 Michael J. Sieler Jr.1
Michael J. Sieler Jr.1 Colleen E. Al-Samarrie1
Colleen E. Al-Samarrie1 Kristin D. Kasschau1Mike L. Kent1,2,3
Kristin D. Kasschau1Mike L. Kent1,2,3 Thomas J. Sharpton1,4*
Thomas J. Sharpton1,4*- 1Department of Microbiology, Oregon State University, Corvallis, OR, United States
- 2Department of Biomedical Sciences, Oregon State University, Corvallis, OR, United States
- 3Zebrafish International Resource Center, University of Oregon, Eugene, OR, United States
- 4Department of Statistics, Oregon State University, Corvallis, OR, United States
Introduction: As climate change increases global water temperatures, ecologists expect intestinal helminth infection ranges to expand and increase the health burden on aquatic organisms. However, the gut microbiome can interact with these parasites to influence infection outcomes, raising the possibility that its response to increasing temperatures may help buffer against increased infection burden or worsen infection outcomes (e.g., inflammatory bowel disease). To evaluate this hypothesis, we sought to determine if the microbiome is resistant or resilient to the stressors of increased water temperature, helminth exposure, and their combination, and whether this variation linked to infection outcomes.
Methods: We leveraged the zebrafish (Danio rerio) model organism to measure how these variables relate to the temporal dynamics of the gut microbiome. In particular, we exposed adult zebrafish to Pseudocapillaria tomentosa, parasitic capillarid with a direct life cycle, across three different water temperatures (28°C, 32°C, 35°C), and analyzed fecal microbiome samples at five time points across 42 days.
Results: Our findings show that parasite exposure and water temperature independently alter gut-microbiome diversity. Moreover, water temperature moderates the association between parasite infection and the gut microbiome. Consistent with this observation, yet counter to prevailing expectations, we find that increasing water temperature reduces P. tomentosa infection worm development and overall abundance in zebrafish. The decline in worm burden at 35°C may be due to either direct thermal inhibition of P. tomentosa development or temperature-mediated interactions with the host microbiome and immune response.
Discussion: Overall, our results indicate that water temperature alters the contextual landscape of the gut microbiome and shapes its response to an intestinal parasite in zebrafish. To our knowledge, this represents the first report of elevated temperature constraining nematode development in a fish host, underscoring that climate change may impose unanticipated, context-dependent impacts on vertebrate gut microbiomes and health outcomes.
Introduction
The steady increase in global temperatures due to climate change challenges vertebrate health (Acevedo-Whitehouse and Duffus, 2009). These threats to vertebrate health take on many forms, including the expected expansion of infectious agents (Mas-Coma et al., 2008; El-Sayed and Kamel, 2020). Of particular concern are the increased infection burdens faced by aquatic organisms experiencing increasing water temperatures (Sydeman et al., 2015). Due in part to the varied coincident effects of climate, the impacts of a warming climate on aquatic organisms are anticipated to be nonuniform in effect (Tomanek, 2010; Sydeman et al., 2015) and vary biogeographically (Ackerly et al., 2010; Sydeman et al., 2015), which in turn complicates harm mitigation and conservation efforts (Duarte et al., 2020). Consequently, there’s an urgent need to better understand climate change’s contextual impacts on organisms depending on the unique environmental conditions of the ecosystems they inhabit.
In recent years researchers have considered that climate change may also elicit harm to vertebrates by disrupting the composition of their gut microbiome (Greenspan et al., 2020). While prior work has shown that varying temperatures impacts gut microbiome composition across a variety of vertebrate host species (Fontaine and Kohl, 2023), less is known about how coincident variables, such as parasite or pathogen exposure, collide with temperature to drive variation in the gut microbiome. Recent work in fish underscores that intestinal parasites alone can restructure community composition and host physiology, even without a thermal component (Zhao et al., 2024; Kashinskaya et al., 2023; Kumar et al., 2024). Yet, to our knowledge, no study has investigated how rising temperature and intestinal parasite exposure together shape both gut-microbiome dynamics and infection outcomes in a vertebrate host. Whether warming amplifies parasite-induced dysbiosis, buffers the host via microbiome-mediated resistance, or simply constrains the parasite itself remains unknown. Filling this gap is critical for forecasting disease risk under climate change and for pinpointing microbiome traits that promote host resilience. These potential interaction effects are important to quantify, because it may be that they elicit even greater effects on the gut microbiome than anticipated by investigations of temperature alone, and could possibly result in increased frequency of dysbiotic disorders. It’s important to elucidate these interactions because increasing work points to the gut microbiome as a key determinant of whether vertebrate physiology is able to buffer against stress (Fontaine et al., 2022; Fontaine and Kohl, 2023), and whether temperature induced perturbations to the gut microbiome may sensitize individuals to subsequent stressors (Fassarella et al., 2021).
To answer these questions, we evaluated the gut microbiome’s temporal response to an exogenous stressor across a gradient of environmental conditions. To do so, we levered the zebrafish (Danio rerio) model organism to measure how gut microbiomes differ across fish reared to adulthood at one of three water temperature conditions (28°C, 32°C, or 35°C; Figure 1). Zebrafish are highly thermal tolerant, capable of existing across a wide spectrum of temperature ranges from 4°C to 40°C (López-Olmeda and Sánchez-Vázquez, 2011), but living outside their thermal optimum can come at a physiological and microbial cost (López-Olmeda and Sánchez-Vázquez, 2011; Wang et al., 2022). While much is known about the thermal range of zebrafish, the effects of altered water temperature on their gut microbiome structure has not been elucidated. We also sought to determine if water temperature affected how the microbiome and host responds to exposure to and infection by intestinal nematode Pseudocapillaria tomentosa, a common source of disease in aquariums, specifically zebrafish facilities (Moravec, 1984; Moravec et al., 1999; Kent et al., 2018; Marandi et al., 2025). P. tomentosa is known to cause high mortality and disrupt the gut microbiome (Kent et al., 2018; Gaulke et al., 2019). Yet, it remains unclear whether and how water temperature mediates interactions between the host-microbiome system and P. tomentosa (Marandi et al., 2025). A key advantage of P. tomentosa is its direct life cycle capability, in which infective eggs larvate in ambient water and can be acquired orally by the host, without requiring an intermediate or paratenic host (Kent et al., 2018; Kent et al., 2019). Although, P. tomentosa may use paratenic hosts (e.g., oligochaete worms) in natural settings, these are not required in the controlled laboratory conditions used here (Kent et al., 2018; Kent et al., 2019; Marandi et al., 2025). This feature enables us to disentangle temperature effects on host-microbiome-parasite interactions from confounding mechanisms such as the temperature-sensitive loss of intermediate hosts that commonly constrain parasites with indirect life cycles (Molnár et al., 2013; Marandi et al., 2025). Overall, our study sought to clarify the environmentally dependent context of a gut microbiome’s resistance and sensitivity to climate change-relevant stressors.
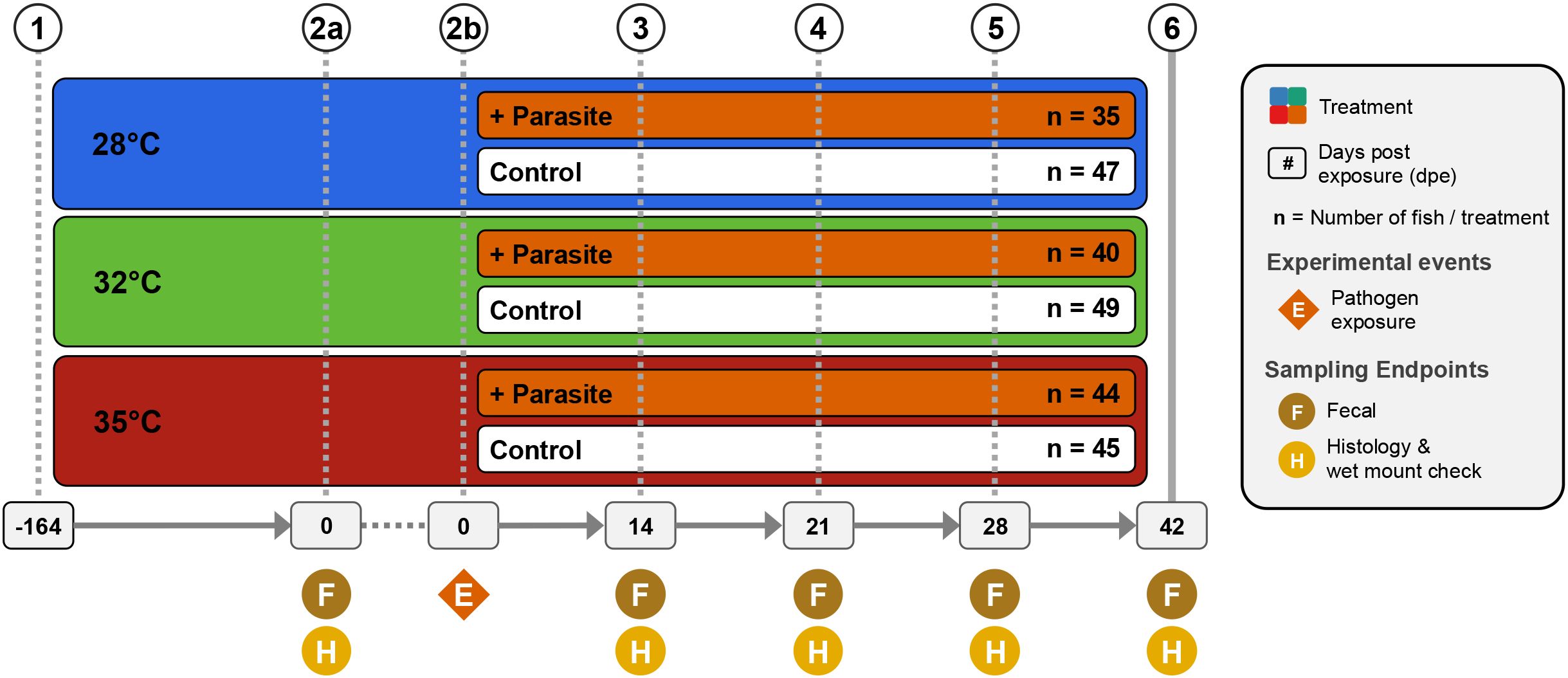
Figure 1. Experimental design showing treatments and husbandry events during the course of the study. Symbols indicate when an experimental event occurred at each time point (1). 260 zebrafish were assigned and acclimated to one of three water temperature groups (e.g., 28°C, 32°C, or 35°C) and reared from 0 to 164 days post fertilization (dpf). (2a) At 164 dpf (or 0 days post exposure; dpe), fecal collections were collected from a random selection of five fish per tank (n = 60). Additionally, histological and wet mount assessments were conducted on selected fish to assess presence of infection and infection burden. (2b) Afterwards, a cohort of fish from each water temperature group were exposed to the nematode Pseudocapillaria tomentosa (4–6). Subsequent fecal samples were collected and histopathological assessments were conducted at 14 dpe (n = 54) (4), 21 dpe (n = 48) (5), 28 dpe (n = 47), and (6) 42 dpe (n = 51).
Results
Water temperature shapes gut microbiome structure
To determine how zebrafish reared across a gradient of increasing water temperatures impacts the structure of the gut microbiome, we reared 260 zebrafish across one of three water temperatures (28°C, 32°C or 35°C) until 206 days-post fertilization (dpf; Figure 1). Additionally, within each temperature cohort, fish were evenly divided into two additional treatment groups: either unexposed or exposed to the intestinal helminthic parasite Pseudocapillaria tomentosa. Microbiome samples were collected at five time points between 164 and 206 dpf. In the parasite exposed cohort, fish were exposed to P. tomentosa following microbiome sampling at 164 dpf, or 0 days post exposure (dpe). Four subsequent microbiome samples were collected at 14 dpe (178 dpf), 21 dpe (185 dpf), 28 dpe (192 dpf), and 42 dpe (206 dpf). Within the parasite unexposed fish cohort, we built generalized linear models (GLM) to determine if water temperature associated with variation in one of four measures of alpha-diversity: Simpson’s Index, Shannon Entropy, richness, and phylogenetic diversity (Supplementary Table S2A.1). An ANOVA test of these GLMs revealed that alpha-diversity varied as a function of temperature for all measures (P<0.05; Figure 2A; Supplementary Table S2A.2), except Shannon Entropy (P>0.05; Supplementary Table S2A.2). A post hoc Tukey test clarified that alpha-diversity scores did not significantly differ between 28°C and 32°C water temperature reared fish for each diversity metric we measured (P>0.05; Supplementary Table S2A.3). However, we observed significant differences in diversity between 28°C and 35°C water temperature reared fish across Simpson’s Index, richness and phylogenetic alpha-diversity measures (P<0.05; Supplementary Table S2A.2), and between 32°C and 35°C water temperature reared fish as measured by richness and phylogenetic diversity metrics (P<0.05; Supplementary Table S2A.2). These results indicate that water temperature associates with fish gut microbiome diversity, and that water temperature may differentially impact particular microbial clades of the gut.
