 Yanmei Li1
†Teng-Yi Huang2
†
Yanmei Li1
†Teng-Yi Huang2
† Yuzhu Mao3
Yanni Chen3
Fan Shi3
Ruixin Peng3
Jinxuan Chen3
Caiying Bai4
Ling Chen3
,5
Kan Wang6
*
Yuzhu Mao3
Yanni Chen3
Fan Shi3
Ruixin Peng3
Jinxuan Chen3
Caiying Bai4
Ling Chen3
,5
Kan Wang6
* Junyan Liu7
*
Junyan Liu7
*- 1Department of Haematology, Guangzhou Women and Children's Medical Center, Guangzhou Medical University, Guangzhou, China
- 2Department of Laboratory Medicine, The Second Affiliated Hospital of Shantou University Medical College, Shantou, China
- 3School of Food Science and Engineering, Guangdong Province Key Laboratory for Green Processing of Natural Products and Product Safety, South China University of Technology, Guangzhou, China
- 4Guangdong Women and Children Hospital, Guangzhou, China
- 5Research Institute for Food Nutrition and Human Health, Guangzhou, China
- 6Research Center for Translational Medicine, The Second Affiliated Hospital, Medical College of Shantou University, Shantou, China
- 7Department of Civil and Environmental Engineering, University of Maryland, College Park, MD, United States
Objective: This study aimed to investigate the effect of environmental conditions including nutrient content, acetic acid concentration, salt concentration, and temperature on the formation of viable but nonculturable (VBNC) state of Pediococcus acidilactici, as well as its control and detection in food system.
Methods: Representing various environmental conditions in different food systems, 16 induction groups were designed for the formation of VBNC state of P. acidilactici. Traditional plate counting was applied to measure the culturable cell numbers, and Live/Dead Bacterial Viability Kit combined with fluorescent microscopy was used to identify viable cells numbers. The inhibition of bacterial growth and VBNC state formation by adjusting the environmental conditions were investigated, and the clearance effect of VBNC cells in crystal cake system was studied. In addition, a propidium monoazide-polymerase chain reaction (PMA-PCR) assay was applied to detect the VBNC P. acidilactici cells in crystal cake food system.
Results: Among the environmental conditions included in this study, acetic acid concentration had the greatest effect on the formation of VBNC state of P. acidilactici, followed by nutritional conditions and salt concentration. Reducing nutrients in the environment and treating with 1.0% acetic acid can inhibit P. acidilactici from entering the VBNC state. In the crystal cake system, the growth of P. acidilactici and the formation of VBNC state can be inhibited by adding 1.0% acetic acid and storing at −20°C. In crystal cake system, the PMA-PCR assay can be used to detect VBNC P. acidilactici cells at a concentration higher than 104 cells/ml.
Conclusion: The VBNC state of P. acidilactici can be influenced by the changing of environmental conditions, and PMA-PCR assay can be applied in food system for the detection of VBNC P. acidilactici cells.
Highlights
1. Acetic acid concentration had the greatest effect on the formation of VBNC state of P. acidilactici, followed by nutritional conditions and salt concentration.
2. Reducing nutrients and treating with 1.0% acetic acid are able to inhibit P. acidilactici from entering into the VBNC state.
3. In the crystal cake food system, the growth of P. acidilactici and the formation of VBNC state can be inhibited by adding 1.0% acetic acid and storing at −20°C.
4. In the crystal cake food system, the PMA-PCR assay can be used to detect VBNC P. acidilactici cells at a concentration higher than 104 cells/ml.
Introduction
Under stress conditions, various species of bacteria can enter a physiologically viable but nonculturable (VBNC) state. Bacteria entering into the VBNC state are a survival strategy to cope with adverse conditions. However, food-borne spoilage and pathogenic bacteria pose a threat to food safety and even public health after entering the VBNC state (Bao et al., 2017a,b; Wen et al., 2020). The presence of VBNC state bacteria can lead to false negative results from the traditional plate counting detection. In addition, VBNC cells retain viability and can cause fatal infections even when completely uncultivable. Bacteria can recover from the VBNC state to an active metabolic state, bringing certain safety risks to human health and food safety (Du et al., 2007; Xu et al., 2011a; Lin et al., 2017; Miao et al., 2017a; Xie et al., 2017a; Hu et al., 2019).
Lactic acid bacteria are widely distributed and are usually found in food products, including meat, milk, and vegetables. When lactic acid bacteria are subjected to external environmental pressures during the production and application process, they are capable of activating a variety of stress reactions and may enter into the VBNC state (Xu et al., 2012a, 2017a; Xie et al., 2017b; Jia et al., 2018; Liu et al., 2018c, 2019). Studies had shown that a variety of lactic acid bacteria are able to enter into the VBNC state, including Lactobacillus plantarum (Liu et al., 2017a; Olszewska and Bialobrzewski, 2019), Lactobacillus acetate (Piao et al., 2019), Lactobacillus lindenii (Liu et al., 2017b), Bifidobacteria, etc. However, few studies on the VBNC of Pediococcus had been reported. Pediococcus acidilactici is a homofermentative bacterium that can grow in a wide range of pH, temperature, and osmotic pressure (Klaenhammer, 1993; Xu et al., 2007; Miao et al., 2017b,c). They are commonly found in fermented vegetables, fermented dairy products, and meat (Barros et al., 2001; Bao et al., 2017c; Xu et al., 2017b).
Lactic acid bacteria contaminated food is mainly caused by incomplete sterilization after fermentation. It is also possible that lactic acid bacteria enter into the VBNC state under environmental pressure and causes false negative detection during shipment. During food storage, transportation, and marketing, bacteria in VBNC state resume activity, causing food spoilage (Lin et al., 2016; Miao et al., 2016; Xu et al., 2016a,b,c). Therefore, the detection of VBNC state bacteria is of great significance. The methods commonly used for the detection of VBNC state bacteria are currently based on micro-optic, immunological, and molecular detection techniques (You et al., 2012; Zhong et al., 2013). Propidium monoazide-polymerase chain reaction (PMA-PCR) is a method for detecting bacteria in VBNC state based on conventional PCR technology developed in recent years (Xu et al., 2008a, 2018; Miao et al., 2018; Zhao et al., 2018a,b). This method overcomes the defect that conventional PCR technology cannot distinguish free DNA released from living bacteria and dead bacteria, and can effectively avoid false positive test results (Xu et al., 2008b; Liu et al., 2018a).
This study aimed to investigate the effect of environmental conditions, including nutrient content, acetic acid concentration, and salt concentration on the formation of VBNC state of P. acidilactici, as well as its control and detection in food system.
Materials and Methods
Bacterial Strains and Culture Conditions
Pediococcus acidilactici BM-PA17927 was stored at −80°C in MRS broth containing 20% (v/v) glycerol. It was streaked on MRS agar plate for 48 h at 37°C. Subsequently, a single colony was inoculated into a culture tube containing 2 ml of MRS broth and cultured at 37°C for 24 h. P. acidilactici was grown overnight in MRS broth at 37°C for 12 h. The bacterial suspension was diluted 1:100 in fresh MRS medium. Subsequently, the cultivation continued to the log phase.
Induction of Entry Into VBNC State
According to the optimal culture conditions of P. acidilactici, and in accordance with the principle of reverse adjustment, conditions which are unfavorable to bacterial growth were selected as candidate factors for inducing VBNC state. Three factors, including nutrition, salt concentration, and acid, were taken as a single variable. Three factors and four levels of orthogonal experiments were designed to reach 16 experimental groups (Table 1). The 16 experimental groups were stored at 4 and − 20°C, respectively. The change trend of the culturable cell number of bacteria was used as an observation index to investigate the influence of environmental pressure on the formation of VBNC state of P. acidilactici. P. acidilactici cultured to logarithmic phase was inoculated into the induction culture, and the concentration of P. acidilactici in the induction culture was guaranteed to be approximately 1 × 108 cells/ml.
