 Lijun Wang1
Lijun Wang1 Yang Jiao1
Yang Jiao1 Aonan Zhao1
Aonan Zhao1 Xiaomeng Xu1Guanyu Ye1Yichi Zhang1
Xiaomeng Xu1Guanyu Ye1Yichi Zhang1 Ying Wang1
Ying Wang1 Yulei Deng1
Yulei Deng1 Wei Xu1*
Wei Xu1* Jun Liu1,2*
Jun Liu1,2*- 1Department of Neurology and Institute of Neurology, Ruijin Hospital, Shanghai Jiao Tong University School of Medicine, Shanghai, China
- 2CAS Center for Excellence in Brain Science and Intelligence Technology, Ruijin Hospital, Shanghai Jiao Tong University School of Medicine, Shanghai, China
Objective: The study aimed to clarify the association of the 21 single nucleotide polymorphisms (SNPs) with Alzheimer’s disease (AD) in the population of southern China.
Methods: A case-control study was conducted with a total sample size of 490 subjects (246 patients with AD and 244 age- and gender-matched healthy controls) enrolled in this study. Twenty-one selected SNPs were detected using SNaPshot assay and polymerase chain reaction (PCR) technique. Then, we assessed how these SNPs correlated with AD susceptibility.
Results: The results showed that rs3764650 of ABCA7 was closely correlated with risen AD morbidity in the allele [P = 0.010, odds ratio (OR) = 1.43, 95% confidence interval (CI) 1.09–1.89], dominant (P = 0.004, OR = 1.71, 95% CI 1.19–2.46), and additive (P = 0.012, OR = 1.42, 95% CI 1.08–1.86) models. However, rs4147929 of ABCA7 was related to higher AD risk in the allele (P = 0.006, OR = 1.45, 95% CI 1.11–1.89), dominant (P = 0.012, OR = 1.59, 95% CI 1.11–2.27), and additive (P = 0.010, OR = 1.40, 95% CI 1.08–1.81) models. In addition, the frequencies of the G-allele at rs3764650 (P = 0.030) and the A-allele at rs4147929 (P = 0.001) in AD were statistically higher in APOE ε4 carriers in comparison to non-carriers.
Conclusion: This study demonstrated that the G-allele at rs3764650 and the A-allele at rs4147929 appeared at higher risk for developing AD, particularly in APOE ε4 carriers. Moreover, it was observed that rs3764650 and rs4147929 of ABCA7 were linked to AD. More in-depth research with a relatively large sample is needed to make the results more convincing.
Introduction
Alzheimer’s disease (AD), an eye-catching neurodegenerative disorder in the elderly, has an increased burden dramatically on the global economic development and health care systems (Alzheimer’s Association, 2021). It is characterized by progressive deterioration of cognitive function and function impairment resulting from extracellular deposition of β-amyloid (Aβ) plaques and neuronal accumulation of neurofibrillary tangles formed by hyperphosphorylated tau protein and brain atrophy (Blennow et al., 2006; De Strooper and Karran, 2016; DeTure and Dickson, 2019; Andrews et al., 2020). Genetic factors play a crucial role in the pathogenesis of AD (Jansen et al., 2019; Kunkle et al., 2019). The most strongly and consistently associated with AD risk gene is apolipoprotein E (APOE) (Liu et al., 2013; Yamazaki et al., 2019; Serrano-Pozo et al., 2021). Previous genome-wide association studies (GWASs) have found and defined up to 20 AD susceptibility loci, including ABCA7, BIN1, CLU, CR1, PICALM, SORL1, and so on (Harold et al., 2009; Lambert et al., 2009, 2013; Steinberg et al., 2015; Vardarajan et al., 2015; Giri et al., 2016; Kunkle et al., 2019). Recently, studies have reported that GRN, TMEM106B, Complement C7, RBFOX1 genes are associated with AD in various cohorts (Viswanathan et al., 2009; Lee et al., 2011; Rutherford et al., 2012; Lu et al., 2014; Jun et al., 2016; Xu et al., 2017; Kunkle et al., 2019; Hu et al., 2021). In addition, hippocampal sclerosis of aging (HS-Aging) is a common, high morbidity brain disorder that occurs in the elderly with a clinical course similar to AD. ABCC9 and KCNMB2 have previously been shown to be associated with HS-Aging (Nelson et al., 2014; Nho and Saykin, 2016; Katsumata et al., 2017; Dugan et al., 2021). However, most of these studies were performed statistically limited in Caucasian populations, and the inheritance of AD in other populations is relatively limited. Repeating the GWAS results in different ethnic groups can help identify SNPs that are associated with AD (Chanock et al., 2007). However, the results in other populations of European descent were inconsistent with those in the southern Chinese population. Therefore, how these candidate loci and AD in the southern Chinese population are related was still not clear. Consequently, in the present study, 21 SNPs were selected from the above studies to investigate how genes affected the AD morbidity of the southern Chinese.