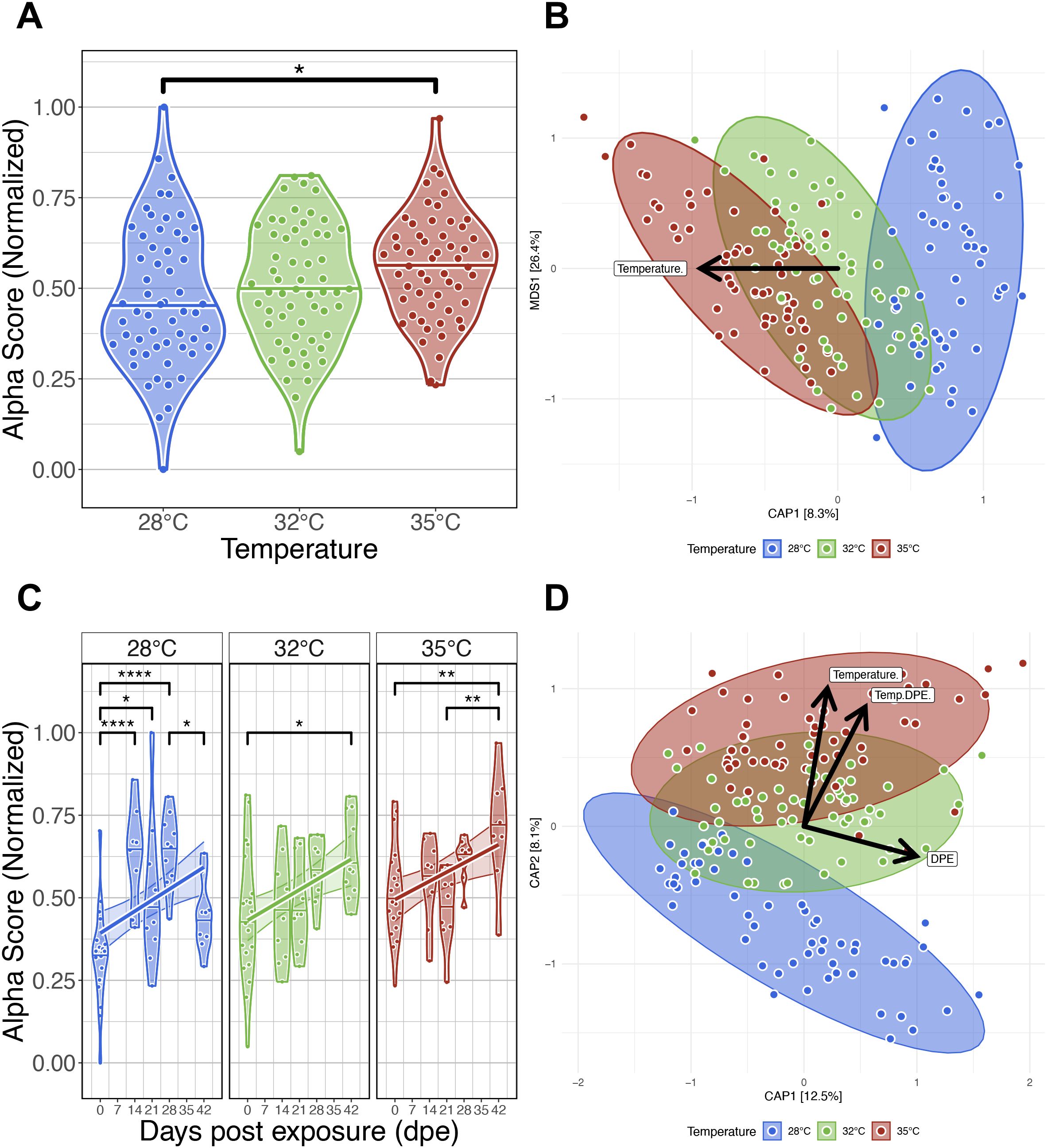
Figure 2. Effects of water temperature on zebrafish gut microbiomes. (A) Simpson’s Index of diversity shows that gut microbiome diversity significantly differs between fish reared at 28°C and 35°C water temperatures. (B) Capscale ordination based on the Bray-Curtis dissimilarity of gut microbiome composition constrained on the main effect of temperature. The analysis shows that gut microbiome composition significantly differs between fish reared at different water temperatures. (C) Simpson’s Index of diversity shows microbial gut diversity increases with time from 0 days post exposure (dpe) to 42 dpe, irrespective of water temperature. (D) Capscale ordination of gut microbiome composition based on the Bray-Curtis dissimilarity constrained on the main effects of water temperature and time (days post exposure, dpe), and their interaction. The analysis shows that shows that gut microbiome composition differs between fish across time depending on water temperature. Ribbons and ellipses indicate 95% confidence interval. Only statistically significant relationships are shown. A “*” indicates statistical significance below the “0.05” level. *p < 0.05, **p < 0.01, ****p < 0.0001. Black arrows indicate direction of greatest change in the indicated by covariates.
To evaluate how temperature associates with microbiome composition in parasite unexposed fish, we quantified dissimilarity amongst all samples and generated distance matrices using the Bray-Curtis, Canberra and half-weighted UniFrac distance metrics. Using permutational multivariate analysis of variance (PERMANOVA), we assessed whether increasing water temperatures explained variance in gut microbial community composition. A PERMANOVA test indicated that microbial communities were significantly stratified by water temperature as measured by all beta-diversity metrics (PERMANOVA, P<0.05; Figure 2B; Supplementary Table S2B.1). These results indicate that microbial communities of fish reared at the same water temperature are more consistent in composition to one another than fish reared at different water temperatures. Additionally, we assessed beta-dispersion, a measure of variance, in the gut microbiome community compositions for each water temperature group. We find the beta-dispersion levels did not significantly differ between the water temperature groups (P>0.05; Supplementary Table S2B.2). These results indicate that fish reared at different water temperatures are consistent in community composition.
Next, we compared our results across five time points between 0- and 42 dpe to determine how water temperature impacts the successional trajectories of gut microbiome diversity and composition. Linear regression revealed gut microbial alpha-diversity was significantly associated with the main effect of time for each alpha-diversity metric we assessed (P<0.05; Figure 2C; Supplementary Table S2C.1, 2). Moreover, we found a temperature dependent effect on time as measured by richness and phylogenetic diversity metrics (P<0.05; Supplementary Table S2C.1, 2). A post hoc Tukey test clarified that microbiome diversity significantly differed between 0- and 42 dpe fish reared at 28°C as measured by richness and phylogenetic diversity (P<0.05; Supplementary Table S2C.3), between 0- and 42 dpe fish reared at 32°C as measured by all alpha-diversity metrics (P<0.05; Supplementary Table S2C.3), and between 0- and 42 dpe fish reared at 35°C as measured by Shannon Entropy and Simpson’s Index (P<0.05; Supplementary Table S2C.3). These results indicate that gut microbial alpha-diversity increases over time irrespective of water temperature.
A PERMANOVA test detected significant clustering of microbial gut community composition based on the interaction of water temperature and time as measured by all beta-diversity metrics (PERMANOVA, P<0.05; Figure 2D; Supplementary Table S2D.1). These results indicate that microbial communities of fish reared at the same water temperature are more consistent in composition to one another across time than fish reared at different water temperatures. Moreover, a pairwise analysis of beta-dispersion found significantly elevated levels of dispersion between fish reared across different temperatures and time as measured by all beta-diversity metrics (P<0.05; Supplementary Table S2D.2). These results indicate that gut microbial community composition varies inconsistently between water temperature groups in a time-dependent manner. Collectively, these results indicate that zebrafish gut microbiomes communities stratify by temperature, and the trajectory of gut microbiome successional development varies depending on water temperature.
Infection burden is highest in fish reared at lower water temperatures
Next, we evaluated infection outcomes of zebrafish reared at different water temperatures and exposed to the intestinal helminthic parasite Pseudocapillaria tomentosa. To determine whether water temperature affects infection burden, we exposed zebrafish to 50 P. tomentosa eggs per liter of tank water at 164 days post-fertilization (dpf). Infection outcomes were assessed using wet mount and histological evaluation at 0, 14, 21, 28, and 42 days post-exposure (dpe). We built a negative binomial general linear model to compare infection burden (total worm counts) between fish reared at different water temperatures (Supplementary Table S3B.1). The regression analysis found a statistically significant effect of temperature on infection burden (P < 0.05; Figure 3A; Supplementary Table S3A.2). However, we did not find a statistically significant interaction effect between water temperature and time on infection burden (P > 0.05; Supplementary Table S3B.3).
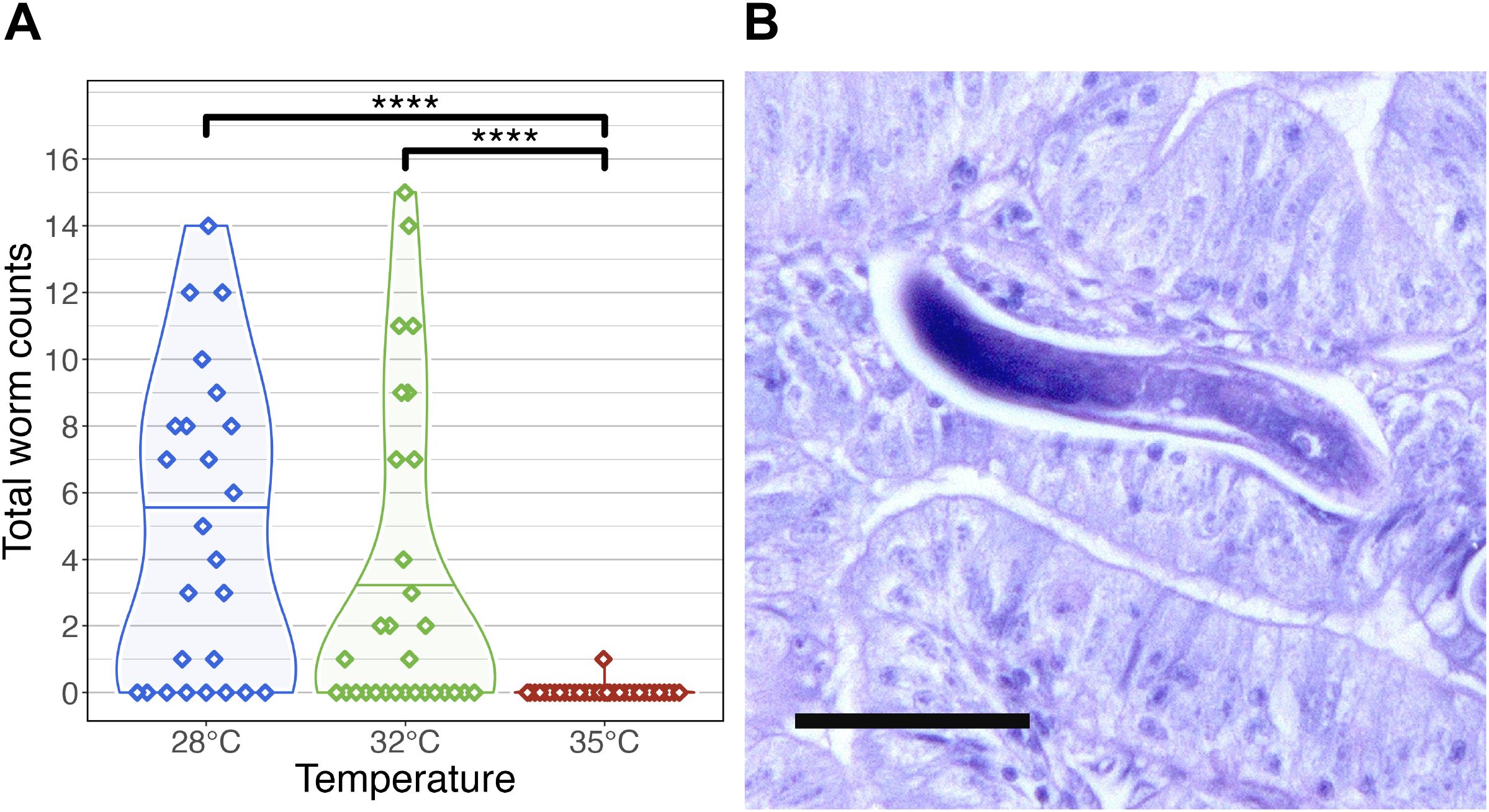
Figure 3. Infection outcomes in zebrafish exposed to Pseudocapillaria tomentosa. (A) Infection outcome analysis of fish exposed to P. tomentosa (n = 89) by temperature. Fish reared at 28°C and 32°C water temperatures had significantly different infection burden to fish reared at 35°C water temperature. Only one fish in our microbiome analysis reared at 35°C was identified as being positively infected by wet mount. Only statistically significant relationships are shown. ****p < 0.0001. (B) Histological sections stained with H&E stain in zebrafish exposed to P. tomentosa examined at 35°C at 21 days post exposure. Arrow = larval worms, sagittal and cross sections. Bar = 50 µm.
Across time points, fish reared at 28°C exhibited the highest mean infection burden (4.86 worms per fish), followed by those at 32°C (3.6 worms per fish). Notably, at 14 dpe, fish at 32°C had a slightly higher infection burden (3.3 worms per fish) than those at 28°C (2.3 worms per fish). Tukey’s post hoc test revealed that infection burden was significantly higher in fish reared at 28°C and 32°C compared to those at 35°C (P < 0.05; Figure 3A; Supplementary Table S3B.3). Only a single larval worm was detected by wet mount in two fish from the 35°C group, while histological examination revealed a slightly higher prevalence, with larval worms observed in 9 out of 32 fish at this temperature (Figure 3B; Supplementary Table S3B.1). These results indicate that infection burden is highest at lower water temperatures. Because P. tomentosa completes a direct life cycle with no intermediate host, this steep decline at 35°C suggests a direct upper thermal limit on egg hatching or early larval survival in addition to. Alternatively, this could be driven by temperature-mediated changes in host immune response or microbiome resistance. We also examined the development of mature female worms across temperature conditions. At 28°C, mature female worms were first detected at 28 dpe in 7 fish, whereas at 32°C, mature females were only observed in 4 fish. Interestingly, at 14 dpe, a single mature female was identified in a fish reared at 32°C, marking the earliest recorded instance of worm maturation at this temperature.
Additionally, we compared the sensitivity of infection detection between histology and wet mount methods on a subset of fish selected for microbiome analysis (n = 120; Supplementary Figure S3C; Supplementary Table S3C.1). McNemar’s test revealed significant differences in detection sensitivity under specific conditions. At 35°C and 21 dpe, histology identified significantly more infections than wet mount (χ² = 4.17, P < 0.05; Supplementary Figure S3C; Supplementary Table S3C.3), with 6 samples testing positive by histology alone compared to 0 by wet mount alone. No statistically significant differences were observed at other temperature and dpe combinations (P > 0.05; Supplementary Table S3C.3). In cases where all samples were concordant (e.g., 28°C at 28 dpe and 35°C at 28 dpe), McNemar’s test could not be computed due to the absence of discordant pairs. These findings suggest that histological methods may be more sensitive than wet mounts, particularly at higher temperatures and intermediate time points. Collectively, these results suggest that higher water temperatures may have a protective effect against infection burden, limiting worm establishment and development in zebrafish.
