 Bárbara Lobão
Bárbara Lobão Diana Lourenço
Diana Lourenço Ana Giga
Ana Giga Pedro Mendes-Bastos
Pedro Mendes-Bastos- 1Instituto Português de Reumatologia, Lisboa, Portugal
- 2Centro Hospitalar de Setúbal, Setúbal, Portugal
- 3Janssen Portugal, Oeiras, Portugal
- 4Dermatology Center, Hospital CUF Descobertas, Lisboa, Portugal
Introduction: Psoriasis (PsO) is a chronic skin condition driven by immune mediators like TNFα, INFγ, IL-17, and IL-23. Psoriatic arthritis (PsA) can develop in PsO patients. Although psoriatic lesions may apparently resolve with therapy, subclinical cutaneous inflammation may persist. The role of tissue-resident memory T-cells (TRM), and regulatory T cells (Tregs) that also contribute to chronic inflammation are being explored in this context. This systematic review explores TRM and Tregs in psoriatic disease (PsD) and its progression.
Methods: A systematic review, following Preferred Reporting Items for Systematic Reviews and Meta-Analyses (PRISMA) guidelines, was performed using Pubmed® and Web of Science™ databases on June 3rd 2023, using patient/population, intervention, comparison, and outcomes (PICO) criteria limited to the English language.
Results: A total of 62 reports were identified and included. In PsO, chronic inflammation is driven by cytokines including IL-17 and IL-23, and cellular mediators such as CD8+ and CD4+ T cells. TRM contributes to local inflammation, while Tregs may be dysfunctional in psoriatic skin lesions. Secukinumab and guselkumab, which target IL-17A and the IL-23p19 subunit, respectively, have different effects on CD8+ TRM and Tregs during PsO treatment. Inhibition of IL-23 may provide better long-term results due to its impact on the Treg to CD8+ TRM ratio. IL-23 may contribute to inflammation persisting even after treatment. In PsA, subclinical enthesitis is perceived as an early occurence, and Th17 cells are involved in this pathogenic process. Recent EULAR guidelines highlight the importance of early diagnosis and treatment to intercept PsA. In PsA, CD8+ TRM cells are present in synovial fluid and Tregs are reduced in peripheral blood. The progression from PsO to PsA is marked by a shift in immune profiles, with specific T-cells subsets playing key roles in perpetuating inflammation. Early intervention targeting TRM cells may hold promising, but clinical studies are limited. Ongoing studies such as IVEPSA and PAMPA aim to improve our knowledge regarding PsA interception in high-risk PsO patients, emphasizing the need for further research in this area.
Conclusion: Early intervention is crucial for PsO patients at high risk of PsA; T cells, particularly type 17 helper T cells, and CD8+ cells are key in the progression from PsO-to-PsA. Early targeting of TRM in PsD shows promise but more research is needed.
1 Introduction
Psoriasis (PsO) is a chronic inflammatory skin condition characterized by thickened red plaques and silvery lamellar scales, primarily affecting the scalp, trunk and extensor surfaces of the limbs (1). The pathogenesis of PsO involves both environmental and genetic factors, with communication between various immune cell types via cytokines such as tumor necrosis factor alpha (TNFα), interferon gamma (IFN-γ), IL-17, IL-22, and IL-23, leading to the establishment of a self-sustaining inflammatory cycle (2, 3). Thus, PsO is a recurring immune-mediated skin disorder characterized by epidermal hyperplasia and the presence of extensive dermal inflammatory infiltrates, which are recognized as the key histologic features of psoriatic lesions (4). PsO is a T cell-mediated inflammatory disease and accumulating evidence suggests that T-helper (Th) cells play a pivotal role in its pathogenesis. Various subtypes of Th cells are involved in inflammatory responses (5). In psoriasis, intraepidermal T cells are predominantly CD8+ and represent key effector cells driving disease. These pathogenic cells produce IL-17 and the neutralization of CD8+T cells effectively prevents psoriasis development in vivo (6). Dermal CD4+ T cells produce inflammatory cytokines, such as IL-17A, IL-22, and IFN-γ, and are regarded as the main pathogenic T cell subpopulation (6).
One of the primary challenges encountered in PsO management lies in the reoccurrence of lesions at previously affected anatomical sites, either during inadequate treatment of the disease or after discontinuation of treatment after resolution of lesions. Existing evidence substantiates the idea of localized immune “memory” involving specific T cell populations that persists within cleared skin (7–11).
Psoriatic arthritis (PsA) develops in up to 30% of patients with PsO (12). Furthermore, over 90% PsA patients initially experience arthritis in the context of pre-existing PsO (13).
PsO condition runs a chronic course with recurrent disease flares and infrequent periods of prolonged drug-free remission (14). Immune inflammatory pathways associated with IL-23, IL-17 and TNF have been identified as also being relevant in PsA, which has led to the development of therapeutic agents targeting these cytokines (15). However, the great variability of response across different therapies suppressing specific inflammatory pathways is not yet fully understood (14, 15).
Strong evidence suggests that the initial phase of PsA is closely associated with early enthesitis, with subsequent inflammation spreading to involve the synovium (16). Studies have shown that many patients with PsO who have not yet experienced musculoskeletal complaints bear a significantly greater burden of subclinical articular inflammation when compared to healthy individuals (17, 18). In the progression from PsO to PsA there is a notable correlation with the development of synovitis and tenosynovitis, again tied to the synovio-enthesal complex (19). Growing evidence indicates that the pathogenesis of PsA may be rooted in autoimmune mechanisms. This is supported by significant links to class I human leukocyte antigen alleles, the development of ectopic lymphoid structures in synovial tissues containing T and B cell clusters, and the presence of clonally proliferating CD8+ T cells in both synovial tissue and fluid (20, 21). Nevertheless, the precise immunological mechanisms responsible for reducing immune tolerance and triggering the transition from PsO to PsA remain largely elusive (12, 22).
The recent discovery of a specialized subset of T cells known as tissue-resident memory T cells (TRM cells) holds great potential for understanding the mechanism underlying of some chronic immune-mediated inflammatory diseases (9–11, 14). TRM are a subset of memory T cells that play a crucial role in providing immune surveillance in specific tissues. They are identified by expression of CD69, a core signature marker of TRM cells, and CD103 (2, 11, 23, 24). Under normal physiological conditions, TRM cells reside in peripheral tissue sites (25).
Unlike other T cells, most TRM cells permanently reside in tissues and do not circulate in the bloodstream. However, recent studies suggest that a small fraction of TRM cells can exit tissues and persist in the blood, migrate to distant tissues or lymph nodes, or transform into other types of memory cell (26–28). TRM can either differentiate from circulating precursors expressing the killer cell lectin like receptor G1 (KLRG1), or from central memory T cells (TCM) and effector memory T cells (TEM) in peripheral tissues. TEM migrates to inflamed peripheral tissues, where they promptly display effector functions, whereas TCM migrate to T cells regions within secondary lymphoid organs. There, they subsequently transform into TEM and carry out effector functions. Within the local microenvironment, signals and cytokines originating from the tissue, such as transforming growing factor-beta 1 (TGFβ1), have the capacity to drive the tissue residency program in precursor cells (29–33). TRM are primarily found in epithelial tissues such as the skin, lungs, and gut, where they functions to protect against infections and cancer. However, as previously mentioned, they may also play a role in disease through driving chronic inflammation (34, 35). TRM have been implicated in heightened responses within inflamed environments due to their potential for crosstalk with other immune cells. The production of cytokines that recruit immune cells into inflamed tissues and the expression of genes involved in recruiting innate immune cells support the hypothesis that TRM cells play a critical role in driving inflammation, contributing to the immune-inflammatory state (31, 36).
Human TRM cells in the skin express the skin-homing chemokine receptors CCR4 and CCR10 as well as the tissue homing molecules CD103 and cutaneous lymphocyte antigen (CLA) (37). Skin’s TRM cells actively search for pathogens by migrating within the epidermal compartment, utilizing several dynamic dendritic projections to squeeze between keratinocytes (38). After skin immunization, it has been suggested that the immune system may distribute antigen-reactive T cells both to the lymph nodes, as TCM cells, and to the peripheral tissues, as TRM cells (39).
While earlier studies primarily focused on CD8+ TRM cells, it is essential to note that CD4+ TRM cells also exist (40). CD4+ T cells assist in fostering the development of cytotoxic memory CD8+ T cells following infection (2).
A growing body of evidence suggests that TRM cells can be found in the inflamed joints of patients with PsA (41). These cells are believed to play a crucial role in driving chronic inflammation and contributing to flares of this condition, similar to their role in driving recurrence of psoriatic skin lesions (14, 35, 42). However, T cells and their role in disease pathogenesis in entheses are poorly characterized. Nonetheless. healthy human entheseal CD4+ and CD8+ T cells express high levels of transcripts suggestive of tissue residency (41).
In addition to TRM cells, regulatory T-cells (Tregs) represent another T-cell population that has gained attention in PsD. Tregs are a specialized immunosuppressive lineage of adaptative lymphocytes that have been shown to exert systemic effects in protecting tissues from. Tregs can suppress the activities of other immune cells mainly by secretion of IL-10 and TGFβ (43).
However, the literature regarding Tregs in patients with PsD is somewhat limited. Treg abnormalities such as decreased expression of CD39 and CD74, increased expression of IL-6Rα21, reduced suppressive functional capacity, and enhanced propensity to differentiate into cells that produce interleukin (IL)-17, have been observed in PsO patients. Only a few studies have investigated the role of Tregs in the pathogenesis of PsA and in-depth characterization of intra-articular Tregs is lacking (44). This systematic review aimed to assess the existing evidence regarding the role of TRM and Tregs in PsO and PsA, and also in the progression from PsO to PsA.
2 Methods
This review adheres to the guidelines set by the Preferred Reporting Items for Systematic reviews and Meta-Analysis (PRISMA) (45). The PubMed® and the Web of Science™ databases were searched on June 3rd, 2023, using the strings “(psoriatic arthritis OR psoriasis OR nails OR enthesitis OR peripheral arthritis OR axial disease OR dactylitis OR scalp) AND tissue-resident memory T-cells”; psoriatic arthritis AND (regulatory T cells NOT tissue-resident memory T cells)”; “psoriasis AND psoriatic disease AND disease interception.” The search was restricted to papers written in English and involving human subjects. Results from these searches were combined and duplicate entries were removed. Two authors independently reviewed the abstracts for all publications identified, and in case of discrepancies, a third author made the final determination regarding the publication for this review. Reviews, case reports, and case series were excluded from consideration.
3 Results
A comprehensive flow chart with the results of the literature search is illustrated in Figure 1. Initially, 443 records were exported from PubMed and Web of Sciences databases. Following elimination of duplicates and records lacking available abstracts, a total of 288 publications were retained. Of these, 220 were deemed ineligible for inclusion based on the application of selection criteria (145 by title, 75 by abstract review). Eighteen additional studies were identified through reference tracking, and citation. Ultimately, 62 studies were included in this review as summarized in Table 1.