Materials and Methods
Study Design and Participants
A sum of 246 patients with AD (137 females and 109 males, average age ± SD: 71.26 ± 8.46 years) were collected from September 2016 to September 2020 from the neurology outpatient clinic at Ruijin Hospital affiliated with Shanghai Jiao Tong University School of Medicine. Patients with AD dementia fulfilled the National Institute of Neurological and Communicative Disorders and Stroke–Alzheimer’s Disease and Related Disorders Association (NINCDS-ADRDA) criteria for probable AD (Dubois et al., 2007). Global cognitive abilities were measured on a scale of 0 (severely impaired) to 30 points (no impaired) using the Mini-mental Status Evaluation (MMSE) test and at least one other cognitive deficit beyond memory impairment. All participants involved were assessed by more than two neurologists with profound experience and received a couple of standard tests, including but not limited to physical examinations, medical history, and neuropsychological and neuroimaging examinations. Conversely, participants recorded with other neurological disorders, which could cause dementia were excluded (McKhann et al., 2011; Janelidze et al., 2018). So, a final total of 244 healthy controls (HC) matched for age and gender (137 females and 107 males, average age ± SD: 71.10 ± 8.31) were voluntarily enrolled in Shanghai, China. Healthy subjects were strictly evaluated by doctors to confirm that they have no symptoms of cognitive decline and do not comply with the criteria for mild cognitive impairment (MCI) or AD dementia (McKhann et al., 2011; Jack et al., 2018). Demographic information of the participants was presented in Table 1. This research was authorized by the Committee on Medical Ethics of Ruijin Hospital affiliated with Shanghai Jiao Tong University School of Medicine. A signed informed consent by the Declaration of Helsinki was submitted by the participants.

Table 1. Demographic and clinical characteristics of subjects included in the study.
DNA Extraction and Genotype Analysis
The genomic DNA was extracted from blood (2 ml) stored in an ethylene diamine tetraacetie acid (EDTA) anticoagulation tube using the phenol-chloroform-isopropyl alcohol method. Polymerase chain reaction (PCR) and extension primers scheme were achieved through Primer 5 software (PREMIER Biosoft International, Version 5.00). PCR materials were conducted by purification with phosphorylase (FastAP, Applied Biosystems) and exonuclease I (EXO I, Applied Biosystems). A consequent extension was applied by the ABI SNaPshot Multiplex Kit (Applied Biosystems). Extended products were purified with FastAP and loaded into ABI3730xl (Applied Biosystems). GeneMapper 4.0 (Applied Biosystems) was used to conduct data analysis. The SNaPshot technique (Applied Biosystems) was utilized for genotyping of SNPs. The following SNPs were tested: rs10792832, rs11136000, rs11218343, rs1990620, rs1990622, rs3173615, rs34860942, rs3764650, rs3792646, rs3818361, rs3851179, rs4147929, rs56081887, rs5848, rs6656401, rs6701713, rs6733839, rs704180, rs744373, rs9331888, and rs9637454. The SNPs rs429358 and rs7412 of the APOE gene were determined by Sanger sequencing. Details of primers were described in Supplementary Table 1.
Statistical Analysis
Statistical evaluations involved in the paper were conducted using SPSS software (two-sided significance level: P-value < 0.05) (version 26.0; IBM SPSS) or PLINK (Purcell et al., 2007) version 1.9.1 The SNP with minor allele frequencies (MAF) < 0.01, call rate < 95%, or not in Hardy-Weinberg equilibrium (HWE) were excluded. Differences in age, education level, and MMSE score between the two groups were examined for continuous variables using the Student’s t-test or non-parametric Mann-Whitney U-test. Dichotomous variables (such as gender, genotype distribution, allele frequency, and HWE) evaluations were conducted by the chi-square test. Then, logistic regression analysis adjusting for age and gender was used to figure out the odds ratio (OR) and 95% confidence intervals (CI) under varied genetic models (allele, dominant, recessive, and additive). It was defined that “A” means the major allele and “a” is the minor allele in the paper. We also defined dominant as 1 (aa + Aa) vs. 0 (AA), recessive as 1 (aa) vs. 0 (AA + Aa), and additive as 0 (AA) vs. 1 (Aa) vs. 2 (aa). Furthermore, the Bonferroni correction method was applied to perform multiple tests with aim to make the conclusion statistically convincing. G*Power software was used to evaluate all the SNPs’ genetic power.
