 Lujuan Gao
Lujuan Gao Yi Sun
Yi Sun Chengyan He3
Chengyan He3- 1Department of Dermatology, Zhongshan Hospital Fudan University, Shanghai, China
- 2Department of Dermatology, Jingzhou Central Hospital, The Second Clinical Medical College, Yangtze University, Jingzhou, China
- 3The Second Clinical Medical College, Yangtze University, Jingzhou, China
- 4Department of Dermatology, Central Hospital of Xiangyang (Affiliated Hospital of Hubei College of Arts and Sciences), Xiangyang, China
Infections of Exophiala spp. and Fusarium spp. are often chronic and recalcitrant. Systemic disseminations, which mostly occur in immunocompromised patients, are often refractory to available antifungal therapies. The conserved target of rapamycin (TOR) orchestrates cell growth and proliferation in response to nutrients and growth factors, which are important for pathogenicity and virulence. INK128 is a second-generation ATP-competitive TOR inhibitor, which binds the TOR catalytic domain and selectively inhibits TOR. In the present study, we investigated the in vitro activities of INK128 alone and the interactions of INK128 with conventional antifungal drugs including itraconazole, voriconazole, posaconazole, and amphotericin B against 18 strains of Exophiala spp. and 10 strains of Fusarium spp. via broth microdilution checkerboard technique system adapted from Clinical and Laboratory Standards Institute broth microdilution method M38-A2. INK128 alone was inactive against all isolates tested. However, favorable synergistic effects between INK128 and voriconazole were observed in 61% Exophiala strains and 60% Fusarium strains, despite Fusarium strains exhibited high MIC values (4–8 μg/ml) against voriconazole. In addition, synergistic effects of INK128/itraconazole were shown in 33% Exophiala strains and 30% Fusarium strains, while synergy of INK128/posaconazole were observed in 28% Exophiala strains and 30% Fusarium strains. The effective working ranges of INK128 were 0.125–2 μg/ml and 1–4 μg/ml against Exophiala isolates and Fusarium isolates, respectively. No synergistic effect was observed when INK128 was combined with amphotericin B. No antagonism was observed in all combinations. In conclusion, INK128 could enhance the in vitro antifungal activity of voriconazole, itraconazole and posaconazole against Exophiala spp. and Fusarium spp., suggesting that azoles, especially voriconazole, combined with TOR kinase inhibitor might provide a potential strategy to the treatment of Exophiala and Fusarium infections. However, further investigations are warranted to elucidate the underlying mechanism and to determine possible reliable and safe application in clinical practice.
Introduction
Exophiala spp. and Fusarium spp. are both increasingly recognized opportunistic pathogen causing cutaneous, subcutaneous and serious invasive infections, especially in immunocompromised and debilitated individuals (Li et al., 2011; Guarro, 2013). Human infection usually occurs as a result of inoculation of the organism through the body surface causing local infection. E. dermatitidis is one of the most common cause of chromoblastomycosis (Li et al., 2011), while Fusarium spp. causes keratitis and onychomycosis, or locally invasive infections (Guarro, 2013). However, disease in immunocompromised patients often manifests with systemic disseminated fungemia, whose prevalence is effectively growing (Li et al., 2011; Guarro, 2013). E. dermatitidis is the leading cause of severe neurotropic phaeohyphomycosis (Li et al., 2011), while fusariosis is, after aspergillosis, the second most common mold infection in humans, among which F. solani species complex and F. oxysporum species complex are responsible for approximately 80% of the cases (Guarro, 2013).
Prompt antifungal treatment is crucial to prevent life-threatening disease. However, fusariosis is mostly refractory to available treatment, with a high mortality rate for systemic disseminations, which is in accordance with the poor in vitro activities of available antifungal drugs against Fusarium spp. (Guarro, 2013). In addition, success rate for Exophiala spp. infection was only 40–70% despite most antifungal drugs showed favorable in vitro activities (Revankar and Sutton, 2010; Kondori et al., 2011; Patel et al., 2013). Optimal treatment remains elusive. Therefore, novel therapeutic strategies are desperately needed.
