 Lei Hao
Lei Hao Yongtao Yang3†
Yongtao Yang3† Qunling Zhan
Qunling Zhan- 1Department of Neurology, The First Branch of The First Affiliated Hospital of Chongqing Medical University, Chongqing, China
- 2Department of Neurology, The First Affiliated Hospital of Chongqing Medical University, Chongqing, China
- 3Department of Neurology, The Fifth People's Hospital of Chongqing, Chongqing, China
Ischemic stroke accounts for 70–80% of all stroke cases. Immunity plays an important role in the pathophysiology of ischemic stroke. Microglia are the first line of defense in the central nervous system. Microglial functions are largely dependent on their pro-inflammatory (M1-like) or anti-inflammatory (M2-like) phenotype. Modulating neuroinflammation via targeting microglia polarization toward anti-inflammatory phenotype might be a novel treatment for ischemic stroke. Mesenchymal stem cells (MSC) and MSC-derived extracellular vesicles (MSC-EVs) have been demonstrated to modulate microglia activation and phenotype polarization. In this review, we summarize the physiological characteristics and functions of microglia in the healthy brain, the activation and polarization of microglia in stroke brain, the effects of MSC/MSC-EVs on the activation of MSC in vitro and in vivo, and possible underlying mechanisms, providing evidence for a possible novel therapeutics for the treatment of ischemic stroke.
Introduction
Stroke is a major disease leading to long-term disability and high near-term mortality in the world. Ischemic stroke caused by arterial occlusion accounts for 70–80% of all stroke cases, and intracerebral hemorrhage (ICH) is responsible for 10–20% (1). Currently, intravenous (IV) thrombolysis with tissue-type plasminogen activator (t-PA) and endovascular thrombectomy constitute the standard treatments for acute ischemic strokes. However, only 3.4 and 5.2% of all patients with acute ischemic stroke (AIS) are suitable for the treatments due to the extremely narrow therapeutic time window limit (2). Most patients with stroke, who are not eligible for the treatments, are still subjected to neurological deficits such as sensory/motor and/or cognitive impairment (3). To date, there is still no effective treatment to improve stroke patient outcomes in the subacute to chronic phases (4). There is an urgent need for developing efficacious therapies for ischemic stroke.
Immunity has been demonstrated to play a key role in the pathophysiology of ischemic stroke. Modulation of immune responses after ischemic stroke provides novel therapeutic strategies for patients who are not fit for IV thrombolysis and endovascular thrombectomy (5). As the resident immune cells in the central nervous system (CNS), microglia continuously monitor the brain tissue and are one of the first cells to respond to brain injury. The resident microglia are rapidly activated and alter their phenotypes and functions when responding to changes in the local CNS microenvironment after cerebral ischemia. These cells exert either a pro- (M1-like) or anti-inflammatory (M2-like) role according to distinct phenotypes. M1-like microglia aggravate brain damage (6, 7), whereas M2-like microglia enhance brain restoration (8, 9). Both M1 and M2-like microglia have high plasticity and are capable of shifting to other phenotypes according to the cerebral environment (10). Therefore, modulating neuroinflammation via targeting the polarization of microglia to a neuroprotective phenotype might be suitable for developing novel therapeutics for stroke.
Many preclinical studies demonstrated the immunomodulatory properties of MSC in ischemic stroke (11–14). It has been demonstrated that MSC secretion-based paracrine effects are the main mechanism of MSC-mediated therapeutic effects rather than direct cellular replacement (15). MSC may exert their functions in a paracrine manner related to the production of extracellular vesicles (EVs). EVs are membranous particles originating from numerous types of cells, which are enriched with various proteins, lipids, and nucleic acids and act as important mediators of intercellular communication (16). Therapeutic use of MSC-EVs has been attractive in recent years due to their several advantages, including lower risk of immunogenicity and microvascular thrombosis, easy crossing to the blood–brain barrier, easy high production of genetically modified EVs, and great ability to transfer active particles to a target due to a higher surface to volume ratio (17).
MSC mainly exerts immunosuppressive effects on the immune system. It has been demonstrated that MSC can suppress the proliferation, activation, and secretion of pro-inflammatory cytokines of various immune cells such as NK cells, macrophages, and T and B cells (18–21). Mounting evidence demonstrated that MSC and MSC-EVs may improve neurological function by modulating microglial activation and polarization in stroke. In this review, we summarize the effects of MSC and MSC-EVs on microglia in ischemic stroke.
Definition of microglia
Microglia originate from erythromyeloid progenitors in the yolk sac. Microglia precursor cells migrate to the brain and differentiate into microglia during embryological development, of which cardinal functions include synaptic remodeling and maturation (22). In adults, microglia are widely distributed across the whole brain as the resident immune cells of the CNS. The main function of microglia is to maintain brain homeostasis by constantly surveying the CNS microenvironment. Microglia are rapidly activated and migrate to the lesion area after ischemic stroke, which is also called microgliosis. Resident microglia are the main source of microgliosis (23). A study showed that the local residual microglia rather than microglia progenitors mainly contribute to the microglia proliferation (24). Ischemic stroke can promote rapid and complex changes in microglial morphologies and functions.
Morphology of microglia in the healthy and stroke brain
In the brain of healthy adults, the density of microglia is regionally specific. There are more microglia in the hippocampus and the olfactory bulb and fewer microglia in the fiber bundle (25). Resting ramified microglia, associated with immune surveillance and maintenance of homeostasis, have much more long, thin, and highly branched mobile processes under physiological conditions. When stimulated by environmental changes, ramified microglia are transformed to be amoeboid with an enlarged cell body and shorter processes (26). Morphological transformation is usually accompanied by functional responses such as migration, antigen presentation, and phagocytosis (27). There are various morphologies of microglia in different brain regions (28). For example, compared with white matter, microglia in the gray matter have smaller somas and process extensions. Microglia produce effects on CNS surveillance via constantly extending and retracting their branches (29). When microglia cells migrate from gray matter to white matter, their morphology may change from ramified to amoeboid shape with rounded cell bodies and processes along the direction of the fibers (30, 31). The number and morphology of microglia are also different in different regions such as the cortex, the amygdala, the hippocampus, and the preoptic area (POA) between sexes (32–35). Phagocytic microglia are significantly more in the hippocampus of female rats than in male rats 2–3 days after birth (33). Fewer microglia were observed in the female rat pup POA than male at postnatal 2 days (36). Neonatal males had much more activated ameboid microglia in POA than females (36). Microglia from female rats had smaller cell bodies, more process branch points, and greater process length than males (36). Adult male mice had a higher density of Iba+ microglia in the hippocampus, cortex, and amygdala than female mice (32).
Microglia will transform into a more spherical shape and coupe with toxic stimuli when exposed to challenges in the microenvironment (29, 37). Microglial cells can exhibit four types of distinct morphology, namely, ramified, intermediate, amoeboid, and round, when activated after stroke (38). Ramified microglia represent the resting state mainly stay distally to the ischemic core or in the contralateral hemisphere, while intermediate microglia have larger cell bodies and shorter bumps. Conversely, amoeba microglia have larger bodies and shorter processes, or even no processes, similar to round microglia in the lesion center (5, 38). The total volume of reactive microglia gradually increased and reached a peak on the 4th day after ischemic stroke (23). In the first few days, only sparse reactive microglia were found in the ischemic core region. In the later stage, more reactive microglia accumulate in the core zone. The density of microglia with a hypertrophic cell body and no ramified processes was the highest in the accumulation zone surrounding the ischemic core. Reactive microglia in the marginal zone between the accumulation zone and outside the non-ischemic area exhibited fewer short and stout processes than resting microglia (23). In another study, Iba1-positive microglia showed different morphological appearances in the control and the ischemic hemisphere (39). Ramified microglia with long thin processes mostly occurred in the normal area, whereas intermediate microglia with swollen processes were localized in the infarction border zone. Amoeboid microglia with round shape and no processes were mostly observed in the ischemic core between 3 and 7 days after stroke (39).
The function of microglia in the healthy and stroke brain
Microglia actively and continuously monitor the cerebral microenvironment by extending and retracting their processes in adult brain (40). The main role of microglia in a healthy brain is a spectrum of surveilling functions, including normal phagocytosis and synaptic pruning. When stimulated by cell injury, microglia rapidly activate and shift to pro- or anti-inflammatory phenotype. Both spatial and temporal dimensions are very important in microglia dynamics because microglia present with pro- or anti-inflammatory phenotype depending on specific pathophysiological stages and brain regions after stroke (41). Pro-inflammatory microglia (M1-like) secrete detrimental cytokines and molecules such as interleukin (IL)-6, IL-1β, NO, TNF-α, IL-12, and IL-23 and express surface markers such as CD16 and CD32, which aggravate brain injury. However, anti-inflammatory microglia (M2-like) secrete anti-inflammatory cytokines such as IL-4, IL-10, transforming growth factor-β (TGF-β), IL-13, and growth factors such as vascular endothelial growth factor (VEGF) and brain-derived neurotrophic factor (BDNF) and express markers such as CD206,arginase1 (Arg1), TGF-β, insulin growth factor 1 (IGF-1), and IL-10, which promote neurological recovery (40, 42, 43). The M2-like microglia are further categorized as M2a, M2b, and M2c depending on their cellular function (5, 44, 45). M2a microglia, induced by IL-13 or IL-4, are involved in tissue regeneration and repair. M2a microglia also produce significant amounts of Arg1, Ym-1, CD206, and Fizz1 (5, 45). Microglia of M2b phenotype, activated by immune complexes such as Fcγ receptors, Toll-like receptors (TLRs), or IL-1R, exert immunoregulatory effects (5, 44, 46). The M2c phenotype microglia, deactivated by IL-10, TGF-β, and glucocorticoids, promote tissue regeneration when the inflammation reaction recedes (5, 47, 48). The function of microglia is also different between sexes. Most genes associated with inflammatory processes, including regulation of cell migration and cytokine production, are more expressed in male mice microglia compared with females (49). Male microglia are more prone to inflammatory reactions than female microglia due to transcriptionally activated NF-κB (49). However, female microglia have much more expression of genes related to cell plasticity, inflammatory response control, and repair than male microglia (49).
