 Yasmeen Faroqi-Shah
Yasmeen Faroqi-Shah- Department of Hearing and Speech Sciences, University of Maryland, College Park, MD, United States
The language production deficit in post-stroke agrammatic aphasia (PSA-G) tends to result from lesions to the left inferior frontal gyrus (LIFG) and is characterized by a triad of symptoms: fragmented sentences, errors in functional morphology, and a dearth of verbs. Despite decades of research, the mechanisms underlying production patterns in PSA-G have been difficult to characterize. Two major impediments to progress may have been the view that it is a purely morphosyntactic disorder and the (sometimes overzealous) application of linguistic theory without interceding psycholinguistic evidence. In this paper, empirical evidence is examined to present an integrated portrait of language production in PSA-G and to evaluate the assumption of a syntax-specific syndrome. In light of extant evidence, it is proposed that agrammatic language production results from a combination of morphosyntactic, phonomotor, and processing capacity limitations that cause a cumulative processing bottleneck at the point of articulatory planning. This proposed Synergistic Processing Bottleneck model of PSA-G presents a testable framework for future research. The paper ends with recommendations for future research on PSA-G.
Introduction
Agrammatism, which refers to morphosyntatic impairment in post-stroke aphasia (PSA), has been a poster-child for the neurocognitive modularity of morphosyntactic processes. Agrammatic aphasia has also been a testing ground for linguistic theories and inferences about Broca's area function (e.g., Grodzinsky, 2000; Patel et al., 2008). Given that there are differences in the use of the diagnostic label of agrammatism, we clarify at the outset that this paper focuses on language production in agrammatic post-stroke aphasia (PSA-G). While morphosyntactic impairments are also acquired from other etiologies such as neurodegeneration in primary progressive aphasias (PPA), this paper focuses on PSA-G because there is a significantly larger body of empirical and theoretical research on agrammatic post-stroke aphasia. Consequently, it allows for a more extensive synthesis of morphosyntactic production deficits following stroke. Further, by focusing on a single etiology, we avoid the (yet unconfirmed) assumption that the same neurocognitive mechanism underlies morphosyntactic impairment in both post-stroke and progressive aphasias. For instance, it is likely that domain-general bilateral neural circuitry is recruited to compensate for language deficits much earlier in the evolution of PPA (compared to PSA) given its insidious disease progression (Sonty et al., 2003; Canu et al., 2020). Asyntactic comprehension, which refers to the pattern of worse understanding of syntactically complex and semantically reversible sentences compared to syntactically simple sentences, is not uniquely and consistently associated with agrammatic production (Caramazza and Zurif, 1976; Miceli et al., 1983; Kolk and van Grunsven, 1985). It is found across a variety of aphasia subtypes, as well as in neurotypical speakers under high cognitive load (Caplan et al., 2007, 2013). Hence this paper views morphosyntactic production deficits as the core component of PSA-G.
Despite over four decades of research on PSA-G, there is not yet a comprehensive understanding of what kinds of deviations from normal sentence production mechanism(s) give rise to the symptom cluster of agrammatic language production. While there have been some mechanistic accounts of PSA-G, such as Pick's (1913) Economy of effort and Kolk's (1995) Time-based approach, most extant accounts of PSA-G are linguistic descriptions of a single symptom (e.g., Thompson, 2003). While these descriptive accounts have provided symptom details (e.g., grammatical functions of words are more impaired than their lexical functions, Boye and Bastiaanse, 2018), such accounts do not explain why a symptom occurs in PSA-G.
The goal of this paper is to describe a mechanistic model of language production in PSA-G as a way move forward from the current stalemate. The model, which is referred to as the Synergistic Processing Bottleneck model, views PSA-G's morphosyntactic deficit as part of a broader clinical profile and is developed from an integration of empirical findings on neurotypical and agrammatic sentence production. It provides a testable framework for future research. In the build-up to the model, this paper is organized as follows: First, issues of empirical rigor in PSA-G research are raised. Next, current empirical evidence on PSA-G symptoms is integrated and critically evaluated to constrain the symptoms that must be accommodated in any theory of PSA-G. Next, the complex clinical picture of PSA in which morphosyntactic deficits occur is presented as a rationale for broadening the theoretical view of PSA-G. Following this, extant theories of PSA-G are briefly discussed in their ability to account for the clinical profile of PSA-G. Finally, in the Discussion section, the Synergistic Processing Bottleneck model is presented as a synthesis of the syntactic and non-syntactic symptoms of PSA-G with current understanding of how sentence production unfolds in neurologically healthy speakers. The paper ends with recommendations for future agrammatism research.
Empirical rigor in PSA-G research
Issues of empirical rigor and reproducibility have been identified in PSA-G research by several authors (Caplan, 1995, 2001; Mauner, 1995; Caramazza et al., 2001; Martin, 2006; Faroqi-Shah, 2020). These issues have significantly weakened the inferences that could be made from the data and has partly contributed to the current standstill in agrammatism theories. The goal of this section is to highlight criteria for scientific rigor that are particularly relevant to the study of PSA-G.
Diagnosis of PSA-G
The first and most important criterion is the diagnosis of PSA-G. A scoping review found that two-thirds of studies do not report any language scores to document agrammatism; and nearly half of the studies do not operationally define the condition, using proxies such as non-fluent or Broca's aphasia (Faroqi-Shah, 2020). There is no explicit consensus on what constitutes “agrammatism” (Berndt and Caramazza, 1981; Miceli et al., 1989; Martin, 2006; Thompson and Bastiaanse, 2012; den Ouden et al., 2019). While early descriptions solely focused on language production symptoms (Pick, 1913; Kleist, 1916; Goodglass and Berko, 1960), the term has evolved (for some authors) to include a sentence comprehension deficit (Berndt and Caramazza, 1981; Grodzinsky, 1984; Avrutin, 2000). The notion of a comprehension deficit as a core symptom of agrammatic production deficit has surprisingly persisted despite numerous studies showing the dissociation between sentence production and comprehension deficits (Caramazza and Zurif, 1976; Berndt and Caramazza, 1981; Miceli et al., 1983; Kolk and van Grunsven, 1985; Martin et al., 1989). In fact, in their classic study, Caramazza and Zurif (1976) reported that both people with (Broca's aphasia) and without (conduction aphasia) agrammatic production showed the same pattern of sentence comprehension deficit.
Additionally, but to a lesser extent, there are different views on the association between agrammatic production and a slow rate of speech (non-fluency). For example, De Villiers (1974) analyzed the speech of non-fluent aphasic speakers with “varying degrees of grammatical impairment ranging from almost intact to severely impaired.” (p. 38). Similarly, Saffran et al. (1989) described the narrative language of speakers who produced “sparse halting speech” (i.e., nonfluent) and divided these speakers into “agrammatic” and “non-fluent non-agrammatic” speakers (p. 446). However, this nuanced yet important distinction between non-fluency and agrammatism seems to have been lost to overzealous theoretical syntacticians in later years. In some studies of PSA-G, participants are recruited based on their non-fluent speech, but there is no further characterization of the nature of morphosyntactic production errors (as noted by Faroqi-Shah, 2020; for example see O'Grady and Lee, 2001). The importance of this relationship between non-fluency and agrammatism will be discussed in a later section.
In clinical settings, it is important to note that physicians may identify aphasia in a neurological examination, however they do not possess the specialized training or standardized assessments to diagnose and differentiate between motor speech and (subtypes of) linguistic deficits. Speech-language pathologists (SLP) are qualified and specially trained to delineate the nuances of communication impairments in aphasia such as speech versus language deficits. In the absence of speech-language pathologists in countries where such a profession does not exist, it is critical to operationalize the inclusionary and exclusionary criteria for PSA-G participants. This was elegantly achieved by Menn and Obler (1990) when comparing PSA-G speakers across fourteen languages: “Agrammatic by clinical standards was defined as being moderately non-fluent, having slow and halting speech, with three or four words being the usual maximum uninterrupted string” (p. 14).
In light of the above ambiguities in operationally defining and diagnosing PSA-G, a critical step in assessing the internal and external validity of any study is whether the authors operationally defined agrammatism, described how participants were diagnosed as PSA-G, and characterized the morphosyntactic profile of participant symptoms. It is important to note whether authors define PSA-G as a purely production disorder, or also assume difficulties in complex sentence comprehension. In short, a study that provides relevant language and clinical characteristics of the participants has greater validity than one that just uses generic terms (e.g., non-fluent aphasia).
Experimental design and inferencing
A second criterion in determining empirical rigor of the findings is the inferential strength of the experimental design. Double dissociations are a key inferential tool in neuropsychological research (Caramazza, 1984; Crawford et al., 2003). This refers to the demonstration that two individuals (or clinical groups) show deficits that are the inverse of one another. For instance, a double dissociation between verb and noun deficits has been shown in agrammatic and anomic aphasia (Miceli et al., 1988; Zingeser and Berndt, 1990; Lee and Thompson, 2011a). However, a majority of PSA-G studies have no comparison group or use a neurotypical “control” group (Faroqi-Shah, 2020). When studies compare PSA-G with a neurotypical group, it shows a single dissociation in which one cannot delineate the unique characteristics of agrammatism from the general impact of aphasia. The inferential power of studies that do include an aphasic comparison (e.g., fluent aphasia or anomic aphasia) may be further weakened if they do not meet the first criterion of accurate diagnosis. An example of this paradox is the frequently cited case study of a double dissociation between regular and irregular past tense in non-fluent and fluent aphasia (Ullman et al., 1997). “Fluent aphasia” is an obscure diagnosis which includes PSA profiles as disparate as anomic aphasia (with mild word retrieval issues) and Wernicke's aphasia (with severe semantic, phonological and self-monitoring challenges). Another challenge in double dissociation studies is matching (or statistically addressing) overall aphasia severity across groups. When composite language scores on standardized tests are used, such as the Aphasia Quotient on the Western Aphasia Battery (Kertesz, 2006), persons with PSA-G tend to score more severely than the comparison group due to their severe production difficulties (see for example the PSA-G vs. non-PSA-G in Faroqi-Shah et al., 2020).
Mediating, moderating, and confounding factors
Another interpretive over-simplification in agrammatism research is the assumption that experimental task performance directly measures the underlying linguistic deficit(s) without other mediating or moderating factors. PSA respond well to speech-language therapy (Brady et al., 2016) and may have engaged in different intervention programs as well as self-guided (or care-giver guided) language practice before partaking in the research study. Thus, researchers rarely measure “pure agrammatism.” For instance, Script Training is a popular and effective intervention for sentence production deficits (Cherney et al., 2008) which might result in the overuse of structural templates such as I am x (x = happy/hungry/eating) or Noun is Verbing (Mom is calling, Dog is eating, etc.). Even before intervention is initiated, for example in the acute phase of stroke, there are significant cognitive and fatigue issues that could mask agrammatism (Adamson et al., 2004; Engelter et al., 2006; Nys et al., 2007; Cumming et al., 2013). In fact, stroke results in a variety of cognitive deficits, which can be severe in PSA (Murray, 2012; Chapman and Hallowell, 2021; Faroqi-Shah et al., 2022b). Short-term memory and working memory have been particularly identified as influencing sentence production and comprehension in PSA and in neurotypical speakers (Caplan et al., 2013; Wright et al., 2014; Fyndanis et al., 2018; but see Ivanova and Ferreira, 2019). Cognitive deficits may limit the overall processing capacity for linguistic computations. Perceptual and motor impairments occur in nearly 74 and 85% of stroke survivors respectively and can be persistent (Mayo et al., 1999; Hazelton et al., 2022). There are also psychological effects of stroke such as depression, anxiety, and post-traumatic stress disorder, which are incident in nearly 70% of stroke survivors (Kauhanen et al., 2000; Assayag et al., 2022; Pompon et al., 2022; Skajaa et al., 2022). This is illustrated in Figure 1.
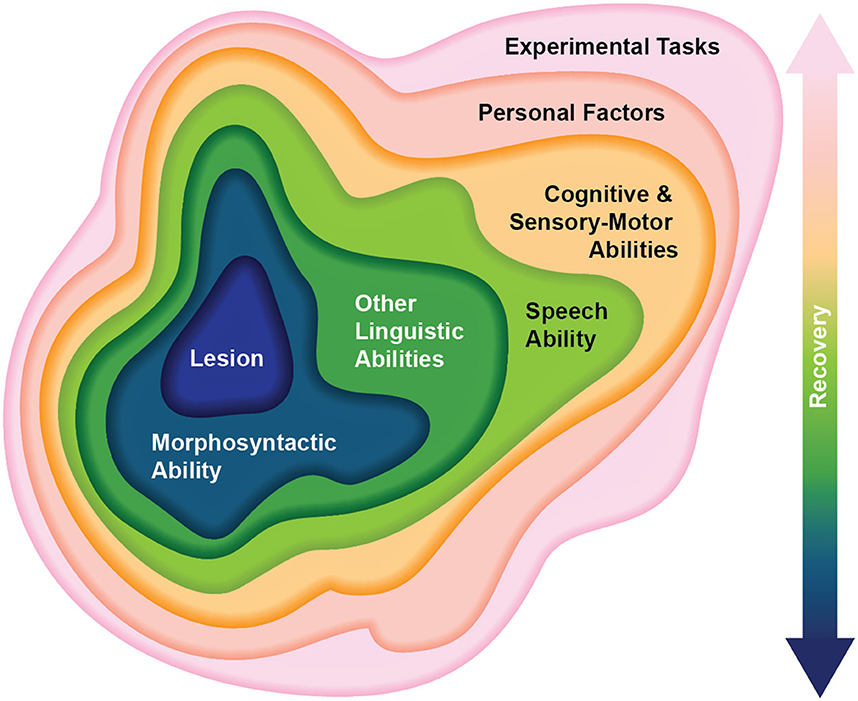
Figure 1. Epistemological framework of the complex clinical picture of post-stroke agrammatic aphasia. The key point is that numerous mediating, moderating, confounding and covarying factors intervene between the deficit (morphosyntactic ability) and the dependent variables measured in experimental tasks (outer layer). From the inner layer: Lesion refers to the anatomical integrity of gray and white matter, including structural and functional connectivity; Morphosyntactic ability is the key deficit of interest in agrammatic aphasia that researchers are trying to understand; Other linguistic abilities refer to concomitant language impairments in semantic, phonological, and orthographic domains; Speech ability represents concomitant non-linguistic impairments in motor control and execution (apraxia of speech and dysarthria) that can impact verbal production of language; Cognitive and Sensory-Motor abilities include any number of non-linguistic impairments resulting from stroke, such as short/long term memory, processing speed, processing capacity, attention, cognitive control, perception (including visual and auditory) and motor control (including limb paresis and praxis); Personal factors include a post-stroke mental disorders, fatigue, cultural and linguistic differences, education, task familiarity; Experimental tasks yield the dependent variables that researchers use to make inferences about morphosyntactic ability; and Recovery refers to the evolving severity of linguistic and non-linguistic abilities that is moderated by neurological recovery, speech-language therapy, and self-generated strategies. The sequence of the intervening layers is flexible.
To summarize, PSA-G is one aspect of a multidimensional clinical profile, and numerous variables intervene between the actual morphosyntactic deficit and the empirical measure(s) obtained by researchers. These variables may affect experimental measures based on whether they are covariates, confounds, moderators, or mediators. A covariate affects the outcome variable but is not related to the independent variable (e.g., limb paresis for a keyboard response); a confound is associated with both the independent and dependent variables (e.g., short-term memory deficit), but does not drive the association between them; a mediator is a causal variable, such that the independent variable causes it, which in turn drives the dependent variable; and a moderator is not on the causal pathway but interacts with the independent variable in a way that drives the outcome (e.g., speech-language treatment) (Morrow et al., 2022). Currently, we lack a clear understanding of which (and how) different variables interact in PSA-G. This knowledge will not only improve how researchers statistically address and interpret outcome measures, but will also improve our understanding of individual variability in PSA-G. It is important to demonstrate how confounds from understanding task demands, memory demands, or lexical retrieval difficulties were addressed in data analysis and interpretation. For example, to address the fact that verb retrieval failures could confound the accuracy of producing verb inflections, Faroqi-Shah and Thompson (2004) analysis of the production of verb inflections only included verbs that were correctly named by each participant in a separate confrontation naming task. At minimum, a study should document screening of intuitive variables based on the experimental task, such as hearing loss and short-term memory for auditory comprehension and apraxia of speech (AoS) for verbal production (for example, see Szupica-Pyrzanowska et al., 2017).
Measurement reliability
The fourth criterion in demonstrating empirical rigor is the reliability of subjective measures of key dependent variables. When reporting accuracy of verbal productions, it is important to operationally define a correct response, describe how phonemic/semantic paraphasias were scored, and report how bias in scoring was addressed. For narrative language analyses, there are multiple sources of subjectivity, including transcription, utterance segmentation, and error coding. Ideally, studies should describe their reliability procedures including procedures for resolution of discrepancies, and should report inter-rater reliability (for sample studies that report reliability/consensus, see Rochon et al., 2000; Matchin and Hickok, 2020).
Summary of empirical rigor in PSA-G research
The four empirical issues listed in this section highlight the need to closely examine the methods adopted by PSA-G studies before drawing conclusions about what constitute the core symptoms of PSA-G and what mechanisms underlie the morphosyntactic impairment in PSA-G. In the following sections of this paper, to the extent possible, the above-mentioned criteria were applied to evaluate and select studies for discussion. That is, studies that provided a clear operational definition of agrammatic aphasia, detailed language and clinical characteristics of participants, reported double dissociations with appropriate statistical treatment, and addressed potential confounds in measurement of the dependent variable, were prioritized over studies that used vague terms (e.g., non-fluent aphasia) without language measures, and did not present a non-agrammatic PSA comparison (individual or group). Interpretations from meta-analyses were given more weight over individual studies as they wash out study-specific differences, and effects are computed over a larger number of participants. In the next section, empirical research is evaluated using these criteria in an attempt to identify the essential components of PSA-G.
Establishing the core morphosyntactic findings in PSA-G
“Boy… girl… cookie jar… mother… water… wash dishes.” This utterance illustrates the triad of deficits that characterize PSA-G: fragmented utterances, errors on functional morphemes, and missing verbs (Tissot et al., 1973; De Villiers, 1974; Goodglass, 1976; Saffran et al., 1989; Zingeser and Berndt, 1990; Goodglass et al., 1993; Bastiaanse and Thompson, 2012). These three symptoms are not mutually exclusive—a sentence may be fragmented because it is missing the verb and/or functional morphemes. Recent empirical support for this cluster of symptoms comes from a principal components analysis of 27 perceptual features of spoken language in an unselected group of English-speaking PSA (Casilio et al., 2019). The analysis found that four of the 27 features clustered together in aphasic speakers: stereotypies and automatisms, short and simplified utterances, omission of function words, and omission of bound morphemes. However, there is heterogeneity among PSA-G for the extent of these individual symptoms (Miceli et al., 1989; Rochon et al., 2000; Dickey and Thompson, 2009).
Following the seminal work of Caramazza and Zurif (1976), who reported difficulties in the comprehension of syntactically complex and semantically reversible sentences in Broca's and conduction aphasia (e.g., The cow that the monkey is scaring is yellow), some researchers included comprehension deficit as an additional symptom. This paper will refer to this comprehension pattern as asyntactic comprehension to avoid confusion with the triad of agrammatic production symptoms. This section will examine empirical findings for the above four symptoms to identify core symptoms that a theory of PSA-G should accommodate.
Sentence production
Three types of tasks have been utilized in PSA-G literature: (1) narrative samples elicited using story retell, picture descriptions, or personal experiences, (2) constrained elicitation of entire sentences or parts of sentences, and (3) arrangement of written word or phrase “anagrams” to construct a sentence. Comparisons across these tasks allow us to compare PSA-G's performance across different cognitive demands (and hence processing load), and delineate syntactic knowledge (anagram task) from performance deficits.
The following generalizations can be made by comparing findings across studies. First, about 30–50% of PSA-G's utterances are syntactically well-formed (Saffran et al., 1989; Rochon et al., 2000; Hsu and Thompson, 2018). Second, very severely impaired PSA-G individuals produce a predominance of 1-2 word fragments (“telegraphic speech”) that does not show any semblance of word order. Such individuals also overuse stereotypical and automatic utterances (e.g., Oh God, I don't know) (Ishkhanyan et al., 2017). Third, the canonical word order of the speaker's language is preserved and is often over-used (Bates et al., 1987; Menn and Obler, 1990; Bastiaanse and Edwards, 2004). This has been interpreted as preservation of language-specific usage patterns (see also Bates et al., 1991; Gahl and Menn, 2016). Fourth, although their use of non-canonical sentence structures is limited, it is not clear that this pattern is unique to PSA-G because the same has been reported across aphasia categories and across elicitation tasks (Bates et al., 1991; Edwards, 1998; Faroqi-Shah and Thompson, 2003; Man et al., 2019). Fifth, word order errors or role reversals have been reported in both anagram and constrained picture descriptions. “A key unresolved question” about these errors is whether it reveals a failure of “function assignment” (Bock and Levelt, 1994). Function assignment refers to event conceptualization and ability to assign thematic functions to entities. Evidence points to relatively preserved function assignment based on the finding that the incidence of word order errors is low in anagram tasks (about 8.5% in: von Stockert and Bader, 1976; Saffran et al., 1980; Scholes, 1982). Further, when authors report error patterns in picture-sentence elicitation tasks, function assignment seems to be preserved. For example, Faroqi-Shah and Thompson (2003) compared passive sentence production between individuals with PSA-G and Wernicke's aphasia and varied the amount of lexical cues provided. Both groups showed similar accuracy of passive sentences across conditions. As is evident from the excerpts below, they also showed awareness of their role reversals. In short, there is no clear evidence suggesting a function assignment deficit in PSA-G.
Broca's #7: “The (the the) guy is helping the bicyclist… The other way around… The man is… The man is quaching the priest eh the bicycle.” (Target: The cyclist is helped by the hunter).
Wernicke #4: “Wife is going to cover the husband. That doesn't sound right. How do you do that?” (Target: The wife is covered by the husband).
