 Lourdes Migura-Garcia1,2*
Lourdes Migura-Garcia1,2* Juan J. González-López3
Juan J. González-López3 Jaime Martinez-Urtaza4
Jaime Martinez-Urtaza4 J. R. Aguirre Sánchez5,6
J. R. Aguirre Sánchez5,6 A. Moreno-Mingorance3A. Perez de Rozas1
A. Moreno-Mingorance3A. Perez de Rozas1 Ursula Höfle6Y. Ramiro6
Ursula Höfle6Y. Ramiro6 Narjol Gonzalez-Escalona7
Narjol Gonzalez-Escalona7- 1Centre de Recerca en Sanitat Animal, Institut de Recerca i Tecnologia Agroalimentàries, Barcelona, Spain
- 2Research and Control of Emerging and Re-emerging Swine Diseases in Europe, OIE Collaborating Centre, CReSA, IRTA, Barcelona, Spain
- 3Servei de Microbiologia, Hospital Universitari Vall d’Hebron, Universitat Autònoma de Barcelona, Barcelona, Spain
- 4Centre for Environment, Fisheries and Aquaculture Science (CEFAS), Weymouth, United Kingdom
- 5Centro de Investigación en Alimentación y Desarrollo, Culiacán, Mexico
- 6Health and Biotechnology (SaBio) Group, Instituto de Investigación en Recursos Cinégéticos IREC (CSIC-UCLM-JCCM), Ciudad Real, Spain
- 7Food and Drug Administration, Silver Spring, MD, United States
Colistin has become the last-line antimicrobial for the treatment of multidrug resistant (MDR) Enterobacterales in human medicine. To date, several colistin resistance genes have been described. Of them mcr-1 is disseminated worldwide in Escherichia coli of human and animal origin. The aim of this study was to characterize mcr-mediated resistance plasmids from E. coli of animal origin in Spain. From our strain collection, 70 E. coli of pig origin collected between 2005 and 2014 (10 per year, except for years 2009–2010–2013) were randomly selected and screened for the presence of mcr-genes. Additionally, 20 E. coli isolated in 2011 from white storks (Ciconia ciconia) from the same urban household waste landfill associated colony were also included. Whole genome sequencing of mcr-positive isolates was carried out on a MiSeq (Illumina). Hybrid whole genome sequencing strategy combining nanopore and Illumina technologies were performed in a selection of isolates to close the genomes and plasmids and identify the presence of antimicrobial resistance genes. Minimum inhibitory concentration (MIC) was used to assess the susceptibility to colistin. Mating experiments were carried out to evaluate transferability of the mcr-genes. A total of 19 mcr-1 and one mcr-4 positive isolates were detected, 15 from pigs distributed during the study period, and five from storks collected in 2011. No other mcr-variants were found. The MICs for colistin ranged between 4 and >4 mg/L. High diversity of STs were detected among the mcr-1 positive E. coli isolates, with only ST-10 shared between pigs and white storks. Except for one isolate, all were genotypic and phenotypically MDR, and five of them also harbored cephalosporin resistance genes (blaCTX–M–14, blaSHV–12, and three blaCMY–2). mcr-1 genes were mobilizable by conjugation, associated with IncX4, IncHI2, and IncI2 plasmids. In our study, mcr-1 genes have been circulating in pig farms since 2005 harbored by a variety of E. coli clones. Its persistence may be driven by co-selection since plasmids containing mcr-1 also exhibit resistance to multiple drugs used in veterinary medicine. Furthermore, this is the first report of the presence of mcr-1 gene in isolates from white storks in Spain. This finding highlights the potential importance of wildlife that forage at urban household waste landfills in the transmission and spread of colistin resistance genes.
Introduction
Antimicrobials have been used for many decades for the treatment of infectious diseases. This continued use, and in some cases, overuse has increased the emergence of resistant bacteria (Wall et al., 2016). The case of colistin is of particular interest. Colistin was introduced in human medicine in the early 1950s. Due to its toxicity when applied systemically, it was restricted to ophthalmic and topical use (Koch-Weser et al., 1970; Falagas et al., 2009). However, the emergence of pan-resistant enterobacteria causing infections in hospital settings has revived the use of colistin as the last-line treatment option. On the contrary, colistin has been prescribed in animal husbandry for the treatment of infections caused by Enterobacterales since the 1960s, especially in pig production. Furthermore, several studies carried in different European countries have identified the used of medicated feed mills supplemented with colistin for the prevention and/or treatment of post-weaning diarrhea (Trauffler et al., 2014; Cameron-Veas et al., 2016; Sjolund et al., 2016).
Resistance to colistin was attributed to chromosomal mutations that resulted in the modification in the lipid A of the LPS, reducing the binding of colistin (Rhouma et al., 2017). It was not until 2015 (Liu et al., 2016), when a plasmid-mediated mechanism of colistin resistance (mcr-1) was described for the first time, that the concern of its use reached the scientific community. The mcr-1 was first isolated in China from an Escherichia coli of pig origin and was harbored in a conjugative plasmid, facilitating its transfer to other enterobacteria. The potential spread of plasmid mediated mcr-genes from animals to humans rang the alarms of the Authorities, and the European Medicine Agency (EMA) reviewed the use of colistin in the European Union. Back in 2015, the use of colistin in swine production in Spain was estimated in 50 mg/PCU (population correction units) and a reduction target to 5 mg/PCU was set up for the next following years (EMA, 2016).
Currently, different variants of genes encoding colistin resistance have been identified (Xavier et al., 2016; Borowiak et al., 2017; Carattoli et al., 2017; Yin et al., 2017). However, mcr-1 has been isolated from enterobacteria of different origins, humans, livestock, wildlife, and the environment (Prim et al., 2016; Caltagirone et al., 2017; El Garch et al., 2018). Furthermore, in 90% of the cases, mcr-1 has been associated to IncX4, IncHI2, and IncI2 mobilizable plasmids (Matamoros et al., 2017; Elbediwi et al., 2019) facilitating its worldwide distribution (Li et al., 2017).
In addition, the emissions of antimicrobial resistant bacteria derived from human activities could have a negative effect in the environment and particularly in wildlife (Arnold et al., 2016). This is especially true for species, such as the white stork (Ciconia ciconia), that have adapted to using solid urban waste landfills as a reliable continuous food source. Spain holds the largest breeding population of storks in Europe, and due to availability of waste has also become a wintering area for storks from northern and central Europe. During the nesting period, storks are spatially bound to the surrounding areas. In some cases, these areas are waste landfills that represent a source of contamination (Gomez et al., 2016).
The aim of this study was to characterize the mechanisms involved in conferring resistance to colistin in isolates of animal origin in Spain. In particular, E. coli causing post-weaning diarrhea between consecutive batches of animals in pig farms, and in commensal E. coli isolated from white storks during the breeding season. Additionally, combining Illumina and Nanopore technology, we have completely sequenced the chromosome and closed whole plasmids of a set of isolates annotating all the genes, including those associated with antimicrobial resistance.
Materials and Methods
Study Design
From our strain collection, 70 E. coli of pig origin were randomly selected for screening the presence of colistin resistance genes. For each year, they represented 10 isolates obtained from diagnostic samples of post-weaning diarrhea collected from five different farms in consecutive batches of animals (one isolate per batch) between 2005 and 2014 (10 per year). Isolates from years 2009, 2010, and 2013 could not be found. Additionally, 20 E. coli obtained from white storks (C. ciconia) in 2011 from the same urban household waste landfill associated colony were also screened for the presence of mcr-genes. They represented fecal droppings collected at the side of different nesting platforms within the same colony. Ten of them were isolated in MacConkey agar and the other ten in MacConkey agar supplemented with cefotaxime (4 mg/L).
This study uses strains obtained from fecal samples of pigs with post-weaning diarrhea collected by the farm veterinarian and sent for diagnostics to our laboratory. Handling and sampling of the storks was carried out following all applicable international, national, and/or institutional guidelines for the care and ethical use of animals, specifically directive 2010/63/EU and Spanish laws 9/2003 and 32/2007, and Royal decrees 178/2004 and 1201/2005.
Detection of mcr Genes
All isolates were tested by PCR methods for the presence of the mcr-1, mcr-2, mcr-3, mcr-4, and mcr-5 genes as previously described (Rebelo et al., 2018). No other mechanisms of resistance were tested.
