 Hengameh Chloé Mirsepasi-Lauridsen1
Hengameh Chloé Mirsepasi-Lauridsen1 Katleen Vrankx2
Katleen Vrankx2 Jørgen Engberg3
Jørgen Engberg3 Alice Friis-Møller4Jørn Brynskov5Inge Nordgaard-Lassen6
Alice Friis-Møller4Jørn Brynskov5Inge Nordgaard-Lassen6 Andreas Munk Petersen4,6
Andreas Munk Petersen4,6 Karen Angeliki Krogfelt1*
Karen Angeliki Krogfelt1*- 1Department of Bacteria, Parasites and Fungi, Statens Serum Institut, Copenhagen, Denmark
- 2Applied Maths NV, Sint-Martens-Latem, Belgium
- 3Department of Clinical Microbiology, Slagelse Hospital, Slagelse, Denmark
- 4Department of Clinical Microbiology, Hvidovre University Hospital, Hvidovre, Denmark
- 5Department of Gastroenterology, Herlev University Hospital, Herlev, Denmark
- 6Department of Gastroenterology, Hvidovre University Hospital, Hvidovre, Denmark
Inflammatory Bowel disease (IBD) is traditionally divided into Crohn's disease (CD) and ulcerative colitis (UC). UC is a relapsing non-transmural inflammatory disease that is restricted to the colon and is characterized by flare-ups of bloody diarrhea. CD is a chronic, segmental localized granulomatous disease that can affect any part of the entire gastrointestinal tract. Ileo-anal pouch is a procedure restoring functionality of the rectum after a colectomy. IBD is a multifactorial disease and flares of IBD are probably triggered by changes in the intestinal microbiota followed by an abnormal immune response. In this study, we aim to analyze the intestinal bacterial diversity in IBD patients during various stages of disease compared with healthy controls. Permission for human experiments and recruitment of participants was obtained from the Ethic Committee for Copenhagen County hospitals (Permission no. KA-03019, Permission no. KA-20060159). Stools from 26 healthy controls, 42 CD, 38 UC and 18 pouch patients were collected. Stool DNA extraction was performed using Qiagen, DNA mini stool kit Denmark. DGGE-PCR amplifying the V2-V3 region of 16S-rDNA gene of the bacteria was amplified by universal primers HDA1 and HDA2. Analysis of DGGE was performed blinded using BioNumerics version 7.5. After normalization, a DGGE gel band matching was performed. The similarities between profiles were calculated with a ranked Pearson correlation coefficient based on the band matching results using band intensities. Simpson's index of diversity and Pielou's species evenness were calculated. Based on the Shannon Diversity Index, UC patients had lower species diversity and bacterial evenness in comparison to healthy persons, p < 0.05. However, only CD and disease pouch patients had lower species diversity compared to those with inactive disease and healthy controls. Well-functioning pouch patients had decreased species evenness in comparison to diseased pouch patients and control group. During the active disease stage in CD and pouch, the patients have a low bacterial diversity in their gut when compared to the inactive stage. In UC patients, a generally low diversity was observed at all stages of the disease compared to healthy controls.
Introduction
Crohn's disease (CD) and ulcerative colitis (UC) are the two main forms of inflammatory bowel disease (IBD), characterized by periods of remission and relapses (1). CD is a chronic inflammatory condition that involves any part of the gastrointestinal tract, while UC is a chronic inflammation localized in the colon (1). The etiology of IBD is still unknown, but many studies indicate a combination of nutrition (2, 3), intestinal dysbacteriosis (4, 5), and abnormal immune response (6–8). During the last two decades, the focus has been on the role of microbiota as a driver of the inflammatory process in IBD.
One theory suggests that living in an industrialized country with high hygiene standards lowers the exposure to complex microbial communities during the development of the immune system. As a result, the immune system is less able to tolerate exposure to environments rich on microbiota resulting in an inappropriate immune activation (6). Humans live in a symbiotic relationship with the gut microbiome, in which humans provide the ideal environment for the microbiota to flourish and bacteria in return ferment the food carbohydrates to short fatty acids, synthesize certain vitamins and degrade dietary oxalates (2–5).
Studies show that IBD with active disease is strongly associated with an overall increased prevalence of proteobacteria (9, 10) and drop in intestinal species richness, more specifically a significant drop in firmicutes (11, 12), Enterobacteriaceae, Bacteroidales, and Clostridiales (13) of Clades IV and XIVa (13). Decreased prevalence of butyrate-producing bacteria such as Clostridiales species in the gut explains the decreased amount of short chain fatty acids in fecal samples from IBD patients (14, 15). Butyrate serves as a major source of energy for colonic epithelial cells (16) and as an inhibitor of pro-inflammatory cytokine expression in the intestinal mucosa through a mechanism that involves hyper-acetylation of histones and suppression of NF-κB signaling (17). Butyrate reinforces the mucosal barrier by inducing production of mucin and antimicrobial peptides, and by strengthening epithelial barrier integrity through directly increasing the expression of tight junction proteins (18). More recent studies indicate a link between IBD and a decreased prevalence of butyrate-producing Faecalibacterium prausnitzii (firmicute), belonging to Clostridia cluster IV species (19, 20), increased prevalence of adherent-invasive E. coli (AIEC) in ileal CD (21, 22) and increased prevalence of E. coli with extra intestinal pathogenic E. coli properties (23–25). It has been shown that fecal microbiota transplantation (FMT) is an effective treatment of UC, indicating that colon microbiota play an essential role in UC (26). A subdivision of IBD into three phenotypes as ileal CD, colonic CD, and UC according to genetic composition has been suggested (27).