Gut microbiome response to parasite exposure varies across water temperature
To investigate how parasite exposure affects the gut microbiome under varying water temperatures, we analyzed fecal samples from exposed and control fish at multiple time points. P. tomentosa is known to alter the zebrafish gut microbiome (Gaulke et al., 2019), but it remains unclear how increasing water temperatures affect this response. We collected fecal samples for microbiome analysis of fish in the parasite exposed cohort at 14-, 21-, 28-, and 42 dpe. Similar to our parasite unexposed fish microbiome analyses, we built generalized linear models (GLM) to determine if temperature, time or their combination associated with variation in measures of microbial diversity and composition of parasite exposed fish (Supplementary Table S4A.1). An ANOVA test of these GLMs revealed that alpha-diversity varied as a function of temperature for all measures (P<0.05; Figure 4A; Supplementary Table S4A.2). A post hoc Tukey test clarified that gut microbial diversity between 28°C and 32°C water temperature reared fish significantly differed for all alpha-diversity metrics (P<0.05; Supplementary Table S4A.3), and gut microbial diversity differed between 28°C and 35°C water temperature reared fish as measured by Simpson’s Index. However, we did not find significant differences in gut microbial diversity between 32°C and 35°C water temperature reared fish for all alpha-diversity metrics, or between 28°C and 35°C water temperature reared fish as measured by Shannon Entropy, richness and phylogenetic diversity metrics. These results indicate that moderate increases in water temperature promotes gut microbial diversification in parasite exposed fish, but diversification of gut microbes plateaus in parasite exposed fish reared at higher water temperatures.
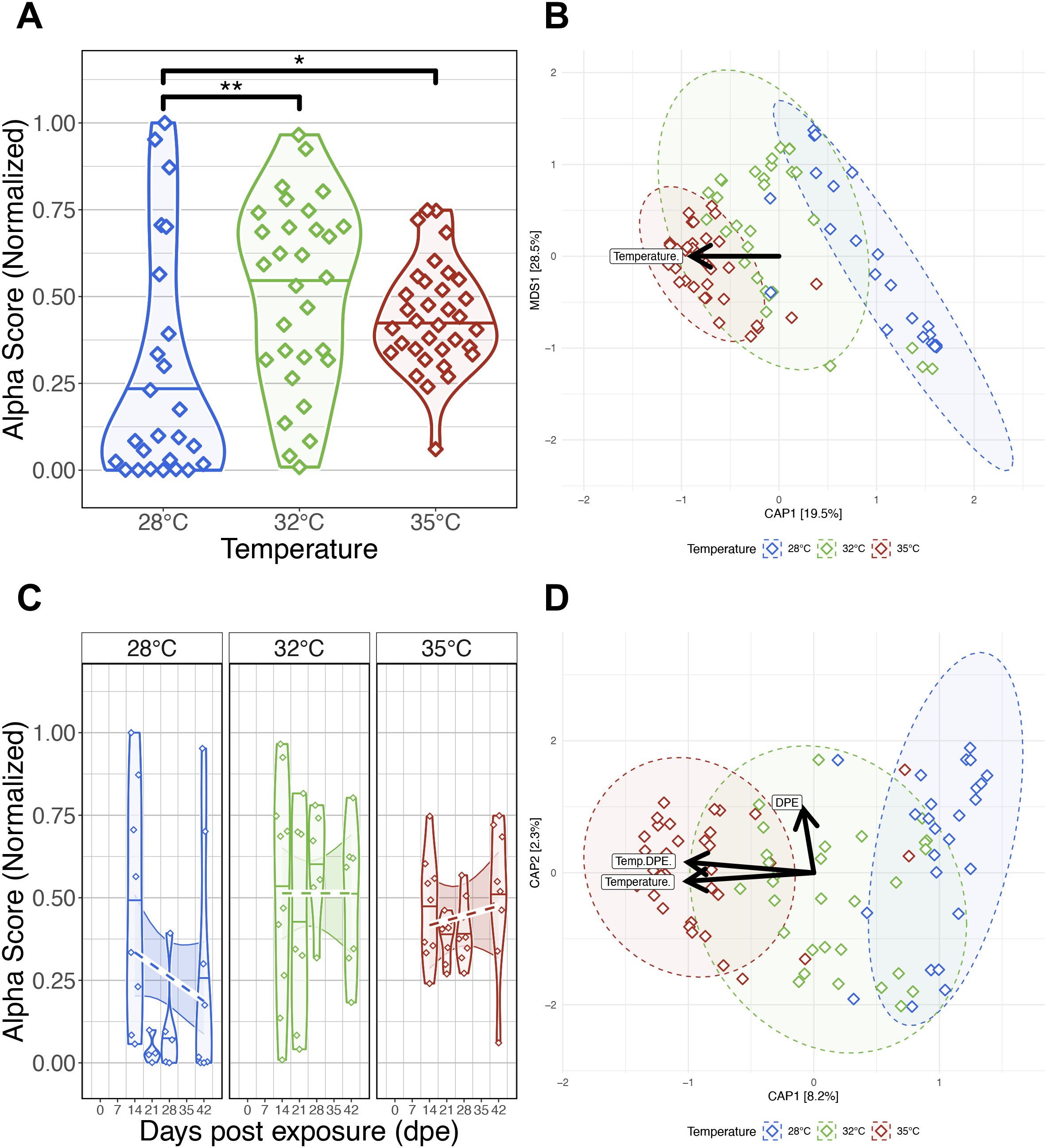
Figure 4. Effects of Pseudocapillaria tomentosa exposure on zebrafish gut microbiomes reared at different water temperatures. (A) Simpson’s Index of diversity shows that gut microbiome diversity significantly differs between fish reared at 28°C water temperature to fish reared at 32°C and 35°C water temperatures. (B) Capscale ordination based on the Bray-Curtis dissimilarity of gut microbiome composition constrained on the main effect of temperature. The analysis shows that gut microbiome composition significantly differs between parasite exposed fish reared at different water temperatures. (C) Simpson’s Index of diversity shows microbial gut diversity decreases with time from 0 days post exposure (dpe) to 42 dpe in parasite exposed fish reared at 28°C water temperature. (D) Capscale ordination of gut microbiome composition based on the Canberra dissimilarity constrained on the main effects of water temperature and time (days post exposure, dpe), and their interaction. The analysis shows that shows that gut microbiome composition differs between parasite exposed fish across time depending on water temperature. Ribbons and ellipses indicate 95% confidence interval. Only statistically significant relationships are shown. A “*” indicates statistical significance below the “0.05” level. Black arrows indicate direction of greatest change in the indicated covariates. A “*” indicates statistical significance below the “0.05” level. *p < 0.05, **p < 0.01.
For each beta-diversity metrics we considered, PERMANOVA tests found that temperature significantly explained the variation in microbiome composition in parasite exposed fish (PERMANOVA, P<0.05; Figure 4B; Supplementary Table S4B.1). These results indicate that gut microbiome communities of parasite exposed fish reared at the same water temperature are more similar to each other than fish reared at different water temperatures. Additionally, we found beta-dispersion levels were significantly elevated between water temperature groups (P<0.05; Supplementary Table S4B.2). A post hoc Tukey test clarified that beta-diversity dispersion levels were highest in fish reared at 28°C, followed by fish reared at 32°C and 35°C water temperatures (P<0.05; Supplementary Table S4B.3). These results indicate that that gut microbiome communities of parasite exposed fish reared at lower water temperatures are more inconsistent in composition than parasite exposed fish reared at higher water temperatures.
Next, we compared our results across five time points to evaluate how parasite exposure and water temperature impacted gut microbiome diversity and composition. An ANOVA test did not find significant main effects of time as measured by Shannon Entropy and Simpson’s Index (P>0.05; Supplementary Table S4C.2), but found marginally significant main effects of time as measured by richness and phylogenetic diversity (P=0.064 and P=0.078; Supplementary Table S4C.2). Moreover, linear regression did not reveal significant interaction effects between temperature and time across all alpha-diversity metrics (P>0.05; Figure 4C; Supplementary Table S4C.2). These results indicate increasing water temperatures generally do not consistently impact microbial gut diversification over time in parasite exposed fish, and particular microbial clades appear more sensitive to the effects of time depending on temperature.
PERMANOVA tests found that community composition was best explained by the interaction effects between temperature and time using the Canberra beta-diversity metric (PERMANOVA, P<0.05; Figure 4D; Supplementary Table S4D.1), but a significant interaction effect was not observed using the Bray-Curtis and half-weighted UniFrac dissimilarity metrics (P>0.05; Supplementary Table S4D.1). Given how these metrics weigh the importance of rarer (e.g., Canberra) versus abundant (e.g., Bray-Curtis) microbial community members, these results indicate that abundant members of the microbiome community are more robust to the effects of temperature across time in parasite exposed fish, while rarer taxa are more sensitive to the effects of time depending on temperature. Moreover, a pairwise analysis of beta-dispersion found significantly elevated levels of dispersion between fish reared across different temperatures and time as measured by all beta-diversity metrics (P<0.05; Supplementary Table S4D.2). These results indicate that parasite exposure inconsistently impacts gut microbial community composition across time depending on temperature (P<0.05; Supplementary Table S4D.2). Collectively, these results indicate that parasite exposure can impact gut microbiome diversity and composition, and these impacts are greatest at lower temperatures.
Gut microbiome response has a non-linear relationship with infection burden
Given the differences we observed in gut microbiome diversity and composition across parasite exposed fish reared at different water temperatures, we further investigated whether gut microbiomes of parasite exposed fish vary depending on presence of infection and infection burden. Linear regression did not find significant main effects of presence of infection or significant interaction effects between presence of infection and temperature on gut microbial alpha-diversity for all metrics we measured (P>0.05; Figure 5A; Supplementary Table S5A.1, 2). These results indicate that gut microbial diversity does not differ as a function of presence of infection. Moreover, a PERMANOVA analysis found microbial community composition was best explained by presence of infection as measured by Canberra (PERMANOVA, P<0.05; Figure 5B; Supplementary Table S5B.1), but a significant result was not observed by the other beta-diversity metrics we measured. Additionally, we did not find statistically significant results between the interaction effects of water temperature and presence of infection on gut microbial community composition. These results indicate that rarer members of microbial communities of parasite exposed fish vary by presence or absence of infection, but abundant microbes do not. However, we did detect elevated levels of beta-dispersion across fish reared at different water temperatures depending on presence of infection (P<0.05; Supplementary Table S5B.2). These results indicate that gut microbiome composition inconsistently varies between fish depending on presence of infection and water temperature.
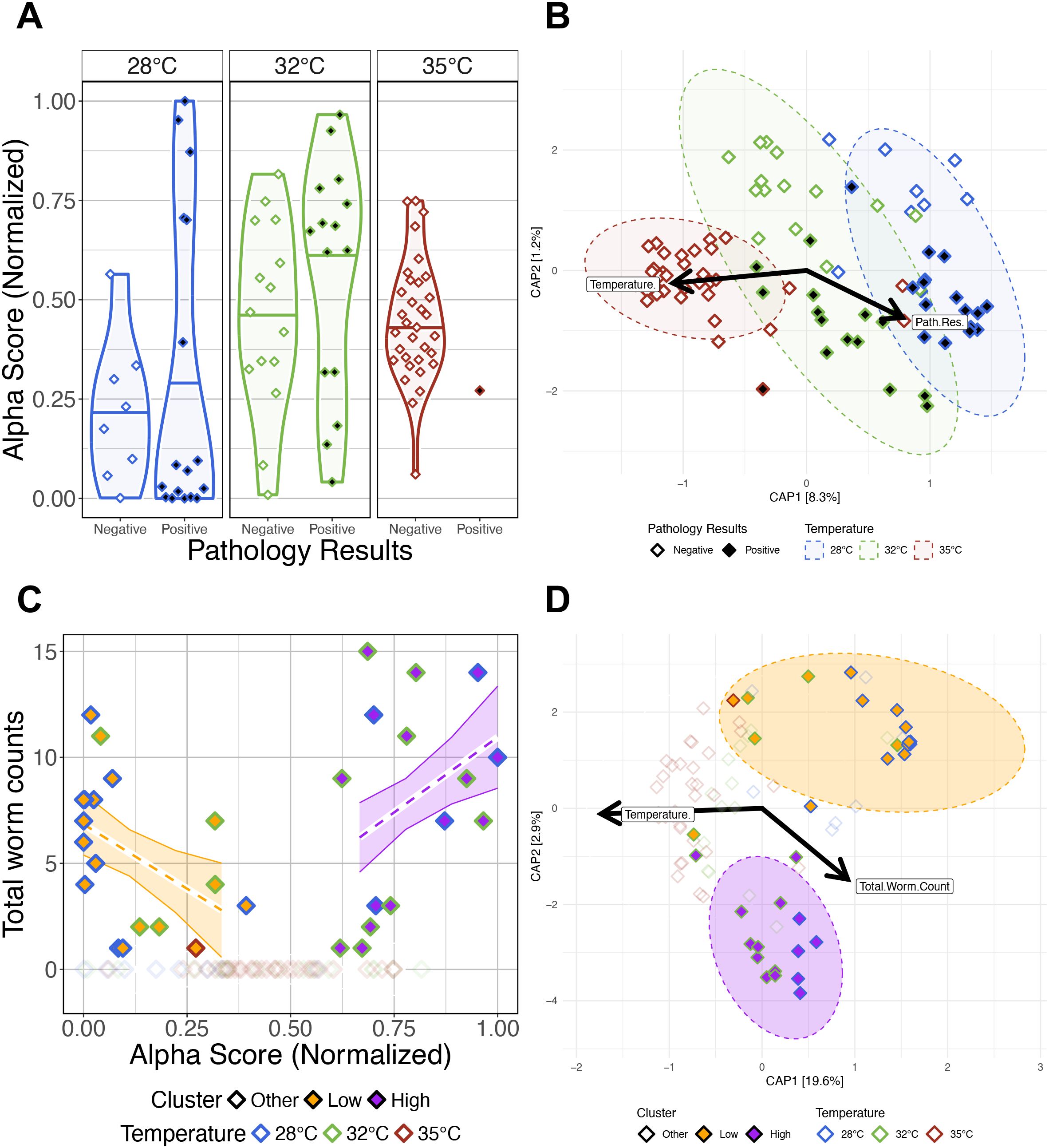
Figure 5. The impacts of presence of infection and infection burden on the gut microbiomes of Pseudocapillaria tomentosa exposed zebrafish. (A) Simpson’s Index for diversity of parasite exposed fish. Gut microbial alpha-diversity does not significantly differ between fish reared at the same water temperature depending on presence of infection. (B) Capscale ordination based on the Canberra dissimilarity of gut microbiome composition of parasite exposed fish constrained on the main effects of temperature and pathology result. The analysis shows that gut microbiome composition significantly differs between positively infected fish reared at different water temperatures. (C) Infection burden (total worm counts) is positively correlated with lowest or highest alpha diversity scores in positively infected fish. (D) Capscale ordination based on the Bray-Curtis dissimilarity of gut microbiome composition constrained on the main effects of water temperature and infection burden. The analysis shows that gut microbiome composition significantly differs between clusters of Low, High and Other fish. Samples points are colored by water temperature, and filled by “Cluster” grouping. Samples with at least one detectable worm and an alpha-diversity score less than 0.5 are categorized as Low (orange fill), samples with at least one detectable worm and an alpha-diversity score greater than 0.5 are categorized as High (purple fill), and samples with no observable infection are categorized as Other (white and transparent fill). Ribbons and ellipses indicate 95% confidence interval. Only statistically significant relationships are shown. Black arrows indicate statistically significant covariates and direction of greatest change in the indicated covariates.