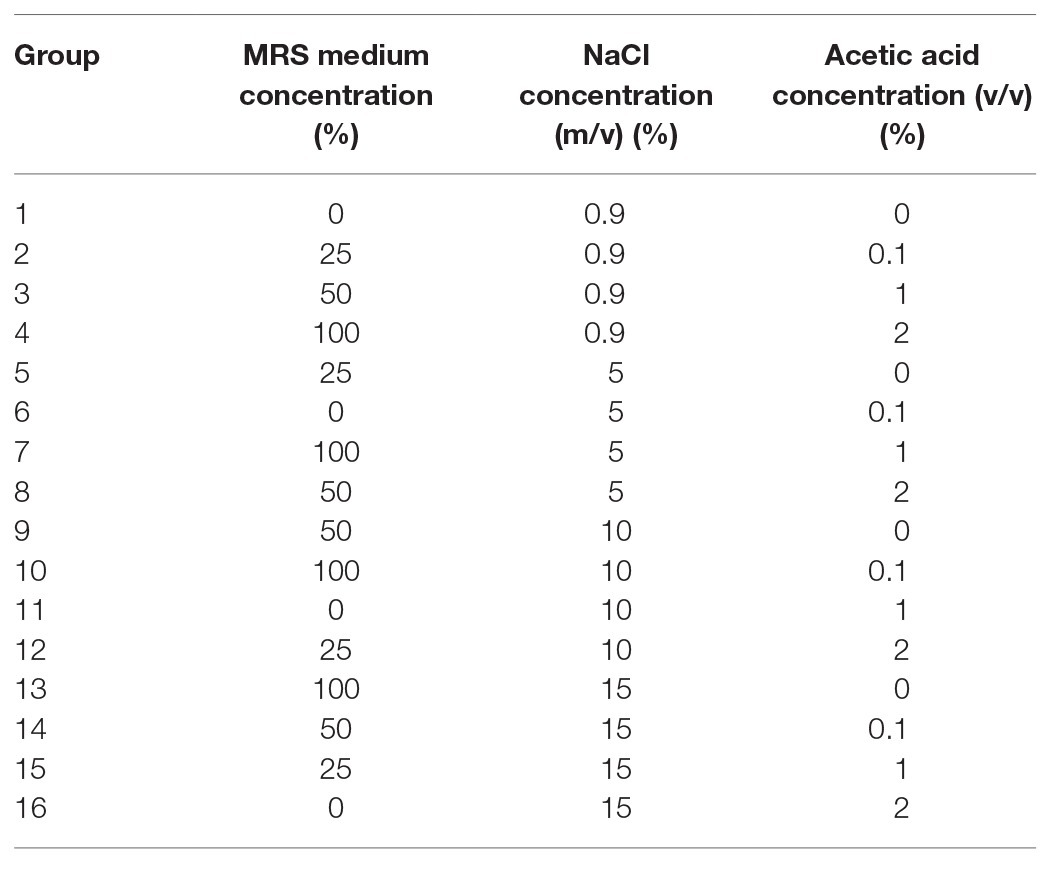
Table 1. The experimental methods of orthogonal array design of VBNC induction of P. acidilactici.
Culturability and Viability Assays
The induction culture was removed from the refrigerator and placed at room temperature. When the induction culture completely melted, it was serially diluted in 0.9% NaCl, spread on MRS agar plates, and incubated at 37°C for 24 h. When the culturable cell number of P. acidilactici bacteria in the induction culture is 1 cells/ml, it is considered that the cells may enter into the VBNC state (Deng et al., 2015). In order to determine whether P. acidilactici cells in the induction culture enter into the VBNC state, the LIVE/DEAD® BacLight™ Bacterial Viability Kit (Thermo Fisher Scientific, China) was used. The fluorescent probes SYTO 9 and PI were used with fluorescence microscope to distinguish intact from membrane-permeabilized cells. For these assays, 500 μl sample was obtained and centrifuged at 5,000 rpm for 15 min. Subsequently, the cells of P. acidilactici were washed twice with physiological saline (PBS). The supernatant was removed, and the pellet was resuspended in 500 μl of PBS. The cell suspensions were incubated with 1.5 μl of a dye mixture containing SYTO 9 and PI for 30 min at room temperature in the dark. Five microliter of cells incubated with the dye mixture was trapped between a slide and a square coverslip. The slide was placed under a fluorescent microscope for further observation. Live cells were observed under red excitation light with a wavelength of A480 nm/450 nm, and dead cells were observed under blue excitation light with a carrier length of A490 nm/635 nm. The results were then superimposed using Image J software. Under fluorescence microscope, the viable cells showed green, whereas dead cells exhibited red.
Inhibition of VBNC State Formation
According to the key environmental conditions for the formation of P. acidilactici VBNC state, the effect of acid (0.7 and 1.0%) and nutrition (0 and 25%) on the formation of P. acidilactici VBNC state were studied (Tables 2 and 3).
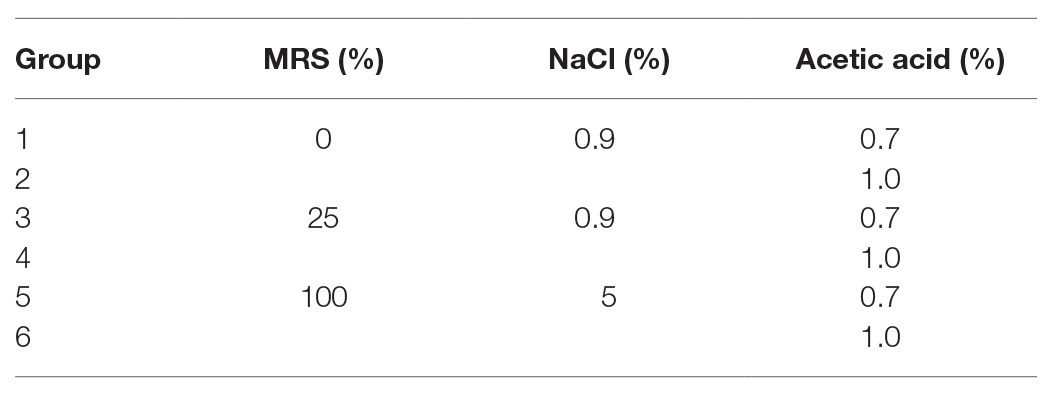
Table 2. Inhibition assay of acid on the formation of VBNC state of P. acidilactici.

Table 3. Inhibition assay of nutrition on the formation of VBNC state of P. acidilactici.
In order to investigate the inhibition of acid and nutrition change on the VBNC state formation of P. acidilactici in food system, the logarithmic phase cells were inoculated into a crystal cake food system with nutrient concentrations of 100, 50, and 25% and a volume fraction of 1.0% acetic acid. At the same time, the initial concentration of P. acidilactici in the food system was guaranteed to be 108 cells/ml and stored at 4 and −20°C, respectively. After 3 days of culture, the formation of VBNC state of P. acidilactici was observed using plate counting method and fluorescence microscopy.
Detection of VBNC State in Food Systems by PMA-PCR Assay
According to the National Food Safety standards (GB4789.3–2016) in China, 25 g of ground crystal cake was added to 225 ml of PBS and sterilized. Subsequently, P. acidilactici VBNC cells were inoculated into PBS. The initial concentrations of P. acidilactici were adjusted to be 106, 105, 104, 103, 102, and 10 cells/ml. Five-hundred microliter of bacterial suspension and PMA (final concentration is 5 μg/ml, when PMA concentration is 5 μg/ml, it can effectively distinguish VBNC state bacteria and dead bacteria) were added to a 1.5 ml centrifuge tube, and they were thoroughly mixed and left at room temperature for 10 min. Subsequently, the centrifuge tube was placed on a crushed ice box, and light treatment was performed for 15 min at a distance of 15 cm from a 650 W halogen lamp to complete the binding of PMA and DNA. The PMA molecules remaining after the treatment are passivated. The PMA-treated bacteria suspension was centrifuged at 10,000 rpm/min for 5 min, and the supernatant was discarded. DNA was then extracted using a bacterial group DNA extraction kit (Dongsheng Biotech, Guangzhou, China). The extracted DNA was detected by PCR. PCR assay was performed in a 25-μl volume and with 0.6 μM primers (pheS-F: CGCAGACAAGTCCAATGCAG; pheS-R: CACGTCGATAAACCACCCCA). The thermal profile for PCR mixtures were 95°C for 5 min, followed by 35 cycles of 95°C for 30 s, 55°C for 30 s, and 72°C for 60 s and a final extension cycle at 72°C for 5 min. The amplified products (5 μl/well) were analyzed by gel electrophoresis in 2% agarose gels and stained with ethidium bromide for 10 min. A negative control was performed using sterile water instead of culture or DNA template, and then visualized by UV transilluminator.
