 Adrian Guerrero-Moreno1
Adrian Guerrero-Moreno1 Christophe Baudouin1,2,3
Christophe Baudouin1,2,3 Stéphane Melik Parsadaniantz1
Stéphane Melik Parsadaniantz1 Annabelle Réaux-Le Goazigo1*
Annabelle Réaux-Le Goazigo1*- 1Sorbonne Université, INSERM, CNRS, Institut de la Vision, IHU FOReSIGHT, Paris, France
- 2CHNO des Quinze-Vingts, IHU FOReSIGHT, INSERM-DGOS CIC 1423, Paris, France
- 3Department of Ophthalmology, Ambroise Paré Hospital, AP-HP, University of Versailles Saint-Quentin-en-Yvelines, Boulogne-Billancourt, France
The cornea is the most densely innervated and sensitive tissue in the body. The cornea is exclusively innervated by C- and A-delta fibers, including mechano-nociceptors that are triggered by noxious mechanical stimulation, polymodal nociceptors that are excited by mechanical, chemical, and thermal stimuli, and cold thermoreceptors that are activated by cooling. Noxious stimulations activate corneal nociceptors whose cell bodies are located in the trigeminal ganglion (TG) and project central axons to the trigeminal brainstem sensory complex. Ocular pain, in particular, that driven by corneal nerves, is considered to be a core symptom of inflammatory and traumatic disorders of the ocular surface. Ocular surface injury affecting corneal nerves and leading to inflammatory responses can occur under multiple pathological conditions, such as chemical burn, persistent dry eye, and corneal neuropathic pain as well as after some ophthalmological surgical interventions such as photorefractive surgery. This review depicts the morphological and functional changes of corneal nerve terminals following corneal damage and dry eye disease (DED), both ocular surface conditions leading to sensory abnormalities. In addition, the recent fundamental and clinical findings of the importance of peripheral and central neuroimmune interactions in the development of corneal hypersensitivity are discussed. Next, the cellular and molecular changes of corneal neurons in the TG and central structures that are driven by corneal nerve abnormalities are presented. A better understanding of the corneal nerve abnormalities as well as neuroimmune interactions may contribute to the identification of a novel therapeutic targets for alleviating corneal pain.
Introduction
The cornea is the most densely innervated tissue in the human body. Corneal innervation is estimated as 300–600 times and 20–40 more innervated than skin and tooth pulp (Muller et al., 2003; Marfurt et al., 2010). The human corneal innervation is organized into four layers: mid stromal nerves, subepithelial plexus, subbasal nerve plexus, and intraepithelial nerve terminals (Muller et al., 2003; Marfurt et al., 2019). The sensory innervation comes from corneal primary afferent neurons whose cell bodies are located in the ophthalmic (V1) branch of the trigeminal ganglion (TG). Corneal neurons represent only 1–5% of total trigeminal neurons (Marfurt and Del Toro, 1987; Launay et al., 2015). The central axons of corneal sensory neurons terminate in two regions of the spinal trigeminal complex (V or Sp5): the sensory trigeminal subnucleus interpolaris/caudalis (Vi/Vc) transition and the subnucleus caudalis/upper cervical cord (Vc/C1) junction regions (Marfurt and Del Toro, 1987; Strassman and Vos, 1993; Meng and Bereiter, 1996). The second order neurons project to the thalamus and synapse with third order neurons projecting to cortical regions (primary somatosensory cortex) (Figure 1). Therefore, any corneal injury or corneal nerve abnormalities may trigger molecular, cellular, and functional changes within the TG and the central nervous system, which may contribute to acute and chronic pain.
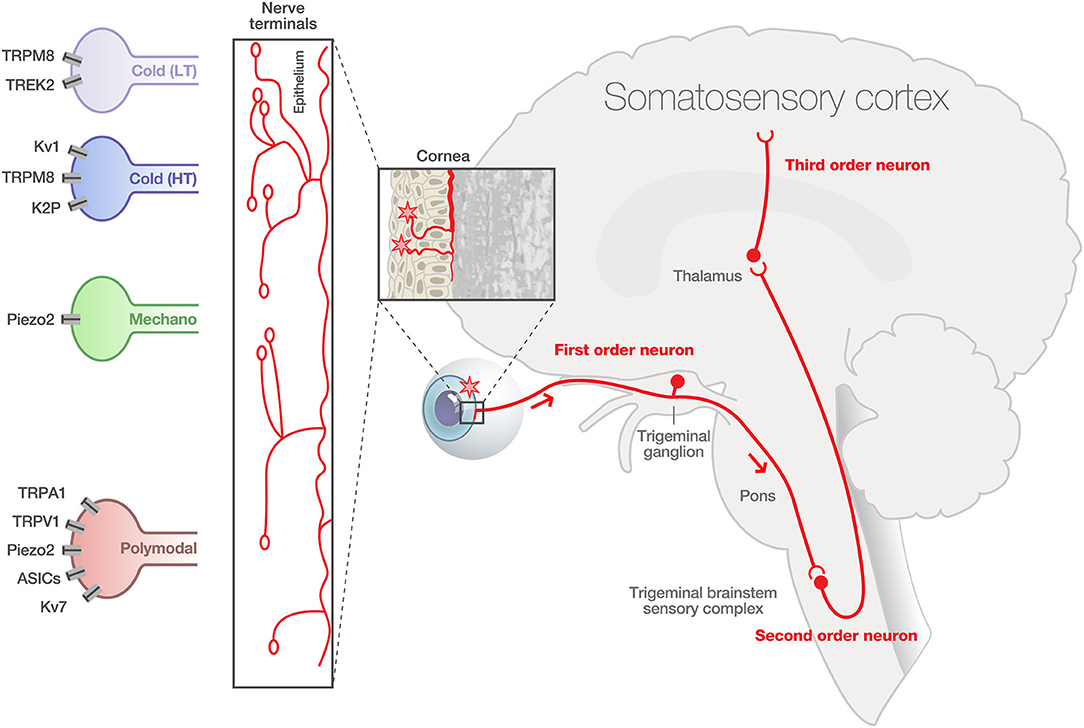
Figure 1. Physiological corneal pain processing. The cornea contained a high density and heterogeneity of A delta and C fibers expressing specialized transient receptor channels, able to respond to diverse environmental stimuli of varying intensities (cold, mechanical, and polymodal). The intraepithelial nerve fibers exhibit distinct morphologies: complex, ramified, and simple. First order sensory neurons innervating the cornea have their cell body in the ophthalmic branch (V1) of the trigeminal ganglion (TG). The central terminals of these pseudo unipolar neurons synapse in the trigeminal brainstem sensory complex with second order nociceptive projection neurons. These neurons project to the thalamus and synapse with third-order neurons projecting to cortical regions (primary somatosensory cortex). LT, low-threshold cold thermoreceptors; HT, high-threshold cold thermoreceptors. Modified from Belmonte et al. (2015) and Rosenthal and Borsook (2016).
Functional Classification of Corneal Fibers
The cornea receives sensitive and autonomous innervation. Corneal nerve fibers have been classified according to different criteria—morphology (intraepithelial nerves), conduction velocity (diameter, myelinated or not), and function/modality (stimulus sensitivity and specificity, electrophysiological properties)—and according to the expression of very specific biochemical markers. Unlike somatic nerve innervation, the cornea lacks A-beta fibers. Cornea is composed of two types of fibers:
• the high speed myelinated Aδ fibers with a larger diameter represent about 20% of the total population of corneal afferent fibers (MacIver and Tanelian, 1993a; Bron et al., 2014).
• slower-conducting unmyelinated C fibers with a small diameter are the most frequent (around 80%) (Gallar et al., 1993; MacIver and Tanelian, 1993a; Hirata and Meng, 2010).
Two sub-populations of C fibers have been identified: peptidergic and non-peptidergic fibers. In rodents, peptidergic fibers contain substance P (SP, 10–20%) or calcitonin gene-related peptide (CGRP, 40–60%). All SP+ terminals are CGRP+ in mice (Ivanusic et al., 2013). Other mediators are also found, such as neurokinin A, serotonin, somatostatin, as well as cholecystokinin or gastrin (Gonzalez-Coto et al., 2014). In humans, to date, only substance P and CGRP have been clearly identified in corneal fibers. A second population of C fibers, the non- peptidergic C fibers, does not express these peptides but has a strong affinity for isolectin B4. In mice, about 20% of corneal neurons are non-peptidergic (Ivanusic et al., 2013).
The majority of corneal nerve fibers are nociceptive and cold thermoreceptors fibers and the remaining fibers are sympathetic or parasympathetic post-ganglion fibers of the autonomic nervous system.
Autonomic nervous fibers of sympathetic (upper cervical ganglion) and parasympathetic system (ciliary ganglion) participate in the regulation of corneal wound healing and re-epithelialization (Jones and Marfurt, 1996; Xue et al., 2018; Xiao et al., 2019). Theses biological processes involve a number of concerted events including cell migration, proliferation, inflammation, and differentiation and extracellular matrix remodeling (Ljubimov and Saghizadeh, 2015). Interestingly, it was recently reported that the mouse autonomous system modulates inflammation and epithelial renewal after corneal abrasion through the activation of distinct local macrophages (Xue et al., 2018; Xiao et al., 2019).
In rodents and cats, these fibers represent about 10% of corneal fibers. Parasympathetic fibers, originating from the ciliary ganglion, contain intestinal vasoactive peptide (VIP), neuropeptide Y, galanin, and Met-enkephalin (Jones and Marfurt, 1998; Muller et al., 2003).
Electrophysiological studies showed that the cornea is innervated by three distinct classes of peripheral sensory nerve fibers:
Mechano-nociceptors (Aδ fibers) are activated only by soft mechanic forces in the order of magnitude closed to that required to damage corneal epithelial cells. In general, mechano-nociceptors only fire when a mechanical stimulus is applied or removed in a phasic way (Belmonte et al., 1991; MacIver and Tanelian, 1993b). Piezo2, a long transmembrane protein and non-selective cation channel mechanically activated (Coste et al., 2010; Shin et al., 2019), has emerged as a marker of mechano-nociceptors in primary sensory neurons.
Cold receptors (Aδ and C fibers) are activated by thermal changes (Gallar et al., 1993; Carr et al., 2003). These are classified into two subgroups (Alcalde et al., 2018).
° High background-low threshold (HB-LT) cold receptors are the principal generators of spontaneous activity (high background) at ocular surface constant temperature. These fibers change their firing frequency according to slight temperature changes (1–2°C, low threshold). Additionally, they detect changes in ocular surface osmolarity (Hirata and Meng, 2010; Parra et al., 2014; Gonzalez-Gonzalez et al., 2017). A hyperosmolar tear film with values close to those found in dry eye patients increases the sensitivity of cold-sensitive neurons.
° Low background-high threshold (LB-HT) cold receptors do not contribute as HB-LT to basal ongoing activity (low background), but instead fire under sharp drops in temperature (>4°C) (Gonzalez-Gonzalez et al., 2017).
TRPM8 (transient receptor potential cation channel subfamily M member 8) is considered as a marker of cold receptors. TRPM8 is activated by menthol and by cold (<23°C) (McKemy et al., 2002; Peier et al., 2002). Due to these characteristics, it is the putative principal sensor of cold.
Indeed, TRPM8 receptors appear to be first activated on the ocular surface after evaporation of the tear film (Belmonte et al., 2017) and mild cooling of the ocular surface has been reported to increase lacrimation via TRPM8 activation of corneal cool primary afferent neurons (Robbins et al., 2012). In the same line, it was found that TRPM8 knockout mice have a lower level of basal tear flow (Parra et al., 2010).
Polymodal nociceptors (Aδ and C fibers) are the most abundant population in the cornea (Gonzalez-Gonzalez et al., 2017). These fibers are activated by different types of stimuli:
° Mechanical: In contrast to pure mechano-nociceptors, polymodal nociceptors have a lower threshold and fire continuously (tonic) while the stimulus is present (Gallar et al., 1993).
° Thermal: They are activated by temperatures above 37°C (Gallar et al., 1993).
° Chemical: There is a broad variety of molecules activating these nociceptors, including protons (pH drops) and inflammatory mediators (prostaglandins, bradykinin, and capsaicin) (Chen et al., 1997).
TRPV1 (transient receptor potential cation channel subfamily V member 1), a non-selective cation channel, is considered as a marker of polymodal sensory fibers (Belmonte et al., 1991). Thus, it represents the principal sensor of hot painful stimulus.
Retrograde tracing anatomical experiments were used to identify the whole population of corneal neurons in the TG and to determine the proportion of the corneal neurons expressing the molecular marker of polymodal, mechanoreceptors and cold cells. This studies have demonstrated by in situ hybridization that Piezo2 represents 28–30% of corneal afferent neurons in guinea pig TGs and do not co-express TRPV1 (Alamri et al., 2015) or TRPM8 (Bron et al., 2014). The co-expression of TRPM8 and TRPV1 occurs in 6% of TRPV1+ cells corresponding to 31% of TRPM8+ cells (Alamri et al., 2015). Interestingly, all SP-reactive neurons were also TRPV1+ in rats (Murata and Masuko, 2006). Therefore, they constitute separate populations of corneal sensory neurons. The proportion of corneal neurons expressing TRPV1 varies depending on the animal species and the tracer used: in rats, 37% when traced with cholera toxin (Murata and Masuko, 2006) and 23% when traced with Fluoro-Gold (Nakamura et al., 2007); in guinea pigs, it reaches 45% of corneal neurons when traced with Fast blue (Alamri et al., 2015). TRPM8 is expressed in 8–15% of corneal neurons in guinea pigs (Bron et al., 2014; Alamri et al., 2015) and 18–22% in mice (Ivanusic et al., 2013; Alcalde et al., 2018).
Anatomical studies focusing on corneal nerve terminals found that corneal polymodal (TRPV1+) and cold (TRPM8+) neurons have distinct nerve terminal morphologies. It was demonstrated that, compared to TRPV1 neurons, TRPM8-IR corneal nerve endings in both guinea pig and mouse corneal epithelia had complex (longer and more branched) morphology (Ivanusic et al., 2013; Alamri et al., 2015, 2018; He et al., 2019) (Figure 1). The presence of Piezo2+ channels has been recently found in corneal nerve fibers in mice (Fernandez-Trillo et al., 2020).