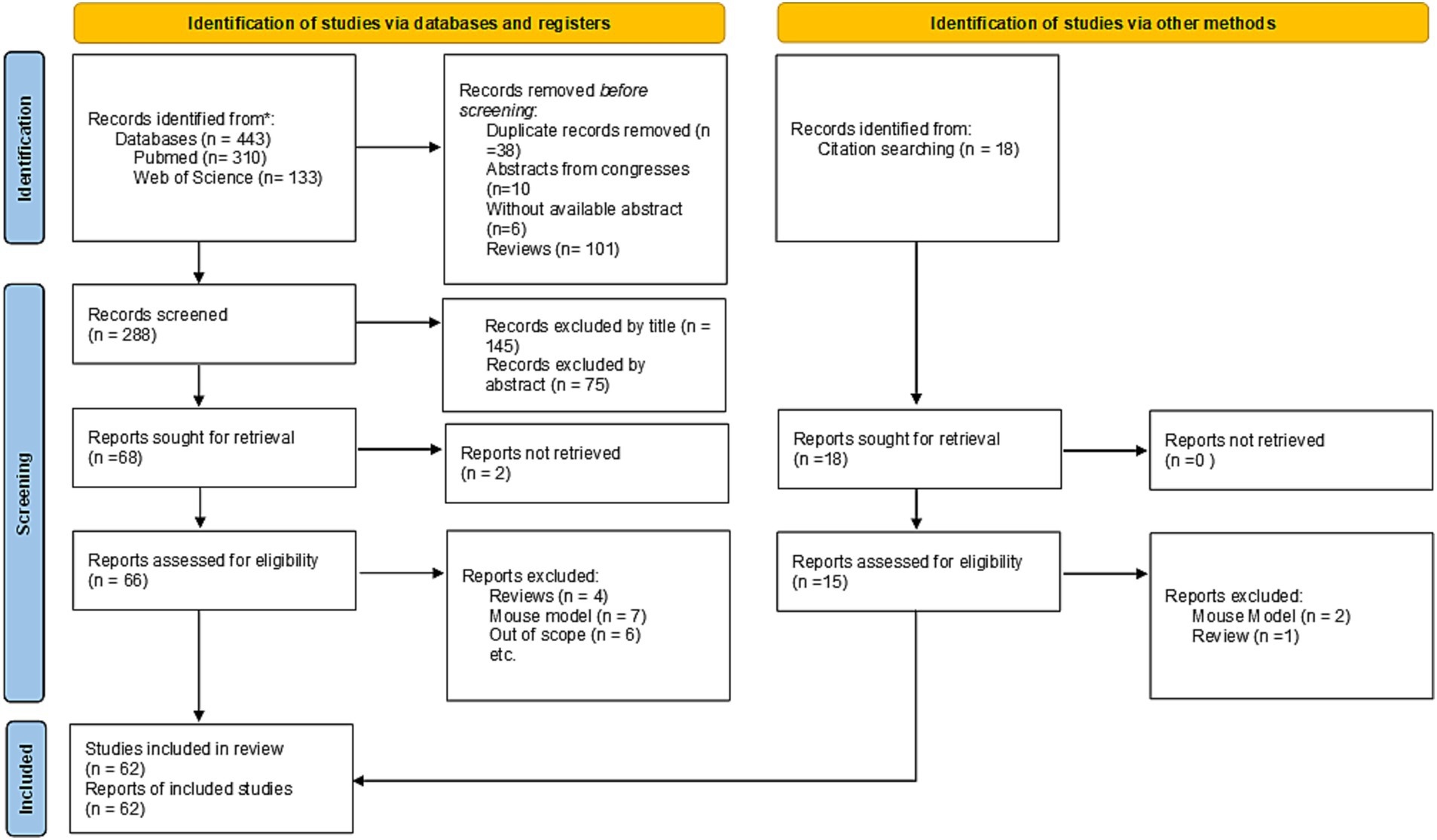
Figure 1. PRISMA flow for systematic review of the literature adapted from Page (45).
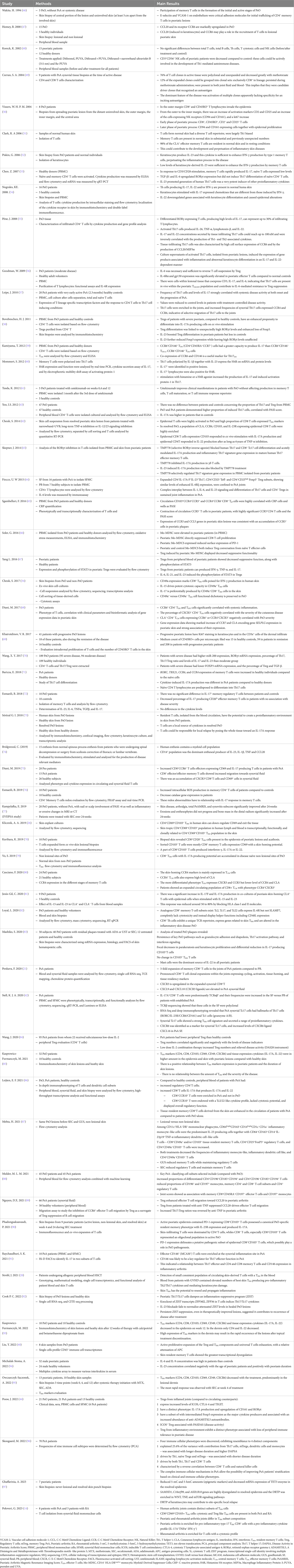
Table 1. Summary of the studies included in the systematic review.
3.1 Psoriasis
Chronic inflammation in PsO is characterized by tissue injury caused by activated immune cells, namely lymphocytes, which, through the production of a distinctive set of cytokines, contribute to disease pathogenesis (Figure 2) (55). The IL-23/IL-17 axis plays a central role in the immunopathogenesis of PsO by stimulating keratinocyte hyperproliferation and perpetuating T cell-mediated inflammation (55).
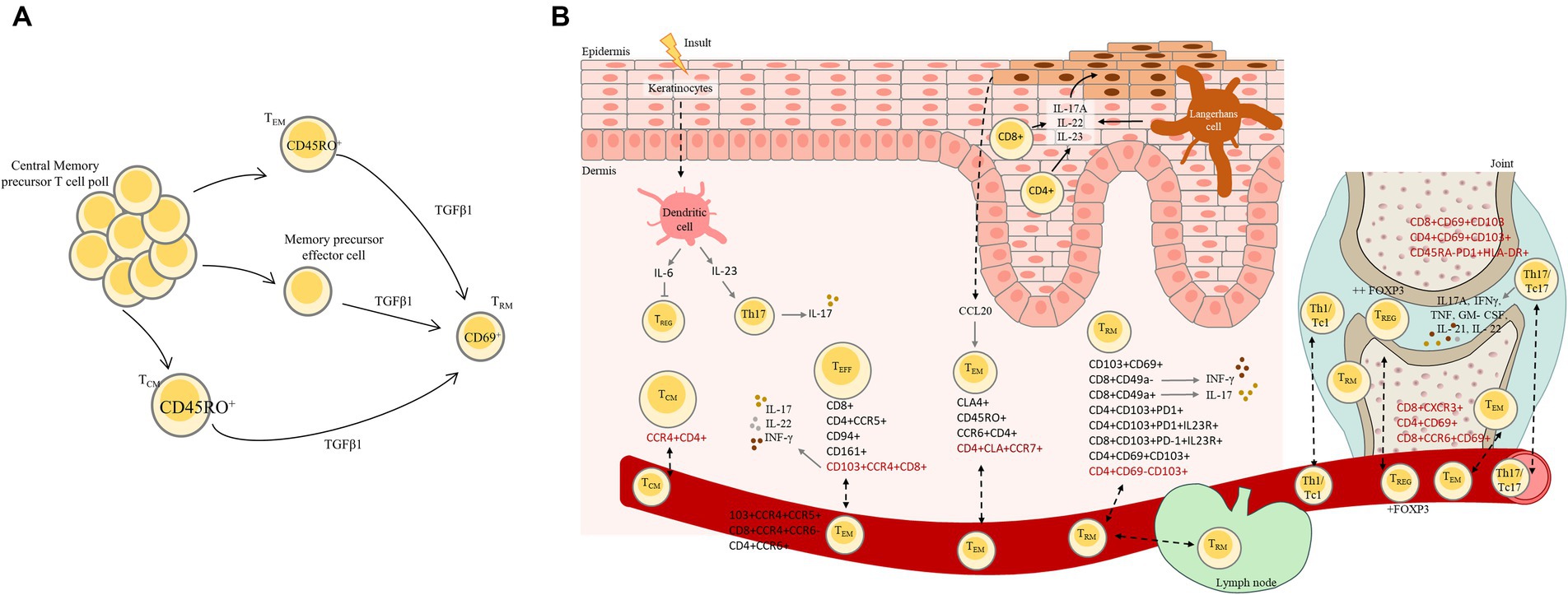
Figure 2. (A) Resident-tissue T cell differentiation from central memory precursor T cell pool. Central memory T cells are typically found in the bloodstream and lymphatic system. In psoriasis, these central memory T cells can undergo differentiation process and transform into resident tissue T cells (TRM) in the skin mainly through the action of TGFβ1. This differentiation process involves changes in the expression of surface markers such as CD45RO that is predominantly expressed on memory T cells and allow to distinguish from naïve T cells, chemokine receptors, and adhesion molecules, allowing central memory T cells to become specialized for residing in skin. (B) Psoriatic disease: initial physiopathology, disease recurrence in resolved psoriasis lesions and transition to PsA. Psoriatic disease involves a complex interplay of immune dysregulation, and environmental triggers. In PsO, activated T cells accumulate in the skin leading to inflammation and the release of cytokines, namely IFN-γ and IL-17, through IL-23 stimulation. These cytokines and immune response in PsO lead to the rapid proliferation of keratinocytes. Psoriasis can relapse in previously resolved lesions due to various factors, and some individuals with PsO may develop PsA as results of shared immune mechanism. Disease recurrence could occur due to the fact that the initial treatment does not resolve the underlying immune dysregulation. This is a complex phenomenon that involves interactions between various immune cells, including TRM and Tregs. TRM cells have been found to be present in the skin; however, they were also found in circulation, indicating that these cells can circulate between lymphoid and non-lymphoid tissues. In psoriatic disease Tregs are dysfunctional or reduced in number, leading to an imbalance between proinflammatory and anti-inflammatory responses that contribute to the recurrence of psoriatic lesions. In this process Langerhans cells are capable of producing inflammatory cytokines such as IL-17A that contribute to the perpetuation of inflammatory response and potentially lead to the recurrence of psoriatic lesions. In PsA, activated T cells produce TNF-α, IL-17, and IL-23. These cytokines play a central role in the inflammatory cascade, leading to joint inflammation, damage, and the clinical features of PsA. CD4+ and CD8+ T cells are recruited to the synovial tissue of affected joints in PsA patients. Specific T cells subsets capable of entering the bloodstream and recirculating between organs are represents in red in the figure. TEM, Effector memory T cells; TCM, Central memory T cells; Treg, Regulatory T cells, TEFF, Effector T cells.