Results and Discussion
Effect of Nutrient, Acid, and Salt on the Formation of VBNC State
Under the conditions corresponding to the experimental groups 3, 4, 6, 8, 11, 12, 15, and 16 (Table 1), the culturable cell numbers of P. acidilactici were reduced to zero (4 and −20°C) within 3 days (Figures 1A,B,D). Under the conditions corresponding to the experimental groups 5, 9, 10, 13, and 14, the culturable cell numbers of P. acidilactici were not able to reduce to zero within 45 days (Figures 1C,E–H). The culturable cell numbers were approximately 103–106 cells/ml and 107 cells/ml at 45 days when stored at 4 and −20°C, respectively. Under the experimental conditions corresponding to the experimental groups 1, 2, and 7, P. acidilactici could enter the VBNC state within 45 days (Table 4; Figure 2). In the conditions corresponding to experimental groups 1 and 7, the VBNC state was entered at 4°C for 31 and 37 days (chart). In 45 days, the culturable cell number of P. acidilactici did not decrease to zero when stored at −20°C. Under the conditions corresponding to group 2 and under the conditions of 4 and −20°C, they entered the VBNC state at 34 and 44 days, respectively.
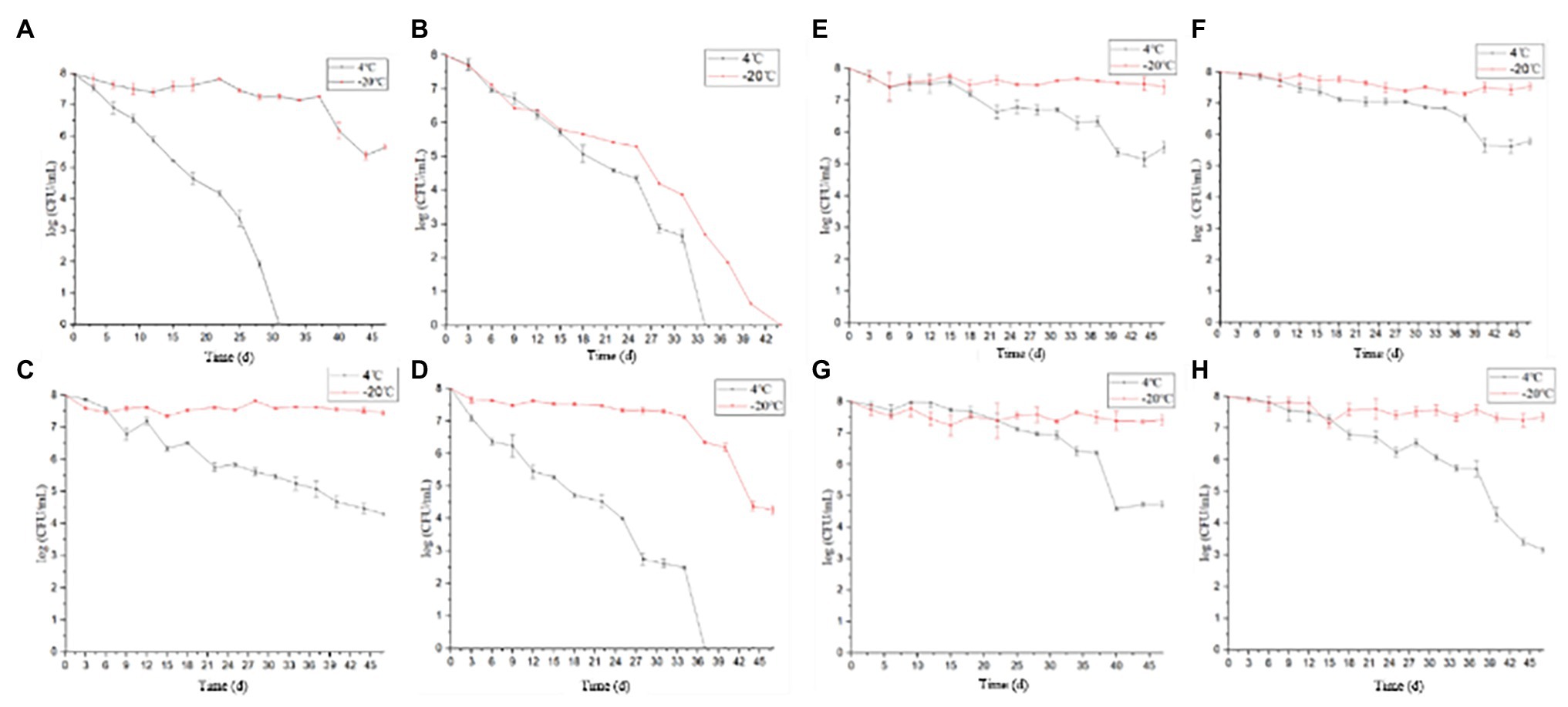
Figure 1. The culturable cell number of Pediococcus acidilactici stored under 16 different conditions (4 and −20°C). (A–H were the culturable cell number tendency of P. acidilactici inoculated in the medium configured according to the methods 1, 2, 5, 7, 9, 10, 13, and 14 and stored at 4 or −20°C, respectively).
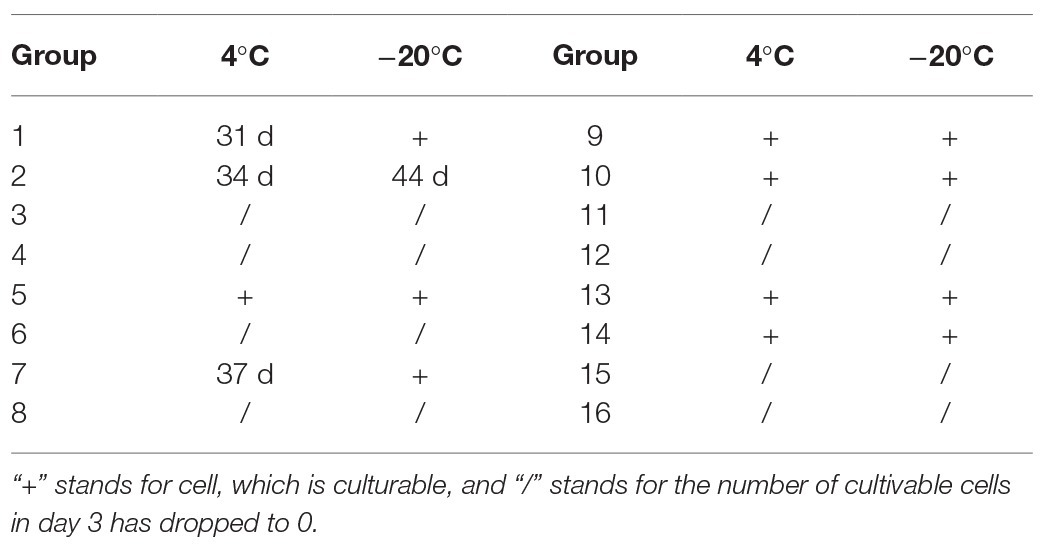
Table 4. The time of culturable cell number of P. acidilactici decreased to 0 stored at different methods.