Accumulating fundamental and clinical studies have reported morphological, structural, and/or functional corneal nerve abnormalities, which may contribute to pain-related sensory abnormalities.
Nociception, Pain, and Neurosensory Abnormalities
Nociception is the neural process of encoding noxious stimuli, whereas pain is defined as an unpleasant sensory and emotional experience associated with, or resembling that associated with, actual or potential tissue damage [the International Association for the Study of Pain (IASP)] (Raja et al., 2020).
Nociceptive pain is a physiological protective warning signal against imminent or ongoing tissue damage. Nociceptive pain starts at the periphery by the activation of nociceptors by physical tissue destruction or by chemical exposure, or thermal processes. Inflammatory pain occurs in the presence of tissue injury and active inflammation. It is the consequence of the activation and alteration of nociceptor function both by immune signals and signals arising from damaged cells (Woolf, 2020).
In this context, proinflammatory mediators participate in the sensitization of the nerves, meaning the reduction of the pain threshold (allodynia) and exacerbated response to a painful stimulus (hyperalgesia). Therefore, sensitization can occur both in the periphery (peripheral sensitization) and in the central nervous system (central sensitization).
Pain can extend beyond its protective usefulness, lasting in the absence of nociceptive stimuli. Neuropathic pain, a chronic pain condition, is defined by the IASP as “pain caused by a lesion or disease of the somatosensory nervous system.” By definition, the origin of the neuropathic pain can be peripheral or central. There is important interest in the field, as the current treatments for alleviating neuropathic pain are often inefficacious and/or produce severe side effects.
In the ocular surface, and more specifically in the cornea, all of these physiological and pathophysiological conditions can occur. Pain can manifest as a result of a noxious stimulus or damage in the ocular surface anatomy (nociceptive pain), or it can result from abnormalities in the ocular surface neurosensory apparatus itself (neuropathic pain).
Neurosensory abnormalities within the corneal nociceptive pathway could lead to pathological conditions such as pain and ocular discomfort that are hallmark of dry eye disease (DED) and the emerging ocular neuropathic pain. Corneal neuropathic pain may have various etiological origins, which include DED, persistent corneal nerve injury, and photorefractive surgeries (Dieckmann et al., 2017; McMonnies, 2017; Aggarwal et al., 2019). Dry eye is defined as a “multifactorial disease of the ocular surface characterized by a loss of homeostasis of the tear film, and accompanied by ocular symptoms, in which tear film instability and hyperosmolarity, ocular surface inflammation and damage, and neurosensory abnormalities play etiological roles" (Craig et al., 2017). The prevalence of DED, which ranges from 5 to 50% of the adult population, increases with age, especially after 50 years, and affects more women than men (Stapleton et al., 2017). DED has gained recognition as a public health problem given its prevalence, morbidity, and cost implications.
Importantly, the cellular and molecular mechanisms underlying the neurosensory abnormalities observed after corneal damage and in DED are currently an open and very active research field.
This increasing understanding of the corneal nerve morphological and functional aspects will bring new insights into their contribution to the physiology and pathophysiology of corneal pain perception and will provide novel opportunities for more efficient therapeutic treatments for this ocular surface diseases.
In the context of other ocular surface diseases, excellent reviews have already provided detailed information about the morphological and functional abnormalities of corneal nerves in herpetic keratitis, neuropathic keratitis, or keratoconus (Cruzat et al., 2017) and diabetes (Markoulli et al., 2018).
The purpose of this review is first to provide an overview of the morphological, molecular, and cellular changes of corneal nerves in rodents after corneal damage and persistent DED. The cellular and molecular changes of corneal neurons in the TG driven by corneal nerve abnormalities are discussed next. Moreover, we depict how peripheral neuroimmune interactions shape the peripheral and the central nervous system. Clinical evidence for corneal nerve abnormalities associated with DED and corneal neuropathic pain is also presented as well as several important questions that remain to be addressed.
Fundamental Studies on Morphologic Abnormalities of Corneal Nerve Terminals
The cornea is a valuable tissue for studying peripheral sensory nerve morphology and function due to its transparency, dense innervation, and accessibility. Corneal nerve structure and function are adversely affected by many ophthalmic and systemic disorders. Experimental descriptive studies exploring the changes of corneal nerve morphology have improved our understanding of how corneal nerve density is altered under DED and corneal injury conditions. Thus, transgenic thy1-yellow fluorescent protein (thy1-YFP) mice, in which corneal nerves express the YFP protein driven by thy1 promoter, represent a useful model for in vivo investigation of peripheral nerve structure (Yu and Rosenblatt, 2007; Namavari et al., 2011; Chaudhary et al., 2012; Sarkar et al., 2012, 2019; Bouheraoua et al., 2019). A decrease in stromal nerve fiber density as well as inflammation were observed in corneas from thy1-YFP mice submitted to experimental DED induced by chronic topical instillation with benzalkonium chloride, a quaternary ammonium used as preservative (Sarkar et al., 2012).
In addition, immunohistochemistry detecting all corneal nerve fibers in tissue samples and in vivo confocal microscopy (IVCM, a non-invasive high-resolution real-time imaging device allowing layer-by-layer analysis of the corneal ultrastructure) have covered aspects of the morphological basis of corneal nerve changes in rodents both in physiological and pathophysiological conditions. A decrease in corneal nerve density was reported in preclinical models of experimental DED induced by prolonged (28 days) (Simsek et al., 2019) or acute (up to 10 days) (Stepp et al., 2018a) scopolamine administration. Similar observations were made in cd25 null mice, which constitute a model of Sjögren Syndrome dry eye (Stepp et al., 2018b).
Tear hyperosmolarity plays a critical role in the initiation and/or perpetuation of DED, which could have consequences with respect to corneal nerve abnormality. Indeed, a reduction in the density of corneal intraepithelial nerves and terminals, in addition to a sensitized ocular surface to hypertonicity, was also recently reported in a murine model of tear hyperosmolarity (Guzman et al., 2020).
Furthermore, surgically induced chronic dry eye models obtained after the unilateral removal of extra-orbital lacrimal gland in mice (Yamazaki et al., 2017) or after the excision of the extraorbital lachrymal gland and Harderian gland had similar findings (Fakih et al., 2019): a reduction in corneal nerve density accompanied with corneal allodynia. Finally, structural abnormalities including a profound loss of nerve density in the sub-basal nerves and increasing of beading and tortuosity of stromal nerve trunks also occurred in a model of corneal injury-induced neuropathic pain (Cho et al., 2019; Pham and Bazan, 2020).
Altogether, these morphological changes in corneal density innervation reported from the fundamental studies above resemble those observed in patients with corneal injury, patients who underwent refractive surgery, and patients with DED and ocular neuropathic pain (see paragraph dedicated to clinical data). One key initiative is to determine the precise underlying molecular mechanisms responsible for these morphological changes.
Recent advances in tissue clearing methods and light-sheet fluorescence microscopy have provided unprecedented access to structural and molecular information from intact tissues. A 3D high resolution imaging of an intact eyeball using a tissue clearing system derived from CLARITY and advanced light sheet microscopy in wild type fluorescently labeled transgenic mice of Prox-1-GFP or Thy1-YFP (Yang et al., 2020) was recently reported. The technology, applicable to various mouse strains (wild type or fluorescently labeled), and a spectrum of ocular components and cell types (i.e., corneal nerves, blood vessels, immune cells.) represent multiple opportunities for further architectural/morphological studies in basal and pathological ocular conditions.
Evidence for Functional Abnormalities of Corneal Nerve Terminals in Experimental Models of Corneal Injury and DED
Corneal nerve dysfunction has been reported in multiple ocular surface conditions including DED, herpetic keratitis, and after surgery (see review McKay et al., 2019). Peripheral sensitization is defined as increased responsiveness and a reduced threshold of nociceptive neurons in the periphery of the stimulation. Such phenomenon can occur in the cornea (see reviews Belmonte et al., 2004, 2017). Indeed, chronic peripheral nerve injury and local inflammation are known to participate in the development of peripheral sensitization. Immune cells locally release cytokines, chemokines, lipids, and growth factors that act on peripheral nociceptors. In turn, nociceptors actively release neuropeptides from their peripheral nerve terminals that modulate the activity of innate and adaptive immune cells (Chiu et al., 2012; Baral et al., 2019).
The development of ocular pain after corneal damage or persistent DED is believed to be due to the abnormal hyperexcitability (sensitization) of corneal nerve terminals. Single corneal nerve terminals and ciliary nerve fiber activity recordings are the two main experimental approaches currently reported to evaluate the functional changes of ongoing or evoked nerve activity in fundamental studies. Single unit extracellular recordings to evaluate polymodal and cold-sensitive corneal nerve terminals have been described using ex vivo isolated corneas in mice and in guinea pigs (Acosta et al., 2013, 2014; Kovacs et al., 2016b; Gonzalez-Gonzalez et al., 2017; Hirata et al., 2017; Alamri et al., 2018; Pina et al., 2019).
Extracellular recordings of ciliary nerve fiber activity have been described in whole eye preparations in vivo in cats (Gallar et al., 1993; Acosta et al., 2001, 2007) and ex vivo in isolated guinea pigs (McLaughlin et al., 2010; Acosta et al., 2013, 2014), rabbits (Beuerman et al., 1992), and mice (Fakih et al., 2019; Joubert et al., 2019). Ex vivo extracellular recordings of ciliary nerve activity being performed in double compartment chambers offer the possibility of applying chemical, mechanical, and thermal stimulations directly to the cornea to specifically study the responses of polymodal nociceptors, mechano-nociceptors, and cold thermoreceptors. By using this approach, a recent study showed a correlation between corneal hypersensitivity (decreased mechanical threshold), higher spontaneous ciliary nerve fiber activity, and the higher responsiveness of corneal polymodal nerve fibers from mice submitted to an acute corneal nerve injury associated with local inflammation (Joubert et al., 2019). The abnormal responses of the polymodal nociceptors (lower activation and decreased latency of the impulse discharge) observed may be a consequence of inflammation, known to play a major role in corneal nerve sensitization (McMahon and Wood, 2006; Gallar et al., 2007; Parra et al., 2014).
Another category of corneal nerve fiber changes their modalities after corneal injury and DED: cold TRPM8 corneal fibers. The contribution of TRPM8 channels to cold transduction in peripheral nerve terminals was confirmed in TRPM8-KO mice. KO animals do not show a response to noxious cold stimulus (Madrid et al., 2006). In lacrimo-deficient guinea pigs, cold nerve terminals exhibited enhanced spontaneous activity (hyperexcitability) and cold response in addition to a reduced cold threshold to cooling ramps compared to nerve terminals recorded from control animals (Kovacs et al., 2016a,b) (Figure 2).
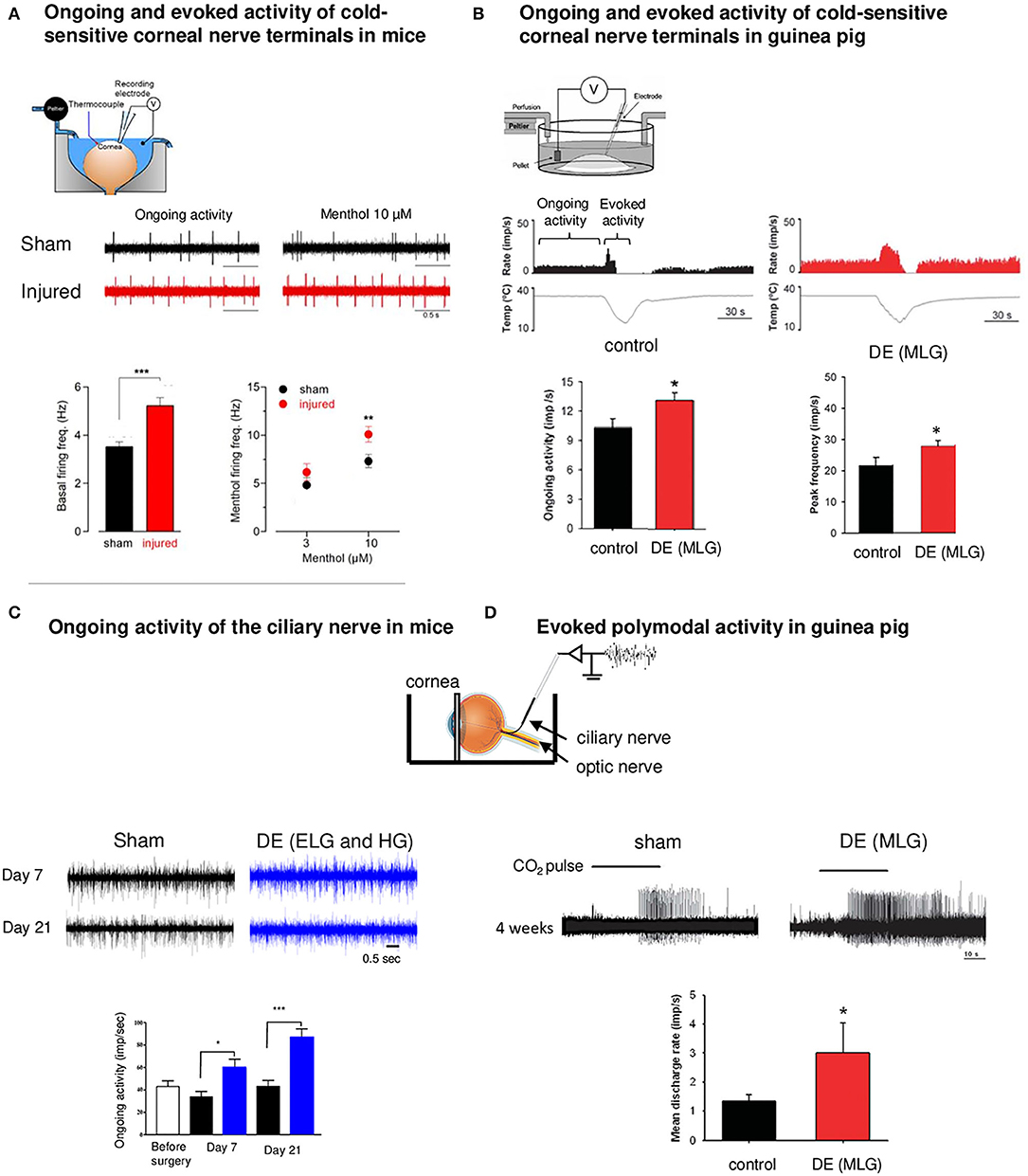
Figure 2. Functional abnormalities of corneal nerves following corneal damage and DE condition. Ex vivo electrophysiological recordings of corneal nerves in models of corneal nerve damage and DE conditions. (A) Electrophysiological set-up. Spontaneous (ongoing) and evoked (menthol 10 μM) activity of cold sensitive corneal nerve terminals in a mouse model of corneal nerve injury (red traces) relative to control (sham animals, black traces). Note the increase in corneal nerve activity for both conditions in the injured cornea. Data plotted as mean ± SEM. *P < 0.05, **P < 0.01, ***P < 0.001. (B) Electrophysiological set-up of ongoing and evoked activity of cold nerve terminals in a guinea pig model of DE obtained after the main lachrymal gland (MLG) excision. Ongoing activity and peak frequency evoked by cooling-ramp were increased 4 weeks after MLG excision. (C) Electrophysiological set-up for extracellular recording of the ciliary nerve. Ongoing ciliary nerve activity 7 days and 21 days after the excision of extraorbital (ELG) and Harderian gland (HG) in mice (blue traces) relative to sham animals (black traces). In both conditions, we noted higher ongoing (spontaneous) firing frequency in DE animals. (D) Activity of polymodal nerve fibers activated by 30 s CO2 pulse in control (sham) and MLG guinea pigs 4 weeks after surgery. The mean discharge rate during the CO2 pulse was higher after the surgery compared to sham. Note the higher responsiveness of polymodal nerve fibers (decreased latency) in DE animals. Modified from Acosta et al. (2013), Kovacs et al. (2016b), Fakih et al. (2019), and Pina et al. (2019).