Do TRM and Treg cells play any additional role in PsO pathophysiology?
The involvement of Effector Memory T (TEM) and Th cells in PsO has been described since 1994 (46). TEM (CD62Llo/CCR7lo) are CD8+ cells that express CD45RO and that have greater cytolytic capacity than central memory T cells (TCM), and express integrins and chemokines necessary for localization to inflamed tissues (50). Th cells are CD4+ and play an important role in adaptive immunity (46). It has been shown that the initial phase of PsO is characterized by heavy epidermal infiltration of CD4+ T cells. Furthermore, these cells play a central role in the progression of PsO (43, 46). Helper CD8+ T cells may induce secretion of cytokines by neighboring cells, such as CCL20 from keratinocytes, which has been associated with the worsening of PsO (82). Additionally, helper CD8+ T are capable of strong expression of CD40L ligand upon activation, which could be a driver of autoinflammatory processes (82).
Treg cells are a subtype of Th cells that are characterized by Foxp3 expression, a transcription factor that is required for their development and function. These cells are crucial for suppressing immune responses mainly through secretion of IL-10 and TGFβ to maintain or restore immune balance. However, in a study by Yang et al., Tregs from psoriatic patients demonstrated a preponderance towards STAT3 phosphorylation when exposed to pro-inflammatory cytokines (such as IL-6, IL-21, and IL-23), resulting in a weakened capacity to suppress T cell mediated inflammation (67). Additionally, Tregs from patients with severe PsO have been shown to have an enhanced propensity to differentiate into IL-17A-producing cells, when compared to healthy controls, which may be triggered by IL-23. Furthermore, IL-23 may reduce Foxp3 expression, without impacting RORγt expression (58). The balance between Foxp3 and RORγt expression may determine whether Tregs maintain an immune-modulatory profile or undergo IL-23 driven differentiation into Th17 cells (i.e., a high RORγt:Foxp3 ratio will promote induction of proinflammatory IL-17A transcription program) (58). Th17 cells, which are IL-17 producing CD4 T cells, mediate inflammation and may also be involved in sustained inflammation later in disease progression (57).
CCL20 is believed to be a key mediator responsible for recruiting TEM and Treg into PsO lesions (47, 55). The presence of CCL20 transcript, primarily expressed by keratinocytes, helps to distinguish lesional from non-lesional skin in psoriatic patients. Further, expression of CCL20 is higher in lesional skin when compared to non-lesional skin and can even be upregulated in non-lesional skin of psoriatic patients when compared to healthy skin from healthy controls. Moreover, CCL20-expressing keratinocytes colocalize with skin-infiltrating T lymphocytes and CCR6, the receptor for CCL20, is expressed at high levels on the skin homing cutaneous lymphocyte associated antigen (CLA+) subset of memory T cells (47, 55). CLA is a ligand for E-selectin and is typically present in T cells that infiltrate into cutaneous sites (51). In addition, CCL20 is associated with endothelial inflammation, which also reflects the state of systemic inflammation that characterizes PsO (47, 55).
IL − 15 has been proposed as a potentially relevant target against the IL-17 response in psoriasis, as it is co-expressed with IL-23 in psoriatic skin lesions. IL-15 is also considered a susceptibility gene to the development of psoriasis. De Jesús-Gil et al identified a synergistic effect between IL-15 and IL-23 on IL-17A/F induced response in CLA+ T cells (81). Of note, this synergistic effect in IL-17A/F production was restricted to skin-related TRM cells in psoriasis patients and it was not observed in healthy controls, and requires the presence of epidermal cells, indicating a cooperation between T cells and epidermal cells (81).
Involvement of specific T cell subsets may differ across different phases of PsO (50). Cytotoxic T cells (CD8+) and Effector T cells (TEFF CD45RO+) infiltrate the epidermis during the early stages of development of PsO skin lesions accompanied by their increase in CD2 and CD25 activation markers (50). The heightened presence of CD8+ T cells in the epidermis at sites of initial formation of psoriatic plaques further supports the role of activated memory T cells in PsO lesions (46).
CCR5+ CD4 T cells also contribute to the formation of the PsO plaques (65). CCR5 and CCL5 genes expression in psoriatic skin lesions highlights the role of CCR5/CCL5 axis in PsO pathogenesis (65). CCR5 has been described as a mediator of CD4 T cell retention in dermal memory T cells clusters. CCL5 is produced by keratinocytes in psoriatic skin and play a role in recruitment of CCR5+ memory T cells (65). Furthermore, expression of CCR5 in psoriatic skin lesions is induced by proinflammatory cytokines such as IFN-γ, TNFα, and CCR5 serves as a chemoattractant for eosinophils, basophils, monocytes and TEFF (CD4+ and CD45RO+), NK cells, and immature dendritic cells (65).
It is well established that severe PsO is associated with systemic inflammation leading to comorbidities such as cardiovascular disorders, metabolic syndrome, among others. Circulating CD103+CCR4+CCR5+ and CCR4+CCR6− CD8+ effector T cells (TEFF) cells were highly correlated with C-reactive protein (CRP) as well as with PASI (Psoriasis Area and Severity Index) score in a study by Sgambelluri et al (65). Another study, by Diani et al., demonstrated that the circulating fraction of CCR6+ CD4+ TEM and TEFF cells also correlates with systemic inflammation in PsO (68). Furthermore, CLA expression on TCM CCR6+ and CCR4+ cells inversely correlates with the PASI score (68). The authors emphasize this finding, hypothesizing that specific subsets of skin-tropic CLA+ CCR7+ memory CD4+ T cells could recirculate toward psoriatic skin in proportion to disease severity (68). Sgambelluri et al. (65) also hypothesized that in chronically inflamed tissue, a subset of differentiated resident cells (CD8 T cells CD103+CCR4+ CD8+ TEFF), is released in the circulation contributing to systemic inflammation (65). However, while the TEM and TEFF subsets of CCR4+ CD4 T cells are associated with PsO severity they do not correlate with systemic inflammation (65). CCR4+ CD4+ T cells, primarily TCM, constitute a subset of cells that efficiently re-enter the circulation from the skin and may relocate to the skin compartment upon antigen re-exposure or in response to inflammatory signals (65). This suggests that involvement of these cells occurs primarily in skin disease, specifically in the context of PsO pathogenesis, and may not have the same potential for impacting systemic inflammation (65).
TEM and TEFF from PsO lesions are chronically activated and poorly suppressed by Treg which, by definition, are immunosuppressive cells (40, 56). In lesional PsO skin, IL-6, a proinflammatory cytokine which signals through Stat3, is produced by keratinocytes, fibroblasts, endothelial cells, macrophages, and Th cells, allowing TEM/TEFF cells to escape Treg-mediated suppression (56). The levels of this proinflammatory cytokine is markedly elevated and expressed by CD31 endothelial cells and expressed by CD11c+ dermal dendritic cells (DC) of PsO lesions (56). Furthermore, IL-6 was necessary and sufficient to reverse human T cell suppression by Treg in an in vitro model (56). Phosphorylation of Stat3 in T cells in response to IL-6 due to presence of IL-6Rα contributed to Th17 differentiation from naïve T cells (56). In lesional PsO skin there are cells that coexpress CD3, IL-17, and IL-6 indicating that Th17 cells are present within psoriatic TEM/TEFF population contributing to IL-6 mediated resistance to Treg suppression (56). Thus, the proinflammatory effect of IL-6 and the resistance to Treg suppression may plays a key role in the pathogenesis of psoriasis. On the other hand the differentiation of Th17 cells, orchestrated by IL-6-indiced Stat3 phosphorylation, further contributes to the inflammatory milieu in psoriatic lesions.
A recent study showed that active psoriatic epidermis contained PD-1 expressing CD8+CD103+ T cells that correlates with disease severity and histopathology (90). Moreover, these cells have a memory resident phenotype, as they express IL-23R and produce IL-17A (90). Thus, the expression of PD-1 in epidermal psoriatic cells could delineate a putative pathogenic subset of epidermal CD8+CD103+ T cells, which possibly play a role in PsO pathogenesis (90).
TEFF are considered circulating precursors of TRM (65). Once TEFF are recruited to the epidermis, they may up-regulate expression of retention signals such as CD103+ and CD69+ and reside there as TRM (65). The presence of T cells expression TRM markers (CD4, CD8, CD103, CD69, CD49, CXCR6) in the epidermis and skin of patients with PsO lesions increased when compared to healthy controls (85). Furthermore, there is a significant positive relationship between the expression of TRM markers in plaque PsO and the duration of skin lesions (85). In human skin epithelia cytotoxic CD8+CD49a− TRM produced interferon-γ (IFN-γ), whereas CD8+CD49a+ TRM produce IL-17, both promoting local inflammation in the skin (34). It was also demonstrated that the presence of IL-23 is important to activate TRM to produce IFN-γ. Therefore, the augmented expression of IL-23 in keratinocytes and cutaneous APC may contribute to the perpetuation of the inflammation process in PsO (52, 53, 87). TRM at barriers surfaces express the markers CD103 and/or CD69, which function to retain them in epithelial tissues (26). Characterization of skin TRM within skin lesions showed that CD8+CD103+ TRM cells, with a small number of CD4+CD103+ TRM cells, were present in the epidermis of PsO and associated with acanthosis (78). These cells represent an effector population capable of maintaining the psoriatic phenotype and driving recurrence of disease at anatomical areas with cutaneous plaques (73, 90). Most of these cells express CD69, which conveys a skin-homing potential and CXCR3, contributing to the perpetuation of inflammation (68). TRM cells are believed to be responsible for the immune memory of PsO skin lesions, may explain occurrence of skin lesions based on the Koebner phenomenon (85). A portion of CD8+CD103+ TRM cells produced IFN-γ, IL-17A or IL-22 (78).
Contrary to the initial belief TRM cells may not fully be characterized as resident in nature, as some may re-enter the recirculation from the skin to other tissues. CD4+CD69+CD103+ TRM in human are capable of downregulating CD69 and exiting the skin (26). Furthermore, a CD4+CD69−CD103+ skin tropic population of cells was detected in lymph nodes and blood, and were transcriptionally, functionally, and clonally related to CD4+CD69+CD103+ TRM in the skin. Additionally, Klicznik et al. demonstrated that CD4+ CD103+ TRM could re-enter circulation and migrate to secondary body sites and reassume a TRM phenotype (CD69+) (26). These findings raise the question of whether recirculating TRM cells derived from the skin may traffic to the musculoskeletal structures and contributes to the pathogenesis of PsA and the progression from PsO to PsA?