In a transient ischemic stroke model, activated Iba1+microglia appeared within 24 h and reached the peak within 4–7 days in the infarct core area. However, in the peri-infarct region, microglia emerged much earlier within ~3.5 h and peaked at 7 days (50). The temporal kinetics of microglia/macrophage polarization was described in a mouse model of middle cerebral artery occlusion (MCAO) (10). Microglia/macrophages initially attracted to the ischemic area exhibit mainly the M2 phenotype (CD206+Iba+ double-positive cells) only for a short period of time within 7 days post ischemia. The early recruitment of M2-like microglia/macrophages may act to clean ischemic tissue and restrict brain damage. M1-like microglia/macrophages (CD16/32+Iba+ double-positive cells) with decreased phagocytosis and enhanced secretion of pro-inflammatory mediators increasingly occurred from day 3 and remained high level until day 14 after ischemia. These cells may exacerbate neuronal demise and impair axon regrowth. Hence, the “helpful” M2-like microglia were first induced in response to ischemic injury during the early, acute phase of ischemic stroke, of which the function is protecting the neurons in the infarct area, phagocytizing cellular debris, and helping to restrict the area of damage. Then, the “harmful” M1-like microglia become more abundant in the next several days, and the neuronal damage caused by ischemia is increased. The shift of M2- to M1-like microglia in the process of chronic inflammation after stroke exacerbate the neuronal injury, leading to reduced neuronal recovery.
Microglia may respond differently between sexes under ischemic stroke. When subjected to pMCAO, male mice exhibited larger ischemic lesions than female mice (49). The progression of damage was significantly lower in male mice after transplantation of female microglia, indicative of the protective effects of female microglia (49). This phenomenon can be probably interpreted by the fact that Ym1 (a marker for microglia anti-inflammatory activation) immunoreactivity in Iba1-positive cells surrounding the ischemic lesion was higher in male mice transplanted with female microglia than in control (49). Hence, sex-specific microglia phenotype is intrinsically independent of the hormonal environment.
With the development of recent technology such as single-cell RNA sequencing and cytometry by time-of-flight mass spectrometry (CyTOF), spatial, temporal, and functional diversity of microglia during development, homeostasis, and disease in mice and humans has been unveiled (51). Varying degrees of pro- and anti-inflammatory markers may coexist on the same microglial cell. Hence, microglia may exhibit multiple phenotypical subtypes in vivo rather than two individual states based on the clustering of transcriptomic data. For example, 14 microglia sub-clusters were discovered in the cortex penumbra at the early stage of ischemic stroke without complete expression of M1 or M2-type marker genes (52). Five distinct microglial subtypes were identified in the mouse model of MCAO, which did not have a complete overlap with classic M1 or M2 subsets at the single-cell level (53). At least six transcriptionally distinct microglial subsets were uncovered in the stroke-aged brain (54). Further research on temporal and regional heterogeneity of activated microglia based on transcriptomic studies will be helpful in better understanding the pathophysiological mechanism of ischemic stroke.
Effects of MSC/MSC-EVs on morphological changes of microglia
MSCs, a population of non-hematopoietic cells, were identified for the first time in the bone marrow (55). Subsequently, MSCs have been successfully isolated and cultured from various tissues in mammals, including circulating blood, umbilical cord blood (UCB), menstrual blood, the placenta, the heart, the adipose tissue, the skeletal muscle, the pancreas, and the dental pulp (56, 57). MSC surface markers include CD105, CD73, and CD90, and more than 95% of the culture do not express CD34, CD45, and CD14 or CD11b, CD79a or CD19, and HLA-DR (58).
MSCs of different origins have been demonstrated to exert immunomodulatory effects on microglia. One study investigated the effects of adipose tissue-derived mesenchymal stem cells (AMSCs) on microglia morphology and involved intracellular molecules in vitro (59). The results showed that AMSCs were capable of inducing a round, flat microglia phenotype (inflammatory) into a ramifying, anti-inflammatory one. Ramified microglia induced by AMSC have decreased secretion of the pro-inflammatory cytokines such as TNF-a and IL-6, increased phagocytic activity, and the upregulation of neurotrophic factors and of the Arg-1 (a marker for M2-like regulatory microglia) (59). AMSC-secreted CSF-1 binds to its receptor, resulting in phosphorylation of PI3K and ERK1/2, followed by activation of small RhoGTPases, actin polymerization, and cell morphology change and migration (59). The ratio of ramified to hypertrophic microglia was significantly increased in the ischemic brain at 12 weeks when MSC-EVs were IV administered in monkeys 24 h and 14 days after ischemic injury (60). Moreover, microglia in the perilesional gray (PG) of MSC-EV-treated monkeys showed significantly greater branching complexity, process intersections, and length compared with control (60). Therefore, MSC-EV treatment could shift microglial morphology from hypertrophic to ramified microglia, which present with stronger surveillance capacity and homeostatic functions (60).
MSC/MSC-EVs modulating microglia activation and phenotype shift in vitro
MSC transplantation is neuroprotective after stroke at least in part via modulating microglia-mediated neuroinflammation. MSC exhibited inhibition of the pro-inflammatory cytokine expression, such as TNF-α, IL-1β, IL-6, and IFNγ, pro-inflammatory enzyme expression, such as iNOS and COX-2, and chemokine expression, such as MCP1 and GRO-α, when co-cultured with LPS, IL-β, or IFNγ-primed HMO6 or BV2 microglia (61–63) (Table 1). However, MSC-produced paracrine factors may account for its therapeutic effect, considering that >99% of transplanted MSC has been entrapped in the lungs. A study on MSC secretome showed that EVs/exosome was the only component that could successfully reproduce most of the beneficial effects exerted by parent cells (71). In vitro, oxygen- and glucose-deprived (OGD) model can activate microglial-mediated inflammatory response and simulate the ischemic/reperfusion microenvironment in vivo. Several in vitro studies demonstrated that exosomes from MSC could promote M1 to M2 microglia shift in OGD conditions (Table 1). Notably, hypoxia preconditioning could increase the production of exosomes by bone marrow MSC (BMSC), whereas inhibiting the secretion of exosomes to a great extent eliminated its effects on M1 to M2 microglia shift in OGD (66). Therefore, exosomes play pivotal roles in the beneficial effects of MSC on microglia. As a marker of MSC-Exos, miRNAs have been considered the most important component and nearly can reproduce most of the exosomal effects on recipient cells (72). MicroRNA (miR) are small, endogenous, non-coding RNA molecules that can selectively hybridize to the 3′ untranslated region (UTR) poly(A) tail of targeted mRNAs, leading to the prevention of their transcription into proteins or promotion of their degradation (73). When co-cultured with primary microglia cells under OGD conditions, exosomes from adipose-derived stem cells (ADSCs) overexpressing miR-30d-5p have a greater effect in inhibiting inflammatory factor expression and promoting M2 microglial shift by inhibiting autophagy (64). MiR-126+ exosomes can also inhibit microglial activation and the expression of inflammatory factors such as TNF-α and IL-1β in the OGD model (65). Human umbilical cord mesenchymal stem cells (hUCMSC)- derived exosomal miR-26b-5p could inhibit M1 polarization of microglia via targeting CH25H to suppress the TLR pathway (68). Rat BMSC-derived exosomal miR-223-3p induced conversion of inflammatory M1 microglia toward M2 microglia via downregulating CysLT2R transcription and expression (67). hUCMSC-exosomal miR-145-5p was demonstrated to attenuate OGD-induced microglial pro-inflammatory activity, evidenced by lowered expression of IL-6, TNF-α, and IL-1β, via suppression of the IRAK1/TRAF6 signaling pathway (70). Compared with EVs derived from other MSC sources, EVs from the umbilical cord have the ability to exert stronger therapeutic immunomodulation and protective effects (74).
