 Zihong Xiong
Zihong Xiong Chao Sun
Chao Sun Shiyong Huang
Shiyong Huang- 1Department of Pediatric Intensive Care Unit, Chengdu Women’s and Children’s Central Hospital, School of Medicine, University of Electronic Science and Technology of China, Chengdu, China
- 2Department of Pediatrics, Chongzhou Health Center for Women and Children, Chongzhou, China
- 3Department of Pediatrics, Southwest Medical University, Luzhou, China
Neural cell death is a critical pathological mechanism underlying the development and progression of central nervous system (CNS) diseases, where programmed cell death (PCD) pathways serve as critical regulatory hubs. In addition to classical apoptosis and autophagy, emerging PCD modalities including necroptosis, pyroptosis, ferroptosis, and cuproptosis exhibit distinct activation patterns in different neurological diseases. Long non-coding RNAs (lncRNAs) have emerged as pivotal regulators of these PCD processes through multiple molecular strategies by modulating chromatin accessibility, assembling signaling complexes, and regulating post-transcriptional processes. These regulatory interactions vary by cellular location and disease stage, influencing cell fate through membrane receptors, kinase cascades, and nuclear transcriptional programs. In various CNS pathologies, specific lncRNAs display dual regulatory capacities—promoting neuronal death by amplifying cytotoxic signals or conferring neuroprotection by inhibiting these pathways. The dynamic lncRNA-PCD interactions offer therapeutic potential through targeted modulation of lncRNA networks to control neuronal survival. Future investigations should prioritize systematic mapping of context-specific lncRNA regulatory networks governing distinct PCD modalities, concurrently advancing spatial epigenomic editing technologies for precise manipulation of these regulatory circuits. Understanding these molecular interactions better will help identify therapeutic targets and guide CNS drug development.
1 Introduction
Long non-coding RNAs (lncRNAs), defined as longer than 200 nucleotidest non-protein-coding transcripts (Nitsche and Stadler, 2017), have transitioned from being mischaracterized as transcriptional noise to emerging as master regulators of gene expression through diverse mechanisms including transcriptional modulation (histone modification, cofactor recruitment), RNA processing (alternative splicing), and post-translational regulation—notably functioning as endogenous RNA sponges within competing endogenous RNA (ceRNA) networks (Khoshnam et al., 2024; Kitagawa et al., 2013; Ruffo et al., 2023; Wu et al., 2021; Yang et al., 2023). Their region-specifically enriched expression in murine hippocampal, cortical, and thalamic structures (Kadakkuzha et al., 2015) and zebrafish CNS (brain/spinal cord) (Kaushik et al., 2013) demonstrates conserved regulation across vertebrate neural systems, underpinning roles in coordinating neural cell proliferation, differentiation, and programmed death (Ghafouri-Fard et al., 2022; Herman et al., 2022; Kleaveland et al., 2018; Wu and Kuo, 2020). This review synthesizes lncRNA-mediated regulation of neural programmed cell death (PCD) by integrating three fundamental dimensions: (1) functional convergence and divergence across apoptosis, autophagy, pyroptosis, ferroptosis, and cuproptosis pathways; (2) shared mechanistic principles spanning ceRNA networks and epigenetic reprogramming; and (3) context-specific dysregulation in neurodegeneration and neurotrauma, thereby addressing the identified knowledge gap in lncRNA-PCD interactomes within the nervous system.
2 LncRNA-mediated programmed neural cell death in central nervous system diseases
2.1 Apoptosis
Apoptosis—an evolutionarily conserved cell death mechanism mediating tissue homeostasis—executes through cytoplasmic shrinkage, nuclear fragmentation, and apoptotic body formation via two core pathways: (1) The mitochondrial pathway, where oxidative stress induces Bax/Bak-mediated mitochondrial pore formation, cytochrome c release, and apoptosome assembly (Apaf-1/pro-caspase-9), culminating in caspase-9 autoactivation and downstream caspase-3/7 proteolysis; (2) The death receptor pathway, initiated by extracellular ligands (FasL/TNF-ɑ) that trimerize receptors to recruit FADD and pro-caspase-8, forming the DISK complex for caspase-8 dimerization-driven autocleavage (Amgalan et al., 2017; Fernández et al., 2015; Riley and Bock, 2022; Schwarzer et al., 2020; Van Opdenbosch and Lamkanfi, 2019). Crucially, while dysregulation of these pathways drives neurodegeneration in cerebral ischemia and chronic disorders, lncRNAs’ regulatory roles in modulating key molecular switches—such as Bax activation thresholds, apoptosome stoichiometry, or DISK assembly kinetics—remain underexplored in neural contexts.
Emerging evidence establishes specific lncRNAs as critical coordinators of cell fate across pathologies. In solid tumors, plasmacytoma variant translocation 1 (PVT1) and p21-associated lncRNA exemplify dual-function regulators orchestrating cellular apoptosis while mediating therapeutic resistance through apoptotic threshold modulation (Ferrer and Dimitrova, 2024; He et al., 2018; Li et al., 2022). In ischemic stroke, MEG3 orchestrates bimodal apoptotic control via hnRNPA1 sequestration to induce Sirt2-dependent mitochondrial dysfunction (Yao et al., 2024), concurrently sequestering cytoplasmic miR-21 to elevate PDCD4 and activate caspase-3 (Yan et al., 2017). The lncRNA KCNQ1OT1 exerts neurotoxic effects by competitively binding miR-296-5p, upregulating Bax expression and enhancing mitochondrial cytochrome c release (Li et al., 2020), while SNHG14 attenuates neuronal injury via miR-98-5p sponging to upregulate BCL2L13 in neuro-2a cells (Zhang et al., 2022). Functional heterogeneity is evident in NEAT1: Global suppression dysregulates synaptic plasticity (Li and Wang, 2023), whereas pharmacological induction by bexarotene elevates NEAT1 to attenuate Pidd1-caspase-2 signaling in traumatic brain injury (Zhong et al., 2017). Neuroprotective mechanisms include N1LR-mediated inhibition of p53 phosphorylation at Ser15 (Wu et al., 2017), and TUG1 promoting neuronal apoptosis through miR-9 sponging and Bcl2l11 upregulation (Chen et al., 2017). The ZFAS1 neuroprotective axis demonstrates miR-582 sequestration to upregulate NOS3 expression (Zhang and Zhang, 2020), while Rian attenuates apoptosis via miR-144-3p/GATA3 signaling (Yao et al., 2020) and Oprm1 reduces cell death through miR-155/GATA3 signaling (Jing et al., 2019). Critically, comparative profiling reveals divergent targeting of death pathways: stroke-associated lncRNAs (MEG3, KCNQ1OT1) converge on mitochondrial apoptosis, whereas neuroprotective lncRNAs (NEAT1, ZFAS1, N1LR) modulate caspase-dependent survival. These systematically cataloged mechanisms establish lncRNAs as hierarchically organized master regulators of neuronal fate (Table 1).
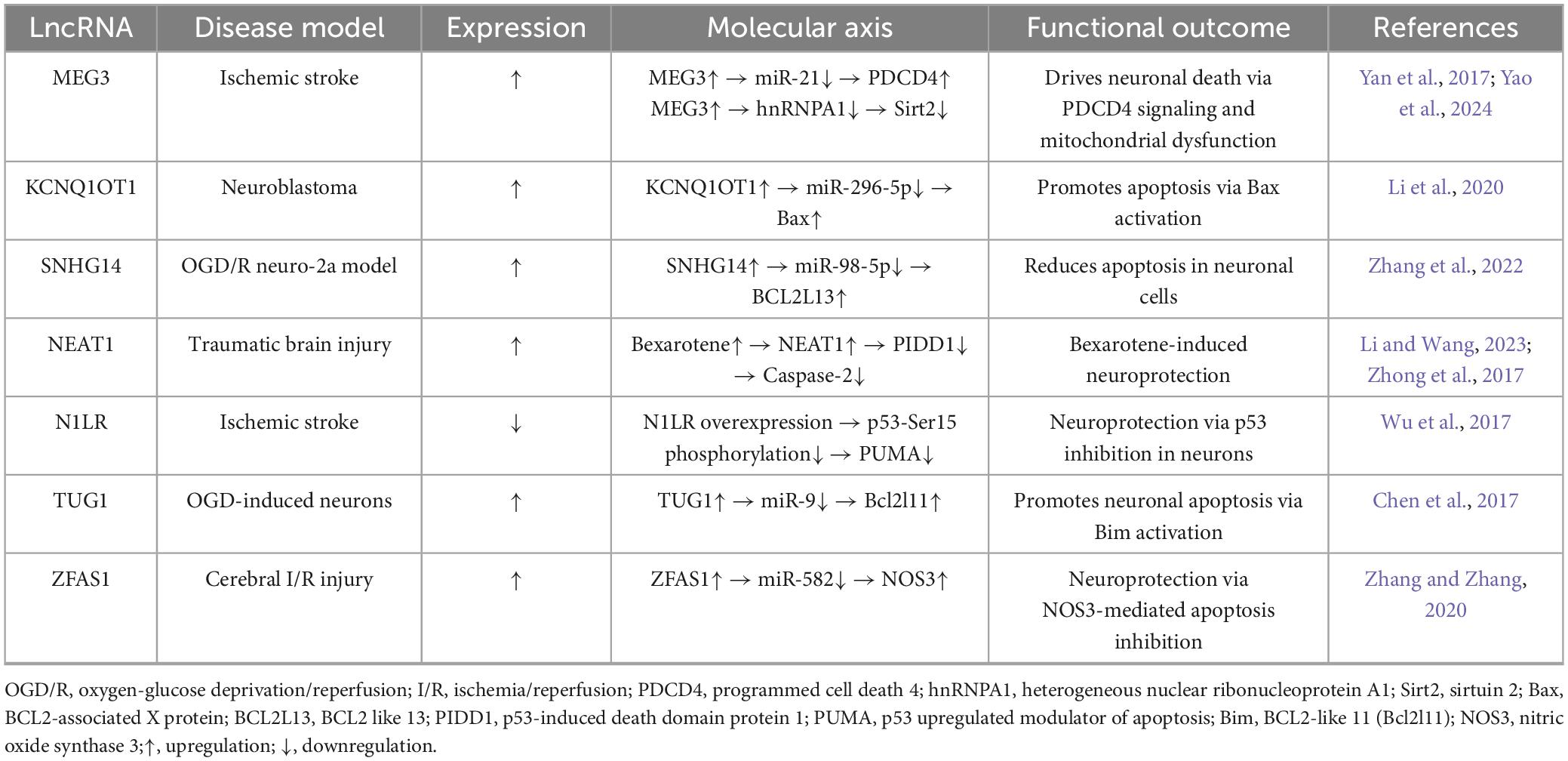
Table 1. Long non-coding RNAs (LncRNA)-driven neuronal apoptosis: targets and mechanisms.