The changes in corneal cold thermoreceptor firing were also observed experimentally after a peripheral corneal axotomy (Pina et al., 2019). Corneal injury evoked electrophysiological alterations in TRPM8 corneal neurons, which manifest by an enhanced sensitivity of the corneal TG neurons to cold (Pina et al., 2019). The application of menthol (10 μM), a TRPM8 agonist, induced a more pronounced increase in the firing rate in corneas of injured mice than in those of sham animals (Pina et al., 2019) (Figure 2).
Furthermore, a study reported that tear fluid hyperosmolarity (325–1,005 mOsm·kg-1), recognized as an important pathogenic factor in dry eye syndrome, increases nerve impulse activity of cold thermoreceptor endings of the cornea (Parra et al., 2014).
Thus, abnormal activity and the responsiveness of peripheral corneal cold thermoreceptors underlie the unpleasant sensations experienced by patients with DED.
In line with these observations, a severe DED induced by the excision of extraorbital lachrymal and Harderian glands in mice provoked corneal mechanical allodynia and corneal inflammation associated with an increase in the ongoing ciliary nerve fiber electrical activity compared to control mice (Fakih et al., 2019) (Figure 2). In addition, during photorefractive keratectomy, another corneal injury condition, functional and morphologic alterations in mechanical, polymodal, and cold sensory nerve fibers of the cornea have been noted, which can lead to postoperative pain (Bech et al., 2018).
In preclinical models of other corneal and conjunctival disorders characterized by an important inflammatory component, such as allergic keratoconjunctivitis (Acosta et al., 2013) or UV Keratitis (Acosta et al., 2014), an inhibition of cold receptors and sensitization of polymodal nociceptors were reported. These results contrast with the preclinical and clinical data from DED condition, in which both populations of corneal fibers (polymodal and thermoreceptors) showed sensitization (abnormal responsiveness and spontaneous firing). These aspects have been recently reviewed (Belmonte, 2019).
Nociceptive nerve terminals contain specific elements for detecting, transmitting, and modulating noxious signals (Waxman, 1999). Among them, the spike initiation zone located in the axon initial segment (AIS) corresponds to the site where action potentials are initiated and represents a critical element in neuronal excitability (Yamada and Kuba, 2016). Structural properties, such as the location relative to the soma and the length of the AIS, can change in an activity-dependent manner that can fine-tune the neuronal output properties (Jones and Svitkina, 2016; Yamazaki et al., 2017). A recent study has not only identified the precise location of the NaV-dependent spike initiation zone in nociceptive corneal nerve terminals in vivo, but has also demonstrated a plasticity in the spike initiation zone (Goldstein et al., 2019). Corneal inflammation shifts the Nav-spike initiation zone toward the terminal end, rendering it hyperexcitable in a model of inflammation-induced peripheral hyperalgesia (Goldstein et al., 2019).
Taken together, corneal nerve damage and inflammation trigger corneal sensory nerve dysfunctions (mostly sensitization); these peripheral nerve abnormalities may account for ocular pain syndromes. The above studies also reinforce the importance of neuroimmune interactions that participate in corneal hypersensitivity, inflammation, and spontaneous and evoked corneal nerve fiber hyperactivity.
Nociceptors and immune cells interact bidirectionally, a communication made possible by the wide spectrum of cells involved and by the receptors and ligands they express (Jain et al., 2020). From these multiple interactions has emerged the notion of neuroimmune interactome, a comprehensive map of the bidirectional ligand-receptor “interactome” between sensory neurons and immune cells (Jain et al., 2020). In the future, peripheral neuroimmune interactome could be constructed during pathological conditions such as corneal inflammation and corneal pain conditions. Enhancing our knowledge of the nociceptor and immune cell communication as well as the plasticity of both actors may contribute to the identification of novel therapeutic targets.
Molecular and Functional Changes in Trigeminal Corneal Neurons After Corneal Injury/Under Pathological Conditions
After peripheral nerve injury, primary sensory neurons show molecular and functional changes, resulting in hyperactivity and hyperexcitability (Berta et al., 2017). These maladaptive changes have been well-documented in dorsal root ganglion neurons in various inflammatory and pain models, but less is known about the possible alterations in the corneal nociceptive trigeminal pathways following ocular surface damage.
Anatomical studies using retrograde-labeled corneal neuron experiments have found a higher expression of TRPV1 in TRPM8+ cold-sensing corneal neurons in the TG in a model of DED, suggesting an enhanced responsiveness of TRPM8+ cells and leading to a cold allodynia (Hatta et al., 2019; Li et al., 2019). Moreover, single cell RT-PCR indicated that all TRPM8+/TRPV1+neurons express substance P (Tac1), while fewer TRPM8+/TRPV1- neurons express Tac1. It was proposed that TRPV1-dependent neuronal sensitization facilitates the release of the neuropeptide substance P from TRPM8+ cold-sensing neurons to signal nociception in response to cold (Li et al., 2019). However, such results are in contradiction with a retrograde tracing study which found that SP and TRPM8 were expressed in different TG neurons, suggesting that TRPM8+ corneal cells are non-peptidergic (He et al., 2019).
Several studies have reported changes in mRNA expression in TG following corneal damage. For example, abnormal SP and TRPM8 gene expression was found associated with corneal hypersensitivity in a model of corneal surgery (epithelium removal and one-third of the anterior stroma with a 2 mm trephine). These molecular and cellular changes occurring in the TG may contribute to the pathogenesis of corneal surgery-induced chronic pain (He et al., 2019). In a model of corneal alkali-burn injury, the increased gene expression of pro-inflammatory cytokines, tumor necrosis factor (TNF alpha), interleukin (IL)-6, SP, and its receptor NK1 (NK1) were reported in the ipsilateral TG (Ferrari et al., 2014). Changes in mRNA expression were also found in TG from an experimental model of DED obtained after 7 days of topical benzalkonium chloride (BAK). This model developed corneal inflammation and corneal hypersensitivity, and an increase in the gene expression of pro-inflammatory cytokines (IL-6 and TNF-alpha) was reported (Launay et al., 2016). Additionally, neuronal activation (FOS), neuronal injury (ATF3), astrocyte (GFAP), and oxidative (INOS2 and NOX4) markers were found to be increased in the ipsilateral TG from animals submitted to a chronic DED characterized by corneal nerve damage, inflammation, and corneal hypersensitivity (Fakih et al., 2019).
Growing evidence indicates that both CGRP and SP play a key role in the development of peripheral sensitization and are implicated in the development of neurogenic inflammation (Chiu et al., 2012), giving a rational to all changes previously described. In this line, SP has a robust effect on monocytes and macrophages and triggers their release of pro-inflammatory cytokines, including IL-1, TNF, and IL-6, via ERK/p38 MAPK-mediated NF-κB activation. Interestingly, both neuropeptides have been found to be abundant in corneal neurons, and a transcriptome signature in the TG using unbiased RNA sequencing revealed an upregulation of tachykinin precursor 1 (Tac1) that encodes SP and Calcb, which encodes CGRP after corneal injury (Pham et al., 2020).
Monitoring c-Fos, ATF3, and c Jun immunopositive cells is a commonly used approach to studying the activation and damage of primary sensory neurons from dorsal root and trigeminal ganglia. cFos, c-Jun, and ATF3 protein expressions were shown to be increased in the ipsilateral TG from animals with corneal damage (De Felipe and Belmonte, 1999; Launay et al., 2016; Fakih et al., 2019; Reaux-Le Goazigo et al., 2019). Both CGRP and ATF3 protein expressions in TG cell bodies increased after injury and returned to a normal level by 1 week, paralleling the time course of changes in nociceptive responses (Hegarty et al., 2018). These increased expressions of FOS, CGRP, and ATF3 in primary sensory neurons may contribute to the activation of central pain pathways in response to sustained ocular stimulation, leading to the centralization of pain (Levine et al., 1993).
Under local inflammation or after tissue damage, cytokines, prostaglandins, nerve growth factor, and bradykinin signals increase TRPV1 expression and/or TRPV1 activity in sensory neurons (Pinho-Ribeiro et al., 2017). In the context of DED, TRPV1 protein levels have been found to increase in the ipsilateral TG, and this TRPV1 upregulation was associated with enhanced eye wipe behavior after hypertonic saline and capsaicin instillation in a rat model for aqueous tear-deficient DE (Bereiter et al., 2018).
In the same way, it has been observed that capsazepine, a TRPV-1 antagonist, prevented dry eye sensitization of cool cells to capsaicin (Hatta et al., 2019) and reduced polymodal responsiveness to acidic stimulation in an allergic eye model (Acosta et al., 2013). Moreover, TRPV-1 pharmacological blockade decreases SP release in cold allodynia (Li et al., 2019).
Neurons in sensory dorsal and trigeminal ganglia are surrounded by satellite glial cells (SGCs) (Hanani, 2005). Activation of SGCs is characterized by GFAP upregulation in the injured trigeminal nerve branch associated with the development of hyperalgesia (Vit et al., 2006; Katagiri et al., 2012), but this has not been observed under non-pathological conditions (Shinoda et al., 2019). Interestingly, increased spontaneous pain behavior and corneal allodynia have been associated with morphological changes (hypertrophy) and upregulation of GFAP protein expression in the SGCs at the level of the ipsilateral trigeminal nerve V1 branch in preclinical models of corneal injury (Launay et al., 2016; Fakih et al., 2019; Reaux-Le Goazigo et al., 2019).
In addition, accumulative evidence reports that immune cells as monocytes/macrophages infiltrate into the TG and become activated, following orofacial pathologies, including peripheral trigeminal nerve trauma and orofacial inflammation (Iwata and Shinoda, 2019). Fundamental studies performed in mice reported that corneal injury induced an increase and activated resident and proliferated macrophages in mouse TGs (Ferrari et al., 2014; Launay et al., 2016; Fakih et al., 2019; Reaux-Le Goazigo et al., 2019). The infiltrated monocytes/macrophages show larger soma and thicker ramifications; such morphological structural changes indicate their activation (Shinoda et al., 2019). The above observations suggest that immune cells and resident glia in the TG could play a significant role in the modulation of ocular pain. The specific role of both the population of cells should be better investigated in the future in the context of ocular pain.
Though RTqPCR and immunohistochemistry have provided some important information about neuronal, glial activations and cellular morphological abnormalities following corneal pain conditions, the major limitation of these techniques is that they lack the ability to monitor the dynamic of these neuronal changes. Functional studies have been performed in trigeminal neurons to better understand their activation and how various corneal nerve injuries may alter their modalities. In vivo electrophysiology works have assessed action potentials from single neurons and/or clusters of neurons in rodent TGs (Lopez de Armentia et al., 2000; Veiga Moreira et al., 2007; Hirata and Meng, 2010; Kurose and Meng, 2013; Hirata et al., 2015; Quallo et al., 2015) under normal and pathological conditions. The literature is not as rich as the one from DRG neurons, but it provides important information about changes in the neuronal properties of corneal neurons under physiological and DE conditions.
Understanding the effect of ocular surface damage on corneal nociceptive neurons is crucial for developing a potential antalgic treatment against corneal pain. To this aim, several studies based on single unit recordings of corneal nociceptive neurons located in the TG have been assessed in preclinical models of DED and corneal injury to provide valuable insight into the normal and pathologic response of the corneal neurons. These studies highlighted the relationship between the electrophysiological signature (both fiber conduction velocity and electrical properties) and the altered responsiveness to various sensory modalities (chemical, thermal, and mechanical) of afferents innervating the cornea.
Thus, in vivo extracellular electrophysiological recordings performed in rat TG single neurons that innervated the cornea, before, and up to 3 h after, the ocular application of continuous hyperosmolar tears demonstrated that dry responses of corneal dry-sensitive neurons were depressed or even completely abolished by the hyperosmolarity of tears (Hirata et al., 2015).
