 Tania Cebrero-Cangueiro1,2,3†
Tania Cebrero-Cangueiro1,2,3† Gema Labrador-Herrera1,2†
Gema Labrador-Herrera1,2† Álvaro Pascual2,4,5
Álvaro Pascual2,4,5 Caridad Díaz6
Caridad Díaz6 Jesús Rodríguez-Baño2,3,4
Jesús Rodríguez-Baño2,3,4 Jerónimo Pachón2,3
Jerónimo Pachón2,3 José P. del Palacio6
José P. del Palacio6 María E. Pachón-Ibáñez1,2*
María E. Pachón-Ibáñez1,2* M. Carmen Conejo5
M. Carmen Conejo5- 1Clinical Unit of Infectious Diseases, Microbiology, and Preventive Medicine, University Hospital Virgen del Rocío, Seville, Spain
- 2Institute of Biomedicine of Seville (IBiS), Virgen del Rocío and Virgen Macarena University Hospitals/Consejo Superior de Investigaciones Científicas (CSIC)/University of Seville, Seville, Spain
- 3Department of Medicine, University of Seville, Seville, Spain
- 4Clinical Unit of Infectious Diseases, Microbiology, and Preventive Medicine, Virgen Macarena University Hospital, Seville, Spain
- 5Department of Microbiology, University of Seville, Seville, Spain
- 6Fundacion Centro de Excelencia en Investigación de Medicamentos Innovadores en Andalucía, MEDINA Foundation, Granada, Spain
Carbapenemase-producing Klebsiella pneumoniae infections are an increasing global threat with scarce and uncertain treatment options. In this context, combination therapies are often used for these infections. The bactericidal and synergistic activity of fosfomycin plus amikacin and gentamicin was studied trough time–kill assays against four clonally unrelated clinical isolates of carbapenemase-producing K. pneumoniae, VIM-1, VIM-1 plus DHA-1, OXA-48 plus CTXM-15, and KPC-3, respectively. The efficacy of antimicrobials that showed synergistic activity in vitro against all the carbapenemase-producing K. pneumoniae were tested in monotherapy and in combination, in a murine peritoneal sepsis model. In vitro, fosfomycin plus amikacin showed synergistic and bactericidal effect against strains producing VIM-1, VIM-1 plus DHA-1, and OXA-48 plus CTX-M-15. Fosfomycin plus gentamicin had in vitro synergistic activity against the strain producing KPC-3. In vivo, fosfomycin and amikacin and its combination reduced the spleen bacterial concentration compared with controls groups in animals infected by K. pneumoniae producing VIM-1 and OXA-48 plus CTX-M-15. Moreover, amikacin alone and its combination with fosfomycin reduced the bacteremia rate against the VIM-1 producer strain. Contrary to the in vitro results, no in vivo efficacy was found with fosfomycin plus amikacin against the VIM-1 plus DHA-1 producer strain. Finally, fosfomycin plus gentamicin reduced the bacterial concentration in spleen against the KPC-3 producer strain. In conclusion, our results suggest that fosfomycin plus aminoglycosides has a dissimilar efficacy in the treatment of this severe experimental infection, when caused by different carbapenemase-producing K. pneumoniae strains. Fosfomycin plus amikacin or plus gentamycin may be useful to treat infections by OXA-48 plus CTX-M-15 or KPC-3 producer strains, respectively.
Introduction
Klebsiella pneumoniae carbapenemase (KPC)-producers (KPC-KP) represent an increasing global threat worldwide and is one of the most important existing pathogens, especially in endemic areas. KPC-KP causes mostly nosocomial infections although they can occur in healthy people. Most common infections among others are urinary tract, pneumonia and bacteraemia (1). Resistance to carbapenems in K. pneumoniae is mainly due to the production of carbapenem-hydrolyzing β-lactamases such as the KPC type (Ambler class A), IMP, VIM and NMD types (Ambler class B) and OXA-48 (Ambler class D), and bacteremia due to these strains is associated with higher rates of treatment failure and death (2). Moreover, carbapenemase producers often show co-resistance to the majority of other antimicrobial agents, leaving scarce and uncertain treatment options, such as tigecycline, colistin, and some aminoglycosides (2–4). Then, the increment of infections by carbapenemase-producing strains prompts the search of new therapies for infections caused by them. Also, awareness of the prevalence and incidence of the specific mechanisms of carbapenem resistance within K. pneumoniae is crucial in the prevention of their spread and selection of appropriate treatment options (5).