The target of rapamycin (TOR), which is a conserved serine/threonine kinase in eukaryotes from yeast to humans and orchestrates cell growth and proliferation in response to nutrients and growth factors, is a promising target for the development of novel antifungal strategy (Crespo and Hall, 2002). It has been demonstrated that the TOR pathway regulates cellular responses to nutrients in yeast cells, including proliferation, translation, transcription, autophagy, ribosome biogenesis, lipid homeostasis, morphogenesis and cellular aggregation, which have important implications for pathogenicity and virulence (Crespo and Hall, 2002; Madeira et al., 2015). Therefore, the TOR signaling cascade is an excellent target for the development of broad-spectrum antifungal agents. INK128 is a second-generation ATP-competitive TOR inhibitor, which binds the TOR catalytic domain and selectively inhibits TOR (Hsieh et al., 2012). Previous study revealed synergistic effects between INK128 and itraconazole (ITC), voriconazole (VRC), posaconazole (POS) against Aspergillus spp. (Gao et al., 2016). Thus, it is reasonable to speculate that INK128 might also have some antifungal activity and interactions with antifungals against Exophiala spp. and Fusarium spp.
The aim of this study was to evaluate the effects of INK128 alone and combined with antifungal agents against Exophiala spp. and Fusarium spp.
Materials and Methods
Fungal Strains
A total of 18 strains of E. dermatitidis and 10 strains of Fusarium spp. (7 strains of F. solani, and 3 strains of F. oxysporum) were studied. All strains were clinical isolates. Fungal identification was determined by microscopic morphology and by molecular sequencing of the internal transcribed spacer (ITS) ribosomal DNA (rDNA), as required. Candida parapsilosis ATCC 22019 was included to ensure quality control.
Antifungals and Chemical Agents
All tested drugs including INK128 (purity ≥ 99%), ITC (purity ≥ 99%), VRC (purity ≥ 99%), POS (purity ≥ 99%), and amphotericin B (AMB; purity ≥ 80%) were purchased in powder form from Selleck Chemicals, Houston, TX, USA and prepared as outlined in the Clinical and Laboratory Standards Institute (CLSI) broth microdilution method M38-A2 (Clinical and Laboratory Standards Institute, 2008). The working concentration ranges of tested drugs were all 0.03–16 μg/ml.
Inoculum Preparation
Conidia harvested from cultures grown for 7 days on Sabouraud dextrose agar (SDA) were suspended in sterile distilled water containing 0.03% Triton and diluted to a concentration of 1–5 × 106 spores/ml, which were than diluted 100 times in RPMI-1640 to achieve a 2-fold suspension more concentrated than the density needed or to approximately 1–5 × 104 spores/ml (Clinical and Laboratory Standards Institute, 2008).
In vitro Antifungal Activity of Individual Tested Agents
The individual minimal inhibitory concentrations (MICs) of INK128, ITC, VRC, POS and AMB were determined according to M38-A2 method (Clinical and Laboratory Standards Institute, 2008). The 96-well plate was inoculated with 100 μl of the inoculum suspension prepared and 100 μl of the serial diluent of tested drugs. Interpretation of results was performed after incubation at 35°C for 48 h for Fusarium spp. and 72 h for Exophiala spp., respectively. All tests were performed in triplicate.
In vitro Interactions of INK128 and Antifungals Agents
The interactions between INK128 and antifungal agents against all strains were tested via the microdilution chequerboard technique, adapted from the CLSI M38-A2 microdilution method (Clinical and Laboratory Standards Institute, 2008). As described, a 50 μl of INK128 with serial dilutions were inoculated in horizontal direction and another 50 μl of azoles or AMB with serial dilutions were inoculated in vertical direction on the 96-well plate, which contained 100 μl prepared inoculum suspension. Interpretation of results was performed after incubation at 35°C for 48 h for Fusarium spp. and 72 h for Exophiala spp., respectively. Drug combination interaction was classified on the basis of the fractional inhibitory concentration index (FICI). The FICI as calculated by the formula: FICI = (Ac/Aa) + (Bc/Ba), where Ac and Bc are the MICs of antifungal drugs in combination, and Aa and Ba are the MICs of antifungal drugs A and B alone (Tobudic et al., 2010). All tests were performed in triplicate.
Interpretation of Results
The MICs were determined as the lowest concentration resulting in complete inhibition of growth (Clinical and Laboratory Standards Institute, 2008). The interaction of INK128 with azoles, or AMB referred to the fractional inhibitory concentration index (FICI), which was classified as follows: FICI of ≤0.5, synergy; FICI of >0.5 to ≤4, no interaction (indifference); FICI of >4, antagonism (Odds, 2003).
Results
In vitro Antifungal Activity of Individual Tested Agent
The MIC ranges of individual tested drugs against E. dermatitidis isolates were >16 μg/ml for INK128, 0.5–1 μg/ml for ITC, 0.25–0.5 μg/ml for VRC, 0.25–1 μg/ml for POS, and 0.5–1 μg/ml for AMB, respectively (Table 1). The MIC ranges against Fusarium spp. are >16 μg/ml for INK128 and ITC, 4–8 μg/ml for VRC and POS, and 2–4 μg/ml for AMB, respectively (Table 2). INK128 individually did not exhibit any significant antifungal activity against all tested strains of Fusarium spp. and Exophiala spp.
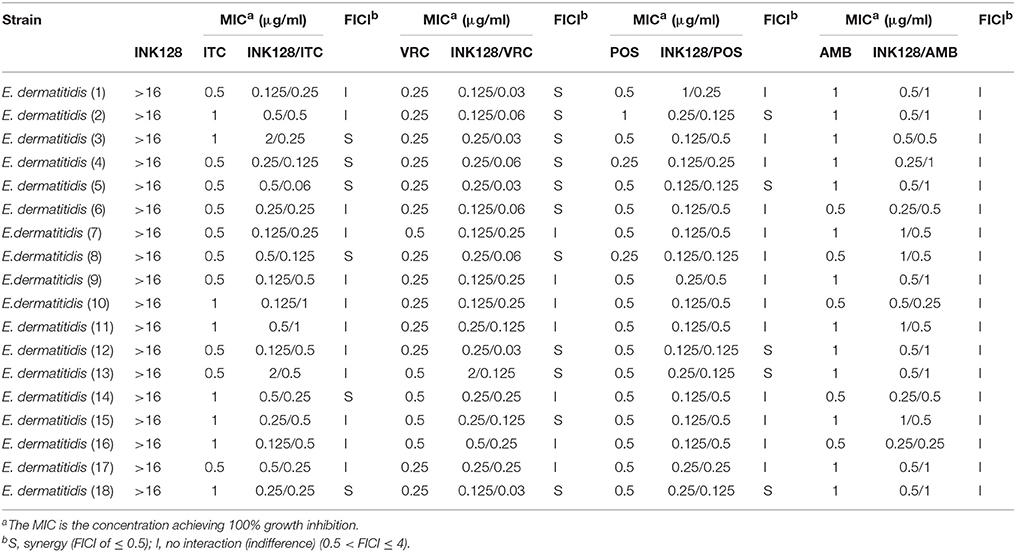
Table 1. MICs and FICIs results with combinations of INK128 with antifungal agents against E. dermatitidis.
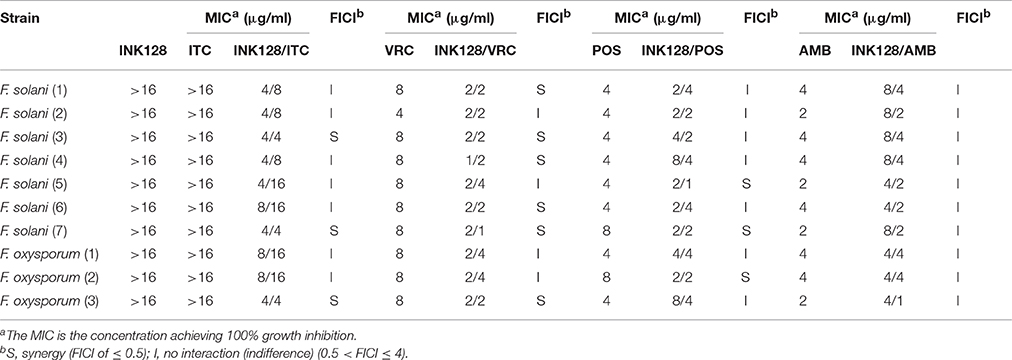
Table 2. MICs and FICIs results with combinations of INK128 with antifungal agents against Fusarium spp.
In vitro Interactions between INK128 and Antifungal Agents
When INK128 was combined with VRC, the MICs of INK128 and VRC against Exophiala spp. decreased to 0.125–2 μg/ml and 0.03–0.25 μg/ml, respectively, demonstrating favorable synergistic effects against 11 (61%) strains of E. dermatitidis (Table 1). Similarly, this INK128/VRC combination also showed favorable synergism against 6 (60%) strains of Fusarium isolates (Table 2), where the MIC ranges of INK128 and VRC decreased to 1–2 μg/ml and 1–4 μg/ml, respectively. The effective working ranges of INK128 in this combination were 0.125–2 μg/ml and 1-2 μg/ml against Exophiala spp. and Fusarium spp., respectively.
When INK128 was combined with POS, the MIC ranges of INK128 and POS against Exophiala spp. decreased to 0.125–1 μg/ml and 0.125–0.5 μg/ml, respectively (Table 1). The MIC ranges of INK128 and POS against Fusarium spp. decreased to 2–8 μg/ml and 1–4 μg/ml, respectively (Table 2). The INK128/POS combination revealed synergistic effects against only 5 (28%) strains of Exophiala spp. and 3(30%) strains of Fusarium spp. (Tables 1, 2). The effective working ranges of INK128 in INK128/POS combination were 0.125–0.25 μg/ml and 2 μg/ml against Exophiala spp. and Fusarium spp., respectively.
When INK128 was combined with ITC, the MIC ranges of INK128 and ITC against Exophiala spp. decreased to 0.125–2 μg/ml and 0.06–1 μg/ml, respectively. The MIC ranges of INK128 and ITC against Fusarium spp. decreased to 4–8 μg/ml and 4–16 μg/ml, respectively. Synergistic effects of the INK128/ITC combination were only observed in only 6 (33%) strains of Exophiala spp. and 3(30%) strains of Fusarium spp. (Tables 1, 2). The effective working ranges of INK128 in INK128/ITC combination were 0.25–2 μg/ml and 4 μg/ml against Exophiala spp. and Fusarium spp., respectively.
No synergistic effect was observed when INK128 was combined with AMB. No antagonism was observed in these combinations.
Discussion
TOR kinase is the central element of TOR signaling pathway, which has been widely investigated for years since its discovery and has been recognized as a central controller of cell growth in eukaryotes (Crespo and Hall, 2002). The pharmaceutical potential of rapamycin, the classical allosteric inhibitor of TOR, was originally discovered in a screen for novel antifungal agents. Rapamycin was found to exhibit potent antifungal effects against a variety of species including Candida spp., Cryptococcus spp., Aspergillus spp., Fusarium spp., Penicillium spp., and dermtophytes (Rohde and Cardenas, 2004). However, rapamycin failed to be used as an antifungal due to its even more potent immunosuppressive property.
INK128, a highly potent, orally active TOR kinase inhibitor, was originally developed for cancer treatment (Hsieh et al., 2012). It has been demonstrated that oral administration of INK128 in mice has high absorption and bioavailability, with doses of 3 mg/kg giving a Cmax of 0.599 μg/ml in plasma (Hsieh et al., 2012). Moreover, previous study has shown that INK128 did not inhibit in vitro peripheral blood lymphocytes proliferation at concentrations of up to 1 μM (0.31 μg/ml), and daily administration of INK128 at a dose up to 5 mg/kg in humanized mice over a 2-week period showed no obvious toxicity (Heredia et al., 2015), suggesting that INK128 is not as immunosuppressive as rapamycin.
Previously, we have demonstrated that INK128 exhibited synergistic effects with ITC (65%), VRC (61%), POS (50%) against Aspergillus spp. (Gao et al., 2016). In the present study, we investigated the in vitro antifungal activity of INK128 alone and combined with antifungal agents against Exophiala spp. and Fusarium spp. The results revealed that INK128 alone was inactive against all strains tested, as was demonstrated against Aspergillus isolates (Gao et al., 2016). Nevertheless, synergistic activities between INK128 and VRC (61%), ITC (33%), POS (28%) were observed in Exophiala spp. Although, the MIC values of ITC, VRC and POS against Fusarium strains were much higher that those against Exophiala spp., synergistic effects between INK128 and VRC (60%), ITC(30%), POS (30%) against Fusarium isolates were comparable to those of Exophiala spp. The effective working ranges of INK128 were 0.125–2 μg/ml and 1–4 μg/ml against Exophiala spp. and Fusarium spp., respectively. No interaction between INK128 and AMB was observed. Moreover, no antagonism was observed.
Synergistic effects between INK128 and ITC or POS against Exophiala spp. and Fusarium spp. were less frequent than those observed in Aspergillus spp. (Gao et al., 2016). However, synergy between INK128 and VRC was comparable among Exophiala spp., Fusarium spp., and previously tested Aspergillus spp. (Gao et al., 2016), despite Fusarium spp. showed relatively poor antifungal susceptibility profile. It is important to note that even with high MICs against VRC (4–8 μg/ml), synergy between INK128 and VRC was observed in up to 60% Fusarium strains, implicating that INK128 could enhance the in vitro susceptibility of VRC-inactive Fusarium strains. The effective working ranges of INK128 against Exophiala spp. were mostly within 0.125–0.25 μg/ml, which could be achieved safely as mentioned above. Compared to Exophiala spp., the working ranges of INK128 against Fusarium spp. were higher, which might associated with the inherently high MICs of Fusarium spp. to most available antifungal medications.
Previous studies have demonstrated synergistic interactions between rapamycin and AMB (70%), ITC (50%), POS (40%) against Mucorales (previously referred to as zygomycetes) through broth microdilution checkerboard procedure (Dannaoui et al., 2009). Futhermore, antagonism of rapamycin/ITC, and indifference to significant antagonism of rapamycin/POS against Mucorales (previously referred to as zygomycetes) have also been reported (Dannaoui et al., 2009; Narreddy et al., 2010). However, there was no interaction between INK128 and AMB against tested Exophiala spp. and Fusarium spp. in the present study, or Aspergillus spp. in the previous study (Gao et al., 2016). No antagonism between INK128 and POS or ITC was observed. The explanation of the difference between INK128 and rapamycin might lie in the different mechanism through which INK128 and rapamycin interact with TOR and antifungals, and different response of tesed species against these antifungal drugs.
In summary, the present study extends previous findings in the combination interactions between TOR inhibitors, especially INK128, and conventional antifungals. INK128 could enhance the in vitro antifungal activity of VRC, ITC, and POS against Exophiala spp. and Fusarium spp. More importantly, the inexistence of immunosuppression of INK128 suggests that the combination of azoles with INK128 might provide a safe alternative strategy to the treatment of Exophiala and Fusarium infections. However, further investigations are warranted to elucidate the underlying mechanism and to determine possible reliable and safe application in clinical practice.
Author Contributions
LG and YS: Conceived and designed the study; CH and YS: Performed all the experiments; LG and QL: Analyzed the data and wrote the manuscript; ML and TZ: Provided general guidance and revised the manuscript.
Funding
This work was supported by grants 31400131 (LG) and 81401677 (YS) from National Natural Science Foundation of China, grant 2015ZSYXQN21 from Outstanding Youth Project of Zhongshan Hospital Fudan University (LG) and grant WJ2015MB281 from Hubei Province Health and Family Planning Scientific Research Project (YS).
Conflict of Interest Statement
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Acknowledgments
We thank Professor Ruoyu Li and Professor Wei Liu from Peking University First Hospital, Research Center for Medical Mycology, Peking University, Beijing; Professor Qiangqiang Zhang from Fudan University Huashan Hospital, Shanghai, and Professor G. Sybren de Hoog from CBS-KNAW Fungal Biodiversity Centre, Utrecht, The Netherlands for kindly provided us with isolates studied.
References
Clinical Laboratory Standards Institute (2008). Reference Method for Broth Dilution Antifungal Susceptibility Testing of Filamentous Fungi. Approved Standard M38-A2. Wayne, PA: CLSI.
Crespo, J. L., and Hall, M. N. (2002). Elucidating TOR signaling and rapamycin action: lessons from Saccharomyces cerevisiae. Microbiol. Mol. Biol. Rev. 66, 579–591. doi: 10.1128/MMBR.66.4.579-591.2002
Dannaoui, E., Schwarz, P., and Lortholary, O. (2009). In vitro interactions between antifungals and immunosuppressive drugs against zygomycetes. Antimicrob. Agents Chemother. 53, 3549–3551. doi: 10.1128/AAC.00184-09
Gao, L., Ding, X., Liu, Z., Wu, Q., Zeng, T., and Sun, Y. (2016). In vitro interactions between target of rapamycin kinase inhibitor and antifungal agents against Aspergillus species. Antimicrob. Agents Chemother. 60, 3813–3816. doi: 10.1128/AAC.02921-15
Guarro, J. (2013). Fusariosis, a complex infection caused by a high diversity of fungal species refractory to treatment. Eur. J. Clin. Microbiol. Infect. Dis. 32, 1491–1500. doi: 10.1007/s10096-013-1924-7
Heredia, A., Le, N., Gartenhaus, R. B., Sausville, E., Medina-Moreno, S., Zapata, J. C., et al. (2015). Targeting of mTOR catalytic site inhibits multiple steps of the HIV-1 lifecycle and suppresses HIV-1 viremia in humanized mice. Proc. Natl. Acad. Sci. U.S.A. 112, 9412–9417. doi: 10.1073/pnas.1511144112
Hsieh, A. C., Liu, Y., Edlind, M. P., Ingolia, N. T., Janes, M. R., Sher, A., et al. (2012). The translational landscape of mTOR signalling steers cancer initiation and metastasis. Nature 485, 55–61. doi: 10.1038/nature10912
Kondori, N., Gilljam, M., Lindblad, A., Jonsson, B., Moore, E. R., and Wenneras, C. (2011). High rate of Exophiala dermatitidis recovery in the airways of patients with cystic fibrosis is associated with pancreatic insufficiency. J. Clin. Microbiol. 49, 1004–1009. doi: 10.1128/JCM.01899-10
Li, D. M., Li, R. Y., de Hoog, G. S., Sudhadham, M., and Wang, D. L. (2011). Fatal Exophiala infections in China, with a report of seven cases. Mycoses 54, e136–e142. doi: 10.1111/j.1439-0507.2010.01859.x
Madeira, J. B., Masuda, C. A., Maya-Monteiro, C. M., Matos, G. S., Montero-Lomeli, M., and Bozaquel-Morais, B. L. (2015). TORC1 inhibition induces lipid droplet replenishment in yeast. Mol. Cell. Biol. 35, 737–746. doi: 10.1128/MCB.01314-14
Narreddy, S., Manavathu, E., Chandrasekar, P. H., Alangaden, G. J., and Revankar, S. G. (2010). In vitro interaction of posaconazole with calcineurin inhibitors and sirolimus against zygomycetes. J. Antimicrob. Chemother. 65, 701–703. doi: 10.1093/jac/dkq020
Odds, F. C. (2003). Synergy, antagonism, and what the chequerboard puts between them. J. Antimicrob. Chemother. 52:1. doi: 10.1093/jac/dkg301
Patel, A. K., Patel, K. K., Darji, P., Singh, R., Shivaprakash, M. R., and Chakrabarti, A. (2013). Exophiala dermatitidis endocarditis on native aortic valve in a postrenal transplant patient and review of literature on E. dermatitidis infections. Mycoses 56, 365–372. doi: 10.1111/myc.12009
Revankar, S. G., and Sutton, D. A. (2010). Melanized fungi in human disease. Clin. Microbiol. Rev. 23, 884–928. doi: 10.1128/CMR.00019-10
Rohde, J. R., and Cardenas, M. E. (2004). Nutrient signaling through TOR kinases controls gene expression and cellular differentiation in fungi. Curr. Top. Microbiol. Immunol. 279, 53–72. doi: 10.1007/978-3-642-18930-2_4
Keywords: INK128, TOR inhitibor, synergy, Fusarium, Exophiala, voriconazole, itraconazole, posaconazole
Citation: Gao L, Sun Y, He C, Li M, Zeng T and Lu Q (2016) INK128 Exhibits Synergy with Azoles against Exophiala spp. and Fusarium spp. Front. Microbiol. 7:1658. doi: 10.3389/fmicb.2016.01658
Received: 02 August 2016; Accepted: 05 October 2016;
Published: 20 October 2016.
Edited by:
Octavio Luiz Franco, Universidade Católica de Brasília, BrazilReviewed by:
Suzana Meira Ribeiro, Universidade Católica Dom Bosco (UCDB), BrazilCamila Guimarães Freitas, Federal Institute of Brasilia, Brazil
Copyright © 2016 Gao, Sun, He, Li, Zeng and Lu. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) or licensor are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Lujuan Gao, gao_lujuan@163.com
Yi Sun, jzzxyysy@163.com
†These authors have contributed equally to this work.
‡Joint first authors.