In this study, the goal was to investigate intestinal bacterial diversity in IBD patients with active and inactive disease compared to healthy controls, using PCR amplification of regions of the 16S ribosomal RNA gene and Denaturing Gradient Gel Electrophoresis (DGGE).
Material and Methods
This study included 97 adult IBD patients with a previously confirmed IBD diagnosis and 31 healthy controls, aged from 25 to 86 and 18 to 67 years, respectively. Symptom scores, stool and blood samples were collected from 38 patients with UC, 15 females and 23 males, 20 with active (UCA) and 18 with inactive (UCI) disease, 41 patients with Crohn's disease, 28 females and 13 males, 21 with active (CDA) and 20 with inactive (CDI) disease, 18 patients with an ileo-anal pouch (UC patients who underwent surgical treatment), 8 females and 10 males, including 7 pouchitis (PA), and 11 well-functioning pouch (PI) patients. The disease activity was measured with the Simple Clinical Colitis Activity Index (SCCAI) (28), the Harvey-Bradshaw Index (HBI) (29), and the Modified Pouchitis Disease Activity Index (MPDAI) (30). All the participants answered the questionnaires regarding medicine or antibiotic usage, travel diarrhea/diarrhea in the last 3 months before participating in the study. Patients or controls, who used antibiotics or corticoides 3 months before the start of the study, were excluded. Overall, 98% of the study group originated from Denmark, while the last 2% were integrated Danish citizens of other ethnicity.
The stool samples were stored at −80°C and thawed before processing. The laboratory staff was blinded to patient histories.
Ethics Statement
Permission for the study was obtained from the Regional Ethics Committee for Copenhagen County Hospitals (Permission no. KA-03019, Permission no. KA-20060159) and all participants gave their informed written consent. Healthy controls were recruited among students and volunteers. Patients and healthy controls completed a questionnaire about their condition and their medication. IBD patients were diagnosed according to standardized criteria (29, 31).
DNA Extraction
DNA extraction was performed according to the instructions of the manufacturer (Qiagen, DNA stool mini kit Denmark) with the following modifications: 100 mg fecal sample was mixed with 1.4 ml ASL buffer in a 2-ml tube and vortexed until the sample was thoroughly homogenized. Samples subsequently mixed with 0.2 g sterile zirconia/silica beads. Hereafter, the samples were processed on a TissueLyser for 6 min at 30 Hz. Lysis was completed at a temperature of 95° C for 5 min. Finally, DNA was extracted according to the instruction of the QIAamp DNA stool MiniKit and eluted in 100 μl elution buffer provided in the kit.
PCR Amplification
The V2-V3 region of the 16S rDNA gene (~200 bp) was amplified using the universal primer set HDA 1 position 338-357: (5′ACT CCT ACG GGA GGC AGC AGT′3) and HDA 2 position 539–561: (5′GTA TTA CCG CGG CTG CTG GCA C–′3) (32). The forward primer, HDA 1, was at the 5′end labeled with GC clamp (5′CGC CCG GGG CGC GCC CCG GGC GGG GCG GGG GCA CGGGGG G ′3). PCRs were performed in a total volume of 50 μL containing 20 μL of 5 PRIME Mastermix, 0.8 μM primer HDA 1-GC, 0.8 μM primer HDA 2, 10 μL of DNA template and, finally, 4 μL RNase free water.
The PCR was performed using the following conditions: preheating at 94°C for 4 min proceeded by 30 cycles of denaturing at 94°C for 30 s, annealing at 56°C for 30 s, elongation at 68°C for 45 s, and finally a single step of 68°C for 7 min. The PCR products were run on a 0.8% agarose gel.
Denaturing Gradient Gel Electrophoresis
PCR fragments were separated by Denaturing Gradient Gel Electrophoresis (DGGE) with DCode System according to the manufacturer's instructions. Eight percent Polyacrylamide (vol/vol) [ratio of acrylamid:bisacrylamide (37.5:1)] was diluted in 0.5xTAE buffer with pH 8.0 using a gradient ranging from 35 to 65% [100% acrylamide corresponds to 7 M urea and 40% (vol/vol) formamide] (33). Gels were cast using a gradient maker and a pump with a flow speed of 5 mL per minute. After polymerization of the gel (2 h), a 3% stacking gel without denaturing chemicals was cast, and an appropriate comb was subsequently inserted and left for 30 min for polymerization. Gels were run at 60°C for 16 h at a constant voltage of 70 V in 0.5x TAE buffer. After electrophoresis, gels were stained with GelRED for 45 min and analyzed using an ultraviolet trans-illuminator.
Data Analyses
Analysis of the DGGE was performed blinded using BioNumerics version 7.5 (Applied Maths NV, Sint-Martens-Latem, Belgium). After normalization, a band matching was performed with a tolerance of 1% and an optimization of 1%. The results from the automatic band matching were checked manually and corrected where necessary. The similarity between profiles was calculated with a Pearson correlation. A dendrogram was then constructed with UPGMA. The reliability of the dendrogram was determined with a cophenetic correlation coefficient. This coefficient compares a similarity matrix derived from the dendrogram with the actual similarity matrix. Reliably separated branches have a high cophenetic correlation. For each profile, the Shannon Diversity Index of diversity and Pielou's species evenness were calculated. Linear discriminant analysis (LDA) was used to analyze differences within patient groups and healthy persons. All statistical analyses were performed after mean-based normalization using analysis of variance (Anova) with post hoc test Bonferroni. The level of significance was set at p ≤ 0.05.