The complex relationship between dendritic cells (DC), keratinocytes, and T cells in PsO is highlighted by the dual production of IL-23, a central driver of inflammation. While primarily produced by DC, IL-23 can also be produced by keratinocytes. This cytokine plays a crucial role in enhancing IFN-γ production by memory T cells and promoting the expansion of pathogenic Th17 (52, 87). Notably, keratinocytes in lesional skin produced markedly higher levels of IL-23 compared to keratinocytes in normal and psoriatic non-lesional skin (52, 79).
In the ECLIPSE study, a head-to-head clinical trial comparing IL-23p19 blockade with guselkumab and IL-17A blockade with secukinumab for the treatment of moderate-to-severe psoriasis in adults (87). Although both guselkumab and secukinumab decreased the frequency of CD4+CD49a−CD103− T cells, they exerted a differential effect on CD8+ TRM and Tregs. By week 4, secukinumab reduced the frequency of Tregs but not CD8+ TRM., whereas guselkumab decreased the frequency of TRM but not of Tregs (87). Therefore, an increase in the ratio of Treg to CD8+ TRM was observed in lesional skin of patients treated with guselkumab compared to patients treated with secukinumab and this differences reached significal significance at week 24 (87). Although skin biopsies were not obtained at later timepoint in the ECLIPSE study, these finds could potentially account for guselkumab achieving superiority versus secukinumab in achieving the primary endpoint of PASI 90 response at week 48 (87).
3.2 Psoriatic arthritis
PsA is an inflammatory, MHC class I allele associated disease, in which injury is likely mediated predominantly by T cells (Figure 2) (42, 49). The enthesis is considered the cardinal lesion in PsA, and recently, Bridgwood C et al. showed that normal spinal enthesis soft tissue has resident myeloid cells capable of being induced to produce IL-23 (74). Naïve CD45RO- CD4+ T lymphocytes are predisposed to differentiate into Th17 cells, characterized by IL-17A and IL-22 production (71). In the very early stages of the disease, which is strongly associated with early enthesitis, IL-17 producing CD4+ Th17 cells, initiate inflammation and may also be involved in sustained inflammation later in disease progression (57). The unregulated of Th17 cell activity is due to intrinsically elevated expression of RORC gene (RAR Related Orphan Receptor C) accompanied by biased Th17 cell development and resistance of Th17 cells to natural antagonists, such as IL-12, IL-4 and IFN-γ (57).
To characterize the network of immune responses and chemokine signaling in the synovial microenvironment that could support the involvement of TRM cell in the pathogenesis of PsA Steel et al. conducted a comprehensive analysis of T cells derived from both peripheral blood and synovial fluid of patients with established PsA. The results showed that IL-17A+CD8+ T cells were predominantly TCRαβ+. Moreover, they were more frequent in the synovial fluid compared to peripheral blood (42). These cells were also polyclonal, and Tc17 cells (IL-17A+CD8+) express hallmarks of both Th17 cells (RORC/IL-23R/CCR6/CD161) and Tc1 cells (granzyme A/B). Synovial Tc17 cells exhibited a strong TRM cells signature (CD45RA−PD1+HLA−DR+) secreting a range of proinflammatory cytokines (IFN-γ, TNF, GM- CSF, IL-21, and IL-22) alongside with IL-17A (42). Retention of these TRM cells in the joints could be mediated by increased levels of CXCR6 ligand CXCL16 (42). CXCL16 possesses chemotactic and angiogenic properties and can be produced as a soluble mediator or as a transmembrane-bound chemokine by monocytes, macrophages, and dendritic cells. Furthermore, CXCL16 was shown to enhance recruitment CXCR6 expressing T cells derived from inflamed tissue (42). Diani et al. demonstrated that the proportion of CXCR6+CD8+T cell effectors expressing CD69, which are capable of trafficking to synovial fluid, was increased in the circulation of PsA patients and correlated with serum CRP levels (75). Furthermore, the number of IL-17 producing cells was also increased in the circulation of PsA patients (75). Moreover, accumulation of CXCR3+ CD8+ T cells, CD4+ T cells and CD69+ cells was found in synovial fluid from PsA patients (21, 75). Taken together, these findings suggest a potential role for CD8+ memory T cell effectors in both systemic and joint manifestations of PsA (75).
According to a study by Povoleri et al. TRM cells may also play a fundamental role in PsA. CD8+CD69+CD103+ TRM cells, such as cytotoxic Treg like TRM, and CD161+CCR6+ type 17-like TRM cells with a pro-inflammatory cytokine profile (IL-17A+TNFα+ IFN-γ+) were identified in the synovial fluid of PsA patients. Only one population of CD4+CD69+CD103+ TRM cells was detected in synovial fluid of PsA patients (14). The significantly larger fraction of IL-17A-secreting tissue-resident and non-resident CD8+ T cells within synovial fluid PsA may contribute to the persistence of the disease (14).
Tregs, particularly those expressing an immunosuppressive profile (i.e., CD4+CD45+ Foxp3+) have also been implicated in the pathogenesis of PsA (84). In a study by Wang et al. PsA patients had lower peripheral Treg counts compared to healthy controls, which was accompanied by an increase in Th17 cells. Also, the absolute number of peripheral Treg cells was significantly negatively correlated with PsA disease activity (84). A very recent study demonstrated that Treg cells derived from the inflammatory microenvironment of inflamed joints in patients with PsA are phenotypically and functionally different from Tregs in the circulation (44). Tregs can down regulate their key transcriptional factor Foxp3 and to assume TEFF phenotype and functions accompanied by the production of pro-inflammatory cytokines including IL-17, upregulation of inhibitory immune receptors, and upregulation of CD161, RORγt and ICOS (44). Treg cells that downregulate Foxp3 exhibit lower expression of inhibitory receptors such as CTLA-2 and TIGIT and produce even more IL-17, displaying the highest expression of CD161 (44). Furthermore, a transcriptional network profile identified through ex vivo signaling protein mapping on T lymphocytes in PsA joints revealed the complex interplay between IL-1, IL-6, and IL-23 signaling and differentiation of Th17 cells and CD4+ Tregs in sustained joint inflammation in PsA (64, 71).
3.3 PsO-PsA progression
The transition from PsO to PsA has gained interest in recent years. However few studies address the roles of T lymphocytes, namely CD4+ and CD8+ cells after PsO onset and before clinical manifestation of PsA. CD4+ TEM cells expressing Th17-associated trafficking receptor CCR6 have been positively correlated with systemic inflammation in patients with psoriatic disease (PsO or PsA) (75).
An analysis of chemokine receptor profiling of circulating cells revealed a reduction in the percentage of CD8+ and CD4+ T cells expressing the Th1/Tc1-associated trafficking receptor CXCR3+, in patients with compared with patients with PsO limited to the and healthy subjects (75).
Another study by Leijten et al. showed that the peripheral blood of patients with PsA, when compared to healthy donors, was characterized by an increase in regulatory CD4+T cells and IL-17A and IL-22 coproducing CD8+ T cells (86). Further, CD8+CCR10+ T cells were enriched in the peripheral blood of PsA patients compared to PsO patients (86). These cells are specific TEM, with a Tc2/22-like cytokine profile (CD8+T cells) and regulatory function. They also co-express skin-homing receptors CCR4 and CLA, along with high levels of DNAX accessory molecule 1. Additionally, these cells were detected under both inflammatory and homeostatic conditions in skin, but were not enriched in synovial fluid (86). Furthermore, this CD8+ T cell subset exhibit a transcriptomic profile comparable to that observed in TRM cells, that originated in the skin, but not in the joint (86).
PsA patients also exhibited a decline in the percentage of circulating Th1/Tc1 IFN-γ− producing CD8+ and CD4+ T cells compared to patients with PsO (75). Findings from Diani et al supported the concept of CXCR3+ cells recruitment to inflamed psoriatic tissue. Based on analysis of the T cell characteristics present in both peripheral blood and synovial fluid mononuclear cells, which demonstrated accumulation of CXCR3+ T cells. Notably, a substantial increase in the levels of CXCR3 ligand, CXCL10, was identified within synovial fluid (75).
The use of machine learning for analysis of peripheral blood immune cell phenotype of consecutive PsO and PsA patients allowed discrimination of immune profiles (88). Key PsA-classifying cell subsets identified included increased proportions of differentiated CD4+CD196+CD183−CD194+ and CD4+CD196−CD183−CD194+ T-cells and reduced proportions of CD196+ and CD197+ monocytes, memory CD4+ and CD8+ T-cell subsets and CD4+ Treg cells (88). The reductions in these T cell subsets could be explained by migration of these cells to the joints and entheses of PsA patients, possibly contributing to disease progression from PsO to PsA.
4 Discussion
Over the past decade, the critical involvement of T cells in the development of PsO skin lesions has been extensively elucidated. Our understanding of the distinct contributions of various T cell subsets to the pathogenesis of PsO has significantly advanced. In particularly, T cells producing IL-17A, play a central role in driving PsO (34, 42, 62, 63, 68, 75, 76) and mature CD8+ T cells could be a putative link to systemic inflammation (75).
Activated T cells can migrate into the epidermis and recognize epidermal autoantigens and possibly progress toward differentiating into TRM (7). That TRM are found in the systemic circulation suggests that re-activated TRM are capable of retrograde migration from non-lymphoid tissue, such as the skin. In turn, these circulating TRM retain the capacity to return to the skin. Furthermore, these cells were also identified in patients with chronic inflammatory diseases, such as psoriatic disease, leading to the hypothesis that TRM regress from inflamed tissue as well (36). The presence of TRM in both tissue and circulating compartment could have implications for development of novel therapies targeting chronic inflammation and circulating TRM could provide a vital diagnostic tool as biomarkers of disease activity and even potentially predict resistance to therapy (36). Additionally, complete TRM suppression appears to be required for achieving disease remission (85).
Towards this, there is a paucity of research assessing the dynamic changes in TRM counts, pathogenic versus non-pathogenic function induced by available psoriasis therapies (95). Compared to TNF inhibition and methotrexate, IL-17A inhibition achieves the most rapid reduction of expression of TRM specific markers. Interestingly, after psoriatic lesions have resolved TRM can still produce IL-17A and the implications for treatment to prevent relapses are unknown (98). It seems that it may vary depending on the therapy. Furthermore, the reduction of the expression of TRM-specific markers was observed predominantly in the dermis, and not in lesional epidermis (95). Thus, how topical drugs may influence TRM quantity in psoriatic lesions remains a relevant question?
Mehta et al. in a study subanalysis of lesional biopsy specimens taken from patients successfully treated with either an IL-23p19 inhibitor, guselkumab, or an IL-17A inhibitor, secukinumab, observed that both CD4+ and CD8+ TRM cells decreased in psoriatic lesions from baseline through week 24 in both groups; however, the decrease in CD8+ TRM cells' frequency was significant only for patients treated with guselkumab (87). IL-23 receptor is reported to be upregulated in the CD8+CD49a−CD103+TRM subsets relative to CD8+CD49a+CD103+TRM (34). Further, CD49a− TRM cells preferentially produce IL-17 and these cells are profoundly expanded in the hypertrophic epidermis of psoriatic lesions (34). Which may, in part, account for the impact of the treatment with guselkumab.