2.2 Autophagy
Autophagy—an evolutionarily conserved lysosomal degradation system comprising three mechanistically distinct forms: microautophagy, macroautophagy, and chaperone-mediated autophagy (Dikic and Elazar, 2018)—facilitates clearance of damaged organelles and protein aggregates through double-membrane autophagosome formation. This catabolic process serves dual homeostatic and stress-responsive functions against oxidative injury, ischemia, and infection (Bernard et al., 2015; Lei and Klionsky, 2021; Mizushima et al., 2008; Siddiqui et al., 2015), governed by the PI3K-AKT1-mTOR signaling axis and core regulators (mTOR, Beclin-1, ATGs, AMPK). Crucially, while macroautophagy plays a defined pathogenic role in neurodegeneration through mediating aberrant protein clearance failure (Nixon, 2013), lncRNAs’ regulatory involvement in modulating these molecular components—particularly through PI3K-AKT1-mTOR pathway interaction—remains uncharacterized in neurological contexts.
Dysfunctional autophagy drives proteinopathic neurodegeneration across CNS disorders: Glial autophagic failure exacerbates amyloidopathy and tauopathy in Alzheimer’s disease (Litwiniuk et al., 2023), while mutant ataxin-3-induced autophagic flux disruption in Machado-Joseph disease (SCA3) is mitigated by Beclin-1 enhancement (Nascimento-Ferreira et al., 2011). LncRNAs regulate neuronal homeostasis via competitive endogenous RNA (ceRNA) mechanisms, exemplified by H19’s context-dependent duality: In adult cerebral ischemia, it sequesters miR-29a to activate ERK1/2-dependent autophagy via DUSP5 suppression, exacerbating injury (Wang et al., 2017); in neonatal hypoxic-ischemic encephalopathy, it sponges miR-29b to promote AKT3-mediated mTOR inhibition, reducing neuronal apoptosis (Zhu et al., 2022). LncRNA-driven PCD networks in Parkinson’s disease demonstrate complementary control: Downregulated SNHG1 activates autophagy via miR-221/222-p27-mTOR signaling (Qian et al., 2019), while BDNF-AS upregulation promotes autophagic apoptosis through miR-125b-5p suppression (Fan et al., 2020), concurrent with NEAT1 stabilizing PINK1 to reinforce autophagic clearance (Yan et al., 2018). This functional dichotomy—SNHG1 impeding neuronal death versus BDNF-AS accelerating degeneration—epitomizes context-dependent lncRNA governance within MPTP-induced Parkinsonian models. In cerebral ischemia, C2dat2 sequesters miR-30d-5p to activate DDIT4-dependent autophagy and apoptosis, potentiating reperfusion injury (Xu et al., 2021), while SNHG12 silencing amplifies mesenchymal stem cell efficacy by activating PI3K/AKT/mTOR signaling (Li et al., 2019). MALAT1 orchestrates microvascular protection through validated complementary routes: Sequestering miR-200c-3p in oxygen-glucose deprivation to trigger SIRT1-mediated survival (Wang et al., 2019), and sponging miR-26b during hypoxia-reoxygenation to enhance ULK2-dependent cytoprotection (Li et al., 2017). Therapeutically, Lethe enhances autophagic clearance in cortical neurons to mitigate sepsis-induced brain injury (Mai et al., 2019), while Cox2 knockdown suppresses NLRP3 inflammasome activation via coordinated autophagy regulation (Xue et al., 2019). This plasticity—H19’s duality and MALAT1’s microenvironment-specific outcomes—confirms lncRNAs as master integrators of programmed cell death. These programmable circuits provide transformative therapeutic strategies, with regulatory hierarchies mapped in Table 2.
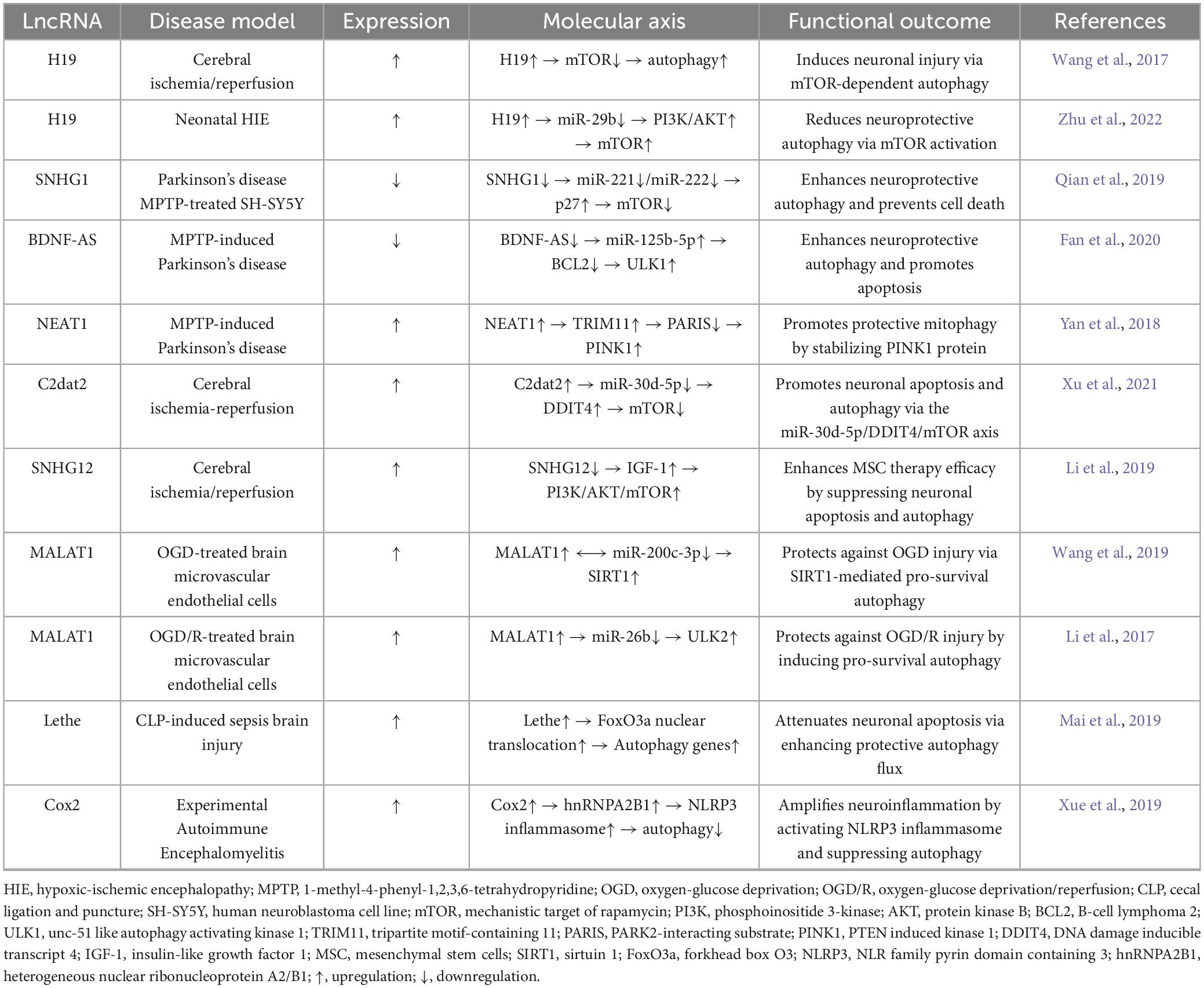
Table 2. Long non-coding RNAs (LncRNA)-driven neuronal autophagy: targets and mechanisms.