In addition to function assignment, PSA-G also show other preserved sentence production abilities. This includes responsiveness to structural priming, a phenomenon that is well-documented in neurotypical speakers (Bock and Loebell, 1990; Pickering and Ferreira, 2008). Cho-Reyes et al. (2016) used the classic structural priming paradigm where speakers repeat a prime sentence (e.g., passive or double-object dative) followed by describing a picture showing an action. Priming of sentence structure is indicated when speakers re-use the syntactic frame of the prime sentence in their picture description. PSA-G speakers not only showed structural priming effects, but also the magnitude of priming was comparable to neurotypical speakers (Cho-Reyes et al., 2016). However, there is considerable variability in the extent to which PSA respond to structural priming and produce complex sentences (den Ouden et al., 2019). Further, visual world paradigms (eye-tracking) show that PSA-G speakers plan their sentences incrementally, starting with the subject noun, just like neurotypical speakers (Lee and Thompson, 2011b; Lee et al., 2015). These studies show that the scope of sentence planning is similar to neurotypical speakers although the timecourse of planning is slower.
To summarize, PSA-G produce incomplete fragments and overuse canonical word order (Bates et al., 1987; Menn and Obler, 1990). There is no strong evidence to indicate that they have a deficit in activating complex syntactic structures from primed sentences, incremental planning, or function assignment (Lee and Thompson, 2011b; Lee et al., 2015; Cho-Reyes et al., 2016).
Functional morphology
Substitutions and omissions of free and bound functional morphemes are a hallmark feature of PSA-G across languages (Bates et al., 1987; Menn and Obler, 1990; Casilio et al., 2019). When comparing PSA-G with other PSA, vulnerability of grammatical morphology seems to be particularly unique to PSA-G (Saffran et al., 1989; Menn and Obler, 1990; Bates et al., 1991; Goodglass et al., 1993; but see Haarmann and Kolk, 1992). Three aspects of functional morphology in PSA-G have been extensively studied. The first is the role of morphological complexity (e.g., Ullman et al., 1997; Lambon Ralph et al., 2005). Much of this work was framed within the dual route model of inflection, which proposed that regular and irregular inflections (e.g., past tense in English) utilize different neural mechanisms. The claim was a double dissociation such that regular inflections can be selectively impaired in PSA-G (e.g., Ullman et al., 1997). A meta-analysis of published data (N = 75) found no difference in the accuracy of regular vs. irregular verb inflections (Faroqi-Shah, 2007). In another study, PSA-G produced affixed verbs in 75% of their responses, showing no specific difficulty with affixation per se (Faroqi-Shah and Thompson, 2004). The meta-analysis and several other studies highlighted the confound between phonological and morphological complexity, not only because morphologically complex stimuli tend to be phonologically complex, but also because of the co-occurrence of both types of deficits in PSA-G (Obler et al., 1999; Kohn and Melvold, 2000; Bird et al., 2003; Braber et al., 2005; Faroqi-Shah et al., 2010; Szupica-Pyrzanowska et al., 2017).
The second topic of extensive research is the semantic and syntactic role subserved by the functional morphemes. Some morphemes refer to a component of the speaker's message, such as numerosity, event time, and pronominal reference, while other morphemes serve a syntactic well-formedness function, such as subject-verb agreement. The following picture of morphological production has emerged in PSA-G. Studies show that verb morphology is less accurately produced than noun morphology (e.g., plural and determiner) (Goodglass et al., 1993). Within verb morphology, tense marking is generally worse than agreement marking and non-finite verbs although there is some inter-individual variability (Wenzlaff and Clahsen, 2004; Faroqi-Shah and Thompson, 2007; Bastiaanse and Thompson, 2012; Fyndanis et al., 2012; Zhang and Hinzen, 2022; but see Burchert et al., 2005). A meta-analysis (N = 143) showed that there is no difference in accuracy among verb tenses, that is past vs. present vs. future tense (Faroqi-Shah and Friedman, 2015). Studies have found that tense comprehension is also impaired (Dickey et al., 2008) and this correlates with the magnitude of tense production deficit (Faroqi-Shah and Dickey, 2009).1
The third line of research has examined usage patterns. In terms of the relative frequency of occurrence of different morphemes, the proportions parallel what is found in neurotypical speakers of that language (De Villiers, 1974; Blackwell and Bates, 1995; Centeno et al., 1996; Centeno, 2007, 2012). That is, the best preserved morphemes are those that are most frequent in the language. This pattern mirrors the overuse of canonical word order described in the previous section (Menn and Obler, 1990; Bates et al., 1991). Beyond language use patterns that drive the overall occurrence of morphemes, Faroqi-Shah and Thompson (2004) found two patterns. First, there was a frequency effect in substitutions of verb inflections: more frequent inflectional variants of a verb (e.g., cooking) were produced for less frequent targets (e.g., cooked) (see also Centeno et al., 1996; Centeno, 2007). Second, individual participants with PSA-G overused a specific verb form. For example, CH overused Verb+ing, RH overused Verb+ed and LD overused the verb stem (Figure 2 in Faroqi-Shah and Thompson, 2004).
To conclude, verb morphology is particularly vulnerable in PSA-G (Goodglass et al., 1993). In languages that mark tense on the verb, the difficulty is found both in production and comprehension (Clahsen and Ali, 2009; Faroqi-Shah and Dickey, 2009). Language use patterns have a major influence on what is produced with an overuse of more frequent morphological forms and little effect of verb regularity or tense type (Faroqi-Shah, 2007; Faroqi-Shah and Friedman, 2015).
Verbs
A double dissociation between verb vs. noun retrieval in agrammatic vs. anomic aphasia was first reported in the late 1980s (Miceli et al., 1988; Zingeser and Berndt, 1990; Bates et al., 1991). Later studies found that verb deficits were not inherently tied to agrammatic (or Broca's) aphasia, and occurred in persons with fluent aphasia as well (Berndt et al., 1997b; Bastiaanse and Jonkers, 1998; Edwards, 1998; Matzig et al., 2009). A meta-analysis (N = 175) of picture naming data found that the pattern of worse verb naming was found across a majority of fluent and non-fluent persons with aphasia (Matzig et al., 2009). Irrespective of the presence of agrammatism, there is an association between reduced verb naming ability (measured with action picture naming) and the production of shorter sentences and impoverished sentence structure (Berndt et al., 1997a; Edwards, 1998; Speer and Wilshire, 2013). In summary, while verb retrieval deficits are not unique to PSA-G, there is nevertheless an association between verb retrieval and sentence elaboration deficits.
To elucidate the source of verb deficits in aphasia, investigators have examined dimensions along which verbs vary, such as transitivity, imageability, instrumentality, and noun homophony (Bastiaanse and Jonkers, 1998; Bird et al., 2000; Kim and Thompson, 2000, 2004; Arévalo et al., 2007; Stavrakaki et al., 2011). The logic is that these variables denote representational complexity of verbs, thus potentially influencing verb breakdown in aphasia. Two variables are particularly relevant to syntactic deficits: verb argument structure and verb weight. Syntactic complexity of a verb is often represented by verb argument structure (VAS), which refers to the number of arguments a verb requires and the number of different argument alternations the verb takes. Thompson's (2003) Argument Structure Complexity Hypothesis proposed that verbs with more complex VAS are more impaired in PSA-G. This pattern is supported in picture naming data, where PSA-G have been compared to comparison groups of Alzheimer's disease and anomic aphasia, showing a double dissociation (Kim and Thompson, 2004; Cho-Reyes and Thompson, 2012). However, in picture naming, VAS is confounded by visual complexity of the pictures because verbs with more complex VAS (e.g., a ditransitive such as giving) are represented by a more complex visual scene compared to scenes that can be named by a verb with a simpler VAS (e.g., barking). Indeed, picture complexity is known to influence verb retrieval in neurotypical speakers (Szekely et al., 2005; Faroqi-Shah et al., 2021). The Argument Structure Complexity Hypothesis has not been borne out in narrative language, where VAS complexity effects have not been found (Webster et al., 2001; Malyutina and den Ouden, 2017). In fact, a large corpus study found that neurotypical and PSA speakers used a variety of verbs with simple and complex VAS. Persons with Broca's aphasia, however, used less complex and diverse VAS elaborations compared to other speaker groups (Malyutina and den Ouden, 2017) and produced fewer adjuncts (Zhang and Hinzen, 2022). If Broca's aphasia is taken as a proxy for PSA-G, then it appears that although verbs are used in sentences, their VAS may not be fully elaborated. Consistent with this, in constrained sentence production tasks where participants are required to retrieve the verb and its arguments to produce a complete sentence, sentences with complex VAS verbs are less well-formed and less complex compared to sentences with simpler VAS verbs (e.g., Dragoy and Bastiaanse, 2010; Malyutina and Zelenkova, 2020). However, it is unclear if this finding is specific to PSA-G because studies either report single dissociations (e.g., Dragoy and Bastiaanse, 2010), insufficiently characterize the morphosyntactic production deficit of PSA-G (“nonfluent” participants in Malyutina and Zelenkova, 2020), or find no differences across aphasia subtypes (Jonkers and Bastiaanse, 1996; Caley et al., 2017; Malyutina and Zelenkova, 2020). While some studies have noted that syntactic complexity has an additive effect with VAS complexity (e.g., Bastiaanse and van Zonneveld, 1998, 2005), other studies have not found this effect (Kok et al., 2007). In an eye-tracking study in which real-time access to verb argument structure information was examined, PSA-G showed spared access to overtly expressed VAS, but showed delays in retrieving VAS information when the argument was not explicitly provided (Mack et al., 2013).
Another dimension of verb complexity is its semantic specificity, referred to as verb weight. At one extreme are light verbs, a specific subset of very frequent, semantically underspecified verbs whose meaning can vary widely according to context (e.g., go, do, make, give). Light verbs are often grammaticalized cross-linguistically (i.e., behave like grammatical morphemes) and take a diverse variety of complements, making them syntactically complex. Heavy verbs, which are semantically more specific, were contrasted with light verbs by Gordon and Dell (2003) in the Division of Labor hypothesis between semantics and syntax. It was proposed that aphasic persons with weaker syntactic abilities would be worse at producing light verbs and vice versa. These predictions were borne out in double dissociations (Kim and Thompson, 2004; Barde et al., 2006). This division of labor between semantic and syntactic complexity of verbs was further supported in a large corpus of 164 persons with aphasia, which found a trade-off (negative correlation) between verb naming in confrontation (most of which are heavy verbs) and (1) light verb use and (2) syntactic productivity (Thorne and Faroqi-Shah, 2016) (replicating Berndt et al., 1997a; Webster et al., 2001).
To summarize the empirical findings on verb deficit in PSA-G, they show a paucity of verbs in narrative language, due to which a verb deficit is recognized as one of the three core symptoms of the agrammatic production (e.g., Tissot et al., 1973). Across PSA subtypes (not just PSA-G), there is a negative association between verb retrieval abilities at the single word level and sentence well-formedness and complexity (Berndt et al., 1997a; Thorne and Faroqi-Shah, 2016). In picture naming tasks where a single word label is elicited, both PSA-G and non-agrammatic PSA show verb retrieval difficulties (e.g., Matzig et al., 2009). In picture naming, there is some evidence of double dissociations between PSA-G and other groups regarding verb argument structure complexity (Cho-Reyes and Thompson, 2012). When verbs are used in sentences and narratives, there is insufficient evidence to suggest that VAS complexity drives verb selection in PSA-G (Jonkers and Bastiaanse, 1996; Malyutina and den Ouden, 2017). However, extrapolating from Broca's aphasia, it is likely that VAS elaboration is limited in PSA-G (Malyutina and den Ouden, 2017). Finally, PSA with syntactic deficits produce fewer light verbs (Thorne and Faroqi-Shah, 2016). It should be pointed out that there are some empirical confounds in investigations of argument structure complexity in PSA-G: the action pictures used as stimuli may differ across VAS types by imageability, visual complexity, or picture name agreement. Additionally, sentences with complex VAS are frequently longer than sentences with simpler VAS. Thus other co-occurring deficits such as lexical retrieval or scope of incremental planning might limit the production of sentences with complex VAS verbs.
Sentence comprehension
Asyntactic comprehension in PSA-G generated immense interest between the 1980s and 2000s (Grodzinsky, 1984, 1988; Zurif et al., 1993; Caplan et al., 2007). Across a variety of comprehension tasks, the following conclusions can be made from studies that were more empirically robust (e.g., Berndt, 1991; Caplan et al., 2007, 2013; Pregla et al., 2022). Asyntactic comprehension is found across neurotypical speakers and across PSA subtypes (Caplan et al., 2007, 2013; see also Wilson and Saygin, 2004; Pregla et al., 2022). In neurotypical speakers and mild aphasia, the pattern of asyntactic comprehension is trigged by the difficulty of the experimental task (Murray et al., 1997; Caplan et al., 2013). Based on these findings, asyntactic comprehension has been attributed to processing/resource limitations rather than a syntactic deficit in agrammatism (Caplan, 2012; Caplan et al., 2013).
Several studies have examined the ability of PSA-G to judge the grammaticality of sentences across a variety of sentence types (Linebarger et al., 1983; Baum, 1989; Wulfeck et al., 1991; Grodzinsky and Finkel, 1996; Devescovi et al., 1997; Kim and Thompson, 2000; Dickey and Thompson, 2009; Faroqi-Shah and Dickey, 2009; Faroqi-Shah et al., 2020). The pattern that emerges from these studies is that grammaticality judgement of most sentence structures is preserved in PSA-G (e.g., wh-questions, verb argument structure violations), the only consistent exception being tense violations (e.g., Dickey et al., 2008), and a few other long-distance dependencies (Baum, 1989).
Summary of core morphosyntactic characteristics of PSA-G
To conclude this section, the most empirically robust findings in PSA-G are impairments in: producing well-formed sentences, elaborating verbs with their arguments in sentence contexts, and producing and comprehending verb tense morphology. Impairments in closely associated processes, such as knowledge of verb argument structure, verb affixation, and function assignment are not implicated in PSA-G. Finally, asyntactic comprehension is not uniquely associated with PSA-G (e.g., Wilson and Saygin, 2004) and is more likely a generic response to higher processing demands (Caplan, 2012).
Re-envisioning PSA-G in a broader cluster of symptoms
A morphosyntactic profile of PSA-G has emerged from the synthesis of evidence in the previous section. However, this is an incomplete portrayal of PSA-G because agrammatic production is one symptom within the broader clinical profile of post-stroke aphasia resulting from left inferior frontal gyrus (LIFG) lesions. Some of these “non-syntactic” aspects are closely associated with PSA-G, such as a slow speaking rate and symptom variability (e.g., Kok et al., 2007; Gordon and Clough, 2020; Gleichgerrcht et al., 2021), while others are characteristic of aphasia in general, such as phonological and cognitive deficits, and use of compensatory strategies to accommodate linguistic deficits (e.g., Braber et al., 2005; Chapman and Hallowell, 2021). Yet others are the consequence of stroke, such as slowed processing speed, perceptuo-motor impairments, and depression (Assayag et al., 2022; Hazelton et al., 2022; Yoo et al., 2022). Figure 1 illustrates the complexity and dynamics of factors at play in PSA-G. The inner layers reflect linguistic attributes most closely associated with PSA-G and the outer layers represent symptoms found across aphasias and stroke survivors. It is very likely that these multiple strata interact in complex ways that are yet to be understood. Therefore it is important to view PSA-G as an amalgamation of cognitive and linguistic symptoms resulting from LIFG lesions. Progress toward a comprehensive theory of PSA-G can be made by accommodating this interaction between the morphosyntactic profile and other non-syntactic behaviors, to the extent that the latter are relatively consistent in PSA-G.
In this section, I will highlight several findings that are inherent in the clinical picture of PSA-G and are overlooked in current theories of agrammatism. Some of these findings question the assumption that PSA-G is a purely morphosyntactic disorder and argue for an expanded view of PSA-G that incorporates other linguistic deficits. Other findings provide insights into cognitive mechanisms that could be implicated as the underlying source of agrammatic aphasia. Evidence for broadening of the linguistic profile of PSA-G will be drawn from the high co-occurrence of slow speech rate along with motoric and phonological deficits. Indications of likely mechanisms that lead to agrammatic language output will be taken from the multiple and synchronized functions subserved by LIFG, and inter- and intra-individual variability documented in PSA-G.
Broca's aphasia, apraxia of speech, and phonological errors
Broca's aphasia is used as a proxy for PSA-G by numerous researchers (e.g., Patel et al., 2008; Boye and Bastiaanse, 2018). To better understand PSA-G, let us sift through the symptoms that constitute Broca's aphasia. Standardized assessments of aphasia characterize Broca's aphasia with the following multidimensional profile: subjective identification of fragmented utterances produced at a slow rate, impaired ability to repeat, and relatively preserved auditory comprehension (Goodglass et al., 2001; Swinburn et al., 2004; Kertesz, 2006). In the Western Aphasia Battery-Revised (WAB-R, Kertesz, 2006), which is the most commonly used standardized test (Kertesz, 2020), Broca's aphasia is identified with scores for fluency, comprehension, repetition, and naming in the range of <5, 4–10, <7.9 and <9 respectively (on a scale of 0–10). Of particular relevance is the lower repetition score, which could occur due to phonological deficits and/or short-term memory limitations. Indeed, phonological errors are widely reported in Broca's aphasia (Trost and Canter, 1974; Monoi et al., 1983; Niemi et al., 1985) and this is relevant in the context of PSA-G for at least three reasons. First, morphologically complex words, which are often challenging for PSA-G, are confounded by phonological complexity. Thus, substitution and omissions of bound morphemes (e.g., kick or kicking for kicks) could be an artifact of phonological challenges, as demonstrated in several studies of PSA-G (Obler et al., 1999; Braber et al., 2005; Lambon Ralph et al., 2005; Faroqi-Shah et al., 2010). In fact, several lesion studies have demonstrated the proximity of lesions associated with syntactic deficits and phonomotor deficits (Borovsky et al., 2007; Faroqi-Shah et al., 2014; Na et al., 2022). This is illustrated in Figure 2.
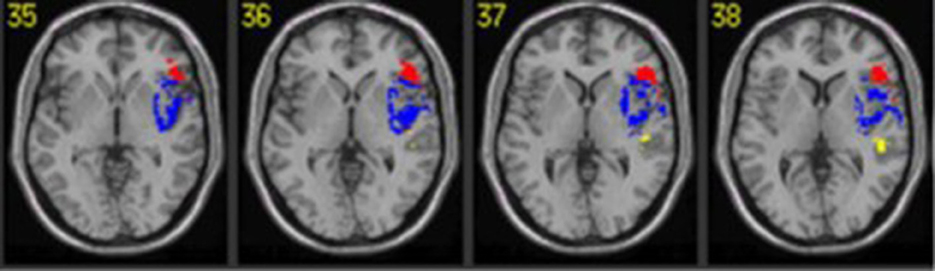
Figure 2. The proximity of lesions for sentence production (red) and phonological (blue) deficits from a voxel wise lesion symptom mapping study of aphasia (Faroqi-Shah et al., 2014). Reprinted with permission from Taylor & Francis Group.
Secondly, errors in repeating multisyllabic words and phoneme distortions are also a hallmark of motor planning difficulty in apraxia of speech, a symptom that co-occurs with and has lesion overlap with Broca's aphasia (Hillis et al., 2004; Richardson et al., 2012; Trupe et al., 2013; Basilakos et al., 2015; Ballard et al., 2016). In fact, AoS is fairly common, occurring in 30% of PSA (Ziegler et al., 2022). Third, den Ouden et al. (2019) reported that PSA-G had more severe apraxia of speech and slower speech rate compared to non-agrammatic PSA. To summarize, PSA-G is one symptom of this broader clinical picture of Broca's aphasia. Importantly, while there is evidence indicating that agrammatic language co-occurs, and is even confounded by, motoric and phonological difficulties, there is no evidence to date indicating that the morphosyntactic deficits of PSA-G occur in isolation without any other linguistic deficits. In the absence of such evidence, it is prudent to question the wisdom of conceptualizing PSA-G as an insular deficit.
Non-fluency
As with the proxy use of Broca's aphasia discussed in the previous section, researchers and clinicians use non-fluent aphasia as a proxy term and often use speech rate to diagnose agrammatism (Gordon and Clough, 2020). The distinction between non-fluency and agrammatic production is important because: (1) a slow rate of speech (non-fluency) could arise from a variety of underlying reasons such as apraxia of speech, difficulty selecting between competing responses (dynamic aphasia), and/or excessive self-monitoring; (2) some fluent aphasias may be on a continuum with Broca's aphasia, and (3) fluency ratings can be unreliable (Gordon, 1998). In the multidimensional classification of aphasia, four subtypes are considered non-fluent (global, Broca's, transcortical motor, and transcortical mixed). Two critical questions are posed here. The first is whether there is a clearly delineated perceptual dichotomy between fluent and non-fluent aphasia given that accurate diagnosis of PSA-G is critical for empirical rigor. The second question is whether we can establish any relationship between PSA-G and slow rate. Evidence for this is evaluated next.
Fluent – non-fluent dichotomy
Despite the ubiquitous use of terms like fluent and non-fluent, there is little published data on their actual rates of speech. In the past few years, two studies analyzed the relationship between rate of speech and other language measures although neither study provides numerical values (Nozari and Faroqi-Shah, 2017; Gordon and Clough, 2020). The data from the 112 unselected PSA in Nozari and Faroqi-Shah (2017) was used to plot the distribution of rate of speech, measured as words per second, in Figure 3. It is noteworthy that the distribution is not bimodal, questioning the dichotomous distinction between fluent and non-fluent. Furthermore, Gordon (2020) reported that most disagreements of aphasia subtyping occurred between Broca's aphasia and two fluent aphasia types: anomic and conduction aphasia (illustrated in Figure 3 in Gordon, 2020). There is considerable overlap in the WAB-R profile scores of Broca's aphasia presented earlier (fluency, comprehension, repetition, and naming: <5, 4–10, <7.9 and <9 respectively) with those for conduction aphasia (>4, 7–10, <6.9 and <10 respectively; Kertesz, 2006). Furthermore, behaviorally, both Broca's and conduction aphasic individuals produce phonological paraphasias with self-corrections and have “functional” comprehension. These two subtypes can thus be viewed on a continuum as they evolve over time with recovery (Pedersen et al., 2004; Flowers et al., 2016). In fact, the overlap and continuity between Broca's and conduction aphasia may explain the similar performance of these two groups in the classic finding of asyntactic comprehension by Caramazza and Zurif (1976). These evidences not only underscore the over-simplification of the non-fluent-fluent dichotomy utilized in agrammatism research (Bates et al., 1991), but also caution against assumptions that a purely syntactic deficit exists in aphasia.