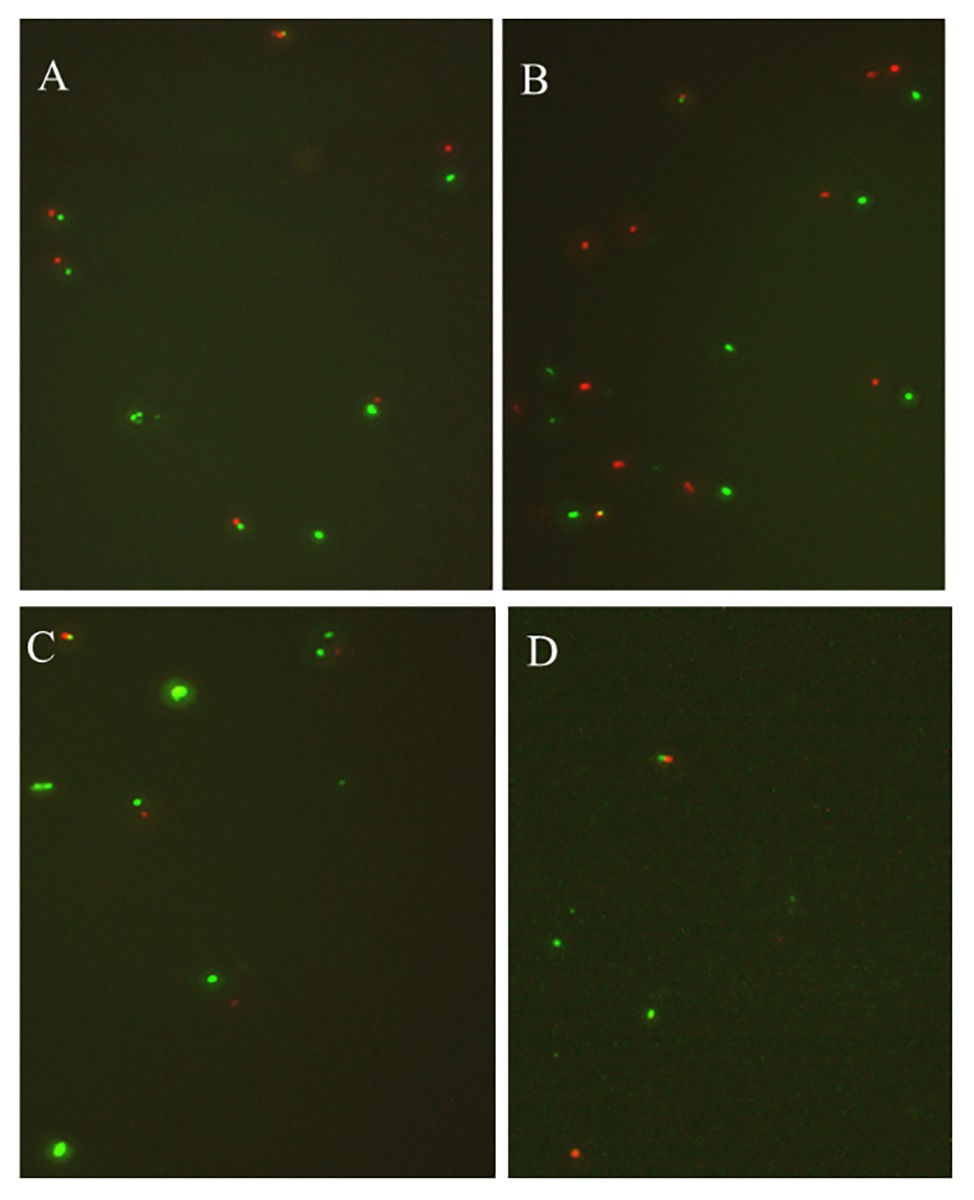
Figure 2. The viability of nonculturable P. acidilactici stored at different conditions with fluorescence microscope (A represents the storage at 4°C in group 1; B and C represent the storage at 4 and −20°C under group 2; D shows storage at 4°C in group 7).
Under the conditions in which the acetic acid concentration is 2%, the medium concentration ranges from 0 to 100%, and the salt concentration ranges from 0.9 to 15%, the culturable and viable cell numbers of P. acidilactici reduced to zero within 3 days (experimental groups 4, 8, 12, and 16). In the induction culture with an acetic acid concentration of 1% (Figure 3), P. acidilactici lost culturability and viability within 3 days when nutrition is inadequate (medium concentration ≤ 50%; experimental groups 3, 11, and 15). However, when the culture medium concentration was 100% and P. acidilactici culture was stored at 4 and −20°C, it could survive for 37 and 45 days, respectively (experimental group 7). This shows that the concentration of nutrients has a certain effect on the survival of P. acidilactici, and sufficient nutrients can enhance the resistance of P. acidilactici to external acidic substances. When the acetic acid concentration was 0.1 and 0%, the culturable cell number of experimental group 6 reduced to zero in 3 days, the culturable cell number of experimental group 2 reduced to 0 in 34 and 44 days, and the culturable cell number of experimental group 1 reduced to 0 in 31 days at 4°C. P. acidilactici in the experimental groups 5, 9, 13, 10, and 14 could survive for more than 45 days. This shows that the acetic acid concentrations used in this study have the most significant effect on the survival of P. acidilactici.

Figure 3. The culturable cell number of P. acidilactici inoculated in the 1.0% (v/v) acetic acid medium containing 100, 50, and 25% nutrients at low temperature for 3 days.
In the experimental groups 5, 9, 10, 13 and 14, with the increase of the salt concentration, the P. acidilactici cells could survive for 45 days, and the culturable cell number of P. acidilactici was not significantly different. The salt concentration used in this experiment had no significant effect on the survival of P. acidilactici. Therefore, among the environmental conditions discussed in this study, acid has the greatest impact on the survival of P. acidilactici, followed by nutrition, and salt concentration has the least effect on the survival of P. acidilactici.
Generally, a variety of environmental conditions can induce bacteria to enter the VBNC state, including the physiological cycle of bacteria (Oliver, 2010), temperature (Pawlowski et al., 2011; Liao et al., 2019), pH (Aurass et al., 2011), osmotic pressure (Aurass et al., 2011), nutritional content (Wong and Wang, 2004; Ramamurthy et al., 2014), high pressure carbon dioxide (Zhao et al., 2013), antibiotic stress (Pasquaroli et al., 2013), water activity (Gruzdev et al., 2012), UV disinfection (Xu et al., 2009; Zhang et al., 2015; Liu et al., 2018b), and antibiotic (Lee and Bae, 2018) can induce bacteria to enter the VBNC state. This study used medium concentration (0, 25, 50, and 100%), sodium chloride concentration (0.9, 5, 10, and 15%), and acetic acid concentration (0, 0.1, 1, and 2%) as single factors to design an orthogonal experiment to explore the ability of environmental conditions to form P. acidilactici VBNC. Among the 16 induction cultures, only in three induction cultures successfully induced P. acidilactici entering into the VBNC state within 45 days. (1) The medium concentration is 0, the sodium chloride mass fraction is 0.9%, and the acetic acid volume fraction is 0. (2) The concentration of the medium is 25%, the mass fraction of sodium chloride is 0.9%, and the volume fraction of acetic acid is 0.1%. (3) Medium concentration is 100%, sodium chloride mass fraction is 5%, and acetic acid volume fraction is 1%. This shows that when the concentration of the medium is less than 25%, the concentration of sodium chloride is 0.9%, and the concentration of acetic acid is less than 0.1%, it is beneficial to the formation of VBNC state of Escherichia coli. That is, nutritional deficiencies and relatively low concentrations of sodium chloride and acetic acid are benificial to VBNC state formation. Under this environmental pressure, P. acidilactici does not die directly, but enters the VBNC state. In addition, when the concentration of the culture medium is 100%, the concentration of sodium chloride is 5%, and the volume fraction of acetic acid is 1%, P. acidilactici can enter the VBNC state. This shows that under the condition of sufficient nutrition, P. acidilactici can resist higher sodium chloride concentration and acid (Xu et al., 2011b, 2012b).
Effect of Temperature on the Formation of VBNC State
In addition to the influence of the composition of the induction culture, temperature also affects the entry of P. acidilactici into VBNC (Table 4). Among the experimental conditions discussed in this study, only the conditions corresponding to the three experimental groups can induce P. acidilactici to enter the VBNC state. In experimental groups 1 and 7, P. acidilactici entered into the VBNC state at 4°C in 31 and 37 days, respectively, but P. acidilactici was still active in 45 days at −20°C. In experimental group 2, P. acidilactici entered into VBNC state after 34 and 44 days, respectively. This shows that −20°C affected more on the survival of P. acidilactici.
Low temperature is the main factor that induces bacteria to enter the VBNC state. Food materials and finished products are mostly kept at low temperatures. During the low temperature storage process, the bacteria are easily affected by low temperature and enter the VBNC state. Food pass the test may contain VBNC state bacteria, which will become an invisible source of contamination and affect food safety (Xu et al., 2010, 2011c; Wang et al., 2011).