2.3 Pyroptosis
Pyroptosis represents a caspase-dependent lytic cell death pathway characterized by gasdermin-mediated membrane pore formation and proinflammatory cytokine release. Its activation occurs through two distinct routes: the canonical pathway triggered by NOD-like receptor inflammasomes (NLRP3, NLRC4, AIM2, Pyrin) activating caspase-1, and the non-canonical pathway dependent on caspase-4/5/11 (Ai et al., 2024; Bai and Zhang, 2021; Huang et al., 2021; Mi et al., 2022; Paerewijck and Lamkanfi, 2022; Pang and Vince, 2023). Both pathways converge on gasdermin-D (GSDMD) proteolysis, where liberated N-terminal fragments oligomerize to form membrane pores enabling IL-1β/IL-18 maturation and secretion—culminating in inflammatory cell lysis. Crucially, regulatory lncRNAs’ roles in modulating sensor activation thresholds or GSDMD processing remain uncharacterized in neurological contexts. Aberrant pyroptosis drives neuroinflammatory pathogenesis in Alzheimer’s, ischemic stroke, epilepsy, and Parkinson’s disease through persistent cytokine dysregulation and neuronal degeneration (Christgen and Kanneganti, 2020; Lünemann et al., 2021; Yan et al., 2022), positioning this pathway as a pathological driver and therapeutic target whose lncRNA regulatory components warrant investigation.
Emerging evidence establishes lncRNAs as master regulators of pyroptotic networks through spatiotemporally precise, cell-type-specific mechanisms. In microglia, Parkinson’s-associated SNHG1 amplifies pyroptosis via the miR-7/NLRP3 axis (Cao et al., 2018; Zhou et al., 2016), spinal injury-induced F630028O10Rik hijacks TLR4/PI3K/AKT signaling to trigger inflammasome activation (Xu et al., 2020), and diabetic hyperglycemia elevates Fendrr expression to stabilize NLRC4 through HERC2-mediated anti-ubiquitination (Wang L. Q. et al., 2021). Conversely in neurons, ischemia-activated H19 drives NLRP3/NLRP6 inflammasomes via the miR-21/PDCD4 ceRNA network (Wan et al., 2020), while hypoxia-induced MEG3 assembles AIM2 inflammasomes to activate caspase-1 cascades (Liang et al., 2020). Neurovascular dysfunction involves endothelial Xist promoting NLRP3-dependent barrier disruption via pyroptotic cleavage (Guo et al., 2022). Therapeutically, hyperbaric oxygen (HBO) suppresses H19/miR-423-5p/NLRP3 in neural stem cells to alleviate pyroptosis (Ye et al., 2022), whereas protocatechuic aldehyde inhibits Xist/NLRP3 signaling to preserve vascular integrity (Guo et al., 2022)—notably, H19 concurrently initiates microglial pyroptosis propagating neuronal deat (Wan et al., 2020). Collectively, lncRNAs architect compartmentalized neuroinflammation by coordinating microglial inflammasome priming (SNHG1, F630028O10Rik, Fendrr), neuronal pyroptosis execution (H19, MEG3), and endothelial barrier disruption (Xist), establishing a multi-tiered druggable architecture mapped in Table 3.
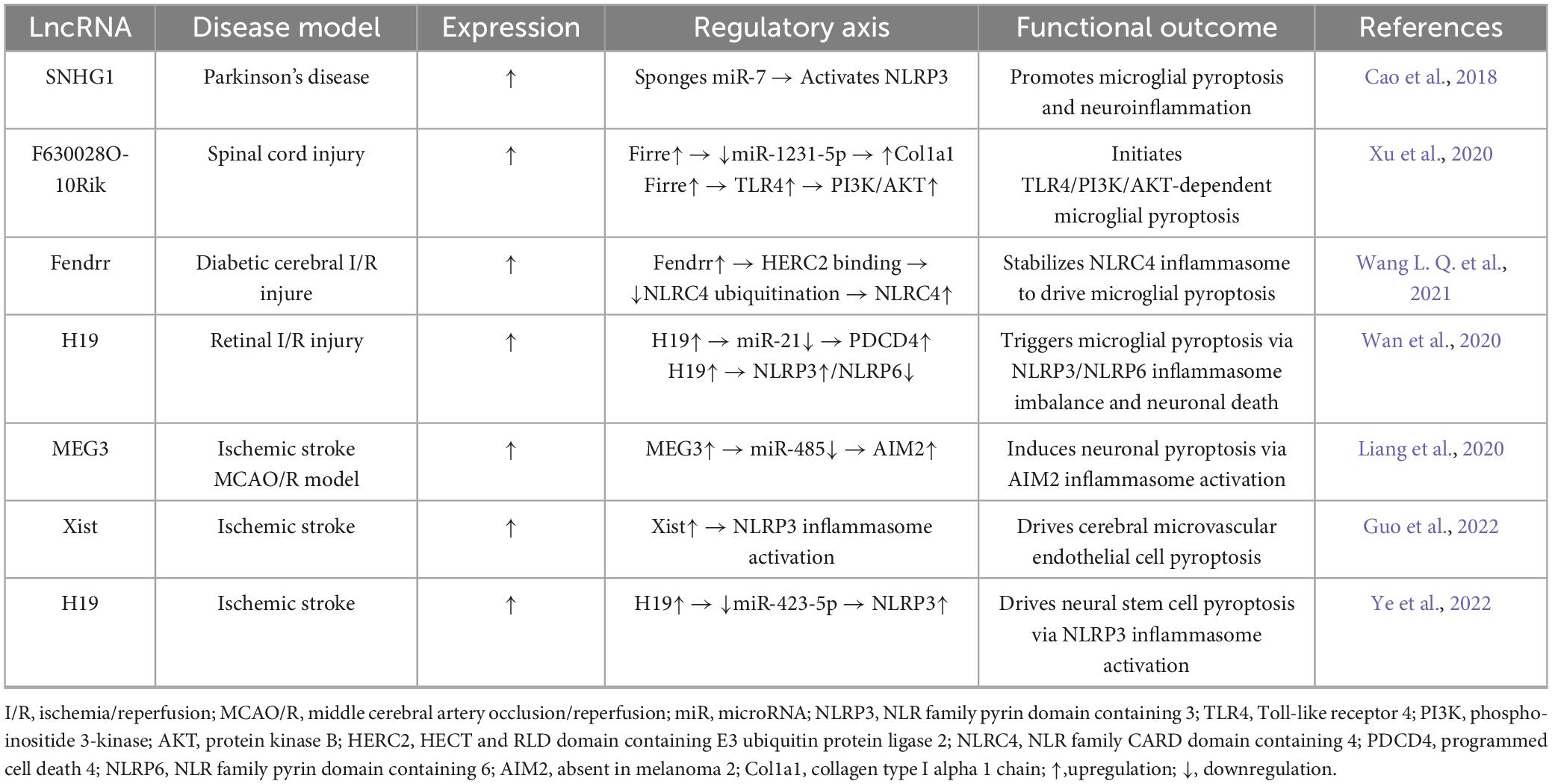
Table 3. Long non-coding RNAs (LncRNA)-driven neuronal pyroptosis: targets and mechanisms.
2.4 Ferroptosis
Ferroptosis—an iron-dependent necrotic process initiated by lipid peroxidation and redox-active iron accumulation (Dixon et al., 2012)—induces oxidative destruction of polyunsaturated fatty acid-rich phospholipid membranes. Its core machinery involves Fe2+-catalyzed Fenton reactions propagating oxidative damage (Jiang et al., 2021), with distinctive mitochondrial ultrastructural changes differentiating it from other death modalities (Zheng and Conrad, 2020). This regulated cell death pathway integrates four interdependent metabolic axes: glutathione-dependent redox balance (Dixon et al., 2012), systemic iron homeostasis (Yang et al., 2016), lipid/amino acid metabolism (Dixon et al., 2015), and mitochondrial dynamics (Gao et al., 2019). Although hepatocarcinoma studies demonstrate GSTZ1-mediated ferroptosis sensitization via NRF2/GPX4 axis suppression (Wang Q. et al., 2021), pathological iron dysregulation predominantly drives neurological degeneration, manifesting as post-traumatic epileptogenesis through astrocyte ferroptosis (Chen et al., 2020) and Alzheimer’s progression potentiated by Aβ-NRF2 axis collapse (Lane et al., 2021). Execution converges on three druggable effector hubs—glutathione peroxidase 4 (GPX4) inactivation enabling lipid peroxidation propagation (Yang et al., 2014), System Xc– dysfunction depleting glutathione reserves (Dixon et al., 2012), and p53-mediated iron metabolism reprogramming (Jiang et al., 2015)—that demonstrate epigenetic susceptibility in cancer models, designating them as priority nodes for lncRNA-driven control in neurodegeneration. Nevertheless, their disease-specific regulatory architecture—particularly lncRNA-calibrated iron homeostasis dynamics—remains uncharted, constituting a fundamental knowledge gap obstructing precision-targeted neurological therapeutics development.
