 Takumi Kawasaki
Takumi Kawasaki Moe Ikegawa
Moe Ikegawa Taro Kawai
Taro Kawai- Laboratory of Molecular Immunobiology, Division of Biological Science, Graduate School of Science and Technology, Nara Institute of Science and Technology (NAIST), Ikoma, Japan
The lungs are constantly exposed to environmental and infectious agents such as dust, viruses, fungi, and bacteria that invade the lungs upon breathing. The lungs are equipped with an immune defense mechanism that involves a wide variety of immunological cells to eliminate these agents. Various types of dendritic cells (DCs) and macrophages (MACs) function as professional antigen-presenting cells (APCs) that engulf pathogens through endocytosis or phagocytosis and degrade proteins derived from them into peptide fragments. During this process, DCs and MACs present the peptides on their major histocompatibility complex class I (MHC-I) or MHC-II protein complex to naïve CD8+ or CD4+ T cells, respectively. In addition to these cells, recent evidence supports that antigen-specific effector and memory T cells are activated by other lung cells such as endothelial cells, epithelial cells, and monocytes through antigen presentation. In this review, we summarize the molecular mechanisms of antigen presentation by APCs in the lungs and their contribution to immune response.
Introduction
The lung is the peripheral tissue that exchanges gas during respiration; therefore, it is exposed to the outer environment, which potentially increases the risk of invasion by viral and bacterial pathogens. Respiratory viruses, including influenza virus and recent coronavirus, induce inflammation and tissue damage, leading to disorders of the lungs. The high infectivity and spreadability of these viruses have caused a worldwide pandemic in recent years and has provoked the argument for recurrent infection and efficacy of vaccination in order to suppress the pandemic. Innate immune cells such as dendritic cells (DCs) and macrophages (MACs) in the lungs form the first line of defense by recognizing the molecular structures common to pathogens, called pathogen-associated molecular patterns, through pattern recognition receptors (1, 2). During the past decade, various types of lung DCs and MACs have been identified and classified according to surface markers, expression genes, and corresponding transcription factors with specialized functions. These DCs and MACs function as antigen-presenting cells (APCs) that engulf pathogens through endocytosis or phagocytosis and present their peptides on major histocompatibility complex class I (MHC-I) or MHC-II protein complex to naïve CD8+ or CD4+ T cells, respectively. Although DCs and MACs are known as professional APCs with a higher expression of co-stimulatory molecules, such as CD80 and CD86, other types of cells such as monocytes and epithelial cells in the lungs also have the potential to present antigens to T cells.
APCs load peptides derived from exogenous antigens on MHC-II and present peptide-MHC-II complex to CD4+ T cells whereas APCs load peptides derived from both endogenous and cytosolic antigens on MHC-I and present peptide-MHC-I complex to CD8+ T cells (Figures 1, 2). In addition, specific APCs take up exogenous antigens, process them, and load peptides onto MHC-I to CD8+ T cells, a process called antigen cross-presentation (3). Lung DCs are largely divided into three major subsets: cDC1s, cDC2s, and plasmacytoid DCs (pDCs). These DCs have been focused on as key regulators of T cell responses (4); however, recent evidence indicates that other types of cells in the lung, such as MACs, monocytes, and epithelial cells, also have antigen presentation capacity to both CD4+ and CD8+ T cells. MACs in the lung are mainly classified into alveolar macrophages (AMs) and interstitial macrophages (IMs). Lung epithelial cells (LECs) consist of alveolar type I (ATI) and alveolar type II (ATII) cells in the alveoli, and the predominant cell types constituting the bronchial airway epithelium include endothelial cells, basal progenitor cells, ciliated cells, secretory club cells, and goblet cells (5, 6). Lung DCs, MAC and LECs express MHC-I and/or MHC-II on their cell surface and potentially present antigen to CD4+ or CD8+ T cells (7).
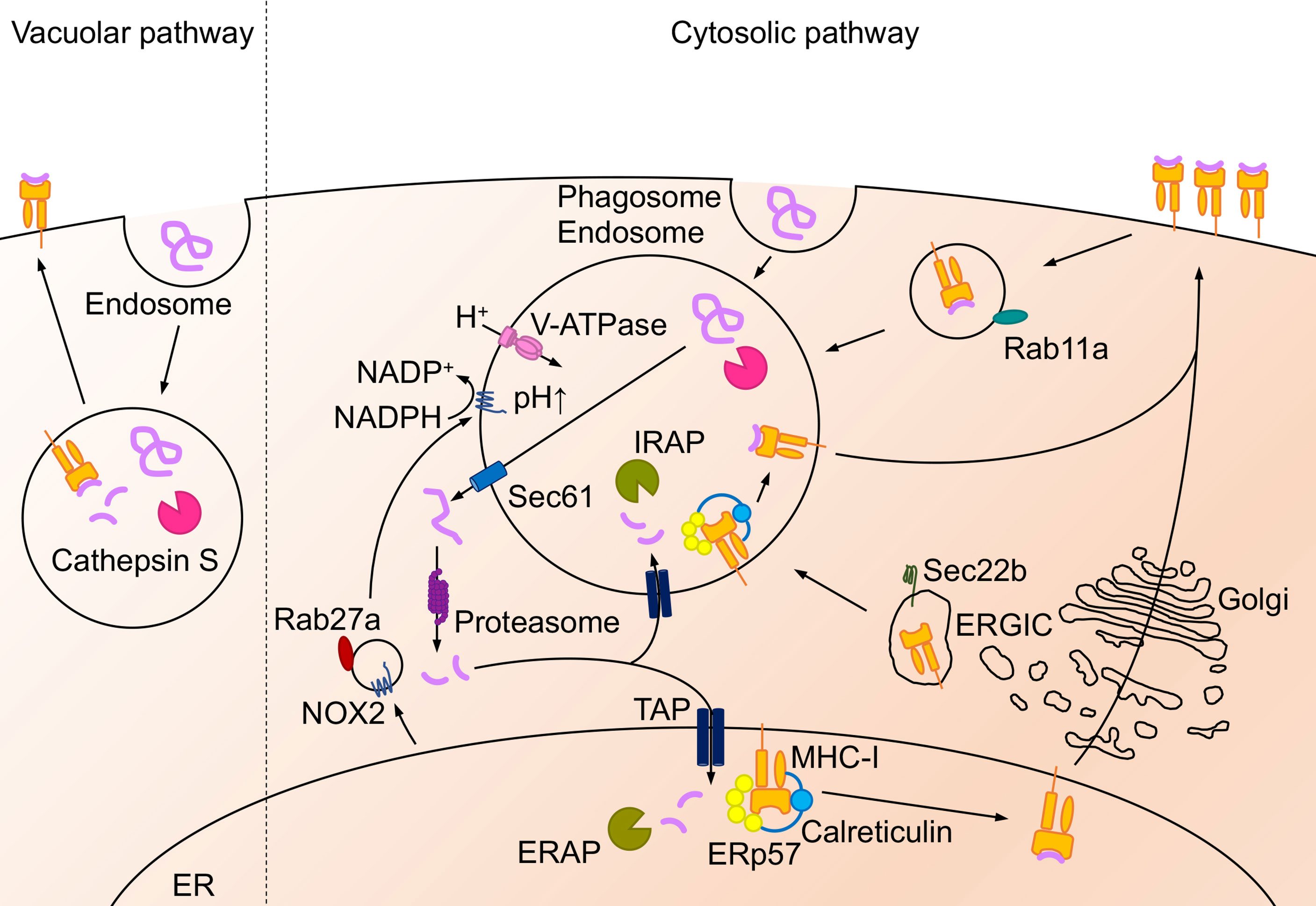
Figure 1 Antigen presentation on MHC-II molecule. Extracellular antigens are endocytosed or phagocytosed, and intracellular antigens are translocated to the late-endosome or the lysosome via autophagosome- or LAMP-2A- mediated autophagy. Then these antigens are degraded by asparaginyl endopeptidase and cathepsin. MHC-II is synthesized in ER and mainly pooled at the plasma membrane as MHC-II-Ii chain complex. When the complex translocates from the ER or the plasma membrane to the acidic compartment, Ii chain is degraded into CLIP and driven out by interaction with H2-M. Afterward, antigen peptides bind to the MHC-II and the peptide-MHC-II complex exports to the cell surface.
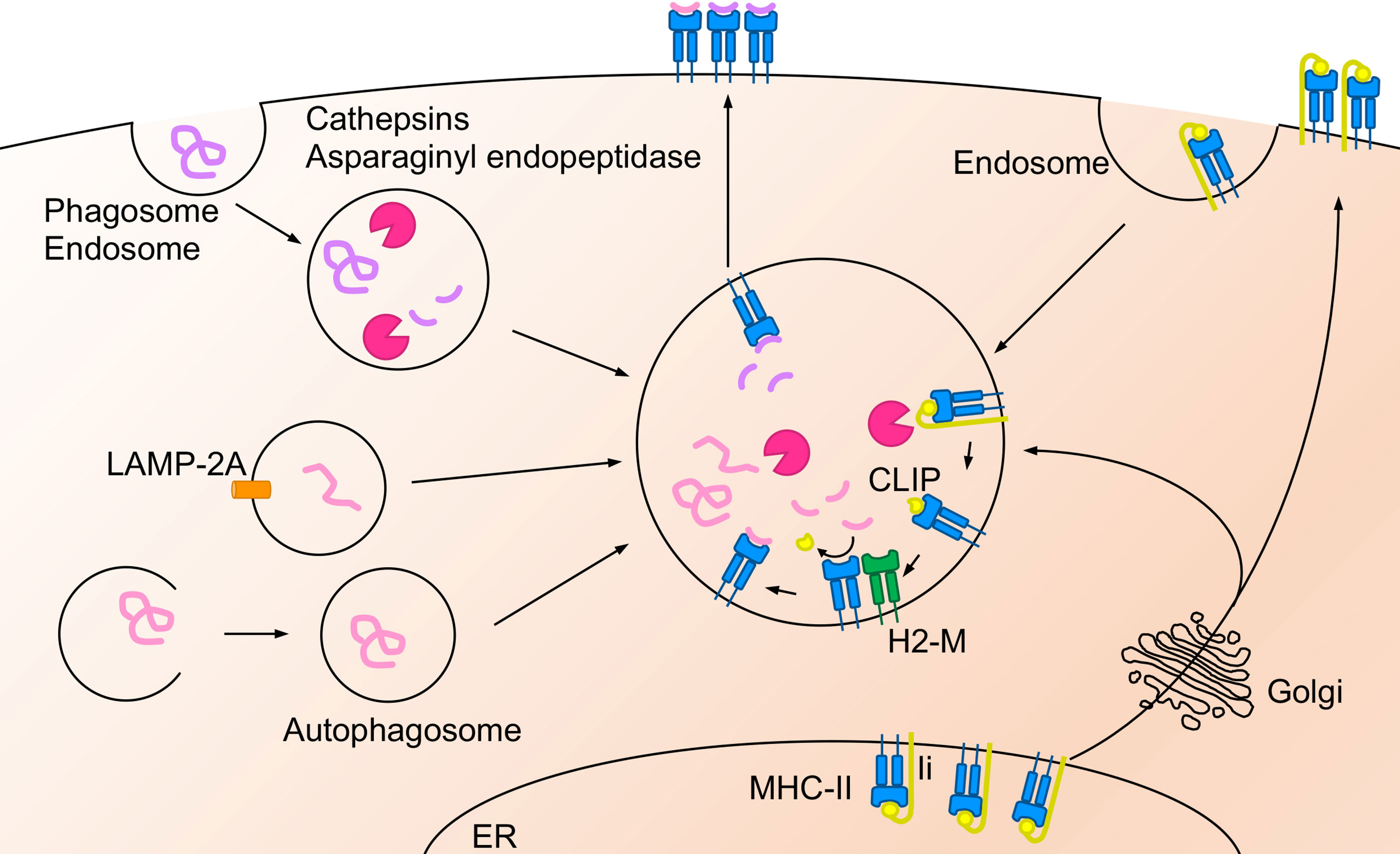
Figure 2 Antigen cross-presentation on MHC-I molecule. Extracellular antigens are presented via “vacuolar pathway” or “cytosolic pathway” in the cross-presentation pathway. In the vacuolar pathway, endocytosed antigen peptides are degraded by cathepsin S and bind to MHC-I in the endosomal compartment. In the cytosolic pathway, endocytosed or phagocytosed extracellular antigens are translocated to the cytosol via Sec61 and degraded by proteasome. The degraded peptides are transported into the ER or the endosome via TAP and trimmed by ERAP (in the ER) or IRAP (in the endosomes). TAP form PLC with MHC-I, ERp57 and calreticulin. Afterward, the trimmed peptides bind to the MHC-I and transported to the cell surface. The MHC-I in the endosomes is recruited from the plasma membrane through Rab11a+ recycle endosome, the ER, or the ERGIC. Antigen degradation regulated by the acidification in the endosome, the phagosome, and the lysosome by V-ATPase. On the other hand, NADPH oxidase NOX2 regulates phagosomal alkalization and is recruited to the phagosomes by Rab27a-dependent pathway.