In a moderate dry eye experimental model obtained after unilateral extraorbital lacrimal gland excision, Kurose et al. demonstrated by a single unit extracellular recording of trigeminal neurons (8–10 weeks after the surgery) that dry eye sensitized corneal cool cells to the TRPM8 agonist menthol and to cool stimulation (Kurose and Meng, 2013). In the same experimental model, Hatta et al. showed that dry eye increased responsiveness to noxious heat and activation by capsaicin through TRPV1 (Hatta et al., 2019) (Figure 3). Pina et al. further demonstrated, in a mouse model of corneal injury, changes in cold sensitivity of corneal neurons in the TG through an enhanced functional expression of TRPM8 channels, suggesting that the increase in ocular dryness sensation and basal tearing rate observed after refractive surgery could be related to a disturbance in the TRPM8 responsiveness of cold sensory neurons (Kovacs et al., 2016b; Pina et al., 2019) (Figure 3).
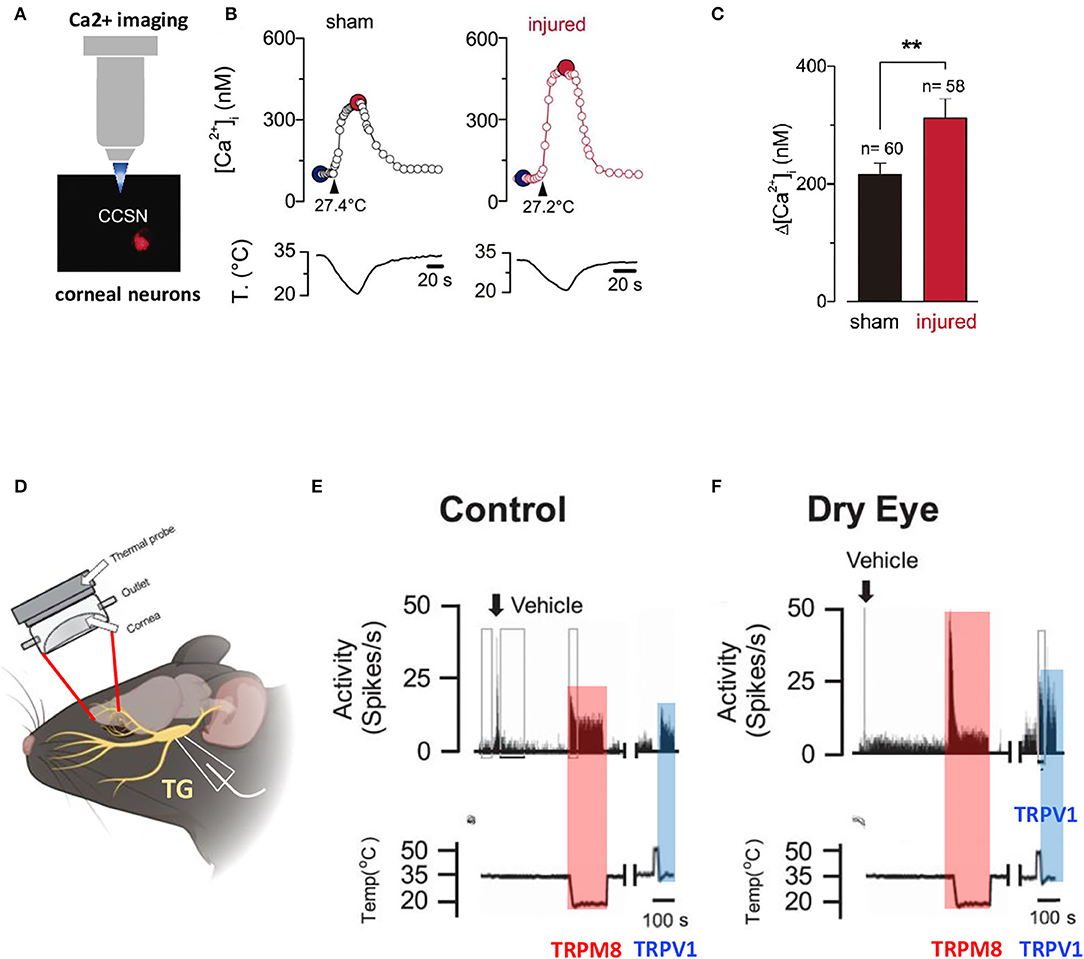
Figure 3. Altered responsiveness of corneal neurons following peripheral nerve damage and after lacrimal gland excision. (A) Altered cold sensitivity of corneal neurons induced by peripheral nerve damage. Calcium imaging on FM 1–43 labeled corneal neurons in culture. (B) Cold-evoked [Ca2+]i response to a temperature ramp of two representative corneal neurons from sham (black trace) and injured (red trace) mice. Large dots indicate the basal (blue) and maximal (red) [Ca2+]i during the cold stimulus. (C) Bar graphs of the mean amplitude of [Ca2+]i rises evoked by cold stimulation in the corneal neurons from sham and injured mice. Modified from Pina et al. (2019). Data plotted as mean ± SEM. **p = 0.0014. (D) Illustration of the custom-made chamber designed with inflow and outflow portals for maintaining a constant environment on the surface of the cornea while recording from neuronal cell bodies located in the TG. (E,F) Effects of vehicle and cool and heat-evoked activity in control (E) and lacrimal gland excision (LGE) animals (F). Neuronal activity recorded from control animals and animals 2 weeks after LGE, following vehicle, cold stimulation (TRPM8 activation), and 3 μM capsaicin (TRPV1 activation) applications to the cornea. Copyright © 2019 the American Physiological Society (Hatta et al., 2019).
Evidence that Corneal Nerve Abnormalities Shape the Central Nervous System in Rodents
Corneal nociceptors, like other primary somatosensory neurons, are pseudo unipolar. They send a peripheral axon to innervate the corneal and a central axon to synapse on second-order neurons at two different locations of the brain stem nuclear complex: the Vi/Vc (trigeminal subnucleus interpolaris/caudalis transition region) and Vc/C1 (caudalis/upper cervical cord junction) areas of the trigeminal subnucleus caudalis region (Figure 1). Persistent ongoing activity in primary nociceptors may generate extensive changes in central pain processing-related structures, leading to maladaptive neuroplasticity. In addition, proinflammatory mediators participate not only in the sensitization of peripheral nerve terminals but also in the transfer of nociceptive information from the periphery to the central nervous system (Grace et al., 2014; Melik Parsadaniantz et al., 2015). More specifically, activated glial cells, which produce various proinflammatory cytokines (TNF alpha, IL-6, IL1 beta), neurotrophic factors, and chemokines, contribute to neuronal excitability and a central sensitization mechanism under trigeminal pain states (Iwata et al., 2011, 2017; Melik Parsadaniantz et al., 2015; Goto et al., 2016).
Recent studies have shown that acute (Launay et al., 2016) and persistent (Fakih et al., 2019) ocular pain conditions induce a higher density of Iba1-immunopositive microglial cells and higher levels of CD68 and ITGAM genes in the ipsilateral trigeminal brainstem sensory complex. Aside from immune cell activation, increased levels of the GFAP protein and gene expression, astrocyte marker in the brain, were reported in this central structure. Moreover, the upregulation of pro-inflammatory markers IL-6 and IL-1β, INOS2 genes, and ATF3 and FOS markers in the trigeminal brainstem sensory complex (TBSC) was associated with the development of corneal hypersensitivity (Fakih et al., 2019) (Figure 4). These findings strongly suggest that neuronal–glial and neuroinflammatory interactions occur after corneal nerve injury in the central nervous system, which may account for the development and persistence of the ocular pain reported in DED patients.
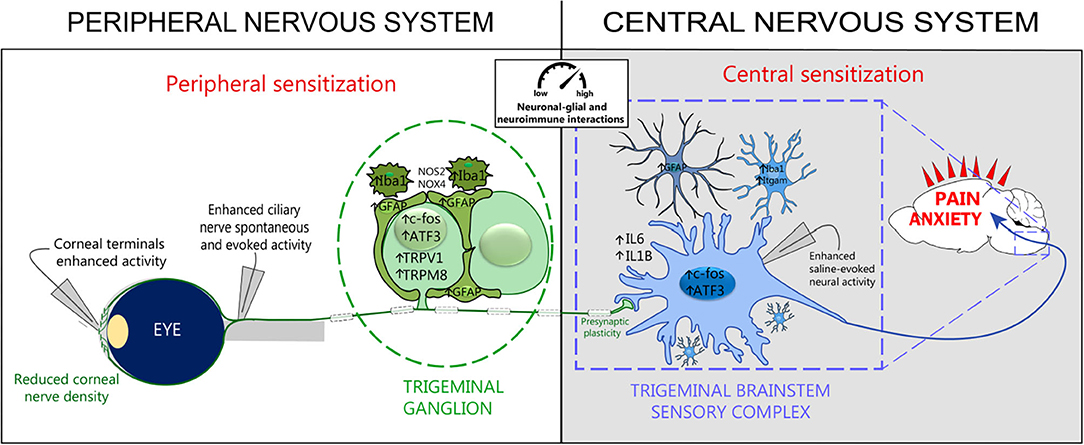
Figure 4. Peripheral and central sensitization associated with persistent dry eye pain. Experimental persistent dry eye triggers corneal nerve abnormalities, neuroimmune interactions, and peripheral and central sensitization. Electrophysiological recordings reveal increased activity in corneal terminals, ciliary nerves, and trigeminal brainstem sensory complex neurons. Histological and molecular studies show neuronal activation (c-Fos) and injury (ATF3), de novo channel production (TRPV1, TRPM8), glial activation (Iba1, GFAP, Itgam), oxidative stress (NOS2, NOX4), proinflammatory responses (IL6, IL1B), and synaptic-enhancer plasticity at the level of TG and TBSC. These cellular and molecular changes lead to pain and promote anxiety-like behaviors. Modified from Guerrero-Moreno et al. (2021).
Persistent ongoing activity in primary nociceptors may also lead to central sensitization, which is defined as “an amplification of neural signaling within the central nervous system that elicits pain hypersensitivity,” (Woolf, 2011) and a functional remodeling of presynaptic sites. Piccolo, one of the components of the presynaptic zone, plays a key role in synaptic plasticity by facilitating/managing the secretion of synaptic vesicles, and increased expression was reported in an orofacial pain model (Thibault et al., 2016). The upregulated expression of Piccolo in the brainstem in mice with persistent ocular pain associated with DED indicated that persistent ongoing corneal nociceptor activity induced profound synaptic reorganization, which may contribute to the chronicity of ocular pain. These central cellular rearrangements (neuroinflammation, astrocytes and microglial activation, and enhancements of excitatory synaptic transmission protein) are in line with the reported sensitization of ocular-responsive neurons of the caudal trigeminal brainstem in persistent tear deficiency in rats (Rahman et al., 2015) (Figure 4). These neuroplastic changes observed in the TG and TBSC under corneal pain are likely to play a role in the establishment and maintenance of central sensitization that is seen in many centralized pain disorders like fibromyalgia and migraine.
There is also a wealth of evidence to show that contralateral structural and molecular changes can occur both in the periphery and the central nervous system in response to a unilateral insult (Koltzenburg et al., 1999; Shenker et al., 2003; Ferrari et al., 2014; Lee et al., 2019).
For example, unilateral corneal damage induces inflammatory responses in contralateral mouse eyes (Yamaguchi et al., 2016; Lee et al., 2019), the TG (Ferrari et al., 2014), and the central nervous system (Launay et al., 2016). Indeed, FOS-like positive neurons were seen in the contralateral trigeminal brainstem after corneal nerve damage (Launay et al., 2016). Contralateral responses were demonstrable not only in rodents, but also in the human eye (Hamrah et al., 2010, 2013; Postole et al., 2016; Yamaguchi et al., 2016), and there are several reasons supporting that these contralateral responses are mediated through neural mechanisms rather than reflecting a systemic effect (Shenker et al., 2003; Gong et al., 2020).
Increased corneal peripheral nociceptive input may result in neuronal activity within the higher-order neurons within the brain, particularly in regions associated with the pain matrix (the thalamus, insula, anterior cingulate cortex, prefrontal cortex, and somatosensory cortex). Only one study has been reported to date in this area of investigation. It showed in a mouse model of corneal alkali burn that corneal spontaneous pain activated the neuropathic central pain matrix (Xiang et al., 2017). Increased phospho-ERK positive neurons were detected in the subnucleus caudalis/upper cervical cord (Vc/V1), the insular cortex, the anterior cingulated cortex, and the rostroventral medulla. Many experiments are still needed to precisely determine the nature of central structures that are recruited during acute and persistent ocular pain.
Clinical Evidence of Corneal Nerve Abnormalities and Dysfunctions
As previously stated, the new definition of DED includes somatosensory abnormalities as a core mechanism (Craig et al., 2017). The measurement of corneal sensitivity in patients could provide the first evidence for somatosensory abnormalities, but practical tools are still lacking to assess hypersensitivity and hyperexcitability, as well as explore other stimuli than simple mechanical responses. The persistence of ocular pain in a subset of patients with dry eye syndromes is a major challenge in the management of ocular pain (Rosenthal and Borsook, 2016; Mehra et al., 2020). It is therefore crucial to identify diagnostic modalities that can accurately predict or identify neuropathic pain.
Corneal sensitivity can be assessed by the Belmonte non-contact gas and the Cochet-Bonnet esthesiometers. The Belmonte non-contact gas esthesiometer allows one to measure corneal sensory abnormalities following mechanical, thermal, and chemical corneal stimulations, i.e., the detection of polymodal function for both A delta and C fibers.
Patients with dry eye exhibit corneal hypoesthesia after mechanical, thermal, and chemical stimulation, and this condition seems to be related to damage to the corneal sensory innervation (Bourcier et al., 2005). Some studies in patients with dry eye or neuropathic pain symptoms have found reduced corneal sensitivity to mechanical, thermal, and chemical stimuli compared to controls (Bourcier et al., 2005), while others have found increased mechanical sensitivity (Spierer et al., 2016). Unfortunately, complex sensing esthesiometers are only scarcely available and are not routine devices, despite their high potential for exploring DED and corneal sensitivity.