Long non-coding RNAs emerge as critical ferroptosis regulators in neurological injury through divergent yet complementary molecular pathways. Clinical-translational evidence reveals PVT1 silencing reduces cerebral infarct volume in acute ischemic stroke models by inhibiting ferroptosis via miR-214/transferrin receptor 1 (TFR1)/tumor protein p53 (TP53) axis (Lu et al., 2020). Conversely, serum exosome-derived NEAT1 promotes cerebrovascular endothelial ferroptosis in septic encephalopathy through concurrent modulation of miR-9-5p/transferrin receptor (TFRC) iron transport and glutamic-oxaloacetic transaminase 1 (GOT1) signaling pathways (Wei et al., 2022). Complementing these, hyperglycemia-exacerbated hypoxia induces Meg3-dependent ferroptosis in brain microvascular endothelia via p53/GPX4 antioxidant defense disruption (Chen et al., 2021). Collectively, these systematically mapped relationships (Table 4) demonstrate lncRNAs orchestrate ferroptosis through three convergent mechanisms: (1) iron homeostasis regulation (PVT1/TFR1, NEAT1/TFRC), (2) antioxidant pathway disruption (Meg3/GPX4), and (3) cellular compartmental specificity (neuronal vs. endothelial targeting), highlighting their roles as tissue-contextual arbiters of iron-dependent cell death.
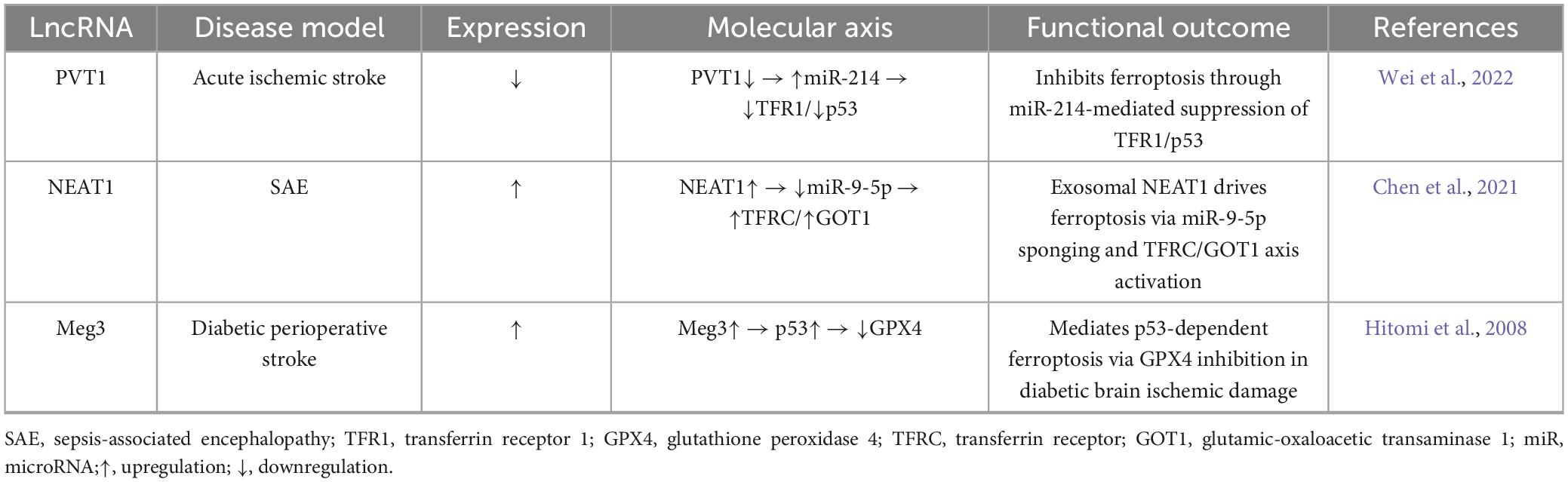
Table 4. Long non-coding RNAs (LncRNA)-driven neuronal ferroptosis: targets and mechanisms.
2.5 Necroptosis
Necroptosis represents a caspase-independent programmed cell death pathway defined by cytoplasmic swelling, plasma membrane rupture, and inflammatory mediator release. Its molecular cascade involves hierarchical activation: RIPK1-RIPK3 interaction initiates necrosome assembly, leading to RIPK3 autophosphorylation and MLKL recruitment (Hitomi et al., 2008; Tran et al., 2024). Subsequent MLKL phosphorylation induces oligomerization and plasma membrane translocation, forming pores that disrupt ionic homeostasis—the hallmark execution event. Beyond this canonical axis, RIPK3 orchestrates alternative signaling via mitochondrial phosphatase PGAM5, which stabilizes RIPK1/MLKL while activating CAMKII isoforms and ROS cascades (Cho et al., 2009; Prasad Panda et al., 2023; Zhan et al., 2022). This cross-compartmental signaling amplifies necroptotic responses, demonstrating the pathway’s capacity for diversification beyond the core RIPK1-RIPK3-MLKL axis while retaining membrane permeabilization as the terminal effector mechanism.
Necroptosis drives neuroinflammation through cytokine release and inflammasome activation, serving as a pathogenic mechanism in multiple sclerosis (MS), amyotrophic lateral sclerosis (ALS), Parkinson’s disease (PD), and Alzheimer’s disease (AD) (Richard and Mousa, 2022; Yuan et al., 2019). Mechanistic studies establish RIPK1-driven necroptosis as clinically relevant in neurological pathologies: In Alzheimer’s disease models, pharmacological inhibition of RIPK1 by necrostatin-1 attenuates tau hyperphosphorylation and Aβ-induced neuroinflammation while rescuing cognitive impairment in APP/PS1 mice (Yang et al., 2017). In parallel, stroke models reveal hypoxia-induced RIPK1/RIPK3/MLKL pathway activation driving neuronal damage, where necrostatin-1 intervention improves neurological recovery in ischemic and hemorrhagic stroke by blocking necroptotic signaling (Chen et al., 2018; Shen et al., 2017). Genetic ablation studies confirm RIP1/RIP3 deficiency protects against acute ischemic injury through dual suppression of necroptosis and neuroinflammation (Zhang et al., 2020), collectively establishing necroptosis inhibition as a promising therapeutic strategy for acute and chronic neurodegenerative conditions.
Notably, cancer biology reveals conserved lncRNA-mediated necroptosis regulation relevant to neurological contexts: Colon cancer studies identify a 6-lncRNA signature modulating TNF-α/NF-κB-driven necroptosis (Liu et al., 2022), gastric adenocarcinoma research shows 12 lncRNAs regulating caspase-8/FADD-mediated death (Luo et al., 2022), and lung adenocarcinoma models reveal 7 prognostic lncRNAs linked to MLKL ubiquitination (Lu et al., 2022). Critically, shared regulatory architecture emerges with identical necroptotic effectors (RIPK1, caspase-8, MLKL) in cancer and neurodegeneration (Najafov et al., 2019; Yuan et al., 2019), lncRNA bridging via cancer-associated regulators exhibiting CNS expression (PVT1 (Zhang et al., 2019), MALAT1 (Huang et al., 2023)), and therapeutic translatability of small-molecule inhibitors effective in both ischemic stroke (necrostatin-1 (Deng et al., 2019)) and cancer. These findings establish lncRNAs as evolutionarily conserved necroptosis modulators whose mechanistic elucidation in oncology provides direct insights for developing targeted neurotherapeutics.
2.6 Cuproptosis
Cuproptosis—a copper-dependent cell death pathway mediated by mitochondrial proteostasis collapse via copper binding to lipoylated enzymes (e.g., dihydrolipoamide S-acetyltransferase, DLAT)—involves oligomerization of lipoylated proteins and iron-sulfur cluster degradation, culminating in membrane damage via proteotoxic mechanisms (Deng et al., 2024; Tsvetkov et al., 2022). Dysregulated cuproptosis contributes to pathological manifestations in specific oncological contexts. In malignancies, dysregulation of ATP7A/B transporters mediates chemotherapy resistance, as evidenced by established prognostic models: ATP7B downregulation correlates with platinum resistance in lung cancer, while ATP7A features in cuproptosis-related gene signatures for hepatocellular carcinoma clinical stratification (Jawed and Bhatti, 2024; Shao et al., 2023). Meanwhile, in Alzheimer’s pathology, nitration modifications of β-amyloid peptides attenuate copper-mediated toxicity by inhibiting copper binding, with these copper-amyloid complexes established as neurotoxic mediators (Zhao et al., 2019). Elucidating CNS-specific cuproptosis mechanisms may reveal multipronged therapeutic opportunities, including copper-chelating agents modulating copper homeostasis (Jiang et al., 2022), metabolic modulators targeting copper-associated pathways (Gao and Zhang, 2023; Kong and Sun, 2023), and exploratory lncRNA-based interventions guided by hepatic malignancy-identified cuproptosis regulatory signatures (Mao et al., 2023). This mechanistic understanding facilitates neuroprotective intervention development for neurological disorders.
Cuproptosis signatures demonstrate significant translational potential in neuro-oncology. Studies by Zhu et al. (2024) develop an 18-gene cluster predicting low-grade glioma (LGG) survival and immunotherapy response, while subsequent research establishes lncRNA-based prognostic models incorporating 9–10 cuproptosis-associated transcripts—including functional regulators like LEF1-AS1 that suppress glioma proliferation and invasion (Chen et al., 2023; Jin et al., 2023; Wang et al., 2022). Bioinformatic analyses of Alzheimer’s datasets reveal dysregulated cuproptosis-associated genes (MTF1, NFE2L2, GLS) and lncRNAs (LY86-AS1, MIR7-3HG) (Zeng et al., 2024). These shared molecular perturbations suggest potential conservation of copper-dependent death pathways across CNS disorders, though mechanistic validation in other neuropathologies remains required. The convergence of cuproptosis-associated signatures in neuropathologies—spanning oncogenesis to proteinopathy—highlights its role as a fundamental cell death mechanism whose tissue-specific regulatory networks require prioritized validation in human neuronal models.