Results
Demographic Characteristics of the Subjects
A total of 246 patients with AD and 244 age- and gender-matched healthy controls were enrolled. No statistically significant difference was found in age and gender between AD and the control group (all P > 0.05). In addition, patients with AD were less educated in comparison to the control group (P < 0.001), which matched well with the previous studies (Xu et al., 2016; Larsson et al., 2017). Patients with AD showed statistically significant lower MMSE scores compared with healthy controls (P < 0.001). Moreover, the patients with AD showed a much higher proportion of APOE ε4 allele carriers than the control subjects (P < 0.001). Detailed information about the participants was summarized in Table 1.
Association Analysis of Single Nucleotide Polymorphisms and Alzheimer’s Disease in Various Genetic Models
The genotype distribution of all SNPs in AD and controls was consistent with HWE. The minimum allele and genotype frequencies for all related SNPs were shown in Supplementary Table 2. Significant statistical differences were noted in the allele frequencies of rs3764650 (P = 0.010) and rs4147929 (P = 0.006) between patients with AD and the healthy ones. Regarding SNP genotype frequencies, it was detected that at rs3764650, the genotypes TT and GT experienced a higher risk for AD than the genotype GG, while at rs4147929, the genotypes GG and AG experienced a higher risk of AD than the genotype AA. Also, statistically significant difference also existed between patients with AD and controls of genotype frequencies in rs3764650 (P = 0.015) and rs4147929 (P = 0.030). The higher APOE level enhanced the risk of developing AD as shown in Table 1. Hence, these data were stratified by APOE ε4 levels to determine whether they impacted the correlation between SNPs and AD susceptibility. Among APOE ε4 carriers, the allele and genotype frequencies of ABCA7 rs3764650 were substantially different between AD and control cases [allele: P = 0.030, odds ratio (OR) = 1.80, 95% confidence interval (CI) 1.05–3.06, genotype: P = 0.034], and allele G was higher in the AD group than that in the control group (Table 2A). After adjusting for age and gender, the dominant and additive models of rs3764650 were nominally significantly related to AD (dominant model: P = 0.016, OR = 2.32, 95% CI 1.17–4.59; additive model: P = 0.043, OR = 1.77, 95% CI 1.02–3.08) (Table 2A). However, after the Bonferroni correction, these associations did not persist. Moreover, among APOE ε4 carriers, the allele and genotype frequencies of ABCA7 rs4147929 showed the obvious difference between patients with AD and healthy controls (allele: P = 0.001, OR = 2.49, 95% CI 1.42–4.37, genotype: P = 0.003), with allele A higher in the case group than that in the control group (Table 2B). After adjusting for age and gender, the dominant and additive models of rs4147929 were related to AD (dominant model: P = 0.002, OR = 3.06, 95% CI 1.52–6.17; additive model: P = 0.004, OR = 2.33, 95% CI 1.31–4.13) (Table 2B). Bonferroni correction was needed to confirm the relationship. To further examine the ABCA7 genetic association of AD, four genetic models including allele, dominant, recessive, and additive models were analyzed by logistic regression (Table 3 and Supplementary Table 3). The results revealed that rs3764650 of ABCA7 was correlated with the AD morbidity in the allele model (P = 0.010, OR = 1.43, 95% CI 1.09–1.89) without adjusting the age and gender, dominant (P = 0.004, OR = 1.71, 95% CI 1.19–2.46), and additive (P = 0.012, OR = 1.42, 95% CI 1.08–1.86) models after adjusting the age and gender. In addition, rs4147929 of ABCA7 was correlated with the risk of developing AD in the allele (P = 0.006, OR = 1.45, 95% CI 1.11–1.89) without adjusting the age and gender, dominant (P = 0.012, OR = 1.59, 95% CI 1.11–2.27) and additive (P = 0.010, OR = 1.40, 95% CI 1.08–1.81) models after adjusting the age and gender. Again, the conclusion also needs to be confirmed by Bonferroni correction.