Next, we investigated how infection burden (i.e., number of intestinal worms detected) impacted parasite exposed fish gut microbiome diversity and composition. We used GLMs to determine if infection burden associated with variation in gut microbial alpha-diversity (Supplementary Table S5C.1). An ANOVA test of these GLMs revealed that alpha-diversity varied as a function of infection burden as measured by Shannon Entropy and Simpson’s Index (P<0.05; Supplementary Table S5C.2.2), but the interaction effects between infection burden and water temperature did not significantly explain variation in alpha-diversity across all measures (P>0.05; Supplementary Table S5C.2.2). These results indicate that gut microbial diversity varies as a function of parasitic worm count. A PERMANOVA analysis found microbial community composition was best explained by infection burden as measured by all beta-diversity metrics (PERMANOVA, P<0.05; Supplementary Table S5C.2.1), but the interaction effect between infection burden and temperature was not significant (P>0.05; Supplementary Table S5C.2.1). These results indicate that higher infection burden drives increased inconsistency in gut microbial composition, regardless of water temperature.
Upon closer inspection of our infection burden results, we observed a non-linear relationship between infection burden and alpha-diversity scores, where highest infection burden associated with either highest or lowest alpha-diversity scores. To further explore this non-linear relationship between gut microbial diversity and infection burden, we grouped parasite exposed fish samples based on their alpha-diversity scores and infection burden. Parasite exposed fish samples with at least one intestinal worm detected were classified as “Low” or “High” if their alpha-diversity score was less than or greater than the median alpha-diversity score, respectively. Fish with zero detected worms were classified as “Other”. When grouping samples either Low or High based on alpha-diversity scores as measured by the Simpson’s index, we find that the samples in the Low group are composed of 67% 28°C and 33% 32°C water temperature reared fish, samples in the High group are composed of 33% 28°C and 67% 32°C water temperature reared fish, and samples in the Other group are composed of 14% 28°C, 27% 32°C, and 59% 35°C water temperature reared fish (Supplementary Table S5C1.0). These results indicate that group membership trends with water temperature. To test this supposition, we used GLMs to determine if infection burden associated with variation in alpha-diversity score grouping (Supplementary Table S5C.1.1). An ANOVA test of these GLMs revealed significant main effects of group for all alpha-diversity measures (P<0.05; Figure 5C; Supplementary Table S5C.2.1), and significant interaction effects between group and alpha-diversity score. Notably, fish in the Low group had a significant negative slope and fish in the High group had a significant positive slope between alpha-diversity and infection burden as measured by Shannon Entropy and Simpson’s Index. These results indicate that parasite exposed fish have diverging gut microbial alpha-diversity responses to high infection burden.
Additionally, we find that these groups of samples - based on high versus low alpha-diversity scores of parasite exposed fish - also formed two distinct clusters in beta-diversity space. A PERMANOVA analysis detected significant clustering between Low, High, and Other groups across each measure of beta-diversity (PERMANOVA, P<0.05; Figure 5D; Supplementary Table S5D.1.1). However, this effect was weakest when considering the Canberra metric. Furthermore, a pairwise analysis of beta-dispersion finds significantly elevated dispersion levels between group membership as measured by Canberra metric, but not the other beta-diversity metrics (Supplementary Table S5D.2). Given that the Canberra metric gives rarer taxa greater importance in its beta-diversity calculations than the other metrics we evaluated, these results suggest there is more consistency in microbial composition among abundant taxa within samples that share Low or High group membership, but not among more rarer taxa. A post-hoc Tukey test also clarified that beta-dispersion levels are significantly different between fish in the High and Other groups compared to fish in the Low group as measured by the Canberra metric (Supplementary Table S5D.3). Together, these results indicate that rarer members of the gut microbiome are less consistently represented across fish in the Low cluster group as compared to fish in the High and Other cluster groups. Collectively, these results indicate that the microbiome response of fish with heaviest infection burden diverge into two distinct trajectories, which may be influenced by water temperature.
Parasite exposure exacerbates water temperature differences in gut microbiome structure
Next, we sought to determine whether the gut microbiomes of zebrafish exposed to the parasite Pseudocapillaria tomentosa respond differentially compared to parasite unexposed control fish across increasing water temperatures. Prior to the parasite exposure at 164 dpf (or 0 dpe), we collected fecal samples from both cohorts of control and parasite exposed fish. Following fecal sample collection, fish in the parasite exposure group were exposed to P. tomentosa. We collected subsequent fecal samples at 14-, 21-, 28- and 42 dpe. Fecal samples were then measured for gut microbial diversity and composition and compared between parasite unexposed and exposed fish. We built generalized linear models (GLM) to determine if parasite exposure as a function of water temperature associated with microbial diversity and composition measures (Supplementary Table S6A.1). Within pre-exposed (i.e., 0 dpe) samples, we did not observe any significant associations between the interaction effect of parasite exposure and water temperature across any of the alpha-diversity measures (P>0.05; Figure 6A; Supplementary Table S6A.2). These results indicate that at 0 dpe prior to parasite exposure, gut microbial diversity measures of fish reared at the same water temperature are not different from one another. Furthermore, PERMANOVA tests revealed significant differences in microbiome composition between control and pre-exposed fish across all beta diversity metrics. Homogeneity of dispersion tests revealed a significant difference in group variability for Bray-Curtis (P<0.05; Figure 6B; Supplementary Table S6B.2), but not for Canberra or Generalized UniFrac. Post hoc Tukey tests indicated no significant pairwise differences in dispersion for any metric (Supplementary Table S6B.3), suggesting that the observed dispersion effect in Bray-Curtis was not driven by specific group outliers. To assess whether these baseline differences in community variation were structured by rearing tank, we tested whether tank explained variation in microbial community composition within each temperature prior to parasite exposure. PERMANOVA tests revealed that tank effects were strong at 32°C across all distance metrics (P < 0.05; Supplementary Figure S6B.1.1). Furthermore, homogeneity of dispersion tests found that tanks did not differ within temperature groups prior to parasite exposure metric combination (P > 0.05; Supplementary Table S6B.1.2), confirming that the significant tank effects at 32°C reflect shifts in community centroids rather than unequal variances. Given that temperature alone consistently explained the largest share of variation, followed by treatment and the more context-dependent tank effects, these results indicate that before parasite exposure microbial communities differ primarily by water temperature, with additional variability introduced by stochastic differences among tanks.
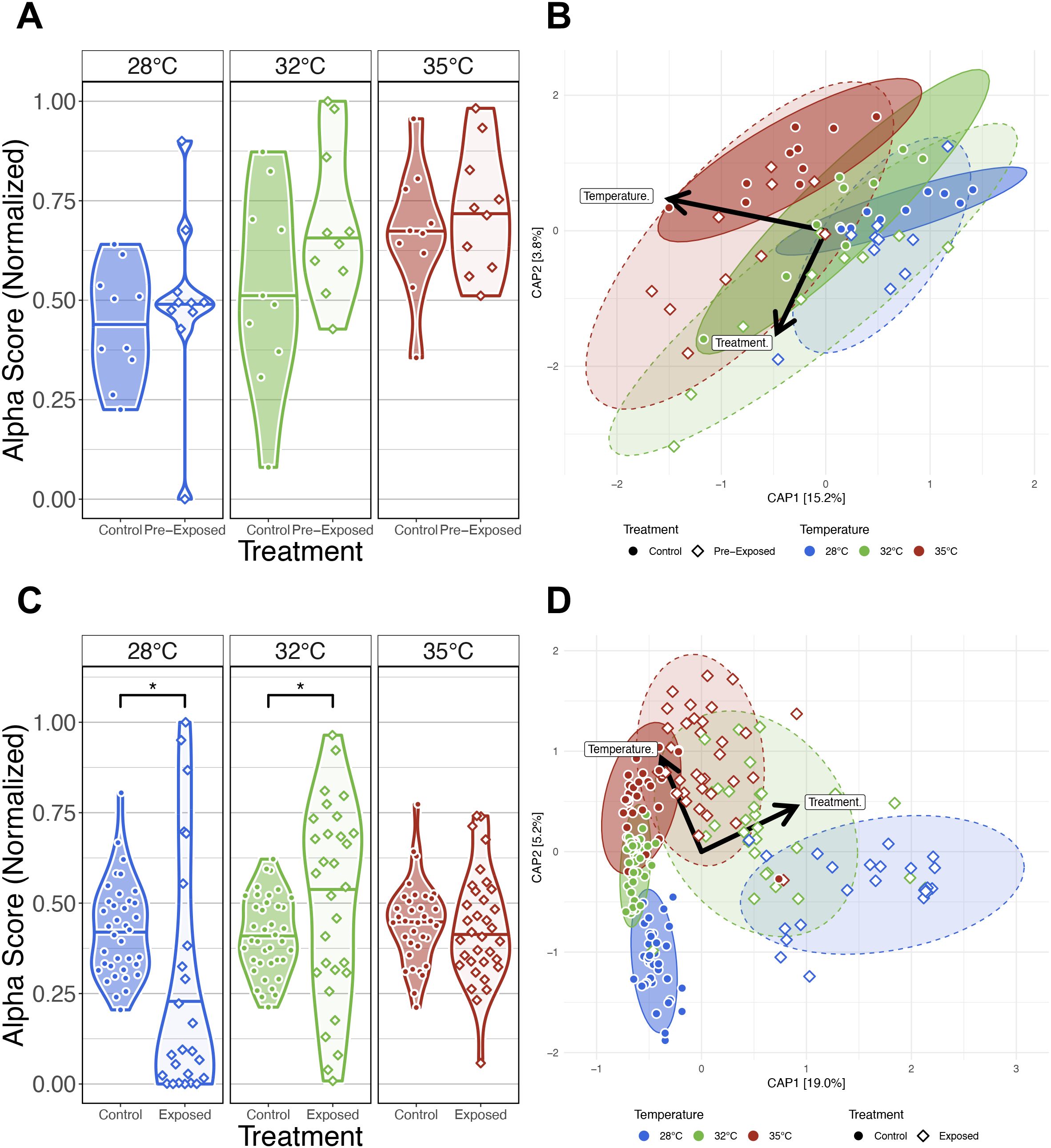
Figure 6. Comparison of the effects of water temperature on the gut microbiome between parasite exposed fish and parasite unexposed fish. (A) Simpson’s Index for diversity of parasite unexposed and pre-exposed fish at 0 days post exposure (dpe). Prior to parasite exposure gut microbial alpha-diversity does not significantly differ between fish reared at the same water temperature. (B) Capscale ordinations based on the Bray-Curtis dissimilarity of gut microbiome composition constrained on the main and interaction effects of temperature and parasite exposure (treatment) of pre-exposure samples at 0 dpe. (C) Simpson’s Index for diversity of parasite unexposed and exposed fish. Gut microbial alpha-diversity significantly differs between parasite exposed fish reared at 28°C and 32°C water temperature relative to unexposed control fish, but gut microbial alpha-diversity does not differ between parasite unexposed and exposed fish reared at 35°C water temperature. (D) Capscale ordinations based on the Bray-Curtis dissimilarity of gut microbiome composition constrained on the main and interaction effects of temperature and parasite exposure (treatment) of post-exposure samples after 0 dpe. The analysis shows gut microbiome composition differs between fish reared at different water temperatures prior to parasite exposure, and parasite exposure further drives these temperature associated differences in microbiome community composition. Ribbons and ellipses indicate 95% confidence interval. Ribbons and ellipses indicate 95% confidence interval. Only statistically significant relationships are shown. A “*” indicates statistical significance below the “0.05” level. Black arrows indicate statistically significant covariates and direction of greatest change in the indicated covariates.
We next compared our results between control and exposed fish across each water temperature to determine how parasite exposure impacts gut microbiome diversity and composition. Linear regression revealed microbial gut alpha-diversity was significantly associated with the interaction effect between temperature and treatment for any alpha-diversity metric we assessed (P<0.05; Figure 6C; Supplementary Table S6C.1, 2). A post hoc Tukey test clarified that microbiome diversity was significantly different between exposure groups of fish reared at 28°C water temperature as measured by Simpson’s Index (P<0.05; Supplementary Table S6C.3), at 32°C water temperature as measured by all alpha-diversity metrics (P<0.05; Supplementary Table S6C.3), and at 35°C as measured by richness and phylogenetic diversity (P<0.05; Supplementary Table S6C.3). These results indicate that gut microbial diversity differs between unexposed and exposed fish depending on water temperature, and parasite exposure uniquely impacts particular microbial clades, rare and abundant taxa depending on water temperature. Additionally, PERMANOVA tests found that microbiome composition differed between control and exposed fish reared at all water temperatures as measured by all beta-diversity metrics (P<0.05; Figure 6D; Supplementary Table S6D.1). These results suggest that the gut microbiomes compositions between control and parasite exposed differed in microbiome community composition regardless of water temperature. Moreover, a pairwise analysis of beta-dispersion found elevated levels of dispersion across all beta-diversity metrics measured, and dispersion levels were highest among parasite exposed fish reared at lower water temperatures (P<0.05; Supplementary Table S6D.2). These results suggest that gut microbiome community composition is less consistent between parasite unexposed and exposed fish reared at lower water temperatures. Collectively, these results demonstrate that water temperature dictates how exposure to parasites alters the temporal trajectory of the gut microbiome.