Effects of Acid and Nutrition on the Survival of P. acidilactici
Under the condition that the medium concentration is 0 (groups 1 and 2), by adding a certain amount of acetic acid (acetic acid volume fraction reaches 0.7 or 1.0%), the culturability of P. acidilactici was 0 after 3 days (Table 5). And bacterial activity test results showed that all bacteria died and could not enter into the VBNC state. When the nutrient concentration was 25 and 100%, and the acetic acid concentration was 7% (in groups 3 and 5), P. acidilactici was culturable at 3 days, but the culturable cell number was significantly reduced, indicating that P. acidilactici may enter into VBNC state. When the acetic acid concentration was 1.0%, P. acidilactici was culturable at 4 and −20°C for 3 days. When the nutrient concentration is 100%, P. acidilactici was culturable and had the potential to enter into the VBNC state (experimental group 6). However, when the nutrient concentration was 0 and 25%, P. acidilactici was not culturable and viable (experimental groups 2 and 4; Figure 4, Table 6). When the acetic acid concentration was 1.0% and the salt concentration was 5%, the nutrient content in the induction culture was reduced to 25% or less. After 3 days, the number of cultivable P. acidilactici and the bacterial activity were 0. During the processing of rice flour products, it is possible to control the formation of lactic acid bacteria VBNC state by minimizing the residual nutrients on the surface of the processing equipment and using acetic acid with a volume fraction of 1.0%.
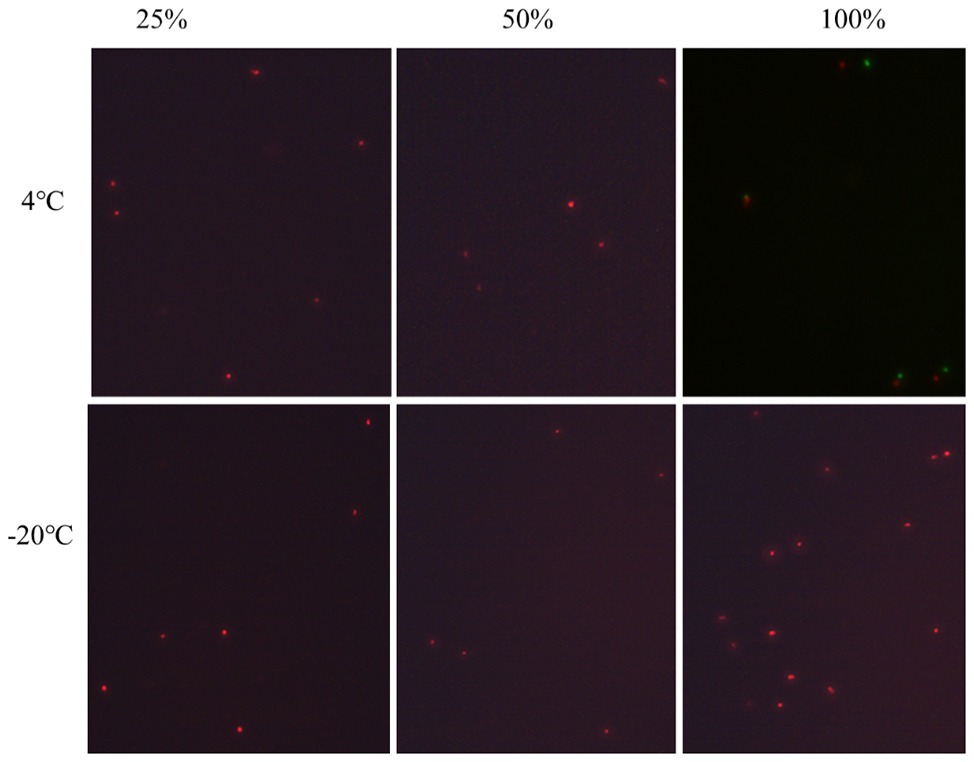
Figure 4. The viability of nonculturable P. acidilactici stored at different conditions with fluorescence microscope.
Elimination of VBNC State in Rice Flour Products
Crystal cake is a traditional Chinese snack, which has the characteristics of rich nutrition. The rich nutrients and sufficient water in crystal cake make it a natural medium for the growth of various pathogenic and spoilage bacteria. In this study, crystal cake was applied as a representative food system to study the inhibition and detection of the VBNC state of P. acidilactici under food processing and storage conditions. P. acidilactici was inoculated in food system with 25, 50, and 100% nutrient content and cultured at 4 and −20°C, respectively. After 3 days of culture, the culturability and activity of P. acidilactici were measured. Only when the nutrient content is 100% and stored at 4°C, P. acidilactici is not culturable and viable, but can survive in other experimental groups. Therefore, adding 1% acetic acid to rice flour products and storing them at −20°C can prevent P. acidilactici in rice flour products from entering the VBNC state and avoid potential threats caused by the presence of P. acidilactici in the VBNC state.
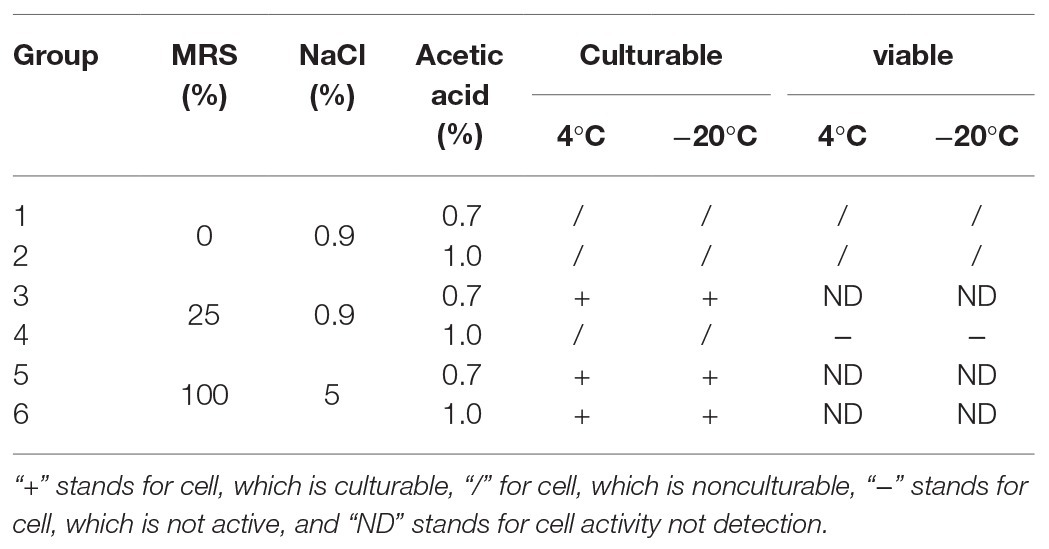
Table 5. Inhibition of acid on the formation of VBNC state of P. acidilactici.
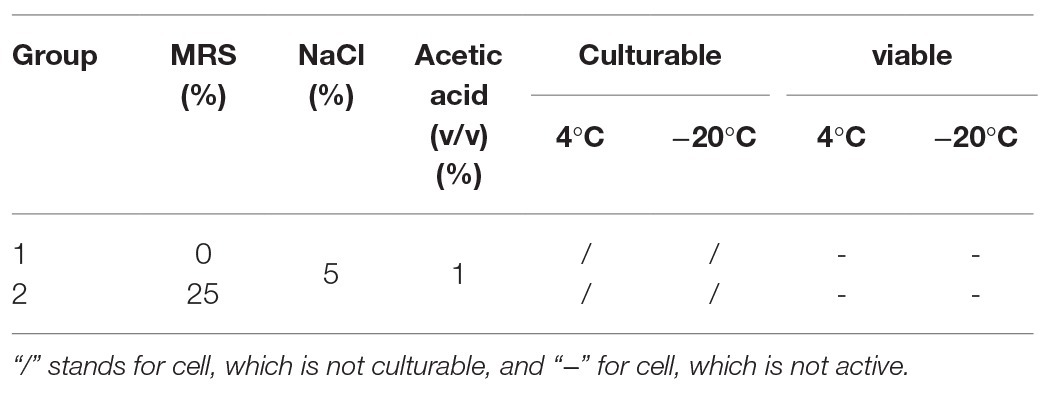
Table 6. Inhibition of nutrition on the formation of VBNC state of P. acidilactici.