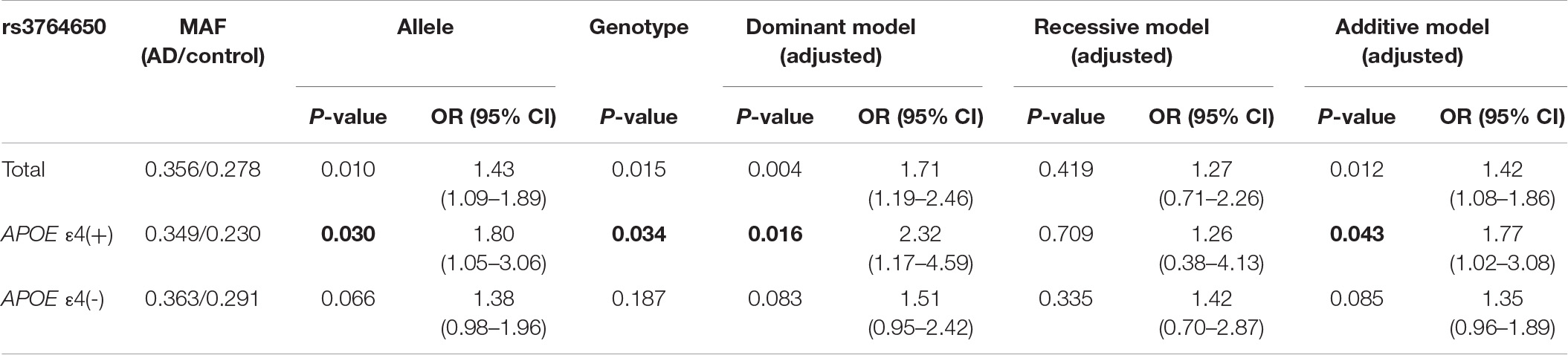
Table 2A. Association of rs3764650 of ABCA7 gene with AD risk stratified by APOE ε4 status.

Table 2B. Association of rs4147929 of ABCA7 gene with AD risk stratified by APOE ε4 status.
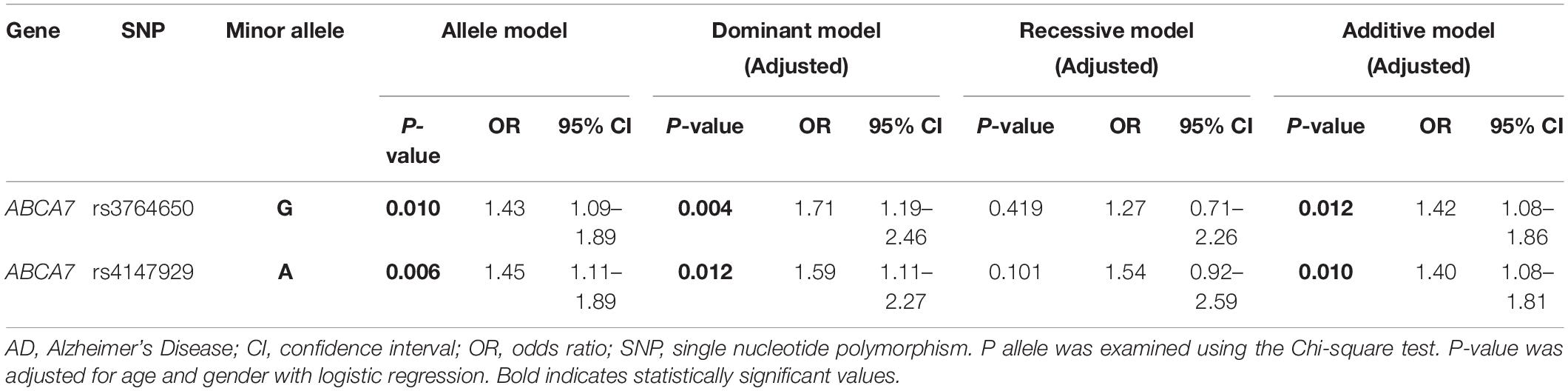
Table 3. Association of SNPs of candidate genes and odds ratio to AD risk.