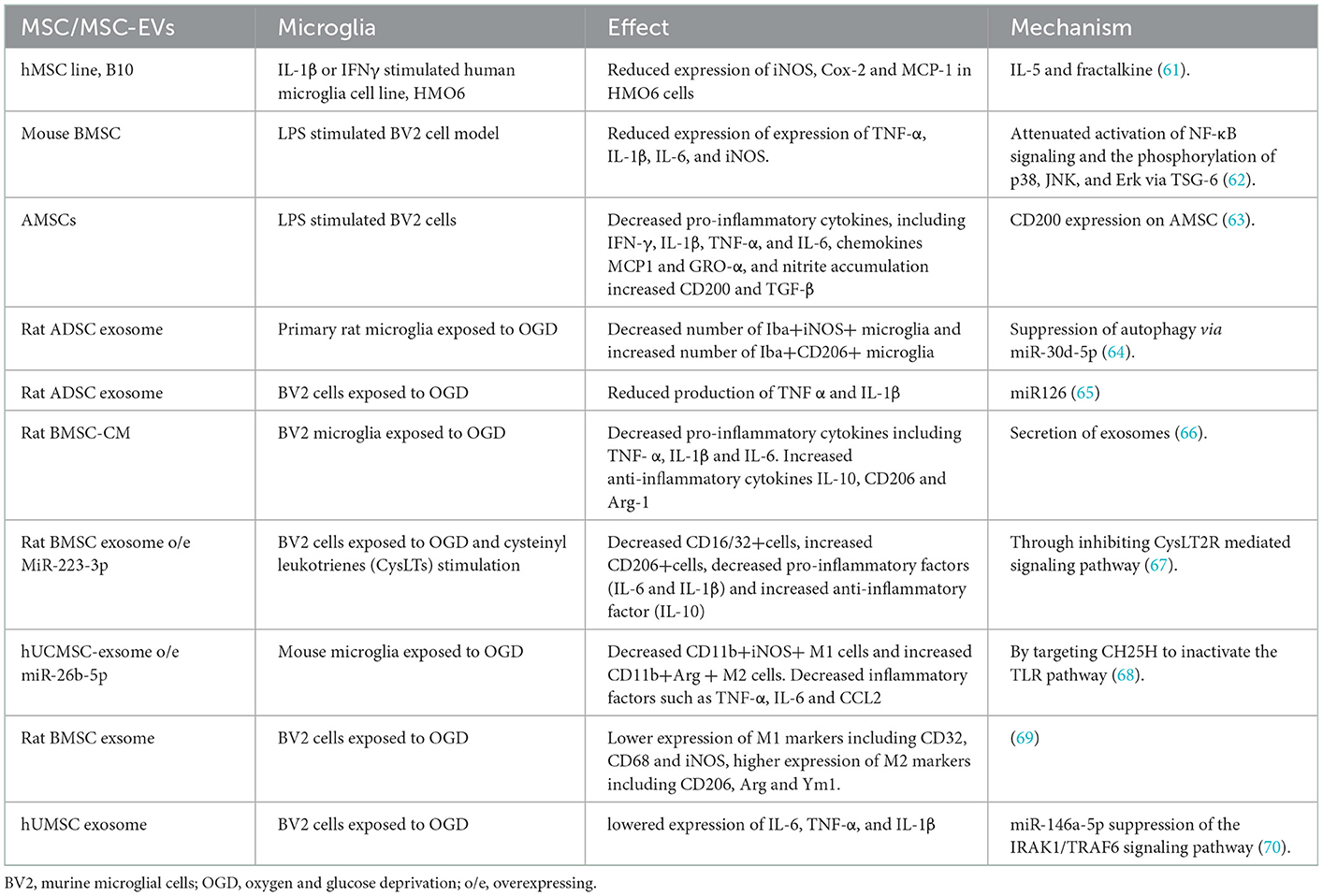
Table 1. MSC modulating microglial activation and polarization in vitro.
MSC/MSC-EVs modulating microglia activation and phenotype shift in vivo
A pile of evidence demonstrated the immunomodulatory effects of MSC or MSC-EVs on microglia in cerebral ischemia model (Table 2). The most commonly used species include rat and mice, whereas monkey was used in one study (60). MSC or MSC-EVs were administrated mainly via IV route, and others include intra-arterial, intracerebral, and intranasal routes. When stem cells were infused in stroke models, most of the animal studies have chosen the acute (within 48 h) and subacute (within 7 days after ischemia) phases as the transplantation time. In one study, hUCMSC was administered IV 2 weeks before cerebral ischemia (87). The number of M1-like microglia (Iba+CD16/32+cells) decreased and M2-like microglia (Iba+CD206+cells) increased 7 days after photothrombotic stroke (87). These results indicated that MSC could also be developed as a prophylactic therapy for patients post-stroke besides therapeutic treatment. Most preclinical studies showed that the number of activated microglia was significantly decreased by MSC or EV administration (15, 61, 63–65, 75, 77–82, 84, 85, 88). Notably, three-dimensional (3D) spheroid cultured MSCs have been demonstrated to exhibit decreased cell size; therefore, these cells are not likely to be entrapped in lung tissue after IV infusion (39). Moreover, the increased expression of chemokine receptor CXCR4 after 3D culture enhanced the homing ability of the MSC to the ischemic brain area. The number of amoeboid microglia decreased significantly 5 days after MCAO. The mRNA expression of microglial markers Iba1 was reduced. 3D-cultured MSC downregulate pro-inflammatory mediators, including IL-12β, Hmgb1, Cxcl10, Ccl2, and IL-1β and increase expression of anti-inflammatory mediators STC1, HGF, and TSG-6 probably via decrease of Mincle expression in microglia cells (39). 3D-cultured MSC has been shown to have enhanced anti-inflammatory effects on microglia, which provide a novel and efficient method for the treatment of immune-mediated disorders. Exosomes derived from MSC of different origins have been demonstrated to exert enhancing effects on microglia polarization from the M1 to M2 phenotype. For example, ADSC-derived exosomes can reduce the number of M1-like microglia (Iba1+/iNOS+cells) and increase the number of M2-like microglia (Iba1+/CD206+cells) at 3 days after MCAO (64). hUCMSCs-exos can also attenuate I/R-induced M1 polarization (68, 70). The number of M1-like microglia (Iba1+/CD32+ cells) was statistically decreased in MCAO rats treated with BMSC-Exos, while the number of M2-like (Iba1+/CD206+cells) microglia was significantly increased (69). Hence, IV-administered BMSC-Exos can shift microglia toward neuroprotective M2 phenotype in the subacute phase of ischemia stroke (69).
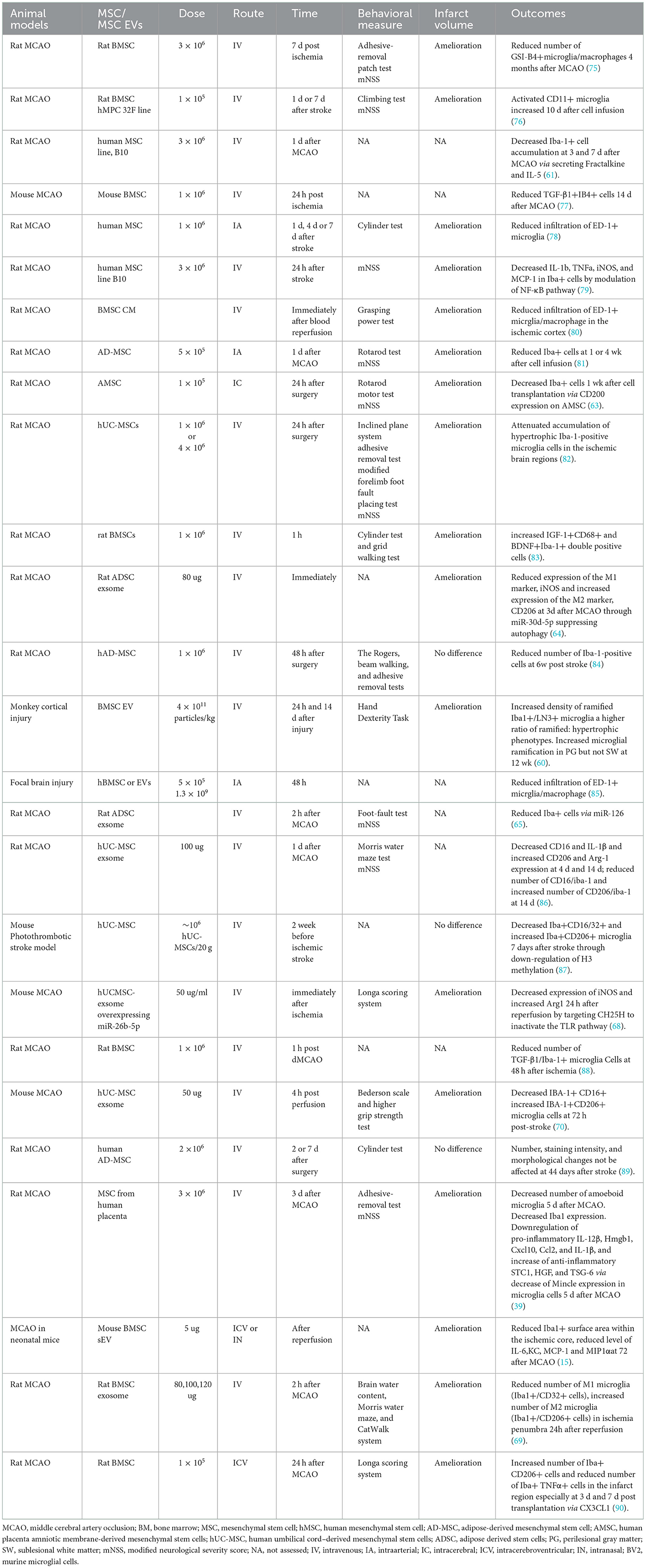
Table 2. Therapeutic use of MSC/MSC EVs in preclinical ischemic stroke animal models.
However, there were some conflicting results. Compared with 7 days post-ischemia, administration of either allogeneic (rMSCs) or xenogeneic (hMSCs) stem cells at 1 day resulted in a significantly greater recovery of motor behavior after MCAO that was possibly related to an increase in activated microglia (CD11) in the infarct region (76). Transplantation of BMSC reduces the proportion of Iba-1+microglia cells expressing TGF-β in the cerebral infarction area 48 h after ischemia (88). IV allogenic BMSC significantly increased the inflammation, evidenced by an increase of TNFαand IL-1βand the number of Iba+ microglia/macrophages in the ischemic core cortex at day 2 after MCAO (83). Hence, MSC delivered at 2 days after MCAO can stimulate but not suppress the immune response (83, 88). It is reasonable that the immunosuppressive capacity of MSC requires a certain amount of time to be induced under inflammatory conditions. The inflammatory reaction will be induced by MSC without being fully activated, followed by exacerbated disease outcome (83). Therefore, the immunosuppressive and anti-inflammatory effects of MSC may be dependent on time. The neurotrophic effects of microglia, evidenced by an increased number of IGF-1+CD68+ and BDNF+Iba-1+ double-positive cells, are most likely responsible for the therapeutic effects of MSC in the acute phase (83). However, in another study, ADMSCs delivered IV at 2 or 7 days post-cerebral ischemia do not have a profound influence on the number or phenotype of Iba+ microglia in the perilesional cortex at 44 days after pMCAO (89). This may be the reason that subpopulations of microglia with different phenotypes were recruited to the perilesion and were involved in tissue repair in different activation stages (89, 91). There are some factors that probably account for these controversial effects of transplanted cells on the number and activation of microglia, including the route, timing, and dose of transplantation, follow-up time, stroke model, and techniques of staining and counting (89).