Gut microbial relative abundance significantly associates with environmental conditions and stressors
Finally, to evaluate how gut microbial abundance is influenced by environmental conditions and stressors (e.g., worm infections), we quantified differential abundance using MaAsLin2. Our analysis revealed 277 unique taxa at the Genus taxonomic level with at least one significant associations between taxon abundance and a covariate (FDR <0.05, Figure 7; Supplementary Table S7A.1). We observed several taxa were significantly associated with the effect of water temperature. Fish reared at 35°C water temperature were enriched for 37 taxa, and depleted of 54 taxa relative to fish reared at 28°C water temperature. Fish reared at 32°C were enriched for 42 taxa, and depleted of 47 taxa relative to fish reared at 28°C water temperature (Figure 7). Notably, Aeromonas and Pseudomonas bacterial abundance significantly associated negatively and positively with the effects of water temperature, respectively. Aeromonas and Pseudomonas are common members of the zebrafish gut microbiome (Semova et al., 2012; Sharpton et al., 2021), and these genera’s bacterial abundance has previously been observed to associate with the effects of water temperature in zebrafish (Wang et al., 2022). These results indicate that gut microbes are differentially selected for across varying water temperatures and time. We also measured how taxon abundance change over time regardless of water temperature. Over the course of 42 days, fish were enriched for 73 taxa and depleted of 36 taxa (Figure 7). Notably, Bosea and Cloacibacterium bacterial abundance were negatively associated with the effects of time. Bosea and Cloacibacterium are common members of the zebrafish gut microbiome (Semova et al., 2012; Stagaman et al., 2020; Sharpton et al., 2021), and were also previously identified as having negative associations with the effects of time in zebrafish (Gaulke et al., 2019). These results indicate that particular members of the gut microbiome associate with time irrespective of water temperature.
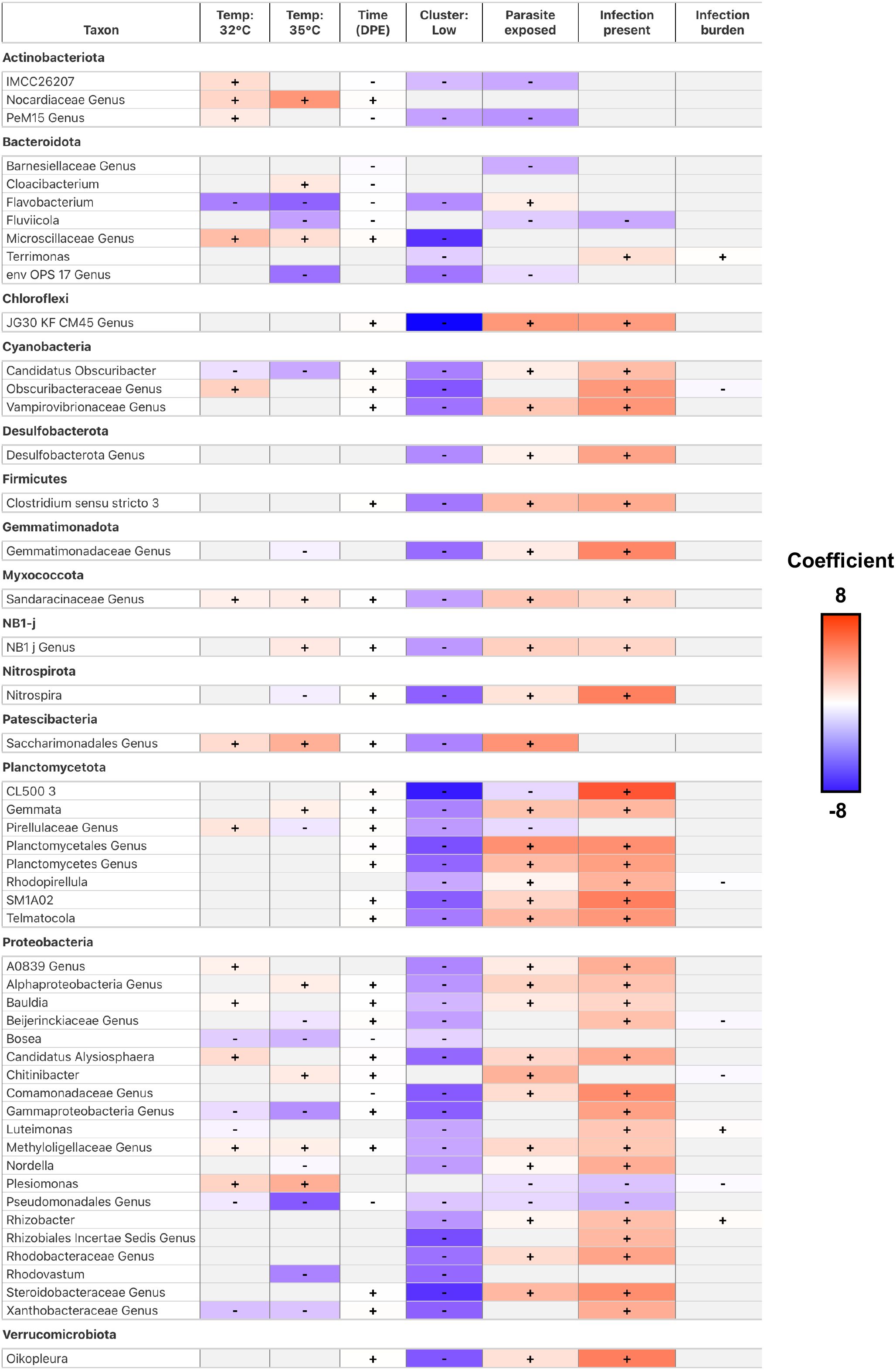
Figure 7. A heatmap of model coefficient values of the top 50 statistically significant abundant gut microbial taxa identified by MaAsLin2. The color of each cell represents the coefficient value and direction (red is positive, blue is negative). A “+” or “-” indicates a statistically significant association was observed between taxon abundance and a covariate. Gray colored cells indicate a significant effect was not observed.
Next, we sought to determine how parasite exposure impacted gut microbial abundance in fish. Fish exposed to P. tomentosa were significantly enriched for 74 taxa, and depleted of 35 taxa relative to parasite unexposed fish (Figure 7). Notably, we find Aeromonas, Chitinibacter, and Flavobacterium are positively associated with parasite exposure, while Plesiomonas, Phreatobacter and Cetobacterium are negatively associated with parasite exposure. With the exception of Phreatobacter and Cetobacterium, these data are consistent with our prior work that found P. tomentosa exposure associated with altered bacterial abundance of these members of the zebrafish gut microbiome (Gaulke et al., 2019). We further investigated the effects of parasite exposure and measured how infection burden or presence of infection impacted gut microbial abundance. Fish with higher infection burden (i.e., number of parasitic worms present) enriched for 49 taxa and were depleted of 13 taxa, while fish that were positively infected enriched for 117 taxa and were depleted of 5 (Figure 7). Notably, we find abundance of members of Cetobacterium, Shewanella, Vibrio and Zooglogea are negatively associated with infection burden. These taxa are identified as common members of the zebrafish gut microbiome (Stephens et al., 2016). Because infection burden varied widely at 28°C and 32°C, we ran temperature-stratified MaAsLin2 models to link genus-level abundance to worm count (FDR < 0.05; Supplementary Table S7A.2.1, 2). At 28°C, 55 genera were significantly associated with burden, whereas 45 genera were significant at 32°C (FDR < 0.05; Supplementary Table S7A.2.3). Eight genera showed concordant responses across the two temperatures and only Novispirillum displayed an opposite trend, indicating strong directional consistency. Shared positive correlates included Bryobacter, Vampirovibrio and Inhella, while Cetobacterium and Shewanella were consistently depleted in heavily infected fish. Temperature-specific effects were evident: 46 genera were unique to 28°C, led by Paraclostridium and Rubrivivax, whereas 36 genera were unique to 32°C, with Novispirillum exhibiting the most significant coefficient (FDR < 0.05; Supplementary Table S7A.2.3). The ten taxa with the smallest q-values further emphasize that nine of the strongest signals arise at 28°C, underscoring the broader taxonomic shift that accompanies high parasite burden at the lower temperature (FDR < 0.05; Supplementary Table S7A.2.4). Together, these data show that parasite burden has a pronounced yet partly temperature-dependent influence on zebrafish gut microbial abundance.
To gauge how strongly the microbiome, temperature, and time predict infection severity, we trained a random-forest regression model using the MaAsLin2-identified genera plus water temperature and days post exposure (DPE). Ten-fold cross-validation showed that the model reduced prediction error by twenty-five percent compared with a mean-only baseline (RMSE = 2.684 worms versus 3.554; Supplementary Table S7B.1.4, 5) and explained thirty-two percent of the variance in worm burden (R² = 0.321; Supplementary Table S7B.1.4). Permutation importance and a one-hundred-run stability screen highlighted a compact predictive core: Plesiomonas, ZOR0006, Cetobacterium, Bryobacter, and Rhizobacter appeared in the top ten predictors in at least eighty-seven percent of runs (Supplementary Table S7B.1.3). In contrast, temperature and DPE entered the top ten in fewer than two percent of runs, indicating that infection intensity is encoded primarily in the abundance patterns of these key genera rather than the measured environmental covariates alone. Together, these findings identify a concise set of microbiome members that both respond to parasite exposure and collectively capture a substantial share of the variation in worm burden.
To deepen our analysis of parasite exposure on the zebrafish gut microbiome, we investigated how taxon relative abundance associated with gut microbiome diversity and composition. Previously, we found that parasite exposed fish reared at 28°C and 32°C water temperatures clustered into two distinct groups of community composition, which associated with high infection burden and either high or low alpha diversity scores. This observation led us to investigate which gut microbiota might be driving the clustering of the gut microbiomes of heavily infection burdened fish. We did not find significantly abundant taxa in the High group. We detected 1 taxon that was significantly enriched and 192 taxa that were significantly depleted among fish in the Low group (Figure 7). Notably, we find Aeromonas was enriched, while Mycobacterium were depleted in the Low group fish. Some species of Mycobacterium are common pathogens in zebrafish facilities (Sieler et al., 2023). These results indicate that the gut microbiome communities of parasite exposed fish experiencing heavy infection burden stratify into two distinct groups represented by the unique depletion of particular members of the gut microbiome. Collectively, these results indicate that environmental conditions associate with altered gut microbial abundance, and the response of specific members of the gut microbiome to environmental stressors varies depending on environmental conditions.
Discussion
The zebrafish is an important model organism for understanding how environmental stressors impact the microbiome (Stagaman et al., 2020; Sharpton et al., 2021). Our work capitalized on the experimental control and scale afforded by the zebrafish model system to investigate how temperature and parasite exposure interact to influence infection and microbiome outcomes. While previous research has investigated how water temperature (Wang et al., 2022) and parasite exposure (Gaulke et al., 2019) independently impact the zebrafish gut microbiome, no studies in any in vivo experimental system, to our knowledge, have examined the microbiome’s temporal response to the combined effects of increasing water temperature and parasite exposure. Overall, we found that water temperature serves as a key contextual variable that dictates the severity of infection, the developmental state of worms, the composition of the gut microbiome in unexposed fish, and how the gut microbiome responds to parasite exposure and infection. These results underscore that the gut microbiome’s response to, and potentially its ability to buffer against, intestinal parasitic infection is influenced by other exogenous factors, in this case, water temperature. Furthermore, our findings challenge current expectations of climate change’s anticipated impact on aquatic organismal parasite burden (Mas-Coma et al., 2008; El-Sayed and Kamel, 2020). Consequently, it is important that we consider going forward how stacking multiple stressors, an experience inherent to life in the Anthropocene, may accelerate the arrival of dysbioses.
We found that parasitic infection burden was highest among zebrafish reared at ambient water temperatures. Given that P. tomentosa has a direct life cycle with no intermediate or paratenic host under laboratory settings, the temperature-linked drop in worm burden and development at 35°C could stem from direct thermal inhibition of egg hatching, initial, or larval development, rather than microbiome-mediated resistance alone. While prior field studies have documented arrested development of P. tomentosa in colder conditions (Moravec, 1983; Moravec, 1984; Marandi et al., 2025), to our knowledge, this study provides the first controlled laboratory evidence that elevated temperatures can suppress parasite development in a fish host. Consistent with our prior work, temporal trends in P. tomentosa infection burden were similar for fish at ambient temperatures of 28°C (Gaulke et al., 2019). However, contrary to expectations that elevated temperatures increase infection burden, we observed the opposite outcome: fish reared at the highest temperatures of 35°C exhibited the lowest infection burden, with only a few larval-stage worms detected. Because parasite eggs were larvated at ambient temperature before being transferred to warmer tanks, we hypothesize that elevated temperatures may have impaired hatching once the eggs were ingested, reducing overall abundance of worms. Nevertheless, at 35°C, worms that did establish infections persisted but remained in an arrested state out to 28- and 42 days post-exposure (dpe), whereas at 28°C and 32°C, worms completed development and mated within 3 to 4 weeks, consistent with previous observations (Kent et al., 2018). Such arrested developmental stages are characteristic of nematodes approaching their upper thermal limit (e.g., Wuchereria bancrofti larvae fail to develop above 31°C in mosquitoes), reinforcing the hypothesis that elevated temperature acts directly on the parasite (Lardeux and Cheffort, 1997). Although Kent et al., 2019 demonstrated that P. tomentosa egg larvation is inhibited at temperatures exceeding 40°C, some eggs still larvated following brief exposures to 40°C for 1 or 8 hours (Kent et al., 2019). These findings suggest that egg hatching may be sensitive to thermal stress but not completely abrogated at extreme temperatures. However, no studies have yet examined how larvation and development respond across a more ecologically relevant thermal range. Controlled in vitro hatching assays across 28-35°C will therefore be essential to disentangle parasite-specific constraints from potential host- or microbiome-mediated effects.
These findings are particularly notable given the broad geographic and thermal distribution of P. tomentosa in natural and captive settings. P. tomentosa is a remarkably cosmopolitan parasite, with natural infections reported in a wide variety of freshwater fish species from Europe, the Middle East, and North and Central America (Marandi et al., 2025). The type locality is France, and it is widespread throughout central Europe (Moravec et al., 1999). There are also reports of natural infections in subtropical climates, including southern Mexico. In temperate regions, P. tomentosa infects hosts living in environments where winter water temperatures drop below 10°C and summer temperatures exceed 30°C. It has also been found in freshwater aquarium fishes, which are typically maintained at 20-28°C (Moravec, 1984; Moravec et al., 1999). These reports suggest a broad thermal range for infection under field conditions. Notably, these observations assume that the many global records of P. tomentosa represent a single species, rather than a complex of cryptic, morphologically indistinguishable species. One relevant field study by Moravec et al., 1983 found that parasite development was seasonally arrested at temperatures below 25°C in the Czech Republic (Moravec, 1983), but no prior work has evaluated outcomes at the warmer limits we tested. Our findings begin to define P. tomentosa’s upper thermal boundaries under controlled conditions.