The Cochet-Bonnet contact esthesiometer is another device that has been widely used to characterize somatosensory disturbances within the human eye and more specifically for evaluations of mechanical nociceptor (A delta fibers) responses. Some studies have reported a reduced sensitivity to mechanical stimuli in DE patients (Adatia et al., 2004; Labbe et al., 2012) exhibiting a decreased corneal nerve density.
The maladaptive processes responsible for corneal pain can take place in either the peripheral or the central nervous system. Functional somatosensory testing, also known as the proparacaine challenge test, is useful for assessing the peripheral or central location of pain generators in patients. This functional test consists of a topical instillation of 0.5% proparacaine (an anesthetic commonly used in ophthalmology) and determines whether pain is abolished or not. The persistence of ocular pain suggests a central location, whereas a peripheral origin is confirmed by pain relief (Goyal and Hamrah, 2016; Crane et al., 2017). Patients with peripheral neuropathic pain would be the principal beneficiaries of topical painkillers, while systemic approaches would be preferred for patients with central neuropathic pain.
A functional assessment of corneal sensitivity is generally combined with a morphological analysis of the corneal nerve. IVCM is a non-invasive technique providing real-time, high-resolution corneal imaging in both humans and laboratory animals. IVCM studies allow for the longitudinal imaging and quantification of cellular changes in, e.g., dendritic and keratocyte cells, and of sub-basal nerve plexus morphology in corneas over time. Excellent reviews have described the broad changes in the sub-basal nerve layer by using IVCM imaging in healthy controls and patients (Cruzat et al., 2017; Al-Aqaba et al., 2019; Labetoulle et al., 2019). Quantification of IVCM images have evidenced that sub-basal nerves undergo a significant decrease in the number and density in patients with dry eye symptoms compared to healthy subjects (Bourcier et al., 2005; Benitez-Del-Castillo et al., 2007; Labbe et al., 2012, 2013; Levy et al., 2017; Nicolle et al., 2018) (Figure 5). Other morphological corneal nerve abnormalities have been observed in DE patients, such as nerve sprouting, increased thickness, tortuosity, and beading (for review Galor et al., 2018). Recent studies in patients with neuropathic corneal pain have also demonstrated decreased corneal nerve density associated with allodynia (Hamrah et al., 2017), photoallodynia (Aggarwal et al., 2015), and post-LASIK neuralgia (Theophanous et al., 2015). Those corneal nerve changes can be quantified automatically by emerging specialized software, representing a very promising tool for clinical assessments (Annunziata et al., 2016; Giannaccare et al., 2019).
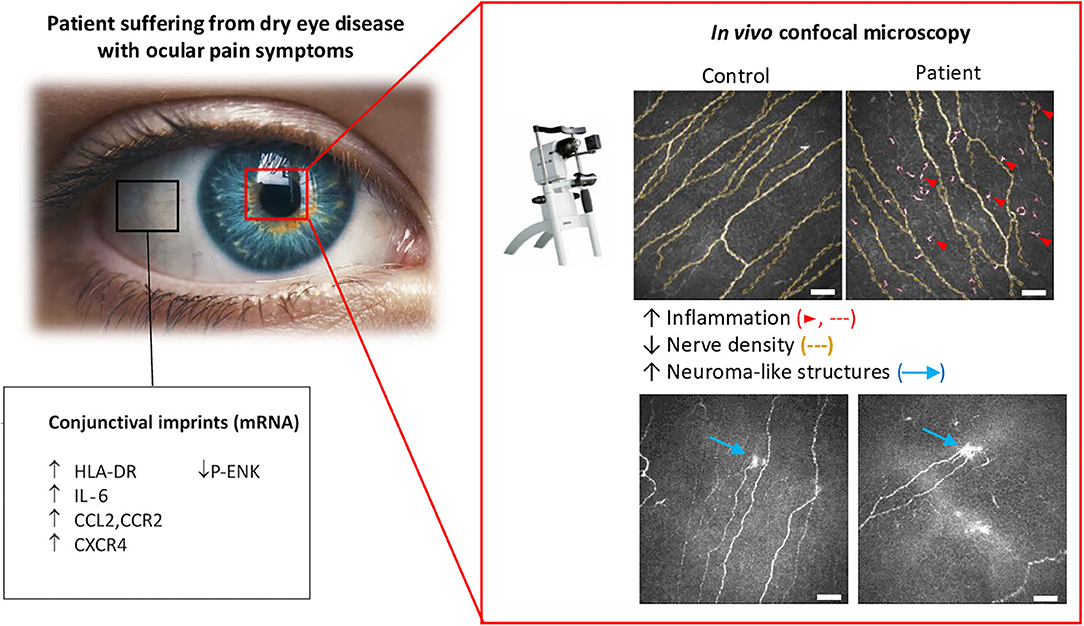
Figure 5. Morphological and molecular changes in the ocular surface from dry eye patients with persistent ocular pain. Conjunctival imprints show higher levels of the proinflammatory markers, HLA-DR, IL-6, and CCL2, and of the chemokine receptors, CCR2 and CXCR4. In contrast, a decrease in the enkephalin precursor P-ENK was found in DE patients. Corneal IVCM imaging showed a decrease in nerve density, which was accompanied by a higher density of hyperreflective (putative inflammatory) cells and the presence of microneuroma-like structures in DED patients with pain syndromes. Modified from Nicolle et al. (2018).
Chronic pain induced after nerve injury is generally associated with prominent morphological and epigenetic changes within the afferent fibers. After nerve damage, destroyed peripheral nerves start to regenerate and form neuromas that exhibit abnormal responsiveness and spontaneous discharges (ectopic firing) (Fawcett and Keynes, 1990). These functional changes are the consequence of altered expressions of ion channel proteins in the soma and in regenerating nerve terminals. Neuroma structures have emerged as other important morphological changes in corneal nerves and have been suggested as clinical imagining biomarkers (hallmark) of corneal neuropathic pain (Aggarwal et al., 2019; Bayraktutar et al., 2020; Moein et al., 2020). Taken together, these studies suggest that a corneal neuroma may represent an important pathological feature of peripheral nerve abnormalities in patients with ocular pain. Interestingly, topical treatment with autologous serum tears reduced corneal nerve abnormalities, improved corneal nerve regeneration, and alleviated corneal pain (Aggarwal et al., 2019). Future directions are needed to determine when and how the morphological changes occur on the ocular surface and which subpopulations of corneal neurons are hit.
In recent years, clinical research regarding neuroimmune crosstalk has focused increasingly on the neuroimmune interactions in the cornea. Among the resident immune cells, epithelial dendritic cells predominantly reside in the basal epithelium of the human cornea. These resident corneal dendritic cells play a key role in maintaining the homeostasis of corneal nerves (Gao et al., 2016). However, corneal damage or corneal nerve abnormalities can disrupt the balance between immune cells and peripheral nerves. Accumulative evidence has revealed, in the cornea from patients with DED and corneal neuropathic pain, increased activated dendritic cells and a close anatomical proximity between peripheral nerves and immune cells (Shetty et al., 2016; Tepelus et al., 2017; Aggarwal et al., 2020).
As stated before, fundamental and clinical studies have well-demonstrated that repeated damage to the ocular surface and corneal nerves per se can cause peripheral and central sensitization mechanisms, explaining the centralized pain in some patients with corneal neuropathic pain. However, the neuronal circuits participating in corneal pain are not fully understood. Over the past decade, brain imaging investigations have shed light on neural correlations with pain perception and modulation. New developments in structural, functional, and neurochemical imaging, such as resting-state connectivity, functional magnetic resonance imaging (fMRI), and γ-aminobutyric acid spectroscopy, have shed light on persistent non-ocular pain (Grachev et al., 2000; Harris and Clauw, 2012; Legarreta et al., 2020).
To date, only one case study from Moulton et al. (2012) has reported by means of fMRI the nature of hemodynamic responses evoked by corneal pain in the human brain. fMRI imaging revealed an activation of the contralateral somatosensory cortex, ventral posteromedial thalamus, and the anterior cingulated cortex during the painful state.
Concluding Remarks and Future Perspectives
The cornea, the most densely innervated tissue in the human body, offers multiple advantages in fundamental and clinical research, as it is an excellent model to study the function and morphology of nociceptive nerve fibers, peripheral nerve damage, nerve regeneration, and sensory abnormalities. Accumulative clinical and animal model studies have identified morphological and functional abnormalities of corneal nerve terminals associated with ocular surface diseases. The last decade has also seen a rapid expansion in our knowledge regarding the nature of the cellular and molecular mechanisms, as well as the neuroimmune interactions that take place in the cornea, the TG, which contains the primary sensory neurons, and higher central structures. However, many questions remain unanswered. Among them, unraveling neuroimmune crosstalk mechanisms leading to inflammation and pain would bring a more comprehensive picture of the entire neuroinflammatory process and represents an exciting and expanding research domain. In the future, there is also an urgent need to elucidate the time window for restoring corneal nerve morphological and functional abnormalities and to identify a more specific target and their signaling pathways in corneal nociceptors, which may offer alternative treatments.
Finally, recent advances in in vivo functional imaging technology, together with automatic frameworks for clinical assessment, bi- and three-photo microscopy imaging, viral-mediated optogenetic control of peripheral nerve expressing light sensitive opsins, chemogenetics, and the development of simple or dual knockout mice, may help in solving the puzzle of these maladaptive changes in the corneal nociceptive pathways.
Author Contributions
AG-M and AR-L equally contributed to drafting the manuscript and the figures. AR-L conceptualized the review. AG-M and AR-L wrote the manuscript. AG-M, AR-L, CB, and SMP contributed to the article and approved the manuscript.
Funding
This work was supported by Sorbonne Université and the Institut National de la Santé et de la Recherche Médicale, ANR, LabEx LIFESENSES (ANR-10-LABX-65), and IHU FOReSIGHT (ANR-18-IAHU-01). AG-M was funded by a H2020-MSCA-ETN program (IT-DED3) (Grant Agreement 765608).
Conflict of Interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
References
Acosta, M. C., Belmonte, C., and Gallar, J. (2001). Sensory experiences in humans and single-unit activity in cats evoked by polymodal stimulation of the cornea. J. Physiol. 534, 511–525. doi: 10.1111/j.1469-7793.2001.t01-1-00511.x
Acosta, M. C., Luna, C., Graff, G., Meseguer, V. M., Viana, F., Gallar, J., et al. (2007). Comparative effects of the nonsteroidal anti-inflammatory drug nepafenac on corneal sensory nerve fibers responding to chemical irritation. Invest. Ophthalmol. Vis. Sci. 48, 182–188. doi: 10.1167/iovs.06-0710
Acosta, M. C., Luna, C., Quirce, S., Belmonte, C., and Gallar, J. (2013). Changes in sensory activity of ocular surface sensory nerves during allergic keratoconjunctivitis. Pain 154, 2353–2362. doi: 10.1016/j.pain.2013.07.012
Acosta, M. C., Luna, C., Quirce, S., Belmonte, C., and Gallar, J. (2014). Corneal sensory nerve activity in an experimental model of UV keratitis. Invest. Ophthalmol. Vis. Sci. 55, 3403–3412. doi: 10.1167/iovs.13-13774
Adatia, F. A., Michaeli-Cohen, A., Naor, J., Caffery, B., Bookman, A., and Slomovic, A. (2004). Correlation between corneal sensitivity, subjective dry eye symptoms and corneal staining in Sjogren's syndrome. Can. J. Ophthalmol. 39, 767–771. doi: 10.1016/S0008-4182(04)80071-1
Aggarwal, S., Colon, C., Kheirkhah, A., and Hamrah, P. (2019). Efficacy of autologous serum tears for treatment of neuropathic corneal pain. Ocul. Surf. 17, 532–539. doi: 10.1016/j.jtos.2019.01.009
Aggarwal, S., Kheirkhah, A., Cavalcanti, B. M., Cruzat, A., Colon, C., Brown, E., et al. (2015). Autologous serum tears for treatment of photoallodynia in patients with corneal neuropathy: efficacy and evaluation with in vivo confocal microscopy. Ocul. Surf. 13, 250–262. doi: 10.1016/j.jtos.2015.01.005
Aggarwal, S., Kheirkhah, A., Cavalcanti, B. M., Cruzat, A., Jamali, A., and Hamrah, P. (2020). Correlation of corneal immune cell changes with clinical severity in dry eye disease: An in vivo confocal microscopy study. Ocul. Surf. doi: 10.1016/j.jtos.2020.05.012. [Epub ahead of print].