As mentioned above, most of the in vivo studies did not demonstrate the real causal effects of MSC or MSC-EVs on microglia in stroke. It was possible that morphological and phenotype transformation of microglia in vivo may be secondary. Moreover, neuroprotective function of MSC on ischemic stroke is comprehensive. The effect of MSC on microglia cannot completely account for the amelioration of functional outcome in ischemic stroke. This is because MSC can also promote macrophage polarization toward anti-inflammatory phenotype (92–94). Besides, infiltration of immune cells such as monocytes and neutrophils to inflammation sites can be prevented by MSC (93, 95). Selective elimination of microglia subtype may help better understand the role of MSC in stroke. Most in vivo studies just observed morphological and phenotype changes of microglia after transplantation of MSC or MSC-EVs in stroke model. Accompanied functional alteration has not been explored in detail. Considering various dynamic states of microglia subtype may be present, determination of microglia by M1 or M2 phenotype based on surface markers is oversimplified and inappropriate. The function of numerous microglia subtype in vivo will be future research focus. Indeed, most of the in vivo studies even cannot reliably distinguish microglia from infiltrating macrophages just by detection of biomarkers. The two kinds of immune cells may be distinguished based on CCR2 expression (96). Combined with the extent of CD45 and CD11b expression, the resident microglia can be characterized as CD11bhi/CD45lo/CCR2−, and infiltrating macrophage as CD11blo/CD45hi/CCR2+ (97). Further research is needed for a precise identification of microglia and macrophage in vivo. Considering that EVs contain proteins, lipids, and nucleic acids, molecular profiles of these EVs will be the focus to determine which components may exert beneficial roles on microglia in ischemic stroke. Besides, the underlying specific molecular mechanisms of MSC in modulating microglia in vivo need to be fully elucidated.
Mechanisms of MSC/MSC-EVs modulating the microglia activation and phenotype shift
Transcription factors
Transcription factors are proteins that can regulate the transcriptional activity of genes by binding to DNA. NF-κB translocates from the cytoplasm into the nucleus in microglia following activation after ischemic stroke. Then, activate microglia were transformed into the M1 phenotype, followed by the production of inflammatory cytokines, resulting in secondary brain injury after stroke (98, 99). Neuroinflammatory responses can be attenuated in ischemic stroke via decreasing hypoxia-induced factor-1α following inhibition of activated NF-κB (p65) upregulation (100). MAPKs are a family of serine/threonine protein kinases, including JNK, ERK, and p38, which also play essential roles in the activation and polarization of microglia (101). MSC-derived soluble factor TSG-6 decreases NF-κB activity and the activation of MAPK signaling in LPS-induced microglia, in which the levels of expression of neuroinflammatory cytokines are reduced (62). B10 transplantation (a human MSC line) specifically decreases the NF-κB protein level in macrophage/microglia in the rat MCAO model (79). It is likely that MSC act on NF-κB signaling by inhibiting TLR2 and CD40 expression at the receptor level, increasing the inhibitory cytokine expression such as IL-4 and IL-10 at the activation level and inhibiting the expression of NF-κB itself (79) (see Figure 1).

Figure 1. Mechanisms of MSC/MSC-EVs modulating the microglia activation.
Members of the signal transducer and activator of transcription (STAT) family of transcription factors have been demonstrated to play critical roles in cellular proliferation, immunity, and inflammation (102, 103). STAT3 can regulate macrophage/microglia polarization to anti-inflammatory phenotype (104, 105). However, some studies indicated that STAT3 could induce M1 microglia polarization in both an MCAO-induced and a bilateral common carotid arteries stenosis (BcaS)-induced model of ischemic stroke (106, 107). Consistently, hMSC reduced microglial STAT3 gene expression and activation of Y705 phosphorylated STAT3 following transplantation in an ouabain-induced brain ischemia rat model, which resulted in decreased microglia activation (108). The effect of STAT3 on microglia is likely contradictory, and further studies are required.
Receptors
CX3CL1/CX3CR1 signaling is reported to regulate the microglia activity. It has been demonstrated that CX3CL1/CX3CR1 inhibits LPS-induced microglial activation and decreases expression of inflammatory factors in microglia such as NO, IL-6, and TNF-α via activating the PI3k/Akt pathway, which effectively protect the neuron (109, 110). The infusion of exogenous CX3CL1 reduced cerebral infarct size, neurological deficits, and caspase-3 activation in an MCAO mouse model (111). CX3CL1 expressed by B10 cells (human mesenchymal lines) inhibited cytokine-induced pro-inflammatory gene expression, including iNOS in a human microglia cell line in vitro (61). In addition, when transplanted in rat MCAO model, B10 cell decreased the accumulation of Iba-1+ microglia and inhibited pro-inflammatory gene expression in the core and ischemic border zone (IBZ) (61). In our study, BMSC intraventricular injection significantly increased the number of M2-type microglia (Iba+/CD206+ cells) and decreased the number of M1-type microglia (Iba+/TNFα+ cells) in the infarct region of MCAO rats, especially at 3 days and 7 days post transplantation (90). The above effects can be reversed via interfering with CX3CL1 administration, indicating that BMSC transplantation could shift microglia from pro-inflammatory type to anti-inflammatory in the infarct region of MCAO rats through triggering the secretion of CX3CL1 (90).
CCL2/CCR2 also plays an important regulatory role in post-stroke inflammation. Overexpressed CCL2 has been shown to increase macrophage infiltration and cerebral infarction area, indicating that CCL2 can promote inflammatory response and impede recovery post-stroke (112, 113). In a study, hUCMSC-derived exosome overexpressing CCR2 promoted M2 microglia/macrophage polarization possibly via competitively binding CCL2 to inhibit the excessive activation and M1 polarization of microglia/macrophages (86).
CD200, a member of the immunoglobulin superfamily, plays a critical role as a membrane glycoprotein in the maintenance of immune homeostasis. IL-4-treated hMSCs significantly reduced microglia secretion of IL-6 and IL-1β when co-cultured with LPS-activated microglia through CD200 expression on hMSCs (108). The immunomodulatory effect of hMSC CD200 expression was effectively inhibited by anti-CD200 antibodies (108). In another study, CD200 was more highly expressed under hypoxic conditions in human placenta amniotic membrane-derived mesenchymal stem cells (AMSCs) (63). When CD200 was silenced, the inhibitory effect of AMSC on pro-inflammatory cytokine expression in co-cultures with LPS-primed BV2 was decreased (63). Moreover, AMSC transplantation significantly decreased microglia activation in the boundary region in MCAO through upregulation of CD200 expression (63).
The receptor proteins IRAK1 and TRAF6 are widely distributed in the cytoplasm and nucleus of various cell types. They are largely involved in Toll-like receptor (TLR)-initiated pathways, which contribute to the expression of pro-inflammatory mediators (70). Overexpression of IRAK1 and TRAF6 can activate NF-κB (114). hUMSC-Exos administration decreased IBA-1+CD16+ (M1 type) and increased IBA-1+CD206+ (M2 type) microglia cells at 72 h post-stroke in the MCAO model (70). The immunomodulatory effect on microglia polarization may be mediated by miR-146a-5p, which binds to 3′UTR of the mRNAs encoding IRAK1 and TRAF6 and downregulates the inflammatory response (70).
Cysteinyl leukotrienes (CysLTs) are potent inflammatory mediators derived from the 5-lipoxygenase metabolites of arachidonic acid after cell necrosis (67). The action of CysLTs is mainly mediated by receptors of CysLT1 and CysLT2 (CysLT1R and CysLT2R), which are activated in various types of cells after brain injury (115). During cerebral ischemia, CysLTs receptors (mainly CysLT1R and CysLT2R) are upregulated in activated microglia (116). In a study, BMSC exosome overexpressing MiR-223-3p promoted M2 microglia transformation of M1 microglia when co-cultured with BV2 cells exposed to OGD in vitro through inhibiting CysLT2R (67). Concomitantly, pro-inflammatory factors such as IL-6 and IL-1β decreased and anti-inflammatory factors such as IL-10 increased (67).
Toll-like receptors (TLR) are signaling receptors in the innate immune system that can promote microglial activation and polarization (113, 117). Microglial activation was induced by TLR4 recognition of LPS on the cell surface after brain injury, followed by the production of pro-inflammatory factors via inhibiting the NF-κB pathway (118, 119). Targeted downregulation of TLR4 expression after ischemia-reperfusion injury can inhibit microglial activation and M1-type polarization, and reduce the release of pro-inflammatory factors (117). hUCMSCs-exos attenuated I/R-induced M1 microglia polarization and inflammatory response. The underlying mechanism maybe that exosomal miR-26b-5p could inhibit CH25H to inactivate the TLR pathway, resulting in the repression of M1 polarization of microglia (68).
Modulation of gene expression
MicroRNAs (miRNAs) can regulate gene expression post-transcriptionally by combining with the 3'-UTR, coding sequence, or 5'UTR of target genes (120). They play key roles in the remodeling process under stroke conditions (121). MSC-derived exosomes are capable of transferring miRNAs to recipient cells and subsequently contribute to microglia polarization after stroke. Several studies elucidated the role of specific miRNAs in microglia polarization. ADSC-derived exosomes mediated the delivery of miR-30d-5p to the microglia and reversed OGD-induced and autophagy-mediated microglial polarization to M1 (64). The in vivo study also demonstrated that ADSC exosomal miR-30d-5p promoted M2 microglia/macrophage in the MCAO model by suppressing autophagy (64). IV administration of miR-126+ exosomes from ADSC post-stroke also inhibited microglial activation and the expression of inflammatory factors in vivo (65). hUCMSCs exosomal miR-26b-5p could repress M1 polarization of microglia by targeting CH25H to inactivate the TLR pathway after cerebral I/R (68). MiR-223-3p from BMSC exosome could promote M1 transformation to M2 microglia when co-cultured with BV2 cells exposed to OGD in vitro (67). As mentioned earlier, miR-146a-5p from hUMSC-Exos could promote the microglial shift from M1 to M2 after administration in the MCAO model via downregulation of IRAK1 and TRAF6 (70).