Results
DNA was extracted from fecal samples and analyzed by DGGE. The individual bands each representing a bacterium were analyzed in BioNumerics. The microbiota was correlated to the diagnosis. Based on the Shannon Diversity Index, UC patients with active and inactive disease had significant lower bacterial diversity in comparison to healthy controls, p < 0.05* (Figure 1A). When comparing the species evenness using Pielou's index, UC patients had decreased species evenness compared with healthy controls, p ≤ 0.05 (Figure 1B). No such difference was found between UC patients with active and inactive disease, respectively.
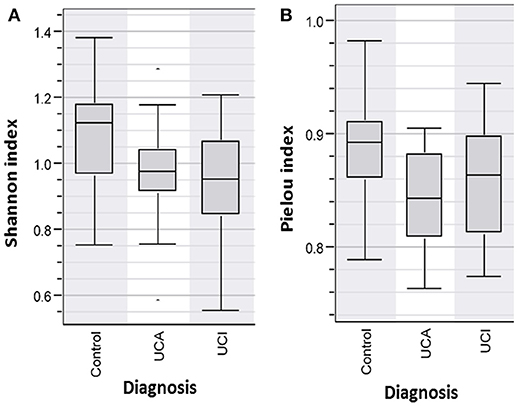
Figure 1. (A) Shannon Diversity Index for patients with active (UCA) and inactive (UCI) Ulcerative colitis vs. healthy person/Control group shows patients group (UCI and UCA) have a significantly decreased bacterial abundance in comparison to controls group, p < 0.05*(B) Pielou index shows significantly decreased bacterial evenness in the patient groups (UCI and UCA) vs. controls, p < 0.05.
Not only a reduced number of bands (low bacterial diversity), but also brighter bands (increased bacterial concentration), were observed using DGGE profiles in UC patients compared to healthy controls. When using cluster analysis (Figure 2A), UC patients with (red) and without (blue) active disease were separated from healthy controls (green). This was confirmed by linear discriminant analysis (LDA; Figure 2B), where the controls in the red circle were separated from patients (Figures 2A,B).
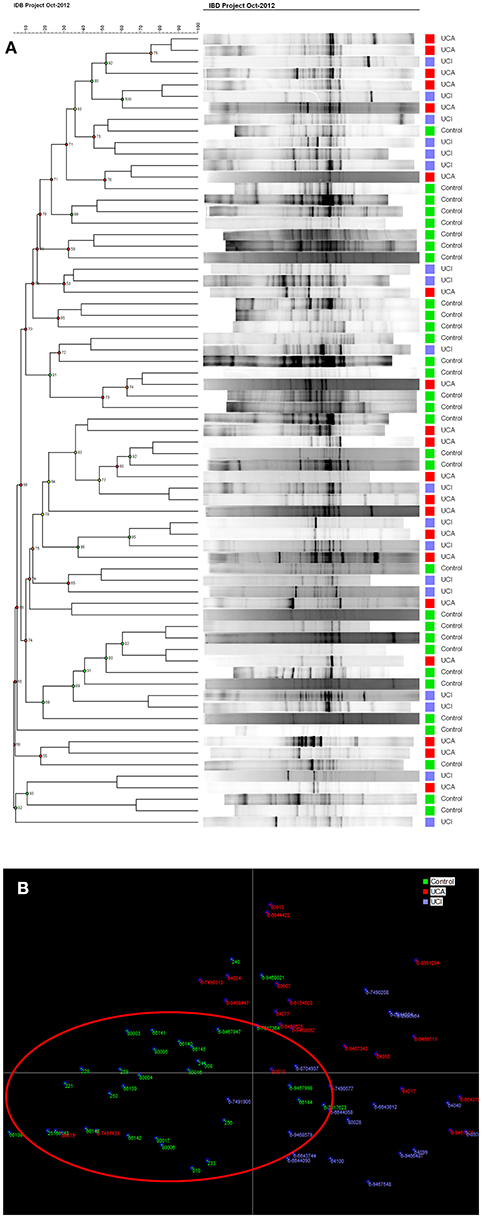
Figure 2. (A) Cluster analysis of control group and patient groups with active (UCA in red)/inactive (UCI in blue) ulcerative colitis. (B) LDA shows that controls in green are mostly gathered in the red circle.
Analysis of the bacterial diversity of CD patients with active (CDA) and inactive (CDI) disease in comparison with controls showed that CD patients with active disease had a significantly lower Shannon Diversity Index in comparison to healthy persons, p < 0.05* (Figure 3A). When comparing the species evenness using Pielou's index, no significant differences were found between patient groups vs. control group/healthy persons, p < 0.083 (Figure 3B).
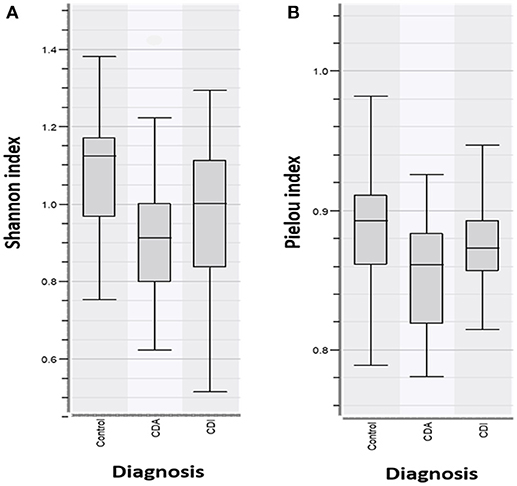
Figure 3. (A) Shannon Diversity Index for CD patients with active (CDA) and inactive disease (CDI) vs. controls show CDA have significantly decreased bacterial abundance in comparison to CDI and controls, p > 0.05*. (B) Pielou index shows no significant differences in species evenness in patient groups vs. controls (p < 0.083).