3 Conclusion
Long non-coding RNAs critically coordinate neural cell death in neurological disorders through multimodal regulatory networks, concurrently modulating caspase-dependent apoptosis and non-apoptotic pathways—including ferroptosis, necroptosis, and cuproptosis—via distinct molecular interactions. Persistent knowledge gaps remain regarding PCD crosstalk (e.g., apoptosis-necroptosis interplay) and context-dependent hierarchical control in CNS pathophysiology (Bertheloot et al., 2021; Ciftci et al., 2024; Malireddi et al., 2019). Notably, emerging evidence reveals functional diversification beyond canonical non-coding roles: a subset of lncRNAs encodes regulatory peptides that orchestrate cellular functions through ribosome-independent mechanisms. This creates a molecular duality that, coupled with precise spatiotemporal expression patterns, positions these molecules as ideal candidates for biomarker development and therapeutic targeting (Bakhti and Latifi-Navid, 2022; Wu et al., 2020; Xing et al., 2021). Future research must address three critical imperatives to leverage this potential: systematically mapping lncRNA-peptide interactomes in neural death pathways using advanced crosslinking immunoprecipitation; resolving region-specific regulatory networks via integrated single-cell/spatial omics; and validating evolutionary conservation of functional motifs through cross-species models. Achieving this will enable CRISPR-engineered organoid platforms to accelerate lncRNA-targeted neurotherapeutic translation.
Author contributions
ZX: Funding acquisition, Writing – original draft. CS: Writing – original draft. SH: Writing – review and editing, Conceptualization.
Funding
The author(s) declare that financial support was received for the research and/or publication of this article. This work was financially supported by grants from the Chengdu Municipal Science and Technology Program (2022-YF05-01279-SN) and the Scientific Research Project of the Chengdu Health Commission (2022414).
Acknowledgments
We would like to thank Yi Qu for her helpful comments during manuscript preparation.
Conflict of interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Generative AI statement
The authors declare that no Generative AI was used in the creation of this manuscript.
Publisher’s note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
References
Ai, Y., Meng, Y., Yan, B., Zhou, Q., and Wang, X. (2024). The biochemical pathways of apoptotic, necroptotic, pyroptotic, and ferroptotic cell death. Mol. Cell 84, 170–179. doi: 10.1016/j.molcel.2023.11.040
Amgalan, D., Chen, Y., and Kitsis, R. N. (2017). death receptor signaling in the heart: Cell survival. apoptosis, and necroptosis. Circulation 136, 743–746. doi: 10.1161/CIRCULATIONAHA.117.029566
Bai, H., and Zhang, Q. (2021). Activation of NLRP3 inflammasome and onset of Alzheimer’s disease. Front. Immunol. 12:701282. doi: 10.3389/fimmu.2021.701282
Bakhti, S. Z., and Latifi-Navid, S. (2022). Non-coding RNA-Encoded peptides/proteins in human cancer: The future for cancer therapy. Curr. Med. Chem. 29, 3819–3835. doi: 10.2174/0929867328666211111163701
Bernard, A., Jin, M., Xu, Z., and Klionsky, D. J. (2015). A large-scale analysis of autophagy-related gene expression identifies new regulators of autophagy. Autophagy 11, 2114–2122. doi: 10.1080/15548627.2015.1099796
Bertheloot, D., Latz, E., and Franklin, B. S. (2021). Necroptosis, pyroptosis and apoptosis: An intricate game of cell death. Cell Mol. Immunol. 18, 1106–1121. doi: 10.1038/s41423-020-00630-3
Cao, B., Wang, T., Qu, Q., Kang, T., and Yang, Q. (2018). Long noncoding RNA SNHG1 promotes neuroinflammation in parkinson’s disease via regulating miR-7/NLRP3 pathway. Neuroscience 388, 118–127. doi: 10.1016/j.neuroscience.2018.07.019
Chen, C., Huang, Y., Xia, P., Zhang, F., Li, L., Wang, E., et al. (2021). Long noncoding RNA Meg3 mediates ferroptosis induced by oxygen and glucose deprivation combined with hyperglycemia in rat brain microvascular endothelial cells, through modulating the p53/GPX4 axis. Eur. J. Histochem. 65:3224. doi: 10.4081/ejh.2021.3224
Chen, D., Xu, Y., Gao, X., Zhu, X., Liu, X., and Yan, D. (2023). A novel signature of cuproptosis-related lncRNAs predicts prognosis in glioma: Evidence from bioinformatic analysis and experiments. Front. Pharmacol. 14:1158723. doi: 10.3389/fphar.2023.1158723
Chen, S., Chen, Y., Zhang, Y., Kuang, X., Liu, Y., Guo, M., et al. (2020). Iron metabolism and ferroptosis in epilepsy. Front. Neurosci. 14:601193. doi: 10.3389/fnins.2020.601193
Chen, S., Wang, M., Yang, H., Mao, L., He, Q., Jin, H., et al. (2017). LncRNA TUG1 sponges microRNA-9 to promote neurons apoptosis by up-regulated Bcl2l11 under ischemia. Biochem. Biophys. Res. Commun. 485, 167–173. doi: 10.1016/j.bbrc.2017.02.043
Chen, Y., Zhang, L., Yu, H., Song, K., Shi, J., Chen, L., et al. (2018). Necrostatin-1 improves long-term functional recovery through protecting oligodendrocyte precursor cells after transient focal cerebral ischemia in mice. Neuroscience 371, 229–241. doi: 10.1016/j.neuroscience.2017.12.007
Cho, Y. S., Challa, S., Moquin, D., Genga, R., Ray, T. D., Guildford, M., et al. (2009). Phosphorylation-driven assembly of the RIP1-RIP3 complex regulates programmed necrosis and virus-induced inflammation. Cell 137, 1112–1123. doi: 10.1016/j.cell.2009.05.037
Christgen, S., and Kanneganti, T. D. (2020). Inflammasomes and the fine line between defense and disease. Curr. Opin. Immunol. 62, 39–44. doi: 10.1016/j.coi.2019.11.007
Ciftci, Y. C., Vatansever, İE., and Akgül, B. (2024). Unraveling the intriguing interplay: Exploring the role of lncRNAs in caspase-independent cell death. Wiley Interdiscip. Rev. RNA. 15:e1862. doi: 10.1002/wrna.1862
Deng, J., Zhuang, H., Shao, S., Zeng, X., Xue, P., Bai, T., et al. (2024). Mitochondrial-Targeted copper delivery for cuproptosis-based synergistic cancer therapy. Adv. Healthc. Mater. 13:e2304522. doi: 10.1002/adhm.202304522
Deng, X. X., Li, S. S., and Sun, F. Y. (2019). Necrostatin-1 prevents necroptosis in brains after ischemic stroke via inhibition of RIPK1-Mediated RIPK3/MLKL signaling. Aging Dis. 10, 807–817. doi: 10.14336/AD.2018.0728
Dikic, I., and Elazar, Z. (2018). Mechanism and medical implications of mammalian autophagy. Nat. Rev. Mol. Cell Biol. 19, 349–364. doi: 10.1038/s41580-018-0003-4
Dixon, S. J., Lemberg, K. M., Lamprecht, M. R., Skouta, R., Zaitsev, E. M., Gleason, C. E., et al. (2012). Ferroptosis: An iron-dependent form of nonapoptotic cell death. Cell 149, 1060–1072. doi: 10.1016/j.cell.2012.03.042
Dixon, S. J., Winter, G. E., Musavi, L. S., Lee, Snijder, B., Rebsamen, M., et al. (2015). Human haploid cell genetics reveals roles for lipid metabolism genes in nonapoptotic cell death. ACS Chem. Biol. 10, 1604–1609. doi: 10.1021/acschembio.5b00245
Fan, Y., Zhao, X., Lu, K., and Cheng, G. (2020). LncRNA BDNF-AS promotes autophagy and apoptosis in MPTP-induced Parkinson’s disease via ablating microRNA-125b-5p. Brain Res. Bull. 157, 119–127. doi: 10.1016/j.brainresbull.2020.02.003
Fernández, A., Ordóñez, R., Reiter, R. J., González-Gallego, J., and Mauriz, J. L. (2015). Melatonin and endoplasmic reticulum stress: Relation to autophagy and apoptosis. J. Pineal Res. 59, 292–307. doi: 10.1111/jpi.12264
Ferrer, J., and Dimitrova, N. (2024). Transcription regulation by long non-coding RNAs: Mechanisms and disease relevance. Nat. Rev. Mol. Cell Biol. 25, 396–415. doi: 10.1038/s41580-023-00694-9