During pathogen infection in the lung, pathogen-specific CD4+ and CD8+ cells are primed in the lung-draining lymph nodes by antigen-presenting DCs that migrate from the infected area in the lung (8, 9). Antigen-presenting DCs encounter naïve CD4+ and CD8+ T cells in the lymph nodes, where antigen-specific T cells are selected, and the proliferation and differentiation by antigen presentation on MHC molecules are induced along with the assistance of co-stimulatory molecules and the local cytokine environment (10, 11). Antigen-specific CD4+ and CD8+ T cells in the lymph nodes migrate to the lungs to directly eliminate infected cells or induce the accumulation of other immunological cells for pathogen clearance. In addition, antigen-specific T cells encounter local APCs in the lungs, including DCs, MACs, monocytes, and LECs, and further differentiate and expand in the lung (12). Parts of antigen-specific cells differentiate into long-lived memory cells, which are divided into three types of population: central memory T (TCM) cells, which are largely found in secondary lymphoid organs; effector memory T (TEF) cells, which systematically circulate, transiently entering peripheral tissue, and resident memory T (TRM) cells, a non-circulating, self-renewing population located in peripheral tissues including the lungs (13, 14). There has been increasing evidence that antigen-specific memory T cell formation through antigen presentation or cytokines is facilitated by various types of lung cells. In this review, we summarize the molecular mechanisms of antigen presentation to MHC-I and MHC-II on APCs and memory T cell formation by APCs during pathogen infection in the lung.
Molecular Basis of Antigen Presentation to CD4+ T Cells
In general, extracellular antigens are endocytosed or phagocytosed by APCs and degraded by proteases such as asparaginyl endopeptidase (15) and cathepsins S, B, H, and L (16–18). Degraded peptides are ultimately presented on MHC-II molecules to prime CD4+ T cells (19) (Figure 1). However, less than 30% of antigens on MHC-II are derived from endogenous antigens, such as cytoplasmic or nuclear antigens (20, 21). Regardless of peptides derived from self or non-self-antigens, these peptides can be presented by APCs, non-professional APCs, or tumor cells mainly via autophagosome- or chaperone-mediated autophagy (22). Antigen degradation is mediated by the fusion of autophagosomes with endosomes and lysosomes in autophagosome-mediated autophagy (23). Antigens degraded by the proteasome in the cytosol are translocated to the late endosome or lysosome, which is enhanced by lysosome-associated membrane protein 2A (LAMP-2A) (24).
Newly synthesized MHC-II forms a complex with the invariant (Ii) chain in the endoplasmic reticulum (ER), and is pooled in the ER or plasma membrane and then respectively, translocated to the endosomes and lysosomes either directly (25) or indirectly though endocytosis (26, 27); however, the complex cannot bind to antigen peptides (28, 29). The Ii chain is degraded into a small fragment called class II-associated Ii chain peptide (CLIP) and binds to MHC-II in the late-endosome or the lysosome (30). The CLIP on MHC-II is driven out by interaction with another nonconventional MHC-II, called HLA-DM in humans and H2-M in mice (30). Then, MHC-II complexes can bind to antigen peptides and be presented on the cell surface (30). The expression of the peptide-MHC-II complex on the cell surface and its turnover by ubiquitination in DCs is essential for their ability to efficiently prime CD4+ T cells (31, 32).
Molecular Basis of Antigen Cross-Presentation Pathway
Specific APCs are thought to take up extracellular antigens through endocytosis or phagocytosis and load peptides onto MHC-I for presentation to CD8+ T cells, a process called antigen cross-presentation (3). The extracellular antigen degradation pathway is mainly divided into the “vacuolar pathway”, through which the peptide is degraded in the endosome, and the “cytosolic pathway” which is responsible for the transport of degraded protein through SEC61 from the endosome to the cytosol (33) (Figure 2).
Vacuolar Pathway of Antigen Cross-Presentation
In the vacuolar pathway, extracellular antigens are endocytosed by APCs and degraded into peptide fragments by proteases in the compartment. Cathepsin S plays a crucial role in antigen degradation in the endosomes of bone marrow-derived DCs (BMDCs) (34). It has been shown that cathepsin S plays a key role in priming CD8+ T cells to Influenza A virus (IAV) peptides loaded on MHC-I in the vacuolar pathway (34). In DCs, cathepsin S is also a crucial protease for MHC-II-dependent presentation to CD4+ T cells (18, 35) whereas cathepsin L in the thymic cortical epithelium (35) and cathepsin F in macrophages likely correspond to proteases in the vacuolar pathway (36). The degraded peptide by cathepsins forms a complex with MHC-I in the endosome, and the peptide-MHC-I complex is transported to the cell surface. However, it is not clear whether cathepsins are required for antigen degradation in all lung APCs during pathogen infection.
Cytosolic Pathway of Antigen Cross-Presentation
In the cytosolic pathway, phagocytosed or endocytosed antigens are translocated from the endosomal compartments to the cytosol via Sec61 (33) and degraded to peptide fragments by the proteasome in the cytosol (37, 38). Phagosomes and endosomes are mainly acidified via V-ATPase for degradation (39), which is regulated by Toll-like receptor (TLR) signals and other maturation signals (40), and restriction of antigen in these compartments by acidification is important for peptide degradation in the cytosolic pathway. DCs lacking the NADPH oxidase NOX2 show enhanced phagosomal acidification and increased antigen degradation, resulting in impaired antigen presentation (41, 42). The recruitment of NOX2 to these compartments is prevented by deficiency of Rab27a, which causes acidification of phagosomes, limiting antigen degradation (43).
The cytosolic pathway is further categorized to two pathways; “ER-dependent pathway” and “Endosomal pathway”. The ER-dependent pathway is the most common route to ER for antigen peptide. Antigen peptides in the cytosol are transported into the ER mainly through transporter associated with antigen processing (TAP) and form peptide-MHC-I complexes in the ER. On the other hand, peptides degraded by the proteasome in the cytosol are transported back to the endosomes through TAP in the endosomal pathway. MHC-I molecules are recycled in the cells. MHC-I molecules in the endosome are transported from the plasma membrane through the Rab11+ recycling endosomes (44) and are also recruited from the ER or the ER-Golgi intermediate compartment (ERGIC) (3, 45). Transported peptides are loaded on MHC-I by the peptide loading complex (PLC) the in the ER or the endosomes (46). PLC consists of TAP, oxidoreductase ERp57, MHC-I heterodimer, and calreticulin (46). PLC is recruited to phagosomes or endosomes via the Sec22b-ERGIC pathway (47). PLC is also recruited from the recycle endosomes after TLR activation (44). In contrast, the N terminal anchor residues of the peptides are trimmed by ER-resident N-aminopeptidases (ERAP1 and ERAP2 in humans, and ERAAP in mice). Insulin regulated aminopeptidase (IRAP), an aminopeptidase similar to ERAP, trims the peptide in the endosomes (48, 49). These peptide trimming proteins are crucial for efficient antigen peptide binding to MHC-I and contribute to cross-presentation (50–52). Although cytosolic peptides shuttle into the ER through TAP1 in the cytosolic pathway, TAP1 blockade in DCs leads to antigen presentation by MHC-I translocation from ERGIC in a Sec22b-dependent manner rather than the Rab11+ recycle-endosome pathway (53).
DCs and MACs in the Lung
DCs in the lung consist of heterogeneous subsets that exert different functions (54, 55). Lung DCs are largely divided into three major subsets and are broadly subdivided into plasmacytoid DCs (pDCs) and conventional DCs (cDCs). Murine cDCs express high levels of integrin CD11c and are further divided into CD103+ DC and CD11b+ DCs. CD103+ DCs and CD11b+ DCs are also referred to as cDC1s and cDC2s, respectively (55–58). Although CD11b and CD11c have been utilized for the separation of DC population, cDCs separation was proposed as two main subsets cDC1s and cDC2s based on the transcription factor expression (59, 60). Interferon regulatory factor 8 (IRF8) and Batf3 drive the development of cDC1s which are separated as XCR1+Cadm1+CD172a− cDC1s (61–69). On the other hand, IRF4 drives the development of cDC2 which are separated as XCR1−Cadm1−CD172a+ cDC1s (67, 69–76). pDCs develop in the presence of transcription factor 4 (E2-2) and the Ets family transcription factor Spi-B (77–79). In the steady-state, cDC1s associate with airway rather than alveoli in the lung (80, 81). cDC2s are located in the airway and lung parenchyma (82–84). Monocyte-derived DCs (moDCs) have been described as another DC population that accumulates in the lungs during inflammation and viral infection (85–87). MoDCs are also known as inflammatory DCs and monocyte derived cells (88–91). These DCs are subdivided based on the presence of surface markers and recent progress in the technology for single-cell RNA sequencing revealed that the cDC2s population in the lung is subdivided based on expression markers with functional differences, whereas pDCs and cDC1s are a unique population (92–94).
MACs in the lungs consist of two major populations: alveolar MACs (AMs) and interstitial MACs (IMs). AMs are located in the alveolar space of the lungs and are in close contact with the type I and II epithelial cells of the alveoli. AMs are the first line of defense against pathogens for host defense in the lung, with a higher engulfment capacity against antigens and pathogens (95). AMs produce cytokines such as TGFβ, IL6, and type I interferon during pathogen infection and inflammation (95, 96). In addition, AMs play a central role in homeostasis and tissue remodeling. Pulmonary surfactant is a mixture of lipids and proteins secreted into the alveolar space by AT II cells. The surfactant is covered with an interface of alveolar epithelial cells in the lungs to reduce the physical tension during breathing. In addition, the engulfment of surfactant and cell debris by AMs is important for the clearance and maintenance of lung homeostasis. Accumulation of pulmonary surfactant in the absence of AMs causes the development of pulmonary alveolar proteinosis (PAP) (97–99). GM-CSF and TGF-β induce PPAR-γ, a crucial transcription factor for AM development (100). Interstitial macrophages (IMs) reside in the parenchyma between the microvascular endothelium and alveolar epithelium. However, compared with AMs, the role of IMs in lung homeostasis remains poorly understood. Like AMs, IMs engulf bacteria and foreign particles and secrete IL-1, IL-6, IL-10, and TNFα (101–104). IMs form a heterogeneous population that is further subdivided based on surface markers with distinct functions (103, 105).
The lung is composed of a complex tissue structure that exchanges gas and is exposed to outer space. The combination of crosstalk between DCs and MACs effectively protects against inhaled pathogens by inducing acquired immunity (Figure 3). cDC1s, cCD2s and IMs express high levels of MHC I/II with co-stimulatory molecules CD80 and CD86 (106). However, AMs express lower levels of MHC-II. Based on the expression of molecules for antigen presentation, it is revealed that each cells display antigen presentation capacity against specific infectious pathogens and allergic materials in the lungs.