Cluster analysis (Figure 4A) showed that generally profiles from CD patients with (CDA, red) active disease could be found in the same cluster as other CDA profiles. The same was seen for CDI and control group. However, many exceptions were found and the clusters were not clearly delineated. This effect was seen more clearly with LDA, where healthy controls (in green) shown in the red circle, were separated from the patients (Figure 4B). No separation could be observed in the CDA and CDI analysis.
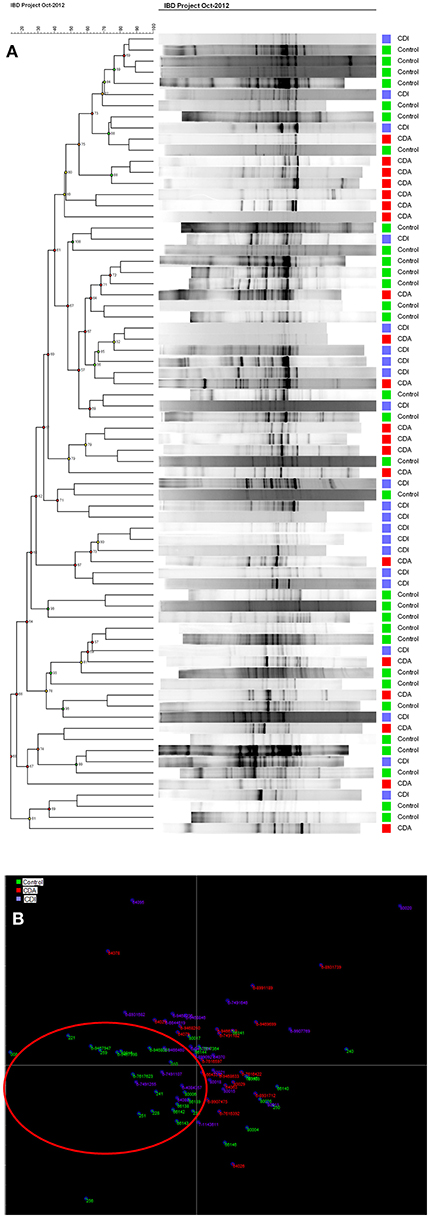
Figure 4. (A) Cluster Analysis of controls and patient groups with active (CDA, in red) and inactive (CDI, in blue) Crohn's disease shows the group clustering in colors (B) LDA shows control in green are mostly separated in the red ring, from CDA in red and CDI in blue.
In comparison to healthy persons and well-functioning pouch patients (PI), diseased pouch patients (PA) had a significantly decreased Shannon Diversity Index, p < 0.05 (Figure 5A). When comparing the species evenness using Pielou's evenness index, a lower evenness was found in the patient groups; however, only the difference between controls and PI reached significance p < 0.05 (Figure 5B).
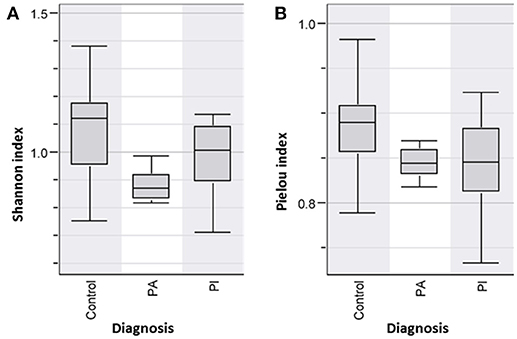
Figure 5. (A) Shannon Diversity Index of pouch patient groups with active (PA) and inactive disease (PI) and controls shows PA has significant decreased bacterial diversity index in comparison to the PI and control group, p < 0.05. The differences were not significant in PI, p-value = 0.06. (B) Species evenness (Pielou index) was lower in the patient groups compared to the healthy controls, however these differences were only significant in PI, p < 0.05.
Cluster analysis (Figure 6A) showed that DGGE profiles from diseased pouch patients (red) and well-functioning pouch patients (blue) were more similar to profiles from other patients, while healthy controls also clustered with other healthy controls (green). However, the clusters were not clearly delineated. A better separation could be seen by LDA, where the majority of controls separated from the patients shown in red circle (Figures 6A,B).
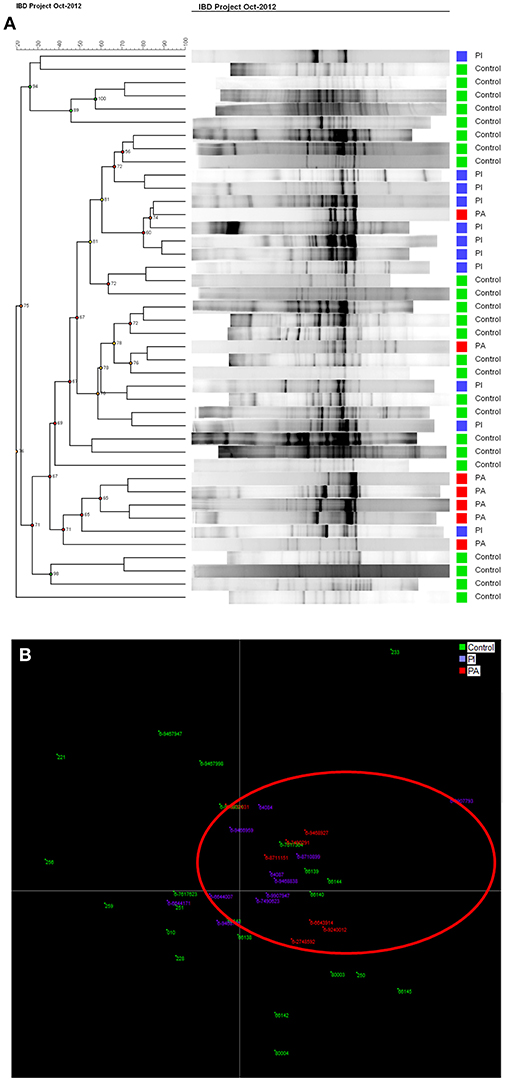
Figure 6. (A) Cluster analysis for pouch patients with active (PA, in red) and inactive (PI, in blue) disease shows controls (green) clustered mostly together. (B) LDA graph shows most of controls in green are separated from PI in blue and PA in red.