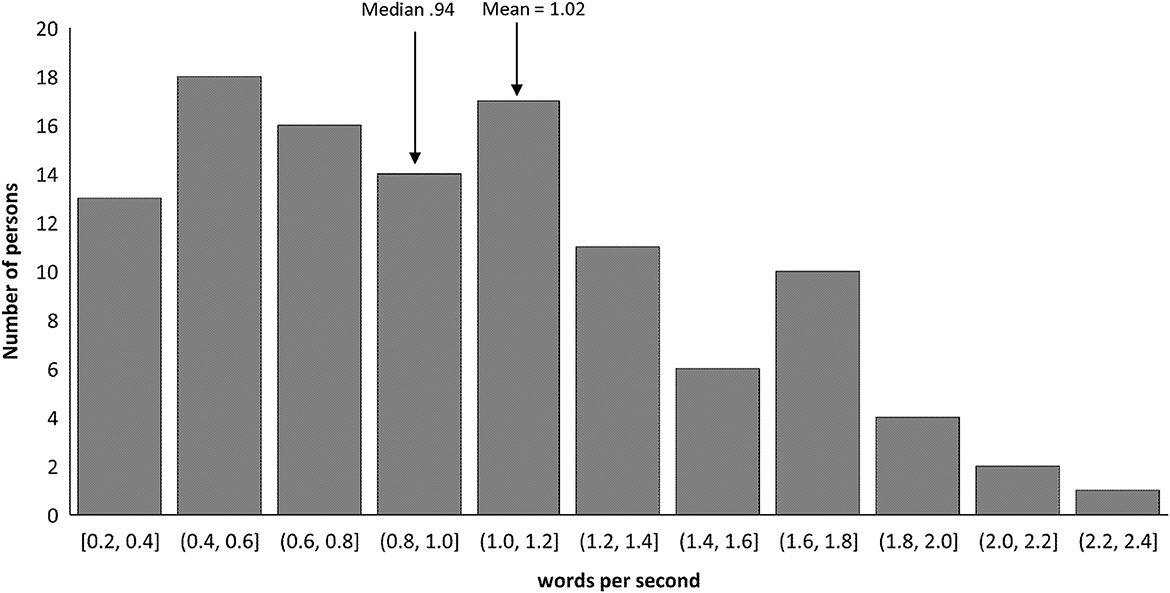
Figure 3. Distribution of rate of speech (words per second) in sample of 112 persons with aphasia (calculated from Nozari and Faroqi-Shah, 2017). The numbers on the x-axis refer to the range of speech rates that are represented in each frequency column.
Relationship between non-fluency and morphosyntactic deficits
A critical question is whether slow speech rate and morphosyntactic deficits are a happenstance co-occurrence or whether there could be a mechanistic relationship between these two symptoms. Here we examine data from studies that examined the relationship between morphosyntactic abilities and some aspect of speaking rate (Nozari and Faroqi-Shah, 2017; Faroqi-Shah et al., 2022a; Salis and DeDe, 2022). In light of the previous critique of the fluent-non-fluent dichotomy, it is noteworthy that all three studies examined morphosyntactic abilities in aphasia as a continuous variable instead of using a categorical diagnosis of PSA-G. Nozari and Faroqi-Shah (2017) examined this question using a path analysis of narrative language samples of 112 persons with PSA (from MacWhinney et al., 2011). Non-fluency (measured as words per second and the WAB-R fluency rating, Kertesz, 2006) was most strongly predicted by morphosyntactic ability (path coefficient = 0.45) and to a smaller extent by lexical abilities, comprehension, and working memory (path coefficients = 0.11 to.13). The results of a new path analysis for morphosyntactic ability as the dependent measure are shown in Figure 4 (using data from Nozari and Faroqi-Shah, 2017). Morphosyntactic ability is represented by the Developmental Sentence Score (DSS, Lee and Canter, 1971). DSS provides a composite measure of an individual's morphosyntactic ability by locating eight types of morphosyntactic elements in the narrative sample and assigning weights to these based on age of acquisition norms (see also Thorne and Faroqi-Shah, 2016). The predictive contribution of speech rate (words/second) was the largest, more so than verb morphology (% past tense), comprehension (WAB-R), and verb retrieval (% verbs) (path coefficients are in Figure 4).

Figure 4. Results of a path analysis (with data from Nozari and Faroqi-Shah, 2017) showing the predictors for morphosyntactic productivity, as measured by the Developmental Syntax Score (Lee and Canter, 1971). For simplicity only the latent variables are shown. Numbers above the arrows represent the path coefficients, *significant predictors.
In another investigation of the relationship between speech fluency and morphosyntactic abilities, we measured the occurrence of disfluencies, such as filled pauses (uh, um, you know, etc.) and silences (Faroqi-Shah et al., 2022a). Disfluencies are interpreted as stalling for time for linguistic planning (Clark and Fox Tree, 2002; Howell, 2007; Salis and DeDe, 2022). Faroqi-Shah et al. (2022a) used relative scores of two continuous language measures, morphosyntactic productivity (DSS, Lee and Canter, 1971), and lexical diversity (Malvern and Richards, 1997), to calculate a difference score (the Standardized Syntax Semantics Difference Score, SSSD). The SSSD was used to categorize participants into those with predominantly morphosyntactic (PSA-G) or lexico-semantic (PSA-LS) deficits. Individuals with predominantly morphosyntactic deficits produced more disfluencies overall (40% disfluencies vs. 29%). Figure 5 shows the proportion of filled pauses as a function of DSS for each group. The stronger association between disfluencies and DSS for the morphosyntactic deficit group indicates that this group is more likely to stall for time when speaking (Figure 5). This suggests an association between non-fluency and morphosyntactic deficit. Pause length is another metric of language fluency. Salis and DeDe (2022) found that longer pauses occurred within longer sentences in the narratives of mildly aphasic and neurotypical speakers. Thus syntactic planning has a time cost and individuals with proficient syntactic competence utilize pauses for syntactic planning. Summarizing across the three studies, the emerging picture is a strong association between morphosyntactic planning and speaking time, as shown in a path analysis (Figure 4), the higher occurrence of disfluencies in people with predominantly morphosyntactic deficits (Faroqi-Shah et al., 2022a) and those who successfully produce longer utterances (Salis and DeDe, 2022). While the latter two findings might appear contradictory, the common theme is that syntactic planning is associated with a high time cost. The implication of timing for a future theory of PSA-G is further analyzed in the Discussion section.
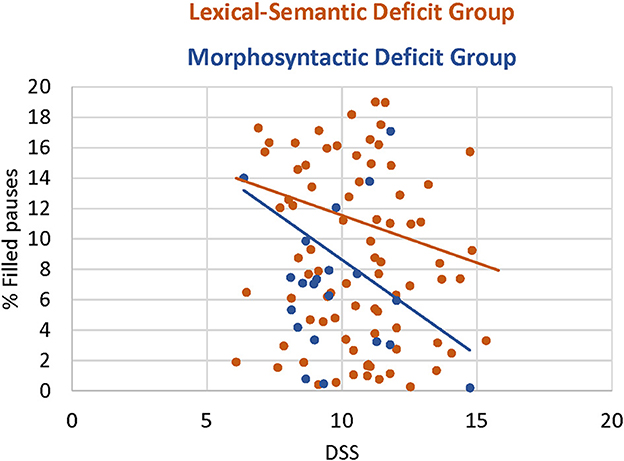
Figure 5. The relationship between filled pauses and deficit group in a narrative language analysis of disfluencies (from Faroqi-Shah et al., 2022a).
Inter- and intra-person variability
Individual variability in morphosyntactic performance is well documented, both within participants due to task demands (Hofstede and Kolk, 1994; Caplan et al., 2007; Kok et al., 2007; Pregla et al., 2022) and across individuals with agrammatic aphasia (Berndt, 1987; Miceli et al., 1989; Rochon et al., 2000; Caramazza et al., 2001; Faroqi-Shah and Thompson, 2004; Drai, 2006). Inter-person variability is unsurprising in light of the complex clinical profile of PSA discussed earlier (Figure 1). It also begs the question of whether PSA-G is a binary clinical condition, or whether agrammatic production lies on a continuum. Arguments in favor of a continuum view are that symptoms such as simplified sentence structure, morphological errors, and impaired comprehension of complex sentences are also found in other aphasia subtypes (Heeschen and Kolk, 1988; Bates et al., 1991; Edwards, 1998; Edwards and Bastiaanse, 1998; Caplan et al., 2007). Several authors have used continuous measures such as the percent of grammatical utterances, mean length of utterance, and DSS to measure morphosyntactic ability (Thompson et al., 2012; Thorne and Faroqi-Shah, 2016). This is not to say that a binary classification of PSA-G is problematic or futile: it is possible that persons who are clinically judged as “agrammatic” are at the extreme end of this continuum. And investigations contrasting groups at the extreme ends will be valuable in delineating the core characteristics of morphosyntactic impairment in aphasia.
Intra-person variability that occurs due to task demands or stimulus properties underscores a fundamental property of morphosyntactic computations: momentary variations in processing load impact the success of the computation. As an illustration of processing load effects on morphosyntactic computation, PSA-G produced more verb inflection errors when required to sequence words into a sentence and inflect the verb compared to just inflecting the verb (Kok et al., 2007; see also Slevc and Martin, 2016). Similarly, passive sentences were more accurately produced and comprehended with passive-bias verbs than with verbs that more commonly occur in active sentences (Gahl, 2002; Menn et al., 2003), and were more accurately produced when passive-morphology was cued (Faroqi-Shah and Thompson, 2003). In light of the inter-and intra-person variability in PSA-G, the logical approach forward is to accommodate the variability into future theoretical accounts of morphosyntactic deficits.
Lesion of the left inferior frontal region
Across studies, agrammatic language production in PSA is unambiguously and consistently associated with large lesions of the left inferior frontal gyrus (LIFG) and underlying white matter connections (Faroqi-Shah et al., 2014; den Ouden et al., 2019; Matchin and Hickok, 2020; Gleichgerrcht et al., 2021). Other lesions, such as those in the left posterior temporal or parietal cortex, have been less consistently implicated for morphosyntactic production deficits (e.g., den Ouden et al., 2019; Gleichgerrcht et al., 2021). The association between LIFG lesions and PSA-G is unsurprising given extensive evidence of the critical role of LIFG for morphosyntactic operations in neurotypical speakers (Embick et al., 2000; Shapiro and Caramazza, 2003; Sahin et al., 2006; Shapiro et al., 2006; Zaccarella et al., 2017). A key point, however, is that the LIFG is not a purely syntactic region, but is involved in several other linguistic operations at various time points during language encoding. This was demonstrated by Sahin et al. (2009) using intracranial recordings as people read or inflected words. LIFG activity occurred sequentially for lexical (~200 milliseconds), morphological (~320 milliseconds), and phonological (~450 milliseconds) processing. Simply put, the LIFG is the end-stage hub for the highly coordinated encoding of lexical, morphosyntactic, and phonological representations for different elements in the sentence (Sahin et al., 2009; see also Zhu et al., 2022). Therefore LIFG lesions could not only affect morphosyntactic computations, but also other linguistic functions that could be critical for sentence production and directly contribute to the manifestation of PSA-G. Correspondingly, LIFG lesions have been implicated for deficits in phonological encoding (Borovsky et al., 2007; Indefrey, 2011; Faroqi-Shah et al., 2014; Flinker et al., 2015; Na et al., 2022), motor planning (Basilakos et al., 2018; Papitto et al., 2020), and word selection (Robinson et al., 1998; Swick et al., 2008; Schnur et al., 2009; Novick et al., 2010; Python et al., 2018).
In fact, the LIFG is an anatomically and functionally heterogeneous region (Amunts et al., 1999; Tettamanti and Weniger, 2006; Clos et al., 2013; Fedorenko and Blank, 2020; Asano et al., 2022) that has been implicated not only for the linguistic encoding mentioned earlier, but also for high level cognitive functions such as selection, sequencing, and inhibition (Fadiga et al., 2009; Schnur et al., 2009; Kunert et al., 2015; Maffei et al., 2020; Kemmerer, 2022). Several authors have argued that the LIFG is a domain- general (or supramodal) high level processing region (Tettamanti and Weniger, 2006; Clos et al., 2013; Fedorenko and Blank, 2020). It is also part of the multiple demand network that helps modulate brain activity when there are high processing load demands (Duncan, 2010). Given LIFG's role in multiple linguistic and cognitive functions, it is important to consider the cumulative impact of the LIFG lesion in PSA-G. For instance, it is likely that individuals with PSA-G have reduced overall processing capacity resulting from their LIFG lesions (Tettamanti and Weniger, 2006; Fedorenko and Blank, 2020). Thus they are unable to handle the high and time-constrained processing demands of sentence production as effectively as other PSA who do not have LIFG lesions. A future theory that accommodates the cumulative impact of LIFG damage on the processing demands of sentence production will better reflect the functional reality of this region and will be a step closer to a mechanistic explanation of PSA-G.
Summary of PSA-G as a broader symptom cluster
To summarize, this section presented evidence of the co-occurrence of non-syntactic deficits in individuals with PSA-G, particularly impairments in phonological encoding, motor planning, and speech fluency. Moreover, LIFG lesions, which are the primary etiology of PSA-G, have also been implicated for these non-syntactic impairments. Based on this evidence, it is prudent to view PSA-G not as an isolated syntactic deficit but as a symptom complex that includes phonomotor difficulties. This blending of morphosyntactic and phonomotor deficits is not an entirely novel idea, and has been proposed earlier [Pick, 1913; Kean, 1977 (cited in Akhutina, 2016); Kolk and Heeschen, 1992]. In light of the evidence presented in this section, there is currently no strong rationale to pursue theoretical accounts of an isolated syntactic deficit in aphasia. To move toward a theoretical account for the broader PSA-G symptom complex, this section drew attention to the widely reported phenomenon of symptom variability in PSA-G. Inter-individual variability may stem from the different extents of syntactic and phonomotor deficits across individuals. Intra-individual (task-based) variability may reflect the (in)ability of the LIFG-lesioned language network to handle the momentary processing and integration demands of the sentence being formulated. The next section evaluates current theories of PSA-G in the context of the broader PSA-G symptom cluster presented here and the core symptoms of PSA-G identified in the preceding section.
Extant theories of PSA-G
Given the non-specificity of asyntactic comprehension across aphasia subtypes (Caramazza and Zurif, 1976; Wilson and Saygin, 2004; Caplan et al., 2007), this section will focus on accounts of agrammatic production. Numerous theories of agrammatism have been proposed in the past four decades. Most extant models view PSA-G as a purely morphosyntactic disorder. Most models also focus on one aspect of the agrammatic symptom cluster such as asyntactic comprehension (Zurif, 1998; Salis and Edwards, 2008), verb complexity (Thompson, 2003; Barde et al., 2006), or verb tense morphology (Friedmann and Grodzinsky, 1997; Faroqi-Shah and Thompson, 2007; Fyndanis et al., 2012). A sample of these theories is given in Table 1.
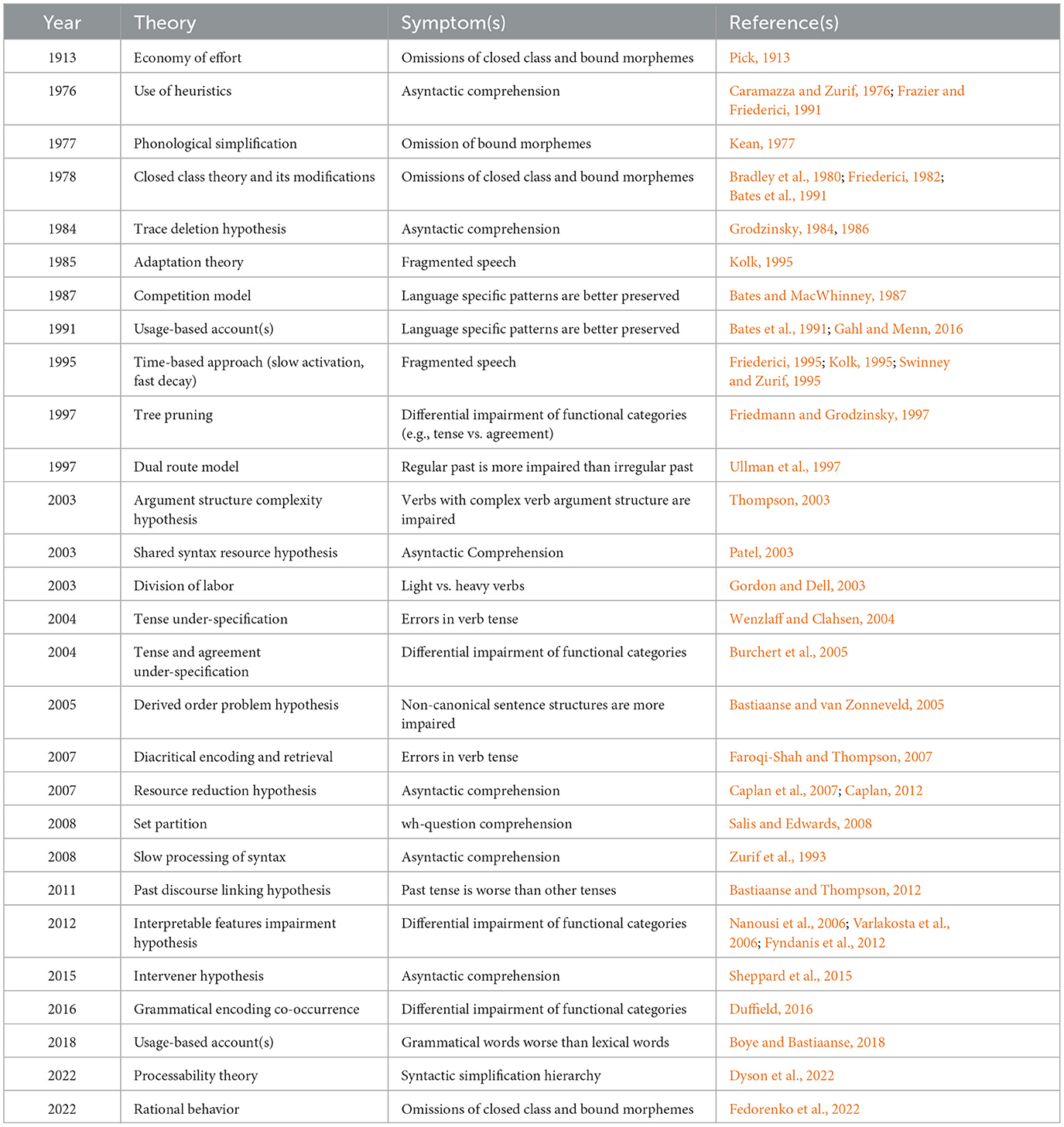
Table 1. An illustrative selection of theories of agrammatism in post-stroke aphasia, presented in chronological order.
Although a detailed discussion of these theories is beyond the scope of the present paper, a few key observations are highlighted. Extant theories fall into two broad genres: linguistic and mechanistic. Studies of PSA-G have been dominated by the application of linguistic theories to specific symptoms (e.g., Grodzinsky, 1984; Friedmann and Grodzinsky, 1997; Boye and Bastiaanse, 2018). These investigations align a linguistic theory with a specific symptom dissociation in PSA-G. In a recent example of this approach, Boye and Bastiaanse (2018) and Boye et al. (2023) used the contrast between grammatical and lexical functions of different words to show that the former word class is deficient in PSA-G (per the ProGram theory, Harder and Boye, 2011). However, PSA-G's dissociation in these two word classes has been documented for decades and formalized in prior theories such as the closed-class theory of agrammatism (Bradley et al., 1980; Biassou et al., 1997). Another example of a linguistic account, which prompted a large body of cross-linguistic investigations of PSA-G, focused on hierarchies of functional categories in the syntactic tree structure (Friedmann and Grodzinsky, 1997). Such linguistically-oriented studies have yielded detailed symptom descriptions of PSA-G (e.g., Wenzlaff and Clahsen, 2004; Burchert et al., 2005; Duman et al., 2007). Besides their focus on single symptoms, linguistically descriptive accounts have done little to advance current understanding of the underlying neurocognitive source of PSA-G. That is, these accounts do not explain why the symptom occurs in PSA-G. Further, most of these accounts lack linking data from neurotypical speakers that attests to the psychological reality of the identified linguistic computation.
Mechanistic theories, in contrast, attempt to explain the observed symptoms using one or more cognitive process(es) or strategies. In general, these theories implicate a unitary cognitive mechanism (or deficit) to explain a cluster of PSA-G symptoms. Some examples of these accounts include Pick's (1913) Economy of Effort, Bates and MacWhinney's (1987) Competition Model, Kolk's (1995) Time-based approach, Ullman's (2001) Declarative Procedural Hypothesis, and Gordon and Dell's (2003) Division of Labor. Unlike linguistic theories, few mechanistic theories adopt a purely syntactic view of agrammatism, thus more easily accommodating the multi-faced clinical picture of PSA-G. The general limitations of this genre of theories include insufficient delineation of how (or why) PSA-G differ from non-agrammatic PSA, and the limited efforts to empirically validate their predictions. Of course, the linguistic-mechanistic distinction of theories is not entirely binary, and some theories incorporate elements of both. For example, the Trace-Deletion Hypothesis for asyntactic comprehension suggests that persons with PSA-G have lost movement-traces and compensate for the absence of linguistic computation by applying heuristic strategies (Grodzinsky, 1984).