The rate of transcription of genes can be influenced by histone post-translational modifications (PTMs) at gene promoters and distal regulatory elements. Histone PTMs can exert negative and positive effects on transcription, including methylation (122). H3K4me1 increased dramatically in mice photothrombotic stroke models receiving LPS, in which CD16/32-positive microglia elevated and CD206-positive microglia decreased surrounding parainfarct region (87). When hUCMSC was administrated, microglial H3K4me1 modification was robustly alleviated, accompanied by a microglial shift from M1 toward M2 phenotype (87). Therefore, inhibiting H3K4 methylation maybe another putative mechanism for MSC to regulate microglia polarization.
Conclusion
Immunity plays an important role in the pathophysiology of stroke. Microglia, as the first responsive immune cells to ischemia, is considered a double-edged sword in the progression of ischemic stroke. Pro-inflammatory microglia aggravate brain injury, while anti-inflammatory microglia are involved in repair after ischemic stroke. Modulating neuroinflammation via driving activated microglia to be neuroprotective might be novel therapeutics for ischemic stroke. An increasing number of evidence indicate that MSC and MSC-EVs could shift microglial phenotype from the pro-inflammatory M1-like state toward the anti-inflammatory M2-like phenotype. The underlying mechanisms involve several facets, including transcription factors, receptors, and gene expression. Further studies are required to elucidate the specific mechanisms of microglia polarization. Besides, cell source, dose, transplantation route, timing, and safety of MSC or MSC-EVs in ischemic stroke need to be further verified.
Author contributions
LH, YY, and XX: acquisition of data and writing-original draft. XG and QZ: designing the review and writing–review and editing. All authors contributed to the article and approved the submitted version.
Funding
This study was funded by the General Program of Chongqing Natural Science Foundation (cstc2020jcyj-msxmX0988).
Conflict of interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Publisher's note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
References
1. Feigin VL, Lawes CMM, Bennett DA, Barker-Collo SL, Parag V. Worldwide stroke incidence and early case fatality reported in 56 population-based studies: a systematic review. The Lancet Neurology. (2009) 8:355–69. doi: 10.1016/S1474-4422(09)70025-0
2. Members WG, Mozaffarian D, Benjamin EJ, Go AS, Arnett DK, Blaha MJ, et al. Executive Summary: Heart Disease and Stroke Statistics−2016 Update: A Report From the American Heart Association. Circulation. (2016) 133:447–54. doi: 10.1161/CIR.0000000000000366
3. Caleo M. Rehabilitation and plasticity following stroke: Insights from rodent models. Neuroscience. (2015) 311:180–94. doi: 10.1016/j.neuroscience.2015.10.029
4. Kanazawa M, Ninomiya I, Hatakeyama M, Takahashi T, Shimohata T. Microglia and Monocytes/Macrophages Polarization Reveal Novel Therapeutic Mechanism against Stroke. Int J Molec Sci. (2017) 18:2135. doi: 10.3390/ijms18102135
5. Xin W-Q, Wei W, Pan Y-L, Cui B-L, Yang X-Y, Bähr M, et al. Modulating poststroke inflammatory mechanisms: Novel aspects of mesenchymal stem cells, extracellular vesicles and microglia. World J Stem Cells. (2021) 13:1030–48. doi: 10.4252/wjsc.v13.i8.1030
6. Tang Y, Le W. Differential roles of M1 and M2 microglia in neurodegenerative diseases. Mol Neurobiol. (2016) 53:1181–94. doi: 10.1007/s12035-014-9070-5
7. Cheng Q, Shen Y, Cheng Z, Shao Q, Wang C, Sun H, et al. Achyranthes bidentata polypeptide k suppresses neuroinflammation in BV2 microglia through Nrf2-dependent mechanism. Ann Translat Med. (2019) 7:575. doi: 10.21037/atm.2019.09.07
8. Zhang L, Zhang J, You Z. Switching of the microglial activation phenotype is a possible treatment for depression disorder. Front Cell Neurosci. (2018) 12:306. doi: 10.3389/fncel.2018.00306
9. Ma L, Niu W, Lv J, Jia J, Zhu M, Yang S. PGC-1alpha-mediated mitochondrial biogenesis is involved in cannabinoid receptor 2 agonist AM1241-induced microglial phenotype amelioration. Cell Mol Neurobiol. (2018) 38:1529–37. doi: 10.1007/s10571-018-0628-z
10. Hu X, Li P, Guo Y, Wang H, Leak R K, Chen S. Microglia/macrophage polarization dynamics reveal novel mechanism of injury expansion after focal cerebral ischemia. Stroke. (2012) 43:3063–70. doi: 10.1161/STROKEAHA.112.659656
11. Acosta SA, Tajiri N, Hoover J, Kaneko Y, Borlongan CV. Intravenous Bone Marrow Stem Cell Grafts Preferentially Migrate to Spleen and Abrogate Chronic Inflammation in Stroke. Stroke. (2015) 46:2616–27. doi: 10.1161/STROKEAHA.115.009854
12. Karlupia N, Manley NC, Prasad K, Schäfer R, Steinberg GK. Intraarterial transplantation of human umbilical cord blood mononuclear cells is more efficacious and safer compared with umbilical cord mesenchymal stromal cells in a rodent stroke model. Stem Cell Res Ther. (2014) 5:45. doi: 10.1186/scrt434
13. Lee SH, Jin KS, Bang OY, Kim BJ, Park SJ, Lee NH, et al. Differential Migration of Mesenchymal Stem Cells to Ischemic Regions after Middle Cerebral Artery Occlusion in Rats. PLoS One. (2015) 10:e0134920. doi: 10.1371/journal.pone.0134920
14. Yoo SW, Chang DY, Lee HS, Kim GH, Park JS, Ryu BY, et al. Immune following suppression mesenchymal stem cell transplantation in the ischemic brain is mediated by TGF-beta. Neurobiol Dis. (2013) 58:249–57. doi: 10.1016/j.nbd.2013.06.001
15. Pathipati P, Lecuyer M, Faustino J, Strivelli J, Phinney DG, Vexler ZS. Mesenchymal Stem Cell (MSC)-Derived Extracellular Vesicles Protect from Neonatal Stroke by Interacting with Microglial Cells. Neurotherapeutics. (2021) 18:1939–52. doi: 10.1007/s13311-021-01076-9
16. Koniusz S, Andrzejewska A, Muraca M, Srivastava AK, Janowski M, Lukomska B. Extracellular Vesicles in Physiology, Pathology, and Therapy of the Immune and Central Nervous System, with Focus on Extracellular Vesicles Derived from Mesenchymal Stem Cells as Therapeutic Tools. Front Cell Neurosci. (2016) 10:109. doi: 10.3389/fncel.2016.00109
17. Dabrowska S, Andrzejewska A, Lukomska B, Janowski M. Neuroinflammation as a target for treatment of stroke using mesenchymal stem cells and extracellular vesicles. J Neuroinflammation. (2019) 16:178. doi: 10.1186/s12974-019-1571-8
18. Kawata Y, Tsuchiya A, Seino S, Watanabe Y, Kojima Y, Ikarashi S, et al. Early injection of human adipose tissue-derived mesenchymal stem cell after inflammation ameliorates dextran sulfate sodium-induced colitis in mice through the induction of M2 macrophages and regulatory T cells. Cell Tissue Res. (2019) 376:257–71. doi: 10.1007/s00441-018-02981-w
19. Moretta A. Natural killer cells and dendritic cells: rendezvous in abused tissues. Nat Rev Immunol. (2002) 2:957–64. doi: 10.1038/nri956
20. Di Nicola M, Carlo-Stella C, Magni M, Milanesi M, Longoni PD, Matteucci P, et al. Human bone marrow stromal cells suppress T-lymphocyte proliferation induced by cellular or nonspecific mitogenic stimuli. Blood. (2002) 99:3838–43. doi: 10.1182/blood.V99.10.3838
21. Glennie S, Soeiro I, Dyson PJ, Lam EW-F, Dazzi F. Bone marrow mesenchymal stem cells induce division arrest anergy of activated T cells. Blood. (2005) 105:2821–7. doi: 10.1182/blood-2004-09-3696
22. Perry VH, Nicoll JA, Holmes C. Microglia in neurodegenerative disease. Nat Rev Neurol. (2010) 6:193–201. doi: 10.1038/nrneurol.2010.17
23. Li T, Pang S, Yu Y, Wu X, Guo J, Zhang S. Proliferation of parenchymal microglia is the main source of microgliosis after ischaemic stroke. Brain. (2013) 136:3578–88. doi: 10.1093/brain/awt287
24. Huang Y, Xu Z, Xiong S, Sun F, Qin G, Hu G, et al. Repopulated microglia are solely derived from the proliferation of residual microglia after acute depletion. Nat Neurosci. (2018) 21:530–40. doi: 10.1038/s41593-018-0090-8
25. Nikodemova M, Kimyon RS, De I, Small AL, Collier LS, Watters JJ. Microglial numbers attain adult levels after undergoing a rapid decrease in cell number in the third postnatal week. J Neuroimmunol. (2015) 278:280–8. doi: 10.1016/j.jneuroim.2014.11.018
26. Wendimu MY, Hooks SB. Microglia phenotypes in aging and neurodegenerative diseases. Cells. (2022) 11:2091. doi: 10.3390/cells11132091
27. Parakalan R, Jiang B, Nimmi B, Janani M, Jayapal M, Lu J, et al. Transcriptome analysis of amoeboid and ramified microglia isolated from the corpus callosum of rat brain. BMC Neurosci. (2012) 13:64. doi: 10.1186/1471-2202-13-64