Discussion
The pathogenesis of IBD is still not clear. However, during the last decades, the main focus has been on an imbalance in the intestinal microbiome in IBD patients that might cause or trigger the disease activity. Next generation sequencing investigation of the gut microbiome provides large amounts of data, which can be difficult to interpret, and it is still quite costly. The advantages of using DGGE (34) compared with next generation sequencing is that DGGE detects highly prevalent (high concentration) intestinal bacteria, while next generation sequencing is random and detects all DNA. Sequencing is biased by the primer annealing, and even small amounts of DNA will be amplified. In this study, we investigated the intestinal microbiota in IBD using 16S rDNA PCR and DGGE enabling the discovery of overall differences in intestinal bacteria community within different IBD groups and controls. When discussing the concept of diversity of a microbial community, the terms species evenness and richness are used. Bacterial richness, also known as the Shannon diversity index is a measurement of the total number of species in the community, while evenness expresses how evenly the individuals in the community are distributed over the different species (35). As the colon contains two-thirds of the total amount of bacteria in the human body (36), it is an exceptional environment, and a perfect symbiosis plays a key role in colon health. UC is localized in the colon and is linked to intestinal bacterial dysbiosis and reduced biodiversity (37). This is also confirmed in our study, where UC patients with and without active disease had significantly reduced Shannon diversity indices and reduced fecal bacterial evenness in comparison with healthy persons (Figure 1). LDA and cluster analysis (Figure 2B) confirm this result, showing a separation between UC patients and healthy controls. Yet, the question of what triggers the disease activity in UC patients remains. If bacteria in the colon are responsible, what are the differences in the intestinal bacteria community in the UC patients with and without active disease? As we see an overall reduced biodiversity in UC patients, the flares of UC might be triggered by the shift between the different bacterial communities in UC patient. The studies using next generation sequencing reveal that a reduced amount of F. praustnitzii (38) and an increased number of Proteobacteriae (39, 40) might have an effect on UC relapses. In order to restore the biodiversity, fecal microbiota transplantation (FMT) was performed and it was shown to be effective to promote remission in UC patients with active disease, confirming the role of the colon microbiota in UC (26).
CD is a chronic inflammatory condition that involves any part of the gastrointestinal tract. Colonic CD has been identified in 60% of the CD patients (41), which often present with diarrhea and an aggressive course of the disease. Intestinal dysbiosis is also linked to colonic CD, with increased prevalence of Faecalibacterium (Firmicutes) and two unidentified genera of Clostridiales and Ruminococceae (42), while ileal patients have a higher prevalence of Proteobacteria compared to healthy subjects. Our results show that CD patients with active disease have a significantly reduced Shannon diversity index in comparison with CD patients with inactive disease and healthy controls (Figure 3A). Decreased species evenness was found in CD patients with and without active disease compared to healthy persons; however, these differences were not significant, p = 0.08 (Figure 3B). Interestingly, a metanalysis performed on FMT in CD, reveals that FMT promotes remission in only 60% of the CD patients (43), as only 60% of the CD patients had colonic CD (41). Reduced bacterial diversity in children with CD was reported previously (40, 44, 45).
Ileal pouch-anal anastomosis is a surgical treatment choice for the majority of UC patients, who fail medical treatment therapy, with the advantage of re-establishment of gastrointestinal continuity and improvement of the quality of life for these patients. Studies show that 40% of patients with ileal pouch-anal anastomosis develop pouchitis during the first year after surgery (46). Furthermore, antibiotic and probiotic treatment appears to be effective to diminish pouchitis (47). Evidence suggests that an abnormal mucosal immune response to altered microbiota in the pouch leads to acute chronic inflammation (48). Our analysis of bacterial diversity in pouch patients in comparison with healthy persons shows that diseased pouch patients (PA) have a significant decreased microbial diversity in comparison to well-functioning pouch patients and healthy persons, p < 0.05 (Figure 5A). Reduced diversity was shown in well-functioning pouch patients in comparison to healthy controls; however, the differences are not significant (Figure 5A). Additionally, our results reveal significantly decreased fecal bacterial evenness in well-functioning pouch patients suggesting that the microbiota in these patients is dominated by just a few species. Maharshak et al. (49) show that pre-pouchititis patients have significantly decreased intestinal bacterial diversity 1 year before pouchitis in comparison to pouch patients, who did not develop pouchitis. The possibility of subclinical inflammation 1 year before pouchitis was confirmed using histological scores, endoscopics and fecal calprotectin (49). This study supports our results showing that well-functioning pouch patients have significantly decreased fecal bacterial evenness and reduced Shannon diversity indices in comparison to healthy persons, which was also confirmed by our LDA analysis (Figure 6B). It could be speculated whether most of the well-functioning pouch patients in our cohort are in the stage of pre-pouchitis, shown by reduced intestinal bacteria evenness in these patients. Increased relapses in pouch patients might be explained by the surgical treatment where the pouch has a “colon like” function, which creates a good environment for bacteria to overgrow and cause infections. This supports the finding that FMT does not promote remission in pouchitis patients (50) as antibiotics do.