Summary of PSA-G theories and considerations for a future theory
The numerous theories of PSA-G that have been proposed (Table 1) broadly fall under a descriptive or a mechanistic label. While the former genre of theories tend to focus on a single symptom, the predictions of the latter genre have not been sufficiently tested. As yet, no theory of agrammatism explains the complete picture of PSA-G findings that has emerged over the past few decades. Of particular relevance for a comprehensive theory are the following observations. First, the theory must accommodate the core morphosyntactic symptoms that are uniquely associated with PSA-G: fragmented sentences, difficulty with functional morphology, especially tense marking, and elaboration of verb argument structure. Second, the theory must accommodate PSA-G's preserved abilities for some syntactic computations such as structural priming, incremental planning, and sensitivity to some syntactic violations. Third, the co-occurrence of non-syntactic deficits such as phonomotor deficits and non-fluency, needs to be accommodated. Conversely, there needs to be strong rationale for proposing an exclusive syntactic deficit. Fourth, symptom variability needs to be accommodated. While inter-individual variability can be easily explained by differences in lesion extent or co-morbidities, within-individual variability from task demands is tricky to explain. Further, a well-founded theory should not only account for the core symptoms of PSA-G but also delineate the mechanism underlying the paradox of other PSA (e.g., Wernicke's and conduction aphasia) who are able to formulate sentences with relatively better sentence structure at a fluent speaking rate. Indeed, there is a critical gap in the current mechanistic understanding of how lexical, grammatical, motoric, and cognitive processes work together to enable fluent sentence production in neurotypical adults and how this breaks down in PSA-G. In summary, there are numerous compelling reasons to re-envision PSA-G within a theoretical framework that accommodates and integrates several findings that are unaddressed by current theories. Other authors have also recently revisited the theoretical framework of PSA-G (Dyson et al., 2022; Fedorenko et al., 2022).
Discussion
The goal of this paper is to develop a theoretical account of neurocognitive mechanism(s) underlying PSA-G that integrates the range of empirical findings and extends our understanding of the condition. The previous sections identified several reasons that have impeded advances in the understanding of PSA-G. A major factor that has stymied progress is the rigor and reproducibility of the empirical evidence, which is weakened by inconsistencies in patient characterization and failure to meet the minimum inferential assumptions of neuropsychological research (Caramazza, 1984; Bezeau and Graves, 2001; Martin, 2006; Gaeta and Brydges, 2020). The second issue is the narrow focus on morphosyntax, when in fact, morphosyntactic deficits do not occur in a vacuum. These are but one of a cluster of co-occurring symptoms in PSA-G, particularly phonomotor deficits and non-fluency (Kean, 1977; Goodglass and Kaplan, 1983; Blumstein, 2000). Further justification for broadening the view of PSA-G beyond a purely syntactic deficit comes from the multifunctional nature of the LIFG, whose lesions are the most consistent etiology of PSA-G. Third, a majority of current PSA-G theories are descriptive, in which linguistic theory is mapped onto any one PSA-G symptom, often lacking an actual explanation of why the symptom occurs. While there are some mechanistic accounts of PSA-G (e.g., Kolk, 1995), there isn't yet a sufficient body of empirical evidence to validate these accounts.
Gaps in the current understanding of neurocognitive mechanisms underlying PSA-G can be bridged by drawing from psycholinguistic findings of how neurotypical speakers formulate sentences. Mechanisms of neurotypical sentence production can be used as a framework within which to compare aphasic performance. This approach was taken by Thompson and colleagues in a series of studies comparing real-time encoding of sentences across agrammatic and neurotypical speakers using eye-tracking methods (Lee and Thompson, 2011b; Mack et al., 2013; Lee et al., 2015; Cho-Reyes et al., 2016). These studies are a valuable first step in uncovering specific aspects of sentence planning, such as incremental encoding of verb arguments. In the ideal world, we would have comparisons of real time performance across speakers who are neurotypical, PSA without agrammatism, and PSA-G. Comparisons across the first two groups would delineate the general impact of aphasia (including word retrieval difficulties), while comparisons across the latter two groups would pinpoint why some aphasic speakers formulate fairly well-formed sentences while PSA-G do not. In the next section, pertinent findings of constituent assembly in neurotypical speakers are presented so that these findings can be integrated into a theory of PSA-G. This is an alternate approach to the linguistic theory approach that is so prevalent in PSA-G research. Neurotypical findings of constituent assembly will be used as a backdrop to present the Synergistic Processing Bottleneck model. The rationale and key assumptions of this model are presented, followed by unanswered questions that await further research.
Integrating psycholinguistic findings to inform a theory of agrammatism
A common approach in testing theories of PSA-G is to compare their accuracy in simple constrained tasks (e.g., sentence completion) with neurotypical speakers whose performance is close to ceiling. In addition to the inferential weakness of this single dissociation approach, the near perfect accuracy of neurotypical speakers misses a key linking element: how do neurotypical speakers operate during sentence production, and how do those neurotypical phenomena inform mechanisms underlying PSA-G. As a way to move forward, this section will overview four key findings pertaining to sentence planning in neurotypical speakers that could inform a theory of PSA-G. The findings were selected to align with the PSA-G symptoms discussed earlier.
Following a classic language production model (Bock and Levelt, 1994), we use the term constituent assembly to refer to syntactic computations that combine two linguistic units, resulting in words (e.g., [stem]+[affix]), phrases, or hierarchical syntactic structures. Some of these computations fulfill an element of the speaker's message (e.g., tense) while others fulfill language-specific well-formedness constraints (e.g., subject-verb agreement for gender or number).
Constituent assembly proceeds incrementally
Several studies have shown that speakers do not plan an entire sentence before they speak. Rather they plan utterances incrementally such that earlier occurring lexical nodes (or “syntactic treelets”) are planned and proceed on to articulatory planning before the next lexical node is planned (Griffin and Bock, 2000; Ferreira and Swets, 2002; Timmermans et al., 2012). This means that, when producing a subject-verb-object type of sentence (e.g., The boy ate a sandwich), the verb phrase is being syntactically planned simultaneously as the subject noun phrase is in some stage of phonological-articulatory planning. This has implications for PSA-G's reduced processing capacity, as was demonstrated by Lee and Thompson (2011b). They examined eye-fixations on adjuncts (e.g., picture of restaurant in the target sentence, The boy ate a sandwich at the restaurant). While neurotypical adults' eye-fixations on the adjunct occurred as they were speaking earlier parts of the sentence, PSA-G's fixations occurred before sentence onset. PSA-G's looks on the adjuncts prior to initiating the sentence show their difficulty in simultaneously planning and speaking, which is required for incremental sentence planning. It is also possible that PSA-G's difficulties with incremental planning are reflected in their slow speech rate and high proportion of disfluencies relative to other PSA (Nozari and Faroqi-Shah, 2017; Faroqi-Shah et al., 2022b).
Constituent assembly hinges on verb retrieval
Constituent assembly proceeds only after obligatory lexical elements are accessed. Evidence comes from the dependence of speech onset times on when the verb (Antón-Méndez, 2020) and its internal arguments (Momma et al., 2016, 2018) become available. For example, Antón-Méndez (2020) manipulated when each picture of a person-action-thing scenario was presented (e.g., pictures of baby, eating, and egg for the target sentence The baby is eating an egg). Sentence initiation times aligned with the presentation of the action picture although speakers had the opportunity to plan the subject phrase incrementally before retrieving the action. Next, a verb (or other lexical node) first needs to be retrieved before its grammatical morphemes can be planned. Evidence for this comes from longer speaking times for phonologically matched grammatical (e.g., is in The bird is flying) vs. lexical verbs (e.g., is in The bird is black) (Lange et al., 2017). In both instances, the lexeme is occurs as the third word in the sentence, but takes longer to articulate as a grammatical element because its planning hinges on the main verb flying. These psycholinguistic realities explain why verb retrieval difficulties are associated with impoverished sentences in PSA (Berndt et al., 1997a; Thorne and Faroqi-Shah, 2016). The crucial role of verb retrieval in PSA is also evident in the path analysis in Figure 4 where verb retrieval, but not noun retrieval predicted sentence production.
Constituent assembly is computationally demanding
There is evidence for the large computational demands and scope of planning of constituent assembly (Allum and Wheeldon, 2007). This is indicated by several findings. First, speakers take longer or make more errors in utterance planning when there is a high processing load or under processing capacity limitations (Ferreira and Swets, 2002; Sikora et al., 2016; Slevc and Martin, 2016; Fyndanis et al., 2018). The fact that neurotypical and PSA speakers take advantage of lexical and syntactic accessibility in structural priming paradigms further points to the computationally intense nature of constituent planning (Faroqi-Shah and Thompson, 2003; Lee et al., 2015; Cho-Reyes et al., 2016). Next, neurotypical speakers as well as those with aphasia show a trade-off between syntactic complexity and lexical-semantic richness in sentence production (Thorne and Faroqi-Shah, 2016; Rezaii et al., 2022). For example, in utterances with low frequency words, neurotypical speakers use high frequency syntactic frames (and vice versa), showing that the computational demands of sentence planning necessitate a balance between syntactic and lexical load (Rezaii et al., 2022). PSA with syntactic deficits produce more semantically specific words while those with lexical-semantic deficits produce semantically lighter words (Thorne and Faroqi-Shah, 2016). Similarly, speakers with a variety of diagnoses show a trade-off between syntactic complexity (or sentence length) and phonological and motor complexity (Silverman and Ratner, 1997; Obler et al., 1999; Marshall and van der Lely, 2006; Walsh and Smith, 2011). These trade-offs indicate that constituent assembly is computationally demanding and is compromised when other linguistic processes require computational resources. For PSA-G in particular, the frequent co-occurrence of phonological/phonetic difficulties (Blumstein, 2000) and apraxia of speech (Trupe et al., 2013) in Broca's aphasia likely diminishes computational resources that are available for constituent assembly.
The LIFG is a core neural hub for constituent assembly
There is a rigorous body of neuroimaging research examining the spatial and temporal correlates of constituent assembly, including the production of words, phrases, and inflectional morphemes (Indefrey et al., 2001; Shapiro et al., 2006; Sahin et al., 2009; Roos and Piai, 2020; Hauptman et al., 2022). These studies have revealed that constituent assembly for production engages a left hemisphere network, with the posterior LIFG and posterior parts of the left superior and middle temporal gyri (LpSTG-MTG) as the syntactic hubs of this network (Matchin and Hickok, 2020). While the LpSTG-MTG region is more consistently associated with verb argument structure (Thompson et al., 2010; Malyutina and den Ouden, 2017), across these production studies, the LIFG is shown to be specifically involved in linear assembly of linguistic elements. As alluded to in earlier sections of this paper, this role of LIFG in constituent assembly is relevant not only because it is the most consistently lesioned region in PSA-G (den Ouden et al., 2019), but is also the end-stage hub for the highly coordinated encoding of lexical, morphosyntactic, and phonological representations for different elements in the sentence (Sahin et al., 2009; see also Zhu et al., 2022).
In summary, several lines of evidence indicate that constituent assembly is a computationally demanding process that proceeds incrementally, hinges on verb retrieval, and engages morphological elaboration only after selection of the lexical head. Production trade-offs between syntactic and lexical complexity indicate that fluent sentence production depends on a precisely timed, synergistic coordination between morphosyntactic, lexical (verb), and phonomotor processes. The LIFG is a critical end-stage hub for this integration between constituent assembly and phonomotor encoding.
The Synergistic Processing Bottleneck model
With the aim of moving the field forward, this section outlines a multicomponent mechanistic model of PSA-G. This model integrates the core morphosyntactic deficit of PSA-G with two non-syntactic components (which admittedly are not mutually exclusive): (a) other linguistic processes inherent in sentence planning (e.g., lexical/phonomotor) and (b) processing capacity. This model is adapted from classic psycholinguistic models of language production in neurotypical speakers (e.g., Bock and Levelt, 1994; Slevc, 2023). Like classic models, this model emphasizes that language production requires a synergistic coordination between content (lexical processes) and structure (morphosyntactic processes). The difference between this model and classic models of language production is that it identifies constituent assembly as the locus of a processing bottleneck for sentence planning (for all speakers). This is illustrated in Figure 6A. The central tenet of the Synergistic Processing Bottleneck (SPB) Model is that sentence production in PSA-G is undermined by a combination of delays in three linguistic processes (constituent assembly, verb activation, phonomotor planning) and processing limitations that cumulatively create a bottleneck at the point of articulatory planning. This is illustrated in Figure 6B. Key terms in the SPB model are defined in Table 2. In light of the core requirements of a future theory of PSA-G listed earlier, the SPB model differs from previous theories (Table 1) in several ways. First, it integrates empirical findings of constituent assembly in neurotypical speakers with PSA-G symptoms to identify a mechanism that can explain the relative impairment and sparing of morphosyntactic abilities in PSA-G. Second, rather than an exclusive syntactic deficit, it proposes a broader view of PSA-G that allows for the co-occurrence of non-syntactic symptoms such as non-fluency. Third, it identifies delays in the timecourse of syntactic and phonomotor processes and processing capacity limitations as the key mechanisms underlying PSA-G. Fourth, it explains symptom variability, particularly intra-person variability in PSA-G, as an interaction between processing capacity and processing load. Yet another difference between prior theories and SPB is the latter's inclusion of a neuroanatomical locus (LIFG). Finally, it provides a mechanism for differentiating the symptoms of PSA-G from non-agrammatic PSA. In the following paragraphs, the rationale for the model will be postulated. Next, PSA-G and non-agrammatic PSA will be contrasted to explain why sentence production deficits are predominant in the former, and testable predictions of the model will be outlined. I will end with implications for future research.
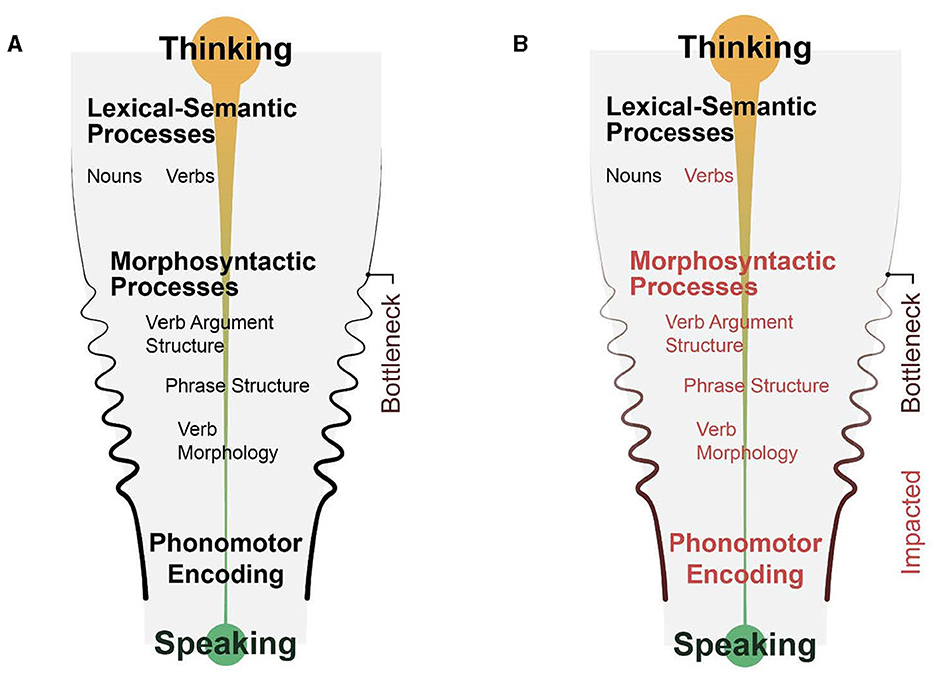
Figure 6. The Synergistic Processing Bottleneck Model of Agrammatism, showing sentence production in (A) neurotypical speakers, and (B) persons with agrammatic aphasia. Morphosyntactic computations (or constituent assembly) are a potential bottleneck due to the high processing load. Computations that are vulnerable in PSA-G due to LIFG lesion are noted in red font. These vulnerabilities exaggerate the processing bottleneck and result in slowly uttered, agrammatic speech output.
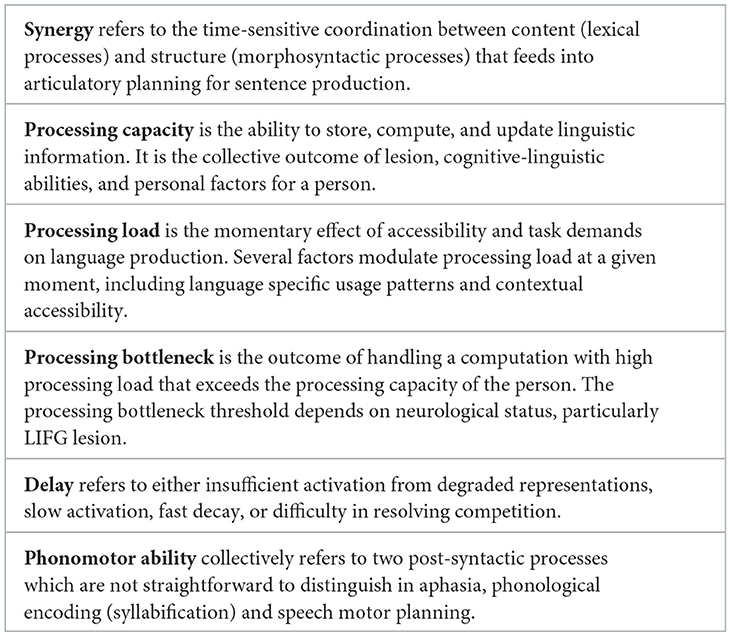
Table 2. Definitions of key terms in the Synergistic Processing Bottleneck model.
Comorbidity of LIFG functions provides a mechanism for the full symptom cluster
The starting point for SPB is the juxtaposition of three LIFG functions that are compromised in PSA-G due to LIFG lesions: constituent assembly, phonomotor planning, and the endstage hub for the highly coordinated encoding of lexical, morphosyntactic, and phonological representations for different elements in the sentence (Sahin et al., 2009; see also Zhu et al., 2022). As for the major lexical categories (nouns and verbs), there is evidence that verb processing selectively engages the LIFG while both nouns and verbs engage left posterior temporal regions (Shapiro et al., 2006; Faroqi-Shah et al., 2018). This explains the IFG's role in integrating linguistic and cognitive representations is also noted in other recent models (Roger et al., 2022). PSA-G is proposed to result from the cumulative effect of LIFG lesions on these three functions. While there is evidence indicating that agrammatic language co-occurs, and is even confounded by, phonomotor difficulties, there is no evidence to date indicating that the morphosyntactic deficits of PSA-G occur in isolation without any other linguistic deficits. In the absence of such evidence, it is better not to view PSA-G as an insular morphosyntactic deficit (see also Fedorenko et al., 2022). With this neuroanatomical backdrop, I will next focus on the cognitive-behavioral mechanisms underlying PSA-G.
Delays and cumulative bottleneck
We speak at an incredibly fast pace of about 150–200 words per minute (Picheny et al., 1986). To achieve this, constituent assembly requires precise synchrony between lexical retrieval and morphosyntactic processes. The encoded sentence constituents are incrementally dispatched for motor planning. The importance of timing for sentence production was first proposed by Kolk (1995). Based on the notion that syntactic trees are built incrementally for sentence production, Kolk (1995) highlighted that synchronized timing of lexical and morphological elements is essential for assembling syntactic trees. Delays in activation of lexical and/or morphological elements could result in agrammatic sentence production. In the SPB model, delay refers to either insufficient activation from degraded representations (Grober, 1984), slow activation (Zurif et al., 1993; Burkhardt et al., 2008), fast decay (Faroqi-Shah et al., 2010), or difficulty in resolving competition (Novick et al., 2010; Mailend et al., 2019) among lexical and/or morphological elements. Although we expect general processing speed delays in all PSA (Faroqi-Shah and Gehman, 2021; Yoo et al., 2022), there is evidence that individuals with PSA-G are particularly slow in real-time activation of lexical and syntactic representations (Freiderici and Kilborn, 1989; Prather et al., 1992, 1997; Zurif et al., 1993; Blumstein and Milberg, 2000; Burkhardt et al., 2008; Love et al., 2008; Ferrill et al., 2012). For example, in a series of studies that compared the timecourse of priming effects in PSA-G and Wernicke's aphasia, Zurif et al. (1993) found slowed activation of lexical and syntactic elements in PSA-G. In contrast, individuals with Wernicke's aphasia demonstrated a normal timecourse of lexical and syntactic activation (Prather et al., 1992, 1997; Zurif et al., 1993). A few authors have implicated a downstream effect of slowed lexical activation on syntactic structure building impairment in PSA-G (e.g., Love et al., 2008; Ferrill et al., 2012) and a few others have identified constituent assembly (Merge) as the locus of slowed activation (e.g., Burkhardt et al., 2008). Another source of activation delays is co-morbid apraxia of speech, which is attributed to delays in resolving competition between motor plans and syllable planning (Haley and Jacks, 2019; Mailend et al., 2019, 2021). The cumulative result of these different delays is an articulatory bottleneck due to which fragmented utterances are spoken slowly (Figure 6B). This is supported by the strong association between speaking rate and syntactic productivity in aphasia (Nozari and Faroqi-Shah, 2017; Faroqi-Shah et al., 2022b; Salis and DeDe, 2022).
Additionally, it is possible that the delays themselves are a circular issue where lexical, morphosyntactic, or motoric representations can decay from the short-term memory buffer before they are integrated. For example, if there is a delay in verb retrieval, the already activated verb's argument(s) may decay before the verb can be integrated with the arguments. Consistent with this, persons with Broca's aphasia use a more restricted variety of verb argument structures than other PSA (Malyutina and den Ouden, 2017). Using eye-tracking methods, PSA-G were also noted to have delayed activation of VAS (Mack et al., 2013). Likewise, there could be delays or decay of referential aspects of a speaker's message (often conveyed by pronouns, functional morphemes, or clausal structures) before constituent assembly is complete. This could account for the pervasive difficulty in expressing and comprehending verb tense morphology in PSA-G (Faroqi-Shah and Dickey, 2009).
The cumulative nature of the articulatory bottleneck implies that articulatory planning of successfully retrieved lexical elements (e.g., nouns) will be uninterrupted in PSA-G. Evidence for this comes from the finding that phonological primes did not facilitate noun picture naming but facilitated verb picture naming in a group of PSA-G who showed a selective verb deficit (Lee and Thompson, 2011a). Noun naming speed was not boosted by phonological primes because nouns did not encounter planning bottlenecks.