TRM has an undeniable ability to accumulate in the skin of psoriatic patients with significant longevity. Therefore, the relationship between TRM accumulation and disease duration is also an important consideration. This could potentially explain the suboptimal therapeutic response in patients with longer disease duration, often accompanied byrapid recurrence of psoriatic lesions at the same anatomical sites (85).
Taken together, these considerations could support early intervention therapeutic in PsO specifically targeting TRM cells to achieve better responses and remission of disease. Eyerich et al. hypothesized that patients with a short disease duration (≤2 years) may exhibit a more rapid and pronounced response to guselkumab treatment (99). Furthermore, these patients may be better positioned to maintain long-term drug-free disease control after guselkumab withdrawal (99). However, the data are sparce and disease modifying trials are warranted (7, 100).
Our extensive understanding of PsA primarily derives from studies conducted on well-established cases, focusing on patients who experienced having the disease for several years. However, our current knowledge of early-stage PsA remains limited, as early intervention represents an under-recognized clinical unmet need in PsA (77). Nevertheless, the most recent EULAR guidelines, which address the transition from PsO to PsA have formulated guidance in the areas of interception of PsA and the clinical management of patients at high risk development of PsA (101). Three distinct stages relevant to PsA considered: patients with PsO at higher risk of developing PsA, those with subclinical PsA and those with clinical manifestations of PsA (101).
In the absence of specific PsA biomarkers, early detection of PsA in PsO patients is both challenging and crucial for timely treatment and inhibiting structural damage (16, 102). Defining a specific immune profile and imaging modalities that could identify patients before onset of clinical features of PsA could potentially facilitate the referral to the rheumatology clinic (16, 85, 88, 93, 101). Moreover, early treatment of PsA may lead to better outcomes for patients due to enhanced therapeutic effectiveness (16, 101). The severity of skin involvement in PsA patients varies. Many but not all PsA patients may have PsO involving the skin requiring systemic treatment (16). Therefore, there is significant interest in the concept of treating the PsO to prevent PsA in a “Treat to Intercept” approach (16). A recent retrospective study published by Singla et al., which included a large cohort of patients with PsO demonstrated that treatment of PsO patients with IL-12/23 or IL-23 inhibitors was associated with reduced risk of inflammatory arthritis when compared with TNF inhibitors. No significant differences were found between IL-17 inhibitors and TNF inhibitors (103). Further studies are needed to confirm these results.
The convergent immunological bridge between the skin and joints is being increasingly recognized (16). Not activated T cells, including CD4+ and CD8+ TRM, are present at under normal conditions in both sites (14, 68, 75, 85, 86, 88, 90, 93, 95).
The increased frequency of IL-17A+CD8+T cells in synovial fluid of patients with PsA adds growing evidence that CD8+ T cells are relevant to the immunopathogenesis of PsA (42). A study by Poloveri et al. showed that CD8+ Treg-like cells harbors various markers of TRM cells, suggesting that these cells may persist in the synovium joints in patients with PsA (14). Moreover, three clusters of TRM cells were found in the psoriatic joints. Joints of PsA patients are enriched for pro-inflammatory type 17: CD8+CD69+CD103+ TRM, characterized by the expression of CD161 and CCR6. The presence of these clusters of cells, all containing IL-17 producing cells suggests that they represent different fates or states of type 17-like TRM, cells. This supports the concept that Tc17/TRM cells in PsA are polyfunctional and able to produce multiple cytokines that drive PsA inflammation (14, 42). In turn, synovial type 17 TRM cells may contribute to joint flares in PsA after treatment withdrawal as in PsO.
These findings lend strength to the idea that there are human CD8+ TRM in circulation (26, 104), suggesting a potential role of these cells in the progression from PsO-PsA (42). This raises the hypothesize that PsO, as a human disease of the barrier tissue may involve a dynamic balance between tissue resident and recirculating memory T cells (105). CD4+ T cells also recirculating between blood and skin and could play a key role in determining the amplitude of skin inflammation in more severe PsO (68).
Some studies that included PsA patients evaluated CD8+ T and CD4+ T cells in the blood, synovial tissue, and synovial fluid showed that there are significant differences between these compartments (42, 75, 86). This is further supported by a study indicating that the subset composition and phenotype of peripheral blood T cells do not mirror that of synovial fluid, suggesting that blood represents a distinct compartment of T cells (23). Additionally, distinct trafficking patterns exist for TRM subsets (23). The molecular profiles of resident and circulating T cells could offer insights into the relationship between the skin and joints (23).
The expression of CXCR6, a marker of TRM, could enhance residence of synovial Tc17 cells in inflamed tissue. This is accompanied by an increase in the levels of CXCL16 in PsA synovium (42). CXCR6 expression at the site of inflammation has been reported in PsO, and CXCL16 has been shown to enhance recruitment of inflamed tissue-derived CXCR 6 expressing T cells (42, 85, 106). A recent study has shown that CXCR6 is the most up regulated chemokine receptor in cutaneous lesions of PsA patients, and its expression correlates positively with PASI score (107). Moreover, CXCL16 blockade or CXCR6 deficiency led to reduced features of arthritis and lower IL-17 production in experimental models (108).
Thus, taken together, the data suggest that increased CXCR6 expression in PsO skin lesions of PsA patients may contribute to the recruitment and persistence of Tc17 cells in their inflamed joints (42, 85, 107).
Few clinical studies have addressed disease interception for PsA through early interventions in high risk PsO patients. The IVEPSA study is an open-label, prospective, exploratory interception study in very early PsA patients with subclinical inflammatory lesions in the joints, treated with an IL17A inhibitor (77). Data from this study demonstrated that inhibition of IL-17 halts the progression of catabolic and anabolic changes in patients' joints (77). Therefore, IVEPSA provides a strong rational for early interventions to intercept PsA. However, selection of high risk patients for early intervention could pose challenges (77). Identification of patients through the use of flow cytometry and machine learning algorithms, while not currently applicable in clinical practice, could offer new perspectives for accurately selecting patients based on their immune profile (88). Similarly, the PAMPA study is an ongoing multicentric, randomized, double-blind, placebo-controlled, interventional, PSA prevention trial at high risk for progression to PsA who are receiving treatment with guselkumab and non-biological standard of care (109). Taken together these studies will contribute to approaches for identifying patients at high risk of progression to PsA and the impact of treatment in preventing this process (109). Due to the limited number of available studies for this systematic literature review, particularly those addressing the transition PsO to PsA, the data on this topic are limited. While there are strong indications regarding the role of TRM and Treg in intercepting in PsA and the potential for targeting them for intercepting PsA, clinical studies are needed to further explore and translate these described mechanisms to patients and potential treatment targets.
5 Conclusion
In conclusion, early intervention in PsA represents an under-recognized clinical need particularly in high-risk PsO patients. Early detection, although challenging without specific biomarkers, is crucial for timely treatment and to prevent permanent structural damage. Research has shed light on the role of T cells, particularly those producing IL-17A, in driving PsO and in the progression PsO to PsA. CD8+ cells and TRM have emerged as significant players in both skin and joint inflammation. Targeting TRM especially in the early stages of disease, appears promising for improving treatment outcomes and maintaining long-term disease control. While these findings offer promising avenues for early intervention and understanding the role TRM and Tregs cells in PsO and PsA, the limited number of available studies highlights the need for further clinical research to translate this hypothesis into effective treatments and improve patient outcomes.
Author contributions
BL: Writing – review & editing. DL: Writing – review & editing. AG: Writing – review & editing. PM-B: Writing – review & editing.
Funding
The author(s) declare financial support was received for the research, authorship, and/or publication of this article. The project received financial support from Janssen for the logistics of expert meetings and editorial support.
Acknowledgments
The authors would like to thank Irina Alho Duarte, from X2SS Portugal, for providing medical writing and editorial support to this manuscript.
Conflict of interest
AG and DL are employees of Janssen Pharmaceutical, Portugal. PM-B has worked as a speaker and/or consultant for AbbVie, Pfizer, Janssen-Cilag, Leo-Pharma, Novartis, Eli-Lilly, Sanofi, Teva, L’Oreal, Pierre Fabre, Cantabria Labs, Bayer, Viatris, Organon, Evelo Biosciences and CS Labs. And works/has worked as a Principal Investigator in Clinical trials supported by AbbVie, Amgen, Biogen, Janssen, Pfizer, Novartis, and Sanofi.