Microbial colonization has an important effect on the instruction and regulation of the immune system (51). Abnormal communication between gut microbial communities and the mucosal immune system has been identified as the core defect that leads to chronic intestinal inflammation (52). Current studies are focusing on restoring a healthy intestinal microbiota using FMT and probiotics to treat the intestinal bacterial dysbiosis. This might open new treatment opportunities for IBD patients treated with immune suppressive medication or for those where medical treatment therapy fails.
Conclusion
This study shows that UC patients with and without active disease have significantly decreased intestinal bacterial diversity and evenness in comparison to healthy persons. However, CD patients and diseased pouch patients had significantly decreased intestinal bacterial diversity, during the active disease compared to healthy persons, CD patients and well-functioning pouch patients without active disease.
The well-functioning pouch patients had significantly decreased species evenness in comparison to diseased pouch patients and healthy persons.
Author Contributions
AP, KK, JE, AF-M, IN-L, HM-L, and JB participated in the design of the study. KV, AP, KK, and HM-L drafted the manuscript. HM-L and KV were responsible for the experimental setting. All authors have read and approved the final manuscript.
Funding
We thank for funding provided to HM-L by the Lundbeck foundation and Med Vet Net Association.
Conflict of Interest Statement
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Acknowledgments
We thank Susanne Jespersen at Statens Serum Institut for helpful discussions, expertise, and Marian Jørgensen for her linguistic review of the manuscript. We thank MD Signe Wildt for helping to recruit patients in the project.
Abbreviations
IBD, Inflammatory Bowel Disease; UC, Ulcerative colitis; UCA, Ulcerative colitis with active disease; UCI, Ulcerative colitis with inactive disease; CD, Crohn's disease; CDA, Crohn's with active disease; CDI, Crohn's with inactive disease; PA, Disease pouch; PI, well-functioning pouch.
References
1. Baumgart DC, Sandborn WJ. Inflammatory bowel disease: clinical aspects and established and evolving therapies. Lancet (2007) 369:1641–57. doi: 10.1016/S0140-6736(07)60751-X
2. Hebuterne X, Filippi J, Al Jaouni R, Schneider S. (2009). Nutritional consequences and nutrition therapy in Crohn's disease. GastroenterolClin Biol. 33(Suppl. 3):S235–44. doi: 10.1016/S0399-8320(09)73159-8
3. Albenberg LG, Wu GD. Diet and the intestinal microbiome: Associations, functions, and implications for health and disease. Gastroenterology (2014) 146:1564–72. doi: 10.1053/j.gastro.2014.01.058
4. Png CW, Linden SK, Gilshenan KS, Zoetendal EG, McSweeney CS, Sly LI, et al. Mucolytic bacteria with increased prevalence in IBD mucosa augment in vitro utilization of mucin by other bacteria. AmJ Gastroenterol. (2010) 105:2420–8. doi: 10.1038/ajg.2010.281
5. Fava F, Danese S. Intestinal microbiota in inflammatory bowel disease: friend of foe? World J Gastroenterol. (2011) 17:557–66. doi: 10.3748/wjg.v17.i5.557
6. Bouma G, Strober W. The immunological and genetic basis of inflammatory bowel disease. Nat Rev Immunol. (2003) 3:521–33. doi: 10.1038/nri1132
7. Keshavarzian A, Fusunyan RD, Jacyno M, Winship D, MacDermott RP, Sanderson IR. Increased interleukin-8 (IL-8) in rectal dialysate from patients with ulcerative colitis: evidence for a biological role for IL-8 in inflammation of the colon. Am J Gastroenterol. (1999) 94:704–12. doi: 10.1111/j.1572-0241.1999.00940.x
8. Nuding S, Fellermann K, Wehkamp J, Stange EF. Reduced mucosal antimicrobial activity in Crohn's disease of the colon. Gut (2007) 56:1240–7. doi: 10.1136/gut.2006.118646
9. Baumgart M, Dogan B, Rishniw M, Weitzman G, Bosworth B, Yantiss R, et al. Culture independent analysis of ileal mucosa reveals a selective increase in invasive Escherichia coli of novel phylogeny relative to depletion of Clostridiales in Crohn's disease involving the ileum. ISMEJ. (2007) 1:403–18. doi: 10.1038/ismej.2007.52
10. Gophna U, Sommerfeld K, Gophna S, Doolittle WF, Veldhuyzen Van Zanten SJO. Differences between tissue-associated intestinal microfloras of patients with Crohn's disease and ulcerative colitis. J Clin Microbiol. (2006) 44:4136–41. doi: 10.1128/JCM.01004-06
11. Frank DN, St Amand AL, Feldman RA, Boedeker EC, Harpaz N, Pace NR. Molecular-phylogenetic characterization of microbial community imbalances in human inflammatory bowel diseases. Proc Natl Acad Sci USA. (2007) 104:13780–5. doi: 10.1073/pnas.0706625104
12. Ott SJ, Musfeldt M, Wenderoth DF, Hampe J, Brant O, Fölsch UR, et al. Reduction in diversity of the colonic mucosa associated bacterial microflora in patients with active inflammatory bowel disease. Gut (2004) 53:685–93. doi: 10.1136/gut.2003.025403
13. Gevers D, Kugathasan S, Denson LA, Vázquez-Baeza Y, Van Treuren W, Ren B, et al. The treatment-naive microbiome in new-onset Crohn's disease. Cell Host Microbe (2014) 15:382–92. doi: 10.1016/j.chom.2014.02.005
14. Vernia P, Gnaedinger A, Hauck W, Breuer RI. Organic anions and the diarrhea of inflammatory bowel disease. Dig Dis Sci. (1988) 33:1353–8. doi: 10.1007/BF01536987
15. Treem WR, Ahsan N, Shoup M, Hyams JS. Fecal Short_Chain Fatty Acids in Children with Inflammatory Bowel Disease. J Pediatr Gastroenterol Nutr. (1994) 18:159–64. doi: 10.1097/00005176-199402000-00007