Gao, L., and Zhang, A. (2023). Copper-instigated modulatory cell mortality mechanisms and progress in oncological treatment investigations. Front. Immunol. 14:1236063. doi: 10.3389/fimmu.2023.1236063
Gao, M., Yi, J., Zhu, J., Minikes, A. M., Monian, P., Thompson, C. B., et al. (2019). Role of mitochondria in ferroptosis. Mol. Cell 73, 354–363.e3. doi: 10.1016/j.molcel.2018.10.042
Ghafouri-Fard, S., Hussen, B. M., Jamali, E., Branicki, W., Taheri, M., and Akbari Dilmaghani, N. (2022). Role of lncRNAs and circRNAs in epilepsy. Ageing Res. Rev. 82:101749. doi: 10.1016/j.arr.2022.101749
Guo, Y., Yang, J. H., He, Y., Zhou, H. F., Wang, Y., Ding, Z. S., et al. (2022). Protocatechuic aldehyde prevents ischemic injury by attenuating brain microvascular endothelial cell pyroptosis via lncRNA Xist. Phytomedicine 94:153849. doi: 10.1016/j.phymed.2021.153849
He, Y., Jing, Y., Wei, F., Tang, Y., Yang, L., Luo, J., et al. (2018). Long non-coding RNA PVT1 predicts poor prognosis and induces radioresistance by regulating DNA repair and cell apoptosis in nasopharyngeal carcinoma. Cell Death Dis. 9:235. doi: 10.1038/s41419-018-0265-y
Herman, A. B., Tsitsipatis, D., and Gorospe, M. (2022). Integrated lncRNA function upon genomic and epigenomic regulation. Mol. Cell 82, 2252–2266. doi: 10.1016/j.molcel.2022.05.027
Hitomi, J., Christofferson, D. E., Ng, A., Yao, J., Degterev, A., Xavier, R. J., et al. (2008). Identification of a molecular signaling network that regulates a cellular necrotic cell death pathway. Cell 135, 1311–1323. doi: 10.1016/j.cell.2008.10.044
Huang, S., Hou, D., Zhang, L., Pei, C., Liang, J., Li, J., et al. (2023). LncRNA MALAT1 promoted neuronal necroptosis in cerebral ischemia-reperfusion mice by stabilizing HSP90. Neurochem. Res. 48, 3457–3471. doi: 10.1007/s11064-023-03991-z
Huang, Y., Xu, W., and Zhou, R. (2021). NLRP3 inflammasome activation and cell death. Cell Mol. Immunol. 18, 2114–2127. doi: 10.1038/s41423-021-00740-6
Jawed, R., and Bhatti, H. (2024). Cuproptosis in lung cancer: Therapeutic options and prognostic models. Apoptosis 29, 1393–1398. doi: 10.1007/s10495-024-01978-x
Jiang, A., Luo, P., Chen, M., Fang, Y., Liu, B., Wu, Z., et al. (2022). A new thinking: Deciphering the aberrance and clinical implication of copper-death signatures in clear cell renal cell carcinoma. Cell Biosci. 12:209. doi: 10.1186/s13578-022-00948-7
Jiang, L., Kon, N., Li, T., Wang, S. J., Su, T., Hibshoosh, H., et al. (2015). Ferroptosis as a p53-mediated activity during tumour suppression. Nature 520, 57–62. doi: 10.1038/nature14344
Jiang, X., Stockwell, B. R., and Conrad, M. (2021). Ferroptosis: Mechanisms, biology and role in disease. Nat. Rev. Mol. Cell Biol. 22, 266–282. doi: 10.1038/s41580-020-00324-8
Jin, J., Li, R., Guo, G., Chen, Y., Li, Z. A., and Zheng, J. (2023). Studies on the therapeutic and prognostic biomarkers of glioma using a novel cuproptosis-related IncRNA signature and validation in glioma. J. Environ. Pathol. Toxicol. Oncol. 42, 53–70. doi: 10.1615/JEnvironPatholToxicolOncol.2023047159
Jing, H., Liu, L., Jia, Y., Yao, H., and Ma, F. (2019). Overexpression of the long non-coding RNA oprm1 alleviates apoptosis from cerebral ischemia-reperfusion injury through the oprm1/miR-155/GATA3 axis. Artif. Cells Nanomed. Biotechnol. 47, 2431–2439. doi: 10.1080/21691401.2019.1626408
Kadakkuzha, B. M., Liu, X. A., McCrate, J., Shankar, G., Rizzo, V., Afinogenova, A., et al. (2015). Transcriptome analyses of adult mouse brain reveal enrichment of lncRNAs in specific brain regions and neuronal populations. Front. Cell Neurosci. 9:63. doi: 10.3389/fncel.2015.00063
Kaushik, K., Leonard, V. E., Kv, S., Lalwani, M. K., Jalali, S., Patowary, A., et al. (2013). Dynamic expression of long non-coding RNAs (lncRNAs) in adult zebrafish. PLoS One 8:e83616. doi: 10.1371/journal.pone.0083616
Khoshnam, S. E., Moalemnia, A., Anbiyaee, O., Farzaneh, M., and Ghaderi, S. (2024). LncRNA MALAT1 and ischemic stroke: Pathogenesis and opportunities. Mol. Neurobiol. 61, 4369–4380. doi: 10.1007/s12035-023-03853-3
Kitagawa, M., Kitagawa, K., Kotake, Y., Niida, H., and Ohhata, T. (2013). Cell cycle regulation by long non-coding RNAs. Cell Mol. Life Sci. 70, 4785–4794. doi: 10.1007/s00018-013-1423-0
Kleaveland, B., Shi, C. Y., Stefano, J., and Bartel, D. P. (2018). A network of noncoding regulatory RNAs acts in the mammalian brain. Cell 174, 350–362.e17. doi: 10.1016/j.cell.2018.05.022
Kong, R., and Sun, G. (2023). Targeting copper metabolism: A promising strategy for cancer treatment. Front. Pharmacol. 14:1203447. doi: 10.3389/fphar.2023.1203447
Lane, D. J. R., Metselaar, B., Greenough, M., Bush, A. I., and Ayton, S. J. (2021). Ferroptosis and NRF2: An emerging battlefield in the neurodegeneration of Alzheimer’s disease. Essays Biochem. 65, 925–940. doi: 10.1042/EBC20210017
Lei, Y., and Klionsky, D. J. (2021). The emerging roles of autophagy in human diseases. Biomedicines 9:1651. doi: 10.3390/biomedicines9111651
Li, K., and Wang, Z. (2023). lncRNA NEAT1: Key player in neurodegenerative diseases. Ageing Res. Rev. 86:101878. doi: 10.1016/j.arr.2023.101878
Li, M. M., Liu, X. H., Zhao, Y. C., Ma, X. Y., Zhou, Y. C., Zhao, Y. X., et al. (2020). Long noncoding RNA KCNQ1OT1 promotes apoptosis in neuroblastoma cells by regulating miR-296-5p/Bax axis. FEBS J. 287, 561–577. doi: 10.1111/febs.15047
Li, R., Wang, X., Zhu, C., and Wang, K. (2022). lncRNA PVT1: A novel oncogene in multiple cancers. Cell Mol. Biol. Lett. 27:84. doi: 10.1186/s11658-022-00385-x
Li, Y., Guo, S., Liu, W., Jin, T., Li, X., He, X., et al. (2019). Silencing of SNHG12 enhanced the effectiveness of MSCs in alleviating ischemia/reperfusion injuries via the PI3K/AKT/mTOR signaling pathway. Front. Neurosci. 13:645. doi: 10.3389/fnins.2019.00645
Li, Z., Li, J., and Tang, N. (2017). Long noncoding RNA Malat1 is a potent autophagy inducer protecting brain microvascular endothelial cells against oxygen-glucose deprivation/reoxygenation-induced injury by sponging miR-26b and upregulating ULK2 expression. Neuroscience 354, 1–10. doi: 10.1016/j.neuroscience.2017.04.017
Liang, J., Wang, Q., Li, J. Q., Guo, T., and Yu, D. (2020). Long non-coding RNA MEG3 promotes cerebral ischemia-reperfusion injury through increasing pyroptosis by targeting miR-485/AIM2 axis. Exp. Neurol. 325:113139. doi: 10.1016/j.expneurol.2019.113139
Litwiniuk, A., Juszczak, G. R., Stankiewicz, A. M., and Urbańska, K. (2023). The role of glial autophagy in Alzheimer’s disease. Mol. Psychiatry 28, 4528–4539. doi: 10.1038/s41380-023-02242-5
Liu, L., Huang, L., Chen, W., Zhang, G., Li, Y., Wu, Y., et al. (2022). Comprehensive analysis of necroptosis-related long noncoding RNA immune infiltration and prediction of prognosis in patients with colon cancer. Front. Mol. Biosci. 9:811269. doi: 10.3389/fmolb.2022.811269
Lu, J., Xu, F., and Lu, H. (2020). LncRNA PVT1 regulates ferroptosis through miR-214-mediated TFR1 and p53. Life Sci. 260:118305. doi: 10.1016/j.lfs.2020.118305
Lu, Y., Luo, X., Wang, Q., Chen, J., Zhang, X., Li, Y., et al. (2022). A novel necroptosis-related lncRNA signature predicts the prognosis of lung adenocarcinoma. Front. Genet. 13:862741. doi: 10.3389/fgene.2022.862741
Lünemann, J. D., Malhotra, S., Shinohara, M. L., Montalban, X., and Comabella, M. (2021). Targeting inflammasomes to treat neurological diseases. Ann. Neurol. 90, 177–188. doi: 10.1002/ana.26158
