 Melinda Magyar1*
Melinda Magyar1* Gábor Sipka1Ildikó Domonkos1Xiaojun Chen2Xu Wang2
Gábor Sipka1Ildikó Domonkos1Xiaojun Chen2Xu Wang2 Guangye Han2Jian-Ren Shen2,3
Guangye Han2Jian-Ren Shen2,3 Petar H. Lambrev1
Petar H. Lambrev1 Győző Garab1,4
Győző Garab1,4- 1Institute of Plant Biology, HUN-REN Biological Research Centre, Szeged, Hungary
- 2State Key Laboratory of Forage Breeding-by-Design and Utilization, Key Laboratory of Photobiology, Institute of Botany, Chinese Academy of Sciences, Beijing, China
- 3Research Institute for Interdisciplinary Science, and Graduate School of Environmental, Life, Natural Science and Technology, Okayama University, Okayama, Japan
- 4Department of Physics, Faculty of Science, University of Ostrava, Ostrava, Czechia
Earlier, we discovered that to reach the maximum chlorophyll-a (Chl-a) fluorescence level of photosystem II (PSII) in diuron-treated samples, and thus PSIIL, the light-adapted charge-separated state, multiple excitations with sufficiently long Δτ waiting times between excitations are required. Experiments of Chl-a fluorescence, elicited by single-turnover saturating flashes (STSFs), revealed that the Δτ1/2 half-waiting times between flashes depended on the lipid content of the reaction centre (RC) and on the temperature. We hypothesized that the gradual formation of PSIIL depends on protonatable residues (and bound water molecules) at the donor side of the RC. Here, we tested this hypothesis: we used diuron-treated isolated PSII core complexes of Thermostichus vulcanus and determined the pH dependence of Δτ1/2 and other Chl-a fluorescence parameters at different temperatures. Data recorded at 5°C revealed moderate and steep increases of Δτ1/2 between pH 5.0 and 6.5, and 6.5 and 8.0, respectively. Much weaker pH dependences were seen at cryogenic temperatures (−20 and −60°C), indicating the role of structural plasticity in the protonation-dependent reorganizations. Similar to the strong pH dependence of Δτ1/2 at 5°C, the magnitude of the F1 fluorescence level, induced by the first STSF, displayed moderate and steep increases between pH 5.0 and 6.5, and 6.5 and 8.0, respectively, at all temperatures. Corroborating data were received from fast Chl-a fluorescence transient measurements. Hence, our data provide experimental support to the hypothesized role of protonation processes in the formation of PSIIL. The protonophores, CCCP and FCCP significantly increased the Δτ1/2 at pH 6.0 and essentially eliminated the pH dependence of the F1 fluorescence level. These data provide experimental support to our hypotheses on the role of protonatable residues in determining the structural dynamics of PSII RCs.
1 Introduction
Photosystem II (PSII) is a large multi-subunit pigment-protein complex embedded in the thylakoid membranes of oxygenic photosynthetic organisms. It plays a fundamental role in light-driven electron transfer, initiating the process of water splitting and oxygen evolution (Barber, 2004; Cardona et al., 2012). PSII catalyses the extraction of electrons from water molecules at the oxygen-evolving complex (OEC) – a process which ultimately supports photosynthetic carbon fixation (Nelson and Yocum, 2006) – using light energy absorbed by its pigment-protein antenna system (Shen, 2015). The catalytic activity of PSII, including charge separation, electron transfer, and water oxidation, relies on finely tuned protonation and deprotonation events that are intricately coupled to its redox chemistry. Altogether, PSII represents the central engine of oxygenic photosynthesis, converting light energy into chemical potential by coupling water oxidation to quinone reduction, thus fuelling life on Earth (Barber, 2004; Shevela et al., 2023).
PSII is organized as a homodimeric supercomplex, where each monomer contains a core complex (CC) composed of several protein subunits and cofactors essential for its function. The reaction centre (RC) is formed by the D1 and D2 proteins, which coordinate all components necessary for the primary charge separation. The integral antenna proteins CP43 and CP47 bind chlorophyll-a (Chl-a) and β-carotene molecules that capture and funnel light energy to the RC. PSII also contains the α and β subunits of cytochrome b559, numerous low-molecular-weight proteins, and the OEC (Umena et al., 2011; Shen, 2015; Suga et al., 2019; Shevela et al., 2023). The RC of PSII contains four Chl-a molecules (a “special pair” forming P680, and two accessory Chls), two pheophytins (Pheo), a non-heme iron, and two plastoquinone molecules (QA and QB), organized along two pseudo-symmetrical branches. Redox-active tyrosine residues (YZ and YD) facilitate electron transfer and connect the RC to the donor side of PSII. The assembly and activity of the OEC are also closely associated with CP43 (Shen, 2015; Suga et al., 2019; Zabret et al., 2021).
The charge separation process is initiated upon illumination of the dark-adapted, open-state PSII (PSIIO), leading to the formation of the radical pair P680+Pheo− (Romero et al., 2017). This is followed by rapid electron transfer from Pheo− to the primary quinone acceptor QA, stabilizing the charge-separated state (P680+QA−). P680+ is then re-reduced by YZ, which in turn oxidizes the Mn4CaO5 cluster of the OEC, advancing the catalytic cycle of water oxidation and generating the S2 state of the OEC. Following these primary reactions, the PSII centres are found in closed state (PSIIC), with all QA reduced. Subsequent electron transfer from QA− to the secondary quinone QB typically occurs over several hundred microseconds under physiological conditions (Shlyk-Kerner et al., 2006). This step can be blocked by herbicides, such as 3-(3′,4′-dichlorophenyl)-1,1′-dimethylurea (DCMU or diuron), which prevent electron flow beyond QA, leading to the stabilization of the charge-separated state. In the presence of DCMU, the lifetime of PSIIC is significantly extended and dominated by charge recombination between QA− and the oxidized OEC (S2) (Tyystjarvi and Vass, 2004).
The redox potential of these cofactors is sensitive to various environmental factors, particularly to the protonation state of surrounding amino acid residues and water molecules. Protonation and deprotonation reactions within PSII are essential for maintaining the proton-coupled electron transfer that drives the catalytic cycle of water oxidation (Siegbahn, 2013; Zaharieva and Dau, 2019). These processes are mediated by complex hydrogen-bonding networks and proton transfer pathways, that link the bulk aqueous phase to the OEC and the quinone sites near and beyond the QB pocket (Saito et al., 2011; Umena et al., 2011; Kaur et al., 2021; Sugo et al., 2022; Li et al., 2024; Yano et al., 2024). These protonation-dependent processes are also reflected in variable Chl-a fluorescence (Crofts and Wraight, 1983; Krieger-Liszkay et al., 2008; Vinyard et al., 2013).
Chl-a fluorescence induction kinetics (ChlF) is one of the most widely used non-invasive techniques to monitor the functioning of PSII and oxygenic photosynthesis across a broad range of biological systems (Govindjee and Papageorgiou, 2004; Papageorgiou and Govindjee, 2011). ChlF induction kinetics provide critical information on electron transfer reactions, PSII activity, plastoquinone redox state, photoprotection, state transitions, responses to environmental stresses, and mechanisms of photoinhibition and repair (Schreiber et al., 1995; Horton and Ruban, 2005; Prásil et al., 2018).
Traditionally, ChlF induction curves – characterized by a rise from the minimal fluorescence level (Fo) to the maximum (Fm) – were interpreted within the widely accepted QA model. According to this model, the variable fluorescence (Fv = Fm – Fo) arises exclusively from the reduction of the primary quinone electron acceptor QA in PSII (Duysens and Sweers, 1963; Strasser et al., 2004; Stirbet and Govindjee, 2012). The multiphasic O–J–I–P fast fluorescence rise is thus ascribed to the progressive closure of PSII reaction centres via QA reduction, while the sigmoidal shape of the O–P rise in the presence of DCMU is interpreted as a manifestation of energetic connectivity between PSII units (Joliot and Joliot, 1964; Lavergne and Trissl, 1995; Stirbet, 2013). However, despite the model’s widespread application, accumulating evidence has highlighted fundamental controversies and limitations of the model (Joliot and Joliot, 1979; Vredenberg, 2011; Schansker et al., 2014; Magyar et al., 2018; Laisk and Oja, 2020). Notably, it has been demonstrated that in DCMU-treated samples, the F1 fluorescence level following the first single-turnover saturating flash (STSF) is significantly lower than Fm, and that Fm cannot be attained solely through QA reduction, induced by the first STSF, but rather requires multiple excitations (Joliot and Joliot, 1979) and sufficiently long Δτ waiting times between the flashes (Magyar et al., 2018). The requirement of Δτ between STSFs indicated the involvement of rate-limiting steps in the dark-to-light transition of PSII.