The remaining author declares that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Publisher’s note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
References
1. Boehncke, WH , and Schön, MP . Psoriasis. Lancet. (2015) 386:983–94. doi: 10.1016/s0140-6736(14)61909-7
2. Emmanuel, T , Mistegård, J , Bregnhøj, A , Johansen, C , and Iversen, L . Tissue-resident memory T cells in skin diseases: a systematic review. Int J Mol Sci. (2021) 22:9004. doi: 10.3390/ijms22169004
3. Griffiths, CEM , Armstrong, AW , Gudjonsson, JE , and Barker, J . Psoriasis. Lancet. (2021) 397:1301–15. doi: 10.1016/s0140-6736(20)32549-6
4. Billi, AC , Gudjonsson, JE , and Voorhees, JJ . Psoriasis: past, present, and future. J Invest Dermatol. (2019) 139:e133–42. doi: 10.1016/j.jid.2019.08.437
5. Hu, P , Wang, M , Gao, H , Zheng, A , Li, J , Mu, D, et al. The role of helper T cells in psoriasis. Front Immunol. (2021) 12:788940. doi: 10.3389/fimmu.2021.788940
6. Di Meglio, P , Villanova, F , Navarini, AA , Mylonas, A , Tosi, I , Nestle, FO, et al. Targeting Cd8(+) T cells prevents psoriasis development. J Allergy Clin Immunol. (2016) 138:274–6.e6. doi: 10.1016/j.jaci.2015.10.046
7. Ben Abdallah, H , Emmanuel, T , Bregnhøj, A , Johansen, C , and Iversen, L . Early intervention and disease memory in psoriasis: a literature review. JEADV Clin Pract. (2022) 1:307–16. doi: 10.1002/jvc2.63
8. Puig, L , Costanzo, A , Munoz-Elias, EJ , Jazra, M , Wegner, S , Paul, CF, et al. The biological basis of disease recurrence in psoriasis: a historical perspective and current models. Br J Dermatol. (2022) 186:773–81. doi: 10.1111/bjd.20963
9. Masopust, D , and Soerens, AG . Tissue-resident T cells and other resident leukocytes. Annu Rev Immunol. (2019) 37:521–46. doi: 10.1146/annurev-immunol-042617-053214
10. Szabo, PA , Miron, M , and Farber, DL . Location, location, location: tissue resident memory T cells in mice and humans. Sci Immunol. (2019) 4:9673. doi: 10.1126/sciimmunol.aas9673
11. Mueller, SN , and Mackay, LK . Tissue-resident memory T cells: local specialists in immune defence. Nat Rev Immunol. (2016) 16:79–89. doi: 10.1038/nri.2015.3
12. FitzGerald, O , Ogdie, A , Chandran, V , Coates, LC , Kavanaugh, A , Tillett, W, et al. Psoriatic arthritis. Nat Rev Dis Primers. (2021) 7:59. doi: 10.1038/s41572-021-00293-y
13. Pennington, SR , and FitzGerald, O . Early origins of psoriatic arthritis: clinical, genetic and molecular biomarkers of progression from psoriasis to psoriatic arthritis. Front Med (Lausanne). (2021) 8:723944. doi: 10.3389/fmed.2021.723944
14. Povoleri, GAM , Durham, LE , Gray, EH , Lalnunhlimi, S , Kannambath, S , Pitcher, MJ, et al. Psoriatic and rheumatoid arthritis joints differ in the composition of Cd8+ tissue-resident memory T cell subsets. Cell Rep. (2023) 42:112514. doi: 10.1016/j.celrep.2023.112514
15. Azuaga, AB , Ramírez, J , and Cañete, JD . Psoriatic arthritis: pathogenesis and targeted therapies. Int J Mol Sci. (2023) 24:4901. doi: 10.3390/ijms24054901
16. McGonagle, DG , Zabotti, A , Watad, A , Bridgewood, C , De Marco, G , Kerschbaumer, A, et al. Intercepting psoriatic arthritis in patients with psoriasis: buy one get one free? Ann Rheum Dis. (2022) 81:7–10. doi: 10.1136/annrheumdis-2021-221255
17. McGonagle, D , Gibbon, W , and Emery, P . Classification of inflammatory arthritis by Enthesitis. Lancet. (1998) 352:1137–40. doi: 10.1016/s0140-6736(97)12004-9
18. Zuliani, F , Zabotti, A , Errichetti, E , Tinazzi, I , Zanetti, A , Carrara, G, et al. Ultrasonographic detection of subclinical Enthesitis and synovitis: a possible stratification of psoriatic patients without clinical musculoskeletal involvement. Clin Exp Rheumatol. (2019) 37:593–9. doi: 10.31138/mjr.33.4.421
19. McGonagle, D , Lories, RJ , Tan, AL , and Benjamin, M . The concept of a "Synovio-Entheseal complex" and its implications for understanding joint inflammation and damage in psoriatic arthritis and beyond. Arthritis Rheum. (2007) 56:2482–91. doi: 10.1002/art.22758
20. Chimenti, MS , Caso, F , Alivernini, S , De Martino, E , Costa, L , Tolusso, B, et al. Amplifying the concept of psoriatic arthritis: the role of autoimmunity in systemic psoriatic disease. Autoimmun Rev. (2019) 18:565–75. doi: 10.1016/j.autrev.2018.11.007
21. Penkava, F , Velasco-Herrera, MDC , Young, MD , Yager, N , Nwosu, LN , Pratt, AG, et al. Single-cell sequencing reveals clonal expansions of pro-inflammatory synovial Cd8 T cells expressing tissue-homing receptors in psoriatic arthritis. Nat Commun. (2020) 11:4767. doi: 10.1038/s41467-020-18513-6
22. Veale, DJ , and Fearon, U . The pathogenesis of psoriatic arthritis. Lancet. (2018) 391:2273–84. doi: 10.1016/s0140-6736(18)30830-4
23. Sathaliyawala, T , Kubota, M , Yudanin, N , Turner, D , Camp, P , Thome, JJ, et al. Distribution and compartmentalization of human circulating and tissue-resident memory T cell subsets. Immunity. (2013) 38:187–97. doi: 10.1016/j.immuni.2012.09.020
24. Iijima, N , and Iwasaki, A . Tissue instruction for migration and retention of Trm cells. Trends Immunol. (2015) 36:556–64. doi: 10.1016/j.it.2015.07.002
25. Gebhardt, T , Mueller, SN , Heath, WR , and Carbone, FR . Peripheral tissue surveillance and residency by memory T cells. Trends Immunol. (2013) 34:27–32. doi: 10.1016/j.it.2012.08.008
26. Klicznik, MM , Morawski, PA , Höllbacher, B , Varkhande, SR , Motley, SJ , Kuri-Cervantes, L, et al. Human Cd4(+)Cd103(+) cutaneous resident memory T cells are found in the circulation of healthy individuals. Sci Immunol. (2019) 4:8995. doi: 10.1126/sciimmunol.aav8995
27. Fonseca, R , Beura, LK , Quarnstrom, CF , Ghoneim, HE , Fan, Y , Zebley, CC, et al. Developmental plasticity allows outside-in immune responses by resident memory T cells. Nat Immunol. (2020) 21:412–21. doi: 10.1038/s41590-020-0607-7
28. Strobl, J , Gail, LM , Kleissl, L , Pandey, RV , Smejkal, V , Huber, J, et al. Human resident memory T cells exit the skin and mediate systemic Th2-driven inflammation. J Exp Med. (2021) 218:e20210417. doi: 10.1084/jem.20210417
29. Enamorado, M , Khouili, SC , Iborra, S , and Sancho, D . Genealogy, dendritic cell priming, and differentiation of tissue-resident memory Cd8(+) T cells. Front Immunol. (2018) 9:1751. doi: 10.3389/fimmu.2018.01751
30. Masopust, D , Vezys, V , Marzo, AL , and Lefrançois, L . Preferential localization of effector memory cells in nonlymphoid tissue. Science. (2001) 291:2413–7. doi: 10.1126/science.1058867
31. Schenkel, JM , and Masopust, D . Tissue-resident memory T cells. Immunity. (2014) 41:886–97. doi: 10.1016/j.immuni.2014.12.007
32. Sallusto, F , Geginat, J , and Lanzavecchia, A . Central memory and effector memory T cell subsets: function, generation, and maintenance. Annu Rev Immunol. (2004) 22:745–63. doi: 10.1146/annurev.immunol.22.012703.104702
33. Herndler-Brandstetter, D , Ishigame, H , Shinnakasu, R , Plajer, V , Stecher, C , Zhao, J, et al. Klrg1(+) effector Cd8(+) T cells lose Klrg1, differentiate into all memory T cell lineages, and convey enhanced protective immunity. Immunity. (2018) 48:716–29.e8. doi: 10.1016/j.immuni.2018.03.015
34. Cheuk, S , Schlums, H , Gallais Serezal, I , Martini, E , Chiang, SC , Marquardt, N, et al. Cd49a expression defines tissue-resident Cd8(+) T cells poised for cytotoxic function in human skin. Immunity. (2017) 46:287–300. doi: 10.1016/j.immuni.2017.01.009
35. Chang, MH , Levescot, A , Nelson-Maney, N , Blaustein, RB , Winden, KD , Morris, A, et al. Arthritis flares mediated by tissue-resident memory T cells in the joint. Cell Rep. (2021) 37:109902. doi: 10.1016/j.celrep.2021.109902
36. Samat, AAK , van der Geest, J , Vastert, SJ , van Loosdregt, J , and van Wijk, F . Tissue-resident memory T cells in chronic inflammation-local cells with systemic effects? Cell. (2021) 10:409. doi: 10.3390/cells10020409
37. Heath, WR , and Carbone, FR . The skin-resident and migratory immune system in steady state and memory: innate lymphocytes, dendritic cells and T cells. Nat Immunol. (2013) 14:978–85. doi: 10.1038/ni.2680
38. Kok, L , Dijkgraaf, FE , Urbanus, J , Bresser, K , Vredevoogd, DW , Cardoso, RF, et al. A committed tissue-resident memory T cell precursor within the circulating Cd8+ effector T cell Pool. J Exp Med. (2020) 217:e2019711. doi: 10.1084/jem.20191711
39. Gaide, O , Emerson, RO , Jiang, X , Gulati, N , Nizza, S , Desmarais, C, et al. Common clonal origin of central and resident memory T cells following skin immunization. Nat Med. (2015) 21:647–53. doi: 10.1038/nm.3860
40. Liu, Y , Wang, H , Taylor, M , Cook, C , Martínez-Berdeja, A , North, JP, et al. Classification of human chronic inflammatory skin disease based on single-cell immune profiling. Sci Immunol. (2022) 7:eab19165. doi: 10.1126/sciimmunol.abl9165
41. Watad, A , Rowe, H , Russell, T , Zhou, Q , Anderson, LK , Khan, A, et al. Normal human Enthesis Harbours conventional Cd4+ and Cd8+ T cells with regulatory features and inducible Il-17a and Tnf expression. Ann Rheum Dis. (2020) 79:1044–54. doi: 10.1136/annrheumdis-2020-217309
42. Steel, KJA , Srenathan, U , Ridley, M , Durham, LE , Wu, SY , Ryan, SE, et al. Polyfunctional, Proinflammatory, tissue-resident memory phenotype and function of synovial interleukin-17a+Cd8+ T cells in psoriatic arthritis. Arthritis Rheumatol. (2020) 72:435–47. doi: 10.1002/art.41156