Processing trade-offs
A comprehensive theory of PSA-G should be able to account for the well-observed inter and intra person variability in agrammatic symptoms. Individuals with PSA vary in their processing capacity (Murray, 2012; Ivanova and Hallowell, 2014; Faroqi-Shah et al., 2022a). In this paper, processing capacity is defined as the ability to store, compute, and update linguistic information. It is the collective outcome of lesion, cognitive-linguistic abilities, and personal factors for any given person (Figure 1). Thus, inter-person variability could be explained by individual differences in processing capacity. Evidence suggests that those with LIFG lesions are particularly vulnerable to processing capacity limitations for language computations (Novick et al., 2010; Robinson et al., 2010; Slevc and Martin, 2016; Stampacchia et al., 2018). For example, Robinson et al. (2010) demonstrated that sentence generation became increasingly difficult for individuals with LIFG lesions as the number of conceptual propositions was increased. This relationship was not found in individuals whose lesions spared the LIFG, showing that LIFG lesions limit processing capacity. Comparable reductions in processing capacity following LIFG lesions have been reported for both linguistic and non-linguistic computations (see also Tettamanti and Weniger, 2006; Stampacchia et al., 2018).
Processing capacity can be viewed as a static ability which interacts with dynamic modulations in processing load for the speaking task at hand. This paper defines processing load as the momentary effect of accessibility and task demands. Several factors modulate processing load at a given moment, including language specific usage patterns and contextual accessibility (Menn and Obler, 1990; Bates et al., 1991; McRae et al., 1997; Gahl and Menn, 2016; Lee, 2020; Goldberg and Ferreira, 2022). Thus for example, planning a sentence with a typical verb-argument (e.g., The policeman arrested the thief ) requires fewer processing resources than with a less typical argument (e.g., The policeman arrested the teacher), which in turn would demand fewer resources than a non-canonical sentence structure (e.g., The teacher was arrested by the policeman). These load effects, however, may be reversed if the context favors the less typical argument, or a non-canonical sentence frame, or with task demands. These momentary variations in processing load will affect the speed with which computations can be completed, which in turn, can exacerbate or alleviate the articulatory bottleneck. The likelihood of a processing bottleneck specifically in PSA-G is supported by the finding that non-fluent PSA's production of well-formed sentences is more accurate and faster when using more frequent subject nouns compared to less frequent subject nouns (see also Robinson et al., 2010; Speer and Wilshire, 2013). In contrast, fluent PSA do not show a facilitation of sentence structure based on subject noun frequency. Thus, task-related intra-person variability in sentence production is captured by processing load effects. Overuse of canonical word order, frozen phrases, and frequent verb forms (Bates et al., 1987; Faroqi-Shah and Thompson, 2004; Ishkhanyan et al., 2017) could be a strategy to deal with processing limitations.
Several authors have proposed processing accounts for PSA, including those for comprehension (Caplan et al., 2007; Burkhardt et al., 2008; Avrutin and Baauw, 2013), production (Kolk, 1995; Dyson et al., 2022), and overall symptoms (Hula and McNeil, 2008). The SPB model proposes that when the aggregate of a person's processing capacity and processing load during sentence production falls below a threshold, it results in agrammatic language production. The computational demands of constituent assembly trip up the sentence production machinery in PSA-G. The SPB framework thus accommodates inter- and intra-individual variability in sentence production. The SPB model differs from previous processing capacity accounts of PSA-G (Kolk, 1995; Kok et al., 2007; Caplan et al., 2013) by identifying specific syntactic and non-syntactic vulnerabilities, incorporating LIFG symptoms, including a timing component, and embracing the heterogeneity inherent in PSA-G.
Key differences between PSA-G and PSA-LS
The language sample below lucidly conveys that fluent sentences can be produced when speakers experience lexical failures. It was spoken at a rate of 140 words/minute (i.e., within normal limits) by a person with a clinical profile of Wernicke's aphasia (per the WAB-R, Kertesz, 2006). She is describing the picnic picture scene that is part of the WAB-R. The transcriber's notes are in square brackets.
“Okay. That will be, um, see here weeding, whiting, reading [weeding, whiting = target approximations to reading]. The cat [=dog]…is eating here, back packing [=picnicking?] and he's speaking at a ball [=target not sure, no ball in the picture]. He's got a book he's reading here, and they've got a fly [=kite] up there and I can see they're really reading.”
In the SPB framework, PSA with lexico-semantic deficits (PSA-LS) are expected to show lexical retrieval and phonological delays and difficulties, particularly for nouns. However, their preserved LIFG, their relatively spared morphosyntactic and motor abilities, and adequate linguistic processing capacity allow them to avoid an end-stage processing bottleneck in sentence production. This results in fluent sentence production that is mostly grammatically well-formed although the sentences may contain lexical errors and paragrammatisms. Self-monitoring appears to be a vulnerability that is unique to severe PSA-LS (Sampson and Faroqi-Shah, 2011).
Summary and predictions of the SPB
The SPB model is a mechanistic model of sentence production that uses the neurotypical language production framework (Figure 6A) to identify key vulnerabilities in PSA-G (Figure 6B). The SPB model accounts for the broad symptom cluster of PSA-G in the following way. LIFG lesions impair constituent assembly, verb activation, phonomotor planning, and processing capacity. In PSA-G, constituent assembly and phonomotor encoding unfold over a delayed timecourse compared to persons without LIFG lesions. The reduced processing capacity particularly impacts constituent assembly, which is a computationally demanding process that proceeds incrementally. As a result, sentences may be fragmented, and verb argument structure may be incomplete. Neurotypical findings of constituent assembly explain why verb retrieval impairments in aphasia affect sentence formulation (Antón-Méndez, 2020) and the realization of grammatical morphemes that need a lexical head (Lange et al., 2017; Boye and Bastiaanse, 2018). Grammatical morphemes that convey referential aspects of a speaker's message (e.g., verb tense; Bastiaanse and Thompson, 2012; Fyndanis et al., 2012) are additionally vulnerable from the slow timecourse of sentence planning. The interplay between reduced processing capacity resulting from LIFG lesions and processing load demands of the sentence being produced explains a variety of phenomena reported in PSA-G. First, it explains PSA-G's relatively preserved performance for some morphosyntactic computations such as incremental sentence planning, subject-verb agreement, function assignment, and grammaticality judgement (e.g., Clahsen and Ali, 2009; Lee et al., 2015). Secondly, it provides a mechanism for performance variability based on task demands or stimulus manipulations, such as structural priming (Faroqi-Shah and Thompson, 2003; Cho-Reyes et al., 2016). Third, the overuse of frequent structural and morphological elements occurs due to the lower processing demands of accessible sentence structures (Bates et al., 1991; Centeno et al., 1996; Ishkhanyan et al., 2017). Fourth, processing capacity limitations explain trade-offs between linguistic features such as between morphological and phonological complexity (Obler et al., 1999; Faroqi-Shah et al., 2010) and syntactic and semantic complexity of verbs (Thorne and Faroqi-Shah, 2016). This view of morphosyntactic planning on a processing continuum also accommodates the continuous nature of morphosyntactic ability in aphasia and reports of simplified sentence structure in other aphasia subtypes (Saffran et al., 1989; Edwards, 1998).
The SPB model provides an empirically testable framework for future investigations of PSA-G. The key predictions are outlined here, beginning with three mechanistic predictions and followed by three expectations pertaining to the pattern of linguistic deficits.
First, given the LIFG's role as the end-stage hub for integrating the incrementally unfolding lexical, syntactic, and phonological representations, the SPB model underscores the observation that there is no evidence (as yet) of a complete and isolated morphosyntactic deficit that impacts all aspects of morphosyntax with the sparing of other linguistic process (e.g., lexical-semantic). In other words, persons with PSA-G (from LIFG lesions) will also have at least some level of phonomotor and lexical deficits, the latter will be particularly evident for verbs (e.g., Miceli et al., 1988).
Second, relative to PSA-LS, PSA-G will show a delayed timecourse for morphosyntactic, verb, and phonomotor planning (Prather et al., 1992, 1997; Burkhardt et al., 2008; Mack et al., 2013). The delayed timecourse will be evident in online paradigms such as priming with different stimulus-onset asynchronies, eye-tracking, and neurophysiological responses using electroencephalography and magnetoencephalography.
Third, while we expect all PSA to show a reduced processing capacity relative to neurotypical speakers (Hula and McNeil, 2008; Caplan, 2012), processing capacity reductions will be steeper in PSA-G relative to PSA-LS, and will interact with processing load manipulations (e.g., Ivanova and Hallowell, 2014). Further, compared to other PSA, PSA-G will show larger benefits (in planning constituents, verbs, and phonomotor details) from manipulations that over-rule processing load, such as priming and usage frequency (Lee and Thompson, 2011a; Speer and Wilshire, 2013; Cho-Reyes et al., 2016; Boye et al., 2023).
The next three predictions are portrayed in the results of the path analysis in Figure 4. An “articulatory bottleneck” will be evident in the speaking rate and disfluencies of PSA-G. The relationship between non-fluency and constituent assembly will be stronger in PSA-G compared to PSA-LS, as shown in Figure 5 (Faroqi-Shah et al., 2022b). It should be noted that the non-fluency-constituent assembly connection does not preclude agrammatic output from showing up in the written modality. This is because processing capacity limitations and activation delays will still impact constituent assembly in writing tasks.
The fifth prediction is a challenge with activating verb representations and fulfilling VAS in PSA-G, which will be evident in online paradigms (Faroqi-Shah et al., 2010; Mack et al., 2013) as well as narrative language (Malyutina and den Ouden, 2017; see also Figure 4). Similar to the prediction for non-fluency, the relationship between verb activation and constituent assembly will be stronger for PSA-G compared to PSA-LS.
Finally, the vulnerability of grammatical morphology will interact with the verb deficit and show processing load effects. When a verb is a lexical head for a grammatical morpheme (as in tense marking on verbs in English), any delays or degradation of verb activation will have a downstream effect on the retrieval and planning of the associated grammatical morpheme(s). Processing load effects in the production of grammatical morphology will include trade-offs with phonological and syntactic complexity and usage frequency effects (Obler et al., 1999; Faroqi-Shah and Thompson, 2004).
The totality of the SPB model markedly differs from prior PSA-G theories (Table 1). As stated earlier, there is some intersection with other theories. These overlapping elements between SPB and a few other theories are contrasted in Table 3.
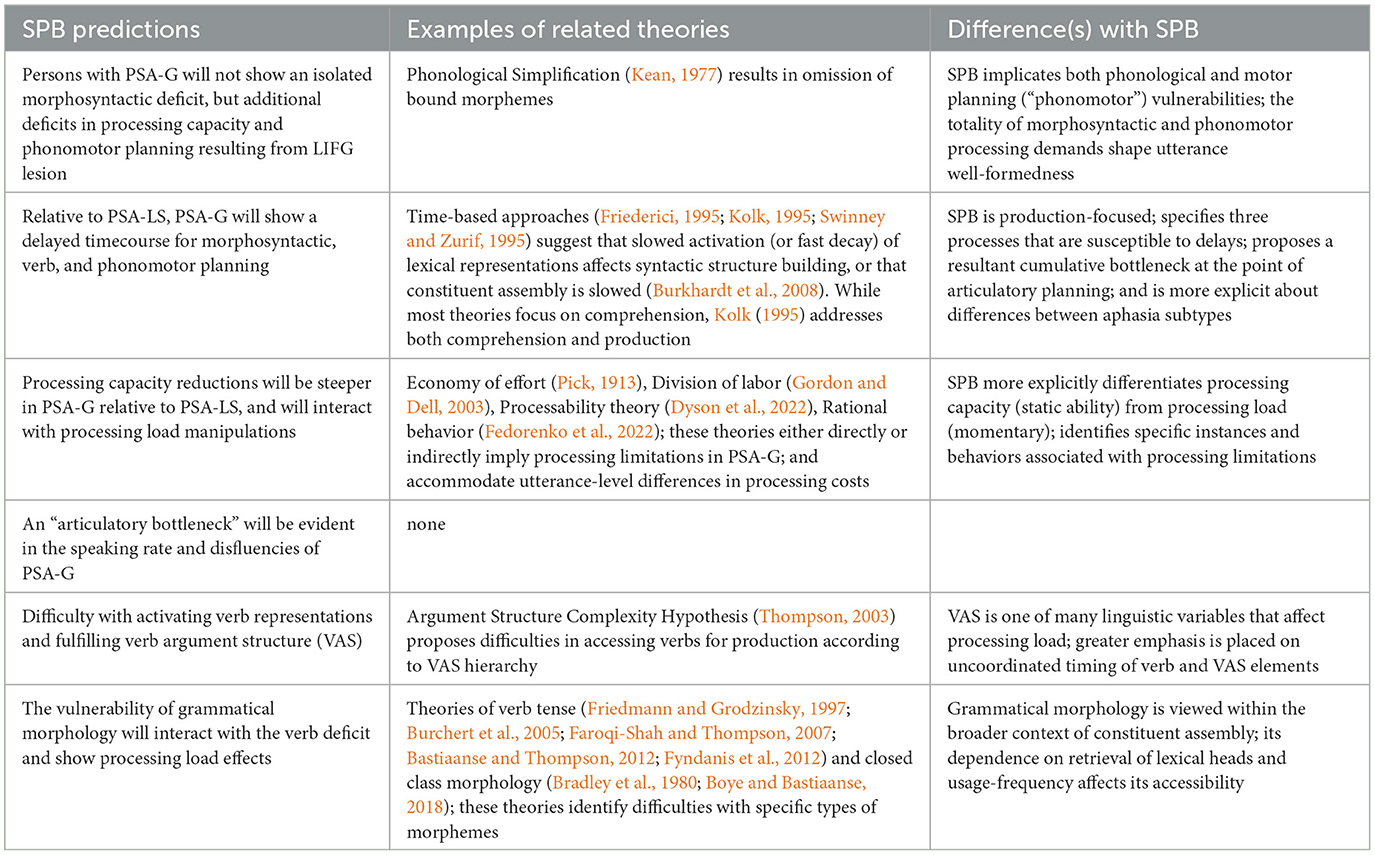
Table 3. The relationship between extant theories of agrammatism and the Synergistic Processing Bottleneck (SPB) model.
Implications for future research
The SPB model was developed from an integration of empirical findings of neurotypical and agrammatic sentence production. Thus, it is only as good as the data it was derived from. Some details of the SPB model are yet to be developed. As detailed earlier, characterization of PSA-G is somewhat murky not only from issues related to empirical rigor, but also from a disproportionate focus on syntactic theory at the cost of uncovering underlying mechanisms. To move forward, it is crucial to test the predictions of the SPB model outlined in the previous section as well as to fill in gaps where findings are inconsistent or insufficient. Here I highlight some unresolved questions and provide suggestions for the conduct of PSA-G research.
Unresolved questions
Much is unknown about the neurocognitive dynamics of sentence production in neurotypical speakers. Improved future understanding of the neurotypical mechanisms underlying sentence production can be used to further refine the SPB model. Empirical evidence in support of slow timing in PSA-G primarily comes from comprehension or lexical priming tasks (Freiderici and Kilborn, 1989; Prather et al., 1992; Burkhardt et al., 2008; Love et al., 2008). This needs to be tested and validated for speech production. Recent studies linking speech timing with morphosyntactic production in aphasia (Nozari and Faroqi-Shah, 2017; Faroqi-Shah et al., 2022b; Salis and DeDe, 2022) are the first step forward in empirically showing a connection between syntactic planning and time cost. While processing capacity limitations are demonstrated in individuals with LIFG damage, we lack an understanding of the aphasia deficit profile of these individuals. Future research can verify and complete the triangular relationship between delayed activation, processing limitations and PSA-G. Given the central role of verbs in sentence construction, we need to resolve some of the inconsistencies in verb deficits in PSA-G (Matzig et al., 2009), particularly in explicating their time course of activation and VAS elaboration. The SPB model does not address how LIFG damage affects the role of the temporal lobe syntactic hub (Fedorenko et al., 2018; Matchin and Hickok, 2020) and the functional connectivity between these two hubs. We currently lack exact knowledge of which factors are mediators, confounds, and moderators of language production in PSA-G, thus limiting our understanding of underlying mechanisms and individual differences. In sum, we hope that researchers can use the SPB model to spur future research that moves away from the syntax-centric view of PSA-G.
Guidelines for experimental rigor
Methodological rigor and replicability are the foundations of science. The issues of empirical rigor pertaining to PSA-G research were noted in an earlier section and were based on the key shortcomings noted in Faroqi-Shah (2020) scoping review of agrammatism research. It is not clear if some of the same empirical issues exist (or do not exist) in research on other topics in aphasiology. The following recommendations focus on participant description and addressing methodological confounds in PSA-G research that were noted by Faroqi-Shah (2020) and may (or may not) be applicable to aphasiology in general. These guidelines do not cover statistical treatment of data (e.g., Bezeau and Graves, 2001).
1. Etiology. First and foremost, aphasia is a health condition that arises from a medical etiology. Details about the etiology (e.g., type, number, location of strokes), time-post onset, and specifically, diagnosis of aphasia should be provided for participants in empirical studies.
Diagnosis of PSA-G. An accurate diagnosis of agrammatism allows for replication and cross-language comparisons. It is important to operationally define and characterize the agrammatic impairment in study participants for internal validity as well as generalizability of findings. While physicians may diagnose aphasia, the characterization of speech-language behavior lies mainly within the scope of practice of speech-language pathologists (American Speech-Language-Hearing Association, 2016). In countries where the SLP profession does not exist, researchers could operationally define their understanding of PSA-G for their language (see Menn and Obler, 1990) and provide supplementary qualitative details, such as language samples. Researchers should document the diagnostic process, including the standardized assessments administered by a licensed SLP, and criteria used to determine the presence (vs. absence) of PSA-G. Researchers may also consider using the core outcome dataset for aphasia recommended by the ROMA consensus (Research Outcome Measurement in Aphasia, Wallace et al., 2019).
2. Language Sample Analysis (LSA). Diagnosis of PSA-G should be supported by a LSA, which refers to quantitative measures derived from narrative language. LSA is more ecologically valid than constrained sentence tasks, it has been conventionally used to document language characteristics in clinical populations (MacWhinney et al., 2011), and is particularly useful in documenting the core morphosyntactic features of PSA-G. In PSA-G research, narratives have been typically elicited from descriptions of picture scenarios such as the “Cookie theft picture” (Goodglass et al., 2001), re-telling of the “Cinderella” (or “Red Riding Hood”) story, and elicitation of a personal narrative such as their “stroke story” (Edwards, 1998; Rochon et al., 2000; MacWhinney et al., 2011; Hsu and Thompson, 2018). A minimum of a 150-word language sample has been recommended for LSA (Saffran et al., 1989). While some studies have provided comparisons of quantitative measures between PSA-G and neurotypical adults (Rochon et al., 2000; Hsu and Thompson, 2018), studies that provide agrammatic vs. non-agrammatic aphasic comparisons are particularly helpful in diagnosing PSA-G (Saffran et al., 1989; Faroqi-Shah, 2020; see also Mack et al., 2021 for primary progressive aphasia). Faroqi-Shah (2020) examined the diagnostic accuracy of various measures derived from the tools available at Talkbank (https://aphasia.talkbank.org/, MacWhinney et al., 2011), identified five core measures that differentiated PSA-G from non-agrammatic aphasia with 89% classification accuracy, and provided cut-off scores. Given the intensive time commitment for LSA, at minimum, perceptual ratings of narrative language could be provided. Casilio et al. (2019) developed a 27-item perceptual rating scale for narrative language in aphasia and identified four items in this scale clustered together and marked agrammatic behavior.
As noted at several instances in this paper, it is worth considering morphosyntactic production impairment in PSA on a continuum rather than a categorical diagnosis, especially depending on the research questions. Given the limitations in sentence production imposed by lexical deficits (e.g., Berndt et al., 1997a; Faroqi-Shah and Thompson, 2003; Speer and Wilshire, 2013), it is important for a continuous measure of morphosyntactic ability to consider the confound of lexical abilities. Thorne and Faroqi-Shah (2016) addressed this by calculating a difference score (the Standardized Syntax Semantics Difference Score, SSSD) based on morphosyntactic and lexical productivity. Participants were then categorized into those with predominantly morphosyntactic (PSA-G) or lexico-semantic (PSA-LS) deficits (see also Faroqi-Shah et al., 2022a).
3. Comparison groups. The rigor of characterizing PSA-G will be enhanced by striving for double dissociations by including an aphasic non-agrammatic comparison group. As noted earlier, it is crucial to provide details of the linguistic and clinical characteristics of this comparison group, including procedures for matching groups.
4. Mediating, moderating, and confounding variables. It is important to document non-linguistic mediating and moderating variables, particularly, an oro-motor examination for co-morbid speech conditions (Ziegler et al., 2022). It is also important to provide a cognitive profile with aphasia-friendly assessments that address the verbal limitations of the participant (e.g., Ivanova and Hallowell, 2014; Faroqi-Shah et al., 2022b). Sensory screenings should be conducted and reported, depending on the experimental demands (hearing, vision, color vision, field cuts). Motor abilities, particularly, with reference to hand-use if any keyboard, writing, or gesturing response is used. Authors should document language moderators such as bilingual status, literacy/education, and word retrieval scores.
5. Experimental accommodations for aphasia. Details of experimental manipulations that are unique to aphasia must be provided. For example, did any participants have hemiparesis that might have affected their keyboard responses (if used), which hand was used for responding, were verbal instructions supplemented by written instructions, how was comprehension of task instructions determined, what was the protocol for practice items, etc.
In summary, research methods that consider the multidimensional nature of aphasia will yield a more accurate picture of PSA-G and help advance the field.
Conclusion
Following a critical examination of current evidence on PSA-G, this paper questions the assumption of the existence of an insular morphosyntactic deficit in PSA. Instead, this paper proposes a broader view of PSA-G as a cluster of morphosyntactic and phonomotor deficits arising from LIFG damage. The LIFG is the hub for synergistically coordinating message content with structure. Structure building (constituent assembly) is computationally demanding and can stall fluent speech output (articulatory bottleneck), especially in persons with limited processing capacity. The SPB model attempts to provide a comprehensive account of PSA-G and can be fine-tuned with future evidence.
Data availability statement
The original contributions presented in the study are included in the article/supplementary material, further inquiries can be directed to the corresponding author.
Author contributions
The author confirms being the sole contributor of this work and has approved it for publication.
Funding
This study was funded by NIH grant 1R21 DC018916-01 to YF-S.