In the search of optimal treatments for infections by these carbapenemase-producing strains, combination among antibiotics is being explored (6, 7), together with newer options like plazomicin, eravacycline and cefiderocol (8). The most commonly in vitro active and potentially useful drugs remain ceftazidime-avibactam and newer inhibitors combinations, gentamicin, amikacin, colistin, tigecycline, and fosfomycin (9). Many in vitro studies and some in vivo studies have investigated the effects of double and triple combinations of drugs with different mechanisms of action (10, 11). Synergistic actions have been convincingly demonstrated for carbapenem-resistant Enterobacteriaceae (particularly K. pneumoniae), Acinetobacter baumannii, and Pseudomonas aeruginosa. In this regard, the study of Erdem et al. showed in vitro that double carbapenem antibiotics plus colistin could be a potential alternative to treat colistin and carbapenem-resistant K. pneumoniae (7). Another in vitro study against multidrug-resistant K. pneumoniae producing KPC-2, KPC-3, NDM-1, OXA-48 and VIM-1 carbapenemases points to polymyxin B in combination with minocycline, rifampicin or fosfomycin as potentially therapy of interest (12). Nevertheless, there are only a few in vivo experimental studies that evaluate antimicrobials in combination against carbapenemase producers K. pneumoniae strains. Hagihara et al., found that meropenem plus amikacin was effective against KPC-, IMP- and OXA-48-producing K. pneumoniae infections, except for NDM type carbapenemase producing (13). Also, the combination of meropenem plus colistin was found to be bactericidal against a KPC-producing K. pneumoniae experimental osteomyelitis in rabbits (14). Several studies have suggested fosfomycin combination with other antimicrobials to be explored because of its in vitro activity. In this regard, Yu et al., demonstrated in vitro synergy with fosfomycin plus imipenem, ertapenem and tigecycline against KPC-producing K. pneumoniae (15). Also, fosfomycin plus gentamicin were synergist in vitro against KPC-3 producing strains (16). Nevertheless, and although the combination of fosfomycin to antimicrobials has been found to be active in vitro, there are not in vivo studies to evaluate this therapeutic alternative, except fosfomycin plus colistin combination in experimental osteomyelitis due to KPC-producing K. pneumoniae, which achieved a reduction the bacterial bone concentration (17).
The aim of the present study was evaluate the efficacy of the fosfomycin plus amikacin or gentamicin in a murine peritoneal sepsis model using four K. pneumoniae clinical strains producers of the currently most prevalent carbapenemases.
Materials and Methods
Bacterial Strains
In this study, we tested four genetically unrelated clinical isolates of KPC-KP: a VIM-1 ST 1603 clone producing isolate, a VIM-1 with the acquired AmpC type beta-lactamase DHA-1 ST 11 producing isolate, a OXA-48 ST 11 clone with the extended spectrum beta-lactemase (ESBL) CTX-M-15, and a KPC-3 ST 512 clone with the broad spectrum beta-lactamases TEM-1 and SHV-11 (18). The identification of the isolates, the presence of carbapenemase genes, and genes coding for other beta-lactamases and the absence of genetic relatedness among the isolates was confirmed by a Microflex LT- MALDI Biotyper mass spectrometer, PCR and sequencing, and PFGE analysis, respectively, as described previously (18). Two of the strains, KPC-3 and VIM-1 (DHA-1) were multidrug-resistant while the other two were not.
Drugs
For the in vitro assays, antimicrobials were used as standard laboratory powders (Sigma-Aldrich, Madrid, Spain). For in vivo experiments, clinical formulations of antimicrobials were used: fosfomycin intravenous 1 g; (Laboratorios ERN, S.A; Barcelona, Spain), Amikacin Normon 125 mg/ml (Laboratorios Normon, S.A; Madrid, Spain) and gentamicin, genta-gobens 80 mg, (Laboratorios Normon, S.A; Madrid, Spain).
In vitro Studies
Antimicrobial Susceptibility Testing
The minimal inhibitory concentrations (MIC) values were tested. MICs of fosfomycin, gentamicin and amikacin were determined by broth microdilution method as recommended by the Clinical Laboratory Standard Institute (CLSI) (19), using Mueller Hinton broth II (MHB) (Becton Dickinson & Co., Sparks, MD, United States) supplemented or not with 25 mg/L of glucose-6-phosphate (G-6-P) (Sigma-Aldrich, Madrid, Spain) and agar dilution method using Mueller Hinton agar supplemented with G-6-P for fosfomycin. MIC of fosfomycin was also determined by the broth microdilution method using MHB supplemented with G-6-P. MIC results were interpreted according to the European Committee on Antimicrobial Susceptibility Testing (EUCAST) (http://www.eucast.org/clinical_breakpoints/) breakpoints for all antibiotics (20). Studies were performed in triplicate to ensure reproducibility.
Time-Kill Curves
Time-kill methodology was used to study the in vitro interactions between fosfomycin plus amikacin and gentamicin. The antibiotic concentrations used for susceptible strains corresponded to the value of their MIC obtained by the broth microdilution method, whereas the concentrations used for resistant strains were those of the susceptibility breakpoints recommended by EUCAST. Experiments were carried out in MHB supplemented with G-6-P with a starting inoculum of 1 × 106 cfu/mL and the drugs alone and in combination. Tubes were incubated at 37°C, with shaking and samples were taken at 0, 1 3, 6, and 24 h, serially diluted, plated (Eddy Jet, IUL S.A., Barcelona, Spain) and incubated at 37°C (16, 21). Bacterial colonies were counted after 24 h using an automatic colony counter (Flash & Go, IUL S.A., Barcelona, Spain). Experiments were performed at least three times on separate occasions. Bactericidal activities of single drugs or combination were defined as a decrease ≥ 3 log10 cfu/mL from the starting inoculum, bacteriostatic effect was defined as no change respect to the initial bacterial concentration during the 24 h. Synergy was defined as a decrease ≥ 2 log10 cfu/mL for the drugs combination compared with the most active single agent (22).
Animals
Immunocompetent C57BL/6J female mice weighing 16–20 g (7–9 weeks old) were used (Production and Experimentation Animal Center, University of Seville, Seville, Spain). Animals had a sanitary status of murine pathogen free and were assessed for genetic authenticity. Mice were housed in an individually ventilated cage system under specific pathogen-free conditions, with water and food ad libitum. This study was carried out in strict accordance with the recommendations in the Guide for the Care and Use of Laboratory Animals (23). In vivo experiments were approved by the Committee on the Ethics of Animal Experiments of the University Hospital Virgen Macarena, Seville, Spain (CI 1961). Procedures were performed under sodium thiopental (B. Braun Medical S.A., Spain) anesthesia, and all efforts were made to minimize suffering.
