glutamate release in the prefrontal cortex and nucleus accumbens following methamphetamine self-administration and during reinstatement in rats. Neuropsychopharmacology 39, 811–822. doi: 10.1038/npp.2013.231
Proitsi, P., Kim, M., Whiley, L., Simmons, A., Sattlecker, M., Velayudhan, L., et al. (2017). Association of blood lipids with Alzheimer’s disease: a comprehensive lipidomics analysis. Alzheimers Dement. 13, 140–151.
Reggiani, C., Coppens, S., Sekhara, T., Dimov, I., Pichon, B., Lufin, N., et al. (2017). Novel promoters and coding first exons in DLG2 linked to developmental disorders and intellectual disability. Genome Med. 9:67.
Rist, M. J., Roth, A., Frommherz, L., Weinert, C. H., Kruger, R., Merz, B., et al. (2017). Metabolite patterns predicting sex and age in participants of the Karlsruhe Metabolomics and Nutrition (KarMeN) study. PLoS One 12:e0183228. doi: 10.1371/journal.pone.0183228
Rodríguez Cerdeira, C., Sánchez-Blanco, E., Sánchez-Blanco, B., and González-Cespón, J. L. (2017). Protein biomarkers of mood disorders. Int. J. Immunopathol. Pharmacol. 30, 7–12.
Ruehle, S., Rey, A. A., Remmers, F., and Lutz, B. (2012). The endocannabinoid system in anxiety, fear memory and habituation. J. Psychopharmacol. 26, 23–39. doi: 10.1177/0269881111408958
Shima, N., Miyawaki, I., Bando, K., Horie, H., Zaitsu, K., Katagi, M., et al. (2011). Influences of methamphetamine-induced acute intoxication on urinary and plasma metabolic profiles in the rat. Toxicology 287, 29–37. doi: 10.1016/j.tox.2011.05.012
Su, H., Chen, T., Jiang, H., Zhong, N., Du, J., Xiao, K., et al. (2020). Intermittent theta burst transcranial magnetic stimulation for methamphetamine addiction: a randomized clinical trial. Eur. Neuropsychopharmacol. 31, 158–161. doi: 10.1016/j.euroneuro.2019.12.114
Su, H., Zhong, N., Gan, H., Wang, J., Han, H., Chen, T., et al. (2017). High frequency repetitive transcranial magnetic stimulation of the left dorsolateral prefrontal cortex for methamphetamine use disorders: a randomised clinical trial. Drug Alcohol Depend. 175, 84–91. doi: 10.1016/j.drugalcdep.2017.01.037
Su, M. F., Liu, M. X., Li, J. Q., Lappin, J. M., Li, S. X., Wu, P., et al. (2018). Epidemiological characteristics and risk factors of methamphetamine-associated psychotic symptoms. Front. psychiatry 9:489. doi: 10.3389/fpsyt.2018.00489
Tasic, L., Pontes, J. G. M., Carvalho, M. S., Cruz, G., Dal Mas, C., Sethi, S., et al. (2017). Metabolomics and lipidomics analyses by1H nuclear magnetic resonance of schizophrenia patient serum reveal potential peripheral biomarkers for diagnosis. Schizophr. Res. 185, 182–189. doi: 10.1016/j.schres.2016.12.024
Wu, H. C., Chen, C. M., Chen, Y. C., Fung, H. C., Chang, K. H., and Wu, Y. R. (2018). DLG2, but not TMEM229B, GPNMB, and ITGA8 polymorphism, is associated with Parkinson’s disease in a Taiwanese population. Neurobiol. Aging 64, 158.e1–158.e6.
Yang, W., Yang, R., Luo, J., He, L., Liu, J., and Zhang, J. (2018). Increased absolute glutamate concentrations and glutamate-to-creatine ratios in patients with methamphetamine use disorders. Front. psychiatry 9:368. doi: 10.3389/fpsyt.2018.00368
Yang, X., Wang, Y., Li, Q., Zhong, Y., Chen, L., Du, Y., et al. (2018). The main molecular mechanisms underlying methamphetamine- induced neurotoxicity and implications for pharmacological treatment. Front. Mol. Neurosci. 11:186. doi: 10.3389/fnmol.2018.00186
Zhang, K., Wang, Q., Jing, X., Zhao, Y., Jiang, H., Du, J., et al. (2016). miR-181a is a negative regulator of GRIA2 in methamphetamine-use disorder. Sci. Rep. 6:35691.
Zhen, L., Shao, T., Luria, V., Li, G., Li, Z., Xu, Y., et al. (2018). EphB2 deficiency induces depression–like behaviors and memory impairment: involvement of NMDA 2B receptor dependent signaling. Front. Pharmacol. 9:862. doi: 10.3389/fphar.2018.00862
Zheng, C.-Y., Seabold, G. K., Horak, M., and Petralia, R. S. (2011). MAGUKs, synaptic development, and synaptic plasticity. Neuroscientist 17, 493–512. doi: 10.1177/1073858410386384
Zheng, P., Fang, Z., Xu, X.-J., Liu, M.-L., Du, X., Zhang, X., et al. (2016). Metabolite signature for diagnosing major depressive disorder in peripheral blood mononuclear cells. J. Affect. Disord. 195, 75–81. doi: 10.1016/j.jad.2016.02.008
Zheng, T., Liu, L., Shi, J., Yu, X., Xiao, W., Sun, R., et al. (2014). The metabolic impact of methamphetamine on the systemic metabolism of rats and potential markers of methamphetamine abuse. Mol. Biosyst. 10, 1968–1977. doi: 10.1039/c4mb00158c
Zhong, N., Jiang, H., Du, J., Zhao, Y., Sun, H., Xu, D., et al. (2016). The cognitive impairments and psychological wellbeing of methamphetamine dependent patients compared with health controls. Prog. Neuropsychopharmacol. Biol. Psychiatry 69, 31–37. doi: 10.1016/j.pnpbp.2016.04.005
Keywords: methamphetamine abuse, metabolomics, bioinformatics, regulatory network, glutamate metabolic pathway
Citation: Peng S, Su H, Chen T, Li X, Du J, Jiang H and Zhao M (2021) The Potential Regulatory Network of Glutamate Metabolic Pathway Disturbance in Chinese Han Withdrawal Methamphetamine Abusers. Front. Genet. 12:653443. doi: 10.3389/fgene.2021.653443
Received: 14 January 2021; Accepted: 01 March 2021;
Published: 23 March 2021.
Edited by:
Cunyou Zhao, Southern Medical University, ChinaCopyright © 2021 Peng, Su, Chen, Li, Du, Jiang and Zhao. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Min Zhao, ZHJtaW56aGFvQHNtaGMub3JnLmNu