Our earlier experiments of Chl-a fluorescence transients elicited by trains of STSFs revealed the role of subtle conformational changes and suggested the involvement of dielectric relaxation processes (Sipka et al., 2019; Sipka et al., 2021), as proposed earlier for the formation of the light-adapted charge-separated state of purple bacterial RCs (Malferrari et al., 2013; Sipka et al., 2022). These processes govern the transition of PSII from its closed state (PSIIC) to a light-adapted charge-separated state (PSIIL), which is characterized by enhanced stabilization of charges compared to PSIIC (Sipka et al., 2021). We also showed that the Δτ1/2 half-waiting times and the stepwise fluorescence increments, F1-F2-F3-…-Fm, depended on the lipid composition of the RC matrix and the temperature (Magyar et al., 2022; Magyar et al., 2023; Magyar et al., 2024). However, our understanding concerning the exact physical mechanisms underlying the dark-to-light transition of PSII RC and the molecular environmental factors on which these transitions depend are still rudimentary. In particular, the pH dependence of these transitions has not been studied – although numerous investigations showed that variations in the pH significantly influence the structure and function of PSII. Acidic pH has been shown to accelerate donor-side photoinhibition, impair electron transfer from the OEC, and lead to altered fragmentation patterns of the D1 protein, often accompanied by the formation of hydroxyl radicals (Hideg et al., 1994; Spetea et al., 1997; Krieger-Liszkay et al., 2008). Low pH also facilitates the dissociation of extrinsic proteins (PsbO, PsbP, and PsbQ), leading to inactivation of the OEC (Shen and Inoue, 1991). It has been demonstrated that at an acidic pH of 3.0, the water oxidation activity declines, accompanied by release of Ca2+ and Mn ions essential for OEC stability (Ono and Inoue, 1988). pH modulates S-state transitions, especially those beyond S2, thereby affecting the efficiency of the water-splitting cycle (Bernat et al., 2002; Suzuki et al., 2005). The pH dependence of oxygen evolution in native PSII membranes is bell-shaped, with maximum at around pH of 6.5 (Vass and Styring, 1991; Semin et al., 2015). Further, protonophores such as CCCP (carbonyl cyanide m-chlorophenyl hydrazone) and FCCP (carbonyl cyanide-p-trifluoromethoxyphenylhydrazone), which disrupt transmembrane proton gradients (Heytler and Prichard, 1962; Volkov et al., 1998), act as ADRY reagents, accelerating the deactivation reactions of the OEC (Renger, 1972). Remarkably, the photoreduction of QA has been shown to be largely insensitive to the variations of pH between 6.0 and 8.0 (Demeter et al., 1985; Krieger et al., 1995).
In an earlier work, we hypothesized that the gradual formation of PSIIL is governed by dielectric relaxation processes in the RC matrix, due to the presence of polarizable groups, protonatable residues and bound water molecules, at the donor side of the RC (Sipka et al., 2022). Here we tested this hypothesis by investigating the pH dependence of Δτ1/2 and the F1 level of ChlF, in the range between pH 5.0 and 8.0, on DCMU-treated PSII CC isolated from Thermostichus (T.) vulcanus. Our experiments, performed at different temperatures (5, −20 and −60°C) and in the presence and absence of FCCP or CCCP, provide evidence on the role of protonation/deprotonation processes in the structural dynamics of PSII RCs and in the gradual formation of PSIIL, in particular.
2 Materials and methods
2.1 Source material
A thermophilic cyanobacterial strain, T. vulcanus, originally isolated from a hot spring in Yunomine, Japan (Koike and Inoue, 1983), was used in this study. Cells were cultivated photoautotrophically in batch cultures using BG11 medium (pH 7.0) at 50°C, under continuous white fluorescent light of 50–100 μmol photons m−2 s−1 photon flux density (Shen et al., 2011). Cultures were maintained with constant aeration on a gyratory shaker operating at 100 rpm.
2.2 Sample preparation
PSII CCs of T. vulcanus were isolated following previously described protocols (Shen and Inoue, 1993; Shen and Kamiya, 2000; Shen et al., 2011; Kawakami and Shen, 2018). In the final purification steps, dimers and monomers were separated from crude PSII using an anion exchange column. The column buffer contained 30 mM MES-NaOH (pH 6.0), 3 mM CaCl2, and 0.03% β-DDM, and a linear gradient ranging from 170 to 300 mM NaCl was applied. After collecting the monomeric and dimeric fractions, polyethylene glycol (PEG) 1,450 was added to a final concentration of 13%, followed by centrifugation to precipitate and concentrate the samples. The resulting pellets were resuspended in a buffer containing 30 mM MES–NaOH (pH 6.0), 20 mM NaCl, and 3 mM CaCl2 and stored in liquid nitrogen or at −80°C until further use. For spectroscopic analyses, isolated PSII CCs were diluted in a reaction buffer containing 5% glycerol, 20 mM MES (pH 6.0), 20 mM NaCl and 3 mM CaCl2.
2.3 Chlorophyll-a fluorescence transients
Chl-a fluorescence transients were recorded following a dark adaptation period of 5 min at room temperature, followed by an additional 5 min dark adaptation at the given temperatures, on which the measurements were performed.
For PAM measurements, the Chl concentration of PSII CC was diluted to ∼10 μg mL−1 in the reaction medium, and to ∼20 μg mL−1 in case of the fast fluorescence measurements. DCMU, dissolved in dimethyl sulfoxide (DMSO) was added to all samples immediately before the measurements to a final concentration of 40 μM (with final DMSO concentration kept below 1%). CCCP, also dissolved in DMSO, was added to a final concentration of 5 μM; FCCP, dissolved in ethanol, was used at a final concentration of 5 μM (unless stated otherwise in the text).
2.3.1 PAM fluorescence
Chl-a fluorescence was measured using a Multi-Color PAM (Phase Amplitude Modulation) fluorometer (MC-PAM; Walz, Effeltrich, Germany). Fluorescence transients were induced by STSFs generated by a xenon flash unit (Excelitas LS-1130-3 Flashpac with FX-1163 Flashtube with reflector, Wiesbaden, Germany) with a pulse duration of 1.5 μs duration at half-peak intensity. The modulated measuring light was applied at low intensity, non-actinic level, with a frequency of 1 kHz, which limited the temporal resolution of the fluorescence kinetics to several milliseconds. Samples were placed on the sample holder of a thermoluminescence device for temperature control. Flash timing was precisely managed using a digital pulse generator (525 Six Channel Digital Pulse/Delay Generator, Berkeley Nucleonics, California, United States), and kinetic traces were acquired using the MC-PAM’s dedicated software.
The increments of Chl-a fluorescence from F1-to-F2 were measured by applying a second STSF at varying delay times (Δτ) after the first flash. These fluorescence increments were normalized to the maximum fluorescence level (Fm), determined using multiple-turnover saturating laser flashes. The normalized values (F1,2-Fo)/(Fm-Fo), were plotted as a function of Δτ. The half-rise times (Δτ1/2), representing the Δτ at which 50% of the maximum F1-to-F2 increment is reached, were calculated using a logistic fitting function.
2.3.2 Fast Chl fluorescence measurements
Fast fluorescence transient measurements were performed using a Handy-PEA fluorometer (Hansatech Instruments Ltd., UK), with an excitation photon flux density of 3,380 μmol photons m-2 s-1. The resulting fluorescence induction curves were analysed and fitted using a sum of three exponentials.
2.4 Gel electrophoresis
The integrity of PSII CC dimers at different pHs was checked by blue native polyacrylamide gel electrophoresis on a 5%–13% (w/v) gradient gel at 6°C according to (Schagger et al., 1994). The amount of samples loaded on each lane was 0.4 μg of Chl. The native gel was stained with Coomassie Blue G-250.
3 Results
To elucidate the role of polarizable groups and protonatable residues in the gradual, light-induced formation of PSIIL and the structural dynamic features of the RC matrix in charge-separated state, we performed STSF-induced and fast Chl-a fluorescence measurements on isolated DCMU-treated PSII CCs at various protonation states, via adjusting the pH of the reaction medium between 5.0 and 8.0 and using protonophores. Our major aim was to determine the pH-dependence of the ChlF fluorescence parameter Δτ1/2, which has been shown to be correlated with the structural dynamics of closed PSII RCs (see Introduction). In addition, we also monitored the response of other ChlF parameters to variations in the pH and the temperature.