28. Kongsui R, Beynon SB, Johnson SJ, Walker FR. Quantitative assessment of microglial morphology and density reveals remarkable consistency in the distribution and morphology of cells within the healthy prefrontal cortex of the rat. J Neuroinflammation. (2014) 11:182. doi: 10.1186/s12974-014-0182-7
30. Arcuri C, Mecca C, Bianchi R, Giambanco I, Donato R. The pathophysiological role of microglia in dynamic surveillance, phagocytosis and structural remodeling of the developing CNS. Front Mol Neurosci. (2017) 10:191. doi: 10.3389/fnmol.2017.00191
31. Torres-Platas SG, Comeau S, Rachalski A, Bo GD, Cruceanu C, Turecki G, et al. Morphometric characterization of microglial phenotypes in human cerebral cortex. J Neuroinflammation. (2014) 11:12. doi: 10.1186/1742-2094-11-12
32. Guneykaya D, Ivanov A, Hernandez DP, Haage V, Wojtas B, Meyer N, et al. Transcriptional and translational differences of microglia from male and female brains. Cell Reports. (2018) 24:2773–83 e6. doi: 10.1016/j.celrep.2018.08.001
33. Nelson LH, Warden S, Lenz KM. Sex differences in microglial phagocytosis in the neonatal hippocampus. Brain Behav Immun. (2017) 64:11–22. doi: 10.1016/j.bbi.2017.03.010
34. Schwarz JM, Sholar PW, Bilbo SD. Sex differences in microglial colonization of the developing rat brain. J Neurochem. (2012) 120:948–63. doi: 10.1111/j.1471-4159.2011.07630.x
35. Rahimian R, Cordeau P, Kriz J. Brain response to injuries: when microglia go sexist. Neuroscience. (2019) 405:14–23. doi: 10.1016/j.neuroscience.2018.02.048
36. Lenz KM, Nugent BM, Haliyur R, McCarthy MM. Microglia are essential to masculinization of brain and behavior. J Neurosci. (2013) 33:2761–72. doi: 10.1523/JNEUROSCI.1268-12.2013
37. Nimmerjahn A, Kirchhoff F, Helmchen F. Resting microglial cells are highly dynamic surveillants of brain parenchyma in vivo. Science. (2005) 308:1314–8. doi: 10.1126/science.1110647
38. Lehrmann E, Christensen T, Zimmer J, Diemer NH, Finsen B. Microglial and macrophage reactions mark progressive changes and define the penumbra in the rat neocortex and striatum after transient middle cerebral artery occlusion. J Compar Neurol. (1997) 386:461–76. doi: 10.1002/(SICI)1096-9861(19970929)386:3<461::AID-CNE9>3.0.CO
39. Li Y, Dong Y, Ran Y, Zhang Y, Wu B, Xie J. Three-dimensional cultured mesenchymal stem cells enhance repair of ischemic stroke through inhibition of microglia. Stem Cell Res Ther. (2021) 12:358. doi: 10.1186/s13287-021-02416-4
40. Lyu J, Xie D, Bhatia TN, Leak RK, Hu X, Jiang X. Microglial/Macrophage polarization and function in brain injury and repair after stroke. CNS Neurosci Ther. (2021) 27:515–27. doi: 10.1111/cns.13620
41. Salter MW, Stevens B. Microglia emerge as central players in brain disease. Nat Med. (2017) 23:1018–27. doi: 10.1038/nm.4397
42. Qin C, Zhou L-Q, Ma X-T, Hu Z-W, Yang S, Chen M, et al. Dual Functions of Microglia in Ischemic Stroke. Neurosci Bull. (2019) 35:921–33. doi: 10.1007/s12264-019-00388-3
43. Jurga AM, Paleczna M, Kuter KZ. Overview of General and Discriminating Markers of Differential Microglia Phenotypes. Front Cell Neurosci. (2020) 14:198. doi: 10.3389/fncel.2020.00198
44. Chhor V, Charpentier TL, Lebon S, Oré M-V, Celador IL, Josserand J, et al. Characterization of phenotype markers and neuronotoxic potential of polarised primary microglia in vitro. Brain Behav Immun. (2013) 32:70–85. doi: 10.1016/j.bbi.2013.02.005
45. Latta CH, Sudduth TL, Weekman EM, Brothers HM, Abner EL, Popa GJ, et al. Determining the role of IL-4 induced neuroinflammation in microglial activity and amyloid-beta using BV2 microglial cells and APP/PS1 transgenic mice. J Neuroinflammation. (2015) 12:41. doi: 10.1186/s12974-015-0243-6
46. Mecha M, Feliú A, Carrillo-Salinas FJ, Rueda-Zubiaurre A, Ortega-Gutiérrez S, de Sola RG, et al. Endocannabinoids drive the acquisition of an alternative phenotype in microglia. Brain Behav Immun. (2015) 49:233–45. doi: 10.1016/j.bbi.2015.06.002
47. Fumagalli S, Perego C, Pischiutta F, Zanier ER, De Simoni M-G. The ischemic environment drives microglia and macrophage function. Front Neurol. (2015) 6:81. doi: 10.3389/fneur.2015.00081
48. Kim E, Cho S. Microglia and monocyte-derived macrophages in stroke. Neurotherapeutics. (2016) 13:702–18. doi: 10.1007/s13311-016-0463-1
49. Villa A, Gelosa P, Castiglioni L, Cimino M, Rizzi N, Pepe G, et al. Sex-specific features of microglia from adult mice. Cell Rep. (2018) 23:3501–11. doi: 10.1016/j.celrep.2018.05.048
50. Ito D, Tanaka K, Suzuki S, Dembo T, Fukuuchi Y. Enhanced expression of Iba1, ionized calcium-binding adapter molecule 1, after transient focal cerebral ischemia in rat brain. Stroke. (2001) 32:1208–15. doi: 10.1161/01.STR.32.5.1208
51. Masuda T, Sankowski R, Staszewski O, Prinz M. Microglia Heterogeneity in the Single-Cell Era. Cell Rep. (2020) 30:1271–81. doi: 10.1016/j.celrep.2020.01.010
52. Guo K, Luo J, Feng D, Wu L, Wang X, Xia L, et al. Single-Cell RNA sequencing with combined use of bulk RNA sequencing to reveal cell heterogeneity and molecular changes at acute stage of ischemic stroke in mouse cortex penumbra area. Front Cell Developm Biol. (2021) 9:624711. doi: 10.3389/fcell.2021.624711
53. Zheng K, Lin L, Jiang W, Chen L, Zhang X, Zhang Q, et al. Single-cell RNA-seq reveals the transcriptional landscape in ischemic stroke. J Cerebral Blood Flow Metabol. (2022) 42:56–73. doi: 10.1177/0271678X211026770
54. Li X, Lyu J, Li R, Jain V, Shen Y, Del Águila Á, et al. Single-cell transcriptomic analysis of the immune cell landscape in the aged mouse brain after ischemic stroke. J Neuroinflammation. (2022) 19:83. doi: 10.1186/s12974-022-02447-5
55. Friedenstein AJ, Piatetzky-Shapiro II, Petrakova KV. Osteogenesis in transplants of bone marrow cells. Journal of embryology and experimental morphology. (1966) 16:381–90. doi: 10.1242/dev.16.3.381
56. Zou Z, Zhang Y, Hao L, Wang F, Liu D, Su Y, et al. More insight into mesenchymal stem cells and their effects inside the body. Expert Opin Biol Ther. (2010) 10:215–30. doi: 10.1517/14712590903456011
57. Hao L, Zou Z, Tian H, Zhang Y, Zhou H, Liu L. Stem cell-based therapies for ischemic stroke. Biomed Res Int. (2014) 2014:468748. doi: 10.1155/2014/468748
58. Dominici M, Blanc KL, Mueller I, Slaper-Cortenbach I, Marini F, Krause D, et al. Minimal criteria for defining multipotent mesenchymal stromal cells. The International Society for Cellular Therapy position statement Cytotherapy. (2006) 8:315–7. doi: 10.1080/14653240600855905
59. Neubrand VE, Pedreño M, Caro M, Forte-Lago I, Delgado M, Gonzalez-Rey E. Mesenchymal stem cells induce the ramification of microglia via the small RhoGTPases Cdc42 and Rac1. Glia. (2014) 62:1932–42. doi: 10.1002/glia.22714
60. Go V, Bowley BGE, Pessina MA, Zhang ZG, Chopp M, Finklestein SP, et al. Extracellular vesicles from mesenchymal stem cells reduce microglial-mediated neuroinflammation after cortical injury in aged Rhesus monkeys. GeroScience. (2020) 42:1–17. doi: 10.1007/s11357-019-00115-w
61. Sheikh AM, Nagai A, Wakabayashi K, Narantuya D, Kobayashi S, Yamaguchi S, et al. Mesenchymal stem cell transplantation modulates neuroinflammation in focal cerebral ischemia: contribution of fractalkine and IL-5. Neurobiol Dis. (2011) 41:717–24. doi: 10.1016/j.nbd.2010.12.009
62. Liu Y, Zhang R, Yan K, Chen F, Huang W, Lv B, et al. Mesenchymal stem cells inhibit lipopolysaccharide-induced inflammatory responses of BV2 microglial cells through TSG-6. J Neuroinflammation. (2014) 11:135. doi: 10.1186/1742-2094-11-135
63. Kong T, Park J-M, Jang JH, Kim C-Y, Bae S-H, Choi Y, et al. Immunomodulatory effect of CD200-positive human placenta-derived stem cells in the early phase of stroke. Exp Mol Med. (2018) 50:e425. doi: 10.1038/emm.2017.233
64. Jiang M, Wang H, Jin M, Yang X, Ji H, Jiang Y, et al. Exosomes from MiR-30d-5p-ADSCs reverse acute ischemic stroke-induced, autophagy-mediated brain injury by promoting m2 microglial/macrophage polarization. Cell Physiol Biochem. (2018) 47:864–78. doi: 10.1159/000490078
65. Geng W, Tang H, Luo S, Lv Y, Liang D, Kang X, et al. Exosomes from miRNA-126-modified ADSCs promotes functional recovery after stroke in rats by improving neurogenesis and suppressing microglia activation. Am J Transl. Res. (2019) 11:780–92.