Discussion
The interaction between rs3764650 and rs4147929 of ABCA7 and the risk factors of AD in southern China was validated in the paper. Differences among APOE ε4 carriers were more dominant when allele and genotype distributions were stratified by APOE ε4 status. It was found that individuals with a heterozygote GT at rs3764650 had a higher susceptibility to AD, while individuals with heterozygote AG at rs4147929 suffered higher AD morbidity. But the relationship between the remaining SNPs and AD was not replicated in the southern Chinese group. Similarly, varied populations could suffer different AD risk genetic variants. Last but not least, the relatively limited sample compared to recent consortium-based GWAS may lead to the dismissal of replication.
A highly conserved protein, a part of the ABCB family of ATP-binding cassette (ABC) transporters, was encoded by the ABCA7, which was known as ATP-binding cassette subfamily A member 7 (Takahashi et al., 2005). The ABCA7 also plays a role in the transportation of cholesterol across membranes (Le Guennec et al., 2016), particularly in the hippocampal CA1 neurons and microglia (Kim et al., 2005), suggesting a role in amyloid clearance and fibril formation (Chan et al., 2008; Aikawa et al., 2018). Recently, some studies investigated the interaction between ABCA7 rs3764650 polymorphism and the morbidity of AD. However, the results were controversial (Hollingworth et al., 2011; Allen et al., 2012; Lambert et al., 2013; Reitz et al., 2013). Our study confirmed that ABCA7 rs3764650 might be important genetic factor in the pathophysiology of AD. Furthermore, previous researchers found that the ABCA7 rs4147929 might be a predisposing factor for late-onset AD (Hollingworth et al., 2011; Lambert et al., 2013; Talebi et al., 2020). In line with that, this study provides supportive evidence for the relationship between rs4147929 and AD. However, it should be noted that the conclusion needs further verification. Meanwhile, the shortcoming of our study is that there is no distinction between early-onset and late-onset AD, and further subgroup research stratified by age of onset will be carried out in the future.
The research plays an important role in figuring out the basic genes of AD in the Asian population and offers useful findings that genetic risk factors varied in different populations. It should be noted that there are certain limitations in the study. Most importantly, the relatively small-sized sample due to its single-centered nature may affect the validity. A consequent meta-analysis of the Asian population with a bigger sample size should be conducted. Secondly, follow-up studies are required to evaluate more participants’ loci for AD susceptibility. Simultaneously, the participants involved here should be followed regularly to find their cognitive variations and figure out the association between genetic polymorphisms and clinical performance. Last but not least, we will conduct a comprehensive neuropsychological battery to assess cognitive function in the future study.
Conclusion
In summary, this study suggested that the G-allele at rs3764650 and the A-allele at rs4147929 appeared at higher risk for developing AD in the southern Chinese population, particularly in APOE ε4 carriers. In addition, rs3764650 and rs4147929 of ABCA7 were observed to be associated with AD. Further investigations on the role of the risk genes in the pathogenesis of AD are essential in the following research.
Data Availability Statement
The original contributions presented in the study are included in the article/Supplementary Material, further inquiries can be directed to the corresponding author/s.
Ethics Statement
The studies involving human participants were reviewed and approved by this research was permitted by the Committee on Medical Ethics of Ruijin Hospital Affiliated to Shanghai Jiao Tong University School of Medicine. The patients/participants provided their written informed consent to participate in this study.
Author Contributions
JL and WX conceived and designed the studies. LW analyzed the results and wrote the manuscript. LW, YJ, AZ, XX, GY, YZ, YW, and YD performed the research. All authors contributed to the article and approved the submitted version.
Funding
This work was supported by the National Natural Science Foundation of China (82071415 and 81873778).
Conflict of Interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Publisher’s Note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
Acknowledgments
We thank all the patients and healthy controls who participated in this study.
Supplementary Material
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fnagi.2022.819499/full#supplementary-material
Footnotes
References
Aikawa, T., Holm, M. L., and Kanekiyo, T. (2018). Abca7 and pathogenic pathways of alzheimer’s disease. Brain Sci. 8:27.