3.1 pH dependence of Δτ1/2 of the F1-to-F2 transition
To probe the effect of pH, between 5.0 and 8.0, on Δτ1/2, associated with the F1-to-F2 increment of ChlF, we performed double-STSF induced ChlF experiments on DCMU-treated PSII CC of T. vulcanus at different temperatures, 5, −20 and −60°C. Figure 1A shows that, at 5°C, the Δτ1/2 half-waiting time values, marked with dashed vertical lines, exhibited characteristic pH dependence: moderate and strong increases of Δτ1/2 in the pH ranges between 5.0 and 6.5, and between 6.5 and 8.0, respectively. As shown in Figures 1B,C and Table 1, the pH dependence of Δτ1/2 is less pronounced at cryogenic temperatures. These data, on one hand, are in harmony with the proposed role of protonatable residues in the gradual formation of PSIIL, and indicate, on the other hand, that the protonation-dependent reorganizations are hindered at low temperatures.
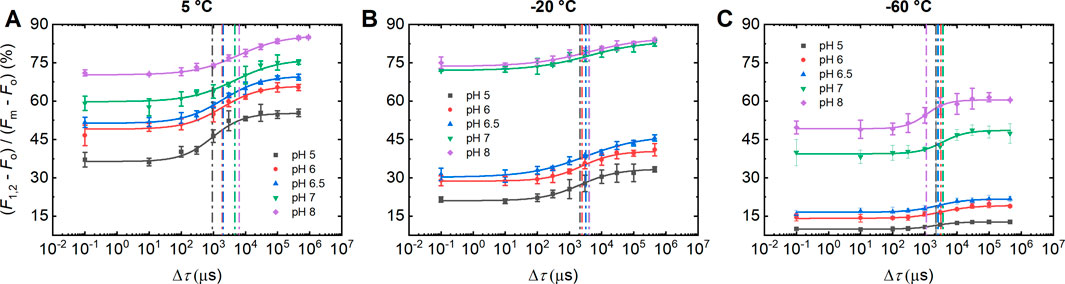
Figure 1. Dependence of the F1-to-F2 Chl-a fluorescence increments of DCMU-treated PSII CC of T. vulcanus on the waiting time (Δτ) between the first and the second STSF at different pH, measured at 5°C (A), −20°C (B) and −60°C (C). Continuous lines represent logistic-function fits of the data points, which represent mean values ±SD (n = 3–7). Dotted vertical lines mark the Δτ1/2 half-waiting time values. (Data on 5 and -20°C, and −60°C were obtained on two different batches.)
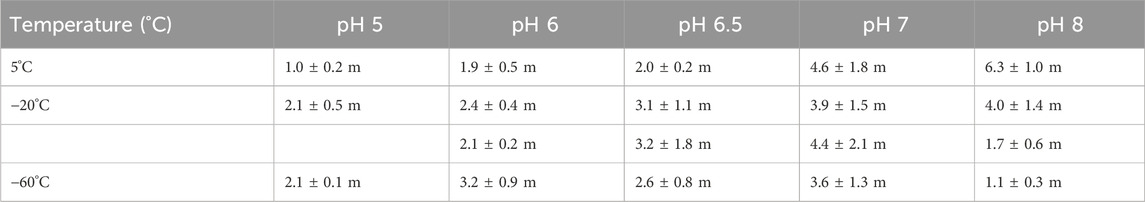
Table 1. Dependences of double-STSF induced F1-to-F2 Δτ1/2 half-rise times of DCMU-treated PSII CC of T. vulcanus at different pHs and temperatures, using the data in Figure 1. The 2 sets of data at −20°C indicate the results of two parallel measurements on two different batches.
Note that Table 1 contains two data sets at −20°C, obtained on two different batches of PSII CC. These data show that different batches display similar trends of their pH dependences in the physiologically relevant pH range, between 6.0 and 7.0, but the actual Δτ1/2 values vary from batch-to-batch (Supplementary Figure S1). Note that Supplementary Table S1 also displays the corresponding P parameter of the logistic fit function and the Fv/Fm values, which testify that the functional activity of PSII, including photochemistry and structural dynamics, is retained in all samples and under all experimental conditions used here. To test the stability of PSII CC at different pHs between pH 5.0 and 8.0, we performed Blue Native PAGE, which revealed no instability of the complexes (Supplementary Figure S2).
It is an interesting, unexpected finding of our experiments with double STSFs that the magnitude of the F1 fluorescence increment exhibited a strong dependence on the pH of the reaction medium (Figure 1). These changes were more pronounced at cryogenic temperatures, which consistently indicated that the largest changes occurred upon increasing the pH from 6.5 to 7.0.
3.2 pH dependence of ChlF amplitudes induced by trains of STSFs at different temperatures
To further investigate the pH-dependence of the F1 fluorescence level of ChlF and to test the behavior of the additional increments, beyond the Fo-to-F1 rise, associated with the PSIIO-to-PSIIC, we conducted ChlF measurements at different temperatures, using trains of STSFs. As can be seen in Figure 2, F1 increases in a stepwise manner with gradually increasing the pH between 5.0 and 8.0. These data show that the transitions from PSIIC-to-PSIIL in all cases proceed through distinct, quantifiable steps and indicate that the protonation state of key residues within PSII modulates the charge stabilization and recombination processes and possibly the structural dynamics of the RCs.
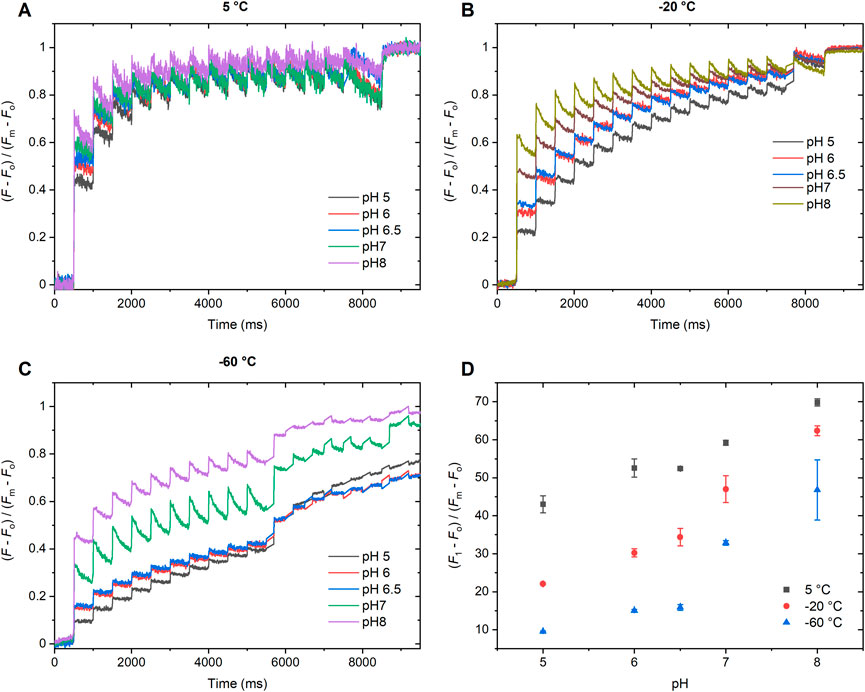
Figure 2. pH-dependent variations of the STSF-induced Chl-a fluorescence transients of DCMU-treated PSII CC of T. vulcanus at 5°C (A), −20°C (B) and −60°C (C). (Each transient represents the average obtained from 3 independent experiments on the same batch; data on 5 and -20, and −60°C were obtained on two different batches). Panel (D) depicts the dependence of the STSF-induced F1 fluorescence level on the pH at different temperatures. (For a more detailed pH dependence at −20°C between 6.5 and 7.0, see Supplementary Figure S4).
As in Figure 1, the variations were more prominent by lowering the temperature. Earlier, we reported similar temperature dependent variations in the Fo-to-F1 rise in DCMU-treated PSII CC of T. vulcanus at pH 6.0 and in spinach thylakoid membranes at pH 7.5 (Magyar et al., 2023). In general, the temperature dependence of the transients recorded on PSII CC at different pHs (Figures 2A–C) followed similar trends as reported earlier for pH 6.0 (Magyar et al., 2023).
It was interesting to observe that, similar to the pH dependence of Δτ1/2 shown above (Figure 1), the largest pH-dependent changes occurred in the narrow range between 6.5 and 7.0. This may indicate the role of a (few) specific residue(s) with pK value(s) in this range. The sharp pH dependence can most prominently be seen at cryogenic temperatures, most strongly at −60°C (Figures 2C,D). It is equally interesting to note that similar, albeit less marked pH dependences are present in the F1-to-F2 increments of ChlF (Supplementary Figure S3). This pH dependence of the increments appeared to persist throughout the entire transient, as it can be inferred from the pH dependences of the number of STSFs required to reach the 50% and 80% of the F1-to-Fm rise at −20°C and 30% at −60°C (Supplementary Figure S3).