Luo, L., Li, L., Liu, L., Feng, Z., Zeng, Q., Shu, X., et al. (2022). A necroptosis-related lncRNA-Based signature to predict prognosis and probe molecular characteristics of stomach adenocarcinoma. Front. Genet. 13:833928. doi: 10.3389/fgene.2022.833928
Mai, C., Qiu, L., Zeng, Y., and Jian, H. G. (2019). LncRNA Lethe protects sepsis-induced brain injury via regulating autophagy of cortical neurons. Eur. Rev. Med. Pharmacol. Sci. 23, 4858–4864. doi: 10.26355/eurrev_201906_18073
Malireddi, R. K. S., Kesavardhana, S., and Kanneganti, T. D. (2019). ZBP1 and TAK1: Master regulators of NLRP3 inflammasome/pyroptosis, apoptosis, and necroptosis (PAN-optosis). Front. Cell Infect. Microbiol. 9:406. doi: 10.3389/fcimb.2019.00406
Mao, Z., Nie, Y., Jia, W., Wang, Y., Li, J., Zhang, T., et al. (2023). Revealing prognostic and immunotherapy-sensitive characteristics of a novel cuproptosis-related LncRNA model in hepatocellular carcinoma patients by genomic analysis. Cancers (Basel) 15:544. doi: 10.3390/cancers15020544
Mi, L., Min, X., Chai, Y., Zhang, J., and Chen, X. (2022). NLRP1 inflammasomes: A potential target for the treatment of several types of brain injury. Front. Immunol. 13:863774. doi: 10.3389/fimmu.2022.863774
Mizushima, N., Levine, B., Cuervo, A. M., and Klionsky, D. J. (2008). Autophagy fights disease through cellular self-digestion. Nature 451, 1069–1075. doi: 10.1038/nature06639
Najafov, A., Mookhtiar, A. K., Luu, H. S., Ordureau, A., Pan, H., Amin, P. P., et al. (2019). TAM kinases promote necroptosis by regulating oligomerization of MLKL. Mol. Cell 75, 457–468.e4. doi: 10.1016/j.molcel.2019.05.022
Nascimento-Ferreira, I., Santos-Ferreira, T., Sousa-Ferreira, L., Auregan, G., Onofre, I., Alves, S., et al. (2011). Overexpression of the autophagic beclin-1 protein clears mutant ataxin-3 and alleviates Machado-Joseph disease. Brain 134(Pt 5), 1400–1415. doi: 10.1093/brain/awr047
Nitsche, A., and Stadler, P. F. (2017). Evolutionary clues in lncRNAs. Wiley Interdiscip. Rev. RNA. 8:e1376. doi: 10.1002/wrna.1376
Nixon, R. A. (2013). The role of autophagy in neurodegenerative disease. Nat. Med. 19, 983–997. doi: 10.1038/nm.3232
Paerewijck, O., and Lamkanfi, M. (2022). The human inflammasomes. Mol. Aspects Med. 88:101100. doi: 10.1016/j.mam.2022.101100
Pang, J., and Vince, J. E. (2023). The role of caspase-8 in inflammatory signalling and pyroptotic cell death. Semin. Immunol. 70:101832. doi: 10.1016/j.smim.2023.101832
Prasad Panda, S., Kesharwani, A., Prasanna Mallick, S., Prasanth, D., Kumar Pasala, P., and Bharadwaj Tatipamula, V. (2023). Viral-induced neuronal necroptosis: Detrimental to brain function and regulation by necroptosis inhibitors. Biochem. Pharmacol. 213:115591. doi: 10.1016/j.bcp.2023.115591
Qian, C., Ye, Y., Mao, H., Yao, L., Sun, X., Wang, B., et al. (2019). Downregulated lncRNA-SNHG1 enhances autophagy and prevents cell death through the miR-221/222 /p27/mTOR pathway in Parkinson’s disease. Exp. Cell Res. 384:111614. doi: 10.1016/j.yexcr.2019.111614
Richard, R., and Mousa, S. (2022). Necroptosis in Alzheimer’s disease: Potential therapeutic target. Biomed. Pharmacother. 152:113203. doi: 10.1016/j.biopha.2022.113203
Riley, J. S., and Bock, F. J. (2022). Voices from beyond the grave: The impact of apoptosis on the microenvironment. Biochim. Biophys. Acta Mol. Cell Res. 1869:119341. doi: 10.1016/j.bbamcr.2022.119341
Ruffo, P., De Amicis, F., Giardina, E., and Conforti, F. L. (2023). Long-noncoding RNAs as epigenetic regulators in neurodegenerative diseases. Neural Regen. Res. 18, 1243–1248. doi: 10.4103/1673-5374.358615
Schwarzer, R., Laurien, L., and Pasparakis, M. (2020). New insights into the regulation of apoptosis, necroptosis, and pyroptosis by receptor interacting protein kinase 1 and caspase-8. Curr. Opin. Cell Biol. 63, 186–193. doi: 10.1016/j.ceb.2020.02.004
Shao, R. N., Bai, K. H., Huang, Q. Q., Chen, S. L., Huang, X., and Dai, Y. J. (2023). A novel prognostic prediction model of cuprotosis-related genes signature in hepatocellular carcinoma. Front. Cell Dev. Biol. 11:1180625. doi: 10.3389/fcell.2023.1180625
Shen, H., Liu, C., Zhang, D., Yao, X., Zhang, K., Li, H., et al. (2017). Role for RIP1 in mediating necroptosis in experimental intracerebral hemorrhage model both in vivo and in vitro. Cell Death Dis. 8:e2641. doi: 10.1038/cddis.2017.58
Siddiqui, M. A., Mukherjee, S., Manivannan, P., and Malathi, K. (2015). RNase L cleavage products promote switch from autophagy to apoptosis by caspase-mediated cleavage of beclin-1. Int. J. Mol. Sci. 16, 17611–17636. doi: 10.3390/ijms160817611
Tran, H. T., Kratina, T., Coutansais, A., Michalek, D., Hogan, B. M., Lawlor, K. E., et al. (2024). RIPK3 cleavage is dispensable for necroptosis inhibition but restricts NLRP3 inflammasome activation. Cell Death Differ. 31, 662–671. doi: 10.1038/s41418-024-01281-x
Tsvetkov, P., Coy, S., Petrova, B., Dreishpoon, M., Verma, A., Abdusamad, M., et al. (2022). Copper induces cell death by targeting lipoylated TCA cycle proteins. Science 375, 1254–1261. doi: 10.1126/science.abf0529
Van Opdenbosch, N., and Lamkanfi, M. (2019). Caspases in cell death, inflammation, and disease. Immunity 50, 1352–1364. doi: 10.1016/j.immuni.2019.05.020
Wan, P., Su, W., Zhang, Y., Li, Z., Deng, C., Li, J., et al. (2020). LncRNA H19 initiates microglial pyroptosis and neuronal death in retinal ischemia/reperfusion injury. Cell Death Differ. 27, 176–191. doi: 10.1038/s41418-019-0351-4
Wang, J., Cao, B., Han, D., Sun, M., and Feng, J. (2017). Long non-coding RNA H19 induces cerebral ischemia reperfusion injury via activation of autophagy. Aging Dis. 8, 71–84. doi: 10.14336/AD.2016.0530
Wang, L. Q., Zheng, Y. Y., Zhou, H. J., Zhang, X. X., Wu, P., and Zhu, S. M. (2021). LncRNA-Fendrr protects against the ubiquitination and degradation of NLRC4 protein through HERC2 to regulate the pyroptosis of microglia. Mol. Med. 27:39. doi: 10.1186/s10020-021-00299-y
Wang, Q., Bin, C., Xue, Q., Gao, Q., Huang, A., Wang, K., et al. (2021). GSTZ1 sensitizes hepatocellular carcinoma cells to sorafenib-induced ferroptosis via inhibition of NRF2/GPX4 axis. Cell Death Dis. 12:426. doi: 10.1038/s41419-021-03718-4
Wang, L., Li, Y., Wang, Y., Li, J., Sun, Y., Chen, J., et al. (2022). Identification of cuproptosis-related lncRNAs for prognosis and immunotherapy in glioma. J. Cell Mol. Med. 26, 5820–5831. doi: 10.1111/jcmm.17603
Wang, S., Han, X., Mao, Z., Xin, Y., Maharjan, S., and Zhang, B. (2019). MALAT1 lncRNA induces autophagy and protects brain microvascular endothelial cells against oxygen-glucose deprivation by binding to miR-200c-3p and upregulating SIRT1 expression. Neuroscience 397, 116–126. doi: 10.1016/j.neuroscience.2018.11.024
Wei, X. B., Jiang, W. Q., Zeng, J. H., Huang, L. Q., Ding, H. G., Jing, Y. W., et al. (2022). Exosome-Derived lncRNA NEAT1 exacerbates sepsis-associated encephalopathy by promoting ferroptosis through regulating miR-9-5p/TFRC and GOT1 axis. Mol. Neurobiol. 59, 1954–1969. doi: 10.1007/s12035-022-02738-1
Wu, J., Yu, H., Huang, H., Shu, P., and Peng, X. (2021). Functions of noncoding RNAs in glial development. Dev. Neurobiol. 81, 877–891. doi: 10.1002/dneu.22848
Wu, P., Mo, Y., Peng, M., Tang, T., Zhong, Y., Deng, X., et al. (2020). Emerging role of tumor-related functional peptides encoded by lncRNA and circRNA. Mol. Cancer 19:22. doi: 10.1186/s12943-020-1147-3