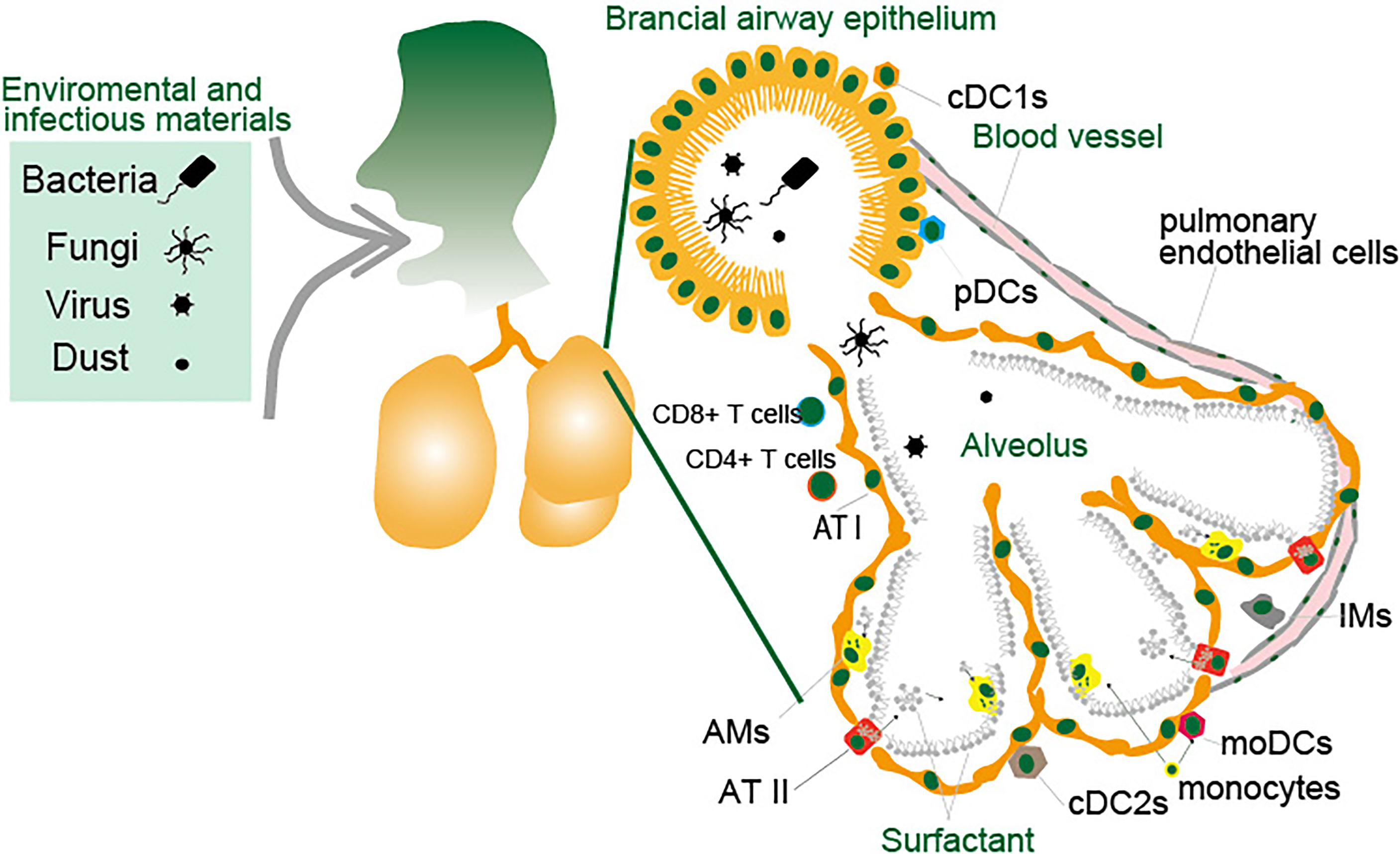
Figure 3 Antigen presenting cells in the lung. The lungs are constantly exposed to environmental and infectious agents such as dust, viruses, fungi, and bacteria that invade the lungs upon breathing. The lungs are protected by various types of immune cells and epithelial cells. Lung DCs are largely divided into three major subsets and are broadly subdivided into pDCs, cDC1s and cDC2s. MACs in the lungs consist of two major populations: AMs and IMs. LECs consist of ATI and ATII cells in the alveoli, and the endothelial cells and other types of cells constituting the bronchial airway epithelium. Monocytes migrate to the lungs in response to inflammatory stimuli in a CCR2-dependent manner and these cells differentiate to moDCs or AMs. Small blood vessels allow oxygen to be extracted from the air into the blood, and carbon dioxide to be released from the blood into the air. The cells lining the inner surface of blood vessels are the pulmonary endothelial cells. These cells function as APCs that engulf pathogens through endocytosis or phagocytosis and present their peptides on major MHC-I or MHC-II protein complex to CD8+ or CD4+ T cells.
Antigen Presentation by pDCs
pDCs are professional cells that secrete type I IFN through the stimulation of innate immune receptors. It is widely accepted that the production of type I IFN by pDCs in the lungs is important for host defense against pathogens. An Aspergillus fumigatus infection model in the lung demonstrated that pDCs are essential for host defense and neutrophil effector activity (107). Antigen presentation by pDCs in the lungs is controversial during pathogen infection. Resting pDCs are weak antigen-presenting cells, but appear to be functionally specialized for their ability to capture and present viral antigens to CD4+ T cells in the presence of CpG DNA or virus stimulation (108, 109). Transplantation of pDCs in an IAV infection model showed that pDCs infected with IAV promote antigen presentation to CD8+ T cells (110). In contrast, ablation of pDCs does not have a significant impact on the production of IAV-specific CD8+ T cells and viral clearance, indicating that pDCs have weak or no antigen cross-presentation capacity in vivo (111). Other groups have shown that pDCs in other peripheral tissues cooperate with cDC2s to promote their maturation and cross-presentation activity and induce antiviral CD8+ T cells, suggesting that pDCs indirectly induce antigen-specific CD8+ T cells (112, 113).
Antigen Presentation by cDC2s
cDC2s are localized in the lungs under a steady-state condition, and a large number of cDC2s are accumulated in the lungs in response to inflammation induced by viral infection (114) or antigen immunization (115). IAV infection induces accumulation of cDC2s, and the depletion of these cells reduces the number of virus-specific CD8+ cells and mortality (85–87). These results indicate that accumulated cDC2s migrate to the lymph nodes and present antigens to CD8+ T cells. However, cDC1 analysis using Batf3-deficient mice indicated that cDC2s have a weak cross-presentation capacity in vivo and support the proliferation of CD8+ T cells in the lung during IAV infection (116). Initial antigen-specific T cell differentiation is induced in the tissue-draining lymph nodes, and lung cDC2s are less migratory than cDC1s (117). During the inflammation, cDC2s in the lungs have shown to prime CD4+ Th2 cells but not CD8+ T cells responses (69, 75, 76). cDC2s also have shown to prime CD4+ Th17 cells response during Aspergillus fumigatus infection (74). T follicular helper (Tfh) cells are a subset of CD4+ T cells that promote antibody production during vaccination. cDC2s carry antigen into the lymph node where cDC2-dependent Tfh cells prime antibody-mediated protection from IAV challenge (67). cDC2s also locate in lymphoid organ, skin intestine and others organs as same with lung cDC2s, and cDC2s in the other organs efficiently promote the differentiation of CD4+ T cells into effector helper T cells during infection with Nippostrongylus brasiliensis, Aspergillus fumigatus or Citrobacterior rodentium (70–73). These results suggest that cDC2s are more specialized in polarizing CD4+ T helper cell responses and providing help to B cells, rather than in inducing CD8+ T cells activation.
cDC2s consist of heterogeneous subpopulations although it is unclear whether the same subpopulation of cDC2s induces both Th2 and Th17 cells (71, 74, 94). Single-cell RNA and cytometry by time-of-flight (CyTOF) analyses revealed that cDC2s consist of five distinct clusters. Ly-6C+CD301b– cDC2s promote Th17 differentiation, and CD200+cDC2s induce the differentiation of Th2 but not Th17 cells (94). In addition, there are conflicting reports on how moDCs and CD11b+DCs interact with and regulate T cell responses (118). A recent report indicated that inflammatory cDC2s (inf-cDC2s) express the Fc receptor CD64 shared with moDCs and IRF8 shared with cDC1s and are infiltrated to present antigen to CD4+ and CD8+ T cells during respiratory virus infection (92). TNFR2− cDC2 subpopulation drives moDCs maturation to generate T follicular helper (Tfh) cells in the lung (119).
Antigen Cross-Presentation by cDC1s
Many studies have shown the importance of cDC1s in the initiation of antiviral T cell response following influenza infection. Particular subsets of cDC1s, such as CD8α+ and CD103+ cDC1s, play specific roles in naïve T cell activation and differentiation (10, 120–122). CD8α+ cDC1s in the spleen and lymphoid organs are known as the cross-presenting subset (123–125). CD103+ cDC1s are migratory DCs that cross-present antigens in peripheral tissues, including the lungs (126, 127). Both CD103+ cDC1s in the lungs and CD8α+ cDC1s in lymph nodes share the expression of various genes, including transcription factors IRF8, BATF3, and ID2, and both of these DC subtypes are developed in the presence of Flt3 (128).
cDC1s directly present antigen to naive CD4+ T cells (129) and cDC1s could prime Th2 and Th17 differentiation by producing IL4, IL12, IL13 and IL17 induction during allergic airway inflammation (130, 131). A mouse model of invasive pulmonary aspergillosis infection showed cDC1s induces Th17 response by producing IL-2 in the lung (132). Other reports postulate that cDC1s promote airway tolerance by the induction of FoxP3+ Tregs in antigen induced airway inflammation (133) or by inducing IL-10 without Treg-induction (134). Although cDC1s can present antigens and stimulate CD4+ T cells, they are well known for their ability to cross-present antigens to CD8+ T cells (127, 132). Lung cDC1s preserve viral antigens in their endocytic compartments and control the induction of virus-specific CD8+ T cells through antigen cross-presentation (116, 135). Lung cDC1s migrate to mediastinal LNs after viral infection, where they directly present antigens to naïve CD8+ T cells or transfer captured antigens to CD8α+ cDC1s, which present antigens and activate naïve CD8+ T cells (86, 136, 137). In addition to cDC1s, cDC2s have the potential to migrate to mediastinal LNs (MLNs) (117), however, cDC2s do not present antigens efficiently in the MLNs (138). The cytotoxic activity of CD8+ T cells plays a critical role in viral clearance in the lungs. Initial virus-specific CD8+ T cells in the LNs are induced by cDC1s migrating from the infected lung, and the virus-specific CD8+ T cells then traffic back to the infected lung to mediate their effector function (10, 11, 139).
Antigen Presentation by moDCs
Chemokine receptor CCR2- and Ly6C-expressing inflammatory monocytes infiltrate into the lung during pathogen infection including Aspergillus fumigatus (140) and IAV (141), and differentiate rapidly into moDCs. MoDCs in other organs are also capable of presenting antigen and priming to CD4+ T cells (142, 143) and CD8+ T cells (88). However, the precise function of moDCs to regulate T cells response in lung is controversial. CCR2-deficient mice impair moDCs recruitment and exhibit reduction of effecter CD8+ T cell response in the lung after IAV infection (85). moDCs depletion by CD11c-cre-Irf4f/f mice reduces CD8+ memory precursor cells and TRM cells during IAV infection (144). MoDCs in the lung prime IFN-γ-producing antigen-specific CD4+ T cells in pulmonary aspergillosis (140). MoDCs also promote Th1 and Th17 cell polarization through antigen presentation during allogeneic responses (118) and induce Th2 type CD4+ cells during house dust mite allergy (145). Report using CD26 as a maker for separation of moDCs indicated that moDCs have poor capacity to migrate to lymph node and prime CD4+ T cells and CD8+ T cells (92, 117, 146).
Antigen Presentation by Macrophages
AMs develop during embryogenesis, and then predominantly maintain their populations by self-renewal (147–149) and are specialized in the removal and recycling of surfactant molecules. Although AMs are the most abundant immune cells in the lungs and have been suggested to play a functional role in antigen presentation during tuberculosis and Cryptococcus neoformans infection in humans (150, 151), supportive evidence for antigen presentation by AMs has not been reported in mice. Certain IM subsets have been contributed to lung immune homeostasis by spontaneously producing the immunosuppressive cytokine IL-10 and preventing the development of aberrant type 2 allergic responses against inhaled allergens (101, 104). IMs are separated by a distinct subpopulation based on the surface expression pattern (103, 152) and single-cell RNA sequencing (153, 154), some of which express antigen-presenting genes and may mediate antigen presentation to CD4+ T cells in the lungs. Accumulated Ly-6C+ monocytes develop to exudative macrophages (exMACs) during Cryptococcus (155), Streptococcus (156) and IAV infection (157). ExMACs produce high levels of TNF-α and NOS2 and stimulate the proliferation of memory CD4+ T cells (157).