43. Yoo, IS , Lee, JH , Song, ST , Kim, JH , Lee, HJ , and Kang, SW . T-helper 17 cells: the driving force of psoriasis and psoriatic arthritis. Int J Rheum Dis. (2012) 15:531–7. doi: 10.1111/j.1756-185X.2012.01813.x
44. Pouw, JN , Olde Nordkamp, MAM , van Kempen, T , Concepcion, AN , van Laar, JM , van Wijk, F, et al. Regulatory T cells in psoriatic arthritis: an Il-17a-producing, Foxp3(Int)Cd161 + Rorgammat + Icos + phenotype, that associates with the presence of Adamtsl5 autoantibodies. Sci Rep. (2022) 12:20675. doi: 10.1038/s41598-022-24924-w
45. Page, MJ , McKenzie, JE , Bossuyt, PM , Boutron, I , Hoffmann, TC , Mulrow, CD, et al. The Prisma 2020 statement: an updated guideline for reporting systematic reviews. BMJ. (2021) 372:n71. doi: 10.1136/bmj.n71
46. Wakita, H , and Takigawa, M . E-selectin and vascular cell adhesion Molecule-1 are critical for initial trafficking of helper-inducer/memory T cells in psoriatic plaques. Arch Dermatol. (1994) 130:457–63. doi: 10.1001/archderm.1994.01690040061008
47. Homey, B , Dieu-Nosjean, MC , Wiesenborn, A , Massacrier, C , Pin, JJ , Oldham, E, et al. Up-regulation of macrophage inflammatory Protein-3 alpha/Ccl20 and cc chemokine receptor 6 in psoriasis. J Immunol. (2000) 164:6621–32. doi: 10.4049/jimmunol.164.12.6621
48. Koreck, A , Suranyi, A , Szony, BJ , Farkas, A , Bata-Csorgo, Z , Kemeny, L, et al. Cd3+Cd56+ Nk T cells are significantly decreased in the peripheral blood of patients with psoriasis. Clin Exp Immunol. (2002) 127:176–82. doi: 10.1046/j.1365-2249.2002.01721.x
49. Curran, SA , FitzGerald, OM , Costello, PJ , Selby, JM , Kane, DJ , Bresnihan, B, et al. Nucleotide sequencing of psoriatic arthritis tissue before and during methotrexate administration reveals a complex inflammatory T cell infiltrate with very few clones exhibiting features that suggest they drive the inflammatory process by recognizing autoantigens. J Immunol. (2004) 172:1935–44. doi: 10.4049/jimmunol.172.3.1935
50. Vissers, WH , Arndtz, CH , Muys, L , Van Erp, PE , de Jong, EM , and van de Kerkhof, PC . Memory effector (Cd45ro+) and cytotoxic (Cd8+) T cells appear early in the margin zone of spreading psoriatic lesions in contrast to cells expressing natural killer receptors, Which Appear Late. Br J Dermatol. (2004) 150:852–9. doi: 10.1111/j.1365-2133.2004.05863.x
51. Clark, RA , Chong, B , Mirchandani, N , Brinster, NK , Yamanaka, K , Dowgiert, RK, et al. The vast majority of Cla+ T cells are resident in Normal skin. J Immunol. (2006) 176:4431–9. doi: 10.4049/jimmunol.176.7.4431
52. Piskin, G , Sylva-Steenland, RM , Bos, JD , and Teunissen, MB . In vitro and in situ expression of Il-23 by keratinocytes in healthy skin and psoriasis lesions: enhanced expression in psoriatic skin. J Immunol. (2006) 176:1908–15. doi: 10.4049/jimmunol.176.3.1908
53. Chen, Z , Tato, CM , Muul, L , Laurence, A , and O'Shea, JJ . Distinct regulation of Interleukin-17 in human T helper lymphocytes. Arthritis Rheum. (2007) 56:2936–46. doi: 10.1002/art.22866
54. Nograles, KE , Zaba, LC , Guttman-Yassky, E , Fuentes-Duculan, J , Suarez-Farinas, M , Cardinale, I, et al. Th17 cytokines interleukin (Il)-17 and Il-22 modulate distinct inflammatory and keratinocyte-response pathways. Br J Dermatol. (2008) 159:1092–102. doi: 10.1111/j.1365-2133.2008.08769.x
55. Pene, J , Chevalier, S , Preisser, L , Venereau, E , Guilleux, MH , Ghannam, S, et al. Chronically inflamed human tissues are infiltrated by highly differentiated Th17 lymphocytes. J Immunol. (2008) 180:7423–30. doi: 10.4049/jimmunol.180.11.7423
56. Goodman, WA , Levine, AD , Massari, JV , Sugiyama, H , McCormick, TS , and Cooper, KD . Il-6 signaling in psoriasis prevents immune suppression by regulatory T cells. J Immunol. (2009) 183:3170–6. doi: 10.4049/jimmunol.0803721
57. Leipe, J , Grunke, M , Dechant, C , Reindl, C , Kerzendorf, U , Schulze-Koops, H, et al. Role of Th17 cells in human autoimmune arthritis. Arthritis Rheum. (2010) 62:2876–85. doi: 10.1002/art.27622
58. Bovenschen, HJ , van de Kerkhof, PC , van Erp, PE , Woestenenk, R , Joosten, I , and Koenen, HJ . Foxp3+ regulatory T cells of psoriasis patients easily differentiate into Il-17a-producing cells and are found in Lesional skin. J Invest Dermatol. (2011) 131:1853–60. doi: 10.1038/jid.2011.139
59. Kamiyama, T , Watanabe, H , Iijima, M , Miyazaki, A , and Iwamoto, S . Coexpression of Ccr6 and Cd146 (Mcam) is a marker of effector memory T-helper 17 cells. J Dermatol. (2012) 39:838–42. doi: 10.1111/j.1346-8138.2012.01544.x
60. Mommert, S , Gschwandtner, M , Koether, B , Gutzmer, R , and Werfel, T . Human memory Th17 cells express a functional histamine H4 receptor. Am J Pathol. (2012) 180:177–85. doi: 10.1016/j.ajpath.2011.09.028
61. Tsuda, K , Yamanaka, K , Kondo, M , Matsubara, K , Sasaki, R , Tomimoto, H, et al. Ustekinumab improves psoriasis without altering T cell cytokine production, differentiation, and T cell receptor repertoire diversity. PLoS One. (2012) 7:e51819. doi: 10.1371/journal.pone.0051819
62. Cheuk, S , Wiken, M , Blomqvist, L , Nylen, S , Talme, T , Stahle, M, et al. Epidermal Th22 and Tc17 cells form a localized disease memory in clinically healed psoriasis. J Immunol. (2014) 192:3111–20. doi: 10.4049/jimmunol.1302313
63. Skepner, J , Ramesh, R , Trocha, M , Schmidt, D , Baloglu, E , Lobera, M, et al. Pharmacologic inhibition of Rorgammat regulates Th17 signature gene expression and suppresses cutaneous inflammation in vivo. J Immunol. (2014) 192:2564–75. doi: 10.4049/jimmunol.1302190
64. Fiocco, U , Martini, V , Accordi, B , Caso, F , Costa, L , Oliviero, F, et al. Ex vivo signaling protein mapping in T lymphocytes in the psoriatic arthritis joints. J Rheumatol Suppl. (2015) 93:48–52. doi: 10.3899/jrheum.150636
65. Sgambelluri, F , Diani, M , Altomare, A , Frigerio, E , Drago, L , Granucci, F, et al. A role for Ccr5(+)Cd4 T cells in cutaneous psoriasis and for Cd103(+) Ccr4(+) Cd8 Teff cells in the associated systemic inflammation. J Autoimmun. (2016) 70:80–90. doi: 10.1016/j.jaut.2016.03.019
66. Soler, DC , Young, AB , Fiessinger, L , Galimberti, F , Debanne, S , Groft, S, et al. Increased, but functionally impaired, Cd14(+) Hla-Dr(−/low) myeloid-derived suppressor cells in psoriasis: a mechanism of dysregulated T cells. J Invest Dermatol. (2016) 136:798–808. doi: 10.1016/j.jid.2015.12.036
67. Yang, L , Li, B , Dang, E , Jin, L , Fan, X , and Wang, G . Impaired function of regulatory T cells in patients with psoriasis is mediated by phosphorylation of Stat3. J Dermatol Sci. (2016) 81:85–92. doi: 10.1016/j.jdermsci.2015.11.007
68. Diani, M , Galasso, M , Cozzi, C , Sgambelluri, F , Altomare, A , Cigni, C, et al. Blood to skin recirculation of Cd4(+) memory T cells associates with cutaneous and systemic manifestations of psoriatic disease. Clin Immunol. (2017) 180:84–94. doi: 10.1016/j.clim.2017.04.001
69. Khairutdinov, VR , Mikhailichenko, AF , Belousova, IE , Kuligina, ES , Samtsov, AV , and Imyanitov, EN . The role of intradermal proliferation of T-cells in the pathogenesis of psoriasis. An Bras Dermatol. (2017) 92:41–4. doi: 10.1590/abd1806-4841.20175765
70. Wang, XY , Chen, XY , Li, J , Zhang, HY , Liu, J , and Sun, LD . Mir-200a expression in Cd4+ T cells correlates with the expression of Th17/Treg cells and relevant cytokines in psoriasis vulgaris: a case control study. Biomed Pharmacother. (2017) 93:1158–64. doi: 10.1016/j.biopha.2017.06.055
71. Baricza, E , Marton, N , Kiralyhidi, P , Kovacs, OT , Kovacsne Szekely, I , Lajko, E, et al. Distinct in vitro T-helper 17 differentiation capacity of peripheral naive T cells in rheumatoid and psoriatic arthritis. Front Immunol. (2018) 9:606. doi: 10.3389/fimmu.2018.00606
72. Esmaeili, B , Mansouri, P , Meysamie, A , and Izad, M . Evaluation of Il-17 producing memory regulatory and effector T cells expressing Cd26 molecule in patients with psoriasis. Iran J Allergy Asthma Immunol. (2018) 17:453–63. doi: 10.18502/ijaai.v17i5.303
73. Gallais Sérézal, I , Classon, C , Cheuk, S , Barrientos-Somarribas, M , Wadman, E , Martini, E, et al. Resident T cells in resolved psoriasis steer tissue responses that stratify clinical outcome. J Invest Dermatol. (2018) 138:1754–63. doi: 10.1016/j.jid.2018.02.030
74. Bridgewood, C , Watad, A , Russell, T , Palmer, TM , Marzo-Ortega, H , Khan, A, et al. Identification of myeloid cells in the human Enthesis as the Main source of local Il-23 production. Ann Rheum Dis. (2019) 78:929–33. doi: 10.1136/annrheumdis-2018-214944
75. Diani, M , Casciano, F , Marongiu, L , Longhi, M , Altomare, A , Pigatto, PD, et al. Increased frequency of activated Cd8(+) T cell effectors in patients with psoriatic arthritis. Sci Rep. (2019) 9:10870. doi: 10.1038/s41598-019-47310-5
76. Esmaeili, B , Mansouri, P , Doustimotlagh, AH , and Izad, M . Redox imbalance and Il-17 responses in memory Cd4(+) T cells from patients with psoriasis. Scand J Immunol. (2019) 89:e12730. doi: 10.1111/sji.12730
77. Kampylafka, E , Simon, D , d'Oliveira, I , Linz, C , Lerchen, V , Englbrecht, M, et al. Disease interception with Interleukin-17 inhibition in high-risk psoriasis patients with subclinical joint inflammation-data from the prospective Ivepsa study. Arthritis Res Ther. (2019) 21:178. doi: 10.1186/s13075-019-1957-0
78. Kurihara, K , Fujiyama, T , Phadungsaksawasdi, P , Ito, T , and Tokura, Y . Significance of Il-17a-producing Cd8(+)Cd103(+) skin resident memory T cells in psoriasis lesion and their possible relationship to clinical course. J Dermatol Sci. (2019) 95:21–7. doi: 10.1016/j.jdermsci.2019.06.002