3.3 pH dependences as reflected by the fast fluorescence transients
We recorded fast Chl-a fluorescence transients, under comparable experimental conditions as for STSFs (Sections 1 and 2), using the well-established, widely used technique of fast Chl-a fluorescence transients. In the presence and absence of DCMU, these transients, elicited by intense long flashes, are commonly termed as O-P and O-J-I-P transients, respectively (see Introduction).
It will be shown below that fast ChlF traces reflect the basic attributes of the pH and temperature dependence of the PSIIO-PSIIC-PSIIL transitions. However, it must be emphasized that these experiments are not suited for the determination of the two key parameters of ChlF, Δτ1/2, related to the structural dynamics of PSII, and the magnitude of F1 level, associated with the formation of PSIIC. Nevertheless, these are presented because double-STSF-induced PAM fluorescence measurements and adjustable Δτ waiting times between STSFs, to our knowledge, are not used in other laboratories, and no standardized protocol is currently available for commercially available Chl-a fluorimeters.
As shown in Figures 3A–C, fast ChlF transient measurements on DCMU-treated PSII CC revealed – qualitatively - similar pH and temperature dependent changes as shown above with STSF-induced transients. By fitting the fluorescence transients with a three-exponential model and analyzing the amplitudes and rise-times, we observed a clear trend in the first component – which is evidently most closely correlated with the F1 rise (PSIIO-PSIIC). It can be seen that, in harmony with the data in Figures 1, 2, the amplitude of this component increased with rising the pH between 5.0 and 8.0, while the rise-times became shorter (Figures 3D,E). The gradual increase of the amplitude of the first exponential with gradually increasing the pH, agrees well with the data obtained with STSF-induced ChlF data (Figure 2D). In addition, the fast ChlF measurements revealed that the rate of Fo-to-F1 rise, induced by a continuous light (long flash), also depends heavily on the pH. Evidently, the rise kinetics of F1 at different pHs and temperatures could and should be determined using STSF excitation.
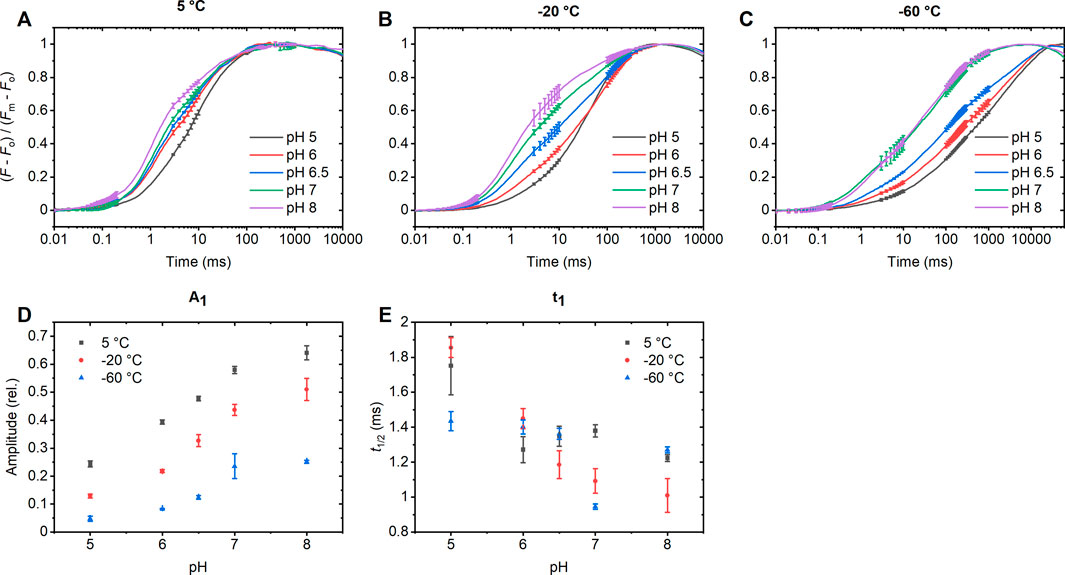
Figure 3. Fast Chl-a fluorescence rise of DCMU-treated PSII CCs of T. vulcanus at 5°C (A), −20°C (B) and −60°C (C), normalized to (Fm - Fo). The curves were fitted with 3 exponentials and the amplitudes (D) and half-rise times (E) of the first component, representing the initial rise, were plotted. In case of the −60°C measurement, the curve was fitted between 50 µs and 700 m. The photon flux density of the excitation was 3,380 mmol photons m–2 s–1. (Since according to the experiments using STSFs, the rises are composed by more than three exponentials, we do not plot the pH dependences of the second and third exponentials.)
3.4 Effect of protonophores
The results of ChlF measurements obtained at different pHs strongly suggest that the structural dynamics PSII RCs depend heavily on polarizability and state of protonation of certain residues of the RC matrix. To substantiate this relationship, we examined the effects of protonophores. To this end, we tested the effects of CCCP and FCCP on the Δτ1/2 parameter and on the Fo-to-F1 rise of ChlF in DCMU-treated PSII CC. These protonophores are known to disrupt proton gradients across membranes by facilitating proton leakage. In the absence of a membrane system, as in our case, they might interact directly with protons within the PSII CC.
When assessing the effect of CCCP and FCCP on the Δτ1/2, we found that they significantly increased the half-waiting time (Figure 4) to values reaching levels comparable to those observed in PSII CC at pH 7.0 and 8.0 at 5°C (cf. Table 1). This suggests that their presence alters the protonation dynamics within PSII, mimicking a higher pH environment. Although CCCP and FCCP are weak acids, they act as proton carriers, binding and releasing protons depending on the local protonation equilibrium (Benz and McLaughlin, 1983). It is shown in Supplementary Figure S5 that FCCP at 2 and 5 µM exerted similar effects, suggesting that the protonophores act at low concentrations.
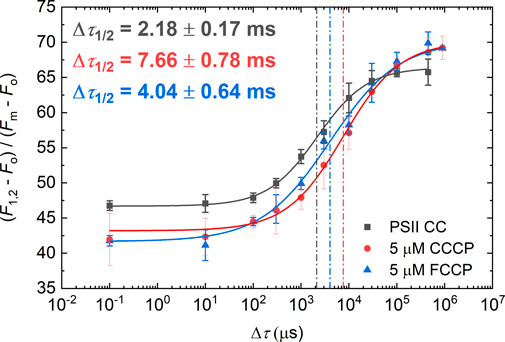
Figure 4. Dependence of the F1-to-F2 Chl-a fluorescence increments of DCMU-treated PSII CC at pH 6.0 on the waiting time (Δτ) between the first and the second STSF in the presence of 5 µM CCCP and FCCP. Continuous lines represent logistic-function fits of the data points, which represent mean values ±SD (n = 3–5). Dotted vertical lines mark the Δτ1/2 half-waiting time values.
As shown in Figure 5, CCCP and FCCP largely eliminated the pH-dependent variations of the F1 level of ChlF at 5°C. (For the effect of FCCP on the kinetic transients, see Supplementary Figure S6). These results corroborate our conclusion on the role of light-induced proton motions inside PSII CC by these uncouplers. (Note that all these effects are observed in the presence of DCMU, which permits only a single stable light-induced charge separation.) In contrast to their effects at 5°C, at −20°C (Supplementary Figure S7), CCCP and FCCP had no discernible effect on the F1 level, suggesting that their activity, based on their ability to shuttle protons between different lipophilic sites (Kasianowicz et al., 1984; Lim et al., 2001; Pansook et al., 2019), is halted at cryogenic temperatures. This is evidently a consequence of the increased rigidity and lack of fluidity in the surrounding environment, which hinders the mobility and proton-shuttling activity of the protonophores (Hazel, 1995; Monteiro et al., 2011).
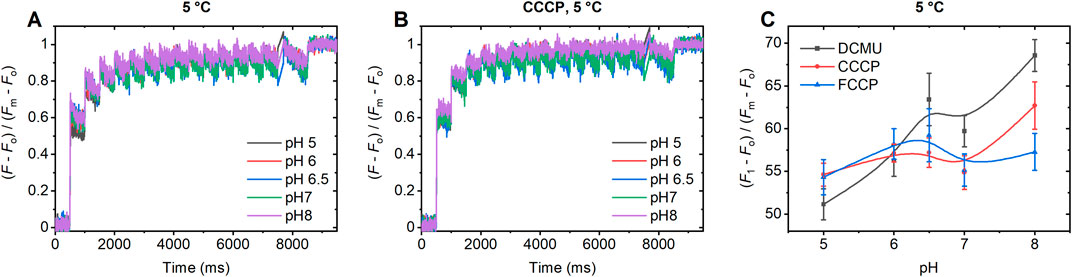
Figure 5. pH-dependent variations of the STSF-induced Chl-a fluorescence transients at 5°C of DCMU-treated PSII CC of T. vulcanus in the absence (A) and presence of CCCP (B) and dependence of the (F1-Fo)/(Fm-Fo) parameter on the pH in the absence and presence of CCCP and FCCP (C).