Antigen Presentation by Monocytes
Two types of monocytes have been identified with different phenotypes and functions: Ly6C+ classical monocytes and Ly6C− non-classical monocytes. Ly6C+ monocytes constitutively enter to lung tissues in the steady state and a large number of these cells migrate to the lungs in response to inflammatory stimuli in a CCR2-dependent manner (158, 159). Ly6C+ monocytes develop to moDCs, IMs, exMACs or monocyte-derived AMs in the lungs during inflammatory stimulation, but in the steady state, monocytes continuously migrate to non-lymphoid organs including lung without differentiating into other types of cells and may exit lung via the lymphatics or undergo local apoptosis and cleared (160). Ly6C+ monocytes have been shown to produce large amounts of IL-1, IL-6, and TNFα, and have an ability to drive adaptive immune responses through antigen presentation (160). Ly6C+ monocytes in other tissues reported that these cells have an ability to present antigen to both CD4+ and CD8+ cells. Ly6C+ monocytes regulate early host response to Aspergillus lung infection by taking up conidia and trafficking them into the draining LN to prime CD4+ T cells (140). Cross-presentation by Ly6C+ inflammatory monocytes in lymphoid organs has been reported in the presence of TLR agonists, especially TLR7 (161). Once recruited into the lungs, Ly6C+ monocytes further differentiate into moDCs and monocyte-derived AMs. Recent evidence have shown that CCR2-deficient mice, which are defective in monocyte trafficking to the lung, exhibit decreased number of virus-specific lung resident memory CD8+ (TRM) cells by the antigen presentation on monocytes (162).
Antigen Presentation by Epithelial and Endothelial Cells
As lung epithelial cells directly interact with the external environment, these cells are thought to be critical regulators of barrier immunity (163, 164). The alveoli are composed of two distinct lung epithelial cell types: AT I cells, which are thin and cover approximately 95% of the internal surface of the lung, and AT II cells, which are cuboidal secreting cells located between type I cells (165). AT I cells are specialized in gas exchange and alveolar fluid regulation, whereas type II cells secrete surfactants and constitute the progenitor cells of the epithelium (166). There is increasing evidence that epithelial cells in the lung contribute to adaptive immune responses in the lungs. AT II cells express MHCII and present antigen. In vitro co-culture experiments AT II cells with antigen specific hybridoma suggested that AT II cells activate CD4+ cells to induce IFNγ in the presence of peptide antigen, and deletion of MHC-II on AT II cells results in a modest worsening of respiratory virus disease following influenza and Sendai virus infections (167). Surfactant Protein C (SPC)lowMHC-IIhigh AT II cells function as APCs to induce CD4+ TRM cells (7). Antigen presenting AT II cells primes naïve CD4+ T cells in vitro and induce regulatory T (Treg) cells (168); however, it is unclear whether AT II cells prime naïve CD4+ T cells in vivo (169). In addition to CD4+ T cells activation, barrier epithelial cells recruit and maintain CD8+ TRM cells near the sites of antigen encounter and reactivate them in the tissues via local antigen presentation (12, 170).
Small blood vessels, known as capillaries, come in close contact with the alveoli, allowing oxygen to be extracted from the air into the blood, and carbon dioxide to be released from the blood into the air. The cells lining the inner surface of these capillaries are known as the pulmonary endothelial cells (171). Lung endothelial cells cross-present malaria antigen to antigen specific reporter cells in vitro and a mouse model of malaria infection by Plasmodium berghi ANKA (PbA) induces IFNγ positive CD8+ T cell. These results demonstrate that lung endothelial cells cross-present malaria antigen to CD8+ T cells, although it is unclear whether these cells activate naive CD8+ T cell in vivo (172).
Perspective and Conclusion
Lungs are protected by various types of APCs that stimulate antigen-specific CD4+ and CD8+ T cells against infectious pathogens. cDC1s and cDC2s work as professional APCs in the lung. Sub-population of cDCs has been investigated by deep separation using single RNA sequence and CyTOF technology and have shown to process and present antigen. In addition, there has been increasing evidence for antigen presentation by resident APCs such as epithelial cells, epithelial cells in the lungs. The relation of pathogen and inflammation model to APCs was shown in Table 1. Although MACs express MHC and costimulatory molecules with higher engulfment capacity, the role of MACs in the lung as APCs is still unclear.
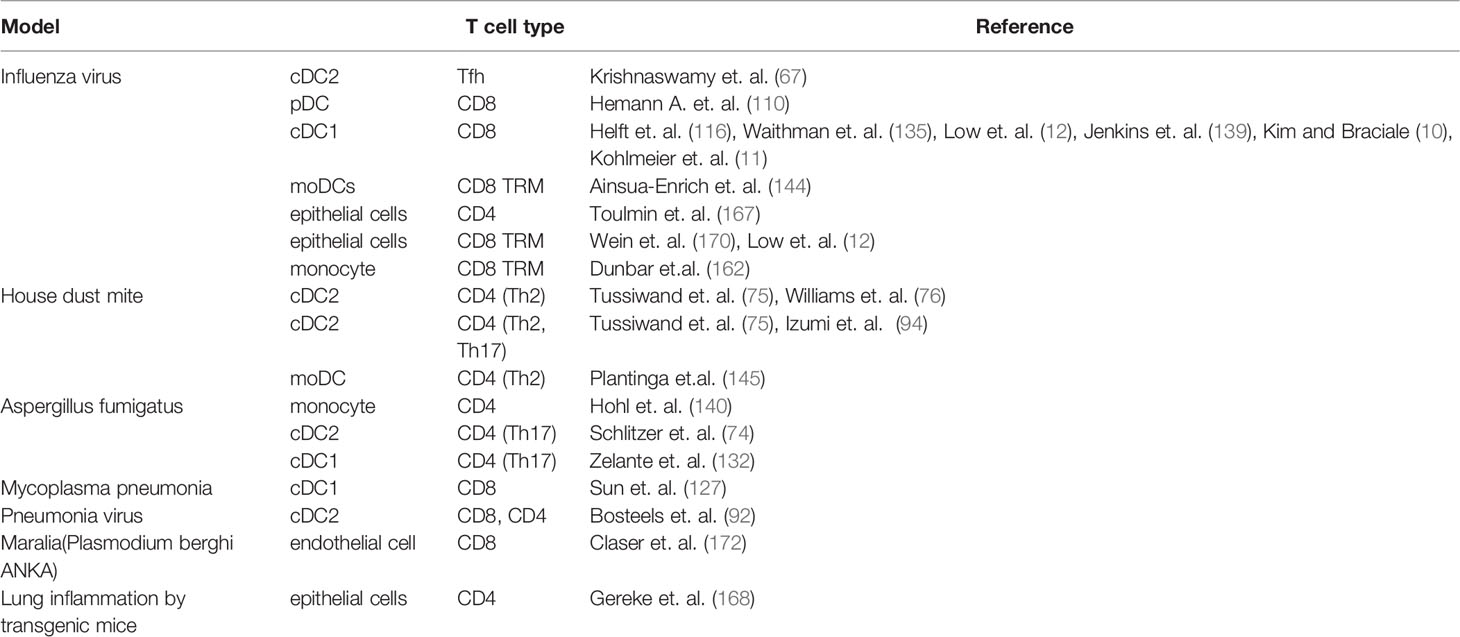
Table 1 Lung APCs and their roles in T cell responses.
To initially prime antigen-specific T cells, antigen-captured DCs and migratory APCs need to traffic to lung-draining LNs where they encounter naïve T cells to select antigen-specific T cells. Following a program of proliferation and differentiation of T cells in LNs, antigen-specific effector or memory T cells migrate back to the infected lung to mediate their effector function (10, 11). At the same time, antigen-specific effector or memory T cells are reactivated by APCs, including monocytes, epithelial cells and endothelial cells in the lungs, with support of cytokine production and the local microenvironment (12). Among the antigen specific memory type cells, CD4+ and CD8+ TRM cells in the lung provide protection against pathogen infection and retain for long time period in the peripheral tissue. Pulmonary antigen encounter is necessary for the establishment of TRM during IAV infection in the lung (173), and antigen presentation by DCs with cytokines such as TGF-β and IL15 is shown to be important for TRM development in the lung (174–176). Various types of APCs in the lungs contribute to pathogen clearance against viruses, fungi, and bacteria; therefore, APCs perform their function depending on the pathogen infection, and further studies are needed to clarify the role of individual APCs in the lungs.
Author Contributions
TakK and MI wrote the manuscript. TarK edited and supervised the manuscript. All authors contributed to the article and approved the submitted version.
Conflict of Interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Publisher’s Note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
Acknowledgments
We thank Chihiro Suzuki for secretarial assistance. This work was supported by JSPS KAKENHI Grant-in-Aid for Scientific Research B (20H03468) and C (19K07608), Grant-in-Aid for Early-Career Scientists (21K14817) and the Takeda Science Foundation. We would like to thank Editage (www.editage.com) for English language editing.