Conflict of interest
The author declares that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Publisher's note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
Footnotes
1. ^This study is frequently cited as evidence of a selective past tense impairment by Bastiaanse and colleagues (e.g., Bastiaanse and Thompson, 2012; Bastiaanse, 2013; Boye et al., 2023) although there was no significant difference across tenses (cf. Figure 2 in Faroqi-Shah and Dickey, 2009).
References
Adamson, J., Beswick, A., and Ebrahim, S. (2004). Is stroke the most common cause of disability? J. Stroke Cerebrovasc. Dis. 13, 171–177. doi: 10.1016/j.jstrokecerebrovasdis.2004.06.003
Akhutina, T. (2016). Luria's classification of aphasias and its theoretical basis. Aphasiology 30, 878–897. doi: 10.1080/02687038.2015.1070950
Allum, P. H., and Wheeldon, L. (2007). Planning scope in spoken sentence production: The role of grammatical units. J. Exp. Psychol.: Learn. Memory Cogn. 33, 791–810.
American Speech-Language-Hearing Association (2016). Scope of Practice in Speech-Language Pathology (Scope of Practice). doi: 10.1044/policy.SP2016-00343
Amunts, K., Schleicher, A., Burgel, U., Mohlberg, H., Uylings, H. B., and Zilles, K. (1999). Broca's region revisited: cytoarchitecture and intersubject variability. J. Compar. Neurol. 412, 319–341. doi: 10.1002/(SICI)1096-9861(19990920)412:2<319::AID-CNE10>3.0.CO;2-7
Antón-Méndez, I. (2020). The role of verbs in sentence production. Front. Psychol. 11, 189. doi: 10.3389/fpsyg.2020.00189
Arévalo, A., Perani, D., Cappa, S. F., Butler, A., Bates, E., and Dronkers, N. (2007). Action and object processing in aphasia: From nouns and verbs to the effect of manipulability. Brain Lang. 100, 79–94. doi: 10.1016/j.bandl.2006.06.012
Asano, R., Boeckx, C., and Fujita, K. (2022). Moving beyond domain-specific vs. domain-general options in cognitive neuroscience. Cortex 154, 259–268. doi: 10.1016/j.cortex.2022.05.004
Assayag, E. B., Tene, O., Korczyn, A. D., Solomon, Z., Bornstein, N. M., Shenhar-Tsarfaty, S., et al. (2022). Posttraumatic stress symptoms after stroke: the effects of anatomy and coping style. Stroke, 53, 1924–1933. doi: 10.1161/STROKEAHA.121.036635
Avrutin, S. (2000). “Comprehension of discourse-linked and non-discourse-linked questions by children and Broca's aphasics,” in Language and the Brain, eds. Y. Grodzinsky, L. Shapiro, and D. Swinney (San Diego: Academic Press) 295–315. doi: 10.1016/B978-012304260-6/50017-7
Avrutin, S., and Baauw, S. (2013). “Processing View on Agrammatism,” in Diagnosing Syntax, eds. L. L.-S. Cheng and N. Corver (Oxford University Press). doi: 10.1093/acprof:oso/9780199602490.003.0019
Ballard, K. J., Azizi, L., Duffy, J. R., McNeil, M. R., Halaki, M., O'Dwyer, N., et al. (2016). A predictive model for diagnosing stroke-related apraxia of speech. Neuropsychologia 81, 129–139. doi: 10.1016/j.neuropsychologia.2015.12.010
Barde, L. H. F., Schwartz, M. F., and Boronat, C. B. (2006). Semantic weight and verb retrieval in aphasia. Brain Lang. 97, 266–278. doi: 10.1016/j.bandl.2005.11.002
Basilakos, A., Rorden, C., Bonilha, L., Moser, D., and Fridriksson, J. (2015). Patterns of poststroke brain damage that predict speech production errors in apraxia of speech and aphasia dissociate. Stroke 46, 1561–1566. doi: 10.1161/STROKEAHA.115.009211
Basilakos, A., Smith, K. G., Fillmore, P., Fridriksson, J., and Fedorenko, E. (2018). Functional characterization of the human speech articulation network. Cereb. Cortex. 28, 1816–1830. doi: 10.1093/cercor/bhx100
Bastiaanse, R. (2013). Why reference to the past is difficult for agrammatic speakers. Clin. Linguist. Phonetics.. doi: 10.3109/02699206.2012.751626. [Epub ahead of print].
Bastiaanse, R., and Edwards, S. (2004). Word order and finiteness in Dutch and English Broca's and Wernicke's aphasia. Brain Lang. 89, 91–107. doi: 10.1016/S0093-934X(03)00306-7
Bastiaanse, R., and Jonkers, R. (1998). Verb retrieval in action naming and spontaneous speech in agrammatic and anomic aphasia. Aphasiology 12, 951–969. doi: 10.1080/02687039808249463
Bastiaanse, R., and van Zonneveld, R. (1998). On the relation between verb inflection and verb position in Dutch agrammatic aphasics. Brain Lang. 64, 165–181. doi: 10.1006/brln.1998.1972
Bastiaanse, R., and van Zonneveld, R. (2005). Sentence production with verbs of alternating transitivity in agrammatic Broca's aphasia. J. Neuroling. 18, 57–66. doi: 10.1016/j.jneuroling.2004.11.006
Bates, E., Friederici, A., and Wulfeck, B. (1987). Grammatical morphology in aphasia: evidence from three languages. Cortex 23, 545–574. doi: 10.1016/S0010-9452(87)80049-7
Bates, E., and MacWhinney, B. (1987). “Competition, variation, and language learning,” in Mechanisms of Language Acquisition, ed B. MacWhinney (Erlbaum), 157–193.
Bates, E., Wulfeck, B., and MacWhinney, B. (1991). Cross-linguistic research is aphasia: An overview. Brain Lang. 41, 123–148. doi: 10.1016/0093-934X(91)90149-U
Baum, S. R. (1989). On-line sensitivity to local and long-distance syntactic dependencies in Broca's Aphasia. Brain Lang. 37, 327. doi: 10.1016/0093-934X(89)90021-7
Berndt, R. S. (1987). “Symptom co-occurence and dissociation in the interpretation of agrammatism,” in The cognitive neuropsychology of language, eds. M. Coltheart, G. Sartori, and R. Job (Hillsdale, New Jersey: Erlbaum) 221–232.
Berndt, R. S. (1991). “Sentence processing in aphasia,” in Acquired Aphasia, ed. M. T. Sarno (New York: Academic Press) 223–269.
Berndt, R. S., and Caramazza, A. (1981). “Syntactic aspects of aphasia,” in Acquired Aphasia, Vol. 1, eds M. T. Sarno (Academic Press), 175.
Berndt, R. S., Haendiges, A. N., Mitchum, C. C., and Sandson, J. (1997a). Verb retrieval in aphasia. 2. Relationship to sentence processing. Brain Lang. 56, 107–137. doi: 10.1006/brln.1997.1728
Berndt, R. S., Mitchum, C. C., Haendiges, A. N., and Sandson, J. (1997b). Verb retrieval in aphasia. 1. Characterizing single word impairments. Brain Lang. 56, 68–106. doi: 10.1006/brln.1997.1727
Bezeau, S., and Graves, R. (2001). Statistical power and effect sizes of clinical neuropsychology research. J. Clin. Exp. Neuropsychol. 23, 399–406. doi: 10.1076/jcen.23.3.399.1181
Biassou, N., Obler, L. K., Nespoulous, J. L., Dordain, M., and Harris, K. S. (1997). Dual processing of open- and closed-class words. Brain Lang. 57, 360–373. doi: 10.1006/brln.1997.1749
Bird, H., Howard, D., and Franklin, S. (2000). Why is a verb like an inanimate object? Grammatical category and semantic category deficits. Brain Lang. 72, 246–309. doi: 10.1006/brln.2000.2292
Bird, H., Ralph, M. A. L., Seidenberg, M. S., McClelland, J. L., and Patterson, K. (2003). Deficits in phonology and past-tense morphology: What's the connection? J. Memory Lang. 48, 502–526. doi: 10.1016/S0749-596X(02)00538-7
Blackwell, A., and Bates, E. (1995). Inducing Agrammatic Profiles in Normals: Evidence for the selective vulnerablity of morphology under cognitive resource limitation. J. Cogn. Neurosci. 7, 228–257. doi: 10.1162/jocn.1995.7.2.228
Blumstein, S., and Milberg, W. P. (2000). “Language deficits in Broca's and Wernicke's aphasia: A singular impairment,” in Langage and the Brain: Representation and Processing, eds. Y. Grodzinsky, L. Shapiro, and D. Swinney (San Diego: Academic Press).
Blumstein, S. E. (2000). “Deficits of speech production and speech perception in aphasia,” in Handbook of Neuropsychology, eds. R., Berndt (The Netherlands: Elsevier Science).
Bock, J. K., and Loebell, H. (1990). Framing sentences. Cognition 35, 1–39. doi: 10.1016/0010-0277(90)90035-I
Bock, K., and Levelt, W. (1994). “Language production: grammatical encoding,” in Handbook of Psycholinguistics, ed. G. M. A (San Diego: Academic Press).
Borovsky, A., Saygin, A. P., Bates, E., and Dronkers, N. (2007). Lesion correlates of conversational speech production deficits. Neuropsychologia 45, 2525–2533. doi: 10.1016/j.neuropsychologia.2007.03.023
Boye, K., and Bastiaanse, R. (2018). Grammatical vs. Lexical Words in Theory and Aphasia: Integrating Linguistics and Neurolinguistics. Glossa J. General Lingu. 3, 1–18. doi: 10.5334/gjgl.436
Boye, K., Bastiaanse, R., Harder, P., and Martínez-Ferreiro, S. (2023). Agrammatism in a usage-based theory of grammatical status: Impaired combinatorics, compensatory prioritization, or both? J. Neuroling. 65, 101108. doi: 10.1016/j.jneuroling.2022.101108
Braber, N., Patterson, K., Ellis, K., and Lambon Ralph, M. A. (2005). The relationship between phonological and morphological deficits in Broca's aphasia: Further evidence from errors in verb inflection. Brain Lang. 92, 278–287. doi: 10.1016/j.bandl.2004.05.012
Bradley, D. C., Garrett, M. F., and Zurif, E. (1980). “Closed class theory of agrammatism,” in Biological Studies of Mental Processes, ed. D. Caplan (London: MIT Press).
Brady, M. C., Kelly, H., Godwin, J., Enderby, P., and Campbell, P. (2016). Speech and language therapy for aphasia following stroke. Cochr. Datab. System. Rev. 2016, CD000425. doi: 10.1002/14651858.CD000425.pub4
Burchert, F., Swoboda-Moll, M., and De Bleser, R. (2005). Tense and Agreement dissociations in German agrammatic speakers: Underspecification vs. hierarchy. Brain Lang. 94, 188–199. doi: 10.1016/j.bandl.2004.12.006
Burkhardt, P., Avrutin, S., Piñango, M. M., and Ruigendijk, E. (2008). Slower-than-normal syntactic processing in agrammatic Broca's aphasia: Evidence from Dutch. J. Neuroling. 21, 120–137. doi: 10.1016/j.jneuroling.2006.10.004
Caley, S., Whitworth, A., and Claessen, M. (2017). Can we separate verbs from their argument structure? A group study in aphasia. Int. J. Commun. Disor. 52, 59–70. doi: 10.1111/1460-6984.12253
Canu, E., Bessi, V., Calderaro, D., Simoni, D., Castelnovo, V., Leocadi, M., et al. (2020). Early functional MRI changes in a prodromal semantic variant of primary progressive aphasia: a longitudinal case report. J. Neurol. 267, 3100–3104. doi: 10.1007/s00415-020-10053-9
Caplan, D. (1995). Issues arising in contemporary studies of disorders of syntactic processing in sentence comprehension in agrammatic patients. Brain Lang. 50, 325–338. doi: 10.1006/brln.1995.1051
Caplan, D. (2001). The measurement of chance performance in aphasia, with specific reference to the comprehension of semantically reversible passive sentences: A note on issues raised by Caramazza, Capitani, Rey, and Berndt (2001) and Drai, Grodzinsky, and Zurif (2001). Brain Lang. 76, 193–201. doi: 10.1006/brln.2000.2354
Caplan, D. (2012). “Resource reduction accounts of syntactically based comprehension disorders,” in Perspectives on agrammatism, eds. R. Bastiaanse, and C. K. Thompson (Hove: Psychology Press) 48–62. doi: 10.4324/9780203120378-9
Caplan, D., Michaud, J., and Hufford, R. (2013). Short-term memory, working memory, and syntactic comprehension in aphasia. Cogn. Neuropsychol. 30, 77–109. doi: 10.1080/02643294.2013.803958
Caplan, D., Waters, G., DeDe, G., Michaud, J., and Reddy, A. (2007). A study of syntactic processing in aphasia I: Behavioral (psycholinguistic) aspects. Brain Lang. 101, 103–150. doi: 10.1016/j.bandl.2006.06.225
Caramazza, A. (1984). The logic of neuropsychological research and the problem of patient classification in aphasia. Brain Lang. 21, 9–20. doi: 10.1016/0093-934X(84)90032-4
Caramazza, A., Capitani, E., Rey, A., and Berndt, R. S. (2001). Agrammatic Broca's aphasia is not associated with a single pattern of comprehension performance. Brain Lang. 76, 158–184. doi: 10.1006/brln.1999.2275
Caramazza, A., and Zurif, E. (1976). Dissociation of algorithmic and heuristic processes in language comprehension: evidence from aphasia. Brain Lang. 3, 572–582. doi: 10.1016/0093-934X(76)90048-1
Casilio, M., Rising, K., Beeson, P., Bunton, K., and Wilson, S. M. (2019). Auditory-perceptual rating of connected speech in aphasia. Am. J. Speech-Language Pathol. 28, 550–568. doi: 10.1044/2018_AJSLP-18-0192
Centeno, J. (2012). “Morphosyntactic features in the spoken language of Spanish-English bilinguals with aphasia,” in Aspects of Multilingual Aphasia, eds. M. R. Gitterman, M. Goral, and L. K. Obler (Clevedon, UK: Multilingual Matters) 207–223. doi: 10.21832/9781847697554-015
Centeno, J., Obler, L. K., Cairns, H., Garro, L., and Merrifield, P. (1996). Frequency effects on verb inflection use by agrammatic Spanish-speaking subjects. Brain Lang. 57, 47–50.
Centeno, J. G. (2007). Canonical features in the inflectional morphology of Spanish-speaking individuals with agrammatic speech. Adv. Speech-Lang. Pathol. 9, 162–172. doi: 10.1080/14417040601006913
Chapman, L. R., and Hallowell, B. (2021). The unfolding of cognitive effort during sentence processing: pupillometric evidence from people with and without Aphasia. J. Speech, Langu. Hear. Res. 64, 4900–4917. doi: 10.1044/2021_JSLHR-21-00129
Cherney, L. R., Halper, A. S., Holland, A. L., and Cole, R. (2008). Computerized Script Training for Aphasia: Preliminary Results. Am. J. Speech-Lang. Pathol. 17, 19–34. doi: 10.1044/1058-0360(2008/003)
Cho-Reyes, S., Mack, J. E., and Thompson, C. K. (2016). Grammatical encoding and learning in agrammatic aphasia: Evidence from structural priming. J. Mem. Lang. 91, 202–218. doi: 10.1016/j.jml.2016.02.004
Cho-Reyes, S., and Thompson, C. K. (2012). Verb and sentence production and comprehension: Northwestern Assessment of Verbs and Sentences (NAVS). Aphasiology 26, 1250–1277. doi: 10.1080/02687038.2012.693584
Clahsen, H., and Ali, M. (2009). Formal features in aphasia: Tense, agreement, and mood in English agrammatism. J. Neuroling. 22, 436–450. doi: 10.1016/j.jneuroling.2009.02.003
Clark, H. H., and Fox Tree, J. E. (2002). Using uh and um in spontaneous speaking. Cognition 84, 73–111. doi: 10.1016/S0010-0277(02)00017-3
Clos, M., Amunts, K., Laird, A. R., Fox, P. T., and Eickhoff, S. B. (2013). Tackling the multifunctional nature of Broca's region meta-analytically: Co-activation-based parcellation of area 44. NeuroImage 83, 174–188. doi: 10.1016/j.neuroimage.2013.06.041
Crawford, J. R., Garthwaite, P. H., and Gray, C. (2003). Wanted: fully operational definitions of dissociations in single case studies. Cortex 39, 357–370. doi: 10.1016/S0010-9452(08)70117-5
Cumming, T. B., Marshall, R. S., and Lazar, R. M. (2013). Stroke, cognitive deficits, and rehabilitation: still an incomplete picture. Int. J. Stroke 8, 38–45. doi: 10.1111/j.1747-4949.2012.00972.x
De Villiers, J. (1974). Quantative aspects of agrammatism in aphasia. Cortex 10, 36–54. doi: 10.1016/S0010-9452(74)80037-7
den Ouden, D. B., Malyutina, S., Basilakos, A., Bonilha, L., Gleichgerrcht, E., Yourganov, G., et al. (2019). Cortical and structural-connectivity damage correlated with impaired syntactic processing in aphasia. Hum. Brain Mapp. 40, 2153–2173. doi: 10.1002/hbm.24514
Devescovi, A., Bates, E., D'Amico, S., Hernandez, A., Marangolo, P., Pizzamiglio, L., et al. (1997). Grammaticality judgements in patients with and without expressive agrammatism. Aphasiology 11, 543–579. doi: 10.1080/02687039708248490
Dickey, M. W., Milman, L. H., and Thompson, C. K. (2008). Judgment of functional morphology in agrammatic aphasia. J. Neuroling. 21, 35–65. doi: 10.1016/j.jneuroling.2007.08.001
Dickey, M. W., and Thompson, C. K. (2009). Automatic processing of wh- and NP-movement in agrammatic aphasia: Evidence from eyetracking. J. Neurolinguist. 22, 563–583. doi: 10.1016/j.jneuroling.2009.06.004
Dragoy, O., and Bastiaanse, R. (2010). Verb production and word order in Russian agrammatic speakers. Aphasiology 24, 28–55. doi: 10.1080/02687030802586902
Drai, D. (2006). “Evaluating deficit patterns of broca aphasics in the presence of high intersubject variability,” in Broca's Region, eds. Y. Grodzinsky and K. Amunts (Oxford: Oxford University Press). doi: 10.1093/acprof:oso/9780195177640.003.0007
Duffield, C. J. (2016). Frequency, activation, and the production of verb inflection in agrammatic aphasia. Aphasiology 30, 1283–1303. doi: 10.1080/02687038.2015.1105357
Duman, T. Y., Aygen, G., Ozgirgin, N., and Bastiaanse, R. (2007). Object scrambling and finiteness in Turkish agrammatic production. J. Neuroling. 20, 306–331. doi: 10.1016/j.jneuroling.2007.01.001
Duncan, J. (2010). The multiple-demand (MD) system of the primate brain: mental programs for intelligent behaviour. Trends Cogn. Sci. 14, 172–179. doi: 10.1016/j.tics.2010.01.004
Dyson, B., Håkansson, G., and Ballard, K. J. (2022). A developmental approach to assessing and treating agrammatic aphasia. Am. J. Speech-Lang. Pathol. 31, 1188–1204. doi: 10.1044/2022_AJSLP-21-00240
Edwards, S. (1998). Single words are not enough: verbs, grammar and fluent aphasia. Int. J. Lang. Commun. Disor. 33, 190–195. doi: 10.3109/13682829809179421
Edwards, S., and Bastiaanse, R. (1998). Diversity in the lexical and syntactic abilities of fluent aphasic speakers. Aphasiology 12, 99–117. doi: 10.1080/02687039808250466
Embick, D., Marantz, A., Miyashita, Y., O'Neil, W., and Sakai, K. L. (2000). A syntactic specialization for Broca's area. Proc. Nat. Acad. Sci. 97, 6150–6154. doi: 10.1073/pnas.100098897
Engelter, S. T., Gostynski, M., Papa, S., Frei, M., Born, C., Ajdacic, V., et al. (2006). Epidemiology of aphasia attributable to first ischemic stroke. Stroke 37, 1379–1384. doi: 10.1161/01.STR.0000221815.64093.8c
Fadiga, L., Craighero, L., and D'Ausilio, A. (2009). Broca's area in language, action, and music. Ann. NY. Acad. Sci. 1169, 448–458. doi: 10.1111/j.1749-6632.2009.04582.x
Faroqi-Shah, Y. (2007). Are regular and irregular verbs dissociated in non-fluent aphasia?: A meta-analysis. Brain Res. Bull. 74, 1–13. doi: 10.1016/j.brainresbull.2007.06.007
Faroqi-Shah, Y. (2020). Diagnostic criteria for agrammatism: a critical analysis and empirical validation. Academy of Aphasia. Virtual.
Faroqi-Shah, Y., and Dickey, M. W. (2009). On-line processing of tense and temporality in agrammatic aphasia. Brain Lang. 108, 97–111. doi: 10.1016/j.bandl.2008.10.003
Faroqi-Shah, Y., and Friedman, L. (2015). Production of verb tense in agrammatic aphasia: a meta-analysis and further data. Behav. Neurol. 2015, 983870. doi: 10.1155/2015/983870
Faroqi-Shah, Y., and Gehman, M. (2021). The role of processing speed and cognitive control on word retrieval in aging and aphasia. J. Speech, Langu. Hear. Res. 64, 949–964. doi: 10.1044/2020_JSLHR-20-00326
Faroqi-Shah, Y., Kevas, Y., and Li, R. (2021). Lexical category differences in bilingual picture naming: Implications for models of lexical representation. Bilingualism: Lang. Cogn. 24, 849–863. doi: 10.1017/S1366728921000213
Faroqi-Shah, Y., Kling, T., Solomon, J., Liu, S., Park, G., and Braun, A. R. (2014). Lesion analysis of language production deficits in aphasia. Aphasiology 28, 258–277. doi: 10.1080/02687038.2013.853023
Faroqi-Shah, Y., Lynch, R., and Bernstein Ratner, N. (2022a). Uhhmm..disfluencies as a window into lexical and syntactic deficits in aphasia. Paper presented at the Academy of Aphasia, Philadelphia, PA.