66. Yu H, Xu Z, Qu G, Wang H, Lin L, Li X, et al. Hypoxic preconditioning enhances the efficacy of mesenchymal stem cells-derived conditioned medium in switching microglia toward anti-inflammatory polarization in ischemia/reperfusion. Cell Mol Neurobiol. (2021) 41:505–24. doi: 10.1007/s10571-020-00868-5
67. Zhao Y, Gan Y, Xu G, Hua K, Liu D. Exosomes from MSCs overexpressing microRNA-223-3p attenuate cerebral ischemia through inhibiting microglial M1 polarization mediated inflammation. Life Sci. (2020) 260:118403. doi: 10.1016/j.lfs.2020.118403
68. Li G, Xiao L, Qin H, Zhuang Q, Zhang W, Liu L, et al. Exosomes-carried microRNA-26b-5p regulates microglia M1 polarization after cerebral ischemia/reperfusion. Cell cycle. (2020) 19:1022–35. doi: 10.1080/15384101.2020.1743912
69. Liu X, Zhang M, Liu H, Zhu R, He H, Zhou Y, et al. Bone marrow mesenchymal stem cell-derived exosomes attenuate cerebral ischemia-reperfusion injury-induced neuroinflammation and pyroptosis by modulating microglia M1/M2 phenotypes. Exp Neurol. (2021) 341:113700. doi: 10.1016/j.expneurol.2021.113700
70. Zhang Z, Zou X, Zhang R, Xie Y, Feng Z, Li F, et al. Human umbilical cord mesenchymal stem cell-derived exosomal miR-146a-5p reduces microglial-mediated neuroinflammation via suppression of the IRAK1/TRAF6 signaling pathway after ischemic stroke. Aging. (2021) 13:3060–79. doi: 10.18632/aging.202466
71. Zhang ZG, Buller B, Chopp M. Exosomes - beyond stem cells for restorative therapy in stroke and neurological injury. Nat Rev Neurol. (2019) 15:193–203. doi: 10.1038/s41582-018-0126-4
72. Ailawadi S, Wang X, Gu H, Fan G-C. Pathologic function and therapeutic potential of exosomes in cardiovascular disease. Biochim Biophys Acta. (2015) 1852:1–11. doi: 10.1016/j.bbadis.2014.10.008
73. Guo H, Ingolia NT, Weissman JS, Bartel DP. Mammalian microRNAs predominantly act to decrease target mRNA levels. Nature. (2010) 466:835–40. doi: 10.1038/nature09267
74. Cai J, Wu J, Wang J, Li Y, Hu X, Luo S, et al. Extracellular vesicles derived from different sources of mesenchymal stem cells: therapeutic effects and translational potential. Cell Biosci. (2020) 10:69. doi: 10.1186/s13578-020-00427-x
75. Li Y, Chen J, Zhang CL, Wang L, Lu D, Katakowski M, et al. Gliosis and brain remodeling after treatment of stroke in rats with marrow stromal cells. Glia. (2005) 49:407–17. doi: 10.1002/glia.20126
76. Yang M, Wei X, Li J, Heine LA, Rosenwasser R, Iacovitti L. Changes in host blood factors and brain glia accompanying the functional recovery after systemic administration of bone marrow stem cells in ischemic stroke rats. Cell Transplant. (2010) 19:1073–84. doi: 10.3727/096368910X503415
77. Xin H, Chopp M, Shen LH, Zhang RL, Zhang L, Zhang ZG, et al. Multipotent mesenchymal stromal cells decrease transforming growth factor beta1 expression in microglia/macrophages and down-regulate plasminogen activator inhibitor 1 expression in astrocytes after stroke. Neurosci Lett. (2013) 542:81–6. doi: 10.1016/j.neulet.2013.02.046
78. Ishizaka S, Horie N, Satoh K, Fukuda Y, Nishida N, Nagata I. Intra-arterial cell transplantation provides timing-dependent cell distribution and functional recovery after stroke. Stroke. (2013) 44:720–6. doi: 10.1161/STROKEAHA.112.677328
79. Wang H, Nagai A, Sheikh AM, Liang XY, Yano S, Mitaki S, et al. Human mesenchymal stem cell transplantation changes proinflammatory gene expression through a nuclear factor-kappaB-dependent pathway in a rat focal cerebral ischemic model. J Neurosci Res. (2013) 91:1440–9. doi: 10.1002/jnr.23267
80. Tsai M-J, Tsai S-K, Hu B-R, Liou D-Y, Huang S-L, Huang M-C, et al. Recovery of neurological function of ischemic stroke by application of conditioned medium of bone marrow mesenchymal stem cells derived from normal and cerebral ischemia rats. J Biomed Sci. (2014) 21:5. doi: 10.1186/1423-0127-21-5
81. Oh S-H, Choi C, Chang D-J, Shin D-A, Lee N, Jeon I, et al. Early neuroprotective effect with lack of long-term cell replacement effect on experimental stroke after intra-arterial transplantation of adipose-derived mesenchymal stromal cells. Cytotherapy. (2015) 17:1090–103. doi: 10.1016/j.jcyt.2015.04.007
82. Lin W, Hsuan YC-Y, Lin M-T, Kuo T-W, Lin C-H, Su Y-C, et al. Human umbilical cord mesenchymal stem cells preserve adult newborn neurons and reduce neurological injury after cerebral ischemia by reducing the number of hypertrophic microglia/macrophages. Cell Transplant. (2017) 26:1798–810. doi: 10.1177/0963689717728936
83. Li X, Huang M, Zhao R, Zhao C, Liu Y, Zou H, et al. Intravenously delivered allogeneic mesenchymal stem cells bidirectionally regulate inflammation and induce neurotrophic effects in distal middle cerebral artery occlusion rats within the first 7 days after stroke. Cell Physiol Biochem. (2018) 46:1951–70. doi: 10.1159/000489384
84. Gómez-de Frutos MC, Laso-García F, Diekhorst L, Otero-Ortega L, Fuentes B, Jolkkonen J, et al. Intravenous delivery of adipose tissue-derived mesenchymal stem cells improves brain repair in hyperglycemic stroke rats. Stem Cell Res Therapy. (2019) 10:212. doi: 10.1186/s13287-019-1322-x
85. Dabrowska S, Andrzejewska A, Strzemecki D, Muraca M, Janowski M, Lukomska B. Human bone marrow mesenchymal stem cell-derived extracellular vesicles attenuate neuroinflammation evoked by focal brain injury in rats. J Neuroinflammation. (2019) 16:216. doi: 10.1186/s12974-019-1602-5
86. Yang HC, Zhang M, Wu R, Zheng HQ, Zhang LY, Luo J, et al. C-C chemokine receptor type 2-overexpressing exosomes alleviated experimental post-stroke cognitive impairment by enhancing microglia/macrophage M2 polarization. World J Stem Cells. (2020) 12:152–67. doi: 10.4252/wjsc.v12.i2.152
87. Feng YW, Wu C, Liang FY, Lin T, Li WQ, Jing YH, et al. hUCMSCs Mitigate LPS-induced trained immunity in ischemic stroke. Front Immunol. (2020) 11:1746. doi: 10.3389/fimmu.2020.01746
88. Liang Z-H, Gu J-J, Yu W-X, Guan Y-Q, Khater M, Li X-B. Bone marrow mesenchymal stem cell transplantation downregulates plasma level and the microglia expression of transforming growth factor beta1 in the acute phase of cerebral cortex ischemia. Chronic Dis Translat Med. (2020) 6:270–80. doi: 10.1016/j.cdtm.2020.05.005
89. Bakreen A, Juntunen M, Dunlop Y, Ugidos IF, Malm T, Miettinen S, et al. Additive behavioral improvement after combined cell therapy and rehabilitation despite long-term microglia presence in stroke rats. Int J Molec Sci. (2021) 22:1512. doi: 10.3390/ijms22041512
90. Bai X-J, Hao L, Guo Y-E, Shi X-B, Wu W-P. Bone marrow stromal cells reverse the microglia type from pro-inflammatory tumour necrosis factor a microglia to anti-inflammatory CD206 microglia of middle cerebral artery occlusion rats through triggering secretion of CX3CL1. Folia neuropathologica. (2021) 59:20–31. doi: 10.5114/fn.2021.105129
91. Ladwig A, Walter HL, Hucklenbroich J, Willuweit A, Langen K-J, Fink GR, et al. Osteopontin augments M2 microglia response and separates M1- and M2-polarized microglial activation in permanent focal cerebral ischemia. Mediators Inflamm. (2017) 2017:7189421. doi: 10.1155/2017/7189421
92. Vasandan AB, Jahnavi S, Shashank C, Prasad P, Kumar A, Prasanna SJ. Human Mesenchymal stem cells program macrophage plasticity by altering their metabolic status via a PGE(2)-dependent mechanism. Sci Rep. (2016) 6:38308. doi: 10.1038/srep38308
93. Wang G, Cao K, Liu K, Xue Y, Roberts AI, Li F, et al. Kynurenic acid, an IDO metabolite, controls TSG-6-mediated immunosuppression of human mesenchymal stem cells. Cell Death Differ. (2018) 25:1209–23. doi: 10.1038/s41418-017-0006-2