79. Vo, S , Watanabe, R , Koguchi-Yoshioka, H , Matsumura, Y , Ishitsuka, Y , Nakamura, Y, et al. Cd8 resident memory T cells with interleukin 17a-producing potential are accumulated in disease-Naïve Nonlesional sites of psoriasis possibly in correlation with disease duration. Br J Dermatol. (2019) 181:410–2. doi: 10.1111/bjd.17748
80. Casciano, F , Diani, M , Altomare, A , Granucci, F , Secchiero, P , Banfi, G, et al. Ccr4(+) skin-tropic phenotype as a feature of central memory Cd8(+) T cells in healthy subjects and psoriasis patients. Front Immunol. (2020) 11:529. doi: 10.3389/fimmu.2020.00529
81. de Jesús-Gil, C , Ruiz-Romeu, E , Ferran, M , Sagristà, M , Chiriac, A , García, P, et al. Il-15 and Il-23 synergize to trigger Th17 response by Cla(+) T cells in psoriasis. Exp Dermatol. (2020) 29:630–8. doi: 10.1111/exd.14113
82. Loyal, L , Warth, S , Jurchott, K , Molder, F , Nikolaou, C , Babel, N, et al. Slamf7 and Il-6r define distinct cytotoxic versus helper memory Cd8(+) T cells. Nat Commun. (2020) 11:6357. doi: 10.1038/s41467-020-19002-6
83. Mashiko, S , Edelmayer, RM , Bi, Y , Olson, LM , Wetter, JB , Wang, J, et al. Persistence of inflammatory phenotype in residual psoriatic plaques in patients on effective biologic therapy. J Invest Dermatol. (2020) 140:1015–1025.e4. doi: 10.1016/j.jid.2019.09.027
84. Wang, J , Zhang, SX , Hao, YF , Qiu, MT , Luo, J , Li, YY, et al. The numbers of peripheral regulatory T cells are reduced in patients with psoriatic arthritis and are restored by low-dose Interleukin-2. Ther Adv Chronic Dis. (2020) 11:2040622320916014. doi: 10.1177/2040622320916014
85. Kasprowicz-Furmanczyk, M , Czerwinska, J , Placek, W , and Owczarczyk-Saczonek, A . Assessment of the tissue resident memory cells in Lesional skin of patients with psoriasis and in healthy skin of healthy volunteers. Int J Environ Res Public Health. (2021) 18:12. doi: 10.3390/ijerph182111251
86. Leijten, EF , van Kempen, TS , Olde Nordkamp, MA , Pouw, JN , Kleinrensink, NJ , Vincken, NL, et al. Tissue-resident memory Cd8+ T cells from skin differentiate psoriatic arthritis from psoriasis. Arthritis Rheumatol. (2021) 73:1220–32. doi: 10.1002/art.41652
87. Mehta, H , Mashiko, S , Angsana, J , Rubio, M , Hsieh, YM , Maari, C, et al. Differential changes in inflammatory mononuclear phagocyte and T-cell profiles within psoriatic skin during treatment with Guselkumab vs Secukinumab. J Invest Dermatol. (2021) 141:1707–1718.e9. doi: 10.1016/j.jid.2021.01.005
88. Mulder, MLM , He, X , van den Reek, J , Urbano, PCM , Kaffa, C , Wang, X, et al. Blood-based immune profiling combined with machine learning discriminates psoriatic arthritis from psoriasis patients. Int J Mol Sci. (2021) 22:10990. doi: 10.3390/ijms222010990
89. Nguyen, DX , Baldwin, HM , Ezeonyeji, AN , and Butt, MR . Ehrenstein MR. regulatory T cells enhance Th17 migration in psoriatic arthritis which is reversed by anti-Tnf. iScience. (2021) 24:102973. doi: 10.1016/j.isci.2021.102973
90. Phadungsaksawasdi, P , Fujiyama, T , Kurihara, K , Ito, T , Honda, T , and Tokura, Y . Pd-1 expression defines epidermal Cd8(+)Cd103(+) T cells preferentially producing Il-17a and using skewed Tcr repertoire in psoriasis. J Invest Dermatol. (2021) 141:2426–2435.e5. doi: 10.1016/j.jid.2021.03.011
91. Raychaudhuri, SK , Abria, C , and Raychaudhuri, SP . Phenotype and pathological significance of Mcam(+) (Cd146(+)) T cell subset in psoriatic arthritis. Mol Biol Rep. (2021) 48:6787–96. doi: 10.1007/s11033-021-06678-2
92. Cook, CP , Taylor, M , Liu, Y , Schmidt, R , Sedgewick, A , Kim, E, et al. A single-cell transcriptional gradient in human cutaneous memory T cells restricts Th17/Tc17 identity. Cell Rep Med. (2022) 3:100715. doi: 10.1016/j.xcrm.2022.100715
93. Kasprowicz-Furmanczyk, M , Czerwinska, J , Placek, W , and Owczarczyk-Saczonek, A . The effect of the Long-term Calcipotriol/betamethasone Dipropionate local therapy on tissue resident memory cells markers in psoriatic eruptions. Int J Environ Res Public Health. (2022) 19:12. doi: 10.3390/ijerph19148345
94. Michalak-Stoma, A , Bartosińska, J , Raczkiewicz, D , Kowal, M , Kozak, J , Gujski, M, et al. Multiple cytokine analysis of Th1/Th2/Th9/Th17/Th22/Treg cytokine pathway for individual immune profile assessment in patients with psoriasis. Med Sci Monit. (2022) 28:e938277. doi: 10.12659/msm.938277
95. Owczarczyk-Saczonek, A , Kasprowicz-Furmanczyk, M , Czerwinska, J , Krajewska-Wlodarczyk, M , and Placek, W . The effect of therapy on Trm in psoriatic lesions. Postep Dermatol Alergol. (2022) 39:209–20. doi: 10.5114/ada.2021.113125
96. Skougaard, M , Ditlev, SB , Stisen, ZR , Coates, LC , Ellegaard, K , and Kristensen, LE . Four emerging immune cellular blood phenotypes associated with disease duration and activity established in psoriatic arthritis. Arthritis Res Ther. (2022) 24:262. doi: 10.1186/s13075-022-02956-x
97. Ghaffarinia, A , Ayaydin, F , Poliska, S , Manczinger, M , Bolla, BS , Flink, LB, et al. Psoriatic resolved skin epidermal keratinocytes retain disease-residual transcriptomic and Epigenomic profiles. Int J Mol Sci. (2023) 24:20. doi: 10.3390/ijms24054556
98. Owczarczyk Saczonek, A , Krajewska-Włodarczyk, M , Kasprowicz-Furmańczyk, M , and Placek, W . Immunological memory of psoriatic lesions. Int J Mol Sci. (2020) 21:625. doi: 10.3390/ijms21020625
99. Eyerich, K , Weisenseel, P , Pinter, A , Schäkel, K , Asadullah, K , Wegner, S, et al. Il-23 blockade with Guselkumab potentially modifies psoriasis pathogenesis: rationale and study protocol of a phase 3b, randomised, double-blind, multicentre study in participants with moderate-to-severe plaque-type psoriasis (guide). BMJ Open. (2021) 11:e049822. doi: 10.1136/bmjopen-2021-049822
100. Schäkel, K , Reich, K , Asadullah, K , Pinter, A , Jullien, D , Weisenseel, P, et al. Early disease intervention with Guselkumab in psoriasis leads to a higher rate of stable complete skin clearance ('Clinical super Response'): week 28 results from the ongoing phase Iiib randomized, double-blind, parallel-group, guide study. J Eur Acad Dermatol Venereol. (2023) 37:2016–27. doi: 10.1111/jdv.19236
101. Zabotti, A , De Marco, G , Gossec, L , Baraliakos, X , Aletaha, D , Iagnocco, A, et al. Eular points to consider for the definition of clinical and imaging features suspicious for progression from psoriasis to psoriatic arthritis. Ann Rheum Dis. (2023) 82:ard-2023-224148–1170. doi: 10.1136/ard-2023-224148
102. Scher, JU , Ogdie, A , Merola, JF , and Ritchlin, C . Preventing psoriatic arthritis: focusing on patients with psoriasis at increased risk of transition. Nat Rev Rheumatol. (2019) 15:153–66. doi: 10.1038/s41584-019-0175-0
103. Singla, S , Putman, M , Liew, J , and Gordon, K . Association between biological immunotherapy for psoriasis and time to incident inflammatory arthritis: a retrospective cohort study. Lancet Rheumatol. (2023) 5:e200–7. doi: 10.1016/S2665-9913(23)00034-6
104. Watanabe, R , Gehad, A , Yang, C , Scott, LL , Teague, JE , Schlapbach, C, et al. Human skin is protected by four functionally and phenotypically discrete populations of resident and recirculating memory T cells. Sci Transl Med. (2015) 7:279ra39. doi: 10.1126/scitranslmed.3010302
105. Mackay, LK , Rahimpour, A , Ma, JZ , Collins, N , Stock, AT , Hafon, ML, et al. The developmental pathway for Cd103(+)Cd8+ tissue-resident memory T cells of skin. Nat Immunol. (2013) 14:1294–301. doi: 10.1038/ni.2744
106. Günther, C , Carballido-Perrig, N , Kaesler, S , Carballido, JM , and Biedermann, T . Cxcl16 and Cxcr6 are upregulated in psoriasis and mediate cutaneous recruitment of human Cd8+ T cells. J Invest Dermatol. (2012) 132:626–34. doi: 10.1038/jid.2011.371
107. Johnsson, H , Cole, J , Siebert, S , McInnes, IB , and Graham, G . Cutaneous lesions in psoriatic arthritis are enriched in chemokine transcriptomic pathways. Arthritis Res Ther. (2023) 25:73. doi: 10.1186/s13075-023-03034-6
108. Slauenwhite, D , Gebremeskel, S , Doucette, CD , Hoskin, DW , and Johnston, B . Regulation of cytokine polarization and T cell recruitment to inflamed paws in mouse collagen-induced arthritis by the chemokine receptor Cxcr6. Arthritis Rheumatol. (2014) 66:3001–12. doi: 10.1002/art.38816
109. Haberman, RH , MacFarlane, KA , Catron, S , Samuels, J , Blank, RB , Toprover, M, et al. Efficacy of Guselkumab, a selective Il-23 inhibitor, in preventing arthritis in a multicentre psoriasis at-risk cohort (Pampa): protocol of a randomised, double-blind, placebo controlled multicentre trial. BMJ Open. (2022) 12:e063650. doi: 10.1136/bmjopen-2022-063650
Keywords: psoriasis, psoriatic arthritis, disease interception, resident-memory T cells, regulatory T cells, IL-17, IL-23
Citation: Lobão B, Lourenço D, Giga A and Mendes-Bastos P (2024) From PsO to PsA: the role of TRM and Tregs in psoriatic disease, a systematic review of the literature. Front. Med. 11:1346757. doi: 10.3389/fmed.2024.1346757
Edited by:
Manuel Pedro Pereira, Charité – University Medicine Berlin, GermanyReviewed by:
Luis Puig, Autonomous University of Barcelona, SpainDemetrios Christou, Charité – University Medicine Berlin, Germany
Copyright © 2024 Lobão, Lourenço, Giga and Mendes-Bastos. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Pedro Mendes-Bastos, cG1lbmRlc2Jhc3Rvc0BnbWFpbC5jb20=