It is interesting to remark that no significant changes in the fluorescence transients were observed upon treatment of the samples with NH4Cl or nigericin (either in the presence or absence of K+ ions), when tested at pH 6 (Supplementary Figure S8). Despite their known effects on proton transport and pH modulation, neither compound substantially affected the multi-STSF fluorescence transients in PSII CC. This might be explained by their limited accessibility to specific sites involved in the protonation/deprotonation processes.
4 Discussion
The major aim of this work was to test the hypothesis set in our previous work, on the role of polarizable groups at the donor side of PSII in the gradual formation of PSIIL, the light-induced charge-separated state of the RC of PSII, and in the structural dynamics of the RC matrix, in general (Sipka et al., 2022). The processes involved in the formation of PSIIL are of substantial interest because of the enhanced stabilization of charges in PSIIL compared to PSIIC (Sipka et al., 2021). Our hypothesis on the mechanism of formation of PSIIL was largely inspired by the experimental findings of Allen et al. (2023), who used wild-type and mutant purple bacterial RCs (bRCs) to demonstrate a strong correlation between the stabilization of charges in the light adapted state of the bRC and polarizable amino acid residues near its primary special pair (P) molecules. Data presented by these authors pointed to the involvement of a hydrogen-bond network and bound water molecules, forming a proton transfer pathway around P.
Our investigations provide experimental evidence for the role of protonation/deprotonation processes in the gradual formation of PSIIL. In particular, we have shown that the Δτ1/2 half-waiting time, a key parameter characteristic of the stepwise light-induced PSIIC-to-PSIIL transition, depends strongly on the pH of the reaction medium. Here, Δτ1/2 was determined for the first step of this transition, manifested in the F1-to-F2 Chl-a fluorescence level increment (Figure 1). (Earlier we have shown that Δτ1/2 is involved in all later steps tested during the Fv induction period (Magyar et al., 2023).) It was interesting to observe that upon increasing the pH from 6.5 to 7.0 and 8.0, the value of Δτ1/2 at 5 °C dramatically increased (Table 1). These data indicate that deprotonation of some residues, with pK between 6.5 and 7.0, strongly hampers the subtle structural reorganization occurring in the RC matrix. This conclusion was substantiated by the effect of the protonophores CCCP and FCCP, which led to increased Δτ1/2 values, in a similar manner as elevating the pH from 6.5 to 7.0 (Figures 4, 5). This effect of protonophores is most likely related to their ability to accelerate the deactivation reactions of the OEC, similar to other ADRY reagents (Renger, 1972). They act as mobile proton carriers and can interact with PSII by altering the local availability of protons, disrupting structured water networks, and affecting the protonation state of residues involved in proton-coupled electron transfer (Shioi et al., 1976; Nemson and Melis, 1990; Samuilov and Barsky, 1993). By disrupting structured water networks and protonation states of key residues, protonophores provide insights into the electrostatic environment of PSII. It is well known that CCCP modifies the donor side reactions of PSII (Homann, 1971; Renger, 1972). Although these protonophores are acidic, their function is not dictated by a static proton load; rather, they dynamically bind and release protons depending on the surrounding pH gradient. However, the exact site of the effect of these protonophores and the identity of the residues, with pK values between 6.5 and 7.0, remain to be clarified.
In addition to the marked pH dependence of Δτ1/2, we also found, unexpectedly, that the F1 level of the Chl-a fluorescence increased dramatically upon increasing the pH between 5.0 and 8.0; again, the increase was most prominent in the narrow pH range between 6.5 and 7.0 (Figures 1, 2). It is noteworthy that the overall pattern of ChlF transients remained the same at all pHs and temperatures. All transients displayed F1<Fm and to reach Fm, multiple STSFs (or multi-turnover flashes) were required. Furthermore, all samples under all conditions possessed high Fv/Fm values (about equal or larger than 0.8 – see Supplementary Table S1). This suggests that the pH and temperature dependences of the Fo-to-F1 rise and the Δτ1/2 half-waiting time might be governed by similar physical mechanism(s), both containing protonation-dependent variations in the structural dynamics of PSIIC. The enhanced Fo-to-F1 rises at high pHs might, in large part, reflect increased levels of the structural plasticity of the RCs; at lower pHs, similar increments can only be induced by additional STSFs (Figure 2, see also Supplementary Figure S4). In this context, it is important to recall that Oja and Laisk (2020) reported a ‘prompt’ rise to approximately 1.8×Fo; this further increased to ∼3×Fo within the sub-ms time range, an increment attributed to protein conformational change. Indeed, at low pH and low temperatures, we detected fluorescence levels reaching only ∼1.8×Fo or lower – with ms time resolution (Figures 1, 2, see also Magyar et al. (2018); Sipka et al. (2021)). As concluded from measurements at cryogenic temperatures, the rise associated with closing the RC, i.e., the reduction of QA was considerably smaller than 1.8×Fo (Schansker et al., 2011; Sipka et al., 2021). In our experiments, at pH 6.0 and −60°C, we reach a quasi-steady state level of 1.13×Fo (corresponding to 15% Fv rise – see Figure 1C). It is important to recall that the photoreduction of QA between pH 6.0 and 8.0 is essentially pH independent (Demeter et al., 1985; Krieger et al., 1995), and occurs at cryogenic temperatures (Kirmaier et al., 1985; Yakovlev and Shuvalov, 2016). These data will aid us to elucidate the physical mechanisms associated with Fv, in general.
The origin of the pH and temperature dependent variations of the F1 level, in particular, remains to be clarified. To this end, it would be important to perform sub-ms time resolution experiments at different temperatures and pHs on DCMU-treated PSII CC. Using PAM fluorimetry, sub-ms (and even sub-µs) fluorescence components have recently been recorded in Chlorella cells (Klughammer et al., 2024). Sub-ms time resolution PAM experiments on DCMU-treated PSII CC, under comparable conditions as applied here, are in preparation in our laboratory. However, they are outside the scope of the present paper, which focuses on the role of protonation/deprotonation processes in the gradual formation of PSIIL. In the present study, sub-ms time resolution measurements are confined to the so-called fast Chl-a, O-P type of, fluorescence transients, using intense long pulses. These data revealed elevated levels and rates of the first exponential rise component of the transient (Figure 3). Hence, they support the notion that the fast rise of ChlF upon the first STSF, i.e., the quasi-stationary F1 level, depends to a large extent on the structural plasticity of PSIIC. The nature of the (most probably subtle) reorganizations at the donor side of PSII RC, marked by the fast transients in ChlF, and the role of the protonation/deprotonation processes, in particular, remain to be clarified.
The enhanced stabilization of charges during PSIIO-to-PSIIL has been proposed to follow a mechanism similar to that in bRCs, with dielectric relaxation processes optimizing the RC matrix’s dielectric properties. Dielectric relaxation processes have earlier been proposed to play important roles in the formation of light adapted states of bRC and PSII (Malferrari et al., 2013; Sipka et al., 2021). Here we established the involvement of protonatable/deprotonatable groups in modulating Fv increments, which are thus thought to be linked to conformational changes and dielectric relaxation. In broad terms, the S-state dependent proton release and fluorescence oscillation patterns, reported by Delosme and Joliot (2002) can be reconciled with our observations–with the assumption that the protonation/deprotonation processes are not confined to the OEC but extend to the donor side of the RC. In general, protonation and deprotonation reactions within PSII are critical for proton-coupled electron transfer (PCET), involving amino acid residues such as histidines and carboxylates that form hydrogen-bonded networks (Saito et al., 2011; Vogt et al., 2015). In particular, residues located near YZ (D1-Tyr161) and the non-heme iron–bicarbonate site modulate the protonation state of redox intermediates like P680+, QA−, and YZ•, thereby influencing PSII activity and recombination kinetics (Ferreira et al., 2004). Redox-active tyrosines, YZ and YD, also exhibit distinct pH-dependent reaction rates (Moser et al., 1995; Reece et al., 2006), further influencing PCET efficiency. However, it is important to note that in our case PCET does not apply, due to the single turnover stable charge separation and the formation of the closed state of the PSII RC upon the first STSF.