Detection of VBNC State of P. acidilactici
Propidium monoazide is a DNA intercalating molecule that can differentiate between live and dead or membrane-damaged bacteria. It can selectively penetrate the damaged cells and form a stable covalent high affinity bonds with DNA, following photo-activation exposure to strong visible light (Andreas et al., 2009). The DNA-PMA bond inhibits PCR amplification of the DNA strands of dead bacteria. It can be known from the results that when the concentration of P. acidilactici in the VBNC state in the crystal cake food system is higher than 104 cells/ml, the P. acidilactici in the VBNC state can be successfully detected using the PMA-PCR technology (Figure 5).
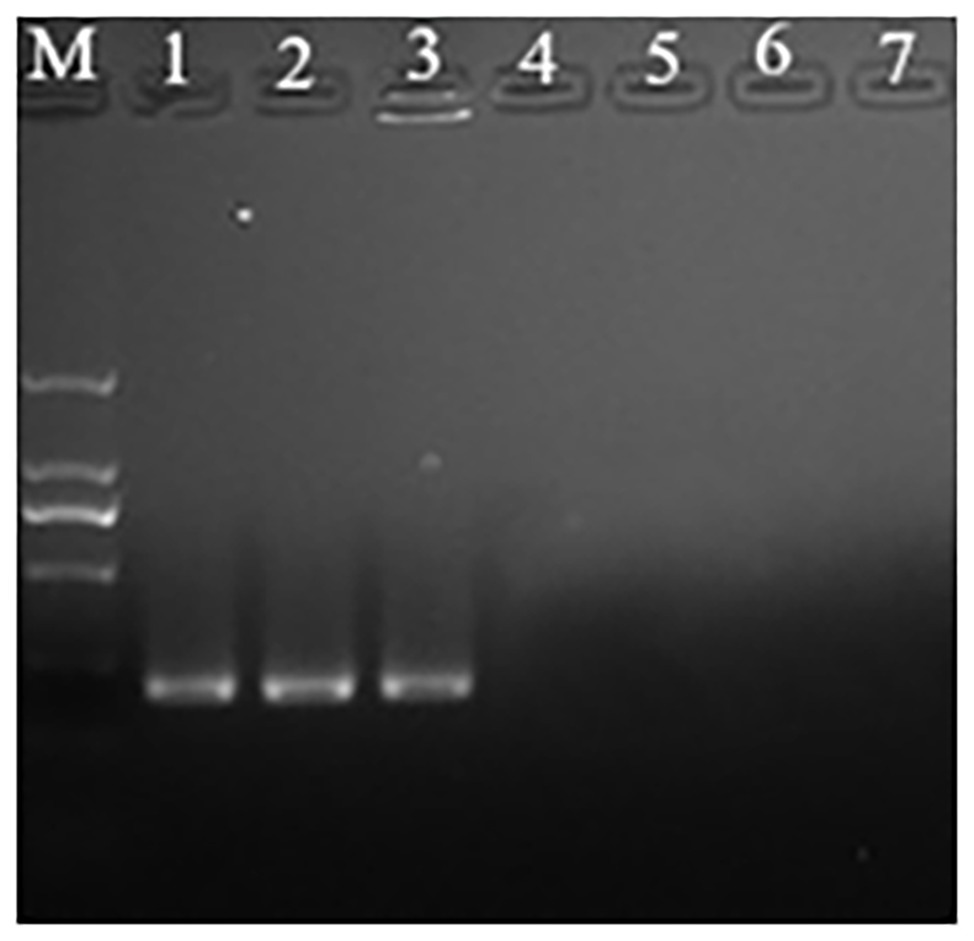
Figure 5. The propidium monoazide-polymerase chain reaction (PMA-PCR) in detection of the viable but nonculturable (VBNC) state of P. acidilactici in crystal cake (Lane M, DNA marker; Lanes 1–6 represent crystal cake with P. acidilactici concentrations of 106, 105, 104, 103, 102, and 10 cells/ml, respectively; Lane 7: negative control).
Conclusion
Among the environmental conditions discussed in this study, acid had the greatest effect on the formation of VBNC state of P. acidilactici, followed by nutrition, and the effect of salt concentration on the survival of P. acidilactici was the least significant. In addition to the influence of the composition of the induction culture, 4°C is more favorable for P. acidilactici to enter into the VBNC state. By reducing the amount of nutrients in the environment and treating with 1.0% acetic acid, the formation of VBNC state can be suppressed. The addition of 1% acetic acid in rice flour products and storage at −20°C can inhibit P. acidilactici in rice flour products from entering into the VBNC state and avoid potential threats caused by the presence of P. acidilactici in the VBNC state. Also, PMA-PCR method can be applied to detect VBNC P. acidilactici cells with concentration higher than 104 cells/ml.
Data Availability Statement
All datasets presented in this study are included in the article.
Author Contributions
JL and KW conceived of the study and participated in its design and coordination. T-YH, YM, YC, FS, RP, and JC performed the experimental work. CB and LC analyzed the data. JL prepared and revised this manuscript. All authors reviewed and approved the final manuscript.
Funding
This work was supported by the National Key R&D Program of China (2016YFD0400203), YangFan Innovative and Entrepreneurial Research Team Project (2014YT02S029), the National Key Research and Development Program of China (2016YFD04012021), the Fundamental Research Funds for the Central Universities (D2191310), Collaborative grant with AEIC (KEO-2019-0624-001-1), and the 111 Project (B17018).
Conflict of Interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
References
Andreas, N., Mazz, A., Masson, L. K., Camper, A. K., and Roland, B. (2009). Selective detection of live bacteria combining propidium monoazide sample treatment with microarray technology. J. Microbiol. Methods 76, 253–261. doi: 10.1016/j.mimet.2008.11.004
Aurass, P., Prager, R., and Flieger, A. (2011). EHEC/EAEC O104:H4 strain linked with the 2011 German outbreak of haemolytic uremic syndrome enters into the viable but non-culturable state in response to various stresses and resuscitates upon stress relief. Environ. Microbiol. 13, 3139–3148. doi: 10.1111/j.1462-2920.2011.02604.x
Bao, X. R., Jia, X. Y., Chen, L. Q., Peters, B. M., Lin, C. -W., Chen, D. Q., et al. (2017c). Effect of polymyxin resistance (pmr) on biofilm formation of Cronobacter sakazakii. Microb. Pathog. 106, 16–19. doi: 10.1016/j.micpath.2016.12.012
Bao, X. R., Yang, L., Chen, L. Q., Li, B., Li, L., Li, Y. Y., et al. (2017a). Virulent and pathogenic features on the Cronobacter sakazakii polymyxin resistant pmr mutant strain s-3. Microb. Pathog. 110, 359–364. doi: 10.1016/j.micpath.2017.07.022
Bao, X. R., Yang, L., Chen, L. Q., Li, B., Li, L., Li, Y. Y., et al. (2017b). Analysis on pathogenic and virulent characteristics of the Cronobacter sakazakii strain BAA-894 by whole genome sequencing and its demonstration in basic biology science. Microb. Pathog. 109, 280–286. doi: 10.1016/j.micpath.2017.05.030
Barros, R. R., Carvalho, G. S., Peralta, J. M., Facklam, R. R., and Teixeira, L. M. (2001). Phenotypic and genotypic characterization of Pediococcus strains isolated from human clinical sources. J. Clin. Microbiol. 39, 1241–1246. doi: 10.1128/JCM.39.4.1241-1246.2001
Deng, Y., Liu, J., Li, L., Fang, H., Tu, J., Li, B., et al. (2015). Reduction and restoration of culturability of beer-stressed and low-temperature-stressed Lactobacillus acetotolerans strain 2011-8. Int. J. Food Microbiol. 206, 96–101. doi: 10.1016/j.ijfoodmicro.2015.04.046
Du, M., Chen, J., Zhang, X., Li, A., Li, Y., and Wang, Y. (2007). Retention of virulence in a viable but nonculturable Edwardsiella tarda isolate. Appl. Environ. Microbiol. 73, 1349–1354. doi: 10.1128/AEM.02243-06
Gruzdev, N., Pinto, R., and Saldinger, S. (2012). Persistence of Salmonella enterica during dehydration and subsequent cold storage. Food Microbiol. 32, 415–422. doi: 10.1016/j.fm.2012.08.003
Hu, B., Pan, Y., Li, Z., Yuan, W., and Deng, L. (2019). EmPis-1L, an effective antimicrobial peptide against the antibiotic-resistant VBNC state cells of pathogenic bacteria. Probiotics Antimicro. Proteins 11, 667–675. doi: 10.1007/s12602-018-9446-3
Jia, X. Y., Hua, J. J., Liu, L., Xu, Z. B., and Li, Y. Y. (2018). Phenotypic characterization of pathogenic Cronobacter spp. strains. Microb. Pathog. 121, 232–237. doi: 10.1016/j.micpath.2018.05.033
Klaenhammer, T. R. (1993). Genetics of bacteriocins produced by lactic acid bacteria. FEMS Microbiol. Rev. 12, 39–85. doi: 10.1111/j.1574-6976.1993.tb00012.x
Lee, S., and Bae, S. (2018). Molecular viability testing of viable but non-culturable bacteria induced by antibiotic exposure. Microb. Biotechnol. 11, 1008–1016. doi: 10.1111/1751-7915.13039
Liao, H., Xiao, W., Xu, L., Ma, Q., Wang, Y., Cai, Y., et al. (2019). Quorum-sensing systems trigger catalase expression to reverse the oxyR deletion-mediated VBNC state in Salmonella typhimurium. Res. Microbiol. 170, 65–73. doi: 10.1016/j.resmic.2018.10.004
Lin, S. Q., Li, L., Li, B., Zhang, X. H., Lin, C. W., Deng, Y., et al. (2016). Development and evaluation of quantitative detection of N-epsilon-carboxymethyl-lysine in Staphylococcus aureus biofilm by LC-MS method. Basic Clin. Pharmacol. Toxicol. 118:33.