In a broader context, many fish pathogens exhibit upper thermal limits to development and infectivity. For example, the ciliate Ichthyophthirius multifiliis, a common aquacultural pathogen, fails to develop above 30°C (Dickerson, 2006). Comparable upper-temperature ceilings have not yet been documented in fish nematodes, but terrestrial filarial worms offer a parallel example of larval development in mosquito vectors halts once temperatures exceed 31°C (Lardeux and Cheffort, 1997). In parasites with indirect or complex life cycles, warming often suppresses prevalence simply by eliminating intermediate or paratenic hosts (Mas-Coma et al., 2008; Kent et al., 2019). As noted previously, P. tomentosa’s direct life cycle rules out temperature-sensitive effects on intermediate hosts, reinforcing that observed patterns likely stem from direct thermal impacts. These results highlight how climate change may suppress, rather than exacerbate, certain infections and challenging expectations in aquatic disease ecology and emphasizing the need to test thermal constraints across a range of pathogens (Kent et al., 2002).
Future research should investigate whether arrested development in P. tomentosa reflects direct thermal limits or host-mediated processes. Beyond direct effects on parasite development, poikilothermic (i.e., animals with variable body temperature and the inability to regulate it) hosts may gain protections against infection through temperature-dependent immune responses or gene expression changes. While studies on zebrafish immunity under elevated temperatures are limited, prior research in teleosts indicates that immune responses are host- and environment-specific, varying with the direction and duration of temperature shifts (Makrinos and Bowden, 2016; Islam et al., 2022). For example, Dittmar et al. found that immune activity was highest at thermal limits and inversely related to acute temperature shifts in three-spine sticklebacks (Dittmar et al., 2014), whereas Bailey et al. observed suppressed immunity and increased parasite burden in rainbow trout exposed to chronic upper optimal thermal ranges (Bailey et al., 2017). Although these studies differ in exposure regimes to ours, they highlight that colonization resistance may be influenced by temperature-sensitive immune responses and gene expression. Future research integrating immune function, gene expression, and histopathological assessments will be crucial to disentangling the host’s role in colonization resistance under chronic parasite exposure and elevated temperatures. Notably, controlled temperature manipulation is already used to mitigate certain aquaculture pathogens, such as Ichthyophthirius multifiliis (Dickerson, 2006), where increasing tank temperature to 30°C can eliminate infections in susceptible fish. Our findings suggest that similar interventions may help mitigate or delay parasite infection in aquaculture settings.
We also found that zebrafish gut microbiome structure stratified depending on the environmental conditions of increasing water temperature. Our results are congruent with previous research that found increased water temperatures altered zebrafish gut microbial diversity and composition (Wang et al., 2022). Moreover, Wang et al. observed that zebrafish reared at different water temperatures manifested distinct liver carbohydrate metabolism profiles and temperature-dependent sensitivity to irradiation. A unique aspect of our study considered how the gut microbiome temporally varies as a function of water temperature. We found that water temperature acts as a filter on initial zebrafish gut microbiome assembly, and these initial differences in assembly between water temperature remained stable across time. Beyond zebrafish, analogous investigations have investigated how temperature variation shapes gut microbiome composition and function in mammals, fish, and other animal species (Sepulveda and Moeller, 2020; Li et al., 2022). In particular, a recent meta-analysis of aquatic organisms’ response to temperature found similar, but inconsistent results to our study, wherein increasing water temperature is associated with both increases and decreases to gut microbial diversity, differences in gut microbiota community composition, and altered gut taxon abundance (Li et al., 2022). Inconsistencies between prior work and ours could be driven by differences in magnitude of the stressor (i.e., press vs pulse (Relman, 2012);), host species (Ley et al., 2008), facility or habitat effects (Roeselers et al., 2011; Breen et al., 2019; Li et al., 2022), or diet (Sieler et al., 2023). Despite these differences, the results of prior studies in conjunction with ours are consistent with the concept of environmental conditions acting as an abiotic filter to shape initial gut microbiome assembly (Costello et al., 2012) and illicit environmentally dependent responses to biotic exogenous stressors.
Finally, we observed a nonlinear relationship between gut-microbiome diversity and infection outcomes, with water temperature moderating these dynamics. Consistent with our prior research on zebrafish infected by P. tomentosa (Gaulke et al., 2019), heavily infected fish displayed dysbiotic microbiomes that matched the Anna Karenina Principle (AKP) expectation of elevated dispersion (Zaneveld et al., 2017). However, once water temperature was included in our model, the AKP signal weakened and at least two alternative stable states emerged. Viewing these patterns through the lens of community-assembly theory helps reconcile this apparent contradiction (Costello et al., 2012). Temperature acts as a selective abiotic filter that deterministically favors taxa possessing traits that confer thermal tolerance, whereas parasite exposure behaves as a largely neutral process, introducing stochastic variation by differentially perturbing communities without a strong trait-based direction. When the selective (e.g., temperature) and neutral (e.g., parasite) forces interact, they generate divergent assembly trajectories that resemble multiple stable states rather than a single AKP-like dysbiosis. This interpretation is supported by the contrasting dispersion trends in unexposed fish versus exposed fish and by the dispersion results. These findings indicate that deterministic (e.g., selective) and stochastic (e.g., neutral) processes jointly shape the assembly of the zebrafish gut microbiome under combined thermal and parasitic stress. Furthermore, our findings underscore the need to consider both individuals’ temporal and spatial contexts and the balance of neutral and selective drivers when assessing microbiome stability (Jeltsch et al., 2025). Moreover, current homeostatic definitions of stability may be insufficient to describe such dynamic shifts (Sommer et al., 2017; Schlomann and Parthasarathy, 2019; Fassarella et al., 2021). Rather, a homeorhetic framework (Waddington, 1942; Chuang et al., 2019), which conceptualizes stability as a change along a stable trajectory rather than a fixed state, may better capture how microbiomes respond to exogenous stressors across environmental gradients, and could reconcile discrepancies in AKP detection across studies.
In conclusion, we found that water temperature alters the contextual landscape of the microbiome to impact its response to an exogenous stressor of an intestinal parasite. Our work revealed that differences in environmental conditions of water temperature were sufficient to temporally change the gut microbiome’s response to parasitic exposure and impact infection outcomes in zebrafish. While the zebrafish gut microbiome differs taxonomically from other animal-microbiome systems, a considerable amount of functional capacity is shared between animals (Rawls et al., 2004). Thus, zebrafish serve as a powerful model for investigating how environmental changes and stressor exposures influence microbiomes and host health. Our findings have important implications for microbiome research in the context of climate change, demonstrating that rising temperatures may have unexpected effects on gut microbiomes and infection outcomes. Future work should further clarify how gut microbiomes and host responses buffer against combined environmental stressors, ultimately shaping health outcomes in vertebrates.
Methods
Fish husbandry
5D strain zebrafish embryos were obtained from the Sinnhuber Aquatic Resource Center at Oregon State University, and reared in our vivarium at Nash Hall (Corvallis, OR, USA). This facility is specific pathogen-free (SPF) and has no known history of Pseudocapillaria tomentosa or other intestinal parasitic infections (Kent et al., 2011). The vivarium is a single pass flow through, using dechlorinated city water. Fish were then randomly divided into twelve 2.8 L tanks. The temperature was recorded daily and the ambient temperature ranged from 27 to 28°C. All other water conditions were monitored weekly, pH was maintained at 7.6, total ammonia was not detected, and conductivity ranged from 102 to 122. Light in the vivarium was provided for 14 hours/day. Fish were fed Gemma Micro 300 (Skretting; Fontaine-les-Vervins, France) at 1.5% body weight twice daily, except on weekends or during exposure to parasitic eggs. One plastic aquatic plant piece, approximately six inches in length, was added to each tank for enrichment. The use of zebrafish in this study was approved by the Institutional Animal Care and Use Committee (IACUC) at Oregon State University (permit number: 5151).
Temperature exposure
At 5 months old, or 206 days post-fertilization (dpf), corresponding to early adulthood in zebrafish, fish were randomly divided into 12 9.5-L tanks (approximately 25 fish/tank). Each tank was outfitted with a 50W (28°C treatment only) or 100W HG-802 Hygger titanium aquarium heater (Hygger, Shenzhen Mago Co., Ltd., Shenzhen City, Guangdong Province, China). Four of the twelve tanks were assigned to each of the temperature treatments: 28°C, 32°C, or 35°C. These temperatures were selected to simulate baseline (28°C), 32°C reflects near-future warming scenarios (+4°C) (Calvin et al., 2023), and upper sublethal thermal limits (35°C) for zebrafish physiology (Sanders et al., 2015). Two tanks for each temperature were held as pathogen negative controls and two tanks were exposed to Pseudocapillaria tomentosa as described below. Fish were acclimated to the prescribed temperature treatments by increasing the heater thermostat settings by 1°C every two days until the final prescribed temperature was achieved. Two temperature logging thermometers, one for the six pathogen negative control tanks and one for the six P. tomentosa exposed tanks, were rotated through the tanks every two days on weekdays to monitor temperature at each temperature treatment. The average range recorded for the water temperature treatments was +/- 1°C.
Pseudocapillaria tomentosa exposure
Pseudocapillaria tomentosa is monoxenous; no invertebrate or vertebrate intermediate host is required for laboratory transmission. We maintain a laboratory population of infected zebrafish at 26-28°C, from which all P. tomentosa eggs used in this study were obtained (Kent lab, OSU; see Martins et al., 2017) (Martins et al., 2017). Eggs were allowed to larvate for 6 days at 28°C, and fish were exposed at 25 larvated eggs/fish. Water flow was turned off for 36h to enhance exposures, while an airstone was provided to each tank to maintain adequate oxygen levels. This was a lower exposure dose than many of our previous studies (Kent et al., 2018). Therefore, we enhanced exposure adding 1 L of water from a stock tank holding infected fish twice a day during the 36 h hour post exposure period. This additional water supplement was created by siphoning water from the bottom of the exposed stock fish tank because the infectious stage is a larvated egg, which sinks in water.
Infection assessment
Exposed and control fish were collected and examined for worm prevalence, abundance and state of development using wet mounts of whole intestines as described in Schuster et al., 2023 (Schuster et al., 2023). After recording observations in wet mounts, the individual intestine was preserved in Dietrich’s solution and intestines of 95 fish were processed for histology prepared as described in Gaulke et al., 2019 (Gaulke et al., 2019). Here we focused on selected samples from fish from the 35°C group as very few worms were detected by wet mounts in this group. Two stepwise sections, 50 um apart, were obtained from each block to enhance the possibility of larval worms.
Fecal collection
Five fish from each tank were randomly selected for fecal sampling at 0 dpe (n=60; 5 samples/tank), prior to parasite exposure. Subsequent fecal sampling took place at 14- (n = 54), 21- (n = 48), 28- (n = 47), and 42 (n = 51) dpe to parasites. Fecal material was collected as previously described (Sieler et al., 2023). In brief, fish were transferred to 1.4 L tanks (1 fish/tank) containing ~ 0.4 L of fish water at least 30 min after the last feeding of the day. Fish were left to defecate overnight and all fecal material was collected from each tank the following morning in a 1.5ml microcentrifuge tube. Fecal samples were immediately spun at 10k rpm for 2 min, excess tank water was removed, and samples were snap frozen on dry ice and stored at -80°C until processing. However, not all fish produced a fecal sample for a variety of reasons. For instance, experiments involving fish have expected mortality, and fish which died prematurely did not produce fecal samples. Additionally, infection conditions may have prevented infected fish from producing a fecal sample. Instances where fish failed to produce a fecal sample are noted in the metadata sheet.
Microbial 16S rRNA library preparation and sequencing
Microbial DNA was extracted from zebrafish fecal samples and 16S rRNA gene sequence libraries were produced and analyzed following previously described methods (Hammer et al., 2024). DNA was isolated from fecal samples using the DNeasy 96 PowerSoil Pro DNA kits (Qiagen, Hilden, Germany), in accordance with the manufacturer’s directions. In brief, samples were subjected to bead beating for 10 minutes using the Qiagen TissueLyser II, spun a max speed in the centrifuge, supernatant was process using 96 well columns, and DNA was eluted with 100µl Tris buffer. The V4 region of the 16S rRNA gene was PCR amplified using dual-index 16S primers and protocols (Kozich et al., 2013). PCR was performed using 1 µl of purified DNA, 2µl of a 5µM mix of the forward and reverse dual-index primers, 5µl of Platinum II Hot-Start PCR Master Mix (Thermo Fisher), Carlsbad, CA), and 2µl water with the following conditions, 94°C, 3m; (94°C, 30s; 50°C, 30s; 68°C, 1m)x 35; 68°C 10m. PCR products were visualized on a 1.5% agarose gel and quantified on the BioTek Synergy H1 Hybrid Multi-Mode Plate Reader using the Quand-iT 1X dsDNA HS Assy kit (Thermo Fisher, Carlsbad, CA, USA). A 100ng aliquot of DNA was selected from each of the 300 samples, the pooled DNA was cleaned using the QIAGEN QIAquick PCR purification kit, and quantified using Qubit HS kit (Carlsbad, CA). The quality of the pooled library was verified on the Agilent TapeStation 4200. The prepared library was submitted to the Oregon State University Center for Quantitative Life Sciences (CQLS) for paired end 2x300 bp read sequencing on an Illumina MiSeq System.
Bioinformatic processing
All microbiome DNA sequence analyses and visualization were conducted in R (v 4.3.3) (R Core Team, 2025). Raw reads were filtered for quality, merged, and assigned using the DADA2 R package (v 1.26.0) as previously described (Callahan et al., 2016). In brief, forward and reverse reads were trimmed at 250 and 225 bp, respectively, subsequently merged into contigs, and subject to amplicon sequence variant (ASV) identification. ASVs unannotated at the Phylum level or identified as non-bacterial were removed, which resulted in 674 remaining detected ASVs. Samples containing reads below the minimum required read count (<5000) were dropped from downstream analysis. The final sample number for microbiome analysis was 260. Phylogenetic analysis was conducted using MOTHUR (v 1.46.1) (Schloss et al., 2009) with default parameters as previously described (Sharpton et al., 2021). Phylogeny was inferred using FastTree2 (Price et al., 2010), an approximately-maximum-likelihood method. Microbiome and sample data were contained in a Phyloseq object using the Phyloseq R package (McMurdie and Holmes, 2013), and the tidyverse (v 2.0.0) (Wickham et al., 2019) and microViz (v 0.12.1) R packages were used for downstream data processing, analyzing, and visualization (Barnett et al., 2021). Code for bioinformatic processing are available at https://github.com/sielerjm/Sieler2025:ZF_Temperature_Parasite/.