Alamri, A., Bron, R., Brock, J. A., and Ivanusic, J. J. (2015). Transient receptor potential cation channel subfamily V member 1 expressing corneal sensory neurons can be subdivided into at least three subpopulations. Front. Neuroanat. 9:71. doi: 10.3389/fnana.2015.00071
Alamri, A. S., Wood, R. J., Ivanusic, J. J., and Brock, J. A. (2018). The neurochemistry and morphology of functionally identified corneal polymodal nociceptors and cold thermoreceptors. PLoS ONE 13:e0195108. doi: 10.1371/journal.pone.0195108
Al-Aqaba, M. A., Dhillon, V. K., Mohammed, I., Said, D. G., and Dua, H. S. (2019). Corneal nerves in health and disease. Prog. Retin. Eye Res. 73:100762. doi: 10.1016/j.preteyeres.2019.05.003
Alcalde, I., Inigo-Portugues, A., Gonzalez-Gonzalez, O., Almaraz, L., Artime, E., Morenilla-Palao, C., et al. (2018). Morphological and functional changes in TRPM8-expressing corneal cold thermoreceptor neurons during aging and their impact on tearing in mice. J. Comp. Neurol. 526, 1859–1874. doi: 10.1002/cne.24454
Annunziata, R., Kheirkhah, A., Aggarwal, S., Hamrah, P., and Trucco, E. (2016). A fully automated tortuosity quantification system with application to corneal nerve fibres in confocal microscopy images. Med. Image Anal. 32, 216–232. doi: 10.1016/j.media.2016.04.006
Baral, P., Udit, S., and Chiu, I. M. (2019). Pain and immunity: implications for host defence. Nat. Rev. Immunol. 19, 433–447. doi: 10.1038/s41577-019-0147-2
Bayraktutar, B. N., Ozmen, M. C., Muzaaya, N., Dieckmann, G., Koseoglu, N. D., Muller, R. T., et al. (2020). Comparison of clinical characteristics of post-refractive surgery-related and post-herpetic neuropathic corneal pain. Ocul. Surf. 18, 641–650. doi: 10.1016/j.jtos.2020.07.006
Bech, F., Gonzalez-Gonzalez, O., Artime, E., Serrano, J., Alcalde, I., Gallar, J., et al. (2018). Functional and morphologic alterations in mechanical, polymodal, and cold sensory nerve fibers of the cornea following photorefractive keratectomy. Invest. Ophthalmol. Vis. Sci. 59, 2281–2292. doi: 10.1167/iovs.18-24007
Belmonte, C. (2019). Pain, dryness, and itch sensations in eye surface disorders are defined by a balance between inflammation and sensory nerve injury. Cornea 38(Suppl 1), S11–S24. doi: 10.1097/ICO.0000000000002116
Belmonte, C., Acosta, M. C., and Gallar, J. (2004). Neural basis of sensation in intact and injured corneas. Exp. Eye Res. 78, 513–525. doi: 10.1016/j.exer.2003.09.023
Belmonte, C., Acosta, M. C., Merayo-Lloves, J., and Gallar, J. (2015). What causes eye pain? Curr. Ophthalmol. Rep. 3, 111–121. doi: 10.1007/s40135-015-0073-9
Belmonte, C., Gallar, J., Pozo, M. A., and Rebollo, I. (1991). Excitation by irritant chemical substances of sensory afferent units in the cat's cornea. J. Physiol. 437, 709–725. doi: 10.1113/jphysiol.1991.sp018621
Belmonte, C., Nichols, J. J., Cox, S. M., Brock, J. A., Begley, C. G., Bereiter, D. A., et al. (2017). TFOS DEWS II pain and sensation report. Ocul. Surf. 15, 404–437. doi: 10.1016/j.jtos.2017.05.002
Benitez-Del-Castillo, J. M., Acosta, M. C., Wassfi, M. A., Diaz-Valle, D., Gegundez, J. A., Fernandez, C., et al. (2007). Relation between corneal innervation with confocal microscopy and corneal sensitivity with noncontact esthesiometry in patients with dry eye. Invest. Ophthalmol. Vis. Sci. 48, 173–181. doi: 10.1167/iovs.06-0127
Bereiter, D. A., Rahman, M., Thompson, R., Stephenson, P., and Saito, H. (2018). TRPV1 and TRPM8 channels and nocifensive behavior in a rat model for dry eye. Invest. Ophthalmol. Vis. Sci. 59, 3739–3746. doi: 10.1167/iovs.18-24304
Berta, T., Qadri, Y., Tan, P. H., and Ji, R. R. (2017). Targeting dorsal root ganglia and primary sensory neurons for the treatment of chronic pain. Expert Opin. Ther. Targets 21, 695–703. doi: 10.1080/14728222.2017.1328057
Beuerman, R. W., Snow, A., Thompson, H., and Stern, M. (1992). Action potential response of the corneal nerves to irritants. Lens Eye Toxic. Res. 9, 193–210.
Bouheraoua, N., Fouquet, S., Marcos-Almaraz, M. T., Karagogeos, D., Laroche, L., and Chedotal, A. (2019). Genetic analysis of the organization, development, and plasticity of corneal innervation in mice. J. Neurosci. 39, 1150–1168. doi: 10.1523/JNEUROSCI.1401-18.2018
Bourcier, T., Acosta, M. C., Borderie, V., Borras, F., Gallar, J., Bury, T., et al. (2005). Decreased corneal sensitivity in patients with dry eye. Invest. Ophthalmol. Vis. Sci. 46, 2341–2345. doi: 10.1167/iovs.04-1426
Bron, R., Wood, R. J., Brock, J. A., and Ivanusic, J. J. (2014). Piezo2 expression in corneal afferent neurons. J. Comp. Neurol. 522, 2967–2979. doi: 10.1002/cne.23560
Carr, R. W., Pianova, S., Fernandez, J., Fallon, J. B., Belmonte, C., and Brock, J. A. (2003). Effects of heating and cooling on nerve terminal impulses recorded from cold-sensitive receptors in the guinea-pig cornea. J. Gen. Physiol. 121, 427–439. doi: 10.1085/jgp.200308814
Chaudhary, S., Namavari, A., Yco, L., Chang, J. H., Sonawane, S., Khanolkar, V., et al. (2012). Neurotrophins and nerve regeneration-associated genes are expressed in the cornea after lamellar flap surgery. Cornea 31, 1460–1467. doi: 10.1097/ICO.0b013e318247b60e
Chen, X., Gallar, J., and Belmonte, C. (1997). Reduction by antiinflammatory drugs of the response of corneal sensory nerve fibers to chemical irritation. Invest. Ophthalmol. Vis. Sci. 38, 1944–1953.
Chiu, I. M., von Hehn, C. A., and Woolf, C. J. (2012). Neurogenic inflammation and the peripheral nervous system in host defense and immunopathology. Nat. Neurosci. 15, 1063–1067. doi: 10.1038/nn.3144
Cho, J., Bell, N., Botzet, G., Vora, P., Fowler, B. J., Donahue, R., et al. (2019). Latent sensitization in a mouse model of ocular neuropathic pain. Transl. Vis. Sci. Technol. 8:6. doi: 10.1167/tvst.8.2.6
Coste, B., Mathur, J., Schmidt, M., Earley, T. J., Ranade, S., Petrus, M. J., et al. (2010). Piezo1 and Piezo2 are essential components of distinct mechanically activated cation channels. Science 330, 55–60. doi: 10.1126/science.1193270
Craig, J. P., Nichols, K. K., Akpek, E. K., Caffery, B., Dua, H. S., Joo, C. K., et al. (2017). TFOS DEWS II definition and classification report. Ocul. Surf. 15, 276–283. doi: 10.1016/j.jtos.2017.05.008
Crane, A. M., Feuer, W., Felix, E. R., Levitt, R. C., McClellan, A. L., Sarantopoulos, K. D., et al. (2017). Evidence of central sensitisation in those with dry eye symptoms and neuropathic-like ocular pain complaints: incomplete response to topical anaesthesia and generalised heightened sensitivity to evoked pain. Br. J. Ophthalmol. 101, 1238–1243. doi: 10.1136/bjophthalmol-2016-309658
Cruzat, A., Qazi, Y., and Hamrah, P. (2017). In vivo confocal microscopy of corneal nerves in health and disease. Ocul. Surf. 15, 15–47. doi: 10.1016/j.jtos.2016.09.004
De Felipe, C., and Belmonte, C. (1999). c-Jun expression after axotomy of corneal trigeminal ganglion neurons is dependent on the site of injury. Eur. J. Neurosci. 11, 899–906. doi: 10.1046/j.1460-9568.1999.00498.x
Dieckmann, G., Goyal, S., and Hamrah, P. (2017). Neuropathic corneal pain: approaches for management. Ophthalmology 124, S34–S47. doi: 10.1016/j.ophtha.2017.08.004
Fakih, D., Zhao, Z., Nicolle, P., Reboussin, E., Joubert, F., Luzu, J., et al. (2019). Chronic dry eye induced corneal hypersensitivity, neuroinflammatory responses, and synaptic plasticity in the mouse trigeminal brainstem. J. Neuroinflamm. 16:268. doi: 10.1186/s12974-019-1656-4
Fawcett, J. W., and Keynes, R. J. (1990). Peripheral nerve regeneration. Annu. Rev. Neurosci. 13, 43–60. doi: 10.1146/annurev.ne.13.030190.000355
Fernandez-Trillo, J., Florez-Paz, D., Inigo-Portugues, A., Gonzalez-Gonzalez, O., Del Campo, A. G., Gonzalez, A., et al. (2020). Piezo2 mediates low-threshold mechanically-evoked pain in the cornea. J. Neurosci. 40, 8976–8993. doi: 10.1523/JNEUROSCI.0247-20.2020
Ferrari, G., Bignami, F., Giacomini, C., Capitolo, E., Comi, G., Chaabane, L., et al. (2014). Ocular surface injury induces inflammation in the brain: in vivo and ex vivo evidence of a corneal-trigeminal axis. Invest. Ophthalmol. Vis. Sci. 55, 6289–6300. doi: 10.1167/iovs.14-13984
Gallar, J., Acosta, M. C., Gutierrez, A. R., and Belmonte, C. (2007). Impulse activity in corneal sensory nerve fibers after photorefractive keratectomy. Invest. Ophthalmol. Vis. Sci. 48, 4033–4037. doi: 10.1167/iovs.07-0012
Gallar, J., Pozo, M. A., Tuckett, R. P., and Belmonte, C. (1993). Response of sensory units with unmyelinated fibres to mechanical, thermal and chemical stimulation of the cat's cornea. J. Physiol. 468, 609–622. doi: 10.1113/jphysiol.1993.sp019791
Galor, A., Moein, H. R., Lee, C., Rodriguez, A., Felix, E. R., Sarantopoulos, K. D., et al. (2018). Neuropathic pain and dry eye. Ocul. Surf. 16, 31–44. doi: 10.1016/j.jtos.2017.10.001
Gao, N., Lee, P., and Yu, F. S. (2016). Intraepithelial dendritic cells and sensory nerves are structurally associated and functional interdependent in the cornea. Sci. Rep. 6:36414. doi: 10.1038/srep36414
Giannaccare, G., Pellegrini, M., Sebastiani, S., Moscardelli, F., Versura, P., and Campos, E. C. (2019). In vivo confocal microscopy morphometric analysis of corneal subbasal nerve plexus in dry eye disease using newly developed fully automated system. Graefes Arch. Clin. Exp. Ophthalmol. 257, 583–589. doi: 10.1007/s00417-018-04225-7
Goldstein, R. H., Barkai, O., Inigo-Portugues, A., Katz, B., Lev, S., and Binshtok, A. M. (2019). Location and plasticity of the sodium spike initiation zone in nociceptive terminals in vivo. Neuron 102, 80–812.e805. doi: 10.1016/j.neuron.2019.03.005
Gong, X., Ren, Y., Fang, X., Cai, J., and Song, E. (2020). Substance P induces sympathetic immune response in the contralateral eye after the first eye cataract surgery in type 2 diabetic patients. BMC Ophthalmol. 20:339. doi: 10.1186/s12886-020-01598-4
Gonzalez-Coto, A. F., Alonso-Ron, C., Alcalde, I., Gallar, J., Meana, A., Merayo-Lloves, J., et al. (2014). Expression of cholecystokinin, gastrin, and their receptors in the mouse cornea. Invest. Ophthalmol. Vis. Sci. 55, 1965–1975. doi: 10.1167/iovs.13-12068
Gonzalez-Gonzalez, O., Bech, F., Gallar, J., Merayo-Lloves, J., and Belmonte, C. (2017). Functional properties of sensory nerve terminals of the mouse cornea. Invest. Ophthalmol. Vis. Sci. 58, 404–415. doi: 10.1167/iovs.16-20033
Goto, T., Oh, S. B., Takeda, M., Shinoda, M., Sato, T., Gunjikake, K. K., et al. (2016). Recent advances in basic research on the trigeminal ganglion. J. Physiol. Sci. 66, 381–386. doi: 10.1007/s12576-016-0448-1
Goyal, S., and Hamrah, P. (2016). Understanding neuropathic corneal pain–gaps and current therapeutic approaches. Semin. Ophthalmol. 31, 59–70. doi: 10.3109/08820538.2015.1114853
Grace, P. M., Hutchinson, M. R., Maier, S. F., and Watkins, L. R. (2014). Pathological pain and the neuroimmune interface. Nat. Rev. Immunol. 14, 217–231. doi: 10.1038/nri3621
Grachev, I. D., Fredrickson, B. E., and Apkarian, A. V. (2000). Abnormal brain chemistry in chronic back pain: an in vivo proton magnetic resonance spectroscopy study. Pain 89, 7–18. doi: 10.1016/S0304-3959(00)00340-7
Guerrero-Moreno, A., Fakih, D., Parsadaniantz, S. M., and Reaux-Le Goazigo, A. (2021). How does chronic dry eye shape peripheral and central nociceptive systems? Neural Regen. Res. 16, 306–307. doi: 10.4103/1673-5374.290895
Guzman, M., Miglio, M., Keitelman, I., Shiromizu, C. M., Sabbione, F., Fuentes, F., et al. (2020). Transient tear hyperosmolarity disrupts the neuroimmune homeostasis of the ocular surface and facilitates dry eye onset. Immunology 161, 148–161. doi: 10.1111/imm.13243
Hamrah, P., Cruzat, A., Dastjerdi, M. H., Pruss, H., Zheng, L., Shahatit, B. M., et al. (2013). Unilateral herpes zoster ophthalmicus results in bilateral corneal nerve alteration: an in vivo confocal microscopy study. Ophthalmology 120, 40–47. doi: 10.1016/j.ophtha.2012.07.036
Hamrah, P., Cruzat, A., Dastjerdi, M. H., Zheng, L., Shahatit, B. M., Bayhan, H. A., et al. (2010). Corneal sensation and subbasal nerve alterations in patients with herpes simplex keratitis: an in vivo confocal microscopy study. Ophthalmology 117, 1930–1936. doi: 10.1016/j.ophtha.2010.07.010
Hamrah, P., Qazi, Y., Shahatit, B., Dastjerdi, M. H., Pavan-Langston, D., Jacobs, D. S., et al. (2017). Corneal nerve and epithelial cell alterations in corneal allodynia: an in vivo confocal microscopy case series. Ocul. Surf. 15, 139–151. doi: 10.1016/j.jtos.2016.10.002
Hanani, M. (2005). Satellite glial cells in sensory ganglia: from form to function. Brain Res. Brain Res. Rev. 48, 457–476. doi: 10.1016/j.brainresrev.2004.09.001
Harris, R. E., and Clauw, D. J. (2012). Imaging central neurochemical alterations in chronic pain with proton magnetic resonance spectroscopy. Neurosci. Lett. 520, 192–196. doi: 10.1016/j.neulet.2012.03.042
Hatta, A., Kurose, M., Sullivan, C., Okamoto, K., Fujii, N., Yamamura, K., et al. (2019). Dry eye sensitizes cool cells to capsaicin-induced changes in activity via TRPV1. J. Neurophysiol. 121, 2191–2201. doi: 10.1152/jn.00126.2018
He, J., Pham, T. L., Kakazu, A. H., and Bazan, H. E. P. (2019). Remodeling of substance p sensory nerves and transient receptor potential melastatin 8 (TRPM8) cold receptors after corneal experimental surgery. Invest. Ophthalmol. Vis. Sci. 60, 2449–2460. doi: 10.1167/iovs.18-26384
Hegarty, D. M., Hermes, S. M., Morgan, M. M., and Aicher, S. A. (2018). Acute hyperalgesia and delayed dry eye after corneal abrasion injury. Pain Rep. 3:e664. doi: 10.1097/PR9.0000000000000664
Hirata, H., and Meng, I. D. (2010). Cold-sensitive corneal afferents respond to a variety of ocular stimuli central to tear production: implications for dry eye disease. Invest. Ophthalmol. Vis. Sci. 51, 3969–3976. doi: 10.1167/iovs.09-4744
Hirata, H., Mizerska, K., Dallacasagrande, V., Guaiquil, V. H., and Rosenblatt, M. I. (2017). Acute corneal epithelial debridement unmasks the corneal stromal nerve responses to ocular stimulation in rats: implications for abnormal sensations of the eye. J. Neurophysiol. 117, 1935–1947. doi: 10.1152/jn.00925.2016
Hirata, H., Mizerska, K., Marfurt, C. F., and Rosenblatt, M. I. (2015). Hyperosmolar tears induce functional and structural alterations of corneal nerves: electrophysiological and anatomical evidence toward neurotoxicity. Invest. Ophthalmol. Vis. Sci. 56, 8125–8140. doi: 10.1167/iovs.15-18383
Ivanusic, J. J., Wood, R. J., and Brock, J. A. (2013). Sensory and sympathetic innervation of the mouse and guinea pig corneal epithelium. J. Comp. Neurol. 521, 877–893. doi: 10.1002/cne.23207
Iwata, K., Imamura, Y., Honda, K., and Shinoda, M. (2011). Physiological mechanisms of neuropathic pain: the orofacial region. Int. Rev. Neurobiol. 97, 227–250. doi: 10.1016/B978-0-12-385198-7.00009-6
Iwata, K., Katagiri, A., and Shinoda, M. (2017). Neuron-glia interaction is a key mechanism underlying persistent orofacial pain. J. Oral Sci. 59, 173–175. doi: 10.2334/josnusd.16-0858
Iwata, K., and Shinoda, M. (2019). Role of neuron and non-neuronal cell communication in persistent orofacial pain. J. Dent. Anesth. Pain. Med. 19, 77–82. doi: 10.17245/jdapm.2019.19.2.77
Jain, A., Hakim, S., and Woolf, C. J. (2020). Unraveling the plastic peripheral neuroimmune interactome. J. Immunol. 204, 257–263. doi: 10.4049/jimmunol.1900818
Jones, M. A., and Marfurt, C. F. (1996). Sympathetic stimulation of corneal epithelial proliferation in wounded and nonwounded rat eyes. Invest. Ophthalmol. Vis. Sci. 37, 2535–2547.