Lin, S. Q., Yang, L., Chen, G., Li, B., Chen, D. Q., Li, L., et al. (2017). Pathogenic features and characteristics of food borne pathogens biofilm: biomass, viability and matrix. Microb. Pathog. 111, 285–291. doi: 10.1016/j.micpath.2017.08.005
Liu, J., Deng, Y., Soteyome, T., Li, Y., Su, J., Li, L., et al. (2018c). Induction and recovery of the viable but Nonculturable state of hop-resistance lactobacillus brevis. Front. Microbiol. 9:2076. doi: 10.3389/fmicb.2018.02076
Liu, J., Li, L., Li, B., Peters, B. M., Deng, Y., Xu, Z., et al. (2017a). Study on spoilage capability and VBNC state formation and recovery of lactobacillus plantarum. Microb. Pathog. 110, 257–261. doi: 10.1016/j.micpath.2017.06.044
Liu, J. Y., Li, L., Li, B., Peters, B. M., Deng, Y., Xu, Z., et al. (2017b). First study on the formation and resuscitation of viable but nonculturable state and beer spoilage capability of lactobacillus lindneri. Microb. Pathog. 107, 219–224. doi: 10.1016/j.micpath.2017.03.043
Liu, L. Y., Lu, Z. R., Li, L., Li, B., Zhang, X., Zhang, X. M., et al. (2018a). Physical relation and mechanism of ultrasonic bactericidal activity on pathogenic E. coli with WPI. Microb. Pathog. 117, 73–79. doi: 10.1016/j.micpath.2018.02.007
Liu, L. Y., Xu, R. R., Li, L., Li, B., Zhang, X., Zhang, X. M., et al. (2018b). Correlation and in vitro mechanism of bactericidal activity on E. coli with whey protein isolate during ultrasonic treatment. Microb. Pathog. 115, 154–158. doi: 10.1016/j.micpath.2017.12.062
Liu, L., Ye, C., Soteyome, T., Zhao, X., and Harro, J. M. (2019). Inhibitory effects of two types of food additives on biofilm formation by foodborne pathogens. Microbiologyopen 8:e00853. doi: 10.1002/mbo3.853
Miao, J., Chen, L. Q., Wang, J. W., Wang, W. X., Chen, D. Q., Li, L., et al. (2017a). Evaluation and application of molecular genotyping on nosocomial pathogen-methicillin-resistant Staphylococcus aureus isolates in Guangzhou representative of southern China. Microb. Pathog. 107, 397–403. doi: 10.1016/j.micpath.2017.04.016
Miao, J., Chen, L. Q., Wang, J. W., Wang, W. X., Chen, D. Q., Li, L., et al. (2017c). Current methodologies on genotyping for nosocomial pathogen methicillin-resistant Staphylococcus aureus (MRSA). Microb. Pathog. 107, 17–28. doi: 10.1016/j.micpath.2017.03.010
Miao, J., Liang, Y. R., Chen, L. Q., Wang, W. X., Wang, J. W., Li, B., et al. (2017b). Formation and development of Staphylococcus biofilm: with focus on food safety. J. Food Saf. 7:e12358. doi: 10.1111/jfs.12358
Miao, J., Peters, B. M., Li, L., Li, B., Zhao, X. H., Xu, Z. B., et al. (2016). Evaluation of ERIC-PCR for fingerprinting methicillin-resistant Staphylococcus aureus strains. Basic Clin. Pharmacol. Toxicol. 118:33.
Miao, J., Wang, W. X., Xu, W. Y., Su, J. Y., Li, L., Li, B., et al. (2018). The fingerprint mapping and genotyping systems application on methicillin-resistant Staphylococcus aureus. Microb. Pathog. 125, 246–251. doi: 10.1016/j.micpath.2018.09.031
Oliver, J. D. (2010). Recent findings on the viable but nonculturable state in pathogenic bacteria. FEMS Microbiol. Rev. 34, 415–425. doi: 10.1111/j.1574-6976.2009.00200.x
Olszewska, M. A., and Bialobrzewski, I. (2019). Mixed species biofilms of lactobacillus plantarum and Listeria innocua show facilitated entrance to the VBNC state during chlorine-induced stress. J. Food Saf. 39:e12651. doi: 10.1111/jfs.12651
Pasquaroli, S., Zandri, G., Vignaroli, C., Vuotto, C., Donelli, G., and Biavasco, F. (2013). Antibiotic pressure can induce the viable but non-culturable state in Staphylococcus aureus growing in biofilms. J. Antimicrob. Chemother. 68, 1812–1817. doi: 10.1093/jac/dkt086
Pawlowski, D. R., Metzger, D. J., Raslawsky, A., Howlett, A., Siebert, G., Karalus, R. J., et al. (2011). Entry of Yersinia pestis into the viable but nonculturable state in a low-temperature tap water microcosm. PLoS One 6:e17585. doi: 10.1371/journal.pone.0017585
Piao, M., Li, Y., Wang, Y., Wang, F., Zhen, T., and Deng, Y. (2019). Induction of viable but putatively non-culturable lactobacillus acetotolerans by thermosonication and its characteristics. LWT-Food Sci. Technol. 109, 313–318. doi: 10.1016/j.lwt.2019.04.046
Ramamurthy, T., Amit, G., Gururaj, P. P., and Sumio, S. (2014). Current perspectives on viable but non-culturable (VBNC) pathogenic bacteria. Front. Public Health 2:103. doi: 10.3389/fpubh.2014.00103
Wang, L., Zhao, X. H., Chu, J., Li, Y., Li, Y. Y., Li, C. H., et al. (2011). Application of an improved loop-mediated isothermal amplification detection of Vibrio parahaemolyticus from various seafood samples. Afr. J. Microbiol. Res. 5, 5765–5771. doi: 10.5897/AJMR11.1237
Wen, S. X., Feng, D. H., Chen, D. Q., Yang, L., and Xu, Z. B. (2020). Molecular epidemiology and evolution of Haemophilus influenzae. Infect. Genet. Evol. 80:104205. doi: 10.1016/j.meegid.2020.104205
Wong, H. C., and Wang, P. (2004). Induction of viable but nonculturable state in Vibrio parahaemolyticus and its susceptibility to environmental stresses. J. Appl. Microbiol. 96, 359–366. doi: 10.1046/j.1365-2672.2004.02166.x
Xie, J. H., Peters, B. M., Chen, L. Q., Chen, D. Q., Li, B., Li, L., et al. (2017a). A 16-year retrospective surveillance report on the pathogenic features and antimicrobial susceptibility of Pseudomonas aeruginosa isolated from Guangzhou representative of southern China. Microb. Pathog. 110, 37–41. doi: 10.1016/j.micpath.2017.06.018
Xie, J. H., Peters, B. M., Li, B., Li, L., Yu, G. C., Xu, Z. B., et al. (2017b). Clinical features and antimicrobial resistance profiles of important Enterobacteriaceae pathogens in Guangzhou representative of southern China, 2001-2015. Microb. Pathog. 107, 206–211. doi: 10.1016/j.micpath.2017.03.038
Xu, Z. B., Gui, Z. Y., Li, L., Li, B., Su, J. Y., Zhao, X. H., et al. (2012b). Expression and purification of gp41-gp36 fusion protein and application in serological screening assay of HIV-1 and HIV-2. Afr. J. Microbiol. Res. 6, 6295–6299. doi: 10.5897/AJMR12.1075
Xu, Z. B., Hou, Y. C., Peters, B. M., Chen, D. Q., Li, B., and Li, L. (2016b). Chromogenic media for MRSA diagnostics. Mol. Biol. Rep. 43, 1205–1212. doi: 10.1007/s11033-016-4062-3
Xu, Z. B., Hou, Y. C., Qin, D., Liu, X. C., Li, B., Li, L., et al. (2016a). Evaluation of current methodologies for rapid identification of methicillin-resistant Staphylococcus aureus strains. Basic Clin. Pharmacol. Toxicol. 118:33.