Faroqi-Shah, Y., Sebastian, R., and van der Woude, A. (2018). Neural representation of word categories is distinct in the temporal lobe: An activation likelihood analysis. Hum. Brain Mapp. 39, 4925–4938. doi: 10.1002/hbm.24334
Faroqi-Shah, Y., Shi, C., and Goodridge, R. (2022b). Short-term memory in aphasia: effects of modality and relationship with Western Aphasia Battery-R performance. Aphasiology 21, 1–31. doi: 10.1080/02687038.2022.2136482
Faroqi-Shah, Y., Slevc, L. R., Saxena, S., Fisher, S. J., and Pifer, M. (2020). Relationship between musical and language abilities in post-stroke aphasia. Aphasiology 34, 793–819. doi: 10.1080/02687038.2019.1650159
Faroqi-Shah, Y., and Thompson, C. K. (2003). Effect of lexical cues on the production of active and passive sentences in Broca's and Wernicke's aphasia. Brain Lang. 85, 409. doi: 10.1016/S0093-934X(02)00586-2
Faroqi-Shah, Y., and Thompson, C. K. (2004). Semantic, lexical, and phonological influences on the production of verb inflections in agrammatic aphasia. Brain Lang. 89, 484–498. doi: 10.1016/j.bandl.2003.12.006
Faroqi-Shah, Y., and Thompson, C. K. (2007). Verb inflections in agrammatic aphasia: Encoding of tense features. J. Memory Lang. 56, 129–151. doi: 10.1016/j.jml.2006.09.005
Faroqi-Shah, Y., Wood, E., and Gassert, J. (2010). Verb impairment in aphasia: a priming study of body part overlap. Aphasiology. 24, 1377–1388. doi: 10.1080/02687030903515362
Fedorenko, E., and Blank, I. A. (2020). Broca's area is not a natural kind. Trends Cogn. Sci. 24, 270–284. doi: 10.1016/j.tics.2020.01.001
Fedorenko, E., Ryskin, R., and Gibson, E. (2022). Agrammatic output in non-fluent, including Broca's aphasia as a rational behavior. Aphasiology 21, 1–20. doi: 10.1080/02687038.2022.2143233
Fedorenko, E., Williams, Z. M., and Ferreira, V. S. (2018). Remaining puzzles about morpheme production in the posterior temporal lobe. Neuroscience, 392, 160–163. doi: 10.1016/j.neuroscience.2018.09.032
Ferreira, F., and Swets, B. (2002). How incremental is language production? Evidence from the production of utterances requiring the computation of arithmetic sums. J. Memory Lang. 46, 57–84. doi: 10.1006/jmla.2001.2797
Ferrill, M., Love, T., Walenski, M., and Shapiro, L. P. (2012). The time-course of lexical activation during sentence comprehension in people with aphasia [Article]. Am. J. Speech-Lang. Pathol. 21, S179–S189. doi: 10.1044/1058-0360(2012/11-0109)
Flinker, A., Korzeniewska, A., Shestyuk, A. Y., Franaszczuk, P. J., Dronkers, N. F., Knight, R. T., et al. (2015). Redefining the role of Broca's area in speech. Proc. Nat. Acad. Sci. 112, 2871–2875. doi: 10.1073/pnas.1414491112
Flowers, H. L., Skoretz, S. A., Silver, F. L., Rochon, E., Fang, J., Flamand-Roze, C., et al. (2016). Poststroke aphasia frequency, recovery, and outcomes: a systematic review and meta-analysis. Arch. Phys. Med. Rehabil. 97, 2188–2201.e2188. doi: 10.1016/j.apmr.2016.03.006
Frazier, L., and Friederici, A. (1991). On deriving the properties of agrammatic comprehension. Brain Lang. 40, 51–66. doi: 10.1016/0093-934X(91)90116-I
Freiderici, A. D., and Kilborn, K. (1989). Temporal constraints on language processing: Syntactic priming in Broca's aphasia. J. Cogn. Neurosci. 1, 262–272. doi: 10.1162/jocn.1989.1.3.262
Friederici, A. D. (1982). Syntactic and semantic processes in aphasic deficits: the availability of prepositions. Brain Lang. 15, 249–258. doi: 10.1016/0093-934X(82)90059-1
Friederici, A. D. (1995). The time course of syntactic activation during language processing: a model based on neuropsychological and neurophysiological data. Brain Lang. 50, 259–281. doi: 10.1006/brln.1995.1048
Friedmann, N., and Grodzinsky, Y. (1997). Tense and agreement in agrammatic production: pruning the syntactic tree. Brain Lang. 56, 397–425.
Fyndanis, V., Arcara, G., Christidou, P., and Caplan, D. (2018). Morphosyntactic production and verbal working memory: evidence from greek aphasia and healthy aging. J. Speech, Langu. Hear. Res. 61, 1171–1187. doi: 10.1044/2018_JSLHR-L-17-0103
Fyndanis, V., Varlokosta, S., and Tsapkini, K. (2012). Agrammatic production: Interpretable features and selective impairment in verb inflection. Lingua 122, 1134–1147. doi: 10.1016/j.lingua.2012.05.004
Gaeta, L., and Brydges, C. R. (2020). An examination of effect sizes and statistical power in speech, language, and hearing research. J. Speech, Langu. Hear. Res. 63, 1572–1580. doi: 10.1044/2020_JSLHR-19-00299
Gahl, S. (2002). Lexical biases in aphasic sentence comprehension: An experimental and corpus linguistic study. Aphasiology 16, 1173–1198. doi: 10.1080/02687030244000428
Gahl, S., and Menn, L. (2016). Usage-based approaches to aphasia. Aphasiology 30, 1361–1377. doi: 10.1080/02687038.2016.1140120
Gleichgerrcht, E., Roth, R., Fridriksson, J., den Ouden, D., Delgaizo, J., Stark, B., et al. (2021). Neural bases of elements of syntax during speech production in patients with aphasia. Brain Lang. 222, 105025. doi: 10.1016/j.bandl.2021.105025
Goldberg, A. E., and Ferreira, F. (2022). Good-enough language production. Trends Cogn. Sci. 26, 300–311. doi: 10.1016/j.tics.2022.01.005
Goodglass, H. (1976). “Agrammatism,” in Studies in Neurolinguistics, Vol. 1, eds H. Whitaker and H. A. Whitaker (Academic Press).
Goodglass, H., and Berko, J. (1960). Agrammatism and inflectional morphology in English. J. Speech-Lang. Hear. Res. 3, 257–267. doi: 10.1044/jshr.0303.257
Goodglass, H., Christiansen, J. A., and Gallagher, R. (1993). Comparison of morphology and syntax in free narrative and structured tests: fluent vs. non-fluent aphasics. Cortex 29, 377–407. doi: 10.1016/S0010-9452(13)80250-X
Goodglass, H., and Kaplan, E. (1983). “Major aphasic syndromes and illustrations of test patterns,” in The assessment of aphasia and related disorders (Philadelphia: Lea and Febiger) 74–100.
Goodglass, H., Kaplan, E., and Barresi, B. (2001). Boston Diagnostic Aphasia Examination, 3rd Edition. Philadelphia: Lippincott Williams and Wilkins.
Gordon, J. K. (1998). The fluency dimension in aphasia. Aphasiology. 12, 673–688. doi: 10.1080/02687039808249565
Gordon, J. K. (2020). Factor analysis of spontaneous speech in Aphasia. J. Speech Lang. Hear. Res. 63, 4127–4147. doi: 10.1044/2020_jslhr-20-00340
Gordon, J. K., and Clough, S. (2020). How fluent? Part B. Underlying contributors to continuous measures of fluency in aphasia. Aphasiology. 34, 643–663. doi: 10.1080/02687038.2020.1712586
Gordon, J. K., and Dell, G. S. (2003). Learning to divide the labor: an account of deficits in light and heavy verb production. Cogn. Sci. 27, 1–40. doi: 10.1207/s15516709cog2701_1
Griffin, M. Z., and Bock, K. (2000). What the eyes say about speaking. Psychol. Sci. 11, 274–279. doi: 10.1111/1467-9280.00255
Grober, E. (1984). The breakdown of word meanings in aphasia. Cortex 20, 557–566. doi: 10.1016/S0010-9452(84)80058-1
Grodzinsky, Y. (1984). The syntactic characterization of agrammatism. Cognition 16, 99–120. doi: 10.1016/0010-0277(84)90001-5
Grodzinsky, Y. (1986). Language deficits and the theory of syntax. Brain Lang. 27, 135–159. doi: 10.1016/0093-934X(86)90009-X
Grodzinsky, Y. (1988). Syntactic representations in agrammatic aphasia: the case of prepositions. Lang. Speech 31, 115–134. doi: 10.1177/002383098803100202
Grodzinsky, Y. (2000). The neurology of syntax: Language use without Broca's area. Behav. Brain Sci. 23, 1–71. doi: 10.1017/S0140525X00002399
Grodzinsky, Y., and Finkel, L. (1996). Severe grammaticality judgement deficits in agrammatism and Wernicke's aphasia. Brain Lang. 55, 50–53.
Haarmann, H. J., and Kolk, H. H. (1992). The production of grammatical morphology in Broca's and Wernicke's aphasics: speed and accuracy factors. Cortex 28, 97–112. doi: 10.1016/S0010-9452(13)80168-2
Haley, K. L., and Jacks, A. (2019). Word-level prosodic measures and the differential diagnosis of apraxia of speech. Clinical Ling. Phonet. 33, 479–495. doi: 10.1080/02699206.2018.1550813
Harder, P., and Boye, K. (2011). Grammaticalization and functional linguistics. The Oxford Handbook of Grammaticalization. doi: 10.1093/oxfordhb/9780199586783.013.0005
Hauptman, M., Blanco-Elorrieta, E., and Pylkkänen, L. (2022). Inflection across categories: Tracking abstract morphological processing in language production with MEG. Cereb. Cortex 32, 1721–1736. doi: 10.1093/cercor/bhab309
Hazelton, C., McGill, K., Campbell, P., Todhunter-Brown, A., Thomson, K., Nicolson, D. J., et al. (2022). Perceptual disorders after stroke: a scoping review of interventions. Stroke 53, 1772–1787. doi: 10.1161/STROKEAHA.121.035671
Heeschen, C., and Kolk, H. (1988). Agrammatism and paragrammatism. Aphasiology. 2, 299–302. doi: 10.1080/02687038808248928
Hillis, A. E., Work, M., Barker, P. B., Jacobs, M. A., Breese, E. L., and Maurer, K. (2004). Re-examining the brain regions crucial for orchestrating speech articulation. Brain 127, 1479–1487. doi: 10.1093/brain/awh172
Hofstede, B. T. M., and Kolk, H. J. J. (1994). The effects of task variation on the production of grammatical morphology in broca's aphasia: a multiple case study. Brain Lang. 46, 278–328. doi: 10.1006/brln.1994.1017
Howell, P. (2007). A model of serial order problems in fluent, stuttered and agrammatic speech. Hum. Move. Sci. 26, 728–741. doi: 10.1016/j.humov.2007.07.004
Hsu, C.-J., and Thompson, C. K. (2018). Manual vs. automated narrative analysis of agrammatic production patterns: the northwestern narrative language analysis and computerized language analysis. J. Speech, Langu. Hear. Res. 61, 373–385. doi: 10.1044/2017_JSLHR-L-17-0185
Hula, W. D., and McNeil, M. R. (2008). Models of attention and dual-task performance as explanatory constructs in aphasia. Semin. Speech Lang. 29, 169–187. doi: 10.1055/s-0028-1082882
Indefrey, P. (2011). The spatial and temporal signatures of word production components: a critical update. Front. Psychol. 2, 1–16. doi: 10.3389/fpsyg.2011.00255
Indefrey, P., Brown, C. M., Hellwig, F., Amunts, K., Herzog, H., and Seitz, R. J. (2001). A neural correlate of syntactic encoding during speech production. PNAS. 98, 5933–5936.
Ishkhanyan, B., Sahraoui, H., Harder, P., Mogensen, J., and Boye, K. (2017). Grammatical and lexical pronoun dissociation in French speakers with agrammatic aphasia: A usage-based account and REF-based hypothesis. J. Neuroling. 44, 1–16. doi: 10.1016/j.jneuroling.2017.02.001
Ivanova, I., and Ferreira, V. S. (2019). The role of working memory for syntactic formulation in language production. J. Exper. Psychol. 45, 1791–1814. doi: 10.1037/xlm0000672
Ivanova, M. V., and Hallowell, B. (2014). A new modified listening span task to enhance validity of working memory assessment for people with and without aphasia. J. Commun. Disor. 52, 78–98. doi: 10.1016/j.jcomdis.2014.06.001
Jonkers, R., and Bastiaanse, R. (1996). The influence of instrumentality and transitivity on action naming in Broca's and anomic aphasia. Brain Lang. 55, 37–39.
Kauhanen, M. L., Korpelainen, J. T., Hiltunen, P., Määtt,ä, R., Mononen, H., Brusin, E., et al. (2000). Aphasia, depression, and non-verbal cognitive impairment in ischaemic stroke. Cerebrov. Dis. 10, 455–461. doi: 10.1159/000016107
Kean, M. -L. (1977). The linguistic interpretation of aphasic syndromes: Agrammatism in Broca's aphasia, an example. Cognition 5, 9–46. doi: 10.1016/0010-0277(77)90015-4
Kemmerer, D. (2022). Revisiting the relation between syntax, action, and left BA44. Front. Hum. Neurosci. 16, 923022. doi: 10.3389/fnhum.2022.923022
Kertesz, A. (2006). Western Aphasia Battery-Revised. San Antonio, TX: Pearson. doi: 10.1037/t15168-000
Kertesz, A. (2020). The Western Aphasia Battery: a systematic review of research and clinical applications. Aphasiology 36, 21–50. doi: 10.1080/02687038.2020.1852002
Kim, M., and Thompson, C. K. (2000). Patterns of comprhension and production of nouns and verbs in agrammatism: Implications for lexical organization. Brain Lang. 74, 1–25. doi: 10.1006/brln.2000.2315
Kim, M., and Thompson, C. K. (2004). Verb deficits in Alzheimer's disease and agrammatism: Implications for lexical organization. Brain Lang. 88, 1–20. doi: 10.1016/S0093-934X(03)00147-0
Kleist, K. (1916). Uber Leitungsafasie und grammatische Stroungen. Monatsschrift fur Psychiatrie Neurol. 16, 118–121.
Kohn, S. E., and Melvold, J. (2000). Effects of morphological complexity on phonological output deficits in fluent and non-fluent aphasia. Brain Lang. 73, 323. doi: 10.1006/brln.2000.2179
Kok, P., van Doorn, A., and Kolk, H. (2007). Inflection and computational load in agrammatic speech. Brain Lang. 102, 273–283. doi: 10.1016/j.bandl.2007.03.001
Kolk, H. (1995). A time-based approach to agrammatic production. Brain Lang. 50, 282–303. doi: 10.1006/brln.1995.1049
Kolk, H., and van Grunsven, M. (1985). Agrammatism as a variable phenomenon. Cogn. Neuropsychol. 2, 347–384. doi: 10.1080/02643298508252666
Kolk, H. H. J., and Heeschen, C. (1992). Agrammatism, paragrammatism and the management of language. Lang. Cogn. Process. 7, 89–129.
Kunert, R., Willems, R. M., Casasanto, D., Patel, A. D., and Hagoort, P. (2015). Music and language syntax interact in broca's area: An fMRI study. PLoS ONE 10, e0141069. doi: 10.1371/journal.pone.0141069
Lambon Ralph, M. A., Braber, N., McClelland, J., and Patterson, K. (2005). What underlies the neuropsychological pattern of irregular < regular past tense production. Brain Lang. 93, 106–119. doi: 10.1016/j.bandl.2004.09.001
Lange, M. V., Messerschmidt, M., Harder, P., Siebner, H. R., and Boye, K. (2017). Planning and production of grammatical and lexical verbs in multi-word messages. PLoS ONE 12, e0186685. doi: 10.1371/journal.pone.0186685
Lee, J. (2020). Effect of lexical accessibility on syntactic production in aphasia: An eyetracking study. Aphasiology 34, 391–410. doi: 10.1080/02687038.2019.1665963
Lee, J., and Thompson, C. (2011a). Phonological Facilitation Effects during Noun and Verb Naming in Agrammatic and Anomic Aphasia. Proc. Soc. Behav. Sci. 23, 180–181. doi: 10.1016/j.sbspro.2011.09.228
Lee, J., and Thompson, C. K. (2011b). Real-time production of arguments and adjuncts in normal and agrammatic speakers. Lang. Cogn. Proc. 26, 985–1021. doi: 10.1080/01690965.2010.496237
Lee, J., Yoshida, M., and Thompson, C. K. (2015). Grammatical planning units during real-time sentence production in speakers with agrammatic aphasia and healthy speakers. J. Speech, Langu. Hear. Res. 58, 1182–1194. doi: 10.1044/2015_JSLHR-L-14-0250
Lee, L. L., and Canter, S. M. (1971). Developmental sentence scoring: a clinical procedure for estimating syntactic development in children's spontaneous speech. J. Speech Hear. Disor. 36, 315–340. doi: 10.1044/jshd.3603.315
Linebarger, M. C., Schwartz, M. F., and Saffran, E. M. (1983). Sensitivity to grammatical structure in so-called agrammatic aphasics. Cognition 13, 361–392. doi: 10.1016/0010-0277(83)90015-X
Love, T., Swinney, D., Walenski, M., and Zurif, E. (2008). How left inferior frontal cortex participates in syntactic processing: Evidence from aphasia. Brain Lang. 107, 203–219. doi: 10.1016/j.bandl.2007.11.004
Mack, J. E., Barbieri, E., Weintraub, S., Mesulam, M. M., and Thompson, C. K. (2021). Quantifying grammatical impairments in primary progressive aphasia: Structured language tests and narrative language production. Neuropsychologia 151, 107713. doi: 10.1016/j.neuropsychologia.2020.107713
Mack, J. E., Ji, W., and Thompson, C. K. (2013). Effects of verb meaning on lexical integration in agrammatic aphasia: Evidence from eyetracking. J. Neuroling. 26, 619–636. doi: 10.1016/j.jneuroling.2013.04.002
MacWhinney, B., Fromm, D., Forbes, M., and Holland, A. (2011). AphasiaBank: methods for studying discourse. Aphasiology 25, 1286–1307. doi: 10.1080/02687038.2011.589893
Maffei, V., Indovina, I., Mazzarella, E., Giusti, M. A., Macaluso, E., Lacquaniti, F., et al. (2020). Sensitivity of occipito-temporal cortex, premotor and Broca's areas to visible speech gestures in a familiar language. PLoS ONE 15, e0234695. doi: 10.1371/journal.pone.0234695
Mailend, M.-L., Maas, E., Beeson, P. M., Story, B. H., and Forster, K. I. (2019). Speech motor planning in the context of phonetically similar words: Evidence from apraxia of speech and aphasia. Neuropsychologia 127, 171–184. doi: 10.1016/j.neuropsychologia.2019.02.018
Mailend, M.-L., Maas, E., Beeson, P. M., Story, B. H., and Forster, K. I. (2021). Examining speech motor planning difficulties in apraxia of speech and aphasia via the sequential production of phonetically similar words. Cogn. Neuropsychol. 38, 72–87. doi: 10.1080/02643294.2020.1847059
Malvern, D., and Richards, B. J. (1997). “A new measure of lexical diversity,” in Evolving models of language, eds. Ryan and A. Wray (Clevedon, UK: Multilingual Matters) 58–71.