Jones, M. A., and Marfurt, C. F. (1998). Peptidergic innervation of the rat cornea. Exp. Eye Res. 66, 421–435. doi: 10.1006/exer.1997.0446
Jones, S. L., and Svitkina, T. M. (2016). Axon initial segment cytoskeleton: architecture, development, and role in neuron polarity. Neural Plast. 2016:6808293. doi: 10.1155/2016/6808293
Joubert, F., Acosta, M. D. C., Gallar, J., Fakih, D., Sahel, J. A., Baudouin, C., et al. (2019). Effects of corneal injury on ciliary nerve fibre activity and corneal nociception in mice: a behavioural and electrophysiological study. Eur. J. Pain 23, 589–602. doi: 10.1002/ejp.1332
Katagiri, A., Shinoda, M., Honda, K., Toyofuku, A., Sessle, B. J., and Iwata, K. (2012). Satellite glial cell P2Y12 receptor in the trigeminal ganglion is involved in lingual neuropathic pain mechanisms in rats. Mol. Pain 8:23. doi: 10.1186/1744-8069-8-23
Koltzenburg, M., Wall, P. D., and McMahon, S. B. (1999). Does the right side know what the left is doing? Trends Neurosci. 22, 122–127. doi: 10.1016/S0166-2236(98)01302-2
Kovacs, I., Dienes, L., Perenyi, K., Quirce, S., Luna, C., Mizerska, K., et al. (2016a). Lacosamide diminishes dryness-induced hyperexcitability of corneal cold sensitive nerve terminals. Eur. J. Pharmacol. 787, 2–8. doi: 10.1016/j.ejphar.2016.05.044
Kovacs, I., Luna, C., Quirce, S., Mizerska, K., Callejo, G., Riestra, A., et al. (2016b). Abnormal activity of corneal cold thermoreceptors underlies the unpleasant sensations in dry eye disease. Pain 157, 399–417. doi: 10.1097/j.pain.0000000000000455
Kurose, M., and Meng, I. D. (2013). Dry eye modifies the thermal and menthol responses in rat corneal primary afferent cool cells. J. Neurophysiol. 110, 495–504. doi: 10.1152/jn.00222.2013
Labbe, A., Alalwani, H., Van Went, C., Brasnu, E., Georgescu, D., and Baudouin, C. (2012). The relationship between subbasal nerve morphology and corneal sensation in ocular surface disease. Invest. Ophthalmol. Vis. Sci. 53, 4926–4931. doi: 10.1167/iovs.11-8708
Labbe, A., Liang, Q., Wang, Z., Zhang, Y., Xu, L., Baudouin, C., et al. (2013). Corneal nerve structure and function in patients with non-sjogren dry eye: clinical correlations. Invest. Ophthalmol. Vis. Sci. 54, 5144–5150. doi: 10.1167/iovs.13-12370
Labetoulle, M., Baudouin, C., Calonge, M., Merayo-Lloves, J., Boboridis, K. G., Akova, Y. A., et al. (2019). Role of corneal nerves in ocular surface homeostasis and disease. Acta Ophthalmol. 97, 137–145. doi: 10.1111/aos.13844
Launay, P. S., Godefroy, D., Khabou, H., Rostene, W., Sahel, J. A., Baudouin, C., et al. (2015). Combined 3DISCO clearing method, retrograde tracer and ultramicroscopy to map corneal neurons in a whole adult mouse trigeminal ganglion. Exp. Eye Res. 139, 136–143. doi: 10.1016/j.exer.2015.06.008
Launay, P. S., Reboussin, E., Liang, H., Kessal, K., Godefroy, D., Rostene, W., et al. (2016). Ocular inflammation induces trigeminal pain, peripheral and central neuroinflammatory mechanisms. Neurobiol. Dis. 88, 16–28. doi: 10.1016/j.nbd.2015.12.017
Lee, H. K., Kim, K. W., Ryu, J. S., Jeong, H. J., Lee, S. M., and Kim, M. K. (2019). Bilateral effect of the unilateral corneal nerve cut on both ocular surface and lacrimal gland. Invest. Ophthalmol. Vis. Sci. 60, 430–441. doi: 10.1167/iovs.18-26051
Legarreta, M. D., Sheth, C., Prescot, A. P., Renshaw, P. F., McGlade, E. C., and Yurgelun-Todd, D. A. (2020). An exploratory proton MRS examination of gamma-aminobutyric acid, glutamate, and glutamine and their relationship to affective aspects of chronic pain. Neurosci. Res. doi: 10.1016/j.neures.2020.03.002. [Epub ahead of print].
Levine, J. D., Fields, H. L., and Basbaum, A. I. (1993). Peptides and the primary afferent nociceptor. J. Neurosci. 13, 2273–2286. doi: 10.1523/JNEUROSCI.13-06-02273.1993
Levy, O., Labbe, A., Borderie, V., Hamiche, T., Dupas, B., Laroche, L., et al. (2017). Increased corneal sub-basal nerve density in patients with Sjogren syndrome treated with topical cyclosporine A. Clin. Exp. Ophthalmol. 45, 455–463. doi: 10.1111/ceo.12898
Li, F., Yang, W., Jiang, H., Guo, C., Huang, A. J. W., Hu, H., et al. (2019). TRPV1 activity and substance P release are required for corneal cold nociception. Nat. Commun. 10:5678. doi: 10.1038/s41467-019-13536-0
Ljubimov, A. V., and Saghizadeh, M. (2015). Progress in corneal wound healing. Prog. Retin. Eye Res. 49, 17–45. doi: 10.1016/j.preteyeres.2015.07.002
Lopez de Armentia, M., Cabanes, C., and Belmonte, C. (2000). Electrophysiological properties of identified trigeminal ganglion neurons innervating the cornea of the mouse. Neuroscience 101, 1109–1115. doi: 10.1016/S0306-4522(00)00440-1
MacIver, M. B., and Tanelian, D. L. (1993a). Free nerve ending terminal morphology is fiber type specific for A delta and C fibers innervating rabbit corneal epithelium. J. Neurophysiol. 69, 1779–1783. doi: 10.1152/jn.1993.69.5.1779
MacIver, M. B., and Tanelian, D. L. (1993b). Structural and functional specialization of A delta and C fiber free nerve endings innervating rabbit corneal epithelium. J. Neurosci. 13, 4511–4524. doi: 10.1523/JNEUROSCI.13-10-04511.1993
Madrid, R., Donovan-Rodriguez, T., Meseguer, V., Acosta, M. C., Belmonte, C., and Viana, F. (2006). Contribution of TRPM8 channels to cold transduction in primary sensory neurons and peripheral nerve terminals. J. Neurosci. 26, 12512–12525. doi: 10.1523/JNEUROSCI.3752-06.2006
Marfurt, C., Anokwute, M. C., Fetcko, K., Mahony-Perez, E., Farooq, H., Ross, E., et al. (2019). Comparative anatomy of the mammalian corneal subbasal nerve plexus. Invest. Ophthalmol. Vis. Sci. 60, 4972–4984. doi: 10.1167/iovs.19-28519
Marfurt, C. F., Cox, J., Deek, S., and Dvorscak, L. (2010). Anatomy of the human corneal innervation. Exp. Eye Res. 90, 478–492. doi: 10.1016/j.exer.2009.12.010
Marfurt, C. F., and Del Toro, D. R. (1987). Corneal sensory pathway in the rat: a horseradish peroxidase tracing study. J. Comp. Neurol. 261, 450–459. doi: 10.1002/cne.902610309
Markoulli, M., Flanagan, J., Tummanapalli, S. S., Wu, J., and Willcox, M. (2018). The impact of diabetes on corneal nerve morphology and ocular surface integrity. Ocul. Surf. 16, 45–57. doi: 10.1016/j.jtos.2017.10.006
McKay, T. B., Seyed-Razavi, Y., Ghezzi, C. E., Dieckmann, G., Nieland, T. J. F., Cairns, D. M., et al. (2019). Corneal pain and experimental model development. Prog. Retin. Eye Res. 71, 88–113. doi: 10.1016/j.preteyeres.2018.11.005
McKemy, D. D., Neuhausser, W. M., and Julius, D. (2002). Identification of a cold receptor reveals a general role for TRP channels in thermosensation. Nature 416, 52–58. doi: 10.1038/nature719
McLaughlin, C. R., Acosta, M. C., Luna, C., Liu, W., Belmonte, C., Griffith, M., et al. (2010). Regeneration of functional nerves within full thickness collagen-phosphorylcholine corneal substitute implants in guinea pigs. Biomaterials 31, 2770–2778. doi: 10.1016/j.biomaterials.2009.12.031
McMahon, S. B., and Wood, J. N. (2006). Increasingly irritable and close to tears: TRPA1 in inflammatory pain. Cell 124, 1123–1125. doi: 10.1016/j.cell.2006.03.006
McMonnies, C. W. (2017). The potential role of neuropathic mechanisms in dry eye syndromes. J. Optom. 10, 5–13. doi: 10.1016/j.optom.2016.06.002
Mehra, D., Cohen, N. K., and Galor, A. (2020). Ocular surface pain: a narrative review. Ophthalmol. Ther. 9, 1–21. doi: 10.1007/s40123-020-00263-9
Melik Parsadaniantz, S., Rivat, C., Rostene, W., and Reaux-Le Goazigo, A. (2015). Opioid and chemokine receptor crosstalk: a promising target for pain therapy? Nat. Rev. Neurosci. 16, 69–78. doi: 10.1038/nrn3858
Meng, I. D., and Bereiter, D. A. (1996). Differential distribution of Fos-like immunoreactivity in the spinal trigeminal nucleus after noxious and innocuous thermal and chemical stimulation of rat cornea. Neuroscience 72, 243–254. doi: 10.1016/0306-4522(95)00541-2
Moein, H. R., Akhlaq, A., Dieckmann, G., Abbouda, A., Pondelis, N., Salem, Z., et al. (2020). Visualization of micro-neuromas by using in vivo confocal microscopy: an objective biomarker for the diagnosis of neuropathic corneal pain? Ocul. Surf. 18, 651–656. doi: 10.1016/j.jtos.2020.07.004
Moulton, E. A., Becerra, L., Rosenthal, P., and Borsook, D. (2012). An approach to localizing corneal pain representation in human primary somatosensory cortex. PLoS ONE 7:e44643. doi: 10.1371/journal.pone.0044643
Muller, L. J., Marfurt, C. F., Kruse, F., and Tervo, T. M. (2003). Corneal nerves: structure, contents and function. Exp. Eye Res. 76, 521–542. doi: 10.1016/S0014-4835(03)00050-2
Murata, Y., and Masuko, S. (2006). Peripheral and central distribution of TRPV1, substance P and CGRP of rat corneal neurons. Brain Res. 1085, 87–94. doi: 10.1016/j.brainres.2006.02.035
Nakamura, A., Hayakawa, T., Kuwahara, S., Maeda, S., Tanaka, K., Seki, M., et al. (2007). Morphological and immunohistochemical characterization of the trigeminal ganglion neurons innervating the cornea and upper eyelid of the rat. J. Chem. Neuroanat. 34, 95–101. doi: 10.1016/j.jchemneu.2007.05.005
Namavari, A., Chaudhary, S., Sarkar, J., Yco, L., Patel, K., Han, K. Y., et al. (2011). In vivo serial imaging of regenerating corneal nerves after surgical transection in transgenic thy1-YFP mice. Invest. Ophthalmol. Vis. Sci. 52, 8025–8032. doi: 10.1167/iovs.11-8332
Nicolle, P., Liang, H., Reboussin, E., Rabut, G., Warcoin, E., Brignole-Baudouin, F., et al. (2018). Proinflammatory markers, chemokines, and enkephalin in patients suffering from dry eye disease. Int. J. Mol. Sci. 19:1221. doi: 10.3390/ijms19041221
Parra, A., Gonzalez-Gonzalez, O., Gallar, J., and Belmonte, C. (2014). Tear fluid hyperosmolality increases nerve impulse activity of cold thermoreceptor endings of the cornea. Pain 155, 1481–1491. doi: 10.1016/j.pain.2014.04.025
Parra, A., Madrid, R., Echevarria, D., del Olmo, S., Morenilla-Palao, C., Acosta, M. C., et al. (2010). Ocular surface wetness is regulated by TRPM8-dependent cold thermoreceptors of the cornea. Nat. Med. 16, 1396–1399. doi: 10.1038/nm.2264
Peier, A. M., Moqrich, A., Hergarden, A. C., Reeve, A. J., Andersson, D. A., Story, G. M., et al. (2002). A TRP channel that senses cold stimuli and menthol. Cell 108, 705–715. doi: 10.1016/S0092-8674(02)00652-9
Pham, T. L., and Bazan, H. E. P. (2020). Docosanoid signaling modulates corneal nerve regeneration: effect on tear secretion, wound healing, and neuropathic pain. J. Lipid Res. doi: 10.1194/jlr.TR120000954. [Epub ahead of print].