References
1. Kawai T, Akira S. The Role of Pattern-Recognition Receptors in Innate Immunity: Update on Toll-Like Receptors. Nat Immunol (2010) 11:373–84. doi: 10.1038/ni.1863
2. Kawasaki T, Kawai T. Discrimination Between Self and Non-Self-Nucleic Acids by the Innate Immune System. Int Rev Cell Mol Biol (2019) 344:1–30. doi: 10.1016/bs.ircmb.2018.08.004
3. Joffre OP, Segura E, Savina A, Amigorena S. Cross-Presentation by Dendritic Cells. Nat Rev Immunol (2012) 12:557–69. doi: 10.1038/nri3254
4. Holt PG. Antigen Presentation in the Lung. Am J Respir Crit Care Med (2000) 162:S151–156. doi: 10.1164/ajrccm.162.supplement_3.15tac2
5. Crystal RG, Randell SH, Engelhardt JF, Voynow J, Sunday ME. Airway Epithelial Cells: Current Concepts and Challenges. Proc Am Thorac Soc (2008) 5:772–7. doi: 10.1513/pats.200805-041HR
6. Zaragosi LE, Deprez M, Barbry P. Using Single-Cell RNA Sequencing to Unravel Cell Lineage Relationships in the Respiratory Tract. Biochem Soc Trans (2020) 48:327–36. doi: 10.1042/BST20191010
7. Shenoy AT, Lyon De Ana C, Arafa EI, Salwig I, Barker KA, Korkmaz FT, et al. Antigen Presentation by Lung Epithelial Cells Directs CD4(+) TRM Cell Function and Regulates Barrier Immunity. Nat Commun (2021) 12:5834. doi: 10.1038/s41467-021-26045-w
8. Banchereau J, Steinman RM. Dendritic Cells and the Control of Immunity. Nature (1998) 392:245–52. doi: 10.1038/32588
9. Henri S, Vremec D, Kamath A, Waithman J, Williams S, Benoist C, et al. The Dendritic Cell Populations of Mouse Lymph Nodes. J Immunol (2001) 167:741–8. doi: 10.4049/jimmunol.167.2.741
10. Kim TS, Braciale TJ. Respiratory Dendritic Cell Subsets Differ in Their Capacity to Support the Induction of Virus-Specific Cytotoxic CD8+ T Cell Responses. PloS One (2009) 4:e4204. doi: 10.1371/journal.pone.0004204
11. Kohlmeier JE, Cookenham T, Miller SC, Roberts AD, Christensen JP, Thomsen AR, et al. CXCR3 Directs Antigen-Specific Effector CD4+ T Cell Migration to the Lung During Parainfluenza Virus Infection. J Immunol (2009) 183:4378–84. doi: 10.4049/jimmunol.0902022
12. Low JS, Farsakoglu Y, Amezcua Vesely MC, Sefik E, Kelly JB, Harman CCD, et al. Tissue-Resident Memory T Cell Reactivation by Diverse Antigen-Presenting Cells Imparts Distinct Functional Responses. J Exp Med (2020) 217:e20192291. doi: 10.1084/jem.20192291
13. Mueller SN, Gebhardt T, Carbone FR, Heath WR. Memory T Cell Subsets, Migration Patterns, and Tissue Residence. Annu Rev Immunol (2013) 31:137–61. doi: 10.1146/annurev-immunol-032712-095954
14. Schenkel JM, Masopust D. Tissue-Resident Memory T Cells. Immunity (2014) 41:886–97. doi: 10.1016/j.immuni.2014.12.007
15. Manoury B, Mazzeo D, Li DN, Billson J, Loak K, Benaroch P, et al. Asparagine Endopeptidase can Initiate the Removal of the MHC Class II Invariant Chain Chaperone. Immunity (2003) 18:489–98. doi: 10.1016/S1074-7613(03)00085-2
16. Lautwein A, Burster T, Lennon-Dumenil AM, Overkleeft HS, Weber E, Kalbacher H, et al. Inflammatory Stimuli Recruit Cathepsin Activity to Late Endosomal Compartments in Human Dendritic Cells. Eur J Immunol (2002) 32:3348–57. doi: 10.1002/1521-4141(200212)32:12<3348::AID-IMMU3348>3.0.CO;2-S
17. Nakagawa TY, Brissette WH, Lira PD, Griffiths RJ, Petrushova N, Stock J, et al. Impaired Invariant Chain Degradation and Antigen Presentation and Diminished Collagen-Induced Arthritis in Cathepsin S Null Mice. Immunity (1999) 10:207–17. doi: 10.1016/S1074-7613(00)80021-7
18. Shi GP, Villadangos JA, Dranoff G, Small C, Gu L, Haley KJ, et al. Cathepsin S Required for Normal MHC Class II Peptide Loading and Germinal Center Development. Immunity (1999) 10:197–206. doi: 10.1016/S1074-7613(00)80020-5
19. Morrison LA, Lukacher AE, Braciale VL, Fan DP, Braciale TJ. Differences in Antigen Presentation to MHC Class I-And Class II-Restricted Influenza Virus-Specific Cytolytic T Lymphocyte Clones. J Exp Med (1986) 163:903–21. doi: 10.1084/jem.163.4.903
20. Hunt DF, Michel H, Dickinson TA, Shabanowitz J, Cox AL, Sakaguchi K, et al. Peptides Presented to the Immune System by the Murine Class II Major Histocompatibility Complex Molecule I-Ad. Science (1992) 256:1817–20. doi: 10.1126/science.1319610
21. Rammensee H, Bachmann J, Emmerich NP, Bachor OA, Stevanovic S. SYFPEITHI: Database for MHC Ligands and Peptide Motifs. Immunogenetics (1999) 50:213–9. doi: 10.1007/s002510050595
22. Crotzer VL, Blum JS. Autophagy and Adaptive Immunity. Immunology (2010) 131:9–17. doi: 10.1111/j.1365-2567.2010.03321.x
23. Simonsen A, Tooze SA. Coordination of Membrane Events During Autophagy by Multiple Class III PI3-Kinase Complexes. J Cell Biol (2009) 186:773–82. doi: 10.1083/jcb.200907014
24. Zhou D, Li P, Lin Y, Lott JM, Hislop AD, Canaday DH, et al. Lamp-2a Facilitates MHC Class II Presentation of Cytoplasmic Antigens. Immunity (2005) 22:571–81. doi: 10.1016/j.immuni.2005.03.009
25. Cresswell P. Invariant Chain Structure and MHC Class II Function. Cell (1996) 84:505–7. doi: 10.1016/S0092-8674(00)81025-9
26. Dugast M, Toussaint H, Dousset C, Benaroch P. AP2 Clathrin Adaptor Complex, But Not AP1, Controls the Access of the Major Histocompatibility Complex (MHC) Class II to Endosomes. J Biol Chem (2005) 280:19656–64. doi: 10.1074/jbc.M501357200
27. McCormick PJ, Martina JA, Bonifacino JS. Involvement of Clathrin and AP-2 in the Trafficking of MHC Class II Molecules to Antigen-Processing Compartments. Proc Natl Acad Sci USA (2005) 102:7910–5. doi: 10.1073/pnas.0502206102
28. Roche PA, Cresswell P. Invariant Chain Association With HLA-DR Molecules Inhibits Immunogenic Peptide Binding. Nature (1990) 345:615–8. doi: 10.1038/345615a0
29. Teyton L, O'Sullivan D, Dickson PW, Lotteau V, Sette A, Fink P, et al. Invariant Chain Distinguishes Between the Exogenous and Endogenous Antigen Presentation Pathways. Nature (1990) 348:39–44. doi: 10.1038/348039a0
30. Cresswell P. Assembly, Transport, and Function of MHC Class II Molecules. Annu Rev Immunol (1994) 12:259–93. doi: 10.1146/annurev.iy.12.040194.001355
31. Kim HJ, Bandola-Simon J, Ishido S, Wong NW, Koparde VN, Cam M, et al. Ubiquitination of MHC Class II by March-I Regulates Dendritic Cell Fitness. J Immunol (2021) 206:494–504. doi: 10.4049/jimmunol.2000975
32. Wilson KR, Jenika D, Blum AB, Macri C, Xu B, Liu H, et al. MHC Class II Ubiquitination Regulates Dendritic Cell Function and Immunity. J Immunol (2021) 207:2255–64. doi: 10.4049/jimmunol.2001426
33. Zehner M, Marschall AL, Bos E, Schloetel JG, Kreer C, Fehrenschild D, et al. The Translocon Protein Sec61 Mediates Antigen Transport From Endosomes in the Cytosol for Cross-Presentation to CD8(+) T Cells. Immunity (2015) 42:850–63. doi: 10.1016/j.immuni.2015.04.008
34. Shen L, Sigal LJ, Boes M, Rock KL. Important Role of Cathepsin S in Generating Peptides for TAP-Independent MHC Class I Crosspresentation In Vivo. Immunity (2004) 21:155–65. doi: 10.1016/j.immuni.2004.07.004
35. Nakagawa T, Roth W, Wong P, Nelson A, Farr A, Deussing J, et al. Cathepsin L: Critical Role in Ii Degradation and CD4 T Cell Selection in the Thymus. Science (1998) 280:450–3. doi: 10.1126/science.280.5362.450
36. Shi GP, Bryant RA, Riese R, Verhelst S, Driessen C, Li Z, et al. Role for Cathepsin F in Invariant Chain Processing and Major Histocompatibility Complex Class II Peptide Loading by Macrophages. J Exp Med (2000) 191:1177–86. doi: 10.1084/jem.191.7.1177
37. Kovacsovics-Bankowski M, Rock KL. A Phagosome-to-Cytosol Pathway for Exogenous Antigens Presented on MHC Class I Molecules. Science (1995) 267:243–6. doi: 10.1126/science.7809629
38. Palmowski MJ, Gileadi U, Salio M, Gallimore A, Millrain M, James E, et al. Role of Immunoproteasomes in Cross-Presentation. J Immunol (2006) 177:983–90. doi: 10.4049/jimmunol.177.2.983
39. Lafourcade C, Sobo K, Kieffer-Jaquinod S, Garin J, van der Goot FG. Regulation of the V-ATPase Along the Endocytic Pathway Occurs Through Reversible Subunit Association and Membrane Localization. PloS One (2008) 3:e2758. doi: 10.1371/journal.pone.0002758
40. Vulcano M, Dusi S, Lissandrini D, Badolato R, Mazzi P, Riboldi E, et al. Toll Receptor-Mediated Regulation of NADPH Oxidase in Human Dendritic Cells. J Immunol (2004) 173:5749–56. doi: 10.4049/jimmunol.173.9.5749
41. Mantegazza AR, Savina A, Vermeulen M, Perez L, Geffner J, Hermine O, et al. NADPH Oxidase Controls Phagosomal pH and Antigen Cross-Presentation in Human Dendritic Cells. Blood (2008) 112:4712–22. doi: 10.1182/blood-2008-01-134791
42. Savina A, Jancic C, Hugues S, Guermonprez P, Vargas P, Moura IC, et al. NOX2 Controls Phagosomal pH to Regulate Antigen Processing During Crosspresentation by Dendritic Cells. Cell (2006) 126:205–18. doi: 10.1016/j.cell.2006.05.035
43. Jancic C, Savina A, Wasmeier C, Tolmachova T, El-Benna J, Dang PM, et al. Rab27a Regulates Phagosomal pH and NADPH Oxidase Recruitment to Dendritic Cell Phagosomes. Nat Cell Biol (2007) 9:367–78. doi: 10.1038/ncb1552
44. Nair-Gupta P, Baccarini A, Tung N, Seyffer F, Florey O, Huang Y, et al. TLR Signals Induce Phagosomal MHC-I Delivery From the Endosomal Recycling Compartment to Allow Cross-Presentation. Cell (2014) 158:506–21. doi: 10.1016/j.cell.2014.04.054
45. Nair-Gupta P, Blander JM. An Updated View of the Intracellular Mechanisms Regulating Cross-Presentation. Front Immunol (2013) 4:401. doi: 10.3389/fimmu.2013.00401
46. Blees A, Januliene D, Hofmann T, Koller N, Schmidt C, Trowitzsch S, et al. Structure of the Human MHC-I Peptide-Loading Complex. Nature (2017) 551:525–8. doi: 10.1038/nature24627
47. Cebrian I, Visentin G, Blanchard N, Jouve M, Bobard A, Moita C, et al. Sec22b Regulates Phagosomal Maturation and Antigen Crosspresentation by Dendritic Cells. Cell (2011) 147:1355–68. doi: 10.1016/j.cell.2011.11.021
48. Saveanu L, Carroll O, Weimershaus M, Guermonprez P, Firat E, Lindo V, et al. IRAP Identifies an Endosomal Compartment Required for MHC Class I Cross-Presentation. Science (2009) 325:213–7. doi: 10.1126/science.1172845
49. Weimershaus M, Maschalidi S, Sepulveda F, Manoury B, van Endert P, Saveanu L. Conventional Dendritic Cells Require IRAP-Rab14 Endosomes for Efficient Cross-Presentation. J Immunol (2012) 188:1840–6. doi: 10.4049/jimmunol.1101504
50. Dick TP, Bangia N, Peaper DR, Cresswell P. Disulfide Bond Isomerization and the Assembly of MHC Class I-Peptide Complexes. Immunity (2002) 16:87–98. doi: 10.1016/S1074-7613(02)00263-7
51. Kanaseki T, Lind KC, Escobar H, Nagarajan N, Reyes-Vargas E, Rudd B, et al. ERAAP and Tapasin Independently Edit the Amino and Carboxyl Termini of MHC Class I Peptides. J Immunol (2013) 191:1547–55. doi: 10.4049/jimmunol.1301043
52. Ortmann B, Copeman J, Lehner PJ, Sadasivan B, Herberg JA, Grandea AG, et al. A Critical Role for Tapasin in the Assembly and Function of Multimeric MHC Class I-TAP Complexes. Science (1997) 277:1306–9. doi: 10.1126/science.277.5330.1306
53. Barbet G, Nair-Gupta P, Schotsaert M, Yeung ST, Moretti J, Seyffer F, et al. TAP Dysfunction in Dendritic Cells Enables Noncanonical Cross-Presentation for T Cell Priming. Nat Immunol (2021) 22:497–509. doi: 10.1038/s41590-021-00903-7