Pharmacokinetic/Pharmacodynamic Analysis
Serum antibiotic concentrations were determined in groups of healthy mice after a single intraperitoneal administration of fosfomycin (500 mg/kg), amikacin (15 mg/kg) or gentamicin (5 mg/kg). In sets of three animals and at 5, 10, 15, 30, 60, 90, 120, 240, 480, and 1,440 min after the administration of each antibiotic, blood samples were obtained from anesthetized mice from the periorbital plexus. Blood samples were immediately centrifuged at 4,500 rpm for 15 min at 4°C, and serum samples were stored at −80°C until its analysis. Serum concentrations of fosfomycin, amikacin and gentamicin were measured using a HPLC-tandem mass spectrometry (LC-MS/MS) (24). Measurement of the fosfomycin, amikacin and gentamicin binding to mice plasma proteins was also performed (25).
The maximum concentration of drug in serum (Cmax), elimination half-life (t1/2), Area Under the concentration-time Curve from 0 to 24 h (AUC0−24), free AUC0−24 (f AUC0−24), AUC0−24/MIC ratio, and f AUC0−24/MIC ratio were calculated using the PKSOLVER program (26). The pharmacodynamic parameters used to assure the efficacy of each antimicrobial Cmax/MIC for amikacin and gentamicin (27) and for fosfomycin this parameter is still not elucidated (28).
Murine Experimental Models
A previously described murine peritoneal sepsis model was used (22). Briefly, groups of C57BL/6J female mice were infected by intraperitoneal (ip) inoculation of 0.5 mL the Minimal Lethal Dose (MLD) of each strain. The inoculum concentrations inoculated were (log10 CFU/ml): 8.97 for VIM-1, 8.94 for VIM-1/DHA-1, 9.35 for OXA 48 plus CTX-M-15, and 8.38 for KPc29 KPC3/TEM-1 and SHV-11. Antimicrobial treatments were initiated 4 h post-inoculation and lasted 72 h. Mice were randomly included into six different therapeutic groups: (i) controls (untreated), (ii) fosfomycin, 500 mg/kg/8 h/ip (1.5 g/Kg/day), (iii) amikacin, 10 mg/kg/12 h/ip (100 mg/Kg/day), (iv) gentamicin, 5 mg/kg/12 h/ip (10 mg/Kg/day), (v) fosfomycin plus amikacin, for three of the strains: VIM-1 producer, VIM-1/DHA-1 producer, and OXA 48 plus CTX M 15 producer, (vi) fosfomycin plus gentamicin, for the KPC-3 TEM 1 plus SHV-11 producer strain. Antimicrobial dosages were chosen after PK/PD analysis to achieve the drugs optimal therapeutic regimen. Antimicrobials therapy lasted 72 h.
Immediately after mice death or sacrificed (sodium thiopental, ip) when the 72 h treatment was completed, aseptic thoracotomies were carried out, and blood samples were obtained for qualitative blood cultures; results were expressed as positive (≥1 cfu present in the plate) or negative. Spleens were aseptically extracted, weighed and homogenized in sterile saline (Stomacher 80; Tekmar Co., Cincinnati, OH, USA) before quantitative cultures (log10 cfu/g) in Columbia agar with 5% sheep blood plates (22).
Previous to the experimental model, in order to discard the toxicity of the treatments, groups of 5 healthy mice received the same dosages of antimicrobials alone or in combination, during 72 h, and were observed for any adverse reaction or body weight loss during 7 days.
Statistical Analysis
Mortality and positive blood cultures were expressed as percentages. Bacterial spleen concentrations (Log10 CFU/g) were expressed as means ± SD. Differences in bacterial concentrations in spleen were compared by analysis of variance (ANOVA) and the Dunnet and Tukey post-hoc-tests. Mortality and blood sterility rates between groups were compared by use of the two-tailed Fisher's-test. A P < 0.05 value was considered as statistically significant. The SPSS v22.0 was used (SPSS Inc., Chicago, IL, USA).
Results
In vitro Results
Antimicrobial Susceptibilities
The MICs of fosfomycin, amikacin and gentamicin for the four clinical strains by the broth microdilution method are shown in Table 1. VIM-1-, OXA-48 plus CTX-M-15-, and KPC-3-producing strains were resistant to fosfomycin, whereas by the dilution in agar method only KPC-3 producer was resistant. Only the KPC-3-producing strain was resistant to amikacin, and the VIM-1-producer strain to gentamicin. No differences in MIC values were obtained when MHB was supplemented with G-6-P.
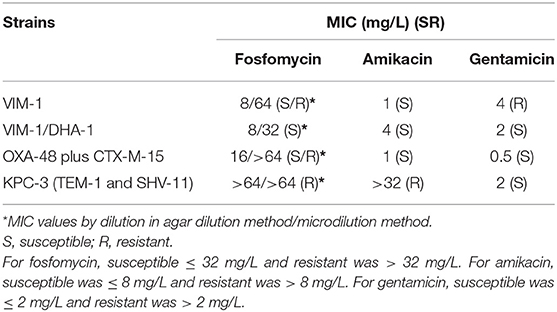
Table 1. MICs for the carbapenem-resistant Klebsiella pneumoniae strains used in the studies.
Time-Kill Curves
The results are shown in Table 2. Fosfomycin plus amikacin was bactericidal and synergistic against VIM-1-, VIM-1 plus DHA-1-, and OXA-48 plus CTX-M-15-producing strains. Fosfomycin plus gentamicin showed synergistic activity only against the KPC-3 producer.