Antimicrobial Susceptibility Testing
All mcr-positive isolates were susceptibility tested using a minimum inhibitory concentration (MIC) broth microdilution (VetMIC GN-mo, National Veterinary Institute, Uppsala, Sweden). Antimicrobials tested were ampicillin (1–128 mg/L), cefotaxime (0.016–2 mg/L), ceftazidime (0.25–16 mg/L), nalidixic acid (1–128 mg/L), ciprofloxacin (0.008–1 mg/L), gentamicin (0.12–16 mg/L), streptomycin (2–256 mg/L), kanamycin (8– to 16 mg/L), chloramphenicol (2–64 mg/L), florfenicol (4–32 mg/L), trimethoprim (1–128 mg/L), sulfamethoxazole (8–1,024 mg/L), tetracycline (1–128 mg/L), and colistin (0.5–4 mg/L). Isolates were considered to be susceptible or resistant based on epidemiological cut-off values defined by EUCAST1.
Mating Experiments
To evaluate the transferability of the genes conferring resistant to colistin, filter-mating experiments were performed with all isolates as donors and E. coli HB101 rifampicin resistant as recipient. Conjugations were carried out as described before (Bielak et al., 2011). Briefly, 500 uL of the mixture of each donor and recipient were placed in a sterile paper filter on a blood agar plate. After overnight incubation, the filter was diluted in PBS and 100 uL were spread on LB agar plates containing rifampicin (100 mg/L) and colistin (2 mg/L).
Whole Genome Sequencing, Contigs Assembly and Annotation
mcr-positive isolates were selected for WGS studies. DNA was extracted using QIAGEN DNeasy® Ultraclean Microbial Kit (Germany) following the manufacture’s recommendations. Short sequencing reads for each strain was generated using MiSeq Illumina sequencing with the MiSeq V3 kit using 2 × 250 base pair paired-end chemistry (Illumina, San Diego, CA, United States) according to manufacturer’s instructions, at 160–180× coverage. The libraries for the MiSeq were constructed using 100 ng of genomic DNA using Nextera DNA Flex kit (Illumina), according to manufacturer’s instructions. The genomes for each strain were de novo assembled using CLC Genomics Workbench v9.5.2 (QIAGEN) using defaults settings except that the minimum contig size threshold was set to 500 bp in length.
Based on results from the MiSeq assemble, five isolates harboring mcr-1 and IncHI2 replicons in the same contig, and therefore, with the two genes presumably located in the same plasmid were selected for further studies. They represented isolates of different origin collected during the study period and with the same replicon family. The whole genomes were sequenced and closed by using a combination of long reads and the short reads generated earlier. The long reads sequences for each strain were generated through nanopore sequencing using a MinION device (Nanopore, Oxford, United Kingdom). The sequencing libraries were prepared using the rapid barcoding sequencing kit RBK004 and run in a FLO-MIN106 (R9.4.1) flow cell, according to manufacturer’s instructions (Nanopore), for 48 h, at 50–130× average coverage. The sequencing library contained DNA fragmented randomly by a transposase present in the Fragmentation Mix of the RBK004 kit, rendering fragments >30 kb. The run was base called live using default settings (Minknow v18.05.5, Albacore). The genomes for each strain were obtained by de novo assembly, using Nanopore data and default settings within CANU program v1.6 (Koren et al., 2017).
A second assembly was generated using a SPAdes v3.11.1 (Bankevich et al., 2012) hybrid assembly (with default settings) using both Nanopore and MiSeq data generated for each strain. The final assembly (FA) was performed by comparing the SPAdes hybrid and CANU assemblies using Mauve (Darling et al., 2004) and filling in the missing regions in the SPAdes assembly with the CANU assembly. The FA sequences were annotated using the NCBI Prokaryotic Genomes Automatic Annotation Pipeline (PGAAP)2 (Tatusova et al., 2016). Antimicrobial resistance genes were annotated using the Comprehensive Antibiotic Resistance Database (CARD v3) (Jia et al., 2017). Plasmid identification was carried out using PlasmidFinder 2.1 (Carattoli et al., 2014).
We performed a Basic Local Alignment Search Tool (BLAST) (Altschul et al., 1990) to detect high similar published plasmids to pGN-295 sequenced herein, and we included pSLK172-1 (CP017632.1), pRDB9 (MH924589), pS38 (KX129782.1), 180-PT54 (CP015833.1), pCHL5009T-88k (CP032939.1), and NRZ14408 (LT599829). We used BLAST Ring Generator (BRIG) to display circular comparisons between the 11 E. coli plasmids (five identified in this study and six downloaded from NCBI with high similarity to our closed plasmids). For this purpose, plasmid pGN-295 was used as reference with an upper identity threshold of 90% and a lower identity threshold of 70% and a nucleotide search using BLASTn (Supplementary Table 3).
Nucleotide Sequence Accession Numbers
The draft genome sequences of the 20 E. coli strains used in our study are available in GenBank under the accession numbers listed in the Supplementary Table 1.
Phylotyping and Phylogenetic Analyses
The Clermont et al. (2013) phylotyping scheme was performed by multiplex PCR for detecting phylotypes A, B1, B2, C, D, and F as described before (Clermont et al., 2013).
The initial identification of the strains was performed using an in silico E. coli MLST approach, based on the information available at EnteroBase for E. coli website3 and using Ridom SeqSphere + software v2.4.0 (Münster, Germany)4 to perform the in silico search. Seven housekeeping genes (dnaE, gyrB, recA, dtdS, pntA, pyrC, and tnaA), described previously for E. coli were used for determining the STs (Wirth et al., 2006).
The phylogenetic relationship of the strains was assessed by a custom core genome multilocus sequence typing (cgMLST) analysis using Ridom SeqSphere + software v2.4.0. We used the cgMLST scheme reported earlier (Lorenz et al., 2017; Gonzalez-Escalona and Kase, 2019). The genome of O157:H7 strain Sakai (NC_002695.1) was used as the reference for the cgMLST. A total of 4,651 genes were used as templates for the analysis of the E. coli strains from this study. We also added nine other genomes of known E. coli strains to establish their phylogenetic context (Supplementary Table 2). A Neighbor-Joining (NJ) tree using the appropriate genetic distances was built after the cgMLST analysis.
In silico Virulence Genes Detection
Each de novo assembled genome was screened in silico for the presence of 102 virulence genes reported for E. coli as described elsewhere (Gonzalez-Escalona and Kase, 2019).
Results
Presence of mcr-Genes
Out of 70 E. coli isolates of pig origin, 14 were positive for the presence of the mcr-1. Additionally one isolate collected in 2007 was positive for mcr-4.2. They were distributed between the study-period (Table 1). No other mcr-variants were detected. Additionally, five isolates of white stork origin obtained from 2011 were also positive, four contained mcr-1, and one contained the mcr-1.2 variant.
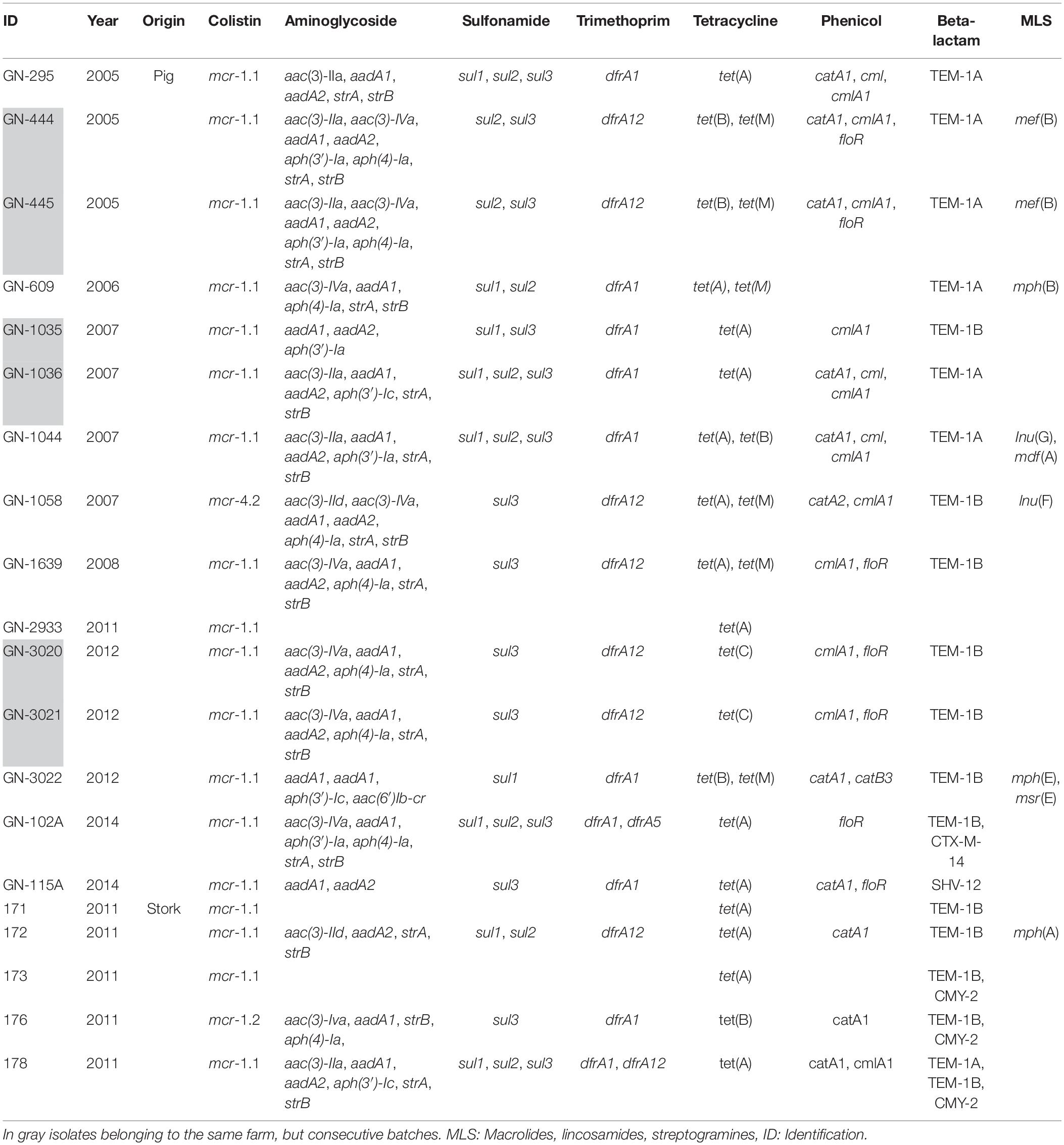
Table 1. Resistance genes for the different antimicrobials families found in isolates bearing mcr-plasmids.