54. Bocchino M, Zanotta S, Capitelli L, Galati D. Dendritic Cells Are the Intriguing Players in the Puzzle of Idiopathic Pulmonary Fibrosis Pathogenesis. Front Immunol (2021) 12:664109. doi: 10.3389/fimmu.2021.664109
55. Guilliams M, Lambrecht BN, Hammad H. Division of Labor Between Lung Dendritic Cells and Macrophages in the Defense Against Pulmonary Infections. Mucosal Immunol (2013) 6:464–73. doi: 10.1038/mi.2013.14
56. Cabeza-Cabrerizo M, Cardoso A, Minutti CM, Pereira da Costa M, Reis e Sousa C. Dendritic Cells Revisited. Annu Rev Immunol (2021) 39:131–66. doi: 10.1146/annurev-immunol-061020-053707
57. Condon TV, Sawyer RT, Fenton MJ, Riches DW. Lung Dendritic Cells at the Innate-Adaptive Immune Interface. J Leukoc Biol (2011) 90:883–95. doi: 10.1189/jlb.0311134
58. Murphy TL, Grajales-Reyes GE, Wu X, Tussiwand R, Briseno CG, Iwata A, et al. Transcriptional Control of Dendritic Cell Development. Annu Rev Immunol (2016) 34:93–119. doi: 10.1146/annurev-immunol-032713-120204
59. Guilliams M, Dutertre CA, Scott CL, McGovern N, Sichien D, Chakarov S, et al. Unsupervised High-Dimensional Analysis Aligns Dendritic Cells Across Tissues and Species. Immunity (2016) 45:669–84. doi: 10.1016/j.immuni.2016.08.015
60. Guilliams M, Ginhoux F, Jakubzick C, Naik SH, Onai N, Schraml BU, et al. Dendritic Cells, Monocytes and Macrophages: A Unified Nomenclature Based on Ontogeny. Nat Rev Immunol (2014) 14:571–8. doi: 10.1038/nri3712
61. Bagadia P, Huang X, Liu TT, Durai V, Grajales-Reyes GE, Nitschke M, et al. An Nfil3-Zeb2-Id2 Pathway Imposes Irf8 Enhancer Switching During cDC1 Development. Nat Immunol (2019) 20:1174–85. doi: 10.1038/s41590-019-0449-3
62. Bajana S, Turner S, Paul J, Ainsua-Enrich E, Kovats S. IRF4 and IRF8 Act in CD11c+ Cells To Regulate Terminal Differentiation of Lung Tissue Dendritic Cells. J Immunol (2016) 196:1666–77. doi: 10.4049/jimmunol.1501870
63. Durai V, Bagadia P, Granja JM, Satpathy AT, Kulkarni DH, Davidson JT, et al. Cryptic Activation of an Irf8 Enhancer Governs cDC1 Fate Specification. Nat Immunol (2019) 20:1161–73. doi: 10.1038/s41590-019-0450-x
64. Edelson BT, Kc W, Juang R, Kohyama M, Benoit LA, Klekotka PA, et al. Peripheral CD103+ Dendritic Cells Form a Unified Subset Developmentally Related to CD8alpha+ Conventional Dendritic Cells. J Exp Med (2010) 207:823–36. doi: 10.1084/jem.20091627
65. Everts B, Tussiwand R, Dreesen L, Fairfax KC, Huang SC, Smith AM, et al. Migratory CD103+ Dendritic Cells Suppress Helminth-Driven Type 2 Immunity Through Constitutive Expression of IL-12. J Exp Med (2016) 213:35–51. doi: 10.1084/jem.20150235
66. Grajales-Reyes GE, Iwata A, Albring J, Wu X, Tussiwand R, Kc W, et al. Batf3 Maintains Autoactivation of Irf8 for Commitment of a CD8alpha(+) Conventional DC Clonogenic Progenitor. Nat Immunol (2015) 16:708–17. doi: 10.1038/ni.3197
67. Krishnaswamy JK, Gowthaman U, Zhang B, Mattsson J, Szeponik L, Liu D, et al. Migratory CD11b(+) Conventional Dendritic Cells Induce T Follicular Helper Cell-Dependent Antibody Responses. Sci Immunol (2017) 2:eaam9169. doi: 10.1126/sciimmunol.aam9169
68. Sichien D, Scott CL, Martens L, Vanderkerken M, Van Gassen S, Plantinga M, et al. IRF8 Transcription Factor Controls Survival and Function of Terminally Differentiated Conventional and Plasmacytoid Dendritic Cells, Respectively. Immunity (2016) 45:626–40. doi: 10.1016/j.immuni.2016.08.013
69. Vander Lugt B, Khan AA, Hackney JA, Agrawal S, Lesch J, Zhou M, et al. Transcriptional Programming of Dendritic Cells for Enhanced MHC Class II Antigen Presentation. Nat Immunol (2014) 15:161–7. doi: 10.1038/ni.2795
70. Bajana S, Roach K, Turner S, Paul J, Kovats S. IRF4 Promotes Cutaneous Dendritic Cell Migration to Lymph Nodes During Homeostasis and Inflammation. J Immunol (2012) 189:3368–77. doi: 10.4049/jimmunol.1102613
71. Gao Y, Nish SA, Jiang R, Hou L, Licona-Limon P, Weinstein JS, et al. Control of T Helper 2 Responses by Transcription Factor IRF4-Dependent Dendritic Cells. Immunity (2013) 39:722–32. doi: 10.1016/j.immuni.2013.08.028
72. Kumamoto Y, Linehan M, Weinstein JS, Laidlaw BJ, Craft JE, Iwasaki A. CD301b(+) Dermal Dendritic Cells Drive T Helper 2 Cell-Mediated Immunity. Immunity (2013) 39:733–43. doi: 10.1016/j.immuni.2013.08.029
73. Satpathy AT, Briseno CG, Lee JS, Ng D, Manieri NA, Kc W, et al. Notch2-Dependent Classical Dendritic Cells Orchestrate Intestinal Immunity to Attaching-and-Effacing Bacterial Pathogens. Nat Immunol (2013) 14:937–48. doi: 10.1038/ni.2679
74. Schlitzer A, McGovern N, Teo P, Zelante T, Atarashi K, Low D, et al. IRF4 Transcription Factor-Dependent CD11b+ Dendritic Cells in Human and Mouse Control Mucosal IL-17 Cytokine Responses. Immunity (2013) 38:970–83. doi: 10.1016/j.immuni.2013.04.011
75. Tussiwand R, Everts B, Grajales-Reyes GE, Kretzer NM, Iwata A, Bagaitkar J, et al. Klf4 Expression in Conventional Dendritic Cells is Required for T Helper 2 Cell Responses. Immunity (2015) 42:916–28. doi: 10.1016/j.immuni.2015.04.017
76. Williams JW, Tjota MY, Clay BS, Vander Lugt B, Bandukwala HS, Hrusch CL, et al. Transcription Factor IRF4 Drives Dendritic Cells to Promote Th2 Differentiation. Nat Commun (2013) 4:2990. doi: 10.1038/ncomms3990
77. Cisse B, Caton ML, Lehner M, Maeda T, Scheu S, Locksley R, et al. Transcription Factor E2-2 is an Essential and Specific Regulator of Plasmacytoid Dendritic Cell Development. Cell (2008) 135:37–48. doi: 10.1016/j.cell.2008.09.016
78. Ghosh HS, Cisse B, Bunin A, Lewis KL, Reizis B. Continuous Expression of the Transcription Factor E2-2 Maintains the Cell Fate of Mature Plasmacytoid Dendritic Cells. Immunity (2010) 33:905–16. doi: 10.1016/j.immuni.2010.11.023
79. Nagasawa M, Schmidlin H, Hazekamp MG, Schotte R, Blom B. Development of Human Plasmacytoid Dendritic Cells Depends on the Combined Action of the Basic Helix-Loop-Helix Factor E2-2 and the Ets Factor Spi-B. Eur J Immunol (2008) 38:2389–400. doi: 10.1002/eji.200838470
80. Lyons-Cohen MR, Thomas SY, Cook DN, Nakano H. Precision-Cut Mouse Lung Slices to Visualize Live Pulmonary Dendritic Cells. J Vis Exp (2017) 122:55465. doi: 10.3791/55465
81. Sung SS, Fu SM, Rose CE Jr., Gaskin F, Ju ST, Beaty SR. A Major Lung CD103 (Alphae)-Beta7 Integrin-Positive Epithelial Dendritic Cell Population Expressing Langerin and Tight Junction Proteins. J Immunol (2006) 176:2161–72. doi: 10.4049/jimmunol.176.4.2161
82. Masten BJ, Lipscomb MF. Comparison of Lung Dendritic Cells and B Cells in Stimulating Naive Antigen-Specific T Cells. J Immunol (1999) 162:1310–7.
83. Masten BJ, Olson GK, Kusewitt DF, Lipscomb MF. Flt3 Ligand Preferentially Increases the Number of Functionally Active Myeloid Dendritic Cells in the Lungs of Mice. J Immunol (2004) 172:4077–83. doi: 10.4049/jimmunol.172.7.4077
84. von Garnier C, Filgueira L, Wikstrom M, Smith M, Thomas JA, Strickland DH, et al. Anatomical Location Determines the Distribution and Function of Dendritic Cells and Other APCs in the Respiratory Tract. J Immunol (2005) 175:1609–18. doi: 10.4049/jimmunol.175.3.1609
85. Aldridge JR Jr, Moseley CE, Boltz DA, Negovetich NJ, Reynolds C, Franks J, et al. TNF/iNOS-Producing Dendritic Cells are the Necessary Evil of Lethal Influenza Virus Infection. Proc Natl Acad Sci USA (2009) 106:5306–11. doi: 10.1073/pnas.0900655106
86. Belz GT, Smith CM, Kleinert L, Reading P, Brooks A, Shortman K, et al. Distinct Migrating and Nonmigrating Dendritic Cell Populations are Involved in MHC Class I-Restricted Antigen Presentation After Lung Infection With Virus. Proc Natl Acad Sci USA (2004) 101:8670–5. doi: 10.1073/pnas.0402644101
87. Legge KL, Braciale TJ. Accelerated Migration of Respiratory Dendritic Cells to the Regional Lymph Nodes is Limited to the Early Phase of Pulmonary Infection. Immunity (2003) 18:265–77. doi: 10.1016/S1074-7613(03)00023-2
88. Cheong C, Matos I, Choi JH, Dandamudi DB, Shrestha E, Longhi MP, et al. Microbial Stimulation Fully Differentiates Monocytes to DC-SIGN/CD209(+) Dendritic Cells for Immune T Cell Areas. Cell (2010) 143:416–29. doi: 10.1016/j.cell.2010.09.039
89. Kool M, Soullie T, van Nimwegen M, Willart MA, Muskens F, Jung S, et al. Alum Adjuvant Boosts Adaptive Immunity by Inducing Uric Acid and Activating Inflammatory Dendritic Cells. J Exp Med (2008) 205:869–82. doi: 10.1084/jem.20071087
90. Leon B, Lopez-Bravo M, Ardavin C. Monocyte-Derived Dendritic Cells Formed at the Infection Site Control the Induction of Protective T Helper 1 Responses Against Leishmania. Immunity (2007) 26:519–31. doi: 10.1016/j.immuni.2007.01.017
91. Wu X, Briseno CG, Durai V, Albring JC, Haldar M, Bagadia P, et al. Mafb Lineage Tracing to Distinguish Macrophages From Other Immune Lineages Reveals Dual Identity of Langerhans Cells. J Exp Med (2016) 213:2553–65. doi: 10.1084/jem.20160600
92. Bosteels C, Neyt K, Vanheerswynghels M, van Helden MJ, Sichien D, Debeuf N, et al. Inflammatory Type 2 cDCs Acquire Features of cDC1s and Macrophages to Orchestrate Immunity to Respiratory Virus Infection. Immunity (2020) 52:1039–56:e1039. doi: 10.1016/j.immuni.2020.04.005
93. Han X, Wang R, Zhou Y, Fei L, Sun H, Lai S, et al. Mapping the Mouse Cell Atlas by Microwell-Seq. Cell (2018) 172:1091–107. e1017. doi: 10.1016/j.cell.2018.02.001
94. Izumi G, Nakano H, Nakano K, Whitehead GS, Grimm SA, Fessler MB, et al. CD11b(+) Lung Dendritic Cells at Different Stages of Maturation Induce Th17 or Th2 Differentiation. Nat Commun (2021) 12:5029. doi: 10.1038/s41467-021-25307-x
95. Hussell T, Bell TJ. Alveolar Macrophages: Plasticity in a Tissue-Specific Context. Nat Rev Immunol (2014) 14:81–93. doi: 10.1038/nri3600
96. Kawasaki T, Ito K, Miyata H, Akira S, Kawai T. Deletion of PIKfyve Alters Alveolar Macrophage Populations and Exacerbates Allergic Inflammation in Mice. EMBO J (2017) 36:1707–18. doi: 10.15252/embj.201695528
97. Guilliams M, De Kleer I, Henri S, Post S, Vanhoutte L, De Prijck S, et al. Alveolar Macrophages Develop From Fetal Monocytes That Differentiate Into Long-Lived Cells in the First Week of Life via GM-CSF. J Exp Med (2013) 210:1977–92. doi: 10.1084/jem.20131199
98. Shibata Y, Berclaz PY, Chroneos ZC, Yoshida M, Whitsett JA, Trapnell BC. GM-CSF Regulates Alveolar Macrophage Differentiation and Innate Immunity in the Lung Through PU.1. Immunity (2001) 15:557–67. doi: 10.1016/S1074-7613(01)00218-7
99. Suzuki T, Sakagami T, Rubin BK, Nogee LM, Wood RE, Zimmerman SL, et al. Familial Pulmonary Alveolar Proteinosis Caused by Mutations in CSF2RA. J Exp Med (2008) 205:2703–10. doi: 10.1084/jem.20080990
100. Schneider C, Nobs SP, Kurrer M, Rehrauer H, Thiele C, Kopf M. Induction of the Nuclear Receptor PPAR-Gamma by the Cytokine GM-CSF is Critical for the Differentiation of Fetal Monocytes Into Alveolar Macrophages. Nat Immunol (2014) 15:1026–37. doi: 10.1038/ni.3005
101. Bedoret D, Wallemacq H, Marichal T, Desmet C, Quesada Calvo F, Henry E, et al. Lung Interstitial Macrophages Alter Dendritic Cell Functions to Prevent Airway Allergy in Mice. J Clin Invest (2009) 119:3723–38. doi: 10.1172/JCI39717
102. Franke-Ullmann G, Pfortner C, Walter P, Steinmuller C, Lohmann-Matthes ML, Kobzik L. Characterization of Murine Lung Interstitial Macrophages in Comparison With Alveolar Macrophages In Vitro. J Immunol (1996) 157:3097–104.