Microbiome diversity metrics
All microbiome analyses were conducted at the genera level unless otherwise noted. We estimated four alpha-diversity metrics for each microbiome fecal sample: Simpson (1949), Shannon (Weaver, 1963), phylogenetic diversity (Faith’s PD (Faith, 1992); ASVs), and richness. We also estimated beta-diversity between each pair of microbiome fecal samples using three metrics. These included Bray-Curtis (Bray and Curtis, 1957), Canberra (Lance and Williams, 1967), and half-weighted generalized UniFrac (Chen et al., 2012).
Statistical Analyses
All statistical analyses were conducted in R (v 4.3.3) (R Core Team, 2025) with a significance level of α = 0.05, and randomization procedures employed a fixed seed (Breen et al., 2019) to ensure reproducibility. Code for statistical analyses are available at https://github.com/sielerjm/Sieler2025:ZF_Temperature_Parasite/.
Using methods previously described (Kundu et al., 2021), we assessed normality of alpha-diversity scores using Shapiro-Wilk test (Shapiro and Wilk, 1965; R Core Team, 2025), transformed non-normal scores using Tukey’s Ladder of Powers (Dodge, 2008; Kundu et al., 2021) and normalized from 0 to 1 (Sieler et al., 2023) before incorporation into linear models. We used generalized linear models (GLM), we assessed the relationship between alpha-diversity score and experimental parameters. Post hoc Tukey Tests evaluated pairwise comparisons of models using the multcomp R package (v 1.4-25) (Hothorn et al., 2016). We corrected for multiple tests using Benjamini-Hochberg correction (Benjamini and Yekutieli, 2001). Two-way ANOVA was used to determine if the expanded models of these GLMs significantly improved the response variable relative to the null model.
Beta-diversity models were generated using methods described previously (Sieler et al., 2023). In brief, we assessed the relationship between experimental parameters and beta-diversity by applying a step-wise model selection approach as implemented in the capscale function (vegan R package v 2.6-4) (Oksanen et al., 2009). Beta diversity was measured using Bray-Curtis, Canberra, and UniFrac distance measures (Bray and Curtis, 1957; Lance, 1971; Lozupone and Knight, 2005; Legendre and Aceres, 2013). Optimal models were subsequently subject to permutation analysis of variance (PERMANOVA) with anova.cca using the vegan R package to determine if the selected model parameters significantly explained the variation in microbiome composition across samples (Oksanen et al., 2009; Anderson, 2017). Differential abundance was measured using MaAsLin2 (Mallick et al., 2021). We assessed beta-diversity dispersion within groups with betadisper using the vegan R package (Anderson, 2006; Anderson et al., 2006; Oksanen et al., 2009).
To assess the relationship between parasite infection outcomes and experimental parameters, we used negative binomial generalized linear models (GLM) with the glm.nb function from the MASS R package (v 7.3-60.0.1) (Ripley and Venables, 2009) and used the negative binomial distribution. We used the negative binomial distribution to account for overdispersion in the count data, a common characteristic of parasite infection data (Poulin, 2013; Kent et al., 2018; Hammer et al., 2024). Significance of main effects and interactions was assessed using two-way ANOVA implemented through the Anova function with the Car R package (v 3.1-2) (Fox et al., 2001). Post-hoc comparisons were conducted using Tukey’s HSD tests via the emmeans package, where we estimated marginal means and performed pairwise contrasts with p-value adjustment using the Tukey method (Lenth, 2017). Detection method comparisons were analyzed on a subset of samples used for microbiome analysis (120 samples, 20 samples/tank, ~10 samples/time point; Supplementary Table S3C.1). To compare detection methods between wet mount and histology, we used McNemar’s test (McNemar, 1947), with discordant pairs (wet only vs histology only) examined at each temperature and DPE combination through 2×2 contingency tables Supplementary Table S3C.2, 3). Using similar methods as described above, we assessed the relationship between infection outcomes and microbiome diversity using GLMs (Supplementary Table S3B.1).
Random forest analysis
To identify microbial features associated with worm burden, we employed a random forest regression approach using the ranger package in R (v 0.17.0) (Wright, 2015). The analysis was restricted to exposed fish samples and utilized only microbial genera that were previously identified as significantly associated with worm burden through MaAsLin2 analysis (q < 0.1). Prior to model training, the microbiome data underwent compositional normalization through total sum scaling followed by log2 transformation with half-minimum replacement for zero values. The dataset was randomly split into training (80%) and testing (20%) sets, with the random forest model trained using 1000 trees and permutation importance. To ensure robust feature selection, we performed a stability analysis by repeating the model training 100 times with different random seeds, tracking the frequency of genera appearing in the top 10 most important features across iterations. Model performance was evaluated through 10-fold cross-validation, comparing the random forest predictions against a null model that predicted the mean worm burden. Performance metrics included root mean squared error (RMSE) and R² values, with variable importance assessed through the percentage increase in mean squared error when each feature was permuted.
Data availability statement
All code generated during this analysis is available in the GitHub repository at the following URL: https://github.com/sielerjm/Sieler2025__ZF_Temperature_Parasite. The raw sequence files generated during the current study are available at the NCBI Sequence Read Archive (SRA), project number: PRJNA1219243.
Ethics statement
The animal study was approved by Institutional Animal Care and Use Committee (IACUC) at Oregon State University (permit number: 5151). The study was conducted in accordance with the local legislation and institutional requirements.
Author contributions
MS: Data curation, Formal Analysis, Investigation, Methodology, Resources, Software, Supervision, Validation, Visualization, Writing – original draft, Writing – review & editing. TS: Conceptualization, Funding acquisition, Methodology, Project administration, Resources, Supervision, Validation, Writing – original draft, Writing – review & editing. CA: Investigation, Methodology, Writing – original draft, Writing – review & editing. KK: Investigation, Methodology, Writing – original draft, Writing – review & editing. MK: Conceptualization, Investigation, Methodology, Project administration, Resources, Supervision, Visualization, Writing – original draft, Writing – review & editing.
Funding
The author(s) declare that financial support was received for the research and/or publication of this article. This work was supported in part by a National Foundation Grant (#2025457) to TJS, and a fellowship to MJS offered by the Oregon Department of Fish and Wildlife.
Acknowledgments
The authors thank the members of the Oregon State University Center for Quantitative Life Sciences for technical assistance with sequencing and maintenance of our computational infrastructure, and Dr. Corbin Schuster and Kelan Elliot for sample collection assistance. We thank Dr. Alexandra Alexiev for her valuable guidance on ecological processes and her constructive feedback on this manuscript.
Conflict of interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Generative AI statement
The author(s) declare that no Generative AI was used in the creation of this manuscript.
Publisher’s note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
Supplementary material
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/frmbi.2025.1605168/full#supplementary-material
Abbreviations
dpe, days post exposure; dpf, days post fertilization.
References
Acevedo-Whitehouse K. and Duffus A. L. J. (2009). Effects of environmental change on wildlife health. Phil Trans. R Soc. B. 364, 3429–3438. doi: 10.1098/rstb.2009.0128
Ackerly D. D., Loarie S. R., Cornwell W. K., Weiss S. B., Hamilton H., Branciforte R., et al. (2010). The geography of climate change: implications for conservation biogeography. Diversity Distributions. 16, 476–487. doi: 10.1111/j.1472-4642.2010.00654.x
Anderson M. J. (2006). Distance-based tests for homogeneity of multivariate dispersions. Biometrics 62, 245–253. doi: 10.1111/j.1541-0420.2005.00440.x
Anderson M. J. (2017). “Permutational multivariate analysis of variance ( PERMANOVA ),” in Wiley statsRef: statistics reference online, 1st ed. Eds. Kenett R. S., Longford N. T., Piegorsch W. W., and Ruggeri F. (Hoboken, NJ, USA: Wiley), 1–15. doi: 10.1002/9781118445112.stat07841
Anderson M. J., Ellingsen K. E., and McArdle B. H. (2006). Multivariate dispersion as a measure of beta diversity. Ecol. Letters. 9, 683–693. doi: 10.1111/j.1461-0248.2006.00926.x
Bailey C., Segner H., Casanova-Nakayama A., and Wahli T. (2017). Who needs the hotspot? The effect of temperature on the fish host immune response to Tetracapsuloides bryosalmonae the causative agent of proliferative kidney disease. Fish Shellfish Immunol. 63, 424–437. doi: 10.1016/j.fsi.2017.02.039
Barnett D., Arts I., and Penders J. (2021). microViz: an R package for microbiome data visualization and statistics. JOSS 6, 3201. doi: 10.21105/joss.03201
Benjamini Y. and Yekutieli D. (2001). The control of the false discovery rate in multiple testing under dependency. Ann. Statist. 29, 1165–1188. doi: 10.1214/aos/1013699998.full
Bray J. R. and Curtis J. T. (1957). An ordination of the upland forest communities of southern wisconsin. Ecol. Monographs. 27, 325–349. doi: 10.2307/1942268
Breen P., Winters A. D., Nag D., Ahmad M. M., Theis K. R., and Withey J. H. (2019). Internal versus external pressures: effect of housing systems on the zebrafish microbiome. Zebrafish 16, 388–400. doi: 10.1089/zeb.2018.1711
Callahan B. J., McMurdie P. J., Rosen M. J., Han A. W., Johnson A. J. A., and Holmes S. P. (2016). DADA2: High-resolution sample inference from Illumina amplicon data. Nat. Methods 13, 581–583. doi: 10.1038/nmeth.3869
Calvin K., Dasgupta D., Krinner G., Mukherji A., Thorne P. W., Trisos C., et al. (2023). “IPCC, 2023: climate change 2023: synthesis report,” in Contribution of Working Groups I, II and III to the Sixth Assessment Report of the Intergovernmental Panel on Climate Change. Eds. Lee H. and Romero J. (First. Intergovernmental Panel on Climate Change (IPCC, IPCC, Geneva, Switzerland). Available online at: https://www.ipcc.ch/report/ar6/syr/ (Accessed June 23, 2025).
Chen J., Bittinger K., Charlson E. S., Hoffmann C., Lewis J., Wu G. D., et al. (2012). Associating microbiome composition with environmental covariates using generalized UniFrac distances. Bioinformatics 28, 2106–2113. doi: 10.1093/bioinformatics/bts342
Chuang J. S., Frentz Z., and Leibler S. (2019). Homeorhesis and ecological succession quantified in synthetic microbial ecosystems. Proc. Natl. Acad. Sci. U.S.A. 116, 14852–14861. doi: 10.1073/pnas.1901055116
Costello E. K., Stagaman K., Dethlefsen L., Bohannan B. J. M., and Relman D. A. (2012). The application of ecological theory toward an understanding of the human microbiome. Science 336, 1255–1262. doi: 10.1126/science.1224203
Dickerson H. W. (2006). “Ichthyophthirius multifiliis and Cryptocaryon irritans (phylum Ciliophora),” in Fish diseases and disorders Volume 1: protozoan and metazoan infections, 2nd ed. Ed. Woo P. T. K. (CABI, UK), 116–153. doi: 10.1079/9780851990156.0116
Dittmar J., Janssen H., Kuske A., Kurtz J., and Scharsack J. P. (2014). Heat and immunity: an experimental heat wave alters immune functions in three-spined sticklebacks ( asterosteus aculeatus). J. Anim. Ecology. 83, 744–757. doi: 10.1111/1365-2656.12175
Dodge Y. (2008). The concise encyclopedia of statistics. 1st. ed. (New York, NY: Springer New York), 192–194. doi: 10.1007/978-0-387-32833-1_136
Duarte C. M., Agusti S., Barbier E., Britten G. L., Castilla J. C., Gattuso J. P., et al. (2020). Rebuilding marine life. Nature 580, 39–51. doi: 10.1038/s41586-020-2146-7
El-Sayed A. and Kamel M. (2020). Climatic changes and their role in emergence and re-emergence of diseases. Environ. Sci. pollut. Res. 27, 22336–22352. doi: 10.1007/s11356-020-08896-w
Faith D. P. (1992). Conservation evaluation and phylogenetic diversity. Biol. Conserv. 61, 1–10. doi: 10.1016/0006-3207(92)91201-3
Fassarella M., Blaak E. E., Penders J., Nauta A., Smidt H., and Zoetendal E. G. (2021). Gut microbiome stability and resilience: elucidating the response to perturbations in order to modulate gut health. Gut 70, 595–605. doi: 10.1136/gutjnl-2020-321747
Fontaine S. S. and Kohl K. D. (2023). The microbiome buffers tadpole hosts from heat stress: a hologenomic approach to understand host–microbe interactions under warming. J. Exp. Biol. 226, jeb245191. doi: 10.1242/jeb.245191
Fontaine S. S., Mineo P. M., and Kohl K. D. (2022). Experimental manipulation of microbiota reduces host thermal tolerance and fitness under heat stress in a vertebrate ectotherm. Nat. Ecol. Evol. 6, 405–417. doi: 10.1038/s41559-022-01686-2
Fox J., Weisberg S., and Price B. (2001). car: Companion to Applied Regression. Available online at: https://CRAN.R-project.org/package=car (Accessed February 11, 2025).
Gaulke C. A., Martins M. L., Watral V. G., Humphreys I. R., Spagnoli S. T., Kent M. L., et al. (2019). A longitudinal assessment of host-microbe-parasite interactions resolves the zebrafish gut microbiome’s link to Pseudocapillaria tomentosa infection and pathology. Microbiome 7, 10. doi: 10.1186/s40168-019-0622-9
Greenspan S. E., Migliorini G. H., Lyra M. L., Pontes M. R., Carvalho T., Ribeiro L. P., et al. (2020). Warming drives ecological community changes linked to host-associated microbiome dysbiosis. Nat. Clim Change 10, 1057–1061. doi: 10.1038/s41558-020-0899-5
Hammer A. J., Gaulke C. A., Garcia-Jaramillo M., Leong C., Morre J., Sieler M. J., et al. (2024). Gut microbiota metabolically mediate intestinal helminth infection in zebrafish. Ed. Rawls J. F. (Washington, DC, USA), e00545–e00524. doi: 10.1128/msystems.00545-24
Hothorn T., Bretz F., Westfall P., Heiberger R. M., Schuetzenmeister A., Scheibe S., et al. (2016). “Package ‘multcomp.’,” in Simultaneous inference in general parametric models Project for Statistical Computing (Vienna, Austria).