Allen, M., Zou, F., Chai, H. S., Younkin, C. S., Crook, J., Pankratz, V. S., et al. (2012). Novel late-onset alzheimer disease loci variants associate with brain gene expression. Neurology 79, 221–228. doi: 10.1212/WNL.0b013e3182605801
Alzheimer’s Association (2021). 2021 Alzheimer’s disease facts and figures. Alzheimers Dement. 17, 327–406. doi: 10.1002/alz.12328
Andrews, S. J., Fulton-Howard, B., and Goate, A. (2020). Interpretation of risk loci from genome-wide association studies of alzheimer’s disease. Lancet Neurol. 19, 326–335. doi: 10.1016/S1474-4422(19)30435-1
Chan, S. L., Kim, W. S., Kwok, J. B., Hill, A. F., Cappai, R., Rye, K. A., et al. (2008). Atp-binding cassette transporter a7 regulates processing of amyloid precursor protein in vitro. J. Neurochem. 106, 793–804. doi: 10.1111/j.1471-4159.2008.05433.x
Chanock, S. J., Manolio, T., Boehnke, M., Boerwinkle, E., Hunter, D. J., Thomas, G., et al. (2007). Replicating genotype-phenotype associations. Nature 447, 655–660.
De Strooper, B., and Karran, E. (2016). The cellular phase of alzheimer’s disease. Cell 164, 603–615.
DeTure, M. A., and Dickson, D. W. (2019). The neuropathological diagnosis of alzheimer’s disease. Mol. Neurodegener. 14:32.
Dubois, B., Feldman, H. H., Jacova, C., DeKosky, S. T., Barberger-Gateau, P., Cummings, J., et al. (2007). Research criteria for the diagnosis of alzheimer’s disease: revising the nincds–adrda criteria. Lancet Neurol. 6, 734–746. doi: 10.1016/S1474-4422(07)70178-3
Dugan, A. J., Nelson, P. T., Katsumata, Y., Shade, L. M. P., Boehme, K. L., Teylan, M. A., et al. (2021). Analysis of genes (tmem106b, grn, abcc9, kcnmb2, and apoe) implicated in risk for late-nc and hippocampal sclerosis provides pathogenetic insights: a retrospective genetic association study. Acta Neuropathol. Commun. 9:152. doi: 10.1186/s40478-021-01250-2
Giri, M., Zhang, M., and Lu, Y. (2016). Genes associated with alzheimer’s disease: an overview and current status. Clin. Interv. Aging 11, 665–681. doi: 10.2147/CIA.S105769
Harold, D., Abraham, R., Hollingworth, P., Sims, R., Gerrish, A., Hamshere, M. L., et al. (2009). Genome-wide association study identifies variants at clu and picalm associated with alzheimer’s disease. Nat. Genet. 41, 1088–1093. doi: 10.1038/ng.440
Hollingworth, P., Harold, D., Sims, R., Gerrish, A., Lambert, J. C., Carrasquillo, M. M., et al. (2011). Common variants at abca7, ms4a6a/ms4a4e, epha1, cd33 and cd2ap are associated with alzheimer’s disease. Nat. Genet. 43, 429–435. doi: 10.1038/ng.803
Hu, Y., Sun, J. Y., Zhang, Y., Zhang, H., Gao, S., Wang, T., et al. (2021). Rs1990622 variant associates with alzheimer’s disease and regulates tmem106b expression in human brain tissues. BMC Med. 19:11. doi: 10.1186/s12916-020-01883-5
Jack, C. R. Jr., Bennett, D. A., Blennow, K., Carrillo, M. C., Dunn, B., Haeberlein, S. B., et al. (2018). Nia-aa research framework: toward a biological definition of alzheimer’s disease. Alzheimers Dement. 14, 535–562. doi: 10.1016/j.jalz.2018.02.018
Janelidze, S., Mattsson, N., Stomrud, E., Lindberg, O., Palmqvist, S., Zetterberg, H., et al. (2018). Csf biomarkers of neuroinflammation and cerebrovascular dysfunction in early alzheimer disease. Neurology. 91, e867–e877. doi: 10.1212/WNL.0000000000006082
Jansen, I. E., Savage, J. E., Watanabe, K., Bryois, J., Williams, D. M., Steinberg, S., et al. (2019). Genome-wide meta-analysis identifies new loci and functional pathways influencing alzheimer’s disease risk. Nat. Genet. 51, 404–413.