Malyutina, S., and den Ouden, D. B. (2017). Task-dependent neural and behavioral effects of verb argument structure features. Brain Lang. 168, 57–72. doi: 10.1016/j.bandl.2017.01.006
Malyutina, S., and Zelenkova, V. (2020). Verb argument structure effects in aphasia are different at single-word vs. sentence level. Aphasiology 34, 431–457. doi: 10.1080/02687038.2019.1697866
Man, G., Meehan, S., Martin, N., Branigan, H., and Lee, J. (2019). Effects of verb overlap on structural priming in dialogue: implications for syntactic learning in Aphasia. J. Speech, Langu. Hear. Res. 62, 1933–1950. doi: 10.1044/2019_JSLHR-L-18-0418
Marshall, C. R., and van der Lely, H. K. J. (2006). A challenge to current models of past tense inflection: The impact of phonotactics. Cognition 100, 302–320. doi: 10.1016/j.cognition.2005.06.001
Martin, R. C. (2006). The neuropsychology of sentence processing: Where do we stand? Cogn. Neuropsychol. 23, 74–95. doi: 10.1080/02643290500179987
Martin, R. C., Wetzel, W. F., Blossom-Stach, C., and Feher, E. (1989). Syntactic loss vs. processing deficit: an assessment of two theories of agrammatism and syntactic comprehension deficits. Cognition 32, 157–191. doi: 10.1016/0010-0277(89)90002-4
Matchin, W., and Hickok, G. (2020). The cortical organization of syntax. Cereb. Cortex 30, 1481–1498. doi: 10.1093/cercor/bhz180
Matzig, S., Druks, J., Masterson, J., and Vigliocco, G. (2009). Noun and verb differences in picture naming: Past studies and new evidence. Cortex 45, 738–758. doi: 10.1016/j.cortex.2008.10.003
Mauner, G. (1995). Examining the empirical and linguistic bases of current theories of agrammatism. Brain Lang. 50, 339–368. doi: 10.1006/brln.1995.1052
Mayo, N., Wood-Dauphinee, S., Ahmed, S., Carron, G., Higgins, J., McEwen, S., et al. (1999). Disablement following stroke. Disab. Rehabil. 21, 258–268. doi: 10.1080/096382899297684
McRae, K., de Sa, V. R., and Seidenberg, M. S. (1997). On the nature and scope of featural representations of word meaning. J. Exp. Psychol. Gen. 126, 99–130. doi: 10.1037/0096-3445.126.2.99
Menn, L., Gahl, S., Holland, A. L., Ramsberger, G., and Jurafsky, D. S. (2003). Beyond canonical form: verb-frame frequency affects verb production and comprehension. Brain Lang. 87, 1–8. doi: 10.1016/s0093-934x(03)00178-0
Menn, L., and Obler, L. (1990). Agrammatic Aphasia: A Cross-Language Narrative Sourcebook. Amsterdam: Benjamins. doi: 10.1075/z.39
Miceli, G., Mazzucchi, A., Menn, L., and Goodglass, H. (1983). Contrasting cases of Italian agrammatic aphasia without comprehension disorder. Brain Lang. 19, 65–97. doi: 10.1016/0093-934X(83)90056-1
Miceli, G., Silveri, C., Romani, C., and Caramazza, A. (1989). Variation in the pattern of omissions and substitutions of grammatical morphemes in the spontaneous speech of so-called agrammatic patients. Brain Lang. 36, 447–492. doi: 10.1016/0093-934X(89)90079-5
Miceli, G., Silveri, M. C., Nocentini, U., and Caramazza, A. (1988). Patterns of dissociation in comprehension and production of nouns and verbs. Aphasiology. 2, 351–358. doi: 10.1080/02687038808248937
Momma, S., Slevc, L. R., and Phillips, C. (2016). The timing of verb selection in Japanese sentence production. J. Exper. Psychol. 42, 813–824. doi: 10.1037/xlm0000195
Momma, S., Slevc, L. R., and Phillips, C. (2018). Unaccusativity in sentence production. Ling. Inquiry 49, 181–194. doi: 10.1162/LING_a_00271
Monoi, H., Fukusako, Y., Itoh, M., and Sasanuma, S. (1983). Speech sound errors in patients with conduction and Broca's aphasia. Brain Lang. 20, 175–194. doi: 10.1016/0093-934X(83)90041-X
Morrow, E. L., Duff, M. C., and Mayberry, L. S. (2022). Mediators, moderators, and covariates: matching analysis approach for improved precision in cognitive-communication rehabilitation research. J. Speech, Langu. Hear. Res. 65, 4159–4171. doi: 10.1044/2022_JSLHR-21-00551
Murray, L. L. (2012). Attention and other cognitive deficits in aphasia: presence and relation to language and communication measures. Am. J. Speech-Lang. Pathol. 21, S51–64. doi: 10.1044/1058-0360(2012/11-0067)
Murray, L. L., Holland, A. L., and Beeson, P. M. (1997). Grammaticality judgements of mildly aphasic individuals under dual-task conditions. Aphasiology 11, 993–1016. doi: 10.1080/02687039708249423
Na, Y., Jung, J., Tench, C. R., Auer, D. P., and Pyun, S.-B. (2022). Language systems from lesion-symptom mapping in aphasia: A meta-analysis of voxel-based lesion mapping studies. NeuroImage. 35, 103038. doi: 10.1016/j.nicl.2022.103038
Nanousi, V., Masterson, J., Druks, J., and Atkinson, M. (2006). Interpretable vs. uninterpretable features: Evidence from six Greek-speaking agrammatic patients. J. Neuroling. 19, 209–238. doi: 10.1016/j.jneuroling.2005.11.003
Niemi, J., Koivuselkä-Sallinen, P., and Hänninen, R. (1985). Phoneme errors in Broca's aphasia: Three Finnish cases. Brain Lang. 26, 28–48. doi: 10.1016/0093-934X(85)90026-4
Novick, J. M., Trueswell, J. C., and Thompson-Schill, S. L. (2010). Broca's area and language processing: Evidence for the cognitive control connection. Langu. Lingu. Compass 4, 906–924. doi: 10.1111/j.1749-818X.2010.00244.x
Nozari, N., and Faroqi-Shah, Y. (2017). Investigating the origin of non-fluency in aphasia: A path modeling approach to neuropsychology. Cortex 95, 119–135. doi: 10.1016/j.cortex.2017.08.003
Nys, G. M. S., van Zandvoort, M. J. E., de Kort, P. L. M., Jansen, B. P. W., de Haan, E. H. F., and Kappelle, L. J. (2007). Cognitive disorders in acute stroke: prevalence and clinical determinants. Cerebrov. Dis. 23, 408–416. doi: 10.1159/000101464
Obler, L. K., Harris, K., Meth, M., Centeno, J., and Mathews, P. (1999). The phonology-morphosyntax interface: affixed words in agrammatism. Brain Langu. 68, 233–240. doi: 10.1006/brln.1999.2101
O'Grady, W., and Lee, M. (2001). The isomorphic mapping hypothesis: Evidence from Korean. Brain Cogn. 46, 226–230. doi: 10.1016/S0278-2626(01)80072-5
Papitto, G., Friederici, A. D., and Zaccarella, E. (2020). The topographical organization of motor processing: An ALE meta-analysis on six action domains and the relevance of Broca's region. Neuroimage 206, 116321. doi: 10.1016/j.neuroimage.2019.116321
Patel, A. (2003). Language, music, syntax and the brain. Nat. Neurosci. 6, 674–681. doi: 10.1038/nn1082
Patel, A., Iversen, J. R., Wassenaar, M., and Hagoort, P. (2008). Musical syntactic processing in agrammatic Broca's aphasia. Aphasiology 22, 776–789. doi: 10.1080/02687030701803804
Pedersen, P., Vinter, K., and Olsen, T. S. (2004). Aphasia after stroke: type, severity and prognosis. Cerebrov. Dis. 17, 35–43. doi: 10.1159/000073896
Picheny, M. A., Durlach, N. I., and Braida, L. D. (1986). Speaking clearly for the hard of hearing. II: Acoustic characteristics of clear and conversational speech. J. Speech Hear. Res. 29, 434–446. doi: 10.1044/jshr.2904.434
Pick, A. (1913). Die agrammatischen Sprachstorungen [The agrammatic speech disorders]. Berlin: Springer-Verlag. doi: 10.1007/978-3-642-71069-8
Pickering, M. J., and Ferreira, V. S. (2008). Structural priming: A critical review. Psychol. Bull. 134, 427–459. doi: 10.1037/0033-2909.134.3.427
Pompon, R. H., Fassbinder, W., McNeil, M. R., Yoo, H., Kim, H. S., Zimmerman, R. M., et al. (2022). Associations among depression, demographic variables, and language impairments in chronic post-stroke aphasia. J. Commun. Disor. 100, 106266. doi: 10.1016/j.jcomdis.2022.106266
Prather, P., Zurif, E. B., Stern, C., and Rosen, J. (1992). Slowed lexical access in non-fluent aphasia: A case study. Brain Lang. 43, 336–348. doi: 10.1016/0093-934X(92)90134-Z
Prather, P. A., Zurif, E., Love, T., and Brownell, H. (1997). Speed of lexical activation in non-fluent Broca's aphasia and fluent Wernicke's aphasia. Brain Lang. 59, 391–411. doi: 10.1006/brln.1997.1751
Pregla, D., Vasishth, S., Lissón, P., Stadie, N., and Burchert, F. (2022). Can the resource reduction hypothesis explain sentence processing in aphasia? A visual world study in German. Brain Lang. 235, 105204. doi: 10.1016/j.bandl.2022.105204
Python, G., Glize, B., and Laganaro, M. (2018). The involvement of left inferior frontal and middle temporal cortices in word production unveiled by greater facilitation effects following brain damage. Neuropsychologia 121, 122–134. doi: 10.1016/j.neuropsychologia.2018.10.026
Rezaii, N., Mahowald, K., Ryskin, R., Dickerson, B., and Gibson, E. (2022). A syntax-lexicon trade-off in language production. Proc. Nat. Acad. Sci. 119, e2120203119. doi: 10.1073/pnas.2120203119
Richardson, J. D., Fillmore, P., Rorden, C., LaPointe, L. L., and Fridriksson, J. (2012). Re-establishing Broca's initial findings. Brain Lang. 123, 125–130. doi: 10.1016/j.bandl.2012.08.007
Robinson, G., Blair, J., and Cipolotti, L. (1998). Dynamic aphasia: an inability to select between competing verbal responses? Brain 121, 77–89. doi: 10.1093/brain/121.1.77
Robinson, G., Shallice, T., Bozzali, M., and Cipolotti, L. (2010). Conceptual proposition selection and the LIFG: neuropsychological evidence from a focal frontal group. Neuropsychologia 48, 1652–1663. doi: 10.1016/j.neuropsychologia.2010.02.010
Rochon, E., Saffran, E. M., Berndt, R. S., and Schwartz, M. F. (2000). Quantitative analysis on aphasic sentence production: further development and new data. Brain Lang. 72, 193–218. doi: 10.1006/brln.1999.2285
Roger, E., Banjac, S., Thiebaut de Schotten, M., and Baciu, M. (2022). Missing links: The functional unification of language and memory (L?M). Neurosci. Biobehav. Rev. 133, 104489. doi: 10.1016/j.neubiorev.2021.12.012
Roos, N. M., and Piai, V. (2020). Across-session consistency of context-driven language processing: A magnetoencephalography study. Euro. J. Neurosci. 52, 3457–3469. doi: 10.1111/ejn.14785
Saffran, E. M., Berndt, R. S., and Schwartz, M. F. (1989). The quantitative analysis of agrammatic production: Procedure and data. Brain Lang. 37, 440–479. doi: 10.1016/0093-934X(89)90030-8
Saffran, E. M., Schwartz, M. F., and Marin, O. S. M. (1980). The word order problem in agrammatism: II. Production. 10, 263. doi: 10.1016/0093-934X(80)90056-5
Sahin, N. T., Pinker, S., Cash, S. S., Schomer, D., and Halgren, E. (2009). Sequential processing of lexical, grammatical, and phonological information within Broca's area. Science 326, 445–449. doi: 10.1126/science.1174481
Sahin, N. T., Pinker, S., and Halgren, E. (2006). Abstract grammatical processing of nouns and verbs in Broca's area: Evidence from FMRI. Cortex 42, 540–562. doi: 10.1016/S0010-9452(08)70394-0
Salis, C., and DeDe, G. (2022). Sentence production in a discourse context in latent aphasia: a real-time study. Am. J. Speech-Lang. Pathol. 31, 1284–1296. doi: 10.1044/2022_AJSLP-21-00232
Salis, C., and Edwards, S. (2008). Comprehension of wh-questions and declarative sentences in agrammatic aphasia: The set partition hypothesis. J. Neuroling. 21, 375–399. doi: 10.1016/j.jneuroling.2007.11.001
Sampson, M., and Faroqi-Shah, Y. (2011). Investigation of self-monitoring in fluent aphasia with jargon. Aphasiology 25, 505–515. doi: 10.1080/02687038.2010.523471
Schnur, T. T., Schwartz, M. F., Kimberg, D. Y., Hirshorn, E., Coslett, H. B., and Thompson-Schill, S. L. (2009). Localizing interference during naming: Convergent neuroimaging and neuropsychological evidence for the function of Broca's area. Proc. Nat. Acad. Sci. 106, 322–327. doi: 10.1073/pnas.0805874106
Scholes, R. J. (1982). The verb-right strategy in agrammatic aphasia. Neuropsychologia 20, 361–363. doi: 10.1016/0028-3932(82)90111-7
Shapiro, K., and Caramazza, A. (2003). Grammatical processing of nouns and verbs in left frontal cortex? Neuropsychologia. 41, 1189–1198. doi: 10.1016/S0028-3932(03)00037-X
Shapiro, K., Moo, L. R., and Caramazza, A. (2006). Cortical signatures of noun and verb production. Proc. Nat. Acad. Sci. USA 103, 1644–1649. doi: 10.1073/pnas.0504142103
Sheppard, S. M., Walenski, M., Love, T., and Shapiro, L. P. (2015). The Auditory Comprehension of Wh-Questions in Aphasia: Support for the Intervener Hypothesis. J. Speech, Langu. Hear. Res. 58, 781–797. doi: 10.1044/2015_JSLHR-L-14-0099
Sikora, K., Roelofs, A., Hermans, D., and Knoors, H. (2016). Executive control in spoken noun-phrase production: Contributions of updating, inhibiting, and shifting. Quart. J. Exper. Psychol. 69, 1719–1740. doi: 10.1080/17470218.2015.1093007
Silverman, S. W., and Ratner, N. B. (1997). Syntactic complexity, fluency, and accuracy of sentence imitation in adolescents. J. Speech, Langu. Hear. Res. 40, 95–106. doi: 10.1044/jslhr.4001.95
Skajaa, N., Adelborg, K., Horváth-Puhó, E., Rothman, K. J., Henderson, V. W., Thygesen, L. C., et al. (2022). Stroke and risk of mental disorders compared with matched general population and myocardial infarction comparators. Stroke 53, 2287–2298. doi: 10.1161/STROKEAHA.121.037740
Slevc, L. R. (2023). “Grammatical Encoding,” in Language Production, eds. R. J. Hartsuiker and K. Strijkers (London: Routledge). doi: 10.4324/9781003145790-2
Slevc, L. R., and Martin, R. C. (2016). Syntactic agreement attraction reflects working memory processes. J. Cogn. Psychol. 28, 773–790. doi: 10.1080/20445911.2016.1202252
Sonty, S. P., Mesulam, M.-M., Thompson, C. K., Johnson, N. A., Weintraub, S., Parrish, T. B., et al. (2003). Primary progressive aphasia: PPA and the language network. Ann. Neurol. 53, 35–49. doi: 10.1002/ana.10390
Speer, P., and Wilshire, C. E. (2013). What's in a sentence? The crucial role of lexical content in sentence production in non-fluent aphasia. Cogn. Neuropsychol. 30, 507–543. doi: 10.1080/02643294.2013.876398
Stampacchia, S., Thompson, H. E., Ball, E., Nathaniel, U., Hallam, G., Smallwood, J., et al. (2018). Shared processes resolve competition within and between episodic and semantic memory: Evidence from patients with LIFG lesions. Cortex 108, 127–143. doi: 10.1016/j.cortex.2018.07.007
Stavrakaki, S., Alexiadou, A., Kambanaros, M., Bostantjopoulou, S., and Katsarou, Z. (2011). The production and comprehension of verbs with alternating transitivity by patients with non-fluent aphasia. Aphasiology 25, 642–668. doi: 10.1080/02687038.2010.542248
Swick, D., Ashley, V., and Turken, and U. (2008). Left inferior frontal gyrus is critical for response inhibition. BMC Neurosci. 9, 102. doi: 10.1186/1471-2202-9-102
Swinburn, K., Porter, G., and Howard, D. (2004). Comprehensive Aphasia Test (1 ed.). Hove: Psychology Press. doi: 10.1037/t13733-000
Swinney, D., and Zurif, E. (1995). Syntactic processing in aphasia. Brain Lang. 50, 225–239. doi: 10.1006/brln.1995.1046
Szekely, A., D'Amico, S., Devescovi, A., Federmeier, K., Herron, D., Iyer, G., et al. (2005). Timed action and object naming. Cortex 41, 7–25. doi: 10.1016/S0010-9452(08)70174-6
Szupica-Pyrzanowska, M., Obler, L. K., and Martohardjono, G. (2017). Morphological vs. phonological explanations for affix errors in agrammatism. Aphasiology 31, 928–950. doi: 10.1080/02687038.2016.1225273
Tettamanti, M., and Weniger, D. (2006). Broca's area: a supramodal hierarchical processor? Cortex 42, 491–494. doi: 10.1016/S0010-9452(08)70384-8
Thompson, C. K. (2003). Unaccusative verb production in agrammatic aphasia: the argument structure complexity hypothesis. J. Neuroling. 16, 151–167. doi: 10.1016/S0911-6044(02)00014-3
Thompson, C. K., and Bastiaanse, R. (2012). “Introduction to agrammatism,” in Perspectives on Agrammatism, eds. R. Bastiaanse and C. K. Thompson (Hove: Psychology Press).
Thompson, C. K., Cho, S., Hsu, C.-J., Wieneke, C., Rademaker, A., Weitner, B. B., et al. (2012). Dissociations between fluency and agrammatism in primary progressive aphasia. Aphasiology 26, 20–43. doi: 10.1080/02687038.2011.584691
Thompson, C. K., den Ouden, D. B., Bonakdarpour, B., Garibaldi, K., and Parrish, T. B. (2010). Neural plasticity and treatment-induced recovery of sentence processing in agrammatism. Neuropsychologia. 48, 3211–3227. doi: 10.1016/j.neuropsychologia.2010.06.036
Thorne, J., and Faroqi-Shah, Y. (2016). Verb production in aphasia: testing the division of labor between syntax and semantics. Sem. Speech Lang. 37, 023–033. doi: 10.1055/s-0036-1571356
Timmermans, M., Schriefers, H., Sprenger, S., and Dijkstra, T. (2012). Describing simple events: The dynamics of incremental grammatical encoding. J. Cogn. Psychol. 24, 441–456. doi: 10.1080/20445911.2011.649254
Trost, J. E., and Canter, G. J. (1974). Apraxia of speech in patients with Broca's aphasia: A study of phoneme production accuracy and error patterns. Brain Lang. 1, 63–79. doi: 10.1016/0093-934X(74)90026-1
Trupe, L. A., Varma, D. D., Gomez, Y., Race, D., Leigh, R., Hillis, A. E., et al. (2013). Chronic apraxia of speech and “Broca's Area”. Stroke J. Cereb. Circ. 44, 740–744. doi: 10.1161/STROKEAHA.112.678508
Ullman, M. T. (2001). The neural basis of lexicon and grammar in first and second language: the declarative/procdeural model. Bilingualism 4, 105–122. doi: 10.1017/S1366728901000220
Ullman, M. T., Corkin, S., Coppola, M., and Hickok, G. (1997). A neural dissociation within language: Evidence that the mental dictionary is part of declarative memory, and that grammatical rules are processed by the procedural system. J. Cogn. Neurosci. 9, 266–276. doi: 10.1162/jocn.1997.9.2.266
Varlakosta, S., Valeonti, N., Kakavoulia, M., Lazaridou, M., Economou, A., and Protopapas, A. (2006). The breakdown of functional categories in Greek aphasia: evidence from agreement, tense and aspect. Aphaisology 20, 723–743. doi: 10.1080/02687030500513703
von Stockert, T. R., and Bader, L. (1976). Some relations of grammar and lexicon in aphasia. Cortex 12, 49–60. doi: 10.1016/S0010-9452(76)80029-9
Wallace, S. J., Worrall, L., Rose, T., Le Dorze, G., Breitenstein, C., Hilari, K., et al. (2019). A core outcome set for aphasia treatment research: The ROMA consensus statement. Int. J. Stroke. 14, 180–185. doi: 10.1177/1747493018806200
Walsh, B., and Smith, A. (2011). Linguistic Complexity, Speech Production, and Comprehension in Parkinson's Disease: Behavioral and Physiological Indices. J. Speech, Langu. Hear. Res. 54, 787–802. doi: 10.1044/1092-4388(2010/09-0085)
Webster, J., Franklin, S., and Howard, D. (2001). An investigation of the interaction between thematic and phrasal structure in non-fluent agrammatic subjects. Brain Lang. 78, 197–200. doi: 10.1006/brln.2001.2460
Wenzlaff, M., and Clahsen, H. (2004). Tense and agreement in German agrammatism. Brain Lang. 89, 57–68. doi: 10.1016/S0093-934X(03)00298-0
Wilson, S. M., and Saygin, A. P. (2004). Grammaticality judgment in aphasia: Deficits are not specific to syntactic structures, aphasic syndromes or lesion sites. J. Cogn. Neurosci. 16, 238–252. doi: 10.1162/089892904322984535
Wright, H. H., Koutsoftas, A. D., Capilouto, G. J., and Fergadiotis, G. (2014). Global coherence in younger and older adults: Influence of cognitive processes and discourse type. Aging, Neuropsychol. Cogn. 21, 174–196. doi: 10.1080/13825585.2013.794894
Wulfeck, B., Bates, E., and Capasso, R. (1991). A crosslinguistic study of grammaticality judgements in Broca's aphasia. Brain Lang. 41, 311–336. doi: 10.1016/0093-934X(91)90158-W
Yoo, H., McNeil, M. R., Dickey, M. W., and Terhorst, L. (2022). Linguistic and nonlinguistic processing speed across age-matched normal healthy controls and individuals with left-hemisphere damage, with and without aphasia. Aphasiology 36, 147–169. doi: 10.1080/02687038.2020.1853966
Zaccarella, E., Schell, M., and Friederici, A. D. (2017). Reviewing the functional basis of the syntactic Merge mechanism for language: A coordinate-based activation likelihood estimation meta-analysis. Neurosci. Biobehav. Rev. 80, 646–656. doi: 10.1016/j.neubiorev.2017.06.011
Zhang, H., and Hinzen, W. (2022). Grammar in ‘agrammatical' aphasia: What's intact? PLoS ONE 17, e0278676. doi: 10.1371/journal.pone.0278676
Zhu, Y., Xu, M., Lu, J., Hu, J., Kwok, V. P. Y., Zhou, Y., et al. (2022). Distinct spatiotemporal patterns of syntactic and semantic processing in human inferior frontal gyrus. Nat. Hum. Behav. 6, 1104–1111. doi: 10.1038/s41562-022-01334-6
Ziegler, W., Aichert, I., Staiger, A., Willmes, K., Baumgaertner, A., Grewe, T., et al. (2022). The prevalence of apraxia of speech in chronic aphasia after stroke: A bayesian hierarchical analysis. Cortex 151, 15–29. doi: 10.1016/j.cortex.2022.02.012
Zingeser, L. B., and Berndt, R. S. (1990). Retrieval of nouns and verbs in agrammatism and anomia [see comments]. Brain Lang. 39, 14–32. doi: 10.1016/0093-934X(90)90002-X
Zurif, E., Swinney, D., Prather, P., Solomon, J., and Bushell, C. (1993). An on-line analysis of syntactic processing in Broca's and Wernicke's aphasia. Brain Lang. 45, 448–464. doi: 10.1006/brln.1993.1054
Keywords: agrammatism, aphasia, syntax, morphology, phonology, processing load, verb
Citation: Faroqi-Shah Y (2023) A reconceptualization of sentence production in post-stroke agrammatic aphasia: the Synergistic Processing Bottleneck model. Front. Lang. Sci. 2:1118739. doi: 10.3389/flang.2023.1118739
Received: 07 December 2022; Accepted: 21 July 2023;
Published: 15 September 2023.
Edited by:
Simona Mancini, Brain and Language, SpainReviewed by:
Svetlana Malyutina, National Research University Higher School of Economics, RussiaChristina Manouilidou, University of Ljubljana, Slovenia
Copyright © 2023 Faroqi-Shah. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Yasmeen Faroqi-Shah, eWZzaGFoQHVtZC5lZHU=