Xu, Z. B., Li, L., Alam, M. J., Zhang, L. Y., Yamasaki, S., and Shi, L. (2008b). First confirmation of integron-bearing methicillin-resistant Staphylococcus aureus. Curr. Microbiol. 2008, 264–268. doi: 10.1007/s00284-008-9187-8
Xu, Z. B., Li, L., Chu, J., Peters, B. M., Harris, M. L., Li, B., et al. (2012a). Development and application of loop-mediated isothermal amplification assays on rapid detection of various types of staphylococci strains. Food Res. Int. 47, 166–173. doi: 10.1016/j.foodres.2011.04.042
Xu, Z. B., Li, L., Shi, L., and Shirliff, M. E. (2011c). Class 1 integron in staphylococci. Mol. Biol. Rep. 38, 5261–5279. doi: 10.1007/s11033-011-0676-7
Xu, Z. B., Li, L., Shirliff, M. E., Peters, B. M., Li, B., Peng, Y., et al. (2011b). Resistance class 1 integron in clinical methicillin-resistant Staphylococcus aureus strains in southern China, 2001-2006. Clin. Microbiol. Infect. 17, 714–718. doi: 10.1111/j.1469-0691.2010.03379.x
Xu, Z. B., Li, L., Shirtliff, M. E., Alam, M. J., Yamasaki, S., and Shi, L. (2009). Occurrence and characteristics of class 1 and 2 integrons in Pseudomonas aeruginosa isolates from patients in southern China. J. Clin. Microbiol. 47, 230–234. doi: 10.1128/JCM.02027-08
Xu, Z. B., Li, L., Shirtliff, M. E., Peters, B. M., Peng, Y., Alam, M. J., et al. (2010). First report of class 2 integron in clinical enterococcus faecalis and class 1 integron in Enterococcus faecium in South China. Diagn. Microbiol. Infect. Dis. 68, 315–317. doi: 10.1016/j.diagmicrobio.2010.05.014
Xu, Z. B., Li, L., Zhao, X. H., Chu, J., Li, B., Shi, L., et al. (2011a). Development and application of a novel multiplex polymerase chain reaction (PCR) assay for rapid detection of various types of staphylococci strains. Afr. J. Microbiol. Res. 2011, 1869–1873. doi: 10.5897/AJMR11.437
Xu, Z. B., Liang, Y. R., Lin, S. Q., Chen, D. Q., Li, B., Li, L., et al. (2016c). Crystal violet and XTT assays on Staphylococcus aureus biofilm quantification. Curr. Microbiol. 73, 474–482. doi: 10.1007/s00284-016-1081-1
Xu, Z. B., Shi, L., Alam, M. J., Li, L., and Yamasaki, S. (2008a). Integron-bearing methicillin-resistant coagulase-negative staphylococci in South China, 2001-2004. FEMS Microbiol. Lett. 278, 223–230. doi: 10.1111/j.1574-6968.2007.00994.x
Xu, Z. B., Shi, L., Zhang, C., Zhang, L. Y., Li, X. H., Cao, Y. C., et al. (2007). Nosocomial infection caused by class 1 integron-carrying Staphylococcus aureus in a hospital in South China. Clin. Microbiol. Infect. 13, 980–984. doi: 10.1111/j.1469-0691.2007.01782.x
Xu, Z. B., Xie, J. H., Peters, B. M., Li, B., Li, L., Yu, G. C., et al. (2017b). Longitudinal surveillance on antibiogram of important gram-positive pathogens in southern China, 2001 to 2015. Microb. Pathog. 103, 80–86. doi: 10.1016/j.micpath.2016.11.013
Xu, Z. B., Xie, J. H., Yang, L., Chen, D. Q., Peters, B. M., and Shirtliff, M. E. (2018). Complete sequence of pCY-CTX, a plasmid carrying a phage-like region and ISEcp1-mediated Tn2 element from Enterobacter cloacae. Microb. Drug Resist. 24, 307–313. doi: 10.1089/mdr.2017.0146
Xu, Z. B., Xu, X. Y., Yang, L., Li, B., Li, L., Li, X. X., et al. (2017a). Effect of aminoglycosides on the pathogenic characteristics of microbiology. Microb. Pathog. 113, 357–364. doi: 10.1016/j.micpath.2017.08.053
You, R., Gui, Z. Y., Xu, Z. B., Shirtliff, M. E., Yu, G. C., Zhao, X. H., et al. (2012). Methicillin-resistance Staphylococcus aureus detection by an improved rapid PCR assay. Afr. J. Microbiol. Res. 6, 7131–7133.
Zhang, S., Ye, C., Lin, H., Lv, L., and Yu, X. (2015). UV disinfection induces a Vbnc state in Escherichia coli and Pseudomonas aeruginosa. Environ. Sci. Technol. 49, 1721–1728. doi: 10.1021/es505211e
Zhao, F., Bi, X., Hao, Y., and Liao, X. (2013). Induction of viable but nonculturable Escherichia coli O157: H7 by high pressure CO2 and its characteristics. PLoS One 8:e62388. doi: 10.1371/journal.pone.0062388
Zhao, X. H., Li, M., and Xu, Z. B. (2018b). Detection of foodborne pathogens by surface enhanced Raman spectroscopy. Front. Microbiol. 9:1236. doi: 10.3389/fmicb.2018.01236
Zhao, X. H., Yu, Z. X., and Xu, Z. B. (2018a). Study the features of 57 confirmed CRISPR loci in 38 strains of Staphylococcus aureus. Front. Microbiol. 9:1591. doi: 10.3389/fmicb.2018.01591
Keywords: Pediococcus acidilactici, viable but nonculturable state, environmental conditions, food system, propidium monoazide-polymerase chain reaction
Citation: Li Y, Huang T-Y, Mao Y, Chen Y, Shi F, Peng R, Chen J, Bai C, Chen L, Wang K and Liu J (2020) Effect of Environmental Conditions on the Formation of the Viable but Nonculturable State of Pediococcus acidilactici BM-PA17927 and Its Control and Detection in Food System. Front. Microbiol. 11:586777. doi: 10.3389/fmicb.2020.586777
Edited by:
Xihong Zhao, Wuhan Institute of Technology, ChinaReviewed by:
Wensen Jiang, Cedars Sinai Medical Center, United StatesSiyuan Liu, University of Saskatchewan, Canada
Copyright © 2020 Li, Huang, Mao, Chen, Shi, Peng, Chen, Bai, Chen, Wang and Liu. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Junyan Liu, jliu81@uthsc.edu; Kan Wang, 120384746@qq.com
†These authors have contributed equally to this work