Wu, Y. Y., and Kuo, H. C. (2020). Functional roles and networks of non-coding RNAs in the pathogenesis of neurodegenerative diseases. J. Biomed. Sci. 27:49. doi: 10.1186/s12929-020-00636-z
Wu, Z., Wu, P., Zuo, X., Yu, N., Qin, Y., Xu, Q., et al. (2017). LncRNA-N1LR enhances neuroprotection against ischemic stroke probably by inhibiting p53 phosphorylation. Mol. Neurobiol. 54, 7670–7685. doi: 10.1007/s12035-016-0246-z
Xing, J., Liu, H., Jiang, W., and Wang, L. (2021). LncRNA-Encoded peptide: Functions and predicting methods. Front. Oncol. 10:622294. doi: 10.3389/fonc.2020.622294
Xu, Q., Guohui, M., Li, D., Bai, F., Fang, J., Zhang, G., et al. (2021). lncRNA C2dat2 facilitates autophagy and apoptosis via the miR-30d-5p/DDIT4/mTOR axis in cerebral ischemia-reperfusion injury. Aging (Albany NY) 13, 11315–11335. doi: 10.18632/aging.202824
Xu, S., Wang, J., Jiang, J., Song, J., Zhu, W., Zhang, F., et al. (2020). TLR4 promotes microglial pyroptosis via lncRNA-F630028O10Rik by activating PI3K/AKT pathway after spinal cord injury. Cell Death Dis. 11:693. doi: 10.1038/s41419-020-02824-z
Xue, Z., Zhang, Z., Liu, H., Li, W., Guo, X., Zhang, Z., et al. (2019). lincRNA-Cox2 regulates NLRP3 inflammasome and autophagy mediated neuroinflammation. Cell Death Differ. 26, 130–145. doi: 10.1038/s41418-018-0105-8
Yan, H., Rao, J., Yuan, J., Gao, L., Huang, W., Zhao, L., et al. (2017). Long non-coding RNA MEG3 functions as a competing endogenous RNA to regulate ischemic neuronal death by targeting miR-21/PDCD4 signaling pathway. Cell Death Dis. 8:3211. doi: 10.1038/s41419-017-0047-y
Yan, W. T., Yang, Y. D., Hu, X. M., Ning, W. Y., Liao, L. S., Lu, S., et al. (2022). Do pyroptosis, apoptosis, and necroptosis (PANoptosis) exist in cerebral ischemia? evidence from cell and rodent studies. Neural Regen. Res. 17, 1761–1768. doi: 10.4103/1673-5374.331539
Yan, W., Chen, Z. Y., Chen, J. Q., and Chen, H. M. (2018). LncRNA NEAT1 promotes autophagy in MPTP-induced Parkinson’s disease through stabilizing PINK1 protein. Biochem. Biophys. Res. Commun. 496, 1019–1024. doi: 10.1016/j.bbrc.2017.12.149
Yang, R., Yang, B., Liu, W., Tan, C., Chen, H., and Wang, X. (2023). Emerging role of non-coding RNAs in neuroinflammation mediated by microglia and astrocytes. J. Neuroinflamm. 20:173. doi: 10.1186/s12974-023-02856-0
Yang, S. H., Lee, D. K., Shin, J., Lee, S., Baek, S., Kim, J., et al. (2017). Nec-1 alleviates cognitive impairment with reduction of Aβ and tau abnormalities in APP/PS1 mice. EMBO Mol. Med. 9, 61–77. doi: 10.15252/emmm.201606566
Yang, W. S., Kim, K. J., Gaschler, M. M., Patel, M., Shchepinov, M. S., and Stockwell, B. R. (2016). Peroxidation of polyunsaturated fatty acids by lipoxygenases drives ferroptosis. Proc. Natl. Acad. Sci. U. S. A. 113, E4966–E4975. doi: 10.1073/pnas.1603244113
Yang, W. S., SriRamaratnam, R., Welsch, M. E., Shimada, K., Skouta, R., Viswanathan, V. S., et al. (2014). Regulation of ferroptotic cancer cell death by GPX4. Cell 156, 317–331. doi: 10.1016/j.cell.2013.12.010
Yao, L., Peng, P., Ding, T., Yi, J., and Liang, J. (2024). m6A-Induced lncRNA MEG3 promotes cerebral ischemia-reperfusion injury via modulating oxidative stress and mitochondrial dysfunction by hnRNPA1/Sirt2 axis. Mol. Neurobiol. 61, 6893–6908. doi: 10.1007/s12035-024-04005-x
Yao, P., Li, Y. L., Chen, Y., Shen, W., Wu, K. Y., and Xu, W. H. (2020). Overexpression of long non-coding RNA Rian attenuates cell apoptosis from cerebral ischemia-reperfusion injury via Rian/miR-144-3p/GATA3 signaling. Gene 737:144411. doi: 10.1016/j.gene.2020.144411
Ye, Y., Feng, Z., Tian, S., Yang, Y., Jia, Y., Wang, G., et al. (2022). HBO alleviates neural stem cell pyroptosis via lncRNA-H19/miR-423-5p/NLRP3 axis and improves neurogenesis after oxygen glucose deprivation. Oxid. Med. Cell Longev. 2022:9030771. doi: 10.1155/2022/9030771
Yuan, J., Amin, P., and Ofengeim, D. (2019). Necroptosis and RIPK1-mediated neuroinflammation in CNS diseases. Nat. Rev. Neurosci. 20, 19–33. doi: 10.1038/s41583-018-0093-1
Zeng, Y., Qian, S., Cao, Y., and Xiao, W. (2024). Unravelling the complex interplay of cuproptosis, lncRNAs, and immune infiltration in Alzheimer’s disease: A step towards novel therapeutic targets. Ann. Hum. Biol. 51:2342531. doi: 10.1080/03014460.2024.2342531
Zhan, Q., Jeon, J., Li, Y., Huang, Y., Xiong, J., Wang, Q., et al. (2022). CAMK2/CaMKII activates MLKL in short-term starvation to facilitate autophagic flux. Autophagy 18, 726–744. doi: 10.1080/15548627.2021.1954348
Zhang, G., Guo, J., Zeng, J., Zhang, X., Chen, R., Wang, G., et al. (2022). LncRNA SNHG14 is beneficial to oxygen glucose deprivation/reoxygenation-induced neuro-2a cell injury via mir-98-5p sequestration-caused BCL2L13 upregulation. Metab. Brain Dis. 37, 2005–2016. doi: 10.1007/s11011-022-01009-1
Zhang, Y., and Zhang, Y. (2020). lncRNA ZFAS1 improves neuronal injury and inhibits inflammation, oxidative stress, and apoptosis by sponging mir-582 and upregulating NOS3 expression in cerebral ischemia/reperfusion injury. Inflammation 43, 1337–1350. doi: 10.1007/s10753-020-01212-1
Zhang, Y., Li, M., Li, X., Zhang, H., Wang, L., Wu, X., et al. (2020). Catalytically inactive RIP1 and RIP3 deficiency protect against acute ischemic stroke by inhibiting necroptosis and neuroinflammation. Cell Death Dis. 11:565. doi: 10.1038/s41419-020-02770-w
Zhang, Y., Yang, G., and Luo, Y. (2019). Long non-coding RNA PVT1 promotes glioma cell proliferation and invasion by targeting miR-200a. Exp. Ther. Med. 17, 1337–1345. doi: 10.3892/etm.2018.7083
Zhao, J., Gao, W., Yang, Z., Li, H., and Gao, Z. (2019). Nitration of amyloid-β peptide (1-42) as a protective mechanism for the amyloid-β peptide (1-42) against copper ion toxicity. J. Inorg. Biochem. 190, 15–23. doi: 10.1016/j.jinorgbio.2018.10.005
Zheng, J., and Conrad, M. (2020). The metabolic underpinnings of ferroptosis. Cell Metab. 32, 920–937. doi: 10.1016/j.cmet.2020.10.011
Zhong, J., Jiang, L., Huang, Z., Zhang, H., Cheng, C., Liu, H., et al. (2017). The long non-coding RNA Neat1 is an important mediator of the therapeutic effect of bexarotene on traumatic brain injury in mice. Brain Behav. Immun. 65, 183–194. doi: 10.1016/j.bbi.2017.05.001
Zhou, Y., Lu, M., Du, R. H., Qiao, C., Jiang, C. Y., Zhang, K. Z., et al. (2016). MicroRNA-7 targets Nod-like receptor protein 3 inflammasome to modulate neuroinflammation in the pathogenesis of Parkinson’s disease. Mol. Neurodegener. 11:28. doi: 10.1186/s13024-016-0094-3
Zhu, H., Wang, L., Chen, J., Shen, H., and Chen, Z. (2022). Mechanisms underlying abnormal expression of lncRNA H19 in neonatal hypoxic-ischemic encephalopathy. Am. J. Perinatol. 39, 844–852. doi: 10.1055/s-0040-1718947
Keywords: long non-coding RNA, central nervous system disease, neural cells, programmed cell death, cell death
Citation: Xiong ZH, Sun C and Huang SY (2025) LncRNA-driven programmed cell death networks: new therapeutic targets for neurological disorders. Front. Mol. Neurosci. 18:1635119. doi: 10.3389/fnmol.2025.1635119
Received: 26 May 2025; Accepted: 07 July 2025;
Published: 24 July 2025.
Edited by:
Robert John Vandenberg, The University of Sydney, AustraliaReviewed by:
Paulina Carriba, Centre for Genomic Regulation (CRG), SpainCopyright © 2025 Xiong, Sun and Huang. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Shiyong Huang, aHN5X2NoZW5nZHVwaWN1QDEyNi5jb20=