 Manlin Xu
Manlin Xu Hongfeng Xie
Hongfeng Xie Juxiang Wu1
Juxiang Wu1 Yucheng Chi
Yucheng Chi- 1Shandong Peanut Research Institute, Qingdao, China
- 2Fujian Agriculture and Forestry University, Fuzhou, China
- 3Open Project Program of Key Laboratory of Tobacco Pest Monitoring Controlling and Integrated Management, Tobacco Research Institute of Chinese Academy of Agricultural Sciences, Qingdao, China
Peanut stripe virus (PStV) belongs to the genus Potyvirus and is the most important viral pathogen of cultivated peanut (Arachis hypogaea L.). The eukaryotic translation initiation factor, eIF4E, and its isoform, eIF(iso)4E, play key roles during virus infection in plants, particularly Potyvirus. In the present study, we cloned the eIF4E and eIF(iso)4E homologs in peanut and named these as PeaeIF4E and PeaeIF(iso)4E, respectively. Quantitative real-time PCR (qRT-PCR) analysis showed that these two genes were expressed during all growth periods and in all peanut organs, but were especially abundant in young leaves and roots. These also had similar expression levels. Yeast two-hybrid analysis showed that PStV multifunctional helper component proteinase (HC-Pro) and viral protein genome-linked (VPg) both interacted with PeaeIF4E and PeaeIF(iso)4E. Bimolecular fluorescence complementation assay showed that there was an interaction between HC-Pro and PeaeIF4E/PeaeIF(iso)4E in the cytoplasm and between VPg and PeaeIF4E/PeaeIF(iso)4E in the nucleus. Silencing either PeaeIF4E or PeaeIF(iso)4E using a virus-induced gene silencing system did not significantly affect PStV accumulation. However, silencing both PeaeIF4E and PeaeIF(iso)4E genes significantly weakened PStV accumulation. The findings of the present study suggest that PeaeIF4E and PeaeIF(iso)4E play important roles in the PStV infection cycle and may potentially contribute to PStV resistance.
Introduction
Peanut is one of the most important oil crops and food legumes in the world. In China, peanuts are grown on 3.5 million hectares of land each year (Xu, 2008). Peanut stripe virus (PStV; genus Potyvirus, family Potyviridae) is one of the most widely distributed peanut viruses constraining peanut production. PStV has been detected in various countries, including China, the US, the Philippines, Thailand, Indonesia, Malaysia, and Korea (Xu et al., 1983; Demski and Lovell, 1985; Saleh et al., 1989; Choi et al., 2001, 2006). Recently, PStV has been reported in India and some African countries, which were possibly caused by exchanges in peanut seed resources (Xu, 2008).
In China, PStV is a very serious infectious disease that afflicts peanut, particularly those grown in northern China. The infection incidence has reached 50% and recently, even 100% in some fields. PStV has also infected various other crops, including soybean (Glycine max), sesame (Sesamum indicum), cowpea (Vigna unguiculata), hyacinth bean (Dolichos lablab), white lupin (Lupinus albus), and patchouli (Pogostemon cablin; Xu, 2008; Singh et al., 2009). To date, no effective method for controlling this virus has been established.
Peanut stripe virus is a member of the genus Potyvirus, an economically significant and one of the largest groups of viruses that infect plants. These viruses are about 10 kb in length, carry a single positive-strand RNA, and contain a 350-kD polyprotein that is translated by a single open reading frame (ORF). The polyprotein is cleaved by three virus-encoded proteases into 10 mature proteins and an additional protein called PIPO, which is embedded in the P3 cistron (Urcuqui-Inchima et al., 2001; Wei et al., 2010).
One of the three virus-encoded proteases is the multifunctional helper component proteinase (HC-Pro), which consists of C-proximal, central, and N-proximal domains. The C-proximal domain separates HC-Pro from the polyprotein precursor via proteolysis (Carrington and Herndon, 1992). HC-Pro contributes to various essential steps that are related to viral replication and infection cycles. HC-Pro is involved in some processes, including virus transmission by aphids (Govier et al., 1977) and virus movement from cell-to-cell (Rojas et al., 1997) to long-distance migration (Saenz et al., 2002). In addition, HC-Pro facilitates the development of virulence and symptom amplification (Atreya et al., 1992; Redondo et al., 2001); it is also a regulator of gene silencing suppression (Llave et al., 2000). HC-Pro interacts with numerous host proteins and some virus proteins, as well as mediates the function of host proteins and other viral proteins (Jin et al., 2007; Ala-Poikela et al., 2011).
Another important region of the potyviral protein is VPg, which is translated into the polyprotein, NIa, which is also known as VPg-Pro. During the potyvirus infection, VPg participates in replication and proteolysis and is composed of N-terminal and C-terminal protease domains (Revers et al., 1999). VPg is a multifunctional protein that plays a crucial role in race-specific replication (translation and RNA synthesis), as well as in cell-to-cell and long-distance movement; it also interacts with host proteins as well as various recessive potyvirus resistance genes in different host species (Lellis et al., 2002; Rajamaki and Valkonen, 2002).
Because viruses have relatively small genomes and a limited number of proteins, they rely on the host-cell environment to complete their infection cycle. The characterization of host proteins, membranes, and nucleic acids, using a model host system, functional genomics, and modern molecular biology methods, help in the understanding of plant–virus interactions (Whitham and Wang, 2004). For example, positive-sense ssRNA viruses replicate in association with host endomembranes (Mackenzie, 2005) and different host factors (Ahlquist et al., 2003; Whitham and Wang, 2004). One of the most important genes is the translation initiation factor, eIF4E, which initiates the translation of mRNA and regulates protein synthesis (Sonenberg et al., 1978; Jackson et al., 2010). eIF4E also interacts with the 5′-terminal cap of mRNA and was initially named the ‘cap-binding protein.’ Moreover, eIF4E and its isoform eIF(iso)4E are functionally redundant and one or both of them interact with HCpro and VPg, which are indispensable for viruses to complete their infection cycle; therefore, abolishing this interaction may prevent the viral infection (Lellis et al., 2002; Browning, 2004; Kang et al., 2005; Jin et al., 2007; Charron et al., 2008; Ala-Poikela et al., 2011; Wang and Krishnaswamy, 2012; Sanfacon, 2015). Based on this concept, silencing or incurring mutation in the gene may disrupt infection. pvr2 is a two-nucleotide substitution of the amino acid of pepper eIF4E and is resistant to PVY (Ruffel et al., 2002). A small number of amino acid substitutions in the tomato eIF4E pot-1 confer resistance against PVY and Tobacco etch virus (TEV) in tomato. Barley rym4 and rym5 are also amino acid substitutions that confer eIF4E resistance to Barley yellow mosaic virus (BaYMV) and Barley mild mosaic virus (BaMMV) in Barley (Kanyuka et al., 2005; Stein et al., 2005). Pepper pvr1(2) contains an eIF4E mutation and pvr6 is an eIF(iso)4E mutation; simultaneous mutations in eIF4E and eIF(iso)4E confer resistance to Chilli veinal mottle virus (ChiVMV) in pepper, and silencing eIF4E and eIF(iso)4E reduces the ChiVMV accumulation (Ruffel et al., 2006; Hwang et al., 2009). In plum, the silencing of eIF(iso)4E results in resistance to Plum pox virus (PPV; Wang et al., 2013; Cui and Wang, 2016). Thus, the dependence of potyviruses on eIF4E and/or eIF(iso)4E varies with each virus–host interaction.
To date, no effective way of controlling PStV such as using genetically resistant varieties has been established, mainly because no resistance genes have been identified. We hypothesize that peanut eIF4E/eIF(iso)4E controls the effect of PStV in peanut. To test this hypothesis, we investigated the effects of silencing the translation initiation factor, eIF4E/eIF(iso)4E, to confer PStV resistance in peanut. Moreover, we examined the interaction between HC-Pro and VPg of PStV with eIF4E/eIF(iso)4E using Y2H and BiFC. We also detected the expression of eIF4E/eIF(iso)4E in different peanut tissues.
Materials and Methods
Cloning and Sequencing of PeaeIF4E and PeaeIF(iso)4E Genes
Total RNA was extracted from peanut (Arachis hypogaea) leaves using TRIzol (Invitrogen, Carlsbad, CA, USA), and cDNA was synthesized using an M-MLV RTase cDNA synthesis kit (Takara, Dalian, China), following the manufacturer’s recommendations. To design primers for cloning the eIF4E and eIF(iso)4E genes of peanut, we compared and downloaded the eIF4E sequences of Medicago truncatula (XM_003593785), Pisum sativum (AY423375), Pisum sativum (DQ641471), Phaseolus vulgaris (EF571276), Phaseolus vulgaris (EF571275), M. tornata (HQ735878), and M. truncatula (HQ735877). The conserved sequences were used in designing the primer pairs PeaeIF4E2-R and PeaeIF4E2-F (Supplementary Table S1) to amplify the peanut eIF4E gene. The PCR products showing the expected lengths were sequenced and compared. The product with the correct sequence was then used in designing primers (Supplementary Table S1) for 5′ amplification of cDNA ends (5′-RACE) and 3′-RACE to obtain the full-length cDNA of PeaeIF4E. A 5′-RACE kit (Invitrogen) was used according to the manufacturer’s instructions to obtain the 5′ terminus of the PeaeIF4E gene. The 5′-RACE eIF4E outer and inner primers and 3′-RACE eIF4E outer and inner primer (Supplementary Table S1) were used to obtain the full-length PeaeIF4E cDNA. The PeaeIF(iso)4E gene was amplified using the primers listed in Supplementary Table S1. Phusion high-fidelity DNA polymerase (Takara, Dalian, China) was used to perform all the PCRs. A gel extraction kit (TianGen, Beijing, China) was used to purify the PCR products, which were then cloned into a pMD-18T easy vector (Takara) for sequencing. DNAMAN 6.0 was used for multiple sequence alignment to homologous proteins of different plant species. MEGA5 with the Equal input model was used for phylogenetic analyses by using the neighbor-joining (NJ) method, and confidence was estimated by using 1,000 bootstrap replicates (Tamura et al., 2011).
Cloning and Sequencing of PStV VPg and HC-Pro
Total RNA extraction and cDNA synthesis of the PStV VPg and HC-Pro genes were similar to the method used in cloning PeaeIF4E and PeaeIF(iso)4E. Based on the reported cDNA sequence of PStV (GenBank Accession No.: KF439722, U05771, and U34972), we designed primers for the amplification of segments that corresponded to PStV VPg and PStV HC-Pro (Supplementary Table S1). Phusion high-fidelity DNA polymerase (Takara) was used for all PCRs. A gel extraction kit (TianGen) was used to purify the PCR products. Then the purified PCR products were cloned into a pMD-18T easy vector (Takara) for sequencing.
qRT-PCR Analysis
Total RNA samples were extracted from the roots, stems, leaves, flower buds, leaf buds of “Huayu 20,” and cDNAs were synthesized using the same method employed in cloning the PeaeIF4E and PeaeIF(iso)4E genes. All peanut tissues were sampled from three different peanut plants as biological replicates. For the analysis of gene silencing in peanut, total RNA was extracted from the leaves of eIF4E-silenced, eIF(iso)4E-silenced, or eIF4E-eIF(iso)4E-double silenced peanuts and used in RT-PCR as previously described. Quantitative real-time PCR (qRT-PCR) reactions were conducted by using a SYBR Premix Ex Taq PCR kit (Takara) on an ABI7500 real-time PCR system (ABI, Foster, CA, USA). The primer pairs YG4E-R/YG4E-F and YG4IE-R/YG4IE-F were used to detect the expression of PeaeIF4E and PeaeIF(iso)4E. The primer pair YGPStV-R/YGPStV-F was used to detect the accumulation of PStV after inoculation. The primer pair actin-R/actin-F was used to amplify the actin gene of A. hypogaea, which was used as a reference. The PCR reaction system consisted of a total volume of 20 μL, which included 2 μL of the RT product, 10 μL of Ex Taq, 0.8 μL of the primers (Supplementary Table S1), and 7.2 μL of DEPC-water. All the reactions were performed in a 96-well optical plate. The PCR conditions were as follows: 94°C for 15 s, 94°C for 6 s, and 60°C for 30 s for a total of 40 cycles. Data analysis was performed by using an ABI7500 real-time PCR system, and standard curves were also constructed.
Subcellular Localization of PStV VPg, PStV HC-Pro, and eIF4E/eIF(iso)4E
The ORF of the target gene without its stop codon was amplified using the Phusion high-fidelity DNA polymerase (Takara) using the corresponding primer pairs, ORF4E-R/ORF4E-F, 4E(isoORF)F/4E(isoORF)R, PV-F/PV-R, and PH-R/PH-F, and then cloned into a pMD-18T easy vector (Takara) for sequencing. After confirmation of the correct clone from pMD-18T, these were then introduced into an entry vector pGWCm by TA cloning, and finally, via LR gateway recombination reaction (Invitrogen), was transferred to the plant expression vector, pHZM03. Plasmid DNA with green fluorescent protein (GFP) was transiently introduced into Arabidopsis protoplasts (Meng, 2012). After incubating for 12–16 h in the dark, GFP expression was visualized using a confocal laser microscope (Leica SP5, Mannheim, Germany).
Yeast Two-Hybrid Assay
Yeast two-hybrid screening was conducted using a Matchmaker Gold Yeast two-hybrid system (Clontech, Mountain View, CA, USA). The coding sequences of PStV VPg, PStV HC-Pro, and eIF4E/eIF(iso)4E were PCR amplified by using Phusion high-fidelity DNA polymerase (Takara) with the primer pair listed in Supplementary Table S1. PStV VPg, PStV HC-Pro were cloned into the prey vector, pGADT7, and eIF4E/eIF(iso)4E were cloned into the bait vector, pGBKT7. Confirmed correct clones were transformed into Escherichia coli DH5α cells for subsequent DNA sequencing. Both the confirmed correct prey and bait vectors were then co-transformed into AH109 yeast cells. SD/-Leu-Trp, SD/-Leu-Trp-His, SD/-Leu-Trp-His-Ade, and SD/-Leu-Trp-His-Ade+X-α-gal (Clontech) were used as selective media to detect any interactions. Yeast that contained both empty pGADT7 and pGBKT7 were used as negative controls, and yeast containing both pGBK-p53 and pGAD-RecT were used as positive controls.
Bimolecular Fluorescence Complementation (BiFC)
The Gateway compatible BiFC vectors pEarleyGate202-NYFP and pEarleyGate202-CYFP were used. DNA fragments corresponding to PStV VPg, PStV HC-Pro, and eIF4E/eIF(iso)4E were introduced individually into the entry vector pGWCm as previously described. pGWCm-VPg and pGWCm-HC-pro were transferred to the pEarleyGate202-CYFP vector, whereas pGWCm-eIF4E and pGWCm-eIF(iso)4E were transferred to the pEarleyGate202-NYFP vector via the LR Gateway recombination reaction (Invitrogen). Plasmid DNA with YFP fusion was introduced into Arabidopsis protoplasts for transient expression (Meng, 2012). After incubation for 12–16 h in the dark, confocal laser-scanning microscopy (Leica SP5) was performed to evaluate YFP expression.
Silencing of Peanut Using the Virus-Induced Gene Silencing (VIGS) Vector System
For VIGS assays, PeaeIF4E and PeaeIF(iso)4E were amplified by using the primers listed in Supplementary Table S1. The PCR products were digested with XhoI and BamHI (New England Biolabs) and ligated into vector ALSV-RNA2, which was digested with the corresponding enzymes. The plasmid constructs were then sequenced (Takara) to confirm that we obtained the correct inserts. Apple latent spherical virus (ALSV)-based VIGS method was used. The constructed ALSV-RNA2 and ALSV-RNA1 were transformed into E. coli DH5α cells for sequencing; then the confirmed correct plasmid was cultured in E. coli DH5α cells using TransGEN plasmid Maxi Kit (TransGEN) for purification. Approximately 1 μg/μL of modified ALSV-RNA2 and ALSV-RNA1 were mechanically inoculated into Chenopodium quinoa plants. Two to three weeks later, symptomatic leaves were collected, homogenized in three volumes of extraction buffer (0.1 M Tris-HCl, pH7.8, 0.1 M NaCl, 5 mM MgCl2), and re-inoculated in C. quinoa plants. Then the total RNA of symptomatic leaves were extracted and diluted to about 0.02 μg/μL to be mechanically inoculated into peanuts (2 weeks after sprouting; Igarashi et al., 2009).
Virus Inoculation
In the present study, the PStV isolate was from Laixi, Qingdao city, Shandong province, China. It was cultured in our laboratory via mechanical inoculation for maintenance. PStV-infected plants were maintained at 25°C with 8 h photoperiod. Two weeks after silenced peanut, PStV was mechanically inoculated into peanut. After infection, the presence of PStV was tested by real-time RT-PCR.
Results
Cloning and Sequencing of PeaeIF4E and PeaeIF(iso)4E Genes
The sequence of the peanut eIF4E gene was amplified by using 5′-RACE and 3′-RACE. The full-length cDNA sequence of the eIF4E gene was deposited in GenBank (Accession No. HE985069). The eIF4E gene was 764-bp long, with a 5′ 39-bp untranslated region, a 696-bp ORF, and a 29-bp 3′ untranslated region, and encoded a putative 231-amino acid polypeptide. Moreover, it had 92% nucleotide sequence identity to its homolog in Pisum sativum cultivar (GenBank Accession No. AY423375.2) and 70% nucleotide identity to its homolog in M. truncatula (GenBank Accession No. BT134162.1).
The peanut eIF(iso)4E gene was isolated as described above. The obtained peanut eIF(iso)4E ORF was 612 bp (GenBank Accession No. KF956378) and was predicted to encode a 203-amino acid polypeptide. The cloned peanut eIF4E and eIF(iso)4E genes were designated as peaeIF4E and peaeIF(iso)4E, respectively. The identity of the peaeIF4E and peaeIF(iso)4E genes at the nucleotide sequence level was 53.55%, whereas that at the amino acid sequence level was 42.67%.
The eIF4E and eIF(iso)4E protein sequences of other plant species were aligned for phylogenetic reconstruction. In the phylogenetic tree, eIF4E and eIF(iso)4E formed two distinct branches (Figure 1). Furthermore, peaeIF4E was clustered with the eIF4E subgroup, whereas peaeIF(iso)4E was classified into the eIF(iso)4E subgroup. Analysis using the conserved domain search service of NCBI confirmed that the two proteins contained the eIF4E family conserved sequence.
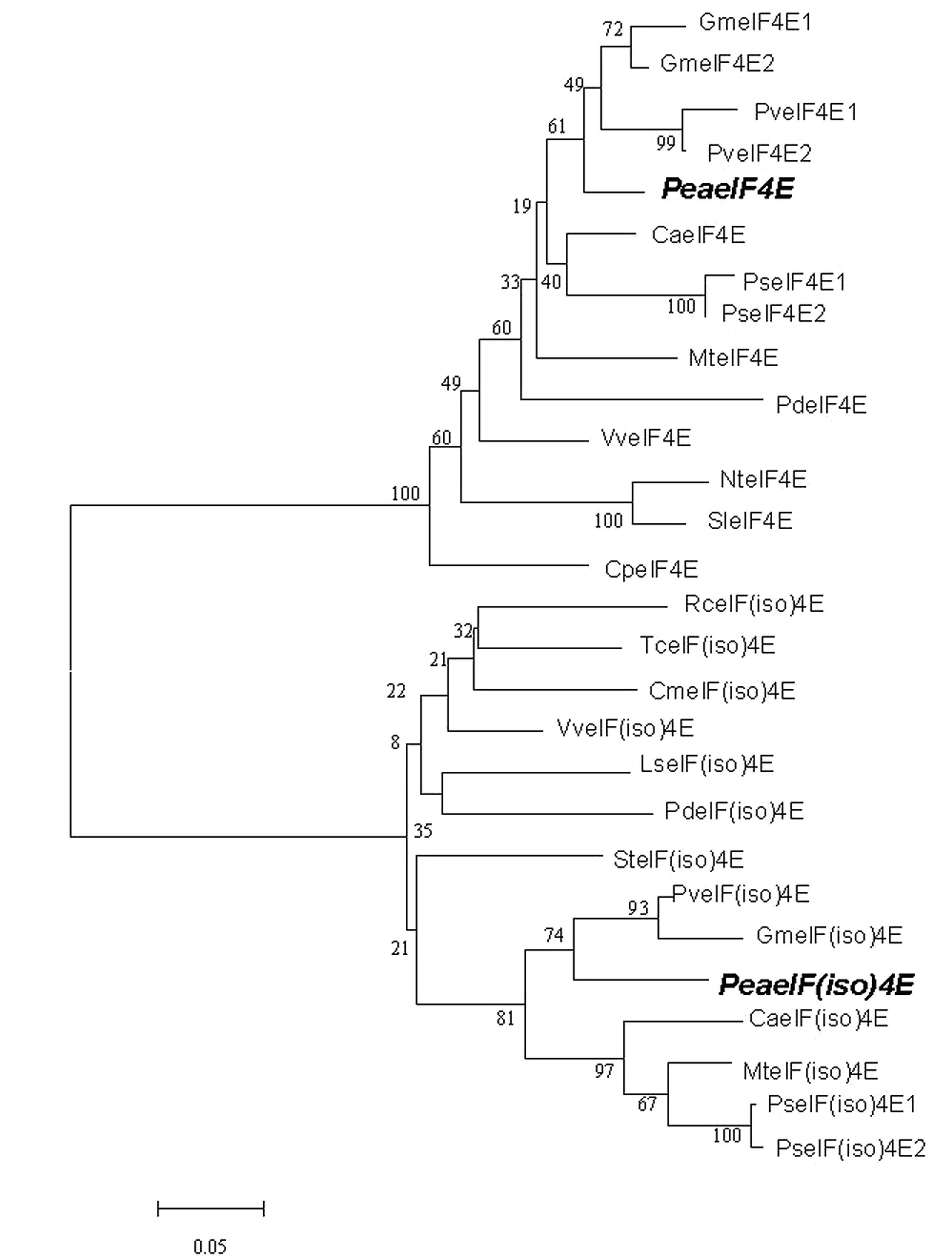
FIGURE 1. Phylogenetic analysis of eIF4E and eIF(iso)4E sequences of different plant species. The phylogenetic tree was constructed using ClustalW (http://www.ebi.ac.uk/clustalw/). The GenBank accession numbers of the amino acid sequences used are listed in Supplementary Table S2. The two peanut sequences are highlighted in bold and italics.
Expression Profiles of PeaeIF4E/PeaeIF(iso)4E in Different Tissues of Peanut
To detect the expression levels of peanut PeaeIF4E and PeaeIF(iso)4E in various tissues, quantitative real-time PCR was performed. RNA was isolated from various tissues such as the roots, stems, leaves, flower buds, and leaf buds of “Huayu 20.” The expression of the peanut actin gene is constant under different conditions and in different tissues (Chi et al., 2012), and was thus selected as a reference. The mRNA transcript levels showed significant differences in different tissues from peanut plants. The expression patterns and the mRNA transcript levels of PeaeIF4E and PeaeIF(iso)4E were similar in all tissues (Figure 2A). The highest transcript levels, for both genes, were observed in the leaf bud and the lowest in flowers (Figure 2A).
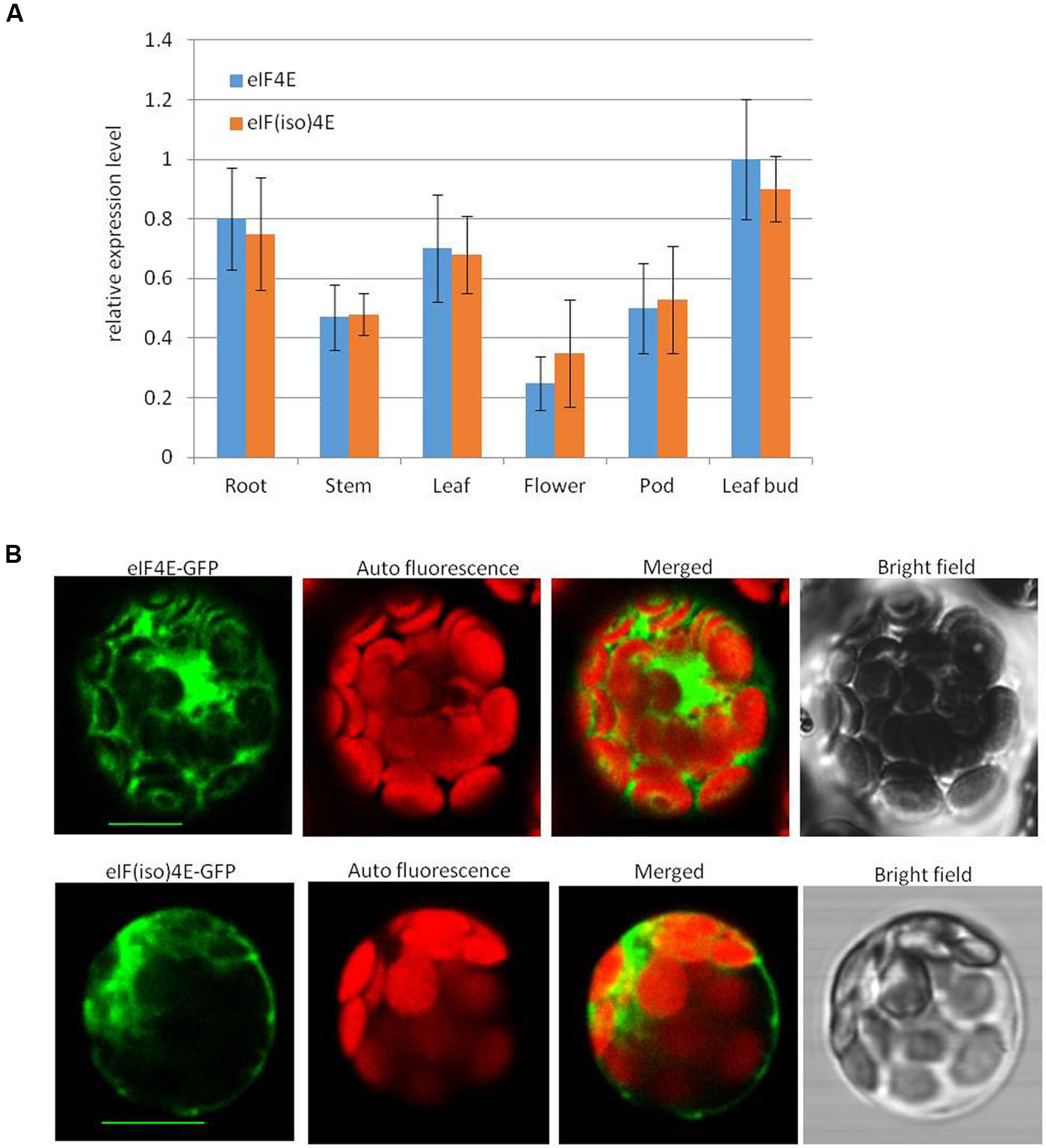
FIGURE 2. mRNA transcript levels (A) of PeaeIF4E and PeaeIF(iso)4E and their subcellular localization (B). Relative mRNA expression levels of peaeIF4E and peaeIF(iso)4E were determined by real-time reverse transcript PCR (RT-PCR). The values represent means of three biological repeats and the value of each biological repeat is the mean of three technical repeats. All values were normalized to the reference gene peanut actin. PeaeIF4E and PeaeIF(iso)4E were fused with green fluorescent protein (GFP) are delivered into protoplasts of Arabidopsis. The GFP fluorescence was observed 12–16 h after transfection. Scale bars = 10 μm.
Subcellular Localization of PeaeIF4E and PeaeIF(iso)4E in Arabidopsis
To test the subcellular localization of PeaeIF4E and PeaeIF(iso)4E, PeaeIF4E and PeaeIF(iso)4E were fused to the GFP by cloning of the ORFs of PeaeIF4E and PeaeIF(iso)4E into the entry vector pGWCm. Recombinant plasmids that expressed the PeaeIF4E-GFP and PeaeIF(iso)4E-GFP fusion proteins were introduced into Arabidopsis protoplasts cell. The Arabidopsis protoplasts cell were cultured in the dark at 23°C for about 12–16 h and observed by the confocal laser scanning microscopy (Leica SP5). The results suggested that the PeaeIF(iso)4E and PeaeIF4E fusion proteins were present in both the nucleus and the cytoplasm (Figure 2B).
Subcellular Localization of VPg and HC-Pro in Arabidopsis
We obtained the VPg and HC-Pro protein cDNAs from PStV by RT-PCR, and their ORFs were fused to GFP as above. The fusion proteins were expressed in Arabidopsis protoplasts as above. The results suggested that the VPg fusion protein was present in the nucleus, and HC-Pro fusion protein was observed in the cytoplasm (Figure 3).
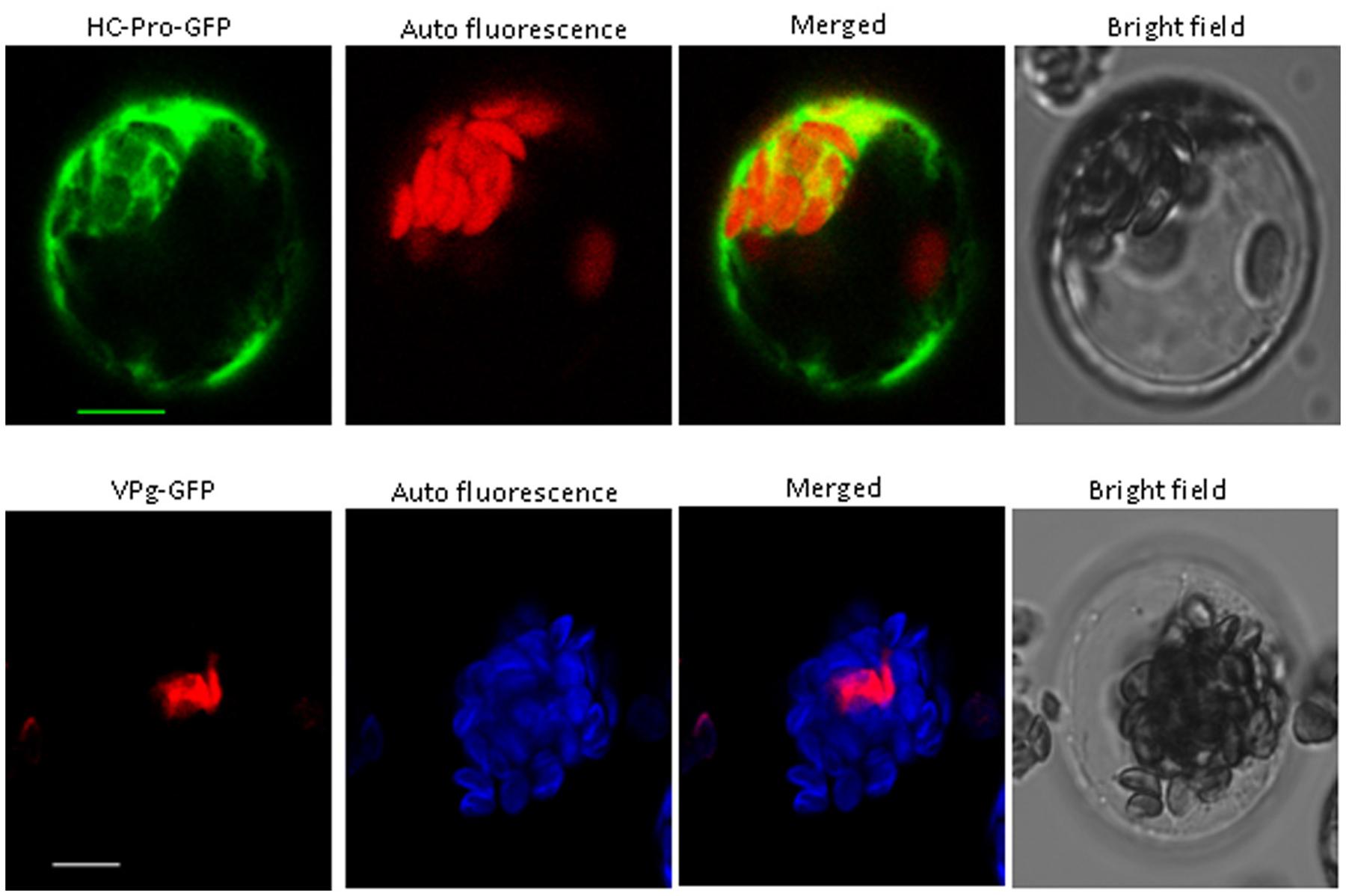
FIGURE 3. Subcellular localization of PStV HC-Pro and PStV VPg. PStV HC-Pro and PStV VPg fused with GFP were transfected into protoplasts of Arabidopsis. The GFP fluorescence was observed 12–16 h after transfection. Scale bars = 10 μm.
Interaction Analysis between eIF4E/eIF(iso)4E and PStV HC-Pro/PStV Vpg
Yeast two-hybrid analysis was used to determine whether there was an interaction between viral proteins and peanut proteins. Yeast two-hybridization showed interactions between VPg and PeaeIF4E/PeaeIF(iso)4E, and between HC-Pro and PeaeIF4E/PeaeIF(iso)4E (Figure 4). The interactions were further confirmed by using BiFC. In this system, the YFP was split into N-terminal and C-terminal fragments, and the PeaeIF4E and PeaeIF(iso)4E were attached to the N-terminal fragment of YFP(eIF4E-NY and eIF(iso)4E-NY). VPg and HC-pro were fused to the C-terminal fragment of YFP(VPg-CY and HC-pro-CY). The eIF(iso)4E-NY+VPg-CY, eIF4E-NY+VPg-CY, eIF4E-NY+HC-pro-CY, and eIF(iso)4E-NY+HC-pro-CY plasmids were then transformed into Arabidopsis protoplasts cells. A nuclear fluorescence signal was, respectively, observed in eIF(iso)4E-NY+VPg-CY and eIF4E-NY+VPg-CY combination and the signal was observed throughout the nucleus(Figure 5C). Cytoplasmic fluorescence signals were observed in the eIF4E-NY+HC-pro-CY and eIF(iso)4E-NY+HC-pro-CY combinations, and the signals were observed throughout the cytoplasm (Figure 5A). As expected, the negative controls, i.e., the combinations of eIF4E-NY+CY and NY+HC-Pro-CY did not emit fluorescence signals (Figure 5B). Taken together, these results show that PeaeIF4E/PeaeIF(iso)4E interacts with VPg in the nucleus, whereas PeaeIF4E/PeaeIF(iso)4E interacts with HC-Pro in the cytoplasm.

FIGURE 4. Yeast two-hybrid assay of protein–protein interaction between the PeaeIF4E/PeaeIF(iso)4E from peanut and PStV-HC-pro/PStV-VPg. Yeast co-transformants were grown on selective medium SD/-Leu-Trp-His-Ade plus X-α-Gal and incubated for 4 days at 30°C (A). Frame (B) corresponds to the clones left (A).
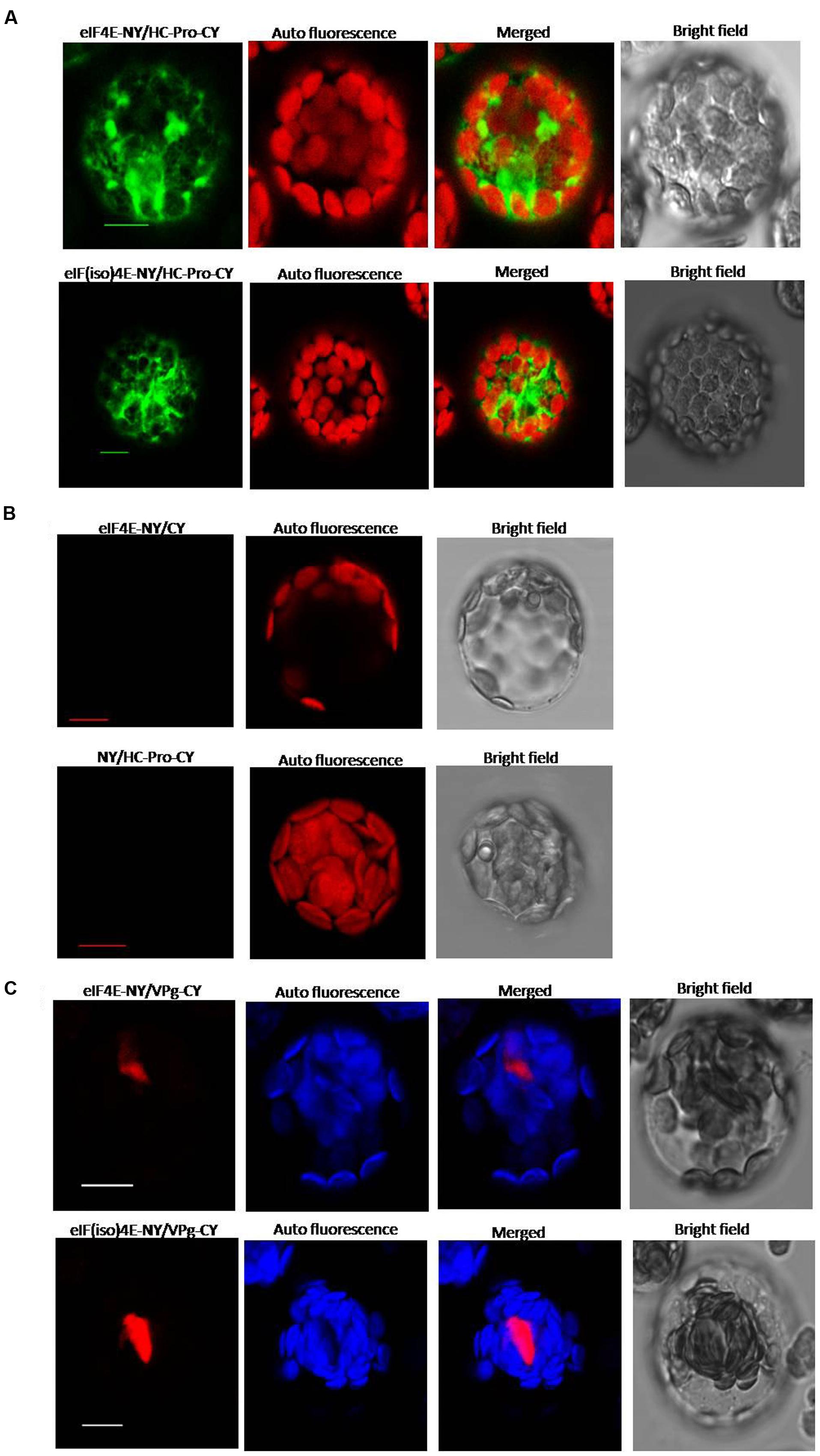
FIGURE 5. Bimolecular fluorescence complementation (BiFC) assay showing interaction between PeaeIF4E/PeaeIF(iso)4E and PStV HC-Pro/PStV VPg. The full-length open reading frame (ORF) of PeaeIF4E/PeaeIF(iso)4E was cloned into the vector pEarleyGate202-NYFP [eIF4E-NY, eIF(iso)4E-NY] and that of PStV HC-Pro/PStV VPg into pEarleyGate202-CYFP (HC-Pro-CY, VPg-CY). The recombinant plasmids were transfected into protoplasts of Arabidopsis. Fluorescence was observed at 14–16 h post-transfection by confocal laser-scanning microscopy. Scale bars = 10 μm. (A) BiFC analysis of PeaeIF4E/PeaeIF(iso)4E and PStV HC-Pro. (B) A range of negative controls. (C) BiFC analysis of PeaeIF4E/PeaeIF(iso)4E and PStV VPg.
Silencing of PeaeIF4E and PeaeIF(iso)4E Genes Confers Resistance against PStV in Peanut
To confirm the role of the PeaeIF4E and PeaeIF(iso)4E genes in PStV infection, gene silencing was performed. The PeaeIF4E (355-nt) and PeaeIF(iso)4E (326-nt) fragments were inserted into the ALSV-RNA2 vector. The recombinant viruses (ALSV-eIF4E and ALSV-eIF(iso)4E) were then inoculated into a peanut. Two weeks after inoculation, real-time RT-PCR analysis was performed, which demonstrated that the expression levels of PeaeIF4E and PeaeIF(iso)4E were significantly lower in the inoculated plants as compared with control (Figure 6A), although no significant phenotypic alterations were observed in the transgenic plants (Figure 6C). The expression level of PeaeIF4E decreased by 60% (upon inoculation with ALSV-eIF4E) while that of PeaeIF(iso)4E decreased by 65% (ALSV-eIF(iso)4E inoculation) as compared with control. When inoculated with ALSV-eIF4E+ALSV-eIF(iso)4E, the expression levels of PeaeIF4E and PeaeIF(iso)4E decreased by 53 and 57%, respectively, as compared with control (Figure 6A). Peanut plants where either the PeaeIF4E or PeaeIF(iso)4E were silenced, showed mosaic symptoms of infection at about 10–14 days after inoculation with PStV. On the other hand, silencing of both PeaeIF4E or PeaeIF(iso)4E caused the symptoms to appear later at about 18–20 days after inoculation with PStV, and the symptoms were milder as compared with plants with only one gene silenced and the control (Figure 6C). Real-time RT-PCR analysis indicated that silencing both PeaeIF4E or PeaeIF(iso)4E reduced PStV accumulation by 70% compared to control plants. No significant differences were observed between plants in which either PeaeIF4E or PeaeIF(iso)4E was silenced as compared with controls (Figure 6B) suggesting that the two isoforms play overlapping or redundant roles in the virus multiplication cycle.
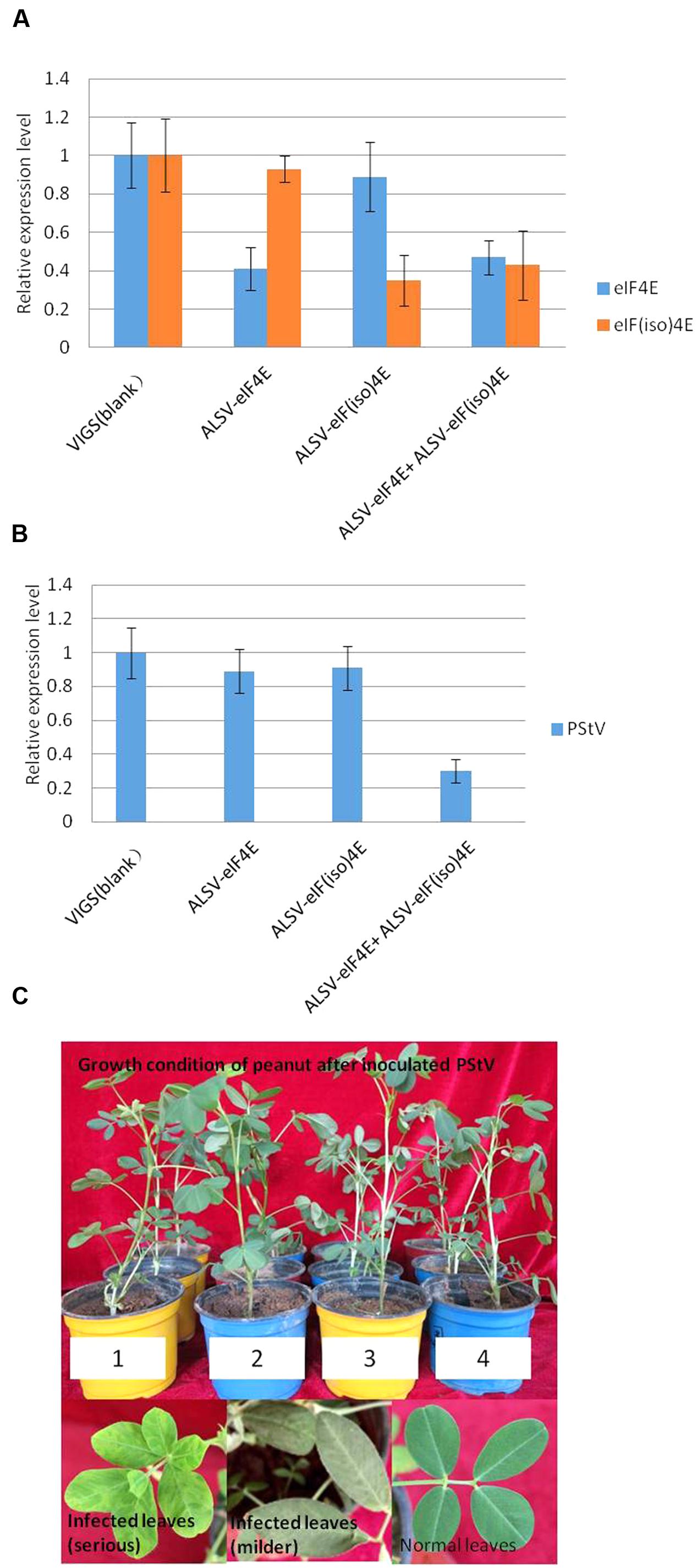
FIGURE 6. Real-time PCR analysis for target gene expression in peanut (A), accumulation of PStV RNA (B) and the growth condition and symptoms of peanut after inoculated PStV (C). Virus-induced gene silencing of PeaeIF4E/PeaeIF(iso)4E in representative plants belonging to each of the four different treatments. For qRT-PCR detection of the expression of PeaeIF4E/PeaeIF(iso)4E, three plants from each group were pooled as one sample and the experiments were performed in triplicate (A). Effects of silencing of PeaeIF4E/PeaeIF(iso)4E on PStV infection. The accumulation of PStV RNA in inoculated peanut plants was detected by RT-PCR 15 days after PStV inoculation (B). The growth condition and symptoms of peanut after inoculated PStV (C). Four different treatments peanuts were mechanically infected by PStV. Three plants from each group were pooled as one sample and the experiments were performed in triplicate, 10–14 days later, PStV disease symptom began to appeared. The peanut growth condition after inoculated with PStV, except the different symptoms on peanut leaves, different treatments peanuts growth condition showed no significant differences compared with control. 1, control group; 2, silencing PeaeIF4E; 3, silencing PeaeIF(iso)4E; 4, silencing both PeaeIF4E and PeaeIF(iso)4E.
Discussion
The cap-binding protein eIF4E/eIF(iso)4E confers resistance to some RNA viruses in specific plant species (Nicaise et al., 2003; Nieto et al., 2006; Ruffel et al., 2006; Hwang et al., 2009). In the present study, we cloned the peanut eIF4E/eIF(iso)4E genes and analyzed their protein sequences. Phylogenetic analyses of these sequences demonstrated that PeaeIF4E and PeaeIF(iso)4E showed high homologies with orthologs from related plant species. PeaeIF4E and PeaeIF(iso)4E were closely related to their homologs from soybean (G. max) and kidney bean (Phaseolus vulgaris). The expression levels of PeaeIF4E and PeaeIF(iso)4E were similar in different peanut tissues with both genes being upregulated in leaf buds and roots and downregulated in flowers (Figure 2). Previous studies have also shown that these two genes are upregulated in young tissues and downregulated in mature tissues of Arabidopsis and plum, which corroborated our results (Rodriguez et al., 1998; Wang et al., 2013).
Confocal microscopy showed that PeaeIF4E and PeaeIF(iso)4E were both localized in the nucleus and the cytoplasm of Arabidopsis cells (Figure 2B). In Chrysanthemum morifolium, eIF(iso)4E was also localized in the nucleus, cytoplasm, and cytomembrane (Song et al., 2013). In Arabidopsis, in quiescent cells, eIF4E was localized in the nucleus, whereas in proliferating cells, this was detected in the cytoplasm. Both in quiescent and proliferating cells, eIF(iso)4E has been observed in the cytoplasm and nucleus (Bush et al., 2009). In mature Arabidopsis cells, PeaeIF4E and PeaeIF(iso)4E were localized to both the nucleus and the cytoplasm, but whether the two proteins have different cellular locations during different growth stages needs further investigation. In animals, eIF4E has been detected in both cytoplasm and nucleus, and about 68% of the eIF4E was detected in mammalian nuclei. eIF4E plays different roles depending on its subcellular location; when it is localized in cytoplasm, it functions in translation initiation. When it is localized in the nucleus, it participates in the export of mRNAs that contain 4E-sensitive elements (SE; Iborra et al., 2001; Culjkovic et al., 2007, 2008). The two peanut proteins that localized in different places may play different roles that require further investigation.
Confocal microscopy showed that the VPg fusion protein was localized to the nucleus, whereas the HC-Pro fusion protein was observed in the cytoplasm. In the case of MDMV (Maize dwarf mosaic virus) and TuMV (Turnip mosaic virus), HC-Pro was detected in the cytoplasm (Li et al., 2001; Zheng, 2011). In the case of Potato virus Y, HC-Pro was localized throughout the cytoplasm, whereas it displays different subcellular localization patterns depending on the cellular environment (del Toro et al., 2014). HC-Pro was also distributed throughout the cytoplasm in CABMV (Cowpea aphid-borne mosaic virus) infected plants (Mlotshwa et al., 2002). In WYMV (Wheat mosaic virus) infected plants, VPg occurred in two forms in the nucleus; one gathered into one or several irregular shape inclusions, whereas the other was evenly distributed across the entire nucleus (Bian, 2013). These results were consistent with the findings of our study as well as with the results of BiFC analysis of HC-Pro and PeaeIF4E/PeaeIF(iso)4E, and that of VPg and PeaeIF4E/PeaeIF(iso)4E. We also observed interactions between VPg and PeaeIF4E/PeaeIF(iso)4E and between HC-Pro and PeaeIF4E/PeaeIF(iso)4E. The interaction between VPg and PeaeIF4E/PeaeIF(iso)4E interactions was observed in the nucleus. These results coincided with the findings on viral protein location in our study. The interaction between VPg and PeaeIF4E/PeaeIF(iso)4E in the nucleus provides additional evidence that both proteins are localized in the nucleus. In potyvirus, the interaction between VPg and eIF4E/eIF(iso)4E plays a major role in cellular transport and localization of RNA (Lellis et al., 2002). HC-Pro and PeaeIF4E/PeaeIF(iso)4E interactions were detected in the cytoplasm but not in the nucleus, which supported the absence of HC-Pro in the nuclei of infected plant cells (Rajamaki and Valkonen, 2003).
The interaction between translation initiation factors and viral proteins is essential for viral replication and infection (Dreher and Miller, 2006; Robaglia and Caranta, 2006). The interaction between eIF4E/eIF(iso)4E and HC-Pro/VPg may be necessary for potyvirus infection and amplification. The silencing of both PeaeIF4E and PeaeIF(iso)4E conferred moderate resistance against PStV in peanut, as evidenced by symptom delay and reduced virus accumulation. This findings strongly suggest that PeaeIF4E and PeaeIF(iso)4E play important roles to facilitate virus infection and that they are functionally interchangeable. The silencing of PeaeIF4E and PeaeIF(iso)4E hindered the interaction between the host and the virus, which in turn prevented infection and viral replication in the host. Furthermore, viral accumulation was lower in gene-silenced peanut plants. Silenced plants with decreased expression of PeaeIF4E and/or PeaeIF(iso)4E did not phenotypically differ from control plants. In tobacco, antisense depletion of either eIF4E and two eIF(iso)4E isoforms displayed normal development, but antisense depletion of both eIF4E and eIF(iso)4E resulted in semi-dwarf phenotype (Combe et al., 2005). It is possible that the remaining low levels of expression of PeaeIF4E and PeaeIF(iso)4E in the silenced plants were sufficient to sustain peanut growth. Alternatively, it is possible that the two genes are dispensable for peanut growth and that some other genes possess complementary functions.
In other virus-host compositions, simultaneous mutations in the eIF4E and eIF(iso)4E genes result in a decrease in viral resistance, such as resistance to Pepper veinal mottle virus (PVMV) and ChiVMV in pepper (Ruffel et al., 2006; Hwang et al., 2009). Furthermore, knocking down the eIF(iso)4E in peach plants results in peach resistance to PPV (Cui and Wang, 2016). Silencing of the eIF(iso)4E gene in plum confers resistance to PPV (Wang et al., 2013) and silencing the eIF4E gene in melon plants confers upon it broad-spectrum viral resistance (Rodriguez-Hernandez et al., 2012). These reports suggest that these viruses probably utilize one or two translation initiation factors during infection. PStV may use either translation initiation factors of PeaeIF4E and PeaeIF(iso)4E during its infection because silencing only one gene does not confer resistance to PStV in peanut. Our study suggests that the eIF4E/eIF(iso)4E gene may be utilized in increasing PStV resistance in peanut by gene silencing, gene mutation, or the TILLING strategy (Ruffel et al., 2002; Kanyuka et al., 2005; Stein et al., 2005; Piron et al., 2010). We have a variety of peanut cultivars that could be employed in TILLING to detect allelic variants of a target gene.
Therefore, the findings of the present study suggest that eIF4E/eIF(iso)4E plays important roles in the PStV infection cycle and may serve as a novel method for increasing the PStV resistance in economically important peanut cultivars. The two genes may also be used as genetic resources for improving PStV resistance in peanut breeding programs.
Author Contributions
MX, HX, JW, and JY: Design of the work, analysis, and interpretation of data. LX and YC: Final approval of the version to be published and agreement to be accountable for all aspects of the work.
Funding
This research was funded by The Youth Scientific Research Foundation of Shandong Academy of Agricultural Sciences (2014QNM01); Natural Science Foundation of Shandong Province (ZR2015YL065, ZR2014CQ025, 2015GNC111029).
Conflict of Interest Statement
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Acknowledgments
We would like to thank Dr. Chengming Fan (The Chinese Academy of Sciences, Beijing, China) for providing Gateway vectors for our localization and BiFC studies, Dr. Qijun Chen (China Agricultural University, Beijing, China) for providing pGWCm, and Dr. Na Chen (Shandong Peanut Research Institute, Qingdao, China) for providing vectors for the Y2H assays.
Supplementary Material
The Supplementary Material for this article can be found online at: http://journal.frontiersin.org/article/10.3389/fmicb.2017.00338/full#supplementary-material
References
Ahlquist, P., Noueiry, A. O., Lee, W. M., Kushner, D. B., and Dye, B. T. (2003). Host factors in positive-strand RNA virus genome replication. J. Virol. 77, 8181–8186. doi: 10.1128/JVI.77.15.8181-8186.2003
Ala-Poikela, M., Goytia, E., Haikonen, T., Rajamaki, M. L., and Valkonen, J. P. (2011). Helper component proteinase of the genus Potyvirus is an interaction partner of translation initiation factors eIF(iso)4E and eIF4E and contains a 4E binding motif. J. Virol. 85, 6784–6794. doi: 10.1128/JVI.00485-11
Atreya, C. D., Atreya, P. L., Thornbury, D. W., and Pirone, T. P. (1992). Site-directed mutations in the potyvirus HC-Pro gene affect helper component activity, virus accumulation, and symptom expression in infected tobacco plants. Virology 191, 106–111. doi: 10.1016/0042-6822(92)90171-K
Bian, L. (2013). Structural and Subcellular Distribution of VPg Protein Encode by Wheat Yellow Masaic Virus. Master thesis, Zhejiang Normal University, Jinhua.
Browning, K. S. (2004). Plant translation initiation factors: it is not easy to be green. Biochem. Soc. Trans. 32(Pt 4), 589–591. doi: 10.1042/BST0320589
Bush, M. S., Hutchins, A. P., Jones, A. M., Naldrett, M. J., Jarmolowski, A., Lloyd, C. W., et al. (2009). Selective recruitment of proteins to 5’ cap complexes during the growth cycle in Arabidopsis. Plant J. 59, 400–412. doi: 10.1111/j.1365-313X.2009.03882.x
Carrington, J. C., and Herndon, K. L. (1992). Characterization of the potyviral HC-pro autoproteolytic cleavage site. Virology 187, 308–315. doi: 10.1016/0042-6822(92)90319-K
Charron, C., Nicolai, M., Gallois, J. L., Robaglia, C., Moury, B., Palloix, A., et al. (2008). Natural variation and functional analyses provide evidence for co-evolution between plant eIF4E and potyviral VPg. Plant J. 54, 56–68. doi: 10.1111/j.1365-313X.2008.03407.x
Chi, X., Hu, R., Yang, Q., Zhang, X., Pan, L., Chen, N., et al. (2012). Validation of reference genes for gene expression studies in peanut by quantitative real-time RT-PCR. Mol. Genet. Genomics 287, 167–176. doi: 10.1007/s00438-011-0665-5
Choi, H. S., Kim, J. S., Cheon, J. U., Choi, J. K., Pappu, S. S., and Pappu, H. R. (2001). First report of Peanut stripe virus (Family Potyviridae) in South Korea. Plant Dis. 85:679. doi: 10.1094/PDIS.2001.85.6.679B
Choi, H. S., Kim, M., Park, J. W., Cheon, J. U., Kim, K. H., Kim, J. S., et al. (2006). Occurrence of Bean common mosaic virus (BCMV) infecting Peanut in Korea. Plant Pathol. J. 22, 97–102. doi: 10.5423/PPJ.2006.22.1.097
Combe, J. P., Petracek, M. E., van Eldik, G., Meulewaeter, F., and Twell, D. (2005). Translation initiation factors eIF4E and eIFiso4E are required for polysome formation and regulate plant growth in tobacco. Plant Mol. Biol. 57, 749–760. doi: 10.1007/s11103-005-3098-x
Cui, H., and Wang, A. (2016). An efficient viral vector for functional genomic studies of Prunus fruit trees and its induced resistance to Plum pox virus via silencing of a host factor gene. Plant Biotechnol. J. 15, 344–356. doi: 10.1111/pbi.12629
Culjkovic, B., Tan, K., Orolicki, S., Amri, A., Meloche, S., and Borden, K. L. (2008). The eIF4E RNA regulon promotes the Akt signaling pathway. J. Cell Biol. 181, 51–63. doi: 10.1083/jcb.200707018
Culjkovic, B., Topisirovic, I., and Borden, K. L. (2007). Controlling gene expression through RNA regulons: the role of the eukaryotic translation initiation factor eIF4E. Cell Cycle 6, 65–69. doi: 10.4161/cc.6.1.3688
del Toro, F., Fernandez, F. T., Tilsner, J., Wright, K. M., Tenllado, F., Chung, B. N., et al. (2014). Potato virus Y HCPro localization at distinct, dynamically related and environment-influenced structures in the cell cytoplasm. Mol. Plant Microbe Interact. 27, 1331–1343. doi: 10.1094/MPMI-05-14-0155-R.F
Demski, J. W., and Lovell, G. R. (1985). Peanut stripe virus and the distribution of peanut seed. Plant Dis. 69, 734–738.
Dreher, T. W., and Miller, W. A. (2006). Translational control in positive strand RNA plant viruses. Virology 344, 185–197. doi: 10.1016/j.virol.2005.09.031
Govier, D. A., Kassanis, B., and Pirone, T. P. (1977). Partial purification and characterization of the potato virus Y helper component. Virology 78, 306–314. doi: 10.1016/0042-6822(77)90101-5
Hwang, J., Li, J., Liu, W. Y., An, S. J., Cho, H., Her, N. H., et al. (2009). Double mutations in eIF4E and eIFiso4E confer recessive resistance to Chilli veinal mottle virus in pepper. Mol. Cells 27, 329–336. doi: 10.1007/s10059-009-0042-y
Iborra, F. J., Jackson, D. A., and Cook, P. R. (2001). Coupled transcription and translation within nuclei of mammalian cells. Science 293, 1139–1142. doi: 10.1126/science.1061216
Igarashi, A., Yamagata, K., Sugai, T., Takahashi, Y., Sugawara, E., Tamura, A., et al. (2009). Apple latent spherical virus vectors for reliable and effective virus-induced gene silencing among a broad range of plants including tobacco, tomato, Arabidopsis thaliana, cucurbits, and legumes. Virology 386, 407–416. doi: 10.1016/j.virol.2009.01.039
Jackson, R. J., Hellen, C. U., and Pestova, T. V. (2010). The mechanism of eukaryotic translation initiation and principles of its regulation. Nat. Rev. Mol. Cell Biol. 11, 113–127. doi: 10.1038/nrm2838
Jin, Y., Ma, D., Dong, J., Li, D., Deng, C., Jin, J., et al. (2007). The HC-pro protein of potato virus Y interacts with NtMinD of tobacco. Mol. Plant Microbe Interact. 20, 1505–1511. doi: 10.1094/MPMI-20-12-1505
Kang, B. C., Yeam, I., Frantz, J. D., Murphy, J. F., and Jahn, M. M. (2005). The pvr1 locus in Capsicum encodes a translation initiation factor eIF4E that interacts with Tobacco etch virus VPg. Plant J. 42, 392–405. doi: 10.1111/j.1365-313X.2005.02381.x
Kanyuka, K., Druka, A., Caldwell, D. G., Tymon, A., McCallum, N., Waugh, R., et al. (2005). Evidence that the recessive bymovirus resistance locus rym4 in barley corresponds to the eukaryotic translation initiation factor 4E gene. Mol. Plant Pathol. 6, 449–458. doi: 10.1111/j.1364-3703.2005.00294.x
Lellis, A. D., Kasschau, K. D., Whitham, S. A., and Carrington, J. C. (2002). Loss-of-susceptibility mutants of Arabidopsis thaliana reveal an essential role for eIF(iso)4E during potyvirus infection. Curr. Biol. 12, 1046–1051. doi: 10.1016/S0960-9822(02)00898-9
Li, X. D., Fan, Z. F., Li, H. F., and Qiu, W. F. (2001). Accumulation and immunolocalization of Maize dwarf mosaic virus HC-Pro in infected maize leaves. Acta Phytopathol. Sin. 31, 310–314.
Llave, C., Kasschau, K. D., and Carrington, J. C. (2000). Virus-encoded suppressor of posttranscriptional gene silencing targets a maintenance step in the silencing pathway. Proc. Natl. Acad. Sci. U.S.A. 97, 13401–13406. doi: 10.1073/pnas.230334397
Mackenzie, J. (2005). Wrapping things up about virus RNA replication. Traffic 6, 967–977. doi: 10.1111/j.1600-0854.2005.00339.x
Meng, Y. Y. (2012). Functional Analysis of CRYs and CIB3 in Flowering and Senescence Regulation in Soybean (Glycine max). Ph. D. thesis, Graduate School of Chinese Academy of Agricultural Sciences, Beijing.
Mlotshwa, S., Verver, J., Sithole-Niang, I., Gopinath, K., Carette, J., van Kammen, A., et al. (2002). Subcellular location of the helper component-proteinase of Cowpea aphid-borne mosaic virus. Virus Genes 25, 207–216. doi: 10.1023/A:1020122104651
Nicaise, V., German-Retana, S., Sanjuan, R., Dubrana, M. P., Mazier, M., Maisonneuve, B., et al. (2003). The eukaryotic translation initiation factor 4E controls lettuce susceptibility to the Potyvirus Lettuce mosaic virus. Plant Physiol. 132, 1272–1282. doi: 10.1104/pp.102.017855
Nieto, C., Morales, M., Orjeda, G., Clepet, C., Monfort, A., Sturbois, B., et al. (2006). An eIF4E allele confers resistance to an uncapped and non-polyadenylated RNA virus in melon. Plant J. 48, 452–462. doi: 10.1111/j.1365-313X.2006.02885.x
Piron, F., Nicolai, M., Minoia, S., Piednoir, E., Moretti, A., Salgues, A., et al. (2010). An induced mutation in tomato eIF4E leads to immunity to two potyviruses. PLoS ONE 5:e11313. doi: 10.1371/journal.pone.0011313
Rajamaki, M. L., and Valkonen, J. P. (2002). Viral genome-linked protein (VPg) controls accumulation and phloem-loading of a potyvirus in inoculated potato leaves. Mol. Plant Microbe Interact. 15, 138–149. doi: 10.1094/MPMI.2002.15.2.138
Rajamaki, M. L., and Valkonen, J. P. (2003). Localization of a potyvirus and the viral genome-linked protein in wild potato leaves at an early stage of systemic infection. Mol. Plant Microbe Interact. 16, 25–34. doi: 10.1094/MPMI.2003.16.1.25
Redondo, E., Krause-Sakate, R., Yang, S. J., Lot, H., Le Gall, O., and Candresse, T. (2001). Lettuce mosaic virus pathogenicity determinants in susceptible and tolerant lettuce cultivars map to different regions of the viral genome. Mol. Plant Microbe Interact. 14, 804–810. doi: 10.1094/MPMI.2001.14.6.804
Revers, F., van der Vlugt, R. A., Souche, S., Lanneau, M., Lot, H., Candresse, T., et al. (1999). Nucleotide sequence of the 3’ terminal region of the genome of four lettuce mosaic virus isolates from Greece and Yemen. Arch. Virol. 144, 1619–1626. doi: 10.1007/s007050050615
Robaglia, C., and Caranta, C. (2006). Translation initiation factors: a weak link in plant RNA virus infection. Trends Plant Sci. 11, 40–45. doi: 10.1016/j.tplants.2005.11.004
Rodriguez, C. M., Freire, M. A., Camilleri, C., and Robaglia, C. (1998). The Arabidopsis thaliana cDNAs coding for eIF4E and eIF(iso)4E are not functionally equivalent for yeast complementation and are differentially expressed during plant development. Plant J. 13, 465–473. doi: 10.1046/j.1365-313X.1998.00047.x
Rodriguez-Hernandez, A. M., Gosalvez, B., Sempere, R. N., Burgos, L., Aranda, M. A., and Truniger, V. (2012). Melon RNA interference (RNAi) lines silenced for Cm-eIF4E show broad virus resistance. Mol. Plant Pathol. 13, 755–763. doi: 10.1111/j.1364-3703.2012.00785.x
Rojas, M. R., Zerbini, F. M., Allison, R. F., Gilbertson, R. L., and Lucas, W. J. (1997). Capsid protein and helper component-proteinase function as potyvirus cell-to-cell movement proteins. Virology 237, 283–295. doi: 10.1006/viro.1997.8777
Ruffel, S., Dussault, M. H., Palloix, A., Moury, B., Bendahmane, A., Robaglia, C., et al. (2002). A natural recessive resistance gene against potato virus Y in pepper corresponds to the eukaryotic initiation factor 4E (eIF4E). Plant J. 32, 1067–1075. doi: 10.1046/j.1365-313X.2002.01499.x
Ruffel, S., Gallois, J. L., Moury, B., Robaglia, C., Palloix, A., and Caranta, C. (2006). Simultaneous mutations in translation initiation factors eIF4E and eIF(iso)4E are required to prevent pepper veinal mottle virus infection of pepper. J. Gen. Virol. 87(Pt 7), 2089–2098. doi: 10.1099/vir.0.81817-0
Saenz, P., Salvador, B., Simon-Mateo, C., Kasschau, K. D., Carrington, J. C., and Garcia, J. A. (2002). Host-specific involvement of the HC protein in the long-distance movement of potyviruses. J. Virol. 76, 1922–1931. doi: 10.1128/JVI.76.4.1922-1931.2002
Saleh, N., Horn, N. M., Reddy, D. V. R., and Middleton, K. J. (1989). Peanut stripe virus in Indonesia. Eur. J. Plant Pathol. 99, 123–127. doi: 10.1007/bf01997480
Sanfacon, H. (2015). Plant translation factors and virus resistance. Viruses 7, 3392–3419. doi: 10.3390/v7072778
Singh, M. K., Chandel, V., Hallan, V., Ram, R., and Zaidi, A. A. (2009). Occurrence of Peanut stripe virus on patchouli and raising of virus-free patchouli plants by meristem tip culture. J. Plant Dis. Prot. 116, 2–6. doi: 10.1007/BF03356278
Sonenberg, N., Morgan, M. A., Merrick, W. C., and Shatkin, A. J. (1978). A polypeptide in eukaryotic initiation factors that crosslinks specifically to the 5’-terminal cap in mRNA. Proc. Natl. Acad. Sci. U.S.A. 75, 4843–4847. doi: 10.1073/pnas.75.10.4843
Song, A., Lou, W., Jiang, J., Chen, S., Sun, Z., Guan, Z., et al. (2013). An isoform of eukaryotic initiation factor 4E from Chrysanthemum morifolium interacts with Chrysanthemum virus B coat protein. PLoS ONE 8:e57229. doi: 10.1371/journal.pone.0057229
Stein, N., Perovic, D., Kumlehn, J., Pellio, B., Stracke, S., Streng, S., et al. (2005). The eukaryotic translation initiation factor 4E confers multiallelic recessive Bymovirus resistance in Hordeum vulgare (L.). Plant J. 42, 912–922. doi: 10.1111/j.1365-313X.2005.02424.x
Tamura, K., Peterson, D., Peterson, N., Stecher, G., Nei, M., and Kumar, S. (2011). MEGA5: molecular evolutionary genetics analysis using maximum likelihood, evolutionary distance, and maximum parsimony methods. Mol. Biol. Evol. 28, 2731–2739. doi: 10.1093/molbev/msr121
Urcuqui-Inchima, S., Haenni, A. L., and Bernardi, F. (2001). Potyvirus proteins: a wealth of functions. Virus Res. 74, 157–175. doi: 10.1016/S0168-1702(01)00220-9
Wang, A., and Krishnaswamy, S. (2012). Eukaryotic translation initiation factor 4E-mediated recessive resistance to plant viruses and its utility in crop improvement. Mol. Plant Pathol. 13, 795–803. doi: 10.1111/j.1364-3703.2012.00791.x
Wang, X., Kohalmi, S. E., Svircev, A., Wang, A., Sanfacon, H., and Tian, L. (2013). Silencing of the host factor eIF(iso)4E gene confers plum pox virus resistance in plum. PLoS ONE 8:e50627. doi: 10.1371/journal.pone.0050627
Wei, T., Zhang, C., Hong, J., Xiong, R., Kasschau, K. D., Zhou, X., et al. (2010). Formation of complexes at plasmodesmata for potyvirus intercellular movement is mediated by the viral protein P3N-PIPO. PLoS Pathog. 6:e1000962. doi: 10.1371/journal.ppat.1000962
Whitham, S. A., and Wang, Y. (2004). Roles for host factors in plant viral pathogenicity. Curr. Opin. Plant Biol. 7, 365–371. doi: 10.1016/j.pbi.2004.04.006
Keywords: peanut, Peanut stripe virus, translation initiation factor 4E, protein–protein interaction, gene silencing
Citation: Xu M, Xie H, Wu J, Xie L, Yang J and Chi Y (2017) Translation Initiation Factor eIF4E and eIFiso4E Are Both Required for Peanut stripe virus Infection in Peanut (Arachis hypogaea L.). Front. Microbiol. 8:338. doi: 10.3389/fmicb.2017.00338
Received: 08 December 2016; Accepted: 17 February 2017;
Published: 10 March 2017.
Edited by:
Brigitte Mauch-Mani, University of Neuchâtel, SwitzerlandReviewed by:
Helene Sanfacon, Agriculture and Agri-Food Canada, CanadaXuefeng Yuan, Shandong Agricultural University, China
Copyright © 2017 Xu, Xie, Wu, Xie, Yang and Chi. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) or licensor are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Jinguang Yang, eWFuZ2ppbmd1YW5nQGNhYXMuY24= Yucheng Chi, ODc2MjY2ODFAMTYzLmNvbQ==