16. Fleming LLFM. Digestion and absorption of fiber carbohydrate in the colon. Am J Gastroenterol. (1986) 81:507–11.
17. Segain J, De Blétière DR, Bourreillea A, Leray V, Gervois N, Rosales C, et al. Butyrate inhibits inflammatory responses through NF B inhibition : implications for Crohn's disease. Gut (2000) 47:397–403. doi: 10.1136/gut.47.3.397
18. Vanhoutvin SALW, Troost FJ, Hamer HM, Lindsey PJ, Koek GH, Jonkers DMAE, et al. Butyrate-induced transcriptional changes in human colonic mucosa. PLoS ONE 4:e6759. doi: 10.1371/journal.pone.0006759
19. Sokol H, Pigneur B, Watterlot L, Lakhdari O, Bermúdez-Humarán LG, Gratadoux J-J, et al. Faecalibacterium prausnitzii is an anti-inflammatory commensal bacterium identified by gut microbiota analysis of Crohn disease patients. Proc Natl Acad Sci USA. (2008) 105:16731–6. doi: 10.1073/pnas.0804812105
20. Sokol H, Seksik P, Furet JP, Firmesse O, Nion-Larmurier I, Beaugerie L, et al. Low counts of faecalibacterium prausnitzii in colitis microbiota. Inflamm Bowel Dis. (2009) 15:1183–9. doi: 10.1002/ibd.20903
21. Martinez-Medina M, Aldeguer X, Lopez-Siles M, Gonzalez-Huix F, Lopez-Oliu C, Dahbi G, et al. Molecular diversity of Escherichia coli in the human gut: new ecological evidence supporting the role of adherent-invasive E. coli (AIEC) in Crohn's disease. InflammBowel Dis. (2009) 15:872–82. doi: 10.1002/ibd.20860
22. Darfeuille-Michaud A, Boudeau J, Bulois P, Neut C, Glasser AL, Barnich N, et al. High prevalence of adherent-invasive Escherichia coli associated with ileal mucosa in Crohn's disease. Gastroenterology (2004) 127:412–21. doi: 10.1053/j.gastro.2004.04.061
23. Mirsepasi-Lauridsen HC, Du Z, Struve C, Charbon G, Karczewski J, Krogfelt KA, et al. Secretion of alpha-hemolysin by Escherichia coli disrupts tight junctions in ulcerative colitis patients. Clin Transl Gastroenterol. (2016) 7:e149. doi: 10.1038/ctg.2016.3
24. Mirsepasi-Lauridsen HC, Halkjaer SI, Mortensen EM, Lydolph MC, Nordgaard-Lassen I, et al. Extraintestinal pathogenic Escherichia coli are associated with intestinal inflammation in patients with ulcerative colitis. Sci Rep. (2015) 6:31152. doi: 10.1038/srep31152
25. Petersen AM, Nielsen EM, Litrup E, Brynskov J, Mirsepasi H, Krogfelt KA. A phylogenetic group of Escherichia coli associated with active left-sided inflammatory bowel disease. BMC Microbiol. (2009) 9:171–9. doi: 10.1186/1471-2180-9-171
26. Costello SP, Soo W, Bryant RV, Jairath V, Hart AL, Andrews JM. Systematic review with meta-analysis: faecal microbiota transplantation for the induction of remission for active ulcerative colitis. Aliment Pharmacol Ther. (2017) 46:213–24. doi: 10.1111/apt.14173
27. Cleynen I, Boucher G, Jostins L, Schumm LP, Zeissig S, Ahmad T, et al. Inherited determinants of Crohn's disease and ulcerative colitis phenotypes: a genetic association study. Lancet (2016) 387:156–67. doi: 10.1016/S0140-6736(15)00465-1
28. Walmsley RS, Ayres RC, Pounder RE, Allan RN. A simple clinical colitis activity index. Gut (1998) 43:29–32. doi: 10.1136/gut.43.1.29
30. Shen B, Achkar JP, Connor JT, Ormsby AH, Remzi FH, Bevins CL, et al. Modified pouchitis disease activity index: a simplified approach to the diagnosis of pouchitis. Dis Colon Rectum (2003) 46:748–53. doi: 10.1007/s10350-004-6652-8
31. Powell-Tuck J, Day DW, Buckell NA, Wadsworth J, Lennard-Jones JE. Correlations between defined sigmoidoscopic appearances and other measures of disease activity in ulcerative colitis. Dig Dis Sci. (1982) 27:533–7. doi: 10.1007/BF01296733
32. Walter J, Tannock GW, Tilsala-Timisjarvi A, Rodtong S, Loach DM, Munro K, et al. Detection and identification of gastrointestinal Lactobacillus species by using denaturing gradient gel electrophoresis and species-specific PCR primers. Appl Environ Microbiol. (2000) 66:297–303. doi: 10.1128/AEM.66.1.297-303.2000
33. Smith B, Li N, Andersen AS, Slotved HC, Krogfelt KA. Optimising bacterial DNA extraction from faecal samples: comparison of three methods. Open Microbiol J. (2011) 5:14–7. doi: 10.2174/1874285801105010014
34. Myers RM, Fischer SG, Lerman LS, Maniatis T. Nearly all single base substitutions in DNA fragments joined to a GC-clamp can be detected by denaturing gradient gel electrophoresis. Nucleic Acids Res. (1985) 13:3131–45. doi: 10.1093/nar/13.9.3131