94. Selleri S, Bifsha P, Civini S, Pacelli C, Dieng MM, Lemieux W, et al. Human mesenchymal stromal cell-secreted lactate induces M2-macrophage differentiation by metabolic reprogramming. Oncotarget. (2016) 7:30193–210. doi: 10.18632/oncotarget.8623
95. Song HB, Park SY, Ko JH, Park JW, Yoon CH, Kim DH, et al. Mesenchymal Stromal Cells Inhibit Inflammatory Lymphangiogenesis in the Cornea by Suppressing Macrophage in a TSG-6-Dependent Manner. Molec Ther. (2018) 26:162–72. doi: 10.1016/j.ymthe.2017.09.026
96. Geissmann F, Manz MG, Jung S, Sieweke MH, Merad M, Ley K. Development of monocytes, macrophages, and dendritic cells. Science. (2010) 327:656–61. doi: 10.1126/science.1178331
97. Calvo B, Rubio F, Fernández M, Tranque P. Dissociation of neonatal and adult mice brain for simultaneous analysis of microglia, astrocytes and infiltrating lymphocytes by flow cytometry. IBRO Reports. (2020) 8:36–47. doi: 10.1016/j.ibror.2019.12.004
98. Xiang B, Xiao C, Shen T, Li X. Anti-inflammatory effects of anisalcohol on lipopolysaccharide-stimulated BV2 microglia via selective modulation of microglia polarization and down-regulation of NF-kappaB p65 and JNK activation. Mol Immunol. (2018) 95:39–46. doi: 10.1016/j.molimm.2018.01.011
99. Jiang C-T, Wu W-F, Deng Y-H, Ge J-W. Modulators of microglia activation and polarization in ischemic stroke (Review). Mol Med Rep. (2020) 21:2006–18. doi: 10.3892/mmr.2020.11003
100. Liu R, Liao X-Y, Pan M-X, Tang J-C, Chen S-F, Zhang Y, et al. Glycine exhibits neuroprotective effects in ischemic stroke in rats through the inhibition of m1 microglial polarization via the NF-kappaB p65/Hif-1alpha signaling pathway. J Immunol. (2019) 202:1704–14. doi: 10.4049/jimmunol.1801166
101. Yang S, Wang H, Yang Y, Wang R, Wang Y, Wu C, et al. Baicalein administered in the subacute phase ameliorates ischemia-reperfusion-induced brain injury by reducing neuroinflammation and neuronal damage. Biomed Pharmacother. (2019) 117:109102. doi: 10.1016/j.biopha.2019.109102
102. Villarino AV, Kanno Y, O'Shea JJ. Mechanisms and consequences of Jak-STAT signaling in the immune system. Nat Immunol. (2017) 18:374–84. doi: 10.1038/ni.3691
103. Banerjee S, Biehl A, Gadina M, Hasni S, Schwartz DM JAK-STAT Signaling as a Target for Inflammatory and Autoimmune Diseases: Current and Future Prospects. Drugs. (2017) 77:521–46. doi: 10.1007/s40265-017-0701-9
104. Cianciulli A, Dragone T, Calvello R, Porro C, Trotta T, Lofrumento DD. IL-10 plays a pivotal role in anti-inflammatory effects of resveratrol in activated microglia cells. Int Immunopharmacol. (2015) 24:369–76. doi: 10.1016/j.intimp.2014.12.035
105. Liu Z-J, Ran Y-Y, Qie S-Y, Gong W-J, Gao F-H, Ding Z-T, et al. Melatonin protects against ischemic stroke by modulating microglia/macrophage polarization toward anti-inflammatory phenotype through STAT3 pathway. CNS Neurosci Ther. (2019) 25:1353–62. doi: 10.1111/cns.13261
106. Qin C, Fan W-H, Liu Q, Shang K, Murugan M, Wu L-J, et al. Fingolimod Protects Against Ischemic White Matter Damage by Modulating Microglia Toward M2 Polarization via STAT3 Pathway. Stroke. (2017) 48:3336–46. doi: 10.1161/STROKEAHA.117.018505
107. Ding Y, Qian J, Li H, Shen H, Li X, Kong Y, et al. Effects of SC99 on cerebral ischemia-perfusion injury in rats: Selective modulation of microglia polarization to M2 phenotype via inhibiting JAK2-STAT3 pathway. Neurosci Res. (2019) 142:58–68. doi: 10.1016/j.neures.2018.05.002
108. McGuckin CP, Jurga M, Miller A-M, Sarnowska A, Wiedner M, Boyle NT, et al. Ischemic brain injury: a consortium analysis of key factors involved in mesenchymal stem cell-mediated inflammatory reduction. Arch Biochem Biophys. (2013) 534:88–97. doi: 10.1016/j.abb.2013.02.005
109. Lyons A, Lynch AM, Downer E J, Hanley R, O'Sullivan JB, Smith A. Fractalkine-induced activation of the phosphatidylinositol-3 kinase pathway attentuates microglial activation in vivo and in vitro. J Neurochem. (2009) 110:1547–56. doi: 10.1111/j.1471-4159.2009.06253.x
110. Mizuno T, Kawanokuchi J, Numata K, Suzumura A. Production and neuroprotective functions of fractalkine in the central nervous system. Brain Res. (2003) 979:65–70. doi: 10.1016/S0006-8993(03)02867-1
111. Cipriani R, Villa P, Chece G, Lauro C, Paladini A, Micotti E, et al. CX3CL1 is neuroprotective in permanent focal cerebral ischemia in rodents. J Neurosci. (2011) 31:16327–35. doi: 10.1523/JNEUROSCI.3611-11.2011
112. Chen Y, Hallenbeck JM, Ruetzler C, Bol D, Thomas K, Berman NEJ, et al. Overexpression of monocyte chemoattractant protein 1 in the brain exacerbates ischemic brain injury and is associated with recruitment of inflammatory cells. J Cerebral Blood Flow Metab. (2003) 23:748–55. doi: 10.1097/01.WCB.0000071885.63724.20
113. Dong R, Huang R, Wang J, Liu H, Xu Z. Effects of microglial activation and polarization on brain injury after stroke. Front Neurol. (2021) 12:620948. doi: 10.3389/fneur.2021.620948
114. Wang Z, Liu F, Wei M, Qiu Y, Ma C, Shen L, et al. Chronic constriction injury-induced microRNA-146a-5p alleviates neuropathic pain through suppression of IRAK1/TRAF6 signaling pathway. J Neuroinflammation. (2018) 15:179. doi: 10.1186/s12974-018-1215-4
115. Wang Y, Yang Y, Zhang S, Li C, Zhang L. Modulation of neuroinflammation by cysteinyl leukotriene 1 and 2 receptors: implications for cerebral ischemia and neurodegenerative diseases. Neurobiol Aging. (2020) 87:1–10. doi: 10.1016/j.neurobiolaging.2019.12.013
116. Shi QJ, Wang H, Liu ZX, Fang SH, Song XM, Lu YB, et al. HAMI 3379, a CysLT2R antagonist, dose- and time-dependently attenuates brain injury and inhibits microglial inflammation after focal cerebral ischemia in rats. Neuroscience. (2015) 291:53–69. doi: 10.1016/j.neuroscience.2015.02.002
117. Tian X, Liu H, Xiang F, Xu L, Dong Z. β-Caryophyllene protects against ischemic stroke by promoting polarization of microglia toward M2 phenotype via the TLR4 pathway. Life Sci. (2019) 237:116915. doi: 10.1016/j.lfs.2019.116915
118. Yao L, Kan EM, Lu J, Hao A, Dheen ST, Kaur C. Toll-like receptor 4 mediates microglial activation and production of inflammatory mediators in neonatal rat brain following hypoxia: role of TLR4 in hypoxic microglia. J Neuroinflammation. (2013) 10:23. doi: 10.1186/1742-2094-10-23
119. Yang M-Y, Yu Q-L, Huang Y-S, Yang G. Neuroprotective effects of andrographolide derivative CX-10 in transient focal ischemia in rat: Involvement of Nrf2/AE and TLR/NF-kappaB signaling. Pharmacol Res. (2019) 144:227–34. doi: 10.1016/j.phrs.2019.04.023
120. Xue Y, Nie D, Wang L-J, Qiu H-C, Ma L, Dong M-X, et al. Microglial Polarization: Novel Therapeutic Strategy against Ischemic Stroke. Aging Dis. (2021) 12:466–79. doi: 10.14336/AD.2020.0701
121. Liu XS, Chopp M, Zhang RL, Zhang ZG. MicroRNAs in cerebral ischemia-induced neurogenesis. J Neuropathol Exp Neurol. (2013) 72:718–22. doi: 10.1097/NEN.0b013e31829e4963
Keywords: mesenchymal stem cell, extracellular vesicles, microglia, ischemic stroke, cell polarization
Citation: Hao L, Yang Y, Xu X, Guo X and Zhan Q (2023) Modulatory effects of mesenchymal stem cells on microglia in ischemic stroke. Front. Neurol. 13:1073958. doi: 10.3389/fneur.2022.1073958
Received: 19 October 2022; Accepted: 28 December 2022;
Published: 18 January 2023.
Edited by:
Dirk M. Hermann, University of Duisburg-Essen, GermanyReviewed by:
Marco Bacigaluppi, San Raffaele Scientific Institute (IRCCS), ItalyZhongmin Zou, Army Medical University, China
Copyright © 2023 Hao, Yang, Xu, Guo and Zhan. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Xiuming Guo,  xmguo813@163.com; Qunling Zhan, aczhan@163.com
xmguo813@163.com; Qunling Zhan, aczhan@163.com
†These authors have contributed equally to this work