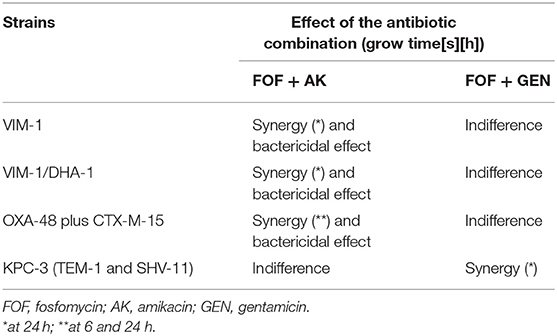
Table 2. Time-kill curves for fosfomycin (FOF) plus amikacin (AMK) and fosfomycin (FOF) plus gentamicin (GEN) against four clinical strains of carbapenemase producing strains.
Pharmacokinetics and Pharmacodynamics
Pharmacokinetic parameters of each antimicrobial are shown in Table 3. Pharmacodynamics profiles are shown in Table 4.
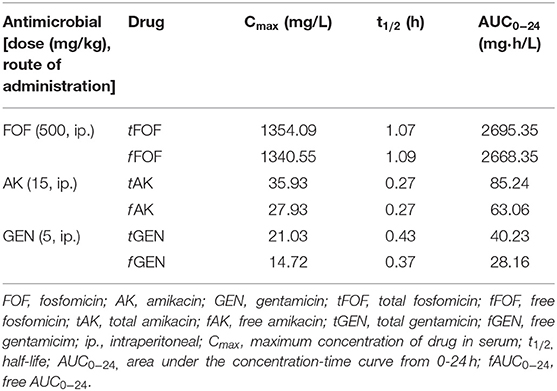
Table 3. Pharmacokinetic profiles of fosfomycin, amikacin and gentamicin in mice serum.
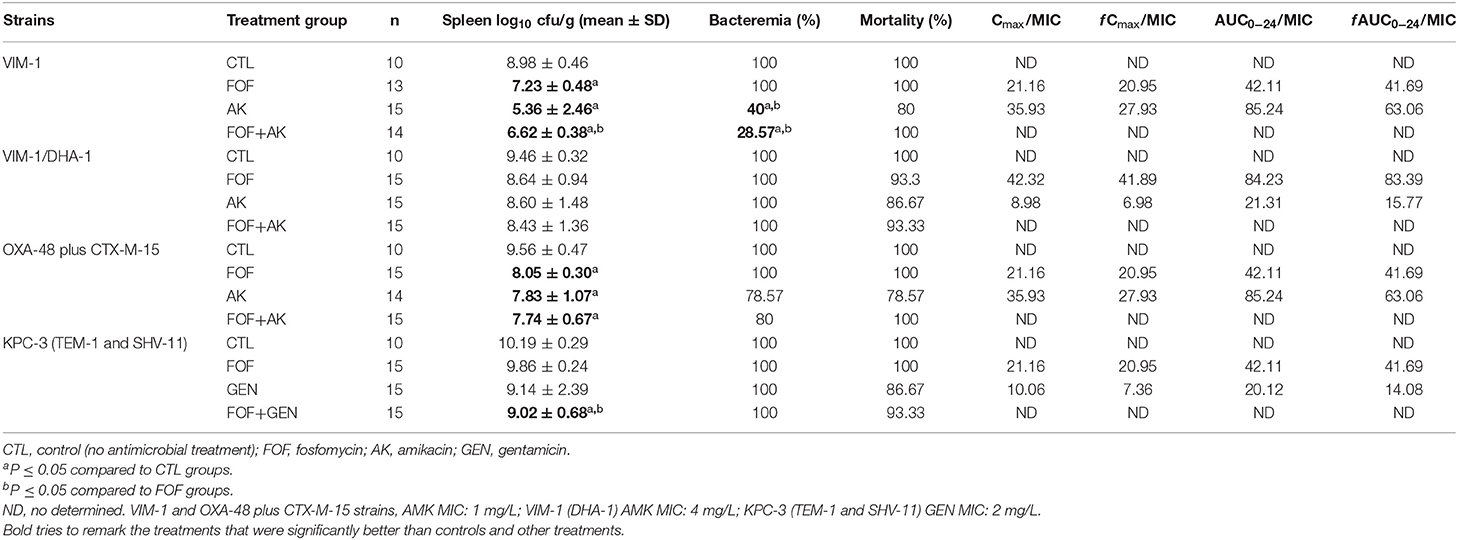
Table 4. In vivo efficacy and pharmacodynamics of fosfomycin, amikacin and gentamicin, alone and in combination, for the experimental peritoneal sepsis model.
In vivo Results: Peritoneal Sepsis Model
The efficacies of the antimicrobials are shown in Table 4. Mortality, bacterial clearance on spleen and bacteremia, are analyzed immediately after the death of mice or at the end of the experiment (72 h of treatment).
Mortality
Mortality in all control groups (non-treated) was 100% within the first 24 h post-infection. Antimicrobials alone or in combination did not significantly reduce mortality against any of the strains.