Islam M. J., Kunzmann A., and Slater M. J. (2022). Responses of aquaculture fish to climate change-induced extreme temperatures: A review. J. World Aquaculture Society. 53, 314–366. doi: 10.1111/jwas.12853
Jeltsch F., Roeleke M., Abdelfattah A., Arlinghaus R., Berg G., Blaum N., et al. (2025). The need for an individual-based global change ecology. IBE 1, 1–18. doi: 10.3897/ibe.1.148200
Kashinskaya E. N., Simonov E. P., Poddubnaya L. G., Vlasenko P. G., Shokurova A. V., Parshukov A. N., et al. (2023). Trophic diversification and parasitic invasion as ecological niche modulators for gut microbiota of whitefish. Front. Microbiol. 14. doi: 10.3389/fmicb.2023.1090899/full
Kent M. L., Bishop-Stewart J. K., Dvm J. L. M., and Spitsbergen J. M. (2002). Pseudocapillaria tomentosa, a Nematode Pathogen, and Associated Neoplasms of Zebrafish (Danio rerio) Kept in Research Colonies. Comp. Med. 52, 354–358.
Kent M., Buchner C., Watral V., Sanders J., LaDu J., Peterson T., et al. (2011). Development and maintenance of a specific pathogen-free (SPF) zebrafish research facility for Pseudoloma neurophilia. Dis. Aquat Org. 95, 73–79. doi: 10.3354/dao02333
Kent M., Gaulke C., Watral V., and Sharpton T. (2018). Pseudocapillaria tomentosa in laboratory zebrafish Danio rerio: patterns of infection and dose response. Dis. Aquat Org. 131, 121–131. doi: 10.3354/dao03286
Kent M. L., Watral V., Villegas E. N., and Gaulke C. A. (2019). Viability of Pseudocapillaria tomentosa eggs exposed to heat, ultraviolet light, chlorine, iodine, and desiccation. Zebrafish 16, 460–468. doi: 10.1089/zeb.2019.1736
Kozich J. J., Westcott S. L., Baxter N. T., Highlander S. K., and Schloss P. D. (2013). Development of a dual-index sequencing strategy and curation pipeline for analyzing amplicon sequence data on the miSeq illumina sequencing platform. Appl. Environ. Microbiol. 79, 5112–5120. doi: 10.1128/aem.01043-13
Kumar V., Roy S., Parida S. N., Bisai K., Dhar S., Jana A. K., et al. (2024). Deciphering the impact of endoparasitic infection on immune response and gut microbial composition of Channa punctata. Front. Cell Infect. Microbiol. 14. doi: 10.3389/fcimb.2024.1296769/full
Kundu P., Torres E. R. S., Stagaman K., Kasschau K., Okhovat M., Holden S., et al. (2021). Integrated analysis of behavioral, epigenetic, and gut microbiome analyses in AppNL-G-F, AppNL-F, and wild type mice. Sci. Rep. 11, 4678. doi: 10.1038/s41598-021-83851-4
Lance G. N. (1971). A note on a new divisive classificatory program for mixed data. Comput. J. 14, 154–155. doi: 10.1093/comjnl/14.2.154
Lance G. N. and Williams W. T. (1967). Mixed-data classificatory programs I - agglomerative systems. Aust. Comput. J. 1, 15–20. doi: 10.1093/comjnl/9.4.373
Lardeux F. and Cheffort J. (1997). Temperature thresholds and statistical modelling of larval Wuchereria bancrofti (Filariidea: Onchocercidae) developmental rates. Parasitology 114, 123–134. doi: 10.1017/S0031182096008359
Legendre P. and Aceres M. D. C. (2013). Beta diversity as the variance of community data: dissimilarity coefficients and partitioning. Ecol. Lett. 16, 951–963. doi: 10.1111/ele.12141
Lenth R. V. (2017). emmeans: Estimated Marginal Means, aka Least-Squares Means. Available online at: https://CRAN.R-project.org/package=emmeans (Accessed February 11, 2025).
Ley R. E., Lozupone C. A., Hamady M., Knight R., and Gordon J. I. (2008). Worlds within worlds: evolution of the vertebrate gut microbiota. Nat. Rev. Microbiol. 6, 776–788. doi: 10.1038/nrmicro1978
Li P., Zhang J., Liu X., Gan L., Xie Y., Zhang H., et al. (2022). The function and the affecting factors of the zebrafish gut microbiota. Front. Microbiol. 13. doi: 10.3389/fmicb.2022.903471/full
López-Olmeda J. F. and Sánchez-Vázquez F. J. (2011). Thermal biology of zebrafish (Danio rerio). J. Thermal Biol. 36, 91–104. doi: 10.1016/j.jtherbio.2010.12.005
Lozupone C. and Knight R. (2005). UniFrac: a new phylogenetic method for comparing microbial communities. Appl. Environ. Microbiol. 71, 8228–8235. doi: 10.1128/aem.71.12.8228-8235.2005
Makrinos D. L. and Bowden T. J. (2016). Natural environmental impacts on teleost immune function. Fish Shellfish Immunol. 53, 50–57. doi: 10.1016/j.fsi.2016.03.008
Mallick H., Rahnavard A., McIver L. J., Ma S., Zhang Y., Nguyen L. H., et al. (2021). Multivariable association discovery in population-scale meta-omics studies. PloS Comput. Biol. 17, e1009442. doi: 10.1371/journal.pcbi.1009442
Marandi A., Jensen A. M., and Von Gersdorff Jørgensen L. (2025). Pseudocapillaria tomentosa (Nematoda: Capillariidae) in fish and its significance in comprehending host-parasite relationships: A review. Curr. Res. Parasitol. Vector-Borne Diseases. 7, 100265. doi: 10.1016/j.crpvbd.2025.100265
Martins M. L., Watral V., Rodrigues-Soares J. P., and Kent M. L. (2017). A method for collecting eggs of Pseudocapillaria tomentosa (Nematoda: Capillariidae) from zebrafish Danio rerio and efficacy of heat and chlorine for killing the nematode’s eggs. J. Fish Diseases. 40, 169–182. doi: 10.1111/jfd.12501
Mas-Coma S., Valero M. A., and Bargues M. D. (2008). Effects of climate change on animal and zoonotic helminthiases. Rev. Sci. Tech. 27, 443–457. doi: 10.20506/rst.27.2.1822
McMurdie P. J. and Holmes S. (2013). phyloseq: an R package for reproducible interactive analysis and graphics of microbiome census data. PloS One 8, e61217. doi: 10.1371/journal.pone.0061217
McNemar Q. (1947). Note on the sampling error of the difference between correlated proportions or percentages. Psychometrika 12, 153–157. doi: 10.1007/BF02295996
Molnár P. K., Dobson A. P., and Kutz S. J. (2013). Gimme shelter – the relative sensitivity of parasitic nematodes with direct and indirect life cycles to climate change. Global Change Biol. 19, 3291–3305. doi: 10.1111/gcb.12303
Moravec F. (1983). Observations on the bionomy of the nematode Pseudocapillaria brevispicula [carp, tench, ponds, parasite. Folia parasitologica. 30, 229–241. doi: 10.3109/10826088309033066
Moravec F. (1984). FIRST RECORD OF THE NEMATODE PSEUDOCAPILLARIA BREVISPICULA. Folia PARASITOLOGICA (PRAHA). 31, 241–245.
Moravec F., Wolter J., and Körting W. (1999). Some nematodes and acanthocephalans from exotic ornamental freshwater fishes imported into Germany. Folia parasitologica. 46, 296–310.
Oksanen J., Kindt R., Legendre P., Hara B., Simpson G., Solymos P., et al. (2009). The vegan package. (Vienna, Austria).
Poulin R. (2013). Explaining variability in parasite aggregation levels among host samples. Parasitology 140, 541–546. doi: 10.1017/S0031182012002053
Price M. N., Dehal P. S., and Arkin A. P. (2010). FastTree 2 – approximately maximum-likelihood trees for large alignments. Poon AFY editor. PloS One 5, e9490. doi: 10.1371/journal.pone.0009490
Rawls J. F., Samuel B. S., and Gordon J. I. (2004). Gnotobiotic zebrafish reveal evolutionarily conserved responses to the gut microbiota. Proc. Natl. Acad. Sci. U.S.A. 101, 4596–4601. doi: 10.1073/pnas.0400706101
R Core Team (2025). R: A Language and Environment for Statistical computing (Vienna, Austria: R Foundation for Statistical Computing, Vienna). Available online at: https://www.R-project.org/ (Accessed February 11, 2025).
Relman D. A. (2012). The human microbiome: ecosystem resilience and health. Nutr. Rev. 70, S2–S9. doi: 10.1111/j.1753-4887.2012.00489.x
Ripley B. and Venables B. (2009). MASS: Support Functions and Datasets for Venables and Ripley’s MASS. Available online at: https://CRAN.R-project.org/package=MASS (Accessed February 11, 2025).
Roeselers G., Mittge E. K., Stephens W. Z., Parichy D. M., Cavanaugh C. M., Guillemin K., et al. (2011). Evidence for a core gut microbiota in the zebrafish. ISME J. 5, 1595–1608. doi: 10.1038/ismej.2011.38
Sanders J. L., Zhou Y., Moulton H. M., Moulton Z. X., McLeod R., Dubey J. P., et al. (2015). The zebrafish, Danio rerio, as a model for Toxoplasma gondii : an initial description of infection in fish. J. Fish Diseases. 38, 675–679. doi: 10.1111/jfd.12393
Schlomann B. H. and Parthasarathy R. (2019). Timescales of gut microbiome dynamics. Curr. Opin. Microbiol. 50, 56–63. doi: 10.1016/j.mib.2019.09.011
Schloss P. D., Westcott S. L., Ryabin T., Hall J. R., Hartmann M., Hollister E. B., et al. (2009). Introducing mothur: open-source, platform-independent, community-supported software for describing and comparing microbial communities. Appl. Environ. Microbiol. 75, 7537–7541. doi: 10.1128/AEM.01541-09
Schuster C. J., Leong C., Kasschau K. D., Sharpton T. J., and Kent M. L. (2023). Early detection of Pseudocapillaria tomentosa by qPCR in four lines of zebrafish, Danio rerio (Hamilton 1882). J. Fish Diseases. 46, 619–627. doi: 10.1111/jfd.13773
Semova I., Carten J. D., Stombaugh J., Mackey L. C., Knight R., Farber SA, et al. (2012). Microbiota regulate intestinal absorption and metabolism of fatty acids in the zebrafish. Cell Host Microbe 12, 277–288. doi: 10.1016/j.chom.2012.08.003
Sepulveda J. and Moeller A. H. (2020). The effects of temperature on animal gut microbiomes. Front. Microbiol. 11. doi: 10.3389/fmicb.2020.00384
Shapiro S. S. and Wilk M. B. (1965). An analysis of variance test for normality (complete samples). Biometrika 52, 591–611. doi: 10.1093/biomet/52.3-4.591
Sharpton T. J., Stagaman K., Sieler M. J., Arnold H. K., and Davis E. W. (2021). Phylogenetic integration reveals the zebrafish core microbiome and its sensitivity to environmental exposures. Toxics 9, 10. doi: 10.3390/toxics9010010
Sieler M. J., Al-Samarrie C. E., Kasschau K. D., Varga Z. M., Kent M. L., and Sharpton T. J. (2023). Disentangling the link between zebrafish diet, gut microbiome succession, and Mycobacterium chelonae infection. Anim. microbiome. 5, 38. doi: 10.1186/s42523-023-00254-8
Sommer F., Anderson J. M., Bharti R., Raes J., and Rosenstiel P. (2017). The resilience of the intestinal microbiota influences health and disease. Nat. Rev. Microbiol. 15, 630–638. doi: 10.1038/nrmicro.2017.58
Stagaman K., Sharpton T. J., and Guillemin K. (2020). Zebrafish microbiome studies make waves. Lab. Anim. (NY). 49, 201–207. doi: 10.1038/s41684-020-0573-6
Stephens W. Z., Burns A. R., Stagaman K., Wong S., Rawls J. F., Guillemin K., et al. (2016). The composition of the zebrafish intestinal microbial community varies across development. ISME J. 10, 644–654. doi: 10.1038/ismej.2015.140
Sydeman W. J., Poloczanska E., Reed T. E., and Thompson S. A. (2015). Climate change and marine vertebrates. Science 350, 772–777. doi: 10.1126/science.aac9874
Tomanek L. (2010). Variation in the heat shock response and its implication for predicting the effect of global climate change on species’ biogeographical distribution ranges and metabolic costs. J. Exp. Biol. 213, 971–979. doi: 10.1242/jeb.038034
Waddington C. H. (1942). Canalization of development and the inheritance of acquired characters. Nature. 150, 563–565. doi: 10.1038/150563a0
Wang B., Zhang S. Q., Dong J. L., Li Y., Jin Y. X., Xiao H. W., et al. (2022). Ambient temperature structures the gut microbiota of zebrafish to impact the response to radioactive pollution. Environ. Pollution. 293, 118539. doi: 10.1016/j.envpol.2021.118539
Wickham H., Averick M., Bryan J., Chang W., McGowan L., François R., et al. (2019). Welcome to the tidyverse. JOSS 4, 1686. doi: 10.21105/joss.01686
Wright M. N. (2015). ranger: A Fast Implementation of Random Forests. Available online at: https://CRAN.R-project.org/package=ranger (Accessed February 11, 2025).
Zaneveld J. R., McMinds R., and Vega Thurber R. (2017). Stress and stability: applying the Anna Karenina principle to animal microbiomes. Nat. Microbiol. 2, 17121. doi: 10.1038/nmicrobiol.2017.121
Keywords: development, infection, helminth, temperature, climate change, Pseudocapillaria tomentosa, zebrafish, gut microbiome
Citation: Sieler MJ Jr., Al-Samarrie CE, Kasschau KD, Kent ML and Sharpton TJ (2025) Modeling the zebrafish gut microbiome’s resistance and sensitivity to climate change and parasite infection. Front. Microbiomes 4:1605168. doi: 10.3389/frmbi.2025.1605168
Received: 02 April 2025; Accepted: 03 July 2025;
Published: 22 July 2025.
Edited by:
Ana Elena Ahuir-Baraja, Universidad CEU Cardenal Herrera, SpainReviewed by:
Hualong Hong, Xiamen University, ChinaQinnan Yang, University of Michigan, United States
Copyright © 2025 Sieler, Al-Samarrie, Kasschau, Kent and Sharpton. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Thomas J. Sharpton, VGhvbWFzLlNoYXJwdG9uQG9yZWdvbnN0YXRlLmVkdQ==