Although the pH dependent variations in Fv parameters can be explained with conformational changes that are influenced by protonation states of key residues, alternative mechanisms may contribute to the observed changes. For instance, shifts in charge recombination kinetics and/or an increased fraction of inefficient centers may affect the kinetic traces (Lavergne and Rappaport, 1998). Indeed, some pH-dependent recombination-associated decays are observed at high pH after the first flash (Figure 2); however, the observed magnitudes and rates offer no explanation for the basic features of the variations either in Δτ1/2 or in the F1 level. Instead, the decays observed after multi-STSF excitation likely stem from the inherent plasticity of PSII rather than S2QA− recombination (Magyar et al., 2018). It is also to be noted that the half-time for S2QA− recombination is 1–2 s, with pH shifts affecting the equilibrium between S2P680 and S1P680+ (Vass and Styring, 1991) – but they remain within the range of our time resolution. Also, the required waiting times and the high Fv/Fm values essentially rule out the presence of inactive centers in sizeable amounts; this holds true also for photoinhibition, which is known to be enhanced under low pH due to destabilization of the OEC (Debus, 1992).
Together, these findings underscore the complex interplay between polarizable groups and protonatable residues, redox processes, and subtle structural reorganizations in PSII. Understanding how these factors are modulated under varying pH conditions and in the presence of protonophores is essential for elucidating the mechanisms underlying water oxidation, photoprotection, and photodamage. These results also raise important questions about the more precise mechanistic origin of Fv, in molecular terms, the role of electric field formation, conformational adjustments, and protein-lipid interactions in PSII function. Addressing these questions is crucial not only for understanding fundamental photosynthetic mechanisms but also for improving the interpretation of Chl fluorescence data across all scales – from molecular studies to ecological monitoring.
Data availability statement
The original contributions presented in the study are included in the article/Supplementary Materal, further inquiries can be directed to the corresponding author.
Author contributions
MM: Writing – original draft, Formal Analysis, Visualization, Methodology, Project administration, Investigation, Data curation, Conceptualization, Validation. GS: Writing – review and editing, Conceptualization, Methodology, Software. ID: Formal Analysis, Visualization, Writing – review and editing. XC: Resources, Writing – review and editing. XW: Resources, Writing – review and editing. GH: Writing – review and editing, Resources, Funding acquisition. J-RS: Funding acquisition, Resources, Writing – review and editing. PL: Funding acquisition, Project administration, Writing – review and editing, Supervision. GG: Supervision, Conceptualization, Writing – original draft, Project administration, Funding acquisition.
Funding
The author(s) declare that financial support was received for the research and/or publication of this article. The authors acknowledge the support from the Hungarian Ministry of Innovation and Technology, National Research, Development and Innovation Fund (ANN-144012 to PHL). GG acknowledges the support from the Czech Science Foundation (GAČR 23-07744S) and the European Union under the LERCO project (CZ.10.03.01/00/22_003/0000003). Studies in the Chinese group are supported from a National Key R&D Program of China (2022YFA0911900, 2022YFC3401800), CAS Project for Young Scientists in Basic Research (YSBR-004), and a National Natural Science Foundation of China (92461307).
Conflict of interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
The author(s) declared that they were an editorial board member of Frontiers, at the time of submission. This had no impact on the peer review process and the final decision.
Generative AI statement
The author(s) declare that no Generative AI was used in the creation of this manuscript.
Publisher’s note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
Supplementary material
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fphbi.2025.1623224/full#supplementary-material
References
Allen, J. P., Chamberlain, K. D., and Williams, J. C. (2023). Identification of amino acid residues in a proton release pathway near the bacteriochlorophyll dimer in reaction centers from Rhodobacter sphaeroides. Photosynth Res. 155, 23–34. doi:10.1007/s11120-022-00968-x
Barber, J. (2004). Engine of life and big bang of evolution: a personal perspective. Photosynth Res. 80, 137–155. doi:10.1023/B:PRES.0000030662.04618.27
Benz, R., and Mclaughlin, S. (1983). The molecular mechanism of action of the proton ionophore FCCP (carbonylcyanide p-trifluoromethoxyphenylhydrazone). Biophys. J. 41, 381–398. doi:10.1016/S0006-3495(83)84449-X
Bernat, G., Morvaridi, F., Feyziyev, Y., and Styring, S. (2002). pH dependence of the four individual transitions in the catalytic S-cycle during photosynthetic oxygen evolution. Biochemistry 41, 5830–5843. doi:10.1021/bi011691u
Cardona, T., Sedoud, A., Cox, N., and Rutherford, A. W. (2012). Charge separation in photosystem II: a comparative and evolutionary overview. Biochim. Biophys. Acta 1817, 26–43. doi:10.1016/j.bbabio.2011.07.012
Crofts, A. R., and Wraight, C. A. (1983). The electrochemical domain of photosynthesis. Biochimica Biophysica Acta 726, 149–185. doi:10.1016/0304-4173(83)90004-6
Debus, R. J. (1992). The manganese and calcium ions of photosynthetic oxygen evolution. Biochim. Biophys. Acta 1102, 269–352. doi:10.1016/0005-2728(92)90133-m
Delosme, R., and Joliot, P. (2002). Period four oscillations in chlorophyll a fluorescence. Photosynth Res. 73, 165–168. doi:10.1023/A:1020430610627
Demeter, S., Rozsa, Z., Vass, I., and Sallai, A. (1985). Thermoluminescence study of charge recombination in Photosystem II at low temperatures. I. Characterization of the Zv and A thermoluminescence bands. Biochimica Biophysica Acta 809, 369–378. doi:10.1016/0005-2728(85)90187-2
Duysens, L., and Sweers, H. (1963). Mechanism of two photochemical reactions in algae as studied by means of fluorescence. University of Tokyo Press.
Ferreira, K. N., Iverson, T. M., Maghlaoui, K., Barber, J., and Iwata, S. (2004). Architecture of the photosynthetic oxygen-evolving center. Science 303, 1831–1838. doi:10.1126/science.1093087
Govindjee, G., and Papageorgiou, G. (2004). Chlorophyll A fluorescence: a signature of photosynthesis. Dordrecht: Springer.
Hazel, J. R. (1995). Thermal adaptation in biological membranes: is homeoviscous adaptation the explanation? Annu. Rev. Physiol. 57, 19–42. doi:10.1146/annurev.ph.57.030195.000315
Heytler, P. G., and Prichard, W. W. (1962). A new class of uncoupling agents--carbonyl cyanide phenylhydrazones. Biochem. Biophys. Res. Commun. 7, 272–275. doi:10.1016/0006-291x(62)90189-4
Hideg, E., Spetea, C., and Vass, I. (1994). Singlet oxygen and free radical production during acceptor- and donor-side-induced photoinhibition. Biochimica Biophysica Acta-Bioenergetics 1186, 143–152. doi:10.1016/0005-2728(94)90173-2
Homann, P. H. (1971). Actions of carbonylcyanide m-chlorophenylhydrazone on electron transport and fluorescence of isolated chloroplasts. Biochimica Biophysica Acta (BBA) - Bioenergetics 245, 129–143. doi:10.1016/0005-2728(71)90015-6
Horton, P., and Ruban, A. (2005). Molecular design of the photosystem II light-harvesting antenna: photosynthesis and photoprotection. J. Exp. Bot. 56, 365–373. doi:10.1093/jxb/eri023
Joliot, A., and Joliot, P. (1964). Kinetic study of the potochemical reaction liberating oxygen during photosynthesis. C R. Hebd. Seances Acad. Sci. 258, 4622–4625.