Bacterial Clearance From Spleen
Fosfomycin alone improved significantly the clearance of bacteria from spleen (CFU/g of tissue) compared with the control groups in mice infected with either VIM-1 (7.23 ± 0.48 vs. 8.98 ± 0.46) or OXA-48 plus CTX-M-15 (8.05 ± 0.30 vs. 9.56 ± 0.47). Amikacin alone was also better than controls in reducing bacterial concentration in spleen against VIM-1 or OXA-48 plus CTX-M-15 strains (5.36 ± 2.46 vs. 8.98 ± 0.46 and 7.83 ± 1.07 vs. 9.56 ± 0.47, respectively). Moreover, the combination of fosfomycin plus amikacin reduced the bacterial concentration compared with the controls for the strains, producing VIM-1 or OXA-48 plus CTX-M-15 (6.62 ± 0.38 vs. 8.98 ± 0.46 and 7.74 ± 0.67 vs. 9.56 ± 0.47, respectively) and fosfomycin plus gentamicin for the strain producing KPC-3 (9.02 ± 0.68 vs. 10.19 ± 0.29). Fosfomycin plus amikacin against the VIM-1 strain reduced the spleen bacterial concentration compared to fosfomycin alone (6.62 ± 0.38 vs. 7.23 ± 0.48). None of the antimicrobials tested alone or in combination reduced the bacterial concentration in spleen against the strain producing VIM-1 (DHA-1).
Bacteremia
Amikacin alone and in combination with fosfomycin showed a significant reduction sterilizing blood cultures of mice infected with the VIM-1 producer (40 and 28.57% vs. 100%, respectively).
Discussion
The results of this study show that the combination of fosfomycin plus aminoglycosides (amikacin and gentamycin) shows efficacy in terms of bacterial clearance from tissue in animals infected with the strains VIM-1, OXA-48 plus CTX-M-15 and KPC-3 producers. Bacteraemia was only significantly reduced against the VIM-1 producer strain with fosfomycin plus amikacin. None of the treatments tested improved mortality compared to those from control groups for any of the studied strains. Finally, there was no significant efficacy in terms of bacterial clearance either from tissue and blood or in the survival in the animals infected with the strain producing VIM-1 (DHA-1). The synergistic activity found between fosfomycin and amikacin is in accordance with other in vitro studies that found a synergistic and bactericidal effect with this combination against fosfomycin susceptible clinical KPC-producing K. pneumoniae isolates, regardless the bacteria susceptibility to amikacin (15).
Fosfomycin dosage used in this study 500 mg/kg every 8 h was chosen because it produces a mean peak serum fosfomycin concentration of 1354.09 ± 217.99 mg/L and a mean AUC0−24 of 2695.35 ± mg*h/L, above to those obtained in humans after a 4 h infusion of 4 g every 8 h (peak of 123 + 16 mg/L; AUC0–24 of 600 mg*h/L) and above dosages proven to be effective in several murine experimental studies using, KPC-3- or OXA-48-producing Escherichia coli (29), Enterococcus faecalis (30). Moreover, with this dose we achieved the PK/PD index that best predicted efficacy for K. pneumoniae strains base on model fit and regression analysis, an unbound AUC0−14/MIC ratio of 41.69 or 83.39, depending on the strains (31). Nevertheless, fosfomycin monotherapy was only effective in reducing tissue concentrations against VIM-1- and OXA-48 plus CTX-M-15-producing strains, but not with the other two tested strains, not even with the fosfomycin susceptible VIM-1/DHA-1-producing strain. These results could be in the line of the described uncertainty as whether fosfomycin monotherapy is efficacious in the treatment of systemic infections other than complicated urinary tract infection and/or acute pyelonephritis (32).
Although the pharmacokinetic/pharmacodynamic target Cmax/MIC ≥ 8 (33) was achieved for all the tested strains, the activity of aminoglycosides (amikacin and gentamicin) in monotherapy was also limited, reducing amikacin only bacterial spleen concentration compared to control groups in mice infected with VIM-1 or OXA-48 plus CTX-M-15 strains, for which the Cmax/MIC value for both strains was = 35.93, predictor of efficacy was achieved. Only in the case of animals infected with the VIM-1 producer strain, amikacin reduced the mortality compared to controls mice. Nevertheless, amikacin is only used alone to treat urinary tract infections (34).
Fosfomycin in combination with amikacin demonstrated in vitro a synergistic effect against the VIM-1-, VIM-1/DHA-1-, and OXA-48 plus CTX-M-15-producing strains, while the combination with gentamycin was the only with synergistic activity against KPC-3-producing strain. These results are in accordance with other studies showing synergy of this combination against KPC-producing K. pneumoniae strains. An in vitro study in KPC-producing K. pneumoniae found a synergistic and bactericidal effect with the combination of fosfomycin with amikacin against fosfomycin susceptible isolates, even when the causative bacteria are resistant to amikacin (15). Other in vitro studies have showed fosfomycin plus amikacin to have additive and synergistic effects against other pathogens such as P. aeruginosa and methicillin-resistant Staphylococcus epidermidis and IMP-8 metallo-β-lactamase-producing K. oxytoca (35, 36).
With regard to the in vivo efficacy of fosfomycin plus amikacin in the experimental murine peritoneal sepsis model, the combination was better than the control groups taking into account the bacterial clearance from spleen against the VIM-1 and the OXA-48 plus CTX-M-15 strain. Moreover, this combination reduced bacteremia compared both with control and fosfomycin groups against the VIM-1 strain. Nevertheless, and besides both antimicrobials achieving the pharmacodynamic values described as optimal for efficacy, f AUC0−24/MIC and Cmax/MIC, no activity is found against the VIM-1/DHA-1 strain, in contrast with the synergy observed in the time-kill studies. Also, and in accordance with the in vitro results fosfomycin plus gentamicin reduced significantly bacteria from spleen compared with control and fosfomycin groups against the KPC-3 strain. Finally, none of the combinations reduced the mortality against any of the tested strains. To our best knowledge, there are no experimental studies in which fosfomycin in combination with amikacin and gentamicin has been evaluated. There are experimental studies that have evaluated fosfomycin in combination with ceftazidime-avibactam combination against MDR P. aeruginosa finding this combination better than either drug alone (37). In other study, they evaluated in a surgical wound infection in mice the activity of fosfomycin in combination with rifampin and tigecycline against Enterococcus faecium and methicillin-resistant Staphylococcus aureus clinical isolates, finding the combination as an alternative treatment to control skin infection (38). Also, Berleur et al., found that fosfomycin plus temocillin reduced bacterial counts in a murine peritonitis model against E. coli strains producing KPC-3 or OXA-48-type carbapenemases (29).