36. Sender R, Fuchs S, Milo R. Revised estimates for the number of human and bacteria cells in the body. PLoS Biol. (2016) 14:1–14. doi: 10.1371/journal.pbio.1002533
37. Kostic AD, Xavier RJ, Gevers D. The microbiome in iflammatory bowel diseases: current status and the future ahead. Gastroenterology (2015) 146:1489–99. doi: 10.1053/j.gastro.2014.02.009
38. Varela E, Manichanh C, Gallart M, Torrejón A, Borruel N, Casellas F, et al. Colonisation by Faecalibacterium prausnitzii and maintenance of clinical remission in patients with ulcerative colitis. Aliment Pharmacol Ther. (2013) 38:151–61. doi: 10.1111/apt.12365
39. Walujkar SA, Dhotre DP, Marathe NP, Lawate PS, Bharadwaj RS, Shouche YS. Characterization of bacterial community shift in human Ulcerative Colitis patients revealed by Illumina based 16S rRNA gene amplicon sequencing. Gut Pathog. (2014) 6:7–13. doi: 10.1186/1757-4749-6-22
41. Hedrick T, Friel C. Colonic crohn disease. Clin Colon Rectal Surg. (2013) 26:84–9. doi: 10.1055/s-0033-1348046
42. Naftali T, Reshef L, Kovacs A, Porat R, Amir I, Konikoff FM, et al. Distinct microbiotas are associated with ileum-restricted and colon-involving crohn's disease. Inflamm Bowel Dis. (2016) 22:293–302. doi: 10.1097/MIB.0000000000000662
43. Xu L, Zhang T, Cui B, He Z, Xiang J, Long C, et al. Clinical efficacy maintains patients' positive attitudes toward fecal microbiota transplantation. Medicine (2016) 95:e4055. doi: 10.1097/MD.0000000000004055
44. Hansen R, Russell RK, Reiff C, Louis P, McIntosh F, Berry SH, et al. Microbiota of de-novo pediatric IBD: increased Faecalibacterium prausnitzii and reduced bacterial diversity in Crohn's but not in ulcerative colitis. Am J Gastroenterol. (2012) 107:1913–22. doi: 10.1038/ajg.2012.335
45. Qin J, Li R, Raes J, Arumugam M, Burgdorf KS, Manichanh C, et al. A human gut microbial gene catalogue established by metagenomic sequencing. Nature (2010) 464:59–65. doi: 10.1038/nature08821
46. Shen B, Lashner BA. Diagnosis and treatment of pouchitis. Gastroenterol Hepatol. (2008) 4:355–61.
47. Segal JP, Ding NS, Worley G, Mclaughlin S, Preston S, Faiz OD, et al. Systematic review with meta-analysis: the management of chronic refractory pouchitis with an evidence-based treatment algorithm. Aliment Pharmacol Ther. (2017) 45:581–92. doi: 10.1111/apt.13905
48. Gionchetti P, Rizzello F, Venturi A, Brigidi P, Matteuzzi D, Bazzocchi G, et al. Oral bacteriotherapy as maintenance treatment in patients with chronic pouchitis: a double-blind, placebo-controlled trial.Gastroenterology (2000) 119:305–9. doi: 10.1053/gast.2000.9370
49. Maharshak N, Cohen NA, Reshef L, Tulchinsky H, Gophna U, Dotan I. Alterations of enteric microbiota in patients with a normal ileal pouch are predictive of pouchitis. J Crohns Colitis (2017) 11:314–20. doi: 10.1093/ecco-jcc/jjw157
50. Schmid M, Frick JS, Malek N, Goetz M. Successful treatment of pouchitis with Vedolizumab, but not fecal microbiota transfer (FMT), after proctocolectomy in ulcerative colitis. Int J Colorectal Dis. (2017) 32:597–8. doi: 10.1007/s00384-017-2761-4
51. Guarner F, Bourdet-Sicard R, Brandtzaeg P, Gill HS, McGuirk P, van Eden W, et al. Mechanisms of disease: the hygiene hypothesis revisited. Nat Clin Pract Gastroenterol Hepatol. (2006) 3:275–84. doi: 10.1038/ncpgasthep0471
Keywords: inflammatory bowel disease, ulcerative colitis, crohn's disease, intestinal microbiome, ileo-anal pouch, dysbiosis, shannon diversity index, pielou index
Citation: Mirsepasi-Lauridsen HC, Vrankx K, Engberg J, Friis-Møller A, Brynskov J, Nordgaard-Lassen I, Petersen AM and Krogfelt KA (2018) Disease-Specific Enteric Microbiome Dysbiosis in Inflammatory Bowel Disease. Front. Med. 5:304. doi: 10.3389/fmed.2018.00304
Received: 21 March 2018; Accepted: 15 October 2018;
Published: 20 November 2018.
Edited by:
Yeong Yeh Lee, University of Science, Malaysia, MalaysiaReviewed by:
Yuji Naito, Kyoto Prefectural University of Medicine, JapanZheng Feei Ma, University of Liverpool, United Kingdom
Copyright © 2018 Mirsepasi-Lauridsen, Vrankx, Engberg, Friis-Møller, Brynskov, Nordgaard-Lassen, Petersen and Krogfelt. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Karen Angeliki Krogfelt, a2FrQHNzaS5kaw==