Pham, T. L., Kakazu, A. H., He, J., Jun, B., Bazan, N. G., and Bazan, H. E. P. (2020). Novel RvD6 stereoisomer induces corneal nerve regeneration and wound healing post-injury by modulating trigeminal transcriptomic signature. Sci. Rep. 10:4582. doi: 10.1038/s41598-020-61390-8
Pina, R., Ugarte, G., Campos, M., Inigo-Portugues, A., Olivares, E., Orio, P., et al. (2019). Role of TRPM8 channels in altered cold sensitivity of corneal primary sensory neurons induced by axonal damage. J. Neurosci. 39, 8177–8192. doi: 10.1523/JNEUROSCI.0654-19.2019
Pinho-Ribeiro, F. A., Verri, W. A. Jr., and Chiu, I. M. (2017). Nociceptor sensory neuron-immune interactions in pain and inflammation. Trends Immunol. 38, 5–19. doi: 10.1016/j.it.2016.10.001
Postole, A. S., Knoll, A. B., Auffarth, G. U., and Mackensen, F. (2016). In vivo confocal microscopy of inflammatory cells in the corneal subbasal nerve plexus in patients with different subtypes of anterior uveitis. Br. J. Ophthalmol. 100, 1551–1556. doi: 10.1136/bjophthalmol-2015-307429
Quallo, T., Vastani, N., Horridge, E., Gentry, C., Parra, A., Moss, S., et al. (2015). TRPM8 is a neuronal osmosensor that regulates eye blinking in mice. Nat. Commun. 6:7150. doi: 10.1038/ncomms8150
Rahman, M., Okamoto, K., Thompson, R., Katagiri, A., and Bereiter, D. A. (2015). Sensitization of trigeminal brainstem pathways in a model for tear deficient dry eye. Pain 156, 942–950. doi: 10.1097/j.pain.0000000000000135
Raja, S. N., Carr, D. B., Cohen, M., Finnerup, N. B., Flor, H., Gibson, S., et al. (2020). The revised international association for the study of pain definition of pain: concepts, challenges, and compromises. Pain. 161, 1976–1982. doi: 10.1097/j.pain.0000000000001939
Reaux-Le Goazigo, A., Poras, H., Ben-Dhaou, C., Ouimet, T., Baudouin, C., Wurm, M., et al. (2019). Dual enkephalinase inhibitor PL265: a novel topical treatment to alleviate corneal pain and inflammation. Pain 160, 307–321. doi: 10.1097/j.pain.0000000000001419
Robbins, A., Kurose, M., Winterson, B. J., and Meng, I. D. (2012). Menthol activation of corneal cool cells induces TRPM8-mediated lacrimation but not nociceptive responses in rodents. Invest. Ophthalmol. Vis. Sci. 53, 7034–7042. doi: 10.1167/iovs.12-10025
Rosenthal, P., and Borsook, D. (2016). Ocular neuropathic pain. Br. J. Ophthalmol. 100, 128–134. doi: 10.1136/bjophthalmol-2014-306280
Sarkar, J., Chaudhary, S., Namavari, A., Ozturk, O., Chang, J. H., Yco, L., et al. (2012). Corneal neurotoxicity due to topical benzalkonium chloride. Invest. Ophthalmol. Vis. Sci. 53, 1792–1802. doi: 10.1167/iovs.11-8775
Sarkar, J., Milani, B., Kim, E., An, S., Kwon, J., and Jain, S. (2019). Corneal nerve healing after in situ laser nerve transection. PLoS ONE 14:e0218879. doi: 10.1371/journal.pone.0218879
Shenker, N., Haigh, R., Roberts, E., Mapp, P., Harris, N., and Blake, D. (2003). A review of contralateral responses to a unilateral inflammatory lesion. Rheumatology 42, 1279–1286. doi: 10.1093/rheumatology/keg397
Shetty, R., Sethu, S., Deshmukh, R., Deshpande, K., Ghosh, A., Agrawal, A., et al. (2016). Corneal dendritic cell density is associated with subbasal nerve plexus features, ocular surface disease index, and serum vitamin d in evaporative dry eye disease. Biomed Res. Int. 2016:4369750. doi: 10.1155/2016/4369750
Shin, K. C., Park, H. J., Kim, J. G., Lee, I. H., Cho, H., Park, C., et al. (2019). The Piezo2 ion channel is mechanically activated by low-threshold positive pressure. Sci. Rep. 9:6446. doi: 10.1038/s41598-019-42492-4
Shinoda, M., Kubo, A., Hayashi, Y., and Iwata, K. (2019). Peripheral and central mechanisms of persistent orofacial pain. Front. Neurosci. 13:1227. doi: 10.3389/fnins.2019.01227
Simsek, C., Kojima, T., Nagata, T., Dogru, M., and Tsubota, K. (2019). Changes in murine subbasal corneal nerves after scopolamine-induced dry eye stress exposure. Invest. Ophthalmol. Vis. Sci. 60, 615–623. doi: 10.1167/iovs.18-26318
Spierer, O., Felix, E. R., McClellan, A. L., Parel, J. M., Gonzalez, A., Feuer, W. J., et al. (2016). Corneal mechanical thresholds negatively associate with dry eye and ocular pain symptoms. Invest. Ophthalmol. Vis. Sci. 57, 617–625. doi: 10.1167/iovs.15-18133
Stapleton, F., Alves, M., Bunya, V. Y., Jalbert, I., Lekhanont, K., Malet, F., et al. (2017). TFOS DEWS II epidemiology report. Ocul. Surf. 15, 334–365. doi: 10.1016/j.jtos.2017.05.003
Stepp, M. A., Pal-Ghosh, S., Tadvalkar, G., Williams, A., Pflugfelder, S. C., and de Paiva, C. S. (2018a). Reduced intraepithelial corneal nerve density and sensitivity accompany desiccating stress and aging in C57BL/6 mice. Exp. Eye Res. 169, 91–98. doi: 10.1016/j.exer.2018.01.024
Stepp, M. A., Pal-Ghosh, S., Tadvalkar, G., Williams, A. R., Pflugfelder, S. C., and de Paiva, C. S. (2018b). Reduced corneal innervation in the CD25 null model of sjogren syndrome. Int. J. Mol. Sci. 19:3821. doi: 10.3390/ijms19123821
Strassman, A. M., and Vos, B. P. (1993). Somatotopic and laminar organization of fos-like immunoreactivity in the medullary and upper cervical dorsal horn induced by noxious facial stimulation in the rat. J. Comp. Neurol. 331, 495–516. doi: 10.1002/cne.903310406
Tepelus, T. C., Chiu, G. B., Huang, J., Huang, P., Sadda, S. R., Irvine, J., et al. (2017). Correlation between corneal innervation and inflammation evaluated with confocal microscopy and symptomatology in patients with dry eye syndromes: a preliminary study. Graefes Arch. Clin. Exp. Ophthalmol. 255, 1771–1778. doi: 10.1007/s00417-017-3680-3
Theophanous, C., Jacobs, D. S., and Hamrah, P. (2015). Corneal neuralgia after LASIK. Optom. Vis. Sci. 92, e233–240. doi: 10.1097/OPX.0000000000000652
Thibault, K., Riviere, S., Lenkei, Z., Ferezou, I., and Pezet, S. (2016). Orofacial neuropathic pain leads to a hyporesponsive barrel cortex with enhanced structural synaptic plasticity. PLoS ONE 11:e0160786. doi: 10.1371/journal.pone.0160786
Veiga Moreira, T. H., Gover, T. D., and Weinreich, D. (2007). Electrophysiological properties and chemosensitivity of acutely dissociated trigeminal somata innervating the cornea. Neuroscience 148, 766–774. doi: 10.1016/j.neuroscience.2007.03.056
Vit, J. P., Jasmin, L., Bhargava, A., and Ohara, P. T. (2006). Satellite glial cells in the trigeminal ganglion as a determinant of orofacial neuropathic pain. Neuron Glia Biol. 2, 247–257. doi: 10.1017/S1740925X07000427
Waxman, S. G. (1999). The molecular pathophysiology of pain: abnormal expression of sodium channel genes and its contributions to hyperexcitability of primary sensory neurons. Pain Suppl. 6, S133–140. doi: 10.1016/S0304-3959(99)00147-5
Woolf, C. J. (2011). Central sensitization: implications for the diagnosis and treatment of pain. Pain 152, S2–15. doi: 10.1016/j.pain.2010.09.030
Woolf, C. J. (2020). Capturing novel non-opioid pain targets. Biol. Psychiatry 87, 74–81. doi: 10.1016/j.biopsych.2019.06.017
Xiang, Y., Zhou, W., Wang, P., Yang, H., Gao, F., Xiang, H., et al. (2017). Alkali burn induced corneal spontaneous pain and activated neuropathic pain matrix in the central nervous system in mice. Cornea 36, 1408–1414. doi: 10.1097/ICO.0000000000001336
Xiao, C., Wu, M., Liu, J., Gu, J., Jiao, X., Lu, D., et al. (2019). Acute tobacco smoke exposure exacerbates the inflammatory response to corneal wounds in mice via the sympathetic nervous system. Commun. Biol. 2:33. doi: 10.1038/s42003-018-0270-9
Xue, Y., He, J., Xiao, C., Guo, Y., Fu, T., Liu, J., et al. (2018). The mouse autonomic nervous system modulates inflammation and epithelial renewal after corneal abrasion through the activation of distinct local macrophages. Mucosal. Immunol. 11, 1496–1511. doi: 10.1038/s41385-018-0031-6
Yamada, R., and Kuba, H. (2016). Structural and functional plasticity at the axon initial segment. Front. Cell. Neurosci. 10:250. doi: 10.3389/fncel.2016.00250
Yamaguchi, T., Hamrah, P., and Shimazaki, J. (2016). Bilateral alterations in corneal nerves, dendritic cells, and tear cytokine levels in ocular surface disease. Cornea 35(Suppl. 1), S65–S70. doi: 10.1097/ICO.0000000000000989
Yamazaki, R., Yamazoe, K., Yoshida, S., Hatou, S., Inagaki, E., Okano, H., et al. (2017). The Semaphorin 3A inhibitor SM-345431 preserves corneal nerve and epithelial integrity in a murine dry eye model. Sci. Rep. 7:15584. doi: 10.1038/s41598-017-15682-1
Yang, Y., Li, G., and Chen, L. (2020). High resolution three-dimensional imaging of the ocular surface and intact eyeball using tissue clearing and light sheet microscopy. Ocul. Surf. 18, 526–532. doi: 10.1016/j.jtos.2020.04.009
Keywords: cornea, trigeminal ganglion, pain, inflammation, peripheral and central sensitization
Citation: Guerrero-Moreno A, Baudouin C, Melik Parsadaniantz S and Réaux-Le Goazigo A (2020) Morphological and Functional Changes of Corneal Nerves and Their Contribution to Peripheral and Central Sensory Abnormalities. Front. Cell. Neurosci. 14:610342. doi: 10.3389/fncel.2020.610342
Received: 25 September 2020; Accepted: 18 November 2020;
Published: 10 December 2020.
Edited by:
Takayoshi Masuoka, Kanazawa Medical University, JapanReviewed by:
Ignacio Alcalde, Instituto Universitario Fernández-Vega, SpainIan Meng, University of New England, United States
Yuichi Hori, Toho University, Japan
Copyright © 2020 Guerrero-Moreno, Baudouin, Melik Parsadaniantz and Réaux-Le Goazigo. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Annabelle Réaux-Le Goazigo, YW5uYWJlbGxlLnJlYXV4QGluc2VybS5mcg==