In summary, our results showed that the fosfomycin plus aminoglycosides combination has a low and dissimilar efficacy in the treatment of severe infections, such as a peritoneal sepsis infection, caused by different KPC-KP strains, reducing only the tissue bacterial concentration against three of the strains and only decreasing bacteremia against the VIM-1 producer strain. Moreover, none of the combinations improved the survival in the infection by any of the KPC-KP strains. Also no activity was found in reducing tissue bacterial concentration or in decreasing bacteremia or mortality in the infection by VIM-1/DHA-1 strain. Because of the lack of available alternatives for these kinds of carbapenemase-producing strains, the fosfomycin plus aminoglycosides combination might be considered for further evaluation against other kind of infections. Overall, the results of the present study suggest that the efficacy of fosfomycin plus aminoglycosides depends on the class of carbapenemase produced by K. pneumoniae.
Data Availability Statement
The raw data supporting the conclusions of this article will be made available by the authors, without undue reservation.
Ethics Statement
In vivo experiments were approved by the Committee on the Ethics of Animal Experiments of the University Hospital Virgen Macarena, Seville, Spain (CI 1961).
Author Contributions
MP-I planned and coordinated the experiments, analyzed the results, and written the manuscript. GL-H and TC-C had performed the in vitro and in vivo experiments. CD and JdP had performed the antibiotic concentrations studies by HPLC tandem mass spectrometry (LC-MS/MS). JR-B, ÁP, and JP had reviewed the manuscript and the experiments. MC obtained the funds to perform the studies and wrote the project, contributed to the performance of the in vitro experiment, and reviewed the results and the manuscript. All authors contributed to the article and approved the submitted version.
Funding
This study was supported by the Consejería de Salud of the Junta de Andalucía (PI-0622-2012) and supported by Plan Nacional de I+D+i 2013-2016 and Instituto de Salud Carlos III, Subdirección General de Redes y Centros de Investigación Cooperativa, Ministerio de Economía, Industria y Competitividad, Spanish Network for Research in Infectious Diseases (REIPI, RD16/0016/0001, RD16/0016/0009)—co-financed by European Development Regional Fund A way to achieve Europe, Operative program Intelligent Growth 2014-2020. MP-I is a researcher belonging to the program Nicolás Monardes (C1-0038-2019), Servicio Andaluz de Salud, Junta de Andalucía, Spain. TC-C was supported by the V Plan Propio of the University of Seville with a postdoctoral contract as research personnel in training. GL-H has a grant from the Ministerio de Ciencia, Innovación y Universidades, Instituto de Salud Carlos III, cofinanced by the European Development Regional Fund (A Way to Achieve Europe) and by the Spanish Network for Research in Infectious Diseases (Grant REIPI RD16/0016/0009).
Conflict of Interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
References
1. Poirel L, Kieffer N, Nordmann P. In vitro evaluation of dual carbapenem combinations against carbapenemase-producing Enterobacteriaceae. J Antimicrob Chemother. (2016) 71:156–61. doi: 10.1093/jac/dkv294
2. Mandell D, Bennet's. Principles and Practice of Infectious Disease. 9th ed. Amterdam: Elsevier (2020).
3. de Oliveira MS, de Assis DB, Freire MP, Boas do Prado GV, Machado AS, Abdala E, et al. Treatment of KPC-producing Enterobacteriaceae: suboptimal efficacy of polymyxins. Clin Microbiol Infect. (2015) 21:179.e1–7. doi: 10.1016/j.cmi.2014.07.010
4. Peleg AY, Hooper DC. Hospital-acquired infections due to gram-negative bacteria. N Engl J Med. (2010) 362:1804–13. doi: 10.1056/NEJMra0904124
5. van Duin D, Doi Y. The global epidemiology of carbapenemase-producing Enterobacteriaceae. Virulence. (2017) 8:460–9. doi: 10.1080/21505594.2016.1222343
6. Kilic U, Koroglu M, Olmez M, Altindis M. Investigation of the in vitro effectiveness of aztreonam/avibactam, colistin/apramycin, and meropenem/apramycin combinations against carbapenemase-producing, extensively drug-resistant Klebsiella pneumoniae strains. Microbial Drug Resist. (2020) 26:1291–7. doi: 10.1089/mdr.2019.0498