Antimicrobial Susceptibility Testing and Presence of Resistance Genes
The 20 isolates exhibited a MIC to colistin ≥4 mg/L. All E. coli were phenotypically resistant to tetracycline confirmed by the presence of tet(A), tet(B), tet(C), tet(M) or the combination of some of these genes, and except for one isolate of pig origin, they were also resistant to ampicillin (Table 1) conferred by TEM-1A or TEM-1B. Furthermore, co-resistance was observed for streptomycin, kanamycin and sulfonamides (67%) with an average of six different genes coding resistance for aminoglycosides. Sixteen isolates exhibited resistance to florfenicol conferred by the presence of a unique gene or the combination of different genes. Nalidixic acid and ciprofloxacin resistance was detected in 14 isolates (56%). The presence of single point mutations in quinolone resistance-determining regions (QRDR) in chromosomal gyrA, parC, or parE genes was confirmed for these isolates. Plasmid mediated quinolone resistance genes were not detected. Except for isolate GN-2933, they were all multidrug resistant (MDR) (phenotypically resistant to at least three different families of antimicrobials). Phenotypic results correlated with the presence of the different resistance genes for each antimicrobial family. As illustrated in Table 1, the number of resistance genes varied depending of the isolate. Resistance to cephalosporins coded by CTX-M-14 and SHV-12 was observed in two isolates of pig origin and was confirmed by the MIC results (MICs for cefotaxime and ceftazidime >2 and >16 mg/L, respectively). CMY-2 was confirmed in three isolates of white stork origin.
Characterization of E. coli Bearing mcr-1 Plasmids
Among pig isolates, phylotyping identified seven isolates belonging to group A, five to group B1, two to group D and one to E. E. coli of white stork origin belonged to group A (n = 3), B1 (n = 1), and E (n = 1). B2 was never detected (Table 2).
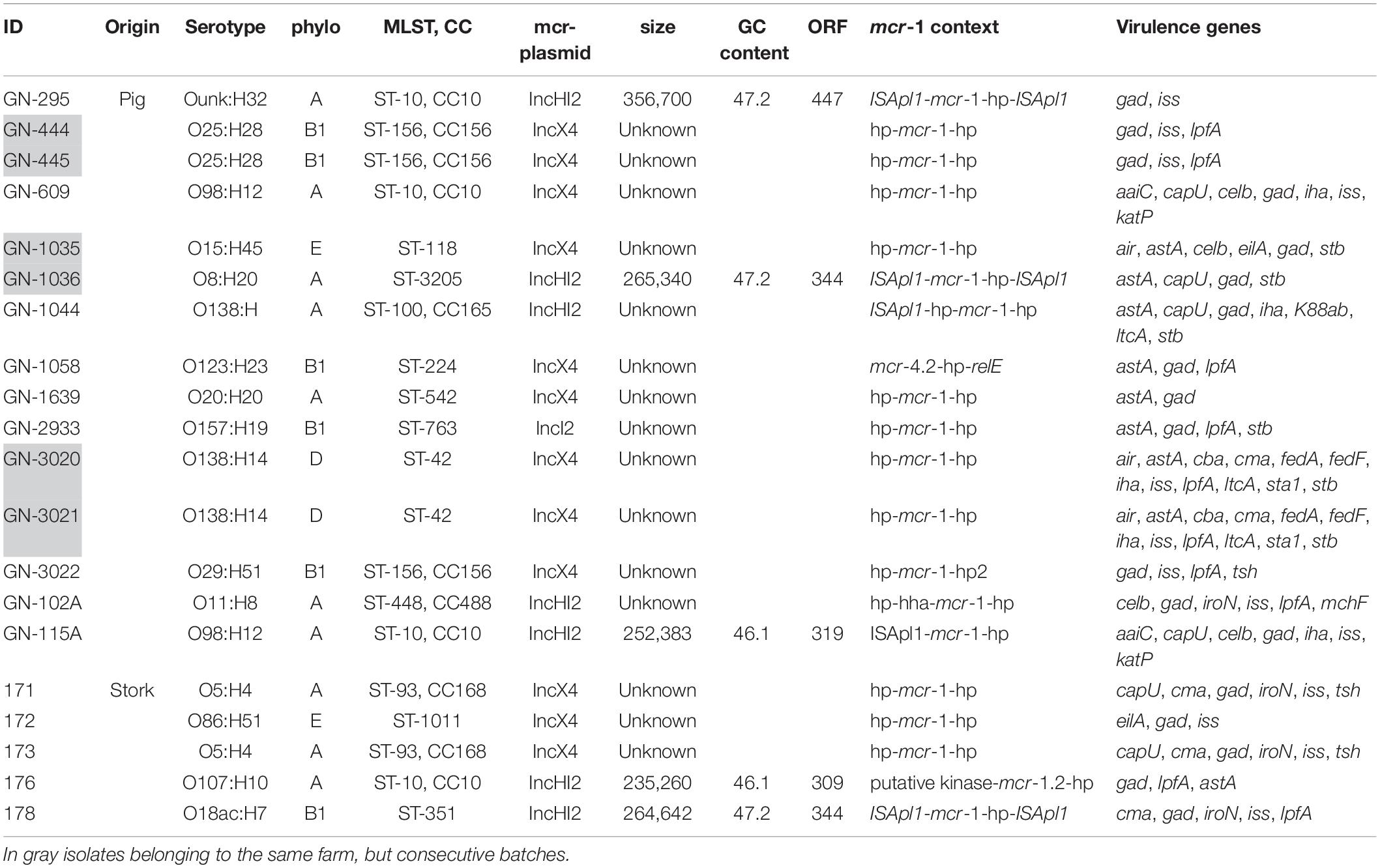
Table 2. Serotype, phylogroup, MLST, plasmid replicon associated to mcr-genes, flanking regions and virulence genes.
WGS identified a high variety of serotypes among the different isolates, with five of them of pig origin typically associated with post-weaning diarrhea, three of serotype O138, one O8, O15, and O157.
There was a polyclonal expansion of E. coli bearing mcr-1 plasmids, with 20 isolates belonging to 14 different MLST types. Some of these ST types, such as ST42, ST118, ST3205, ST763, and ST351 are not very frequent according to the MLST database (see text footnote 3). ST10 was detected in three isolates of pig origin distributed over the years. This was the only ST type shared between E. coli of pig and white stork origin. Phylogenetic studies demonstrated the presence of the same clonal linages of E. coli causing post-weaning diarrhea between different batches of animals in two different occasions; isolates GN-444 and GN-445 in 2005 and GN-3020 and GN-3021 in 2012 (Table 2). Additionally, two isolates obtained from white stork droppings (171 and 173) were clonally related (Figure 1).