103. Gibbings SL, Thomas SM, Atif SM, McCubbrey AL, Desch AN, Danhorn T, et al. Three Unique Interstitial Macrophages in the Murine Lung at Steady State. Am J Respir Cell Mol Biol (2017) 57:66–76. doi: 10.1165/rcmb.2016-0361OC
104. Kawano H, Kayama H, Nakama T, Hashimoto T, Umemoto E, Takeda K. IL-10-Producing Lung Interstitial Macrophages Prevent Neutrophilic Asthma. Int Immunol (2016) 28:489–501. doi: 10.1093/intimm/dxw012
105. Ginhoux F, Guilliams M. Tissue-Resident Macrophage Ontogeny and Homeostasis. Immunity (2016) 44:439–49. doi: 10.1016/j.immuni.2016.02.024
106. Misharin AV, Morales-Nebreda L, Mutlu GM, Budinger GR, Perlman H. Flow Cytometric Analysis of Macrophages and Dendritic Cell Subsets in the Mouse Lung. Am J Respir Cell Mol Biol (2013) 49:503–10. doi: 10.1165/rcmb.2013-0086MA
107. Guo Y, Kasahara S, Jhingran A, Tosini NL, Zhai B, Aufiero MA, et al. During Aspergillus Infection, Monocyte-Derived DCs, Neutrophils, and Plasmacytoid DCs Enhance Innate Immune Defense Through CXCR3-Dependent Crosstalk. Cell Host Microbe (2020) 28:104–16. e104. doi: 10.1016/j.chom.2020.05.002
108. Asselin-Paturel C, Boonstra A, Dalod M, Durand I, Yessaad N, Dezutter-Dambuyant C, et al. Mouse Type I IFN-Producing Cells are Immature APCs With Plasmacytoid Morphology. Nat Immunol (2001) 2:1144–50. doi: 10.1038/ni736
109. Nakano H, Yanagita M, Gunn MD. CD11c(+)B220(+)Gr-1(+) Cells in Mouse Lymph Nodes and Spleen Display Characteristics of Plasmacytoid Dendritic Cells. J Exp Med (2001) 194:1171–8. doi: 10.1084/jem.194.8.1171
110. Hemann EA, Sjaastad LE, Langlois RA, Legge KL. Plasmacytoid Dendritic Cells Require Direct Infection To Sustain the Pulmonary Influenza A Virus-Specific CD8 T Cell Response. J Virol (2015) 90:2830–7. doi: 10.1128/JVI.02546-15
111. Wolf AI, Buehler D, Hensley SE, Cavanagh LL, Wherry EJ, Kastner P, et al. Plasmacytoid Dendritic Cells are Dispensable During Primary Influenza Virus Infection. J Immunol (2009) 182:871–9. doi: 10.4049/jimmunol.182.2.871
112. Brewitz A, Eickhoff S, Dahling S, Quast T, Bedoui S, Kroczek RA, et al. CD8(+) T Cells Orchestrate pDC-XCR1(+) Dendritic Cell Spatial and Functional Cooperativity to Optimize Priming. Immunity (2017) 46:205–19. doi: 10.1016/j.immuni.2017.01.003
113. Fu C, Peng P, Loschko J, Feng L, Pham P, Cui W, et al. Plasmacytoid Dendritic Cells Cross-Prime Naive CD8 T Cells by Transferring Antigen to Conventional Dendritic Cells Through Exosomes. Proc Natl Acad Sci USA (2020) 117:23730–41. doi: 10.1073/pnas.2002345117
114. Osterholzer JJ, Ames T, Polak T, Sonstein J, Moore BB, Chensue SW, et al. CCR2 and CCR6, But Not Endothelial Selectins, Mediate the Accumulation of Immature Dendritic Cells Within the Lungs of Mice in Response to Particulate Antigen. J Immunol (2005) 175:874–83. doi: 10.4049/jimmunol.175.2.874
115. van Rijt LS, Prins JB, Leenen PJ, Thielemans K, de Vries VC, Hoogsteden HC, et al. Allergen-Induced Accumulation of Airway Dendritic Cells is Supported by an Increase in CD31(hi)Ly-6C(Neg) Bone Marrow Precursors in a Mouse Model of Asthma. Blood (2002) 100:3663–71. doi: 10.1182/blood-2002-03-0673
116. Helft J, Manicassamy B, Guermonprez P, Hashimoto D, Silvin A, Agudo J, et al. Cross-Presenting CD103+ Dendritic Cells are Protected From Influenza Virus Infection. J Clin Invest (2012) 122:4037–47. doi: 10.1172/JCI60659
117. Nakano H, Burgents JE, Nakano K, Whitehead GS, Cheong C, Bortner CD, et al. Migratory Properties of Pulmonary Dendritic Cells are Determined by Their Developmental Lineage. Mucosal Immunol (2013) 6:678–91. doi: 10.1038/mi.2012.106
118. Chow KV, Lew AM, Sutherland RM, Zhan Y. Monocyte-Derived Dendritic Cells Promote Th Polarization, Whereas Conventional Dendritic Cells Promote Th Proliferation. J Immunol (2016) 196:624–36. doi: 10.4049/jimmunol.1501202
119. Mansouri S, Patel S, Katikaneni DS, Blaauboer SM, Wang W, Schattgen S, et al. Immature Lung TNFR2(-) Conventional DC 2 Subpopulation Activates moDCs to Promote Cyclic Di-GMP Mucosal Adjuvant Responses In Vivo. Mucosal Immunol (2019) 12:277–89. doi: 10.1038/s41385-018-0098-0
120. Hao X, Kim TS, Braciale TJ. Differential Response of Respiratory Dendritic Cell Subsets to Influenza Virus Infection. J Virol (2008) 82:4908–19. doi: 10.1128/JVI.02367-07
121. Infusini G, Smith JM, Yuan H, Pizzolla A, Ng WC, Londrigan SL, et al. Respiratory DC Use IFITM3 to Avoid Direct Viral Infection and Safeguard Virus-Specific CD8+ T Cell Priming. PloS One (2015) 10:e0143539. doi: 10.1371/journal.pone.0143539
122. Kim TS, Gorski SA, Hahn S, Murphy KM, Braciale TJ. Distinct Dendritic Cell Subsets Dictate the Fate Decision Between Effector and Memory CD8(+) T Cell Differentiation by a CD24-Dependent Mechanism. Immunity (2014) 40:400–13. doi: 10.1016/j.immuni.2014.02.004
123. den Haan JM, Lehar SM, Bevan MJ. CD8(+) But Not CD8(-) Dendritic Cells Cross-Prime Cytotoxic T Cells In Vivo. J Exp Med (2000) 192:1685–96. doi: 10.1084/jem.192.12.1685
124. Hildner K, Edelson BT, Purtha WE, Diamond M, Matsushita H, Kohyama M, et al. Batf3 Deficiency Reveals a Critical Role for CD8alpha+ Dendritic Cells in Cytotoxic T Cell Immunity. Science (2008) 322:1097–100. doi: 10.1126/science.1164206
125. Pooley JL, Heath WR, Shortman K. Cutting Edge: Intravenous Soluble Antigen is Presented to CD4 T Cells by CD8- Dendritic Cells, But Cross-Presented to CD8 T Cells by CD8+ Dendritic Cells. J Immunol (2001) 166:5327–30. doi: 10.4049/jimmunol.166.9.5327
126. Desch AN, Randolph GJ, Murphy K, Gautier EL, Kedl RM, Lahoud MH, et al. CD103+ Pulmonary Dendritic Cells Preferentially Acquire and Present Apoptotic Cell-Associated Antigen. J Exp Med (2011) 208:1789–97. doi: 10.1084/jem.20110538
127. Sun X, Jones HP, Dobbs N, Bodhankar S, Simecka JW. Dendritic Cells are the Major Antigen Presenting Cells in Inflammatory Lesions of Murine Mycoplasma Respiratory Disease. PloS One (2013) 8:e55984. doi: 10.1371/journal.pone.0055984
128. Ginhoux F, Liu K, Helft J, Bogunovic M, Greter M, Hashimoto D, et al. The Origin and Development of Nonlymphoid Tissue CD103+ DCs. J Exp Med (2009) 206:3115–30. doi: 10.1084/jem.20091756
129. Semmrich M, Plantinga M, Svensson-Frej M, Uronen-Hansson H, Gustafsson T, Mowat AM, et al. Directed Antigen Targeting In Vivo Identifies a Role for CD103+ Dendritic Cells in Both Tolerogenic and Immunogenic T-Cell Responses. Mucosal Immunol (2012) 5:150–60. doi: 10.1038/mi.2011.61
130. Conejero L, Khouili SC, Martinez-Cano S, Izquierdo HM, Brandi P, Sancho D. Lung CD103+ Dendritic Cells Restrain Allergic Airway Inflammation Through IL-12 Production. JCI Insight (2017) 2:e90420. doi: 10.1172/jci.insight.90420
131. Nakano H, Free ME, Whitehead GS, Maruoka S, Wilson RH, Nakano K, et al. Pulmonary CD103(+) Dendritic Cells Prime Th2 Responses to Inhaled Allergens. Mucosal Immunol (2012) 5:53–65. doi: 10.1038/mi.2011.47
132. Zelante T, Wong AY, Ping TJ, Chen J, Sumatoh HR, Vigano E, et al. CD103(+) Dendritic Cells Control Th17 Cell Function in the Lung. Cell Rep (2015) 12:1789–801. doi: 10.1016/j.celrep.2015.08.030
133. Khare A, Krishnamoorthy N, Oriss TB, Fei M, Ray P, Ray A. Cutting Edge: Inhaled Antigen Upregulates Retinaldehyde Dehydrogenase in Lung CD103+ But Not Plasmacytoid Dendritic Cells to Induce Foxp3 De Novo in CD4+ T Cells and Promote Airway Tolerance. J Immunol (2013) 191:25–9. doi: 10.4049/jimmunol.1300193
134. Engler DB, Reuter S, van Wijck Y, Urban S, Kyburz A, Maxeiner J, et al. Effective Treatment of Allergic Airway Inflammation With Helicobacter Pylori Immunomodulators Requires BATF3-Dependent Dendritic Cells and IL-10. Proc Natl Acad Sci USA (2014) 111:11810–5. doi: 10.1073/pnas.1410579111
135. Waithman J, Zanker D, Xiao K, Oveissi S, Wylie B, Ng R, et al. Resident CD8(+) and Migratory CD103(+) Dendritic Cells Control CD8 T Cell Immunity During Acute Influenza Infection. PloS One (2013) 8:e66136. doi: 10.1371/journal.pone.0066136
136. Allan RS, Waithman J, Bedoui S, Jones CM, Villadangos JA, Zhan Y, et al. Migratory Dendritic Cells Transfer Antigen to a Lymph Node-Resident Dendritic Cell Population for Efficient CTL Priming. Immunity (2006) 25:153–62. doi: 10.1016/j.immuni.2006.04.017
137. Carbone FR, Belz GT, Heath WR. Transfer of Antigen Between Migrating and Lymph Node-Resident DCs in Peripheral T-Cell Tolerance and Immunity. Trends Immunol (2004) 25:655–8. doi: 10.1016/j.it.2004.09.013
138. GeurtsvanKessel CH, Willart MA, van Rijt LS, Muskens F, Kool M, Baas C, et al. Clearance of Influenza Virus From the Lung Depends on Migratory Langerin+CD11b- But Not Plasmacytoid Dendritic Cells. J Exp Med (2008) 205:1621–34. doi: 10.1084/jem.20071365
139. Jenkins MM, Bachus H, Botta D, Schultz MD, Rosenberg AF, Leon B, et al. Lung Dendritic Cells Migrate to the Spleen to Prime Long-Lived TCF1(hi) Memory CD8(+) T Cell Precursors After Influenza Infection. Sci Immunol (2021) 6:eabg6895. doi: 10.1126/sciimmunol.abg6895