Joliot, P., and Joliot, A. (1979). Comparative-study of the fluorescence yield and of the C550 absorption change at room-temperature. Biochimica Biophysica Acta 546, 93–105. doi:10.1016/0005-2728(79)90173-7
Kasianowicz, J., Benz, R., and Mclaughlin, S. (1984). The kinetic mechanism by which CCCP (carbonyl cyanide m-chlorophenylhydrazone) transports protons across membranes. J. Membr. Biol. 82, 179–190. doi:10.1007/BF01868942
Kaur, D., Khaniya, U., Zhang, Y. Y., and Gunner, M. R. (2021). Protein motifs for proton transfers that build the transmembrane proton gradient. Front. Chem. 9, 660954. doi:10.3389/fchem.2021.660954
Kawakami, K., and Shen, J. R. (2018). Purification of fully active and crystallizable photosystem II from thermophilic cyanobacteria. Methods Enzymol. 613, 1–16. doi:10.1016/bs.mie.2018.10.002
Kirmaier, C., Holten, D., and Parson, W. W. (1985). Temperature and detection-wavelength dependence of the picosecond electron-transfer kinetics measured in rhodopseudomonas-sphaeroides reaction centers - resolution of new spectral and kinetic components in the primary charge-separation process. Biochimica Biophysica Acta 810, 33–48. doi:10.1016/0005-2728(85)90204-x
Klughammer, C., Schlosser, F., and Schreiber, U. (2024). Flash-kinetics as a complementary analytical tool in PAM fluorimetry. Photosynth. Res. 161, 151–176. doi:10.1007/s11120-024-01101-w
Koike, H., and Inoue, Y. (1983). Preparation of oxygen-evolving photosystem II particles from a thermophilic blue-green alga. Oxyg. Evol. Syst. Photosynth. 257–263. doi:10.1016/B978-0-12-372360-4.50034-1
Krieger, A., Rutherford, A. W., and Johnson, G. N. (1995). On the determination of redox midpoint potential of the primary quinone electron-acceptor, Q(a), in photosystem-ii. Biochimica Biophysica Acta-Bioenergetics 1229, 193–201. doi:10.1016/0005-2728(95)00002-z
Krieger-Liszkay, A., Fufezan, C., and Trebst, A. (2008). Singlet oxygen production in photosystem II and related protection mechanism. Photosynth. Res. 98, 551–564. doi:10.1007/s11120-008-9349-3
Laisk, A., and Oja, V. (2020). Variable fluorescence of closed photochemical reaction centers. Photosynth. Res. 143, 335–346. doi:10.1007/s11120-020-00712-3
Lavergne, J., and Rappaport, F. (1998). Stabilization of charge separation and photochemical misses in photosystem II. Biochemistry 37, 7899–7906. doi:10.1021/bi9801210
Lavergne, J., and Trissl, H. W. (1995). Theory of fluorescence induction in photosystem II: derivation of analytical expressions in a model including exciton-radical-pair equilibrium and restricted energy transfer between photosynthetic units. Biophys. J. 68, 2474–2492. doi:10.1016/S0006-3495(95)80429-7
Li, H. J., Nakajima, Y., Nango, E., Owada, S., Yamada, D., Hashimoto, K., et al. (2024). Oxygen-evolving photosystem II structures during S1-S2-S3 transitions. Nature 626, 670–677. doi:10.1038/s41586-023-06987-5
Lim, M. L., Minamikawa, T., and Nagley, P. (2001). The protonophore CCCP induces mitochondrial permeability transition without cytochrome c release in human osteosarcoma cells. FEBS Lett. 503, 69–74. doi:10.1016/s0014-5793(01)02693-x
Magyar, M., Akhtar, P., Sipka, G., Domonkos, I., Han, W., Li, X., et al. (2024). Effects of lipids on the rate-limiting steps in the dark-to-light transition of Photosystem II core complex of Thermostichus vulcanus. Front. Plant Sci. 15, 1381040. doi:10.3389/fpls.2024.1381040
Magyar, M., Akhtar, P., Sipka, G., Han, W., Li, X., Han, G., et al. (2022). Dependence of the rate-limiting steps in the dark-to-light transition of photosystem II on the lipidic environment of the reaction center. Photosynthetica 60, 147–156. doi:10.32615/ps.2022.016
Magyar, M., Sipka, G., Han, W., Li, X., Han, G., Shen, J. R., et al. (2023). Characterization of the rate-limiting steps in the dark-to-light transitions of closed photosystem II: temperature dependence and invariance of waiting times during multiple light reactions. Int. J. Mol. Sci. 24, 94. doi:10.3390/ijms24010094
Magyar, M., Sipka, G., Kovács, L., Ughy, B., Zhu, Q. J., Han, G. Y., et al. (2018). Rate-limiting steps in the dark-to-light transition of Photosystem II - revealed by chlorophyll-a fluorescence induction. Sci. Rep. 8, 2755. doi:10.1038/s41598-018-21195-2
Malferrari, M., Mezzetti, A., Francia, F., and Venturoli, G. (2013). Effects of dehydration on light-induced conformational changes in bacterial photosynthetic reaction centers probed by optical and differential FTIR spectroscopy. Biochim. Biophys. Acta 1827, 328–339. doi:10.1016/j.bbabio.2012.10.009
Monteiro, J. P., Martins, A. F., Lucio, M., Reis, S., Geraldes, C. F., Oliveira, P. J., et al. (2011). Interaction of carbonylcyanide p-trifluoromethoxyphenylhydrazone (FCCP) with lipid membrane systems: a biophysical approach with relevance to mitochondrial uncoupling. J. Bioenerg. Biomembr. 43, 287–298. doi:10.1007/s10863-011-9359-2
Moser, C. C., Page, C. C., Farid, R., and Dutton, P. L. (1995). Biological electron-transfer. J. Bioenergetics Biomembr. 27, 263–274. doi:10.1007/BF02110096
Nelson, N., and Yocum, C. F. (2006). Structure and function of photosystems I and II. Annu. Rev. Plant Biol. 57, 521–565. doi:10.1146/annurev.arplant.57.032905.105350
Nemson, J. A., and Melis, A. (1990). Light-induced oxidation-reduction reactions of photosystem-ii in dichlorophenyl-dimethyl urea (dcmu) inhibited thylakoids. Zeitschrift Fur Naturforschung Sect. C-a J. Biosci. 45, 258–264. doi:10.1515/znc-1990-3-420
Oja, V., and Laisk, A. (2020). Time- and reduction-dependent rise of photosystem II fluorescence during microseconds-long inductions in leaves. Photosynth Res. 145, 209–225. doi:10.1007/s11120-020-00783-2
Ono, T., and Inoue, Y. (1988). Discrete extraction of the Ca atom functional for O-2 evolution in higher-plant photosystem-ii by a simple low Ph treatment. Febs Lett. 227, 147–152. doi:10.1016/0014-5793(88)80886-x
Pansook, S., Incharoensakdi, A., and Phunpruch, S. (2019). Effects of the photosystem II inhibitors CCCP and DCMU on hydrogen production by the unicellular halotolerant cyanobacterium aphanothece halophytica. ScientificWorldJournal 2019, 1030236. doi:10.1155/2019/1030236
Papageorgiou, G. C., and Govindjee, (2011). Photosystem II fluorescence: slow changes - scaling from the past. J. Photochem. Photobiol. B-Biology 104, 258–270. doi:10.1016/j.jphotobiol.2011.03.008
Prásil, O., Kolber, Z. S., and Falkowski, P. G. (2018). Control of the maximal chlorophyll fluorescence yield by the QB binding site. Photosynthetica 56, 150–162. doi:10.1007/s11099-018-0768-x
Reece, S. Y., Hodgkiss, J. M., Stubbe, J., and Nocera, D. G. (2006). Proton-coupled electron transfer: the mechanistic underpinning for radical transport and catalysis in biology. Philos. Trans. R. Soc. Lond. B. 361, 1351–1364. doi:10.1098/rstb.2006.1874
Renger, G. (1972). The action of 2-anilinothiophenes as accelerators of the deactivation reactions in the watersplitting enzyme system of photosynthesis. Biochim. Biophys. Acta 256, 428–439. doi:10.1016/0005-2728(72)90072-2
Romero, E., Novoderezhkin, V. I., and Van Grondelle, R. (2017). Quantum design of photosynthesis for bio-inspired solar-energy conversion. Nature 543, 355–365. doi:10.1038/nature22012
Saito, K., Shen, J. R., Ishida, T., and Ishikita, H. (2011). Short hydrogen bond between redox-active tyrosine Y(Z) and D1-His190 in the photosystem II crystal structure. Biochemistry 50, 9836–9844. doi:10.1021/bi201366j
Samuilov, V. D., and Barsky, E. L. (1993). Interaction of carbonyl cyanide M-chlorophenylhydrazone with the photosystem-ii acceptor side. Febs Lett. 320, 118–120. doi:10.1016/0014-5793(93)80074-5
Schagger, H., Cramer, W. A., and Von Jagow, G. (1994). Analysis of molecular masses and oligomeric states of protein complexes by blue native electrophoresis and isolation of membrane protein complexes by two-dimensional native electrophoresis. Anal. Biochem. 217, 220–230. doi:10.1006/abio.1994.1112
Schansker, G., Tóth, S. Z., Holzwarth, A. R., and Garab, G. (2014). Chlorophyll a fluorescence: beyond the limits of the Q(A) model. Photosynth. Res. 120, 43–58. doi:10.1007/s11120-013-9806-5
Schansker, G., Toth, S. Z., Kovacs, L., Holzwarth, A. R., and Garab, G. (2011). Evidence for a fluorescence yield change driven by a light-induced conformational change within photosystem II during the fast chlorophyll a fluorescence rise. Biochim. Biophys. Acta 1807, 1032–1043. doi:10.1016/j.bbabio.2011.05.022
Schreiber, U., Bilger, W., and Neubauer, C. (1995). “Chlorophyll fluorescence as a nonintrusive indicator for rapid assessment of in vivo photosynthesis,” in Ecophysiology of photosynthesis. Editors E.-D. SCHULZE, and M. M. CALDWELL (Berlin, Heidelberg: Springer Berlin Heidelberg).