Jun, G., Ibrahim-Verbaas, C. A., Vronskaya, M., Lambert, J. C., Chung, J., Naj, A. C., et al. (2016). A novel alzheimer disease locus located near the gene encoding tau protein. Mol. Psychiatry 21, 108–117. doi: 10.1038/mp.2015.23
Katsumata, Y., Nelson, P. T., Ellingson, S. R., and Fardo, D. W. (2017). Gene-based association study of genes linked to hippocampal sclerosis of aging neuropathology: Grn, tmem106b, abcc9, and kcnmb2. Neurobiol. Aging 53, 193.e17–193.e25. doi: 10.1016/j.neurobiolaging.2017.01.003
Kim, W. S., Fitzgerald, M. L., Kang, K., Okuhira, K., Bell, S. A., Manning, J. J., et al. (2005). Abca7 null mice retain normal macrophage phosphatidylcholine and cholesterol efflux activity despite alterations in adipose mass and serum cholesterol levels. J. Biol. Chem. 280, 3989–3995. doi: 10.1074/jbc.M412602200
Kunkle, B. W., Grenier-Boley, B., Sims, R., Bis, J. C., Damotte, V., Naj, A. C., et al. (2019). Genetic meta-analysis of diagnosed alzheimer’s disease identifies new risk loci and implicates abeta, tau, immunity and lipid processing. Nat. Genet. 51, 414–430.
Lambert, J. C., Heath, S., Even, G., Campion, D., Sleegers, K., Hiltunen, M., et al. (2009). Genome-wide association study identifies variants at clu and cr1 associated with alzheimer’s disease. Nat. Genet. 41, 1094–1099. doi: 10.1038/ng.439
Lambert, J. C., Ibrahim-Verbaas, C. A., Harold, D., Naj, A. C., Sims, R., Bellenguez, C., et al. (2013). Meta-analysis of 74,046 individuals identifies 11 new susceptibility loci for alzheimer’s disease. Nat. Genet. 45, 1452–1458. doi: 10.1038/ng.2802
Larsson, S. C., Traylor, M., Malik, R., Dichgans, M., Burgess, S., Markus, H. S., et al. (2017). Modifiable pathways in alzheimer’s disease: mendelian randomisation analysis. BMJ 359:j5375. doi: 10.1136/bmj.j5375
Le Guennec, K., Nicolas, G., Quenez, O., Charbonnier, C., Wallon, D., Bellenguez, C., et al. (2016). ABCA7 rare variants and Alzheimer disease risk. Neurology 86, 2134–2137.
Lee, M. J., Chen, T. F., Cheng, T. W., and Chiu, M. J. (2011). Rs5848 variant of progranulin gene is a risk of alzheimer’s disease in the taiwanese population. Neurodegener. Dis. 8, 216–220. doi: 10.1159/000322538
Liu, C. C., Liu, C. C., Kanekiyo, T., Xu, H., and Bu, G. (2013). Apolipoprotein e and alzheimer disease: risk, mechanisms and therapy. Nat. Rev. Neurol. 9, 106–118. doi: 10.1038/nrneurol.2012.263
Lu, R. C., Wang, H., Tan, M. S., Yu, J. T., and Tan, L. (2014). Tmem106b and apoe polymorphisms interact to confer risk for late-onset alzheimer’s disease in han chinese. J. Neural Transm. 121, 283–287. doi: 10.1007/s00702-013-1106-x
McKhann, G. M., Knopman, D. S., Chertkow, H., Hyman, B. T., Jack, C. R. Jr., Kawas, C. H., et al. (2011). The diagnosis of dementia due to alzheimer’s disease: recommendations from the national institute on aging-alzheimer’s association workgroups on diagnostic guidelines for alzheimer’s disease. Alzheimers Dement. 7, 263–269. doi: 10.1016/j.jalz.2011.03.005
Nelson, P. T., Estus, S., Abner, E. L., Parikh, I., Malik, M., Neltner, J. H., et al. (2014). Abcc9 gene polymorphism is associated with hippocampal sclerosis of aging pathology. Acta Neuropathol. 127, 825–843. doi: 10.1007/s00401-014-1282-2
Nho, K., and Saykin, A. J. (2016). Alzheimer’s disease neuroimaging I, Nelson PT. Hippocampal sclerosis of aging, a common alzheimer’s disease ‘mimic’: risk genotypes are associated with brain atrophy outside the temporal lobe. J. Alzheimers Dis. 52, 373–383. doi: 10.3233/JAD-160077