140. Hohl TM, Rivera A, Lipuma L, Gallegos A, Shi C, Mack M, et al. Inflammatory Monocytes Facilitate Adaptive CD4 T Cell Responses During Respiratory Fungal Infection. Cell Host Microbe (2009) 6:470–81. doi: 10.1016/j.chom.2009.10.007
141. Cao W, Taylor AK, Biber RE, Davis WG, Kim JH, Reber AJ, et al. Rapid Differentiation of Monocytes Into Type I IFN-Producing Myeloid Dendritic Cells as an Antiviral Strategy Against Influenza Virus Infection. J Immunol (2012) 189:2257–65. doi: 10.4049/jimmunol.1200168
142. Nakano H, Lin KL, Yanagita M, Charbonneau C, Cook DN, Kakiuchi T, et al. Blood-Derived Inflammatory Dendritic Cells in Lymph Nodes Stimulate Acute T Helper Type 1 Immune Responses. Nat Immunol (2009) 10:394–402. doi: 10.1038/ni.1707
143. Wuthrich M, Ersland K, Sullivan T, Galles K, Klein BS. Fungi Subvert Vaccine T Cell Priming at the Respiratory Mucosa by Preventing Chemokine-Induced Influx of Inflammatory Monocytes. Immunity (2012) 36:680–92. doi: 10.1016/j.immuni.2012.02.015
144. Ainsua-Enrich E, Hatipoglu I, Kadel S, Turner S, Paul J, Singh S, et al. IRF4-Dependent Dendritic Cells Regulate CD8(+) T-Cell Differentiation and Memory Responses in Influenza Infection. Mucosal Immunol (2019) 12:1025–37. doi: 10.1038/s41385-019-0173-1
145. Plantinga M, Guilliams M, Vanheerswynghels M, Deswarte K, Branco-Madeira F, Toussaint W, et al. Conventional and Monocyte-Derived CD11b(+) Dendritic Cells Initiate and Maintain T Helper 2 Cell-Mediated Immunity to House Dust Mite Allergen. Immunity (2013) 38:322–35. doi: 10.1016/j.immuni.2012.10.016
146. Nakano H, Moran TP, Nakano K, Gerrish KE, Bortner CD, Cook DN. Complement Receptor C5aR1/CD88 and Dipeptidyl Peptidase-4/CD26 Define Distinct Hematopoietic Lineages of Dendritic Cells. J Immunol (2015) 194:3808–19. doi: 10.4049/jimmunol.1402195
147. Hoeffel G, Chen J, Lavin Y, Low D, Almeida FF, See P, et al. C-Myb(+) Erythro-Myeloid Progenitor-Derived Fetal Monocytes Give Rise to Adult Tissue-Resident Macrophages. Immunity (2015) 42:665–78. doi: 10.1016/j.immuni.2015.03.011
148. Sheng J, Ruedl C, Karjalainen K. Most Tissue-Resident Macrophages Except Microglia Are Derived From Fetal Hematopoietic Stem Cells. Immunity (2015) 43:382–93. doi: 10.1016/j.immuni.2015.07.016
149. Yona S, Kim KW, Wolf Y, Mildner A, Varol D, Breker M, et al. Fate Mapping Reveals Origins and Dynamics of Monocytes and Tissue Macrophages Under Homeostasis. Immunity (2013) 38:79–91. doi: 10.1016/j.immuni.2012.12.001
150. Ina Y, Takada K, Yamamoto M, Morishita M, Yoshikawa K. Antigen-Presenting Capacity of Alveolar Macrophages and Monocytes in Pulmonary Tuberculosis. Eur Respir J (1991) 4:88–93.
151. Vecchiarelli A, Dottorini M, Pietrella D, Monari C, Retini C, Todisco T, et al. Role of Human Alveolar Macrophages as Antigen-Presenting Cells in Cryptococcus Neoformans Infection. Am J Respir Cell Mol Biol (1994) 11:130–7. doi: 10.1165/ajrcmb.11.2.8049074
152. Chakarov S, Lim HY, Tan L, Lim SY, See P, Lum J, et al. Two Distinct Interstitial Macrophage Populations Coexist Across Tissues in Specific Subtissular Niches. Science (2019) 363:eaau0964. doi: 10.1126/science.aau0964
153. Bleriot C, Chakarov S, Ginhoux F. Determinants of Resident Tissue Macrophage Identity and Function. Immunity (2020) 52:957–70. doi: 10.1016/j.immuni.2020.05.014
154. Schyns J, Bai Q, Ruscitti C, Radermecker C, De Schepper S, Chakarov S, et al. Non-Classical Tissue Monocytes and Two Functionally Distinct Populations of Interstitial Macrophages Populate the Mouse Lung. Nat Commun (2019) 10:3964. doi: 10.1038/s41467-019-11843-0
155. Osterholzer JJ, Chen GH, Olszewski MA, Zhang YM, Curtis JL, Huffnagle GB, et al. Chemokine Receptor 2-Mediated Accumulation of Fungicidal Exudate Macrophages in Mice That Clear Cryptococcal Lung Infection. Am J Pathol (2011) 178:198–211. doi: 10.1016/j.ajpath.2010.11.006
156. Taut K, Winter C, Briles DE, Paton JC, Christman JW, Maus R, et al. Macrophage Turnover Kinetics in the Lungs of Mice Infected With Streptococcus Pneumoniae. Am J Respir Cell Mol Biol (2008) 38:105–13. doi: 10.1165/rcmb.2007-0132OC
157. Lin KL, Suzuki Y, Nakano H, Ramsburg E, Gunn MD. CCR2+ Monocyte-Derived Dendritic Cells and Exudate Macrophages Produce Influenza-Induced Pulmonary Immune Pathology and Mortality. J Immunol (2008) 180:2562–72. doi: 10.4049/jimmunol.180.4.2562
158. Jakubzick C, Gautier EL, Gibbings SL, Sojka DK, Schlitzer A, Johnson TE, et al. Minimal Differentiation of Classical Monocytes as They Survey Steady-State Tissues and Transport Antigen to Lymph Nodes. Immunity (2013) 39:599–610. doi: 10.1016/j.immuni.2013.08.007
159. Shi C, Pamer EG. Monocyte Recruitment During Infection and Inflammation. Nat Rev Immunol (2011) 11:762–74. doi: 10.1038/nri3070
160. Jakubzick CV, Randolph GJ, Henson PM. Monocyte Differentiation and Antigen-Presenting Functions. Nat Rev Immunol (2017) 17:349–62. doi: 10.1038/nri.2017.28
161. Larson SR, Atif SM, Gibbings SL, Thomas SM, Prabagar MG, Danhorn T, et al. Ly6C(+) Monocyte Efferocytosis and Cross-Presentation of Cell-Associated Antigens. Cell Death Differ (2016) 23:997–1003. doi: 10.1038/cdd.2016.24
162. Dunbar PR, Cartwright EK, Wein AN, Tsukamoto T, Tiger Li ZR, Kumar N, et al. Pulmonary Monocytes Interact With Effector T Cells in the Lung Tissue to Drive TRM Differentiation Following Viral Infection. Mucosal Immunol (2020) 13:161–71. doi: 10.1038/s41385-019-0224-7
163. Krausgruber T, Fortelny N, Fife-Gernedl V, Senekowitsch M, Schuster LC, Lercher A, et al. Structural Cells are Key Regulators of Organ-Specific Immune Responses. Nature (2020) 583:296–302. doi: 10.1038/s41586-020-2424-4
164. Shenoy AT, Wasserman GA, Arafa EI, Wooten AK, Smith NMS, Martin IMC, et al. Lung CD4(+) Resident Memory T Cells Remodel Epithelial Responses to Accelerate Neutrophil Recruitment During Pneumonia. Mucosal Immunol (2020) 13:334–43. doi: 10.1038/s41385-019-0229-2
165. Wong MH, Johnson MD. Differential Response of Primary Alveolar Type I and Type II Cells to LPS Stimulation. PloS One (2013) 8:e55545. doi: 10.1371/journal.pone.0055545
166. Fehrenbach H. Alveolar Epithelial Type II Cell: Defender of the Alveolus Revisited. Respir Res (2001) 2:33–46. doi: 10.1186/rr36
167. Toulmin SA, Bhadiadra C, Paris AJ, Lin JH, Katzen J, Basil MC, et al. Type II Alveolar Cell MHCII Improves Respiratory Viral Disease Outcomes While Exhibiting Limited Antigen Presentation. Nat Commun (2021) 12:3993. doi: 10.1038/s41467-021-23619-6
168. Gereke M, Jung S, Buer J, Bruder D. Alveolar Type II Epithelial Cells Present Antigen to CD4(+) T Cells and Induce Foxp3(+) Regulatory T Cells. Am J Respir Crit Care Med (2009) 179:344–55. doi: 10.1164/rccm.200804-592OC
169. Debbabi H, Ghosh S, Kamath AB, Alt J, Demello DE, Dunsmore S, et al. Primary Type II Alveolar Epithelial Cells Present Microbial Antigens to Antigen-Specific CD4+ T Cells. Am J Physiol Lung Cell Mol Physiol (2005) 289:L274–279. doi: 10.1152/ajplung.00004.2005
170. Wein AN, McMaster SR, Takamura S, Dunbar PR, Cartwright EK, Hayward SL, et al. CXCR6 Regulates Localization of Tissue-Resident Memory CD8 T Cells to the Airways. J Exp Med (2019) 216:2748–62. doi: 10.1084/jem.20181308
171. Niethamer TK, Stabler CT, Leach JP, Zepp JA, Morley MP, Babu A, et al. Defining the Role of Pulmonary Endothelial Cell Heterogeneity in the Response to Acute Lung Injury. Elife (2020) 9:e53072. doi: 10.7554/eLife.53072
172. Claser C, Nguee SYT, Balachander A, Wu Howland S, Becht E, Gunasegaran B, et al. Lung Endothelial Cell Antigen Cross-Presentation to CD8(+)T Cells Drives Malaria-Associated Lung Injury. Nat Commun (2019) 10:4241. doi: 10.1038/s41467-019-12017-8
173. McMaster SR, Wein AN, Dunbar PR, Hayward SL, Cartwright EK, Denning TL, et al. Pulmonary Antigen Encounter Regulates the Establishment of Tissue-Resident CD8 Memory T Cells in the Lung Airways and Parenchyma. Mucosal Immunol (2018) 11:1071–8. doi: 10.1038/s41385-018-0003-x
174. Jung YW, Kim HG, Perry CJ, Kaech SM. CCR7 Expression Alters Memory CD8 T-Cell Homeostasis by Regulating Occupancy in IL-7- and IL-15-Dependent Niches. Proc Natl Acad Sci USA (2016) 113:8278–83. doi: 10.1073/pnas.1602899113
175. Mohammed J, Beura LK, Bobr A, Astry B, Chicoine B, Kashem SW, et al. Stromal Cells Control the Epithelial Residence of DCs and Memory T Cells by Regulated Activation of TGF-Beta. Nat Immunol (2016) 17:414–21. doi: 10.1038/ni.3396
Keywords: lung, antigen presentation, antigen cross presentation, dendritic cells, macrophages
Citation: Kawasaki T, Ikegawa M and Kawai T (2022) Antigen Presentation in the Lung. Front. Immunol. 13:860915. doi: 10.3389/fimmu.2022.860915
Received: 24 January 2022; Accepted: 15 April 2022;
Published: 09 May 2022.
Edited by:
Elodie Segura, Institut Curie, FranceReviewed by:
Natalio Garbi, University of Bonn, GermanyHideki Nakano, National Institute of Environmental Health Sciences (NIH), United States
Copyright © 2022 Kawasaki, Ikegawa and Kawai. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Takumi Kawasaki, kawast01@bs.naist.jp; Taro Kawai, tarokawai@bs.naist.jp