Semin, B. K., Davletshina, L. N., and Rubin, A. B. (2015). Correlation between pH dependence of O2 evolution and sensitivity of Mn cations in the oxygen-evolving complex to exogenous reductants. Photosynth. Res. 125, 95–103. doi:10.1007/s11120-015-0155-4
Shen, J. R. (2015). The structure of photosystem II and the mechanism of water oxidation in photosynthesis. Annu. Rev. Plant Biol. 66, 23–48. doi:10.1146/annurev-arplant-050312-120129
Shen, J. R., and Inoue, Y. (1991). Low pH-induced dissociation of three extrinsic proteins from O2-evolving photosystem II. Plant Cell Physiology 32, 453–457. doi:10.1093/oxfordjournals.pcp.a078101
Shen, J. R., and Inoue, Y. (1993). Binding and functional properties of two new extrinsic components, cytochrome c-550 and a 12-kDa protein, in cyanobacterial photosystem II. Biochemistry 32, 1825–1832. doi:10.1021/bi00058a017
Shen, J. R., and Kamiya, N. (2000). Crystallization and the crystal properties of the oxygen-evolving photosystem II from Synechococcus vulcanus. Biochemistry 39, 14739–14744. doi:10.1021/bi001402m
Shen, J.-R., Kawakami, K., and Koike, H. (2011). “Purification and crystallization of oxygen-evolving photosystem II core complex from thermophilic cyanobacteria,” in Photosynthesis research protocols. Editor R. CARPENTIER (Totowa, NJ: Humana Press).
Shevela, D., Kern, J. F., Govindjee, G., and Messinger, J. (2023). Solar energy conversion by photosystem II: principles and structures. Photosynth Res. 156, 279–307. doi:10.1007/s11120-022-00991-y
Shioi, Y., Takamiya, K., and Nishimura, M. (1976). Light-induced oxidation-reduction reactions of cytochromes in the green sulfur photosynthetic bacterium Prosthecochloris aesturarii. J. Biochem. 80, 811–820. doi:10.1093/oxfordjournals.jbchem.a131342
Shlyk-Kerner, O., Samish, I., Kaftan, D., Holland, N., Sai, P. S., Kless, H., et al. (2006). Protein flexibility acclimatizes photosynthetic energy conversion to the ambient temperature. Nature 442, 827–830. doi:10.1038/nature04947
Siegbahn, P. E. (2013). Water oxidation mechanism in photosystem II, including oxidations, proton release pathways, O-O bond formation and O2 release. Biochim. Biophys. Acta 1827, 1003–1019. doi:10.1016/j.bbabio.2012.10.006
Sipka, G. B., Magyar, M., Mezzetti, A., Akhtar, P., Zhu, Q., Xiao, Y., et al. (2021). Light-adapted charge-separated state of photosystem II: structural and functional dynamics of the closed reaction center. Plant Cell 33, 1286–1302. doi:10.1093/plcell/koab008
Sipka, G., Muller, P., Brettel, K., Magyar, M., Kovacs, L., Zhu, Q., et al. (2019). Redox transients of P680 associated with the incremental chlorophyll-a fluorescence yield rises elicited by a series of saturating flashes in diuron-treated photosystem II core complex of Thermosynechococcus vulcanus. Physiol. Plant 166, 22–32. doi:10.1111/ppl.12945
Sipka, G., Nagy, L., Magyar, M., Akhtar, P., Shen, J. R., Holzwarth, A. R., et al. (2022). Light-induced reversible reorganizations in closed Type II reaction centre complexes: physiological roles and physical mechanisms. Open Biol. 12, 220297. doi:10.1098/rsob.220297
Spetea, C., Hideg, E., and Vass, I. (1997). Low pH accelerates light-induced damage of photosystem II by enhancing the probability of the donor-side mechanism of photoinhibition. Biochimica Biophysica Acta-Bioenergetics 1318, 275–283. doi:10.1016/s0005-2728(96)00145-4
Stirbet, A., and GOVINDJEE, (2012). Chlorophyll a fluorescence induction: a personal perspective of the thermal phase, the J-I-P rise. Photosynth Res. 113, 15–61. doi:10.1007/s11120-012-9754-5
Stirbet, A. (2013). Excitonic connectivity between photosystem II units: what is it, and how to measure it? Photosynth. Res. 116, 189–214. doi:10.1007/s11120-013-9863-9
Strasser, R. J., Tsimilli-Michael, M., and Srivastava, A. (2004). Analysis of the chlorophyll a fluorescence transient, in Chlorophyll a fluorescence: a signature of photosynthesis. Editor G. C. PAPAGEORGIOU (Dordrecht: Springer).
Suga, M., Akita, F., Yamashita, K., Nakajima, Y., Ueno, G., Li, H., et al. (2019). An oxyl/oxo mechanism for oxygen-oxygen coupling in PSII revealed by an x-ray free-electron laser. Science 366, 334–338. doi:10.1126/science.aax6998
Sugo, Y., Saito, K., and Ishikita, H. (2022). Conformational changes and H-bond rearrangements during quinone release in photosystem II. Biochemistry 61, 1836–1843. doi:10.1021/acs.biochem.2c00324
Suzuki, H., Sugiura, M., and Noguchi, T. (2005). pH dependence of the flash-induced S-state transitions in the oxygen-evolving center of photosystem II from Thermosynechoccocus elongatus as revealed by Fourier transform infrared spectroscopy. Biochemistry 44, 1708–1718. doi:10.1021/bi0483312
Tyystjarvi, E., and Vass, I. (2004). “Light emission as a probe of charge separation and recombination in the photosynthetic apparatus: relation of prompt fluorescence to delayed light emission and thermoluminescence,” in Chlorophyll a fluorescence: a signature of photosynthesis. Editor G. C. PAPAGEORGIOU (Dordrecht: Springer).
Umena, Y., Kawakami, K., Shen, J. R., and Kamiya, N. (2011). Crystal structure of oxygen-evolving photosystem II at a resolution of 1.9 Å. Nature 473, 55–60. doi:10.1038/nature09913
Vass, I., and Styring, S. (1991). pH-dependent charge equilibria between tyrosine-D and the S states in photosystem II. Estimation of relative midpoint redox potentials. Biochemistry 30, 830–839. doi:10.1021/bi00217a037
Vinyard, D. J., Ananyev, G. M., and Dismukes, G. C. (2013). Photosystem II: the reaction center of oxygenic photosynthesis. Annu. Rev. Biochem. 82 (82), 577–606. doi:10.1146/annurev-biochem-070511-100425
Vogt, L., Vinyard, D. J., Khan, S., and Brudvig, G. W. (2015). Oxygen-evolving complex of Photosystem II: an analysis of second-shell residues and hydrogen-bonding networks. Curr. Opin. Chem. Biol. 25, 152–158. doi:10.1016/j.cbpa.2014.12.040
Volkov, A. G., Deamer, D. W., Tanelian, D. L., and Markin, V. S. (1998). Liquid interfaces in chemistry and biology. New York: Wiley.
Vredenberg, W. (2011). Kinetic analyses and mathematical modeling of primary photochemical and photoelectrochemical processes in plant photosystems. Biosystems 103, 138–151. doi:10.1016/j.biosystems.2010.10.016
Yakovlev, A. G., and Shuvalov, V. A. (2016). Physical stage of photosynthesis charge separation. Physics-Uspekhi 59, 531–557. doi:10.3367/ufne.2016.02.037701
Yano, J., Kern, J., and Yachandra, V. K. (2024). Structure function studies of photosystem II using X-ray free electron lasers. Annu. Rev. Biophys. 53, 343–365. doi:10.1146/annurev-biophys-071723-102519
Zabret, J., Bohn, S., Schuller, S. K., Arnolds, O., Moller, M., Meier-Credo, J., et al. (2021). Structural insights into photosystem II assembly. Nat. Plants 7, 524–538. doi:10.1038/s41477-021-00895-0
Keywords: chlorophyll-a fluorescence, core complex of photosystem II, protonation, rate-limiting step, structural dynamics, waiting time
Citation: Magyar M, Sipka G, Domonkos I, Chen X, Wang X, Han G, Shen J-R, Lambrev PH and Garab G (2025) The role of protonation processes in the gradual formation of the light-adapted charge-separated state of photosystem II. Front. Photobiol. 3:1623224. doi: 10.3389/fphbi.2025.1623224
Received: 05 May 2025; Accepted: 18 June 2025;
Published: 02 July 2025.
Edited by:
Stefano D.' Alessandro, University of Turin, ItalyReviewed by:
Ganesh Mahadeo Nawkar, Université de Lausanne, SwitzerlandRana Hussein Ali, Goethe University Frankfurt, Germany
Copyright © 2025 Magyar, Sipka, Domonkos, Chen, Wang, Han, Shen, Lambrev and Garab. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Melinda Magyar, bWFneWFyLm1lbGluZGFAYnJjLmh1