Purcell, S., Neale, B., Todd-Brown, K., Thomas, L., Ferreira, M. A., Bender, D., et al. (2007). Plink: a tool set for whole-genome association and population-based linkage analyses. Am. J. Hum. Genet. 81, 559–575. doi: 10.1086/519795
Reitz, C., Jun, G., Naj, A., Rajbhandary, R., Vardarajan, B. N., Wang, L. S., et al. (2013). Variants in the atp-binding cassette transporter (abca7), apolipoprotein e 4,and the risk of late-onset alzheimer disease in african americans. JAMA 309, 1483–1492. doi: 10.1001/jama.2013.2973
Rutherford, N. J., Carrasquillo, M. M., Li, M., Bisceglio, G., Menke, J., and Josephs, K. A. (2012). TMEM106B risk variant is implicated in the pathologic presentation of Alzheimer disease. Neurology 79, 717–718. doi: 10.1212/WNL.0b013e318264e3ac
Serrano-Pozo, A., Das, S., and Hyman, B. T. (2021). Apoe and alzheimer’s disease: advances in genetics, pathophysiology, and therapeutic approaches. Lancet Neurol. 20, 68–80. doi: 10.1016/S1474-4422(20)30412-9
Steinberg, S., Stefansson, H., Jonsson, T., Johannsdottir, H., Ingason, A., Helgason, H., et al. (2015). Loss-of-function variants in abca7 confer risk of alzheimer’s disease. Nat. Genet. 47, 445–447. doi: 10.1038/ng.3246
Takahashi, K., Kimura, Y., Nagata, K., Yamamoto, A., Matsuo, M., and Ueda, K. (2005). Abc proteins: key molecules for lipid homeostasis. Med. Mol. Morphol. 38, 2–12. doi: 10.1007/s00795-004-0278-8
Talebi, M., Delpak, A., Khalaj-Kondori, M., Sadigh-Eteghad, S., Talebi, M., Mehdizadeh, E., et al. (2020). Abca7 and epha1 genes polymorphisms in late-onset alzheimer’s disease. J. Mol. Neurosci. 70, 167–173.
Vardarajan, B. N., Zhang, Y., Lee, J. H., Cheng, R., Bohm, C., Ghani, M., et al. (2015). Coding mutations insorl1and alzheimer disease. Ann. Neurol. 77, 215–227.
Viswanathan, J., Makinen, P., Helisalmi, S., Haapasalo, A., Soininen, H., and Hiltunen, M. (2009). An association study between granulin gene polymorphisms and alzheimer’s disease in finnish population. Am. J. Med. Genet. B Neuropsychiatr. Genet. 150B, 747–750. doi: 10.1002/ajmg.b.30889
Xu, H. M., Tan, L., Wan, Y., Tan, M. S., Zhang, W., Zheng, Z. J., et al. (2017). Pgrn is associated with late-onset alzheimer’s disease: a case-control replication study and meta-analysis. Mol. Neurobiol. 54, 1187–1195. doi: 10.1007/s12035-016-9698-4
Xu, W., Tan, L., Wang, H. F., Tan, M. S., Tan, L., Li, J. Q., et al. (2016). Education and risk of dementia: dose-response meta-analysis of prospective cohort studies. Mol. Neurobiol. 53, 3113–3123. doi: 10.1007/s12035-015-9211-5
Keywords: Alzheimer’s disease, ABCA7, rs4147929, rs3764650, single nucleotide polymorphisms
Citation: Wang L, Jiao Y, Zhao A, Xu X, Ye G, Zhang Y, Wang Y, Deng Y, Xu W and Liu J (2022) Analysis of Genetic Association Between ABCA7 Polymorphism and Alzheimer’s Disease Risk in the Southern Chinese Population. Front. Aging Neurosci. 14:819499. doi: 10.3389/fnagi.2022.819499
Received: 21 November 2021; Accepted: 19 April 2022;
Published: 25 May 2022.
Edited by:
Nilton Custodio, Peruvian Institute of Neurosciences (IPN), PeruReviewed by:
Mario Reynaldo Cornejo-Olivas, National Institute of Neurological Sciences, PeruClaudio Villegas-Llerena, Universidad de San Martín de Porres, Peru
Copyright © 2022 Wang, Jiao, Zhao, Xu, Ye, Zhang, Wang, Deng, Xu and Liu. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Wei Xu, eHcxMTI0NkByamguY29tLmNu; Jun Liu, amx5MDUyMEBob3RtYWlsLmNvbQ==