7. Erdem F, Abulaila A, Aktas Z, Oncul O. In vitro evaluation of double carbapenem and colistin combinations against OXA-48, NDM carbapenemase-producing colistin-resistant Klebsiella pneumoniae strains. Antimicrob Resist Infect Control. (2020) 9:70. doi: 10.1186/s13756-020-00727-4
8. Durante-Mangoni E, Andini R, Zampino R. Management of carbapenem-resistant Enterobacteriaceae infections. Clin Microbiol Infect. (2019) 25:943–50. doi: 10.1016/j.cmi.2019.04.013
9. Sheu CC, Chang YT, Lin SY, Chen YH, Hsueh PR. Infections caused by carbapenem-resistant Enterobacteriaceae: an update on therapeutic options. Front Microbiol. (2019) 10:80. doi: 10.3389/fmicb.2019.00080
10. Rodriguez-Bano J, Gutierrez-Gutierrez B, Machuca I, Pascual A. Treatment of infections caused by extended-spectrum-beta-lactamase-, ampC-, and carbapenemase-producing Enterobacteriaceae. Clin Microbiol Rev. (2018) 31:e00079–17. doi: 10.1128/CMR.00079-17
11. Bonomo RA, Burd EM, Conly J, Limbago BM, Poirel L, Segre JA, et al. Carbapenemase-producing organisms: a global scourge. Clin Infect Dis. (2018) 66:1290–7. doi: 10.1093/cid/cix893
12. Wistrand-Yuen P, Olsson A, Skarp KP, Friberg LE, Nielsen EI, Lagerback P, et al. Evaluation of polymyxin B in combination with 13 other antibiotics against carbapenemase-producing Klebsiella pneumoniae in time-lapse microscopy and time-kill experiments. Clin Microbiol Infect. (2020) 26:1214–21. doi: 10.1016/j.cmi.2020.03.007
13. Hagihara M, Kato H, Yamashita R, Soda M, Watanabe H, Sakanashi D, et al. In vivo study assessed meropenem and amikacin combination therapy against carbapenem-resistant and carbapenemase-producing Enterobacteriaceae strains. J Infect Chemother. (2020) 26:1–7. doi: 10.1016/j.jiac.2019.10.014
14. Tattevin P, Dinh A, Ghout I, Mouton W, Verdier MC, Laurent F, et al. Efficacy of generic meropenem products in combination with colistin in carbapenemase-producing Klebsiella pneumoniae experimental osteomyelitis. Int J Antimicrob Agents. (2020) 56:106152. doi: 10.1016/j.ijantimicag.2020.106152
15. Yu W, Shen P, Bao Z, Zhou K, Zheng B, Ji J, et al. In vitro antibacterial activity of fosfomycin combined with other antimicrobials against KPC-producing Klebsiella pneumoniae. Int J Antimicrob Agents. (2017) 50:237–41. doi: 10.1016/j.ijantimicag.2017.03.011
16. Souli M, Galani I, Boukovalas S, Gourgoulis MG, Chryssouli Z, Kanellakopoulou K, et al. In vitro interactions of antimicrobial combinations with fosfomycin against KPC-2-producing Klebsiella pneumoniae and protection of resistance development. Antimicrob Agents Chemother. (2011) 55:2395–7. doi: 10.1128/AAC.01086-10
17. Cremieux AC, Dinh A, Nordmann P, Mouton W, Tattevin P, Ghout I, et al. Efficacy of colistin alone and in various combinations for the treatment of experimental osteomyelitis due to carbapenemase-producing Klebsiella pneumoniae. J Antimicrob Chemother. (2019) 74:2666–75. doi: 10.1093/jac/dkz257
18. Pachon-Ibanez ME, Labrador-Herrera G, Cebrero-Cangueiro T, Diaz C, Smani Y, Del Palacio JP, et al. Efficacy of colistin and its combination with rifampin in vitro and in experimental models of infection caused by carbapenemase-producing clinical isolates of Klebsiella pneumoniae. Front Microbiol. (2018) 9:912. doi: 10.3389/fmicb.2018.00912
19. CLSI. Performance Standars for Antimicrobial Susceptibility Tests. 29th ed. CLSI supplement M100. Wayne, PA: Clinical and Laboratory Standards Institute (2019).
20. (EUCAST) ECoAST. European Society of Clinical Microbiology and Infectious Diseases. Clinical Breakpoints. EUCAST (2019).
21. Pournaras S, Vrioni G, Neou E, Dendrinos J, Dimitroulia E, Poulou A, et al. Activity of tigecycline alone and in combination with colistin and meropenem against Klebsiella pneumoniae carbapenemase (KPC)-producing Enterobacteriaceae strains by time-kill assay. Int J Antimicrob Agents. (2011) 37:244–7. doi: 10.1016/j.ijantimicag.2010.10.031
22. Parra-Millan R, Vila-Farres X, Ayerbe-Algaba R, Varese M, Sanchez-Encinales V, Bayo N, et al. Synergistic activity of an OmpA inhibitor and colistin against colistin-resistant Acinetobacter baumannii: mechanistic analysis and in vivo efficacy. J Antimicrob Chemother. (2018) 73:3405–12. doi: 10.1093/jac/dky343
23. US I. Committee on Care and Use of Laboratory Animals. Guide for the Care and Use of Laboratory Animals. NIH publication Bethesda, MD: US Dept of Health and Human Services, Public Health Service pv. (2011).
24. Gobin P, Lemaitre F, Marchand S, Couet W, Olivier JC. Assay of colistin and colistin methanesulfonate in plasma and urine by liquid chromatography-tandem mass spectrometry. Antimicrob Agents Chemother. (2010) 54:1941–8. doi: 10.1128/AAC.01367-09
25. Waters NJ, Jones R, Williams G, Sohal B. Validation of a rapid equilibrium dialysis approach for the measurement of plasma protein binding. J Pharm Sci. (2008) 97:4586–95. doi: 10.1002/jps.21317
26. Zhang Y, Huo M, Zhou J, Xie S. PKSolver: an add-in program for pharmacokinetic and pharmacodynamic data analysis in Microsoft Excel. Comput Methods Programs Biomed. (2010) 99:306–14. doi: 10.1016/j.cmpb.2010.01.007
27. Bland CM, Pai MP, Lodise TP. Reappraisal of contemporary pharmacokinetic and pharmacodynamic principles for informing aminoglycoside dosing. Pharmacotherapy. (2018) 38:1229–38. doi: 10.1002/phar.2193
28. Merino-Bohorquez V, Docobo-Perez F, Sojo J, Morales I, Lupion C, Martin D, et al. Population pharmacokinetics and pharmacodynamics of fosfomycin in non-critically ill patients with bacteremic urinary infection caused by multidrug-resistant Escherichia coli. Clin Microbiol Infect. (2018) 24:1177–83. doi: 10.1016/j.cmi.2018.02.005
29. Berleur M, Guerin F, Massias L, Chau F, Poujade J, Cattoir V, et al. Activity of fosfomycin alone or combined with temocillin in vitro and in a murine model of peritonitis due to KPC-3- or OXA-48-producing Escherichia coli. J Antimicrob Chemother. (2018) 73:3074–80. doi: 10.1093/jac/dky283
30. Oliva A, Furustrand Tafin U, Maiolo EM, Jeddari S, Betrisey B, Trampuz A. Activities of fosfomycin and rifampin on planktonic and adherent Enterococcus faecalis strains in an experimental foreign-body infection model. Antimicrob Agents Chemother. (2014) 58:1284–93. doi: 10.1128/AAC.02583-12
31. Lepak AJ, Zhao M, VanScoy B, Taylor DS, Ellis-Grosse E, Ambrose PG, et al. In vivo pharmacokinetics and pharmacodynamics of ZTI-01 (fosfomycin for injection) in the neutropenic murine thigh infection model against Escherichia coli, Klebsiella pneumoniae, and Pseudomonas aeruginosa. Antimicrob Agents Chemother. (2017) 61:e00476–17. doi: 10.1128/AAC.00476-17
32. Doi Y. Treatment options for carbapenem-resistant gram-negative bacterial infections. Clin Infect Dis. (2019) 69(Suppl. 7):S565–75. doi: 10.1093/cid/ciz830
33. Logre E, Enser M, Tanaka S, Dubert M, Claudinon A, Grall N, et al. Amikacin pharmacokinetic/pharmacodynamic in intensive care unit: a prospective database. Ann Intensive Care. (2020) 10:75. doi: 10.1186/s13613-020-00685-5
34. Jayakumar I, Mathaiyan J, Mandal J, Deepanjali S, Sreenivasan SK. Impact of therapeutic drug monitoring on once-daily regimen of amikacin in patients with urinary tract infection: a prospective observational study. Ther Drug Monit. (2020) 42:841–7. doi: 10.1097/FTD.0000000000000800
35. Vergara-Lopez S, Dominguez MC, Conejo MC, Pascual A, Rodriguez-Bano J. Prolonged treatment with large doses of fosfomycin plus vancomycin and amikacin in a case of bacteraemia due to methicillin-resistant Staphylococcus epidermidis and IMP-8 metallo-beta-lactamase-producing Klebsiella oxytoca. J Antimicrob Chemother. (2015) 70:313–5. doi: 10.1093/jac/dku341
36. Cai Y, Fan Y, Wang R, An MM, Liang BB. Synergistic effects of aminoglycosides and fosfomycin on Pseudomonas aeruginosa in vitro and biofilm infections in a rat model. J Antimicrob Chemother. (2009) 64:563–6. doi: 10.1093/jac/dkp224
37. Papp-Wallace KM, Zeiser ET, Becka SA, Park S, Wilson BM, Winkler ML, et al. Ceftazidime-avibactam in combination with fosfomycin: a novel therapeutic strategy against multidrug-resistant Pseudomonas aeruginosa. J Infect Dis. (2019) 220:666–76. doi: 10.1093/infdis/jiz149
38. Simonetti O, Morroni G, Ghiselli R, Orlando F, Brenciani A, Xhuvelaj L, et al. In vitro and in vivo activity of fosfomycin alone and in combination with rifampin and tigecycline against Gram-positive cocci isolated from surgical wound infections. J Med Microbiol. (2018) 67:139–43. doi: 10.1099/jmm.0.000649
Keywords: fosfomycin, aminoglycosides, in vivo, time kill curves, carbapenemase-producing Klebsiella pneumoniae
Citation: Cebrero-Cangueiro T, Labrador-Herrera G, Pascual Á, Díaz C, Rodríguez-Baño J, Pachón J, del Palacio JP, Pachón-Ibáñez ME and Conejo MC (2021) Efficacy of Fosfomycin and Its Combination With Aminoglycosides in an Experimental Sepsis Model by Carbapenemase-Producing Klebsiella pneumoniae Clinical Strains. Front. Med. 8:615540. doi: 10.3389/fmed.2021.615540
Received: 09 October 2020; Accepted: 01 March 2021;
Published: 26 March 2021.
Edited by:
Spyros Pournaras, National and Kapodistrian University of Athens, GreeceReviewed by:
Georgia Vrioni, National and Kapodistrian University of Athens, GreeceKonstantina Dafopoulou, University Hospital of Larissa, Greece
Copyright © 2021 Cebrero-Cangueiro, Labrador-Herrera, Pascual, Díaz, Rodríguez-Baño, Pachón, del Palacio, Pachón-Ibáñez and Conejo. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: María E. Pachón-Ibáñez, bXBhY2hvbi1pYmlzQHVzLmVz
†These authors have contributed equally to this work