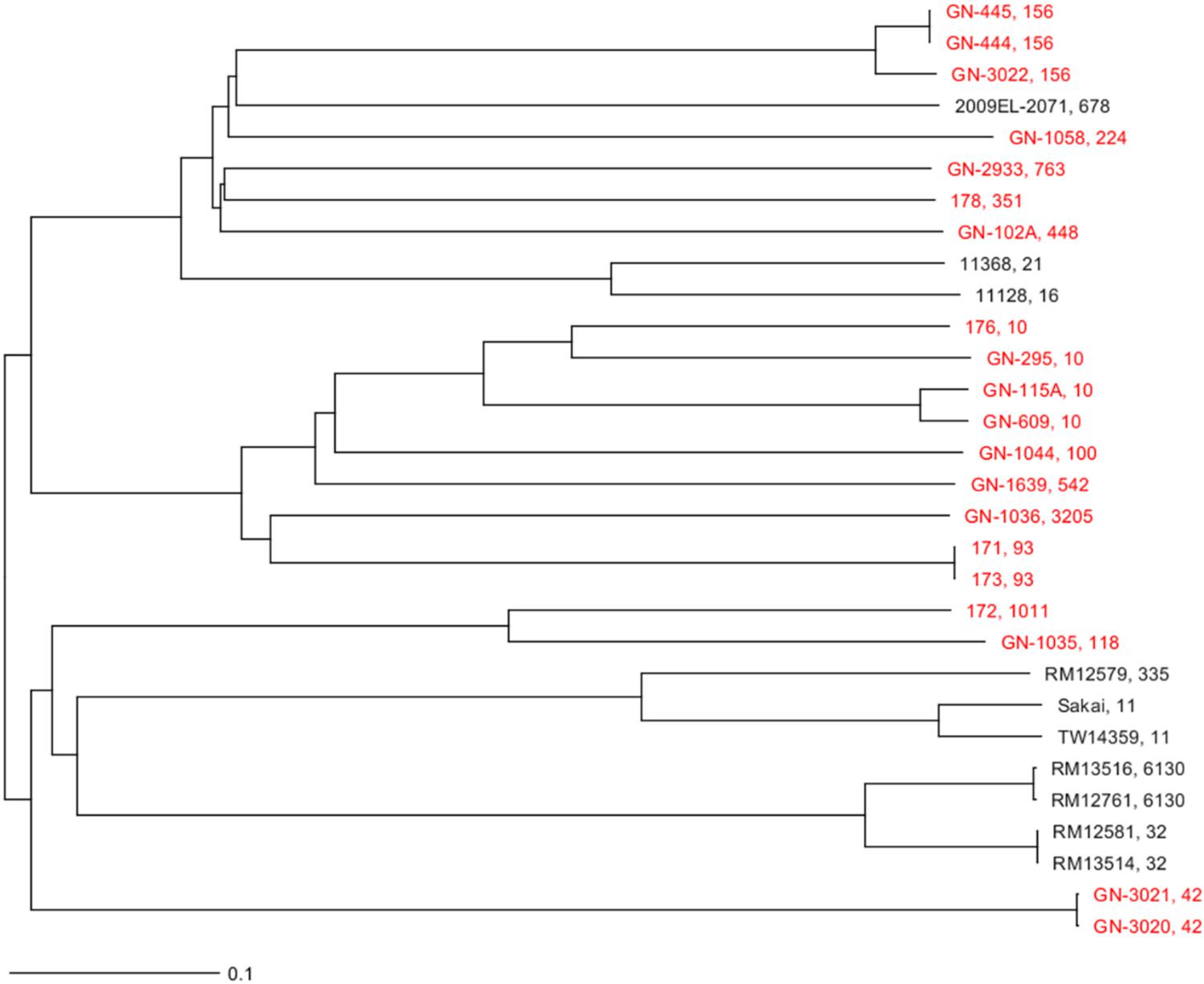
Figure 1. Neighbor joining phylogenetic tree generated by a cgMLST analysis of the strains sequenced in this study and 10 other known E. coli strains (Supplementary Table 2). The genome of E. coli O157:H7 strain Sakai (NC_002695) was used as reference. The final cgMLST analysis was based on 2514 shared loci among those strains. The sequenced strains in this study are in red. Next to the name of the strain is their ST separated by a comma.
WGS also detected a wide variety of virulence genes, ranging from 2 to 12 (Table 2), with no apparent association between number of virulence genes, phylogroup or ST type.
Plasmid Characterization and Genetic Context of the mcr-Genes
All mcr-plasmids (mcr-1 and mcr-4) successfully transferred the colistin resistant gene by conjugation to the recipient strain, demonstrating their ability to move the mcr-genes between isolates.
For all isolates, Illumina sequencing allowed to detect mcr-1 genes in the same contigs as replicons of the families IncX4 (n = 12), IncHI2 (n = 7), and IncI2 (n = 1). In the case of mcr-4.2, it was also associated to IncX4 (Table 2). Additionally, isolates contained a wide variety of replicons of different incompatibility groups presumably in different plasmids, ranging from 3 to 12. IncF and ColRNAI were the more represented (n = 15) followed by IncFIB (n = 11), IncI1 (n = 10), Col(MG8282) (n = 9) and finally, IncHI2A, IncHI2, and col156 (n = 8, for each of them).
The sizes of the five IncHI2 plasmids sequenced in this study varied between 235,260 and 356,700 bp (Table 2) and were all pMLST ST-4. In general, the architecture of the IncHI2 plasmids appeared to present high level of similarity with all of them having the conjugative transfer system, HigB-HigA toxin-antitoxin system for plasmid maintenance, macrolide resistance efflux pump mef(B) and a tellurium resistant operon (Figure 2). Except for p176, they also harbored a mercury resistance operon, which also included the genes for cobalt-zinc-cadmium resistance. In particular, pGN-115A contained the copper resistance gene pcoE. Plasmid p176 also possessed a MazEF toxin-antitoxin system. Additionally, these high molecular weight plasmids also yielded resistance genes for different families of antimicrobials, including aminoglycosides, beta-lactams, tetracycline, phenicols, sulfonamides, and trimethoprim (Table 2). Interestingly, pGN-295 with the largest molecular weight, presented and extra-DNA region highly similar to a previously sequenced plasmid named pCHL5009T-88k and detected in and E. coli of human origin in New Zealand (E. coli NRZ14408). In that region, an IncY replicon was detected.
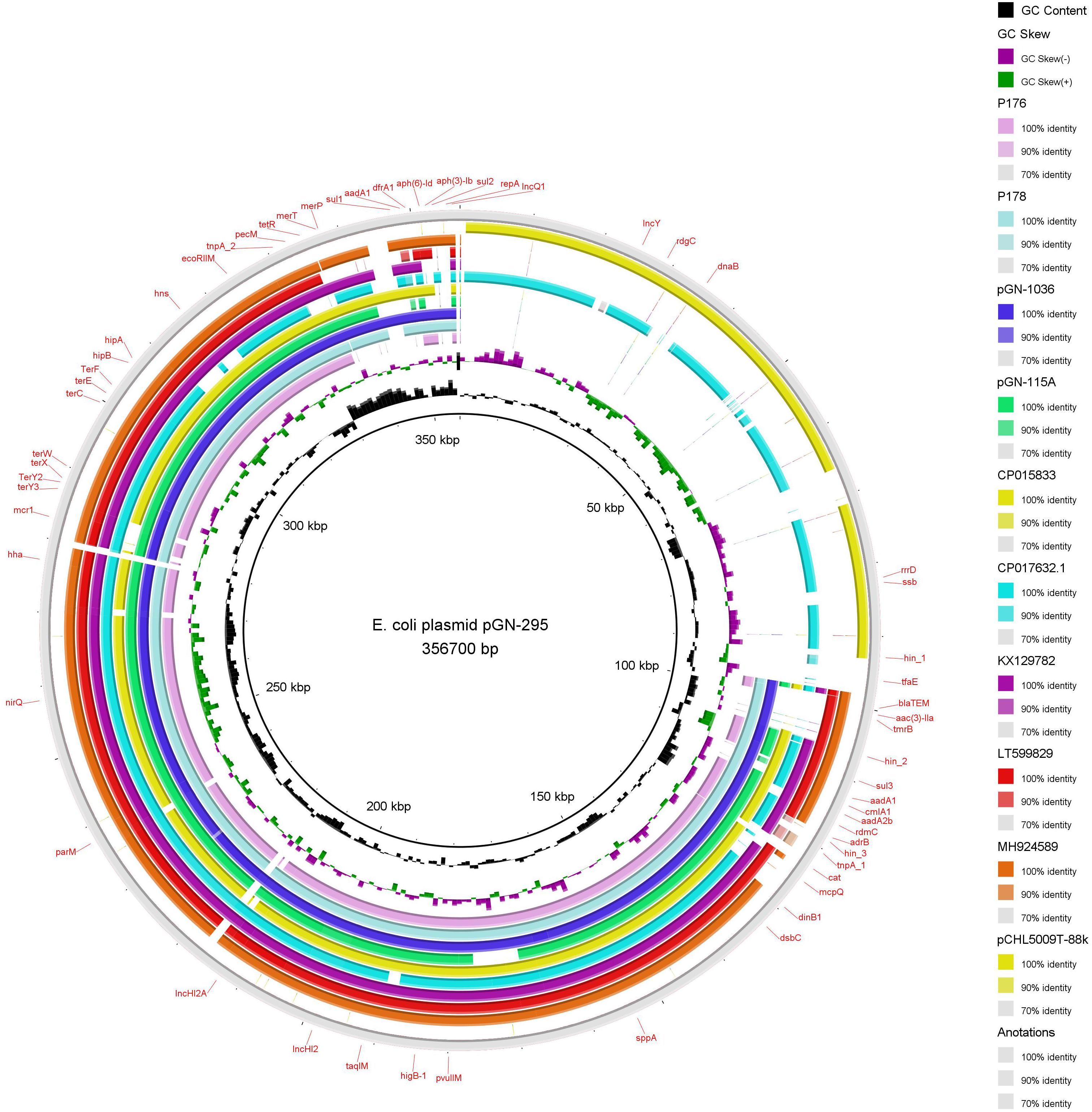
Figure 2. BRIG visualization of multiple E. coli plasmids comparisons. The solid innermost ring (black) correspond to the reference plasmid (pGN-295). After this ring plasmid p176 is shown (light purple). The third ring correspond to p178 (light blue), then pGN-1036 (navy blue) and pGN-115A (green). Additionally high identity plasmids to pGN-295 were download and added to the comparisons. The fifth (mustard) correspond to CP015833 plasmid, next CP07632.1 (aqua), KX129782 (purple), LT599829 (red), MH924589 (brown), pCHL5009T-88k (yellow), and finally we add the reference again (gray). The label in the outer ring represent the annotation on the genes associated to virulence, antibiotic resistance, stress resistance and replicons.
Furthermore, there were three different mcr-1 context or arrangements found in all these families of plasmids (Table 2). mcr-1 was flanked upstream and downstream by the ISApl1 elements, loss of the downstream ISApl1 element, and the complete loss of ISApl1 elements surrounding the mcr-1. Regarding mcr-4.2, following the gene was an ORF coding for a hypothetical protein and a relA gene.
Discussion
Plasmid mediated colistin resistance has been described worldwide in a variety of Enterobacterales of different origins, environment, food producing animals, wildlife and humans (Zurfluh et al., 2016; Guenther et al., 2017; Ovejero et al., 2017; Barlaam et al., 2019; Elbediwi et al., 2019; Lalaoui et al., 2019; Mendes Oliveira et al., 2019; Nang et al., 2019; Zając et al., 2019). In particular, in Spain, mcr-1 has been described circulating in pig farms for many years (Quesada et al., 2016; Garcia-Menino et al., 2018), and our results demonstrate its presence in pig farms in Spain as early as 2005. Although other mcr-gene variants have also been reported in Spain (Carattoli et al., 2017), studies by Garcia-Menino et al. (2018) screening large number of E. coli isolates from diagnostic cases of post-weaning diarrhea detected mcr-1 as the most common variant followed by mcr-4. Our study also confirmed similar results, although more recently described mcr-genes have not been tested herein. Furthermore, in two different occasions, carry over of the same clones causing diarrhea between batches of animals within the same farm was observed. In addition to the consumption of antimicrobials, internal biosecurity including cleaning and disinfection can play an important role in the maintenance of antimicrobial resistant bacteria in the farms (Raasch et al., 2018), facilitating its transmission and persistence between different batches of animals. However, a vertical transmission from sows to their litter cannot be disregarded.
Although we do not have antimicrobial consumption data for these farms, it is interesting to notice that in pig production in Spain, before the finding of the mcr-genes, colistin was the first choice antimicrobial for the treatment of E. coli causing post-weaning diarrhea (Moreno, 2014). The majority of these treatments were prescribed empirically instead of based on antimicrobial susceptibility testing. This fact highlights the importance of performing phenotypic tests to provide a successful therapy for the treatment of disease and to acquire epidemiological surveillance data of the phenotypes circulating in the farms. Generally, National Programs in Europe only include commensal E. coli to monitor antimicrobial resistance from food producing animals (Garcia-Migura et al., 2014), and little is known about epidemiology and resistant mechanisms present in diagnostic samples. Additionally, except for one, all of the isolates herein were MDR, impairing a successful treatment of post-weaning diarrhea and most probable facilitating the proliferation of the pathogen, since the antimicrobials would eliminate beneficial microbiota that could colonize the same niche and compete for the nutrients (Soler et al., 2018).
Wildlife is generally not treated with antibiotics but has been shown to acquire commensal microbiota carrying resistance genes when in contact with anthropized environments and livestock (Gomez et al., 2016; Darwich et al., 2019). To the best of our knowledge, this would be the first report of ST10 E. coli isolates mcr-1 positive described in white storks. The colony from which these originated is located on the premises of a solid urban waste disposal site. Acquisition of these genes is probably through foraging of feed at the landfill. Several studies have reported the presence of mcr-1 in migratory birds (Liakopoulos et al., 2016; Mohsin et al., 2016; Ruzauskas and Vaskeviciute, 2016; Tarabai et al., 2019), including co-resistance to colistin and cephalosporins. In fact, three of our isolates of white stork origin contained mcr-1 and blaCMY–2 genes within the same host. The fact that a large proportion of the European white stork population is migratory may facilitate the spread of the mcr-1 gene between different continents and emphasizes the importance of studies monitoring wildlife as potential reservoirs of resistance genes with an impact in public health along their migration routes. Furthermore, as especially juvenile storks with different geographical origins may share the same areas during wintering in Africa, this could potentially lead to cross contamination between storks from different regions (Flack et al., 2016).
The diversity of clones in terms of serotype, phylotype and ST type encountered among isolates of both, pigs and white stork origin was very high, demonstrating a wide range of hosts harboring mcr-plasmids. The unique ST type shared between isolates of both origins was ST10. ST10 is widely disseminated in humans and animals and frequently associated not only with the presence of cephalosporin resistance genes (Ojer-Usoz et al., 2017), but currently also with the occurrence of mcr-1 (Matamoros et al., 2017; Elbediwi et al., 2019). In fact, two ST10 isolates of pig and white stork origin from our study exhibited co-resistance to colistin and cephalosporins, both critically important antimicrobials to treat severe human infections caused by MDR Gram-negative in hospital settings (World Health Organization [WHO], 2017). Furthermore, ST10 has been identified as a high-risk lineage causing human extraintestinal infections (Manges et al., 2019). In Spain mcr-1 ST10 has also been described in clinical isolates causing severe infections (Lalaoui et al., 2019). The fact that food-producing animals, and wildlife associated to human settlements such as white storks that frequently nest on structures within villages and cities, could be a reservoir and source of potential pathogens with a MDR profile is worrisome, and emphasizes the need to coordinate efforts from human and veterinary sectors within a One Health approach.
Despite the clonal diversity observed among E. coli isolates in this study, only three plasmid incompatibility groups, IncX4, IncHI2, and IncI2 could be directly associated to the presence of the mcr-1 gene. These replicons are well described in the literature for their fidelity toward mcr-1 genes (Matamoros et al., 2017; Sun et al., 2017). Zurfluh et al. (2017) described IncI2 mcr-1 positive plasmids carried by E. coli strains belonging to distinct STs than the one in our study, confirming the high transmissibility of plasmids containing mcr-1 gene. Interestingly, the BLAST of our sequenced IncHI2 plasmids, retrieved similar backbones of small molecular weight plasmids with the absence of mcr-1. Furthermore, a region of pGN-295 containing an IncY replicon matched a plasmid obtained from a patient in New Zealand, pCHL5009T-88k (CP032939.1). It is impossible to find an epidemiological connection between plasmids found so far apart. However, these results suggest that some plasmids might be highly adapted to their host and able to evolve through recombination processes. They are dynamic and plastic structures prone to merge and generate mega-plasmids, providing an advantage for the bacterium.
Mating experiments demonstrated the capability of all the isolates to mobilize the resistant gene to a recipient strain, and WGS data corroborated the presence of the conjugative elements. It is worth noticing that closed IncHI2 plasmids from pigs and white storks also carried a variety of resistance genes for different families of antimicrobials (aminoglycosides, beta-lactams, tetracycline, sulfonamides, phenicols, and trimethoprim). This means that even withdrawing the use of colistin in pig farms, other common antimicrobials used during the rearing cycle in pig production such as tetracycline or beta-lactams could co-select for the presence of colistin resistance, persisting for long periods in food producing animals. Interestingly, these five IncHI2-ST4 plasmids also contained genes encoding resistance for heavy metals (mercury, cobalt, zinc, cadmium, copper and tellurium) and small multidrug resistance efflux transporter (QacE) conferring resistance to quaternary ammonium compounds (QAC). QAC have been commonly used as disinfectants with different applications in health care and the food-industry (Kaskova et al., 2007; Quinn et al., 2015). Resistance to disinfectants presumably confers these clones the capacity to survive under extreme conditions. Furthermore, since mazEF and hipBA toxin–antitoxin systems were detected, these isolates are bound to the plasmid, and losing the plasmid will trigger the pathway of cellular death (Engelberg-Kulka et al., 2005). Strategies to target these systems appear to be a promising therapy to defeat these MDR pathogens due to the bactericidal effect (Rownicki et al., 2018).
The mcr-1 flanking areas described herein have also been reported in studies elsewhere for the three-replicon families (Hadjadj et al., 2017; Zurfluh et al., 2017; Duggett et al., 2018; Zając et al., 2019). The ISApl1 flanking the mcr-1 gene appears to be a very efficient mechanism for “traveling” around, facilitating mcr-1 transposition between a limited number of incompatibility families of plasmids (Matamoros et al., 2017). Same genetic context has been identified in IncHI2 plasmids herein together with the loss of the ISApl1 element downstream the mcr-1 gene or the total absence of ISApl1, anchoring the mcr-1 gene to the plasmid. These differences in the surrounding regions of mcr-1 probably indicate different stages in the evolution of the plasmid once integrating the transposon, becoming more stable while losing the ISApl1 elements (Wang et al., 2018). In the case of mcr-4, recent studies have also detected similar flanking regions to the one described here in E. coli causing post-weaning diarrhea presumably in Italy (MG800338). Again, it is difficult to elucidate an epidemiological relation, unless providers of animals were shared at some point between European countries. More likely, mcr-genes have found a successful plasmid expansion by transposition and conjugation to perpetuate its transmission.
To conclude, we confirmed the polyclonal expansion of IncX4 and IncHI2 plasmids bearing mcr-1 in E. coli causing post-weaning diarrhea in pig farms in Spain since at least 2005. This highlights the role of food producing animals as reservoirs of antimicrobial resistant genes with impact in human health. Additionally, to the best of our knowledge this is the first report of mcr-1 detected in commensal E. coli isolated from white stork as far back as 2011. Anthropogenic pressure is highly associated to the presence of these resistance mechanisms. The study also showed the benefits of applying an advanced strategy combining Illumina and Nanopore sequencing technologies to close the genomes and plasmids as a key element to gain a basic understanding on genetic variation and dynamics of plasmid transfer. Using this innovative approach along with comprehensive analysis within a One Health context, we might be able to design strategies to minimize the emergence and persistence of resistance mechanisms within food producing animals and improve our knowledge about the spread of these mechanisms driven by forces such as migratory movements of wild birds. Hence, efforts should be made to mitigate the effect of the anthropogenic pressure and the released of antimicrobial resistance genes into the environment.
Data Availability Statement
The draft genome sequences of the 20 E. coli strains used in our study are available in GenBank under the accession numbers listed in the Supplementary Table 1.
Ethics Statement
Ethical review and approval was not required for the animal study because this study is performed with E. coli of fecal samples of animal origin stored in our freezer.
Author Contributions
LM-G designed the study. AP, YR, and UH contributed to the collection of samples. NG-E, JG-L, JM-U, JA, AM-M, and LM-G performed the sequencing, analyses, and interpretation. LM-G drafted the first manuscript. All authors contributed with the final writing, reviewing, and editing.
Funding
This work was partially supported by the grant RTI2018-095586-B-C22 from the Ministerio de Economía y Competitividad (Gobierno de España), by Plan Nacional de I+D+i 2013–2016, Instituto de Salud Carlos III and the Spanish Ministry of Health (FIS PI15/00604), and by the CERCA program from Generalitat de Catalunya. Contract of LM-G was supported by INIA and the European Social Fund.
Conflict of Interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
The handling Editor declared a shared affiliation, though no other collaboration, with the authors JG-L and AM-M at time of review.
Supplementary Material
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fmicb.2019.03072/full#supplementary-material
Footnotes
- ^ http://www.eucast.org/
- ^ http://www.ncbi.nlm.nih.gov/genome/annotation_prok
- ^ https://enterobase.warwick.ac.uk/species/index/ecoli
- ^ http://www.ridom.com/seqsphere
References
Altschul, S. F., Gish, W., Miller, W., Myers, E. W., and Lipman, D. J. (1990). Basic local alignment search tool. J. Mol. Biol. 215, 403–410. doi: 10.1016/s0022-2836(05)80360-2
Arnold, K. E., Williams, N. J., and Bennett, M. (2016). ‘Disperse abroad in the land’: the role of wildlife in the dissemination of antimicrobial resistance. Biol. Lett. 12:20160137. doi: 10.1098/rsbl.2016.0137
Bankevich, A., Nurk, S., Antipov, D., Gurevich, A. A., Dvorkin, M., Kulikov, A. S., et al. (2012). SPAdes: a new genome assembly algorithm and its applications to single-cell sequencing. J. Comput. Biol. 19, 455–477. doi: 10.1089/cmb.2012.0021
Barlaam, A., Parisi, A., Spinelli, E., Caruso, M., Taranto, P. D., and Normanno, G. (2019). Global emergence of colistin-resistant Escherichia coli in food chains and associated food safety implications: a review. J. Food Prot. 82, 1440–1448. doi: 10.4315/0362-028x.jfp-19-116
Bielak, E., Bergenholtz, R. D., Jorgensen, M. S., Sorensen, S. J., Hansen, L. H., and Hasman, H. (2011). Investigation of diversity of plasmids carrying the blaTEM-52 gene. J. Antimicrob. Chemother. 66, 2465–2474. doi: 10.1093/jac/dkr331
Borowiak, M., Fischer, J., Hammerl, J. A., Hendriksen, R. S., Szabo, I., and Malorny, B. (2017). Identification of a novel transposon-associated phosphoethanolamine transferase gene, mcr-5, conferring colistin resistance in d-tartrate fermenting Salmonella enterica subsp. enterica serovar Paratyphi B. J. Antimicrob. Chemother. 72, 3317–3324. doi: 10.1093/jac/dkx327
Caltagirone, M., Nucleo, E., Spalla, M., Zara, F., Novazzi, F., Marchetti, V. M., et al. (2017). Occurrence of extended spectrum beta-Lactamases, KPC-Type, and MCR-1.2-producing Enterobacteriaceae from wells, river water, and wastewater treatment plants in Oltrepo Pavese area, Northern Italy. Front. Microbiol. 8:2232. doi: 10.3389/fmicb.2017.02232
Cameron-Veas, K., Moreno, M. A., Fraile, L., and Migura-Garcia, L. (2016). Shedding of cephalosporin resistant Escherichia coli in pigs from conventional farms after early treatment with antimicrobials. Vet. J. 211, 21–25. doi: 10.1016/j.tvjl.2016.02.017
Carattoli, A., Villa, L., Feudi, C., Curcio, L., Orsini, S., Luppi, A., et al. (2017). Novel plasmid-mediated colistin resistance mcr-4 gene in Salmonella and Escherichia coli, Italy 2013, Spain and Belgium, 2015 to 2016. Euro Surveill. 22:30589. doi: 10.2807/1560-7917.es.2017.22.31.30589
Carattoli, A., Zankari, E., Garcia-Fernandez, A., Voldby Larsen, M., Lund, O., Villa, L., et al. (2014). In silico detection and typing of plasmids using PlasmidFinder and plasmid multilocus sequence typing. Antimicrob. Agents Chemother. 58, 3895–3903. doi: 10.1128/aac.02412-14
Clermont, O., Christenson, J. K., Denamur, E., and Gordon, D. M. (2013). The Clermont Escherichia coli phylo-typing method revisited: improvement of specificity and detection of new phylo-groups. Environ. Microbiol. Rep. 5, 58–65. doi: 10.1111/1758-2229.12019
Darling, A. C., Mau, B., Blattner, F. R., and Perna, N. T. (2004). Mauve: multiple alignment of conserved genomic sequence with rearrangements. Genome Res. 14, 1394–1403. doi: 10.1101/gr.2289704
Darwich, L., Vidal, A., Seminati, C., Albamonte, A., Casado, A., Lopez, F., et al. (2019). High prevalence and diversity of extended-spectrum beta-lactamase and emergence of OXA-48 producing Enterobacterales in wildlife in Catalonia. PLoS One 14:e0210686. doi: 10.1371/journal.pone.0210686
Duggett, N. A., Randall, L. P., Horton, R. A., Lemma, F., Kirchner, M., Nunez-Garcia, J., et al. (2018). Molecular epidemiology of isolates with multiple mcr plasmids from a pig farm in Great Britain: the effects of colistin withdrawal in the short and long term. J. Antimicrob. Chemother. 73, 3025–3033. doi: 10.1093/jac/dky292
El Garch, F., de Jong, A., Bertrand, X., Hocquet, D., and Sauget, M. (2018). mcr-1-like detection in commensal Escherichia coli and Salmonella spp. from food-producing animals at slaughter in Europe. Vet. Microbiol. 213, 42–46. doi: 10.1016/j.vetmic.2017.11.014
Elbediwi, M., Li, Y., Paudyal, N., Pan, H., Li, X., Xie, S., et al. (2019). Global burden of colistin-resistant bacteria: mobilized colistin resistance genes study (1980-2018). Microorganisms 7:461. doi: 10.3390/microorganisms7100461
EMA (2016). Sales of Veterinary Antimicrobial Agents in 29 European Countries in 2014. Report No 6. Brussels: EU Publications.
Engelberg-Kulka, H., Hazan, R., and Amitai, S. (2005). mazEF: a chromosomal toxin-antitoxin module that triggers programmed cell death in bacteria. J. Cell Sci. 118(Pt 19), 4327–4332. doi: 10.1242/jcs.02619
Falagas, M. E., Vouloumanou, E. K., and Rafailidis, P. I. (2009). Systemic colistin use in children without cystic fibrosis: a systematic review of the literature. Int. J. Antimicrob. Agents 33, 503.e1–503.e13. doi: 10.1016/j.ijantimicag.2008.10.021
Flack, A., Fiedler, W., Blas, J., Pokrovsky, I., Kaatz, M., Mitropolsky, M., et al. (2016). Costs of migratory decisions: a comparison across eight white stork populations. Sci. Adv. 2:e1500931. doi: 10.1126/sciadv.1500931
Garcia-Menino, I., Garcia, V., Mora, A., Diaz-Jimenez, D., Flament-Simon, S. C., Alonso, M. P., et al. (2018). Swine enteric colibacillosis in Spain: pathogenic potential of mcr-1 ST10 and ST131 E. coli isolates. Front. Microbiol. 9:2659. doi: 10.3389/fmicb.2018.02659
Garcia-Migura, L., Hendriksen, R. S., Fraile, L., and Aarestrup, F. M. (2014). Antimicrobial resistance of zoonotic and commensal bacteria in Europe: the missing link between consumption and resistance in veterinary medicine. Vet. Microbiol. 170, 1–9. doi: 10.1016/j.vetmic.2014.01.013
Gomez, P., Lozano, C., Camacho, M. C., Lima-Barbero, J. F., Hernandez, J. M., Zarazaga, M., et al. (2016). Detection of MRSA ST3061-t843-mecC and ST398-t011-mecA in white stork nestlings exposed to human residues. J. Antimicrob. Chemother. 71, 53–57. doi: 10.1093/jac/dkv314
Gonzalez-Escalona, N., and Kase, J. A. (2019). Virulence gene profiles and phylogeny of Shiga toxin-positive Escherichia coli strains isolated from FDA regulated foods during 2010-2017. PLoS One 14:e0214620. doi: 10.1371/journal.pone.0214620
Guenther, S., Falgenhauer, L., Semmler, T., Imirzalioglu, C., Chakraborty, T., Roesler, U., et al. (2017). Environmental emission of multiresistant Escherichia coli carrying the colistin resistance gene mcr-1 from German swine farms. J. Antimicrob. Chemother. 72, 1289–1292. doi: 10.1093/jac/dkw58510.1093/jac/dkw585
Hadjadj, L., Riziki, T., Zhu, Y., Li, J., Diene, S. M., and Rolain, J. M. (2017). Study of mcr-1 gene-mediated colistin resistance in Enterobacteriaceae isolated from humans and animals in different countries. Genes (Basel) 8:394. doi: 10.3390/genes8120394
Jia, B., Raphenya, A. R., Alcock, B., Waglechner, N., Guo, P., Tsang, K. K., et al. (2017). CARD 2017: expansion and model-centric curation of the comprehensive antibiotic resistance database. Nucleic Acids Res. 45, D566–D573. doi: 10.1093/nar/gkw1004
Kaskova, A., Ondrasovicova, O., Vargova, M., Ondrasovic, M., and Venglovsky, J. (2007). Application of peracetic acid and quarternary ammonium disinfectants as a part of sanitary treatment in a poultry house and poultry processing plant. Zoonoses Public Health 54, 125–130. doi: 10.1111/j.1863-2378.2007.00987.x
Koch-Weser, J., Sidel, V. W., Federman, E. B., Kanarek, P., Finer, D. C., and Eaton, A. E. (1970). Adverse effects of sodium colistimethate. Manifestations and specific reaction rates during 317 courses of therapy. Ann. Intern. Med. 72, 857–868. doi: 10.7326/0003-4819-72-6-857
Koren, S., Walenz, B. P., Berlin, K., Miller, J. R., Bergman, N. H., and Phillippy, A. M. (2017). Canu: scalable and accurate long-read assembly via adaptive k-mer weighting and repeat separation. Genome Res. 27, 722–736. doi: 10.1101/gr.215087.116
Lalaoui, R., Djukovic, A., Bakour, S., Sanz, J., Gonzalez-Barbera, E. M., Salavert, M., et al. (2019). Detection of plasmid-mediated colistin resistance, mcr-1 gene, in Escherichia coli isolated from high-risk patients with acute leukemia in Spain. J. Infect. Chemother. 25, 605–609. doi: 10.1016/j.jiac.2019.03.007
Li, R., Xie, M., Zhang, J., Yang, Z., Liu, L., Liu, X., et al. (2017). Genetic characterization of mcr-1-bearing plasmids to depict molecular mechanisms underlying dissemination of the colistin resistance determinant. J. Antimicrob. Chemother. 72, 393–401. doi: 10.1093/jac/dkw411
Liakopoulos, A., Mevius, D. J., Olsen, B., and Bonnedahl, J. (2016). The colistin resistance mcr-1 gene is going wild. J. Antimicrob. Chemother. 71, 2335–2336. doi: 10.1093/jac/dkw262
Liu, Y. Y., Wang, Y., Walsh, T. R., Yi, L. X., Zhang, R., Spencer, J., et al. (2016). Emergence of plasmid-mediated colistin resistance mechanism MCR-1 in animals and human beings in China: a microbiological and molecular biological study. Lancet Infect. Dis. 16, 161–168. doi: 10.1016/s1473-3099(15)00424-7
Lorenz, S. C., Gonzalez-Escalona, N., Kotewicz, M. L., Fischer, M., and Kase, J. A. (2017). Genome sequencing and comparative genomics of enterohemorrhagic Escherichia coli O145:H25 and O145:H28 reveal distinct evolutionary paths and marked variations in traits associated with virulence & colonization. BMC Microbiol. 17:183. doi: 10.1186/s12866-017-1094-3
Manges, A. R., Geum, H. M., Guo, A., Edens, T. J., Fibke, C. D., and Pitout, J. D. D. (2019). Global extraintestinal pathogenic Escherichia coli (ExPEC) lineages. Clin. Microbiol. Rev. 32:e00135-18, doi: 10.1128/cmr.00135-18
Matamoros, S., van Hattem, J. M., Arcilla, M. S., Willemse, N., Melles, D. C., Penders, J., et al. (2017). Global phylogenetic analysis of Escherichia coli and plasmids carrying the mcr-1 gene indicates bacterial diversity but plasmid restriction. Sci. Rep. 7:15364. doi: 10.1038/s41598-017-15539-7
Mendes Oliveira, V. R., Paiva, M. C., and Lima, W. G. (2019). Plasmid-mediated colistin resistance in Latin America and Caribbean: a systematic review. Travel Med. Infect. Dis. 31, 101459. doi: 10.1016/j.tmaid.2019.07.015
Mohsin, M., Raza, S., Roschanski, N., Schaufler, K., and Guenther, S. (2016). First description of plasmid-mediated colistin-resistant extended-spectrum beta-lactamase-producing Escherichia coli in a wild migratory bird from Asia. Int. J. Antimicrob. Agents 48, 463–464. doi: 10.1016/j.ijantimicag.2016.07.001
Moreno, M. A. (2014). Survey of quantitative antimicrobial consumption per production stage in farrow-to-finish pig farms in Spain. Vet. Rec. Open 1:e000002. doi: 10.1136/vropen-2013-000002
Nang, S. C., Li, J., and Velkov, T. (2019). The rise and spread of mcr plasmid-mediated polymyxin resistance. Crit. Rev. Microbiol. 45, 131–161. doi: 10.1080/1040841x.2018.1492902
Ojer-Usoz, E., Gonzalez, D., and Vitas, A. I. (2017). Clonal Diversity of ESBL-producing Escherichia coli isolated from environmental, human and food samples. Int. J. Environ. Res. Public Health 14:676. doi: 10.3390/ijerph14070676
Ovejero, C. M., Delgado-Blas, J. F., Calero-Caceres, W., Muniesa, M., and Gonzalez-Zorn, B. (2017). Spread of mcr-1-carrying Enterobacteriaceae in sewage water from Spain. J. Antimicrob. Chemother. 72, 1050–1053. doi: 10.1093/jac/dkw533
Prim, N., Rivera, A., Rodriguez-Navarro, J., Espanol, M., Turbau, M., Coll, P., et al. (2016). Detection of mcr-1 colistin resistance gene in polyclonal Escherichia coli isolates in Barcelona, Spain, 2012 to 2015. Euro Surveill. 21:30183. doi: 10.2807/1560-7917.es.2016.21.13.30183
Quesada, A., Ugarte-Ruiz, M., Iglesias, M. R., Porrero, M. C., Martinez, R., Florez-Cuadrado, D., et al. (2016). Detection of plasmid mediated colistin resistance (MCR-1) in Escherichia coli and Salmonella enterica isolated from poultry and swine in Spain. Res. Vet. Sci. 105, 134–135. doi: 10.1016/j.rvsc.2016.02.003
Quinn, M. M., Henneberger, P. K., Braun, B., Delclos, G. L., Fagan, K., Huang, V., et al. (2015). Cleaning and disinfecting environmental surfaces in health care: toward an integrated framework for infection and occupational illness prevention. Am. J. Infect. Control 43, 424–434. doi: 10.1016/j.ajic.2015.01.029
Raasch, S., Postma, M., Dewulf, J., Stark, K. D. C., and Grosse Beilage, E. (2018). Association between antimicrobial usage, biosecurity measures as well as farm performance in German farrow-to-finish farms. Porcine Health Manag. 4, 30. doi: 10.1186/s40813-018-0106-5
Rebelo, A. R., Bortolaia, V., Kjeldgaard, J. S., Pedersen, S. K., Leekitcharoenphon, P., Hansen, I. M., et al. (2018). Multiplex PCR for detection of plasmid-mediated colistin resistance determinants, mcr-1, mcr-2, mcr-3, mcr-4 and mcr-5 for surveillance purposes. Euro Surveill. 23:17–00672. doi: 10.2807/1560-7917.es.2018.23.6.17-00672
Rhouma, M., Fairbrother, J. M., Beaudry, F., and Letellier, A. (2017). Post weaning diarrhea in pigs: risk factors and non-colistin-based control strategies. Acta Vet. Scand. 59:31. doi: 10.1186/s13028-017-0299-7
Rownicki, M., Pienko, T., Czarnecki, J., Kolanowska, M., Bartosik, D., and Trylska, J. (2018). Artificial activation of Escherichia coli mazEF and hipBA toxin-antitoxin systems by antisense peptide nucleic acids as an antibacterial strategy. Front. Microbiol. 9:2870. doi: 10.3389/fmicb.2018.02870
Ruzauskas, M., and Vaskeviciute, L. (2016). Detection of the mcr-1 gene in Escherichia coli prevalent in the migratory bird species Larus argentatus. J. Antimicrob. Chemother. 71, 2333–2334. doi: 10.1093/jac/dkw245
Sjolund, M., Postma, M., Collineau, L., Losken, S., Backhans, A., Belloc, C., et al. (2016). Quantitative and qualitative antimicrobial usage patterns in farrow-to-finish pig herds in Belgium, France, Germany and Sweden. Prev. Vet. Med. 130, 41–50. doi: 10.1016/j.prevetmed.2016.06.003
Soler, C., Goossens, T., Bermejo, A., Migura-Garcia, L., Cusco, A., Francino, O., et al. (2018). Digestive microbiota is different in pigs receiving antimicrobials or a feed additive during the nursery period. PLoS One 13:e0197353. doi: 10.1371/journal.pone.0197353
Sun, J., Xu, Y., Gao, R., Lin, J., Wei, W., Srinivas, S., et al. (2017). Deciphering MCR-2 colistin resistance. MBio 8:e00625-17. doi: 10.1128/mBio.00625-17
Tarabai, H., Valcek, A., Jamborova, I., Vazhov, S. V., Karyakin, I. V., Raab, R., et al. (2019). Plasmid-Mediated mcr-1 colistin resistance in Escherichia coli from a black kite in Russia. Antimicrob. Agents Chemother. 63:e01266-19. doi: 10.1128/aac.01266-19
Tatusova, T., DiCuccio, M., Badretdin, A., Chetvernin, V., Nawrocki, E. P., Zaslavsky, L., et al. (2016). NCBI prokaryotic genome annotation pipeline. Nucleic Acids Res. 44, 6614–6624. doi: 10.1093/nar/gkw569
Trauffler, M., Griesbacher, A., Fuchs, K., and Kofer, J. (2014). Antimicrobial drug use in Austrian pig farms: plausibility check of electronic on-farm records and estimation of consumption. Vet. Rec. 175, 402. doi: 10.1136/vr.102520
Wang, R., van Dorp, L., Shaw, L. P., Bradley, P., Wang, Q., Wang, X., et al. (2018). The global distribution and spread of the mobilized colistin resistance gene mcr-1. Nat. Commun. 9:1179. doi: 10.1038/s41467-018-03205-z
Wall, B. A., Mateus, A., Marshall, L., and Pfeiffer, D. U. (2016). Drivers, Dynamics and Epidemiology of Antimicrobial Resistance in Animal Production. Rome: Food and Agriculture Organization.
Wirth, T., Falush, D., Lan, R., Colles, F., Mensa, P., Wieler, L. H., et al. (2006). Sex and virulence in Escherichia coli: an evolutionary perspective. Mol. Microbiol. 60, 1136–1151. doi: 10.1111/j.1365-2958.2006.05172.x
World Health Organization [WHO] (2017). Critically Important Antimicrobials for Human Medicine, 5th Edn. Geneva: World Health Organization.
Xavier, B. B., Lammens, C., Ruhal, R., Kumar-Singh, S., Butaye, P., Goossens, H., et al. (2016). Identification of a novel plasmid-mediated colistin-resistance gene, mcr-2, in Escherichia coli, Belgium, June 2016. Euro Surveill. 21:30280. doi: 10.2807/1560-7917.es.2016.21.27.30280
Yin, W., Li, H., Shen, Y., Liu, Z., Wang, S., Shen, Z., et al. (2017). Novel plasmid-mediated Colistin resistance gene mcr-3 in Escherichia coli. MBio 8:e00543-17. doi: 10.1128/mBio.00543-17
Zając, M., Sztromwasser, P., Bortolaia, V., Leekitcharoenphon, P., Cavaco, L. M., Ziȩtek-Barszcz, A., et al. (2019). Occurrence and characterization of mcr-1-positive Escherichia coli isolated from food-producing animals in Poland, 2011–2016. Front. Microbiol. 10:1753. doi: 10.3389/fmicb.2019.01753
Zurfluh, K., Klumpp, J., Nüesch-Inderbinen, M., and Stephan, R. (2016). Full-length nucleotide sequences of mcr-1-harboring plasmids isolated from extended-spectrum-β-lactamase-producing Escherichia coli isolates of different origins. Antimicrob. Agents Chemother. 60, 5589–5591. doi: 10.1128/aac.00935-16
Keywords: Escherichia coli, colistin, mcr-plasmids, MinION nanopore, pigs, storks, Spain
Citation: Migura-Garcia L, González-López JJ, Martinez-Urtaza J, Aguirre Sánchez JR, Moreno-Mingorance A, Perez de Rozas A, Höfle U, Ramiro Y and Gonzalez-Escalona N (2020) mcr-Colistin Resistance Genes Mobilized by IncX4, IncHI2, and IncI2 Plasmids in Escherichia coli of Pigs and White Stork in Spain. Front. Microbiol. 10:3072. doi: 10.3389/fmicb.2019.03072
Received: 04 October 2019; Accepted: 19 December 2019;
Published: 17 January 2020.
Edited by:
Daniel Yero, Autonomous University of Barcelona, SpainReviewed by:
Roger Stephan, University of Zurich, SwitzerlandMagdalena Maria Zaja̧c, National Veterinary Research Institute (NVRI), Poland
Min Yue, Zhejiang University, China
Copyright © 2020 Migura-Garcia, González-López, Martinez-Urtaza, Aguirre Sánchez, Moreno-Mingorance, Perez de Rozas, Höfle, Ramiro and Gonzalez-Escalona. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Lourdes Migura-Garcia